resource use and partitioning by two co-occurring freshwater gastropod species
TRANSCRIPT

Arch Hydrobiol 159 2 229ndash251 Stuttgart February 2004
Resource use and partitioning by two co-occurringfreshwater gastropod species
Paola Lombardo1 2 and G Dennis Cooke1 3
Department of Biological Sciences and Water Resources Research InstituteKent State University USA
With 8 figures and 6 tables
Abstract Many trophic generalist gastropods exert predation-based control on theirtrophic resources However most grazing experiments have been carried out withsingle snail species and little is known about snail effects on trophic resources by coe-xisting species Substrate colonization and feeding behavior of Physa spp (Physidae)and Helisoma trivolvis (Planorbidae) were observed in single- and two-species labora-tory trials Spatial overlap between Physa and H trivolvis was derived from distribu-tions of snail individuals on trophic (living macrophytes periphyton and leaf litter)and spatial resources (aquarium space divided into quadrats) Food consumption wasdetermined as biomass changes with respect to snail-devoid controls Snails reducedthe degree of spatial overlap when coexisting though Physa appeared to affect H tri-volvis distribution more than vice-versa regardless of snail density Both species pre-ferred leaf litter as a substrate and both increased nutrient (phosphorus) concentrationof water Periphyton and leaf litter biomass loss was similar in all snail-present aqua-ria with only inedible leaf veins left intact at the end of the 10-day experiment Bio-mass of living macrophytes increased in all Physa-only aquaria and in two-specieshigh density aquaria suggesting that macrophyte enhancement may be possible inhigh-density Physa-dominated snail assemblages
Key words gastropods resource utilization herbivory overlap behavior Physa sppHelisoma trivolvis
Introduction
Aquatic gastropods are efficient grazers (eg Cattaneo amp Kalff 1986) andoften control trophic resources in a predator-like fashion (Rosemond et al
1 Authorsrsquo address Department of Biological Sciences and Water Resources Re-search Institute Kent State University Kent OH 44242-0001 USA2 Current address La Sapienza State University of Rome Dept of Genetics amp Molec-ular Biology ndash Ecology Area Via dei Sardi 70 00185 Rome Italy
DOI 1011270003-913620040159-0229 0003-9136040159-0229 $ 575atilde 2004 E Schweizerbartrsquosche Verlagsbuchhandlung D-70176 Stuttgart
230 Paola Lombardo and G Dennis Cooke
1993 Stoner et al 1995 Muntildeoz et al 2000) Many gastropod species alsoshare general habitat and trophic spectra andor preferences (Brown 1982Lombardo amp Cooke 2002) Freshwater pulmonates are poorer competitorsthan prosobranchs (Brown et al 1998) but have broader trophic spectra(Brown 1991) seem to adapt better to existing trophic conditions (Brendel-berger 1997) and coexist in most natural conditions sometimes greatly over-lapping in habitat or trophic utilization (Brown 1982 1997 Jokinen 1987)
Snails exhibit complex and flexible behaviors when coexisting with otheranimals (e g Rochette et al 1995 Turner et al 1999) Some pulmonatespecies influence resource utilization by coexisting invertebrate grazers evenwhen resources are not limiting (Gresens 1995) Though overlap does notnecessarily lead to competition (Rossi 1985) changes in resource utilizationby coexisting snail species may impact the whole food web Yet direct studiesof interspecific interactions among freshwater snails are uncommon (Brown1982)
Freshwater snails in temperate ecosystems rarely consume living macro-phytes (Broumlnmark 1989) though they are able to do so (Sheldon 1987 Lom-bardo amp Cooke 2002) Indeed macrophyte growth may be enhanced in thepresence of gastropods (Martin et al 1992 review in Broumlnmark amp Ver-maat 1998) through grazing-induced relief from light-limiting epiphytes(Sand-Jensen amp Borum 1991) andor through chemical conditioning ofwater (Underwood 1991) Most studies of snail-macrophyte interactions in-volved single snail species and little is known of effects on plants in multiple-species assemblages In these assemblages normally plant-avoiding snail spe-cies may consume living macrophytes with possible effects on ecosystem-level processes A better knowledge of snail interspecific interactions includ-ing effects on food resources is needed in order to evaluate the mechanisms ofsnail effects on aquatic plants
This investigation addressed resource partitioning by two common fresh-water pulmonate gastropods Physa spp Say (Physidae) and Helisoma trivol-vis Say (Planorbidae) alone and in combination from snail distribution dataon trophic (periphyton living macrophytes and decomposing leaf litter) andspatial resources (aquarium walls divided into quadrats) in indoor aquaria Re-source consumption and effects on nutrient (phosphorus) concentration bysnails also were determined
Materials and methods
The experiment involved a cross-gradient of snail densities and species compositionand was carried out in triplicate (Fig 1) in randomly assigned clear plastic aquariaover 10 days Each aquarium contained 275 liters of untreated tap water (Fig 2) left
Resource use and partitioning by two co-occurring snail species 231
Fig 1 Experimental design Circles in each experimental condition symbolize totaland relative snail biomass number of snail individuals not drawn to scale Low-highdensity crossed treatments were not performed Number of adult snails in low-densitytrials were 8 and 4 for Physa and 2 and 1 for H trivolvis (single-species and two-spe-cies trials respectively) High densities were 3 acute low densities Each experimentalcondition was carried out in triplicate
Fig 2 Schematic representation of an experimental aquarium subdivided in six quad-rats (AndashF) with trophic resources (one C demersum sprig periphyton on a glass tileand decomposing maple leaves) superimposed on quadrats Layout was the same ineach aquarium Snails were added according to the experimental design in Fig 1
232 Paola Lombardo and G Dennis Cooke
undisturbed for 24 hours before the experimentrsquos beginning (t0) to allow chlorine to es-cape Tap water was chosen because of its high calcium content necessary for snailmetabolism (Brown 1991) and low total phosphorus content
Physical layout was the same in all aquaria Trophic resources were three types ofknown snail food periphyton grown on glass tiles living macrophytes and coarse ter-restrial leaf litter Spatial resources were six quadrats of roughly similar size markedby lines on aquaria outer surfaces and labelled A through F (Fig 2) Wall surface waslower for central quadrats (C and D) but snail utilization of walls remained minimaland was not quantified Available surface area for periphyton was less than for macro-phytes and leaf litter but total edible quantity was similar for each food type withcoarser or tougher material assumed inedible for snails (e g Thomas et al 1985) Allperiphytic material was considered edible
Methods for trophic resource preparation and biomass determination followedthose in Lombardo amp Cooke (2002) Periphyton was a thick layer of algae pro-tozoans and unrecognizable detritus developed on rough-surface (ldquopattern 62rdquo)102 mm acute 102 mm glass tiles during a 60-day colonization period with inocula fromthe vegetated littoral zone of East Twin Lake Portage County Northeast Ohio (USA)East Twin Lake is a mesotrophic (TP ~ 20 mgl) clear-water natural lake with abundantaquatic vegetation and a rich diverse snail assemblage throughout its littoral zone
Periphyton was manually scraped with a plastic (non-absorbant) brush after re-trieving the tiles at the end of the experiment (tF) Some periphytic material was una-voidably lost during retrieval because of the very loose attachment to the tiles andsloughing of older algal patches leading to high variability in final biomass for controltiles however statistical power for grazed vs ungrazed comparisons remained intactThe scraped material was collected on 45-mm fiberglass filters and dry-weighed afteroven-desiccation at 60 ƒC for 72 hours An additional set of six tiles was used at the be-ginning of the experiment (t0) to determine initial periphyton biomass using the samemethod Periphyton did not develop on aquarium surfaces in either snail-present orcontrol aquaria
Macrophytes were ~ 15-cm long Ceratophyllum demersum L apical sprigs alsocollected from East Twin Lakersquos littoral zone where this plant was common C de-mersum was covered with a thin biofilm of loosely attached diatoms amorphous detri-tus and protozoans as revealed from a qualitative microscopic examination (100 acutendash 400 acute) The biofilm on C demersum was termed epiphyton to distinguish it from theperiphyton grown on glass tiles Periphyton composition was assumed to be similar tothe epiphyton collected from the same habitat Biomass change for plants was ex-pected to be less marked than for periphyton and maple leaves therefore C demersumbiomass was determined as living weight to allow for more precise t0 and tF determina-tions on the same sprigs Sprigs were partially dried on water-absorbing paper andweighed once they did not release water upon gentle shaking Weighing conditionswere uniform (rdry-wet = 0844 p lt 005 n = 6) Contribution of epiphyton to sprig bio-mass was considered negligible Each aquarium received ~ 5ndash7 g of living C demer-sum (one-way type I ANOVA F = 0656 p gt 065)
Air-dried maple (Acer sp) leaves were rewetted for 48 hours to leach tannins Let-ter-tagged three-leaf packs were assembled with a nylon string at the petioles and in-dividually dry-weighed after oven-desiccation at 60 ƒC for 72 hours Leaf packs were
Resource use and partitioning by two co-occurring snail species 233
conditioned with natural microflora in shallow indoor aquaria for three weeks beforet0 Post-conditioning leaf pack biomass at t0 was determined by adjusting each packrsquospre-conditioning biomass for the average post-conditioning biomass from six addi-tional packs that were dry-weighed at t0 The microflora on leaf packs was not ana-lyzed microscopically but was assumed to be a mixture of microfungi bacteria andto a lesser extent algae Snail individuals and feces were gently removed before finalbiomass determinations
Total phosphorus concentration (TP) in the aquaria was determined (ascorbic acidmethod Clesceri et al 1989) at t0 mid-term (t5 = day 5) and at the end of the experi-ment (t10 = tF) All inert material (e g aquaria glass tiles glassware) was analyticallycleaned in 10 HCl and rinsed with distilled water before use Initial TP in randomlychosen aquaria (after introduction of trophic resources and before introduction ofsnails) was 198 plusmn 49 mgl (average plusmn standard error n = 6) Mid-term and final TPwere determined with one 10-ml sample per aquarium Evaporation which remainedminimal did not affect TP determination
Water temperature was determined from randomly chosen aquaria on three datesand remained at 170 plusmn 05 ƒC (avg plusmn std error n = 3) Daylight diffused into the ex-periment locale Parallel aquarium orientation and equal distance from windows pro-vided uniform light The D L cycle followed the natural ~ 15 09 photoperiod (June)with midday irradiance at the water surface of 90 plusmn 13 mE mndash2 sndash1 (avg plusmn std error n= 6) a level expected to support algal and C demersum growth despite being close tolimiting conditions (e g Sand-Jensen amp Madsen 1991)
Snails were collected from an unnamed pond near West Branch Reservoir (PortageCounty) where macrophytes attached algae and allochthonous macrodetritus of plantorigin were abundant Physa spp included Physa integra Haldeman and Physa gy-rina Say which are difficult to distinguish in vivo but share similar ecological char-acteristics (Clampitt 1970) Though P gyrina can reach a larger size body size of thePhysa individuals used in the experiment was similar The ldquolow-densityrdquo conditionroughly represented a low to intermediate natural density of Physa in macropythe-richlittoral zones in northeast Ohio shallow lakes with potential snail predators (~ 09ndash13individualsg Lombardo 2001) Low-density Physa aquaria received 8 adult individu-als High density was set as 3 acute low density Because H trivolvis individual living bio-mass is three- to four-fold that of Physa (P Lombardo unpubl data from repeatedweight trials) each aquarium received four Physa individuals for every H trivolvis in-dividual to maintain similar snail biomass and hence potential grazing pressure in eachlow- or high-density snail-present aquarium
Snails were fed ad libitum with diverse plant material collected in situ and thenstarved for ~12 hrs before t0 to avoid trophic bias (Brendelberger 1994) Snails werethe last biological material introduced at t0 All individuals (Physa 4ndash8 mm shelllength H trivolvis 7ndash15 mm shell diameter) were placed in quadrat C providing sim-ilar initial access to trophic resources during evening hours Snails were not disturbedor replaced until tF Snail mortality at tF was low with a maximum of two dead indi-viduals recorded in two high-density Physa aquaria All H trivolvis individuals wererecovered alive at tF In one high-density H trivolvis aquarium an additional individ-ual (probably introduced accidentally as a juvenile on C demersum) was observedstarting from t7 This individual was included in distribution and overlap calculations
234 Paola Lombardo and G Dennis Cooke
starting from t7 Food consumption data from the aquarium with the additional individ-ual were similar to the other replicates Egg masses or newborns were not observed
Numbers of snail individuals on each trophic and spatial resource were recordeddaily at 08 00ndash10 00 and 18 00ndash21 00 hours Differences in snail distribution pat-terns on trophic and spatial resources were tested with a repeated-measures ANOVAfollowed by SNK (Student-Newman-Keuls) analysis at p pound 010 for significant outco-mes (Zar 1984) Day zero distributions (all individuals on quadrat C) were excludedfrom statistical analysis
Species overlap on trophic and spatial resources was calculated using the overlapindex a (Levins 1968)
aij =Sh (pi middot pj) a ji =
Sh (pj middot pi)
Sh (pi)2 Sh (pj)
2
where pi is the proportion of species i on the hth resource and pj is the proportion ofspecies j on the hth resource The aij value quantifies the overlap of species i over spe-cies j and is not necessarily symmetrical with the aji overlap of species j over speciesi depending on general resource use by each species [quantified by Sh (pi)
2] Asym-metrical indices are preferred because they better represent natural situations (Slo-bodchikoff amp Schulz 1980) The a values for the twice-daily observations did notfollow any discernible temporal pattern and were averaged over the 10-day periodAveraged a values expressed a general concept and are not to be intended as Mayrsquos(1972) ldquoaverage alphardquo
Distribution-based snail preference for resources was assessed using the modifiedversion of Roarsquos (1992) MANOVA procedure used and described in Lombardo ampCooke (2002) Such a method assesses preference or rejection of a resource relative tothe other resources ldquoValuerdquo of trophic resources determined with this method was assubstrate only
Food consumption by snails was measured as changes in resource biomass ingrazed vs ungrazed aquaria (controls) Biomass change was normalized as finalweight expressed as percent of initial weight Percent changes of less than 100 indi-cated biomass loss and percent changes higher than 100 indicated biomass gainBiomass of food items remained unchanged from initial values in snail absence (two-tailed t-tests for initial vs final biomass in control aquaria tleaf packs = 0123 tmacrophytes
= 0234 tperiphyton = 1645 n1 = n2 = 3 d f = 4 and n s for all three) and any changein snail-present aquaria was ascribed to snail presence Snail biomass changes were notquantified but low mortality and apparent new shell growth suggested non-limitingconditions for snails
Significant differences among treatments for food item biomass and TP were deter-mined with one-way type I ANOVAs SNK tests with p pound 010 were carried out forsignificant ANOVAs (Zar 1984) Linear correlations and t-tests were performed forselected data sets Small sample size (n = 3) precluded testing for normality and equa-lity of variances (e g Zar 1984) but data were neither additive nor scaled Thus pa-rametric analysis on untransformed data was performed also due to negligible testpower difference with non-parametric ANOVAs with equal sample size (Zar 1984)and negligible differences with non-parametric MANOVA outcomes (Roa 1992)
Resource use and partitioning by two co-occurring snail species 235
Results
Snail distribut ion and overlap on resources
Snails accumulated on trophic resources (though gradually on maple leafpacks) and were found on empty quadrats (spatial resources) in lower num-bers (Figs 3 and 4 Table 1) Snails were found in significantly higher num-bers on leaf packs regardless of density and species assemblage (Table 1)Preference for leaf packs was striking for both species (Fig 5) Physarsquos higherrelative presence on leaf packs at high snail density (Table 2) was accompa-nied by a significant reduction in H trivolvisrsquo average percent distribution(Table 3) and preference degree (Fig 5) from single- to two-species aquariarather than reflecting an increased preference by Physa (Table 3 Fig 5) Rela-tive distribution on leaves was independent of snail density in single-speciesaquaria but decreased with increasing density for coexisting snails (Table 4)
Physarsquos initial accumulation on periphyton decreased (with temporary tileabandonment at low density) shortly after an evident reduction to almostldquoclean tilerdquo conditions in periphyton biomass was observed at t3ndash t4 in allsnail-present aquaria (Fig 3) H trivolvis visits on periphyton were relativelyfrequent in single-species aquaria but rare when coexisting with Physa(Fig 4) especially at low density (Table 3) Snails rejected periphyton tiles athigh density but coexistence led to an increase in Physa preference (from neu-
Table 1 Differences in snail distribution (as number of individuals) on spatial (emptyquadrats) and trophic resources tested with repeated-measures ANOVAs followed bySNK multiple comparisons Critical values for F are one-tailed values for numeratord f = 8 and denominator d f = 152 Levels of significance are ns (p gt 010) (ppound 010) (p pound 005) and (p pound 001) Significantly different snail distributions onthe trophic and spatial resources resulting from SNK tests (p pound 010) are symbolizedwith different letters (a = lowest 10-day average snail density on resource progressi-vely higher densities listed alphabetically)
SNK (p pound010)
ANOVA trophic resources empty quadrats
leaf peri- macro-experimental condition F p packs phyton phytes A B C D E F
Physa low density 41046 c b b a ab ab a a abPhysa low density with H trivolvis 60252 c b a a a a a a aH trivolvis low density 16331 d a c ab ab a ab ab abH trivolvis low density with Physa 12221 b a a a a a a a a
Physa high density 121278 c ab b ab ab a a a aPhysa high density with H trivolvis 75880 c a b a a a a a aH trivolvis high density 31430 b a a a a a a a aH trivolvis high density with Physa 8239 c ab ab a a ab ab ab ab
236 Paola Lombardo and G Dennis Cooke
Fig 3 Distribution of Physa on spatial (food-devoid quadrats on aquarium surfaces)and trophic resources as average snail numbers on each trophic substrate and on allquadrats One morning (m) and one evening (e) observation per day on days 1 through10 Total numbers per aquarium given on x axis Number of live individuals on eachobservation normalized to initial numbers (when applicable) for graph comparability
Resource use and partitioning by two co-occurring snail species 237
Fig 4 Distribution of H trivolvis on spatial (food-devoid quadrats on aquarium surfa-ces) and trophic resources during the experiment Explanations as in Fig 3
238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

230 Paola Lombardo and G Dennis Cooke
1993 Stoner et al 1995 Muntildeoz et al 2000) Many gastropod species alsoshare general habitat and trophic spectra andor preferences (Brown 1982Lombardo amp Cooke 2002) Freshwater pulmonates are poorer competitorsthan prosobranchs (Brown et al 1998) but have broader trophic spectra(Brown 1991) seem to adapt better to existing trophic conditions (Brendel-berger 1997) and coexist in most natural conditions sometimes greatly over-lapping in habitat or trophic utilization (Brown 1982 1997 Jokinen 1987)
Snails exhibit complex and flexible behaviors when coexisting with otheranimals (e g Rochette et al 1995 Turner et al 1999) Some pulmonatespecies influence resource utilization by coexisting invertebrate grazers evenwhen resources are not limiting (Gresens 1995) Though overlap does notnecessarily lead to competition (Rossi 1985) changes in resource utilizationby coexisting snail species may impact the whole food web Yet direct studiesof interspecific interactions among freshwater snails are uncommon (Brown1982)
Freshwater snails in temperate ecosystems rarely consume living macro-phytes (Broumlnmark 1989) though they are able to do so (Sheldon 1987 Lom-bardo amp Cooke 2002) Indeed macrophyte growth may be enhanced in thepresence of gastropods (Martin et al 1992 review in Broumlnmark amp Ver-maat 1998) through grazing-induced relief from light-limiting epiphytes(Sand-Jensen amp Borum 1991) andor through chemical conditioning ofwater (Underwood 1991) Most studies of snail-macrophyte interactions in-volved single snail species and little is known of effects on plants in multiple-species assemblages In these assemblages normally plant-avoiding snail spe-cies may consume living macrophytes with possible effects on ecosystem-level processes A better knowledge of snail interspecific interactions includ-ing effects on food resources is needed in order to evaluate the mechanisms ofsnail effects on aquatic plants
This investigation addressed resource partitioning by two common fresh-water pulmonate gastropods Physa spp Say (Physidae) and Helisoma trivol-vis Say (Planorbidae) alone and in combination from snail distribution dataon trophic (periphyton living macrophytes and decomposing leaf litter) andspatial resources (aquarium walls divided into quadrats) in indoor aquaria Re-source consumption and effects on nutrient (phosphorus) concentration bysnails also were determined
Materials and methods
The experiment involved a cross-gradient of snail densities and species compositionand was carried out in triplicate (Fig 1) in randomly assigned clear plastic aquariaover 10 days Each aquarium contained 275 liters of untreated tap water (Fig 2) left
Resource use and partitioning by two co-occurring snail species 231
Fig 1 Experimental design Circles in each experimental condition symbolize totaland relative snail biomass number of snail individuals not drawn to scale Low-highdensity crossed treatments were not performed Number of adult snails in low-densitytrials were 8 and 4 for Physa and 2 and 1 for H trivolvis (single-species and two-spe-cies trials respectively) High densities were 3 acute low densities Each experimentalcondition was carried out in triplicate
Fig 2 Schematic representation of an experimental aquarium subdivided in six quad-rats (AndashF) with trophic resources (one C demersum sprig periphyton on a glass tileand decomposing maple leaves) superimposed on quadrats Layout was the same ineach aquarium Snails were added according to the experimental design in Fig 1
232 Paola Lombardo and G Dennis Cooke
undisturbed for 24 hours before the experimentrsquos beginning (t0) to allow chlorine to es-cape Tap water was chosen because of its high calcium content necessary for snailmetabolism (Brown 1991) and low total phosphorus content
Physical layout was the same in all aquaria Trophic resources were three types ofknown snail food periphyton grown on glass tiles living macrophytes and coarse ter-restrial leaf litter Spatial resources were six quadrats of roughly similar size markedby lines on aquaria outer surfaces and labelled A through F (Fig 2) Wall surface waslower for central quadrats (C and D) but snail utilization of walls remained minimaland was not quantified Available surface area for periphyton was less than for macro-phytes and leaf litter but total edible quantity was similar for each food type withcoarser or tougher material assumed inedible for snails (e g Thomas et al 1985) Allperiphytic material was considered edible
Methods for trophic resource preparation and biomass determination followedthose in Lombardo amp Cooke (2002) Periphyton was a thick layer of algae pro-tozoans and unrecognizable detritus developed on rough-surface (ldquopattern 62rdquo)102 mm acute 102 mm glass tiles during a 60-day colonization period with inocula fromthe vegetated littoral zone of East Twin Lake Portage County Northeast Ohio (USA)East Twin Lake is a mesotrophic (TP ~ 20 mgl) clear-water natural lake with abundantaquatic vegetation and a rich diverse snail assemblage throughout its littoral zone
Periphyton was manually scraped with a plastic (non-absorbant) brush after re-trieving the tiles at the end of the experiment (tF) Some periphytic material was una-voidably lost during retrieval because of the very loose attachment to the tiles andsloughing of older algal patches leading to high variability in final biomass for controltiles however statistical power for grazed vs ungrazed comparisons remained intactThe scraped material was collected on 45-mm fiberglass filters and dry-weighed afteroven-desiccation at 60 ƒC for 72 hours An additional set of six tiles was used at the be-ginning of the experiment (t0) to determine initial periphyton biomass using the samemethod Periphyton did not develop on aquarium surfaces in either snail-present orcontrol aquaria
Macrophytes were ~ 15-cm long Ceratophyllum demersum L apical sprigs alsocollected from East Twin Lakersquos littoral zone where this plant was common C de-mersum was covered with a thin biofilm of loosely attached diatoms amorphous detri-tus and protozoans as revealed from a qualitative microscopic examination (100 acutendash 400 acute) The biofilm on C demersum was termed epiphyton to distinguish it from theperiphyton grown on glass tiles Periphyton composition was assumed to be similar tothe epiphyton collected from the same habitat Biomass change for plants was ex-pected to be less marked than for periphyton and maple leaves therefore C demersumbiomass was determined as living weight to allow for more precise t0 and tF determina-tions on the same sprigs Sprigs were partially dried on water-absorbing paper andweighed once they did not release water upon gentle shaking Weighing conditionswere uniform (rdry-wet = 0844 p lt 005 n = 6) Contribution of epiphyton to sprig bio-mass was considered negligible Each aquarium received ~ 5ndash7 g of living C demer-sum (one-way type I ANOVA F = 0656 p gt 065)
Air-dried maple (Acer sp) leaves were rewetted for 48 hours to leach tannins Let-ter-tagged three-leaf packs were assembled with a nylon string at the petioles and in-dividually dry-weighed after oven-desiccation at 60 ƒC for 72 hours Leaf packs were
Resource use and partitioning by two co-occurring snail species 233
conditioned with natural microflora in shallow indoor aquaria for three weeks beforet0 Post-conditioning leaf pack biomass at t0 was determined by adjusting each packrsquospre-conditioning biomass for the average post-conditioning biomass from six addi-tional packs that were dry-weighed at t0 The microflora on leaf packs was not ana-lyzed microscopically but was assumed to be a mixture of microfungi bacteria andto a lesser extent algae Snail individuals and feces were gently removed before finalbiomass determinations
Total phosphorus concentration (TP) in the aquaria was determined (ascorbic acidmethod Clesceri et al 1989) at t0 mid-term (t5 = day 5) and at the end of the experi-ment (t10 = tF) All inert material (e g aquaria glass tiles glassware) was analyticallycleaned in 10 HCl and rinsed with distilled water before use Initial TP in randomlychosen aquaria (after introduction of trophic resources and before introduction ofsnails) was 198 plusmn 49 mgl (average plusmn standard error n = 6) Mid-term and final TPwere determined with one 10-ml sample per aquarium Evaporation which remainedminimal did not affect TP determination
Water temperature was determined from randomly chosen aquaria on three datesand remained at 170 plusmn 05 ƒC (avg plusmn std error n = 3) Daylight diffused into the ex-periment locale Parallel aquarium orientation and equal distance from windows pro-vided uniform light The D L cycle followed the natural ~ 15 09 photoperiod (June)with midday irradiance at the water surface of 90 plusmn 13 mE mndash2 sndash1 (avg plusmn std error n= 6) a level expected to support algal and C demersum growth despite being close tolimiting conditions (e g Sand-Jensen amp Madsen 1991)
Snails were collected from an unnamed pond near West Branch Reservoir (PortageCounty) where macrophytes attached algae and allochthonous macrodetritus of plantorigin were abundant Physa spp included Physa integra Haldeman and Physa gy-rina Say which are difficult to distinguish in vivo but share similar ecological char-acteristics (Clampitt 1970) Though P gyrina can reach a larger size body size of thePhysa individuals used in the experiment was similar The ldquolow-densityrdquo conditionroughly represented a low to intermediate natural density of Physa in macropythe-richlittoral zones in northeast Ohio shallow lakes with potential snail predators (~ 09ndash13individualsg Lombardo 2001) Low-density Physa aquaria received 8 adult individu-als High density was set as 3 acute low density Because H trivolvis individual living bio-mass is three- to four-fold that of Physa (P Lombardo unpubl data from repeatedweight trials) each aquarium received four Physa individuals for every H trivolvis in-dividual to maintain similar snail biomass and hence potential grazing pressure in eachlow- or high-density snail-present aquarium
Snails were fed ad libitum with diverse plant material collected in situ and thenstarved for ~12 hrs before t0 to avoid trophic bias (Brendelberger 1994) Snails werethe last biological material introduced at t0 All individuals (Physa 4ndash8 mm shelllength H trivolvis 7ndash15 mm shell diameter) were placed in quadrat C providing sim-ilar initial access to trophic resources during evening hours Snails were not disturbedor replaced until tF Snail mortality at tF was low with a maximum of two dead indi-viduals recorded in two high-density Physa aquaria All H trivolvis individuals wererecovered alive at tF In one high-density H trivolvis aquarium an additional individ-ual (probably introduced accidentally as a juvenile on C demersum) was observedstarting from t7 This individual was included in distribution and overlap calculations
234 Paola Lombardo and G Dennis Cooke
starting from t7 Food consumption data from the aquarium with the additional individ-ual were similar to the other replicates Egg masses or newborns were not observed
Numbers of snail individuals on each trophic and spatial resource were recordeddaily at 08 00ndash10 00 and 18 00ndash21 00 hours Differences in snail distribution pat-terns on trophic and spatial resources were tested with a repeated-measures ANOVAfollowed by SNK (Student-Newman-Keuls) analysis at p pound 010 for significant outco-mes (Zar 1984) Day zero distributions (all individuals on quadrat C) were excludedfrom statistical analysis
Species overlap on trophic and spatial resources was calculated using the overlapindex a (Levins 1968)
aij =Sh (pi middot pj) a ji =
Sh (pj middot pi)
Sh (pi)2 Sh (pj)
2
where pi is the proportion of species i on the hth resource and pj is the proportion ofspecies j on the hth resource The aij value quantifies the overlap of species i over spe-cies j and is not necessarily symmetrical with the aji overlap of species j over speciesi depending on general resource use by each species [quantified by Sh (pi)
2] Asym-metrical indices are preferred because they better represent natural situations (Slo-bodchikoff amp Schulz 1980) The a values for the twice-daily observations did notfollow any discernible temporal pattern and were averaged over the 10-day periodAveraged a values expressed a general concept and are not to be intended as Mayrsquos(1972) ldquoaverage alphardquo
Distribution-based snail preference for resources was assessed using the modifiedversion of Roarsquos (1992) MANOVA procedure used and described in Lombardo ampCooke (2002) Such a method assesses preference or rejection of a resource relative tothe other resources ldquoValuerdquo of trophic resources determined with this method was assubstrate only
Food consumption by snails was measured as changes in resource biomass ingrazed vs ungrazed aquaria (controls) Biomass change was normalized as finalweight expressed as percent of initial weight Percent changes of less than 100 indi-cated biomass loss and percent changes higher than 100 indicated biomass gainBiomass of food items remained unchanged from initial values in snail absence (two-tailed t-tests for initial vs final biomass in control aquaria tleaf packs = 0123 tmacrophytes
= 0234 tperiphyton = 1645 n1 = n2 = 3 d f = 4 and n s for all three) and any changein snail-present aquaria was ascribed to snail presence Snail biomass changes were notquantified but low mortality and apparent new shell growth suggested non-limitingconditions for snails
Significant differences among treatments for food item biomass and TP were deter-mined with one-way type I ANOVAs SNK tests with p pound 010 were carried out forsignificant ANOVAs (Zar 1984) Linear correlations and t-tests were performed forselected data sets Small sample size (n = 3) precluded testing for normality and equa-lity of variances (e g Zar 1984) but data were neither additive nor scaled Thus pa-rametric analysis on untransformed data was performed also due to negligible testpower difference with non-parametric ANOVAs with equal sample size (Zar 1984)and negligible differences with non-parametric MANOVA outcomes (Roa 1992)
Resource use and partitioning by two co-occurring snail species 235
Results
Snail distribut ion and overlap on resources
Snails accumulated on trophic resources (though gradually on maple leafpacks) and were found on empty quadrats (spatial resources) in lower num-bers (Figs 3 and 4 Table 1) Snails were found in significantly higher num-bers on leaf packs regardless of density and species assemblage (Table 1)Preference for leaf packs was striking for both species (Fig 5) Physarsquos higherrelative presence on leaf packs at high snail density (Table 2) was accompa-nied by a significant reduction in H trivolvisrsquo average percent distribution(Table 3) and preference degree (Fig 5) from single- to two-species aquariarather than reflecting an increased preference by Physa (Table 3 Fig 5) Rela-tive distribution on leaves was independent of snail density in single-speciesaquaria but decreased with increasing density for coexisting snails (Table 4)
Physarsquos initial accumulation on periphyton decreased (with temporary tileabandonment at low density) shortly after an evident reduction to almostldquoclean tilerdquo conditions in periphyton biomass was observed at t3ndash t4 in allsnail-present aquaria (Fig 3) H trivolvis visits on periphyton were relativelyfrequent in single-species aquaria but rare when coexisting with Physa(Fig 4) especially at low density (Table 3) Snails rejected periphyton tiles athigh density but coexistence led to an increase in Physa preference (from neu-
Table 1 Differences in snail distribution (as number of individuals) on spatial (emptyquadrats) and trophic resources tested with repeated-measures ANOVAs followed bySNK multiple comparisons Critical values for F are one-tailed values for numeratord f = 8 and denominator d f = 152 Levels of significance are ns (p gt 010) (ppound 010) (p pound 005) and (p pound 001) Significantly different snail distributions onthe trophic and spatial resources resulting from SNK tests (p pound 010) are symbolizedwith different letters (a = lowest 10-day average snail density on resource progressi-vely higher densities listed alphabetically)
SNK (p pound010)
ANOVA trophic resources empty quadrats
leaf peri- macro-experimental condition F p packs phyton phytes A B C D E F
Physa low density 41046 c b b a ab ab a a abPhysa low density with H trivolvis 60252 c b a a a a a a aH trivolvis low density 16331 d a c ab ab a ab ab abH trivolvis low density with Physa 12221 b a a a a a a a a
Physa high density 121278 c ab b ab ab a a a aPhysa high density with H trivolvis 75880 c a b a a a a a aH trivolvis high density 31430 b a a a a a a a aH trivolvis high density with Physa 8239 c ab ab a a ab ab ab ab
236 Paola Lombardo and G Dennis Cooke
Fig 3 Distribution of Physa on spatial (food-devoid quadrats on aquarium surfaces)and trophic resources as average snail numbers on each trophic substrate and on allquadrats One morning (m) and one evening (e) observation per day on days 1 through10 Total numbers per aquarium given on x axis Number of live individuals on eachobservation normalized to initial numbers (when applicable) for graph comparability
Resource use and partitioning by two co-occurring snail species 237
Fig 4 Distribution of H trivolvis on spatial (food-devoid quadrats on aquarium surfa-ces) and trophic resources during the experiment Explanations as in Fig 3
238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 231
Fig 1 Experimental design Circles in each experimental condition symbolize totaland relative snail biomass number of snail individuals not drawn to scale Low-highdensity crossed treatments were not performed Number of adult snails in low-densitytrials were 8 and 4 for Physa and 2 and 1 for H trivolvis (single-species and two-spe-cies trials respectively) High densities were 3 acute low densities Each experimentalcondition was carried out in triplicate
Fig 2 Schematic representation of an experimental aquarium subdivided in six quad-rats (AndashF) with trophic resources (one C demersum sprig periphyton on a glass tileand decomposing maple leaves) superimposed on quadrats Layout was the same ineach aquarium Snails were added according to the experimental design in Fig 1
232 Paola Lombardo and G Dennis Cooke
undisturbed for 24 hours before the experimentrsquos beginning (t0) to allow chlorine to es-cape Tap water was chosen because of its high calcium content necessary for snailmetabolism (Brown 1991) and low total phosphorus content
Physical layout was the same in all aquaria Trophic resources were three types ofknown snail food periphyton grown on glass tiles living macrophytes and coarse ter-restrial leaf litter Spatial resources were six quadrats of roughly similar size markedby lines on aquaria outer surfaces and labelled A through F (Fig 2) Wall surface waslower for central quadrats (C and D) but snail utilization of walls remained minimaland was not quantified Available surface area for periphyton was less than for macro-phytes and leaf litter but total edible quantity was similar for each food type withcoarser or tougher material assumed inedible for snails (e g Thomas et al 1985) Allperiphytic material was considered edible
Methods for trophic resource preparation and biomass determination followedthose in Lombardo amp Cooke (2002) Periphyton was a thick layer of algae pro-tozoans and unrecognizable detritus developed on rough-surface (ldquopattern 62rdquo)102 mm acute 102 mm glass tiles during a 60-day colonization period with inocula fromthe vegetated littoral zone of East Twin Lake Portage County Northeast Ohio (USA)East Twin Lake is a mesotrophic (TP ~ 20 mgl) clear-water natural lake with abundantaquatic vegetation and a rich diverse snail assemblage throughout its littoral zone
Periphyton was manually scraped with a plastic (non-absorbant) brush after re-trieving the tiles at the end of the experiment (tF) Some periphytic material was una-voidably lost during retrieval because of the very loose attachment to the tiles andsloughing of older algal patches leading to high variability in final biomass for controltiles however statistical power for grazed vs ungrazed comparisons remained intactThe scraped material was collected on 45-mm fiberglass filters and dry-weighed afteroven-desiccation at 60 ƒC for 72 hours An additional set of six tiles was used at the be-ginning of the experiment (t0) to determine initial periphyton biomass using the samemethod Periphyton did not develop on aquarium surfaces in either snail-present orcontrol aquaria
Macrophytes were ~ 15-cm long Ceratophyllum demersum L apical sprigs alsocollected from East Twin Lakersquos littoral zone where this plant was common C de-mersum was covered with a thin biofilm of loosely attached diatoms amorphous detri-tus and protozoans as revealed from a qualitative microscopic examination (100 acutendash 400 acute) The biofilm on C demersum was termed epiphyton to distinguish it from theperiphyton grown on glass tiles Periphyton composition was assumed to be similar tothe epiphyton collected from the same habitat Biomass change for plants was ex-pected to be less marked than for periphyton and maple leaves therefore C demersumbiomass was determined as living weight to allow for more precise t0 and tF determina-tions on the same sprigs Sprigs were partially dried on water-absorbing paper andweighed once they did not release water upon gentle shaking Weighing conditionswere uniform (rdry-wet = 0844 p lt 005 n = 6) Contribution of epiphyton to sprig bio-mass was considered negligible Each aquarium received ~ 5ndash7 g of living C demer-sum (one-way type I ANOVA F = 0656 p gt 065)
Air-dried maple (Acer sp) leaves were rewetted for 48 hours to leach tannins Let-ter-tagged three-leaf packs were assembled with a nylon string at the petioles and in-dividually dry-weighed after oven-desiccation at 60 ƒC for 72 hours Leaf packs were
Resource use and partitioning by two co-occurring snail species 233
conditioned with natural microflora in shallow indoor aquaria for three weeks beforet0 Post-conditioning leaf pack biomass at t0 was determined by adjusting each packrsquospre-conditioning biomass for the average post-conditioning biomass from six addi-tional packs that were dry-weighed at t0 The microflora on leaf packs was not ana-lyzed microscopically but was assumed to be a mixture of microfungi bacteria andto a lesser extent algae Snail individuals and feces were gently removed before finalbiomass determinations
Total phosphorus concentration (TP) in the aquaria was determined (ascorbic acidmethod Clesceri et al 1989) at t0 mid-term (t5 = day 5) and at the end of the experi-ment (t10 = tF) All inert material (e g aquaria glass tiles glassware) was analyticallycleaned in 10 HCl and rinsed with distilled water before use Initial TP in randomlychosen aquaria (after introduction of trophic resources and before introduction ofsnails) was 198 plusmn 49 mgl (average plusmn standard error n = 6) Mid-term and final TPwere determined with one 10-ml sample per aquarium Evaporation which remainedminimal did not affect TP determination
Water temperature was determined from randomly chosen aquaria on three datesand remained at 170 plusmn 05 ƒC (avg plusmn std error n = 3) Daylight diffused into the ex-periment locale Parallel aquarium orientation and equal distance from windows pro-vided uniform light The D L cycle followed the natural ~ 15 09 photoperiod (June)with midday irradiance at the water surface of 90 plusmn 13 mE mndash2 sndash1 (avg plusmn std error n= 6) a level expected to support algal and C demersum growth despite being close tolimiting conditions (e g Sand-Jensen amp Madsen 1991)
Snails were collected from an unnamed pond near West Branch Reservoir (PortageCounty) where macrophytes attached algae and allochthonous macrodetritus of plantorigin were abundant Physa spp included Physa integra Haldeman and Physa gy-rina Say which are difficult to distinguish in vivo but share similar ecological char-acteristics (Clampitt 1970) Though P gyrina can reach a larger size body size of thePhysa individuals used in the experiment was similar The ldquolow-densityrdquo conditionroughly represented a low to intermediate natural density of Physa in macropythe-richlittoral zones in northeast Ohio shallow lakes with potential snail predators (~ 09ndash13individualsg Lombardo 2001) Low-density Physa aquaria received 8 adult individu-als High density was set as 3 acute low density Because H trivolvis individual living bio-mass is three- to four-fold that of Physa (P Lombardo unpubl data from repeatedweight trials) each aquarium received four Physa individuals for every H trivolvis in-dividual to maintain similar snail biomass and hence potential grazing pressure in eachlow- or high-density snail-present aquarium
Snails were fed ad libitum with diverse plant material collected in situ and thenstarved for ~12 hrs before t0 to avoid trophic bias (Brendelberger 1994) Snails werethe last biological material introduced at t0 All individuals (Physa 4ndash8 mm shelllength H trivolvis 7ndash15 mm shell diameter) were placed in quadrat C providing sim-ilar initial access to trophic resources during evening hours Snails were not disturbedor replaced until tF Snail mortality at tF was low with a maximum of two dead indi-viduals recorded in two high-density Physa aquaria All H trivolvis individuals wererecovered alive at tF In one high-density H trivolvis aquarium an additional individ-ual (probably introduced accidentally as a juvenile on C demersum) was observedstarting from t7 This individual was included in distribution and overlap calculations
234 Paola Lombardo and G Dennis Cooke
starting from t7 Food consumption data from the aquarium with the additional individ-ual were similar to the other replicates Egg masses or newborns were not observed
Numbers of snail individuals on each trophic and spatial resource were recordeddaily at 08 00ndash10 00 and 18 00ndash21 00 hours Differences in snail distribution pat-terns on trophic and spatial resources were tested with a repeated-measures ANOVAfollowed by SNK (Student-Newman-Keuls) analysis at p pound 010 for significant outco-mes (Zar 1984) Day zero distributions (all individuals on quadrat C) were excludedfrom statistical analysis
Species overlap on trophic and spatial resources was calculated using the overlapindex a (Levins 1968)
aij =Sh (pi middot pj) a ji =
Sh (pj middot pi)
Sh (pi)2 Sh (pj)
2
where pi is the proportion of species i on the hth resource and pj is the proportion ofspecies j on the hth resource The aij value quantifies the overlap of species i over spe-cies j and is not necessarily symmetrical with the aji overlap of species j over speciesi depending on general resource use by each species [quantified by Sh (pi)
2] Asym-metrical indices are preferred because they better represent natural situations (Slo-bodchikoff amp Schulz 1980) The a values for the twice-daily observations did notfollow any discernible temporal pattern and were averaged over the 10-day periodAveraged a values expressed a general concept and are not to be intended as Mayrsquos(1972) ldquoaverage alphardquo
Distribution-based snail preference for resources was assessed using the modifiedversion of Roarsquos (1992) MANOVA procedure used and described in Lombardo ampCooke (2002) Such a method assesses preference or rejection of a resource relative tothe other resources ldquoValuerdquo of trophic resources determined with this method was assubstrate only
Food consumption by snails was measured as changes in resource biomass ingrazed vs ungrazed aquaria (controls) Biomass change was normalized as finalweight expressed as percent of initial weight Percent changes of less than 100 indi-cated biomass loss and percent changes higher than 100 indicated biomass gainBiomass of food items remained unchanged from initial values in snail absence (two-tailed t-tests for initial vs final biomass in control aquaria tleaf packs = 0123 tmacrophytes
= 0234 tperiphyton = 1645 n1 = n2 = 3 d f = 4 and n s for all three) and any changein snail-present aquaria was ascribed to snail presence Snail biomass changes were notquantified but low mortality and apparent new shell growth suggested non-limitingconditions for snails
Significant differences among treatments for food item biomass and TP were deter-mined with one-way type I ANOVAs SNK tests with p pound 010 were carried out forsignificant ANOVAs (Zar 1984) Linear correlations and t-tests were performed forselected data sets Small sample size (n = 3) precluded testing for normality and equa-lity of variances (e g Zar 1984) but data were neither additive nor scaled Thus pa-rametric analysis on untransformed data was performed also due to negligible testpower difference with non-parametric ANOVAs with equal sample size (Zar 1984)and negligible differences with non-parametric MANOVA outcomes (Roa 1992)
Resource use and partitioning by two co-occurring snail species 235
Results
Snail distribut ion and overlap on resources
Snails accumulated on trophic resources (though gradually on maple leafpacks) and were found on empty quadrats (spatial resources) in lower num-bers (Figs 3 and 4 Table 1) Snails were found in significantly higher num-bers on leaf packs regardless of density and species assemblage (Table 1)Preference for leaf packs was striking for both species (Fig 5) Physarsquos higherrelative presence on leaf packs at high snail density (Table 2) was accompa-nied by a significant reduction in H trivolvisrsquo average percent distribution(Table 3) and preference degree (Fig 5) from single- to two-species aquariarather than reflecting an increased preference by Physa (Table 3 Fig 5) Rela-tive distribution on leaves was independent of snail density in single-speciesaquaria but decreased with increasing density for coexisting snails (Table 4)
Physarsquos initial accumulation on periphyton decreased (with temporary tileabandonment at low density) shortly after an evident reduction to almostldquoclean tilerdquo conditions in periphyton biomass was observed at t3ndash t4 in allsnail-present aquaria (Fig 3) H trivolvis visits on periphyton were relativelyfrequent in single-species aquaria but rare when coexisting with Physa(Fig 4) especially at low density (Table 3) Snails rejected periphyton tiles athigh density but coexistence led to an increase in Physa preference (from neu-
Table 1 Differences in snail distribution (as number of individuals) on spatial (emptyquadrats) and trophic resources tested with repeated-measures ANOVAs followed bySNK multiple comparisons Critical values for F are one-tailed values for numeratord f = 8 and denominator d f = 152 Levels of significance are ns (p gt 010) (ppound 010) (p pound 005) and (p pound 001) Significantly different snail distributions onthe trophic and spatial resources resulting from SNK tests (p pound 010) are symbolizedwith different letters (a = lowest 10-day average snail density on resource progressi-vely higher densities listed alphabetically)
SNK (p pound010)
ANOVA trophic resources empty quadrats
leaf peri- macro-experimental condition F p packs phyton phytes A B C D E F
Physa low density 41046 c b b a ab ab a a abPhysa low density with H trivolvis 60252 c b a a a a a a aH trivolvis low density 16331 d a c ab ab a ab ab abH trivolvis low density with Physa 12221 b a a a a a a a a
Physa high density 121278 c ab b ab ab a a a aPhysa high density with H trivolvis 75880 c a b a a a a a aH trivolvis high density 31430 b a a a a a a a aH trivolvis high density with Physa 8239 c ab ab a a ab ab ab ab
236 Paola Lombardo and G Dennis Cooke
Fig 3 Distribution of Physa on spatial (food-devoid quadrats on aquarium surfaces)and trophic resources as average snail numbers on each trophic substrate and on allquadrats One morning (m) and one evening (e) observation per day on days 1 through10 Total numbers per aquarium given on x axis Number of live individuals on eachobservation normalized to initial numbers (when applicable) for graph comparability
Resource use and partitioning by two co-occurring snail species 237
Fig 4 Distribution of H trivolvis on spatial (food-devoid quadrats on aquarium surfa-ces) and trophic resources during the experiment Explanations as in Fig 3
238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

232 Paola Lombardo and G Dennis Cooke
undisturbed for 24 hours before the experimentrsquos beginning (t0) to allow chlorine to es-cape Tap water was chosen because of its high calcium content necessary for snailmetabolism (Brown 1991) and low total phosphorus content
Physical layout was the same in all aquaria Trophic resources were three types ofknown snail food periphyton grown on glass tiles living macrophytes and coarse ter-restrial leaf litter Spatial resources were six quadrats of roughly similar size markedby lines on aquaria outer surfaces and labelled A through F (Fig 2) Wall surface waslower for central quadrats (C and D) but snail utilization of walls remained minimaland was not quantified Available surface area for periphyton was less than for macro-phytes and leaf litter but total edible quantity was similar for each food type withcoarser or tougher material assumed inedible for snails (e g Thomas et al 1985) Allperiphytic material was considered edible
Methods for trophic resource preparation and biomass determination followedthose in Lombardo amp Cooke (2002) Periphyton was a thick layer of algae pro-tozoans and unrecognizable detritus developed on rough-surface (ldquopattern 62rdquo)102 mm acute 102 mm glass tiles during a 60-day colonization period with inocula fromthe vegetated littoral zone of East Twin Lake Portage County Northeast Ohio (USA)East Twin Lake is a mesotrophic (TP ~ 20 mgl) clear-water natural lake with abundantaquatic vegetation and a rich diverse snail assemblage throughout its littoral zone
Periphyton was manually scraped with a plastic (non-absorbant) brush after re-trieving the tiles at the end of the experiment (tF) Some periphytic material was una-voidably lost during retrieval because of the very loose attachment to the tiles andsloughing of older algal patches leading to high variability in final biomass for controltiles however statistical power for grazed vs ungrazed comparisons remained intactThe scraped material was collected on 45-mm fiberglass filters and dry-weighed afteroven-desiccation at 60 ƒC for 72 hours An additional set of six tiles was used at the be-ginning of the experiment (t0) to determine initial periphyton biomass using the samemethod Periphyton did not develop on aquarium surfaces in either snail-present orcontrol aquaria
Macrophytes were ~ 15-cm long Ceratophyllum demersum L apical sprigs alsocollected from East Twin Lakersquos littoral zone where this plant was common C de-mersum was covered with a thin biofilm of loosely attached diatoms amorphous detri-tus and protozoans as revealed from a qualitative microscopic examination (100 acutendash 400 acute) The biofilm on C demersum was termed epiphyton to distinguish it from theperiphyton grown on glass tiles Periphyton composition was assumed to be similar tothe epiphyton collected from the same habitat Biomass change for plants was ex-pected to be less marked than for periphyton and maple leaves therefore C demersumbiomass was determined as living weight to allow for more precise t0 and tF determina-tions on the same sprigs Sprigs were partially dried on water-absorbing paper andweighed once they did not release water upon gentle shaking Weighing conditionswere uniform (rdry-wet = 0844 p lt 005 n = 6) Contribution of epiphyton to sprig bio-mass was considered negligible Each aquarium received ~ 5ndash7 g of living C demer-sum (one-way type I ANOVA F = 0656 p gt 065)
Air-dried maple (Acer sp) leaves were rewetted for 48 hours to leach tannins Let-ter-tagged three-leaf packs were assembled with a nylon string at the petioles and in-dividually dry-weighed after oven-desiccation at 60 ƒC for 72 hours Leaf packs were
Resource use and partitioning by two co-occurring snail species 233
conditioned with natural microflora in shallow indoor aquaria for three weeks beforet0 Post-conditioning leaf pack biomass at t0 was determined by adjusting each packrsquospre-conditioning biomass for the average post-conditioning biomass from six addi-tional packs that were dry-weighed at t0 The microflora on leaf packs was not ana-lyzed microscopically but was assumed to be a mixture of microfungi bacteria andto a lesser extent algae Snail individuals and feces were gently removed before finalbiomass determinations
Total phosphorus concentration (TP) in the aquaria was determined (ascorbic acidmethod Clesceri et al 1989) at t0 mid-term (t5 = day 5) and at the end of the experi-ment (t10 = tF) All inert material (e g aquaria glass tiles glassware) was analyticallycleaned in 10 HCl and rinsed with distilled water before use Initial TP in randomlychosen aquaria (after introduction of trophic resources and before introduction ofsnails) was 198 plusmn 49 mgl (average plusmn standard error n = 6) Mid-term and final TPwere determined with one 10-ml sample per aquarium Evaporation which remainedminimal did not affect TP determination
Water temperature was determined from randomly chosen aquaria on three datesand remained at 170 plusmn 05 ƒC (avg plusmn std error n = 3) Daylight diffused into the ex-periment locale Parallel aquarium orientation and equal distance from windows pro-vided uniform light The D L cycle followed the natural ~ 15 09 photoperiod (June)with midday irradiance at the water surface of 90 plusmn 13 mE mndash2 sndash1 (avg plusmn std error n= 6) a level expected to support algal and C demersum growth despite being close tolimiting conditions (e g Sand-Jensen amp Madsen 1991)
Snails were collected from an unnamed pond near West Branch Reservoir (PortageCounty) where macrophytes attached algae and allochthonous macrodetritus of plantorigin were abundant Physa spp included Physa integra Haldeman and Physa gy-rina Say which are difficult to distinguish in vivo but share similar ecological char-acteristics (Clampitt 1970) Though P gyrina can reach a larger size body size of thePhysa individuals used in the experiment was similar The ldquolow-densityrdquo conditionroughly represented a low to intermediate natural density of Physa in macropythe-richlittoral zones in northeast Ohio shallow lakes with potential snail predators (~ 09ndash13individualsg Lombardo 2001) Low-density Physa aquaria received 8 adult individu-als High density was set as 3 acute low density Because H trivolvis individual living bio-mass is three- to four-fold that of Physa (P Lombardo unpubl data from repeatedweight trials) each aquarium received four Physa individuals for every H trivolvis in-dividual to maintain similar snail biomass and hence potential grazing pressure in eachlow- or high-density snail-present aquarium
Snails were fed ad libitum with diverse plant material collected in situ and thenstarved for ~12 hrs before t0 to avoid trophic bias (Brendelberger 1994) Snails werethe last biological material introduced at t0 All individuals (Physa 4ndash8 mm shelllength H trivolvis 7ndash15 mm shell diameter) were placed in quadrat C providing sim-ilar initial access to trophic resources during evening hours Snails were not disturbedor replaced until tF Snail mortality at tF was low with a maximum of two dead indi-viduals recorded in two high-density Physa aquaria All H trivolvis individuals wererecovered alive at tF In one high-density H trivolvis aquarium an additional individ-ual (probably introduced accidentally as a juvenile on C demersum) was observedstarting from t7 This individual was included in distribution and overlap calculations
234 Paola Lombardo and G Dennis Cooke
starting from t7 Food consumption data from the aquarium with the additional individ-ual were similar to the other replicates Egg masses or newborns were not observed
Numbers of snail individuals on each trophic and spatial resource were recordeddaily at 08 00ndash10 00 and 18 00ndash21 00 hours Differences in snail distribution pat-terns on trophic and spatial resources were tested with a repeated-measures ANOVAfollowed by SNK (Student-Newman-Keuls) analysis at p pound 010 for significant outco-mes (Zar 1984) Day zero distributions (all individuals on quadrat C) were excludedfrom statistical analysis
Species overlap on trophic and spatial resources was calculated using the overlapindex a (Levins 1968)
aij =Sh (pi middot pj) a ji =
Sh (pj middot pi)
Sh (pi)2 Sh (pj)
2
where pi is the proportion of species i on the hth resource and pj is the proportion ofspecies j on the hth resource The aij value quantifies the overlap of species i over spe-cies j and is not necessarily symmetrical with the aji overlap of species j over speciesi depending on general resource use by each species [quantified by Sh (pi)
2] Asym-metrical indices are preferred because they better represent natural situations (Slo-bodchikoff amp Schulz 1980) The a values for the twice-daily observations did notfollow any discernible temporal pattern and were averaged over the 10-day periodAveraged a values expressed a general concept and are not to be intended as Mayrsquos(1972) ldquoaverage alphardquo
Distribution-based snail preference for resources was assessed using the modifiedversion of Roarsquos (1992) MANOVA procedure used and described in Lombardo ampCooke (2002) Such a method assesses preference or rejection of a resource relative tothe other resources ldquoValuerdquo of trophic resources determined with this method was assubstrate only
Food consumption by snails was measured as changes in resource biomass ingrazed vs ungrazed aquaria (controls) Biomass change was normalized as finalweight expressed as percent of initial weight Percent changes of less than 100 indi-cated biomass loss and percent changes higher than 100 indicated biomass gainBiomass of food items remained unchanged from initial values in snail absence (two-tailed t-tests for initial vs final biomass in control aquaria tleaf packs = 0123 tmacrophytes
= 0234 tperiphyton = 1645 n1 = n2 = 3 d f = 4 and n s for all three) and any changein snail-present aquaria was ascribed to snail presence Snail biomass changes were notquantified but low mortality and apparent new shell growth suggested non-limitingconditions for snails
Significant differences among treatments for food item biomass and TP were deter-mined with one-way type I ANOVAs SNK tests with p pound 010 were carried out forsignificant ANOVAs (Zar 1984) Linear correlations and t-tests were performed forselected data sets Small sample size (n = 3) precluded testing for normality and equa-lity of variances (e g Zar 1984) but data were neither additive nor scaled Thus pa-rametric analysis on untransformed data was performed also due to negligible testpower difference with non-parametric ANOVAs with equal sample size (Zar 1984)and negligible differences with non-parametric MANOVA outcomes (Roa 1992)
Resource use and partitioning by two co-occurring snail species 235
Results
Snail distribut ion and overlap on resources
Snails accumulated on trophic resources (though gradually on maple leafpacks) and were found on empty quadrats (spatial resources) in lower num-bers (Figs 3 and 4 Table 1) Snails were found in significantly higher num-bers on leaf packs regardless of density and species assemblage (Table 1)Preference for leaf packs was striking for both species (Fig 5) Physarsquos higherrelative presence on leaf packs at high snail density (Table 2) was accompa-nied by a significant reduction in H trivolvisrsquo average percent distribution(Table 3) and preference degree (Fig 5) from single- to two-species aquariarather than reflecting an increased preference by Physa (Table 3 Fig 5) Rela-tive distribution on leaves was independent of snail density in single-speciesaquaria but decreased with increasing density for coexisting snails (Table 4)
Physarsquos initial accumulation on periphyton decreased (with temporary tileabandonment at low density) shortly after an evident reduction to almostldquoclean tilerdquo conditions in periphyton biomass was observed at t3ndash t4 in allsnail-present aquaria (Fig 3) H trivolvis visits on periphyton were relativelyfrequent in single-species aquaria but rare when coexisting with Physa(Fig 4) especially at low density (Table 3) Snails rejected periphyton tiles athigh density but coexistence led to an increase in Physa preference (from neu-
Table 1 Differences in snail distribution (as number of individuals) on spatial (emptyquadrats) and trophic resources tested with repeated-measures ANOVAs followed bySNK multiple comparisons Critical values for F are one-tailed values for numeratord f = 8 and denominator d f = 152 Levels of significance are ns (p gt 010) (ppound 010) (p pound 005) and (p pound 001) Significantly different snail distributions onthe trophic and spatial resources resulting from SNK tests (p pound 010) are symbolizedwith different letters (a = lowest 10-day average snail density on resource progressi-vely higher densities listed alphabetically)
SNK (p pound010)
ANOVA trophic resources empty quadrats
leaf peri- macro-experimental condition F p packs phyton phytes A B C D E F
Physa low density 41046 c b b a ab ab a a abPhysa low density with H trivolvis 60252 c b a a a a a a aH trivolvis low density 16331 d a c ab ab a ab ab abH trivolvis low density with Physa 12221 b a a a a a a a a
Physa high density 121278 c ab b ab ab a a a aPhysa high density with H trivolvis 75880 c a b a a a a a aH trivolvis high density 31430 b a a a a a a a aH trivolvis high density with Physa 8239 c ab ab a a ab ab ab ab
236 Paola Lombardo and G Dennis Cooke
Fig 3 Distribution of Physa on spatial (food-devoid quadrats on aquarium surfaces)and trophic resources as average snail numbers on each trophic substrate and on allquadrats One morning (m) and one evening (e) observation per day on days 1 through10 Total numbers per aquarium given on x axis Number of live individuals on eachobservation normalized to initial numbers (when applicable) for graph comparability
Resource use and partitioning by two co-occurring snail species 237
Fig 4 Distribution of H trivolvis on spatial (food-devoid quadrats on aquarium surfa-ces) and trophic resources during the experiment Explanations as in Fig 3
238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 233
conditioned with natural microflora in shallow indoor aquaria for three weeks beforet0 Post-conditioning leaf pack biomass at t0 was determined by adjusting each packrsquospre-conditioning biomass for the average post-conditioning biomass from six addi-tional packs that were dry-weighed at t0 The microflora on leaf packs was not ana-lyzed microscopically but was assumed to be a mixture of microfungi bacteria andto a lesser extent algae Snail individuals and feces were gently removed before finalbiomass determinations
Total phosphorus concentration (TP) in the aquaria was determined (ascorbic acidmethod Clesceri et al 1989) at t0 mid-term (t5 = day 5) and at the end of the experi-ment (t10 = tF) All inert material (e g aquaria glass tiles glassware) was analyticallycleaned in 10 HCl and rinsed with distilled water before use Initial TP in randomlychosen aquaria (after introduction of trophic resources and before introduction ofsnails) was 198 plusmn 49 mgl (average plusmn standard error n = 6) Mid-term and final TPwere determined with one 10-ml sample per aquarium Evaporation which remainedminimal did not affect TP determination
Water temperature was determined from randomly chosen aquaria on three datesand remained at 170 plusmn 05 ƒC (avg plusmn std error n = 3) Daylight diffused into the ex-periment locale Parallel aquarium orientation and equal distance from windows pro-vided uniform light The D L cycle followed the natural ~ 15 09 photoperiod (June)with midday irradiance at the water surface of 90 plusmn 13 mE mndash2 sndash1 (avg plusmn std error n= 6) a level expected to support algal and C demersum growth despite being close tolimiting conditions (e g Sand-Jensen amp Madsen 1991)
Snails were collected from an unnamed pond near West Branch Reservoir (PortageCounty) where macrophytes attached algae and allochthonous macrodetritus of plantorigin were abundant Physa spp included Physa integra Haldeman and Physa gy-rina Say which are difficult to distinguish in vivo but share similar ecological char-acteristics (Clampitt 1970) Though P gyrina can reach a larger size body size of thePhysa individuals used in the experiment was similar The ldquolow-densityrdquo conditionroughly represented a low to intermediate natural density of Physa in macropythe-richlittoral zones in northeast Ohio shallow lakes with potential snail predators (~ 09ndash13individualsg Lombardo 2001) Low-density Physa aquaria received 8 adult individu-als High density was set as 3 acute low density Because H trivolvis individual living bio-mass is three- to four-fold that of Physa (P Lombardo unpubl data from repeatedweight trials) each aquarium received four Physa individuals for every H trivolvis in-dividual to maintain similar snail biomass and hence potential grazing pressure in eachlow- or high-density snail-present aquarium
Snails were fed ad libitum with diverse plant material collected in situ and thenstarved for ~12 hrs before t0 to avoid trophic bias (Brendelberger 1994) Snails werethe last biological material introduced at t0 All individuals (Physa 4ndash8 mm shelllength H trivolvis 7ndash15 mm shell diameter) were placed in quadrat C providing sim-ilar initial access to trophic resources during evening hours Snails were not disturbedor replaced until tF Snail mortality at tF was low with a maximum of two dead indi-viduals recorded in two high-density Physa aquaria All H trivolvis individuals wererecovered alive at tF In one high-density H trivolvis aquarium an additional individ-ual (probably introduced accidentally as a juvenile on C demersum) was observedstarting from t7 This individual was included in distribution and overlap calculations
234 Paola Lombardo and G Dennis Cooke
starting from t7 Food consumption data from the aquarium with the additional individ-ual were similar to the other replicates Egg masses or newborns were not observed
Numbers of snail individuals on each trophic and spatial resource were recordeddaily at 08 00ndash10 00 and 18 00ndash21 00 hours Differences in snail distribution pat-terns on trophic and spatial resources were tested with a repeated-measures ANOVAfollowed by SNK (Student-Newman-Keuls) analysis at p pound 010 for significant outco-mes (Zar 1984) Day zero distributions (all individuals on quadrat C) were excludedfrom statistical analysis
Species overlap on trophic and spatial resources was calculated using the overlapindex a (Levins 1968)
aij =Sh (pi middot pj) a ji =
Sh (pj middot pi)
Sh (pi)2 Sh (pj)
2
where pi is the proportion of species i on the hth resource and pj is the proportion ofspecies j on the hth resource The aij value quantifies the overlap of species i over spe-cies j and is not necessarily symmetrical with the aji overlap of species j over speciesi depending on general resource use by each species [quantified by Sh (pi)
2] Asym-metrical indices are preferred because they better represent natural situations (Slo-bodchikoff amp Schulz 1980) The a values for the twice-daily observations did notfollow any discernible temporal pattern and were averaged over the 10-day periodAveraged a values expressed a general concept and are not to be intended as Mayrsquos(1972) ldquoaverage alphardquo
Distribution-based snail preference for resources was assessed using the modifiedversion of Roarsquos (1992) MANOVA procedure used and described in Lombardo ampCooke (2002) Such a method assesses preference or rejection of a resource relative tothe other resources ldquoValuerdquo of trophic resources determined with this method was assubstrate only
Food consumption by snails was measured as changes in resource biomass ingrazed vs ungrazed aquaria (controls) Biomass change was normalized as finalweight expressed as percent of initial weight Percent changes of less than 100 indi-cated biomass loss and percent changes higher than 100 indicated biomass gainBiomass of food items remained unchanged from initial values in snail absence (two-tailed t-tests for initial vs final biomass in control aquaria tleaf packs = 0123 tmacrophytes
= 0234 tperiphyton = 1645 n1 = n2 = 3 d f = 4 and n s for all three) and any changein snail-present aquaria was ascribed to snail presence Snail biomass changes were notquantified but low mortality and apparent new shell growth suggested non-limitingconditions for snails
Significant differences among treatments for food item biomass and TP were deter-mined with one-way type I ANOVAs SNK tests with p pound 010 were carried out forsignificant ANOVAs (Zar 1984) Linear correlations and t-tests were performed forselected data sets Small sample size (n = 3) precluded testing for normality and equa-lity of variances (e g Zar 1984) but data were neither additive nor scaled Thus pa-rametric analysis on untransformed data was performed also due to negligible testpower difference with non-parametric ANOVAs with equal sample size (Zar 1984)and negligible differences with non-parametric MANOVA outcomes (Roa 1992)
Resource use and partitioning by two co-occurring snail species 235
Results
Snail distribut ion and overlap on resources
Snails accumulated on trophic resources (though gradually on maple leafpacks) and were found on empty quadrats (spatial resources) in lower num-bers (Figs 3 and 4 Table 1) Snails were found in significantly higher num-bers on leaf packs regardless of density and species assemblage (Table 1)Preference for leaf packs was striking for both species (Fig 5) Physarsquos higherrelative presence on leaf packs at high snail density (Table 2) was accompa-nied by a significant reduction in H trivolvisrsquo average percent distribution(Table 3) and preference degree (Fig 5) from single- to two-species aquariarather than reflecting an increased preference by Physa (Table 3 Fig 5) Rela-tive distribution on leaves was independent of snail density in single-speciesaquaria but decreased with increasing density for coexisting snails (Table 4)
Physarsquos initial accumulation on periphyton decreased (with temporary tileabandonment at low density) shortly after an evident reduction to almostldquoclean tilerdquo conditions in periphyton biomass was observed at t3ndash t4 in allsnail-present aquaria (Fig 3) H trivolvis visits on periphyton were relativelyfrequent in single-species aquaria but rare when coexisting with Physa(Fig 4) especially at low density (Table 3) Snails rejected periphyton tiles athigh density but coexistence led to an increase in Physa preference (from neu-
Table 1 Differences in snail distribution (as number of individuals) on spatial (emptyquadrats) and trophic resources tested with repeated-measures ANOVAs followed bySNK multiple comparisons Critical values for F are one-tailed values for numeratord f = 8 and denominator d f = 152 Levels of significance are ns (p gt 010) (ppound 010) (p pound 005) and (p pound 001) Significantly different snail distributions onthe trophic and spatial resources resulting from SNK tests (p pound 010) are symbolizedwith different letters (a = lowest 10-day average snail density on resource progressi-vely higher densities listed alphabetically)
SNK (p pound010)
ANOVA trophic resources empty quadrats
leaf peri- macro-experimental condition F p packs phyton phytes A B C D E F
Physa low density 41046 c b b a ab ab a a abPhysa low density with H trivolvis 60252 c b a a a a a a aH trivolvis low density 16331 d a c ab ab a ab ab abH trivolvis low density with Physa 12221 b a a a a a a a a
Physa high density 121278 c ab b ab ab a a a aPhysa high density with H trivolvis 75880 c a b a a a a a aH trivolvis high density 31430 b a a a a a a a aH trivolvis high density with Physa 8239 c ab ab a a ab ab ab ab
236 Paola Lombardo and G Dennis Cooke
Fig 3 Distribution of Physa on spatial (food-devoid quadrats on aquarium surfaces)and trophic resources as average snail numbers on each trophic substrate and on allquadrats One morning (m) and one evening (e) observation per day on days 1 through10 Total numbers per aquarium given on x axis Number of live individuals on eachobservation normalized to initial numbers (when applicable) for graph comparability
Resource use and partitioning by two co-occurring snail species 237
Fig 4 Distribution of H trivolvis on spatial (food-devoid quadrats on aquarium surfa-ces) and trophic resources during the experiment Explanations as in Fig 3
238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

234 Paola Lombardo and G Dennis Cooke
starting from t7 Food consumption data from the aquarium with the additional individ-ual were similar to the other replicates Egg masses or newborns were not observed
Numbers of snail individuals on each trophic and spatial resource were recordeddaily at 08 00ndash10 00 and 18 00ndash21 00 hours Differences in snail distribution pat-terns on trophic and spatial resources were tested with a repeated-measures ANOVAfollowed by SNK (Student-Newman-Keuls) analysis at p pound 010 for significant outco-mes (Zar 1984) Day zero distributions (all individuals on quadrat C) were excludedfrom statistical analysis
Species overlap on trophic and spatial resources was calculated using the overlapindex a (Levins 1968)
aij =Sh (pi middot pj) a ji =
Sh (pj middot pi)
Sh (pi)2 Sh (pj)
2
where pi is the proportion of species i on the hth resource and pj is the proportion ofspecies j on the hth resource The aij value quantifies the overlap of species i over spe-cies j and is not necessarily symmetrical with the aji overlap of species j over speciesi depending on general resource use by each species [quantified by Sh (pi)
2] Asym-metrical indices are preferred because they better represent natural situations (Slo-bodchikoff amp Schulz 1980) The a values for the twice-daily observations did notfollow any discernible temporal pattern and were averaged over the 10-day periodAveraged a values expressed a general concept and are not to be intended as Mayrsquos(1972) ldquoaverage alphardquo
Distribution-based snail preference for resources was assessed using the modifiedversion of Roarsquos (1992) MANOVA procedure used and described in Lombardo ampCooke (2002) Such a method assesses preference or rejection of a resource relative tothe other resources ldquoValuerdquo of trophic resources determined with this method was assubstrate only
Food consumption by snails was measured as changes in resource biomass ingrazed vs ungrazed aquaria (controls) Biomass change was normalized as finalweight expressed as percent of initial weight Percent changes of less than 100 indi-cated biomass loss and percent changes higher than 100 indicated biomass gainBiomass of food items remained unchanged from initial values in snail absence (two-tailed t-tests for initial vs final biomass in control aquaria tleaf packs = 0123 tmacrophytes
= 0234 tperiphyton = 1645 n1 = n2 = 3 d f = 4 and n s for all three) and any changein snail-present aquaria was ascribed to snail presence Snail biomass changes were notquantified but low mortality and apparent new shell growth suggested non-limitingconditions for snails
Significant differences among treatments for food item biomass and TP were deter-mined with one-way type I ANOVAs SNK tests with p pound 010 were carried out forsignificant ANOVAs (Zar 1984) Linear correlations and t-tests were performed forselected data sets Small sample size (n = 3) precluded testing for normality and equa-lity of variances (e g Zar 1984) but data were neither additive nor scaled Thus pa-rametric analysis on untransformed data was performed also due to negligible testpower difference with non-parametric ANOVAs with equal sample size (Zar 1984)and negligible differences with non-parametric MANOVA outcomes (Roa 1992)
Resource use and partitioning by two co-occurring snail species 235
Results
Snail distribut ion and overlap on resources
Snails accumulated on trophic resources (though gradually on maple leafpacks) and were found on empty quadrats (spatial resources) in lower num-bers (Figs 3 and 4 Table 1) Snails were found in significantly higher num-bers on leaf packs regardless of density and species assemblage (Table 1)Preference for leaf packs was striking for both species (Fig 5) Physarsquos higherrelative presence on leaf packs at high snail density (Table 2) was accompa-nied by a significant reduction in H trivolvisrsquo average percent distribution(Table 3) and preference degree (Fig 5) from single- to two-species aquariarather than reflecting an increased preference by Physa (Table 3 Fig 5) Rela-tive distribution on leaves was independent of snail density in single-speciesaquaria but decreased with increasing density for coexisting snails (Table 4)
Physarsquos initial accumulation on periphyton decreased (with temporary tileabandonment at low density) shortly after an evident reduction to almostldquoclean tilerdquo conditions in periphyton biomass was observed at t3ndash t4 in allsnail-present aquaria (Fig 3) H trivolvis visits on periphyton were relativelyfrequent in single-species aquaria but rare when coexisting with Physa(Fig 4) especially at low density (Table 3) Snails rejected periphyton tiles athigh density but coexistence led to an increase in Physa preference (from neu-
Table 1 Differences in snail distribution (as number of individuals) on spatial (emptyquadrats) and trophic resources tested with repeated-measures ANOVAs followed bySNK multiple comparisons Critical values for F are one-tailed values for numeratord f = 8 and denominator d f = 152 Levels of significance are ns (p gt 010) (ppound 010) (p pound 005) and (p pound 001) Significantly different snail distributions onthe trophic and spatial resources resulting from SNK tests (p pound 010) are symbolizedwith different letters (a = lowest 10-day average snail density on resource progressi-vely higher densities listed alphabetically)
SNK (p pound010)
ANOVA trophic resources empty quadrats
leaf peri- macro-experimental condition F p packs phyton phytes A B C D E F
Physa low density 41046 c b b a ab ab a a abPhysa low density with H trivolvis 60252 c b a a a a a a aH trivolvis low density 16331 d a c ab ab a ab ab abH trivolvis low density with Physa 12221 b a a a a a a a a
Physa high density 121278 c ab b ab ab a a a aPhysa high density with H trivolvis 75880 c a b a a a a a aH trivolvis high density 31430 b a a a a a a a aH trivolvis high density with Physa 8239 c ab ab a a ab ab ab ab
236 Paola Lombardo and G Dennis Cooke
Fig 3 Distribution of Physa on spatial (food-devoid quadrats on aquarium surfaces)and trophic resources as average snail numbers on each trophic substrate and on allquadrats One morning (m) and one evening (e) observation per day on days 1 through10 Total numbers per aquarium given on x axis Number of live individuals on eachobservation normalized to initial numbers (when applicable) for graph comparability
Resource use and partitioning by two co-occurring snail species 237
Fig 4 Distribution of H trivolvis on spatial (food-devoid quadrats on aquarium surfa-ces) and trophic resources during the experiment Explanations as in Fig 3
238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 235
Results
Snail distribut ion and overlap on resources
Snails accumulated on trophic resources (though gradually on maple leafpacks) and were found on empty quadrats (spatial resources) in lower num-bers (Figs 3 and 4 Table 1) Snails were found in significantly higher num-bers on leaf packs regardless of density and species assemblage (Table 1)Preference for leaf packs was striking for both species (Fig 5) Physarsquos higherrelative presence on leaf packs at high snail density (Table 2) was accompa-nied by a significant reduction in H trivolvisrsquo average percent distribution(Table 3) and preference degree (Fig 5) from single- to two-species aquariarather than reflecting an increased preference by Physa (Table 3 Fig 5) Rela-tive distribution on leaves was independent of snail density in single-speciesaquaria but decreased with increasing density for coexisting snails (Table 4)
Physarsquos initial accumulation on periphyton decreased (with temporary tileabandonment at low density) shortly after an evident reduction to almostldquoclean tilerdquo conditions in periphyton biomass was observed at t3ndash t4 in allsnail-present aquaria (Fig 3) H trivolvis visits on periphyton were relativelyfrequent in single-species aquaria but rare when coexisting with Physa(Fig 4) especially at low density (Table 3) Snails rejected periphyton tiles athigh density but coexistence led to an increase in Physa preference (from neu-
Table 1 Differences in snail distribution (as number of individuals) on spatial (emptyquadrats) and trophic resources tested with repeated-measures ANOVAs followed bySNK multiple comparisons Critical values for F are one-tailed values for numeratord f = 8 and denominator d f = 152 Levels of significance are ns (p gt 010) (ppound 010) (p pound 005) and (p pound 001) Significantly different snail distributions onthe trophic and spatial resources resulting from SNK tests (p pound 010) are symbolizedwith different letters (a = lowest 10-day average snail density on resource progressi-vely higher densities listed alphabetically)
SNK (p pound010)
ANOVA trophic resources empty quadrats
leaf peri- macro-experimental condition F p packs phyton phytes A B C D E F
Physa low density 41046 c b b a ab ab a a abPhysa low density with H trivolvis 60252 c b a a a a a a aH trivolvis low density 16331 d a c ab ab a ab ab abH trivolvis low density with Physa 12221 b a a a a a a a a
Physa high density 121278 c ab b ab ab a a a aPhysa high density with H trivolvis 75880 c a b a a a a a aH trivolvis high density 31430 b a a a a a a a aH trivolvis high density with Physa 8239 c ab ab a a ab ab ab ab
236 Paola Lombardo and G Dennis Cooke
Fig 3 Distribution of Physa on spatial (food-devoid quadrats on aquarium surfaces)and trophic resources as average snail numbers on each trophic substrate and on allquadrats One morning (m) and one evening (e) observation per day on days 1 through10 Total numbers per aquarium given on x axis Number of live individuals on eachobservation normalized to initial numbers (when applicable) for graph comparability
Resource use and partitioning by two co-occurring snail species 237
Fig 4 Distribution of H trivolvis on spatial (food-devoid quadrats on aquarium surfa-ces) and trophic resources during the experiment Explanations as in Fig 3
238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003
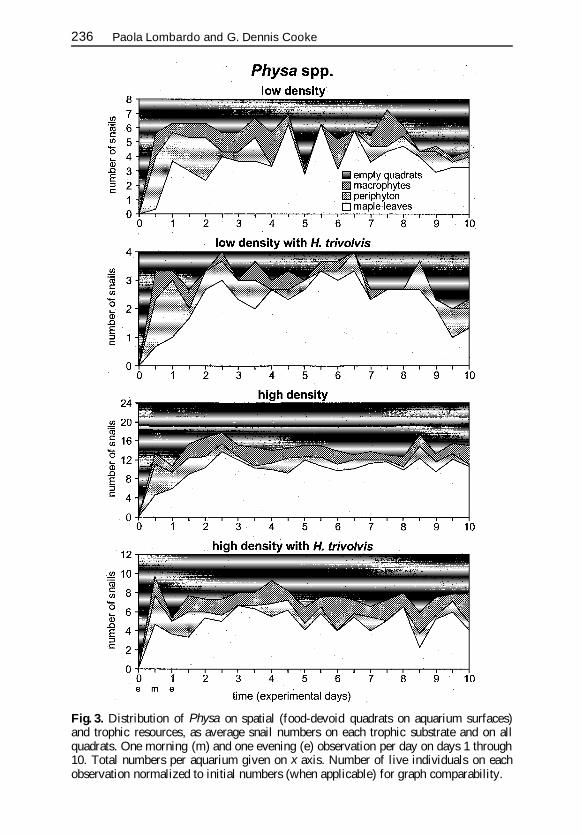
236 Paola Lombardo and G Dennis Cooke
Fig 3 Distribution of Physa on spatial (food-devoid quadrats on aquarium surfaces)and trophic resources as average snail numbers on each trophic substrate and on allquadrats One morning (m) and one evening (e) observation per day on days 1 through10 Total numbers per aquarium given on x axis Number of live individuals on eachobservation normalized to initial numbers (when applicable) for graph comparability
Resource use and partitioning by two co-occurring snail species 237
Fig 4 Distribution of H trivolvis on spatial (food-devoid quadrats on aquarium surfa-ces) and trophic resources during the experiment Explanations as in Fig 3
238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 237
Fig 4 Distribution of H trivolvis on spatial (food-devoid quadrats on aquarium surfa-ces) and trophic resources during the experiment Explanations as in Fig 3
238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

238 Paola Lombardo and G Dennis Cooke
Table 2 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing Physa vs H trivolvispercent relative distribution on spatial (empty quadrats) and trophic resources (averagenumber of snail individuals on each resource out of total number in each experimentalcondition) Positive t values represent Physarsquos average relative distribution higher thanH trivolvisrsquo Levels of significance are n s (p gt 010) (p pound 010) p pound 005) and (p pound001)
low density high density
single species two-species single species two-speciesaquaria aquaria aquaria aquaria
resource t p t p t p t p
maple leaves 1681 1270 n s 1256 n s 2665 periphyton ndash2603 1825 ndash0246 n s ndash 0728 n smacrophytes ndash0369 n s 2121 1947 3301 empty quadrats 1044 n s ndash3342 ndash2075 ndash 3878
Table 3 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing single-species vs two-species percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distributionhigher in two-species than single-species aquaria Other explanations as in Table 2
low density high density
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves 1757 1258 n s ndash0499 n s ndash2059 periphyton 0662 n s ndash 3584 0023 n s 0348 n smacrophytes ndash1899 ndash 2978 2430 ndash0171 n sempty quadrats ndash1809 2812 ndash1073 n s 1843
tral) and a decrease in H trivolvis preference for periphyton (to rejection) atlow density (Fig 5)
Snail presence on living macrophytes remained low (Figs 3 and 4) andtended to be similar to that on empty quadrats (Table 1) except for some Htrivolvis accumulation at low density and some Physa accumulation at highdensity in H trivolvisrsquo presence (Table 3) accompanied by increased prefer-ence (Fig 5) Physarsquos relative presence on macrophytes was significantlyhigher than H trivolvisrsquo in two-species aquaria regardless of snail density (Ta-ble 2) Macrophytes were mildly preferred by Physa in two-species high-den-sity aquaria but were neutral or rejected in all other conditions (Fig 5)
Snail relative presence on empty quadrats tended to be higher in high-den-sity aquaria (Figs 3 and 4 Table 1) Physarsquos and H trivolvisrsquo relative distribu-tions on and rejection degree for quadrats were similar in low-density single-species aquaria (Figs 3ndash5) but H trivolvisrsquo relative presence on quadrats be-came significantly higher than Physarsquos in all other conditions (Table 2) Physa
Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 239
Fig 5 Experiment-long resource (substrate) preference by Physa and H trivolvisfrom distribution data (i e trophic resources treated as substrates or spatial resources)modified after Roarsquos (1992) MANOVA Centered averages plusmn standard error shownDepartures from no-preference patterns were significant in all conditions (Hotelling-based one-tailed F 100 d f = 27 and p 001 for all) Preferred resources (sub-strates) are represented by positive bars Significantly preferred (or rejected) resourcesare those for which the avg plusmn std error interval does not include the x axis or area ofno preference Avg plusmn std error intervals not to be used to infer between average dif-ferences Day zero distributions were excluded from analysis Leav maple leavesperi periphyton macr C demersum sprig Andash F food-devoid quadrats
240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

240 Paola Lombardo and G Dennis Cooke
Table 4 Two-tailed t-tests (n1 = n2 = 20 d f = 38) comparing low-density vs high-density percent relative distribution of Physa and H trivolvis on spatial (empty quad-rats) and trophic resources Positive t values represent average relative distribution ona resource higher in high- than low-density aquaria Other explanations as in Table 2
single-species two-species
Physa H trivolvis Physa H trivolvis
resource t p t p t p t p
maple leaves ndash0861 n s 0429 n s ndash3141 ndash2287 periphyton ndash1285 n s ndash 3389 ndash2058 0740 n smacrophytes ndash0793 n s ndash 1669 3329 2106 empty quadrats 2953 4619 4071 1354 n s
Table 5 Averaged a index on trophic and spatial resources for Physa over H trivolvis(aPhy Hel) and vice-versa (aHel Phy) in single- and two-species trials Alpha values aver-aged over 20 observations (day 1 through day 10) For t-tests n1 = n2 = 20 Criticalvalues for the t-tests are two-tailed values for d f = (n1 + n2) ndash 2 = 38 Levels of signifi-cance are n s (p gt 010) (p pound010) (p pound 005) and (p pound 001)
low density high density
aPhy Hel aHelPhy aPhy Hel aHelPhy
single-species 075 068 t = 0733 n s 092 089 t = 0471 n stwo-species 074 061 t = 1330 n s 073 068 t = 0669 n s
t = 0083 t = 0893 t = 2653 t = 3976n s n s
slightly decreased (low density only p lt 010) and H trivolvis increased theirrelative distribution on quadrats when in each otherrsquos presence (Table 3) Ac-cordingly Physarsquos general rejection for quadrats did not change from single-to two-species aquaria while H trivolvis decreased its degree of rejectionwhen in Physarsquos presence (Fig 5) to the point that trophic substrates as awhole became ldquorejectedrdquo at high density (MANOVA-based preference analy-sis calculated from pooled trophic vs spatial resources statistical significancenot calculable) Snails accumulated on empty quadrats at increasing densityalso in single-species aquaria (Table 4)
General substrate use was similar for Physa and H trivolvis as single-spe-cies overlap of Physa over H trivolvis (aPhy Hel) was statistically the same assingle-species aHel Phy regardless of snail density (Table 5) Both Physa andH trivolvis increased the degree of potential (single-species) mutual spatialoverlap with increasing snail density (two-tailed t-tests from values in Table 5tPhysa = 2153 p lt 005 tH trivolvis = 2912 p lt 001 n1 = n2 = 20 for both) butoverlap tended to decrease when in each otherrsquos presence (single-species vstwo-species aquaria) This decrease was significant at high density (Table 5)
Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 241
Table 6 Results for one-way type I ANOVAs for changes in biomass for leaf packsmacrophytes and periphyton at tF and for mid-term and final nutrient (TP) concentra-tions in the seven experimental conditions Critical values for F are one-tailed valuesfor d f = 614 Levels of significance are n s (p gt010) (p pound 010) (p pound 005) and (p pound 001) SNK multiple comparison tests performed for significant ANOVAsare reported visually in Figs 6 (trophic resources) and 7 (TP)
Variable F p
leaf packs 21859 macrophytes 11167 periphyton 4573
TP mid-term 3435 TP final 6055
Coexistence-induced reduction in averaged a values was more pronounced forH trivolvis than Physa
Snail effect on trophic resources
Periphyton and maple leaves were significantly reduced by Physa and H tri-volvis independently of snail density and assemblage composition though leafconsumption by H trivolvis at low density was lower than in other snail-pre-sent aquaria (Fig 6 and Table 6) Virtually all periphyton and all leaf ediblematerial (i e except for the skeleton of leaf veins) were eaten by snails by tF
Radular damage on macrophytes (C demersum) was not observed and allsprigs appeared in good health at tF but plant response to snail grazing varied(Fig 6) Macrophyte biomass increased significantly in all Physa-only aquariaChange in C demersum biomass (final weight as percent of initial weight) wasweakly but significantly correlated with final number of Physa individuals (r =0690 n = 6 p lt 010 single-species aquaria only) H trivolvis did not haveany detectable net effect on macrophyte biomass but C demersum biomasschange was weakly negatively correlated with final number of H trivolvis in-dividuals (r = ndash0681 n = 6 p lt 010 single-species aquaria only) The effecton C demersum by coexisting snails also depended on snail density (Fig 6)Presence of H trivolvis overrode Physarsquos enhancing effects at low densitywhile Physarsquos enhancement at high density was independent of H trivolvisrsquopresence
Snail effect on total phosphorus concentrat ion (TP)
TP increased from t0 values in most snail-present aquaria (Fig 7 and Table 6)TP at mid-term was significantly higher than initial values (two-tailed t-testtinit vs mid = 3672 n1 = 6 n2 = 21 d f = 25 p lt 001) and then decreased attF except in high-density Physa-only aquaria (two tailed paired t-test
242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

242 Paola Lombardo and G Dennis Cooke
Fig 6 Changes in biomass (expressed as percent of initial biomass present at tF) forthe trophic resources under the experimental conditions (average plusmn standard error)Phy or Hel Physa or H trivolvis single-species aquaria Phy + Hel two-species aqua-ria ctrl controls (no snails) Different letters represent significantly different averages(a = lowest value) according to SNK multiple comparisons of means with p pound 010after significant ANOVAs (Table 6)
Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 243
Fig 7 Mid-term (t5) and final total phosphorus concentration (TP) in the experimentalaquaria (average plusmn standard error) Black triangle indicates initial TP (198 plusmn 49 mglavg plusmn std error n = 6) Other explanations as in Fig 6 ANOVAs in Table 6
Fig 8 Linear correlations between TP and number of snail individuals at tF in single-species aquaria Wide-dash line represents the H trivolvis correlation as number ofPhysa-equivalents (1 adult H trivolvis individual = 4 adult Physa individuals) PhysaTP = 6255 sn ndash 11612 r = 0760 p = 005 H trivolvis TP = 3578 sn + 28799 r =0662 p = 010 H trivolvis as Physa-equivalents TP = 0895 sn + 28799 r = 0662 p= 010 For all correlations n = 6 and d f = 5 Units in correlation equations as ongraph axes
tmid vs fin excluding hi-density Phy = 6896 n1 = n2 = 18 d f = 17 p lt 001) where itincreased by ~25 from mid-term values
Final TP was positively correlated with final number of snail individuals(single-species aquaria only) with higher significance level and significantlysteeper slope for Physa than H trivolvis (t-test for comparison of slope coeffi-cient m t = 17090 d f = 8 p lt 001) (Fig 8) Slope remained significantlysteeper for Physa when snail densities were adjusted to obtain comparable
244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

244 Paola Lombardo and G Dennis Cooke
snail living biomass (t-test for m t = 34226 d f = 8 p lt 001) Percentchange in C demersum biomass was positively weakly correlated with finalTP in Physa-only aquaria but not in H trivolvis or two-species aquaria (rPhysa
= 0701 p lt 010 rH trivolvis = ndash0255 p gt050 rPhy + H triv = 0615 p gt010 n =6 and d f = 5 for all)
Discussion
Coexistence led to changes in distribution and reduction in overlap for bothsnail species but Physa influenced H trivolvis more than vice-versa regard-less of snail density For example H trivolvis reduced its degree of overlapover Physa (i e moved away from Physa) more than vice-versa especially athigh density (Table 5) Overlap reduction depended in part on Physa accumu-lation on leaf packs (low density) or macrophytes (high density) (Table 3 andFig 3) However changes in two-species vs single-species aquaria were morepronounced for H trivolvis which moved away from trophic substrates toquadrats when in Physarsquos presence (Table 3 and Fig 4) H trivolvis displace-ment from trophic substrates in Physarsquos presence was not related to competi-tion for food as H trivolvis can feed on all three trophic resources (Lom-bardo amp Cooke 2002) and total edible material was not depleted at tF(Fig 6)
Uneven responses to coexistence were observed also in other studies Gre-sens (1995) reported diminished foraging activities by chironomids in snailpresence under laboratory non-limiting conditions supporting earlier in situobservations (Cuker 1983) Gresens (1995) argued that snail-induced behav-ioral changes were sufficient to decrease foraging activity in the ldquoless asser-tiverdquo chironomids giving an edge to snails Fratini et al (2000) report ofsimilarly ldquoassertiverdquo snails coexisting with herbivorous crabs in coastal habi-tats Uneven responses to coexistence were observed also for other freshwaterinvertebrates (Costantini amp Rossi 1998) and for snail-snail interactions incoastal (Fletcher amp Creese 1985 Schmitt 1996) and fresh waters (Brown1982) though effects were more pronounced (e g diminished fecundity) dueto lower and probably limiting food quantity Qualitative field observationsof habitat partitioning by coexisting Physa and H trivolvis with Physa ac-cumulating on epiphyton-rich plant material and H trivolvis on sediments (PLombardo pers observ) also support the laboratory results
Highly significant relative snail accumulation on food-devoid quadrats (Ta-ble 4) and increases in a values (Table 5) in single-species aquaria from lowto high density under apparently non-limiting conditions imply occurrence ofdensity-dependent intraspecific interactions in crowded situations for bothPhysa and H trivolvis Temporary abandonment of glass tiles by Physa soon
Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 245
after periphyton appeared to be depleted by t3-t4 was observed at low density(where per capita food availability was higher) but not at high density (Fig 3)suggesting that snails may minimize density-dependent interactions withchanges in behavior perhaps by increasing individual mobility in search ofless crowded substrates or by revisiting recently grazed areas in search of left-over food or less favorite resources
Distribution adjustments by single-species or coexisting Physa and H tri-volvis did not cause major changes in periphyton or leaf pack consumption(Fig 6) Similar leaf pack and periphyton biomass loss in all snail-presentaquaria indicates that snail per capita food consumption may be density-dependent with more snails consuming less food individually supportingother laboratory (Lombardo amp Cooke unpubl) or field observations (Lom-bardo 1989 Cattaneo amp Mousseau 1995) of decreased individual grazingrate at increasing snail density Lower per capita consumption without appre-ciable negative effects on snail growth or mortality also implies that total foodquantity never went below the minimum or ldquomaintenancerdquo level (Eisenberg1970) i e was not limiting
Periphyton biomass was suppressed to almost-zero values in all snail-pre-sent aquaria (Fig 6) indicating that periphyton was a favorite trophic resourcefor both snail species as found elsewhere (e g Brown 1991 Lombardo ampCooke 2002) Most periphyton consumption occurred early with snails aban-doning glass tiles after tiles became ldquocleanrdquo to visual examination at t3-t4(Figs 3 and 4) Tiles may have become a substrate similar to empty quadratsafter early periphyton depletion with consequent avoidance by snails explain-ing the experiment-long low preference or rejection for tiles as a substrate(Fig 5) (resource value in the MANOVA-based preference analysis was assubstrate only as calculations used snail distribution and not food consump-tion data) Tile abandonment by low-density Physa was less pronounced in Htrivolvisrsquo presence (Fig 3) H trivolvis grazing on small-bodied periphytonmay not be as effective as Physarsquos (Barnese et al 1990) suggesting thatPhysa may benefit from left-over periphyton in H trivolvisrsquo presence at mod-erate snail densities
Periphyton removal rate at high snail densities may be too high to maketile revisitation worthwhile for snails as suggested by experiment-long mildrejection of tiles in all high-density aquaria (Fig 5) This apparently activeavoidance of food-devoid tiles supports observations for coastal limpets (Che-lazzi et al 1998) and suggests that snails may be attracted to their trophic re-sources by distant chemoreception though neither this nor earlier studies(Bovbjerg 1968 Broumlnmark 1985) provide conclusive evidence in this re-gard
Food-devoid tiles also represented exposed surfaces that snails may haveactively avoided Although predation on snails did not occur in the laboratory
246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

246 Paola Lombardo and G Dennis Cooke
snails coexisted with several visual predators (mainly fish) in their originalhabitat and may have retained anti-predator behaviors during the experimentincluding hiding in sheltering microhabitats (Turner et al 1999) The veryfew Physa individuals observed on aquarium walls (i e exposed surfaces)and accumulation on shelter-providing leaf packs during morning observationsat low density (Fig 3) support this hypothesis However all Physa individualswere always accounted for during the twice-daily distribution observationswithout disturbance to the aquaria implying that snails did not hide inside theheavily microcolonized maple leaf packs and suggesting that color camou-flaging may be sufficient as protection against visual predators Heavy com-pacted leaves and low oxygen levels likely caused by respiring microdecom-posers (fungi bacteria) at the microhabitat scale may have prevented snails tohide inside the leaf packs until the latter were progressively skeletonized fromthe outer edges though this hypothesis needs a direct verification
High preference for leaf packs (Fig 5) may have been a combination ofpack high value as both a trophic (Fig 6) and a spatial resource for small-bodied snails Initial snail accumulation was lowest on intact leaf packs(Figs 3 and 4) supporting Physarsquos and H trivolvisrsquo mild preference for leaflitter as a trophic resource observed by Lombardo amp Cooke (2002) How-ever most edible leaf material was consumed by tF when only a network ofinedible leaf veins remained intact Though edible material may have beenpresent among veins pack value as a trophic resource likely declined as theoriginal material was being consumed Value as a substrate of such ldquoleaf skele-tonsrdquo may have remained high (as a shelter against potential fish predationTurner et al 1999) Fading value as a trophic resource (Fig 6) but sustainedhigh value as a substrate (Figs 3 through 5) suggest that coarse detritus mayhave an overall higher value as a substrate than as a trophic resource for bothPhysa and H trivolvis
Living C demersum is edible for both snail species (Sheldon 1987 Lom-bardo amp Cooke 2002) but plant biomass increased in Physarsquos presence(Fig 6) Physarsquos enhancing effects on living C demersum may be related to acombination of absence of direct grazing increased plant access to light andnutrients after epiphyton removal by grazing snails (Broumlnmark amp Weisner1992) and phosphorus enrichment of water by snails (Figs 7 and 8 C demer-sum biomass gain positively correlated with TP in Physa-only aquariap lt 010) The weak statistical separation in Fig 7 may have depended in parton the relatively high variability in initial TP (198 plusmn 49 mgl avg plusmn std er-ror) which may have been caused by different short-term initial release ratesfrom trophic resources probably exacerbated by the small aquarium sizeHowever lower P excretion by H trivolvis (Fig 8) supports Rybakrsquos (2002)observations that the magnitude of nutrient excretion by individual snails isspecies-specific and suggests that H trivolvisrsquo potential to enhance C demer-
Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 247
sum by increasing P availability in its immediate environment may be lowerthan Physarsquos H trivolvisrsquo lower ability to solubilize P may be the major causefor the observed absence of enhancing effects on C demersum by H trivolvis(Fig 6) since similar periphyton consumption (Fig 6) and macrophyte aban-donment patterns (Figs 3 and 4) suggest that H trivolvisrsquo potential for epiphy-ton removal is similar to Physarsquos As macrophyte enhancement in snail pres-ence may depend also on factors other than nutrient excretion and epiphytonremoval (Lombardo amp Cooke unpubl) specific studies on snail-P-macro-phyte interactions are needed to address this issue
A weak but significant (p lt 010) positive correlation between number ofPhysa individuals and plant biomass gain suggests that Physa did not graze onliving C demersum supporting other evidence that C demersum is an unpre-ferred plant species for Physa and other gastropods (Sheldon 1987 Pi-nowska 2002) and that Physa does not consume living C demersum tissueunless it is the only available food (Sheldon 1990 Lombardo amp Cooke2002) Low-density Physa tended to abandon macrophytes at roughly thesame time they abandoned glass tiles (Fig 3) suggesting that C demersumvalue as a trophic resource may have declined following the likely epiphytondepletion Preference by Physa and other gastropods for attached algae overliving macrophyte tissue (Broumlnmark 1989 Kornijoacutew et al 1995 Lombardoamp Cooke 2002) supports this hypothesis Epiphyton depletion by Physa mayhave led to increased plant leaf surface access to light nutrients andor CO2
(Sand-Jensen amp Borum 1991 Jones et al 2000) leading to increased plantgrowth (Fig 6) but this hypothesis could not be verified Involvement of P en-richment in the immediate water environment of C demersum was likely asTP was significantly correlated with number of Physa individuals at tF (Fig 8)and changes in plant biomass also were correlated with final TP (plt 010) En-hancement effects and absence of grazing on living C demersum when achoice is available was found also for other freshwater snail species (Under-wood 1991 Pinowska 2002)
Physarsquos different preference for macrophytes under different conditions(Fig 5) may have been associated with a combination of Physarsquos overall pref-erence for attached algae and H trivolvis presence Macrophytes were rejectedin high-density Physa-only aquaria but preferred by Physa in the presence ofH trivolvis (Fig 5) H trivolvisrsquo low effectiveness on small algae (Barnese etal 1990) may have maintained a ldquolawnrdquo of adnate algae for Physa in crowdedconditions but higher total food availability in low-density aquaria may haverendered colonization of other trophic resources (leaf packs) more worthwhilefor Physa once periphyton and epiphyton became scarce
H trivolvis did not have any appreciable effect on plant biomass (Fig 6) Aweakly significant (p lt010) negative correlation between number of H trivol-vis individuals and macrophyte biomass at tF suggests that H trivolvis may re-
248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

248 Paola Lombardo and G Dennis Cooke
sort to grazing on living macrophytes only under conditions of low per capitafood availability as also found for Physa in in situ experiments (Sheldon1987 1990) Neither radular damage nor tissue aging (leaf decoloration orshedding) on C demersum were observed suggesting that when occurringgrazing targeted older decomposing tissues as also observed for other fresh-water snail species (Kornijoacutew et al 1995 Pinowska 2002)
Lower H trivolvis ability to solubilize P (Fig 8) and absence of correla-tion between final TP and changes in plant biomass in H trivolvis-only aqua-ria (pgt 050) suggests that H trivolvis effect on living macrophytes may resultfrom a balance between negative (grazing) and positive forces (P excretion)Season-based plant condition with June sprigs relatively close to the annualpeak in biomass accrual (Best amp Meulemans 1979) also may have con-tributed to counteract H trivolvis grazing on C demersum as also suggestedby H trivolvis ultimately depressing past-peak (November) C demersum bio-mass in multiple-choice preference trials (Lombardo amp Cooke 2002) Alsodifferent factors may have contributed differently and in non-additive ways assuggested by non-linear effects of snail grazing and nutrient enhancement onstream periphyton (Rosemond et al 1993) and lake vegetation (Pinowska2002)
Snail coexistence also led to non-additive effects on macrophyte biomassdepending on snail density Net macrophyte enhancement by Physa at highdensity overrode H trivolvisrsquo (potential) negative effects on C demersumwhile the opposite occurred at low density (Fig 6) suggesting that macro-phytes in natural situations may benefit the most in the presence of Physa-dominated abundant snail assemblages Physarsquos positive effects on macro-phytes (nutrient excretion removal of light-limiting epiphyton and lack of di-rect grazing) may be further enhanced by Physa-driven lower presence ofldquoless assertiverdquo snail species such as H trivolvis Physa would benefit from acontinuous ldquolawnrdquo of small algae left behind by H trivolvis while limiting Htrivolvis colonization (and therefore potential consumption) of living macro-phytes as long as availability of other trophic resources (e g macrodetritus)is sufficient to support H trivolvis Physa-mediated enhancement may be crit-ical in inner macrophyte beds where light may be most limiting and wheresnails tend to accumulate (Marklund et al 2001) Dense vegetation in meso-to eutrophic littoral zones with sufficient nutrients to sustain primary produc-tion sufficient protection of small-bodied Physa from (fish) predation(Broumlnmark 1988) and production of macrodetritus from decaying plantsmay be maintained in the presence of physid-dominated invertebrate assem-blages following a positive-feedback loop (Broumlnmark amp Weisner 1992)
Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 249
Acknowledgements
We thank Ms Christina L Brant (KSU currently at the EHSrsquo Lakes Mgmt Unit ofLake County IL) for her help in the laboratory and two anonymous reviewers fortheir constructive criticism on an earlier version of the manuscript This research ispart of a PhD dissertation prepared at Kent State University by Paola Lombardowho extends her gratitude to the Dissertation Committee members and to Prof DavidW Waller (KSU) for advice on statistical analysis
References
Barnese L E Lowe R L amp Hunter R D (1990) Comparative grazing effi-ciency of pulmonate and prosobranch snails ndash J N Amer Benthol Soc 9 35ndash44
Best E P H amp Meulemans J T (1979) Photosynthesis in relation to growth anddormancy in Ceratophyllum demersum ndash Aquat Bot 6 53 ndash65
Bovbjerg R V (1968) Responses to food in lymnaeid snails ndash Physiol Zool 41412 ndash423
Brendelberger H (1994) Food selection experiments with two freshwater gastro-pods ndash Verh Internat Verein Limnol 25 2341 ndash2345
ndash (1997) Contrasting feeding strategies of two freshwater gastropods Radix peregra(Lymnaeidae) and Bithynia tentaculata (Bithyniidae) ndash Arch Hydrobiol 1401ndash21
Broumlnmark C (1985) Interactions between macrophytes epiphytes and herbivoresan experimental approach ndash Oikos 45 26ndash30
ndash (1988) Effects of vertebrate predation on freshwater gastropods an exclosure exper-iment ndash Hydrobiologia 169 363 ndash 370
ndash (1989) Interactions between epiphytes macrophytes and freshwater snails a re-view ndash J Moll Studies 55 299 ndash311
Broumlnmark C amp Vermaat J (1998) Complex fish-snail-epiphyton interactions andtheir effects on submerged freshwater macrophytes ndash In Jeppesen E Soslashnder-gaard Ma Soslashndergaard Mo amp Christoffersen K (eds) The StructuringRole of Submerged Macrophytes in Lakes ndash Springer-Verlag New York NY pp47ndash68
Broumlnmark C amp Weisner S E B (1992) Indirect effects of fish community struc-ture on submerged vegetation in shallow eutrophic lakes an alternative mecha-nism ndash Hydrobiologia 243244 293 ndash301
Brown K M (1982) Resource overlap and competition in pond snails an experi-mental analysis ndash Ecology 63 412 ndash422
ndash (1991) Mollusca Gastropoda ndash In Thorp J H amp Covich A P (eds) Ecologyand Classification of North American Freshwater Invertebrates ndash Academic PressSan Diego CA pp 285 ndash314
ndash (1997) Temporal and spatial patterns of abundance in the gastropod assemblage of amacrophyte bed ndash Amer Malacol Bull 14 27ndash33
Brown K M Alexander J E amp Thorp J H (1998) Differences in the ecologyand distribution of lotic pulmonate and prosobranch gastropods ndash Amer MalacolBull 14 91ndash 101
250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

250 Paola Lombardo and G Dennis Cooke
Cattaneo A amp Kalff J (1986) The effect of grazer size manipulation on periphy-ton communities ndash Oecologia 69 612 ndash617
Cattaneo A amp Mousseau B (1995) Empirical analysis of the removal rate of peri-phyton by grazers ndash Oecologia 103 249 ndash254
Chelazzi G Santini G amp Della Santina P (1998) Route selection in the fo-raging of Patella vulgata (Mollusca Gastropoda) ndash J Mar Biol Assoc U K 781223 ndash1232
Clampitt P T (1970) Comparative ecology of the snails Physa gyrina and Physa in-tegra (Basommatophora Physidae) ndash Malacologia 10 113 ndash151
Clesceri L S Greenberg A E amp Trussell R R (eds) (1989) Standard Meth-ods for the Examination of Water and Wastewater 17th ed ndash Am Public HealthAssoc Washington DC
Costantini M L amp Rossi L (1998) Competition between two aquatic detritivorousisopods ndash a laboratory study ndash Hydrobiologia 368 17ndash 27
Cuker B E (1983) Competition and coexistence between the grazing snail LymnaeaChironomidae and microcrustacea in an arctic epilithic lacustrine community ndashEcology 64 10ndash15
Eisenberg R M (1970) The role of food in the regulation of the pond snail Lymnaeaelodes ndash Ecology 51 680 ndash684
Fletcher W J amp Creese R G (1985) Competitive interactions between co-occur-ring herbivorous gastropods ndash Mar Biol 86 183 ndash191
Fratini S Cannicci S amp Vannini M (2000) Competition and interactions be-tween Neosarmatium smithi (Crustacea Grapsidae) and Terabralia palustris (Mol-lusca Gastropoda) in a Kenyan mangrove ndash Mar Biol 137 309 ndash316
Gresens S E (1995) Grazer diversity competition and the response of the periphy-ton community ndash Oikos 73 336 ndash346
Jokinen E H (1987) Structure of freshwater snail communities species-area rela-tionships and incidence categories ndash Amer Malacol Bull 5 9ndash19
Jones J I Eaton J W amp Hardwick K J (2000) The influence of periphyton onboundary layer conditions a pH microelectrode investigation ndash Aquat Bot 67191 ndash206
Kornijoacutew R Gulati R D amp Ozimek T (1995) Food preference of freshwater in-vertebrates comparing fresh and decomposed angiosperm and a filamentous algandash Freshwat Biol 33 205 ndash 212
Levins R (1968) Evolution in Changing Environments ndash Princeton University PressPrinceton NJ
Lombardo P (1989) Livelli di predazione e variazioni delle strutture trofiche a basedetrito nel Lago Albano ndash Unpublished Thesis of Laurea La Sapienza State Uni-versity of Rome Rome I
ndash (2001) Effects of freshwater gastropods on epiphyton macrophytes and watertransparency under meso- to eutrophic conditions ndash PhD Dissertation Kent StateUniversity Kent OH
Lombardo P amp Cooke G D (2002) Consumption and preference of selected foodtypes by two freshwater gastropod species ndash Arch Hydrobiol 155 667 ndash685
Marklund O Blindow I amp Hargeby A (2001) Distribution and diel migrationof macroinvertebrates within dense submerged vegetation ndash Freshwat Biol 46913 ndash924
Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003

Resource use and partitioning by two co-occurring snail species 251
Martin T H Crowder L B Dumas C F amp Burkholder J M (1992) Indirecteffects of fish on macrophytes in Bays Mountain Lake evidence for a littoraltrophic cascade ndash Oecologia 89 476 ndash 481
May R M (1972) Will a large complex system be stable ndash Nature 238 413 ndash414Muntildeoz I Real M Guasch H Navarro E amp Sabater S (2000) Resource
limitation by freshwater snail (Stagnicola vulnerata) grazing pressure an experi-mental study ndash Arch Hydrobiol 148 517 ndash532
Pinowska A (2002) Effects of snail grazing and nutrient release on growth ofmacrophytes Ceratophyllum demersum and Elodea canadensis and the filament-ous green alga Cladophora sp ndash Hydrobiologia 479 83ndash 94
Roa R (1992) Design and analysis of multiple-choice feeding-preference experi-ments ndash Oecologia 89 509 ndash515
Rochette R Morissette S amp Himmelman J H (1995) A flexible response to amajor predator provides the whelk Buccinum undatum L with nutritional gains ndashJ Exp Mar Biol Ecol 185 167 ndash180
Rosemond A D Mulholland P J amp Elwood J W (1993) Top-down and bot-tom-up control of stream periphyton effects of nutrients and herbivores ndash Ecol-ogy 74 1264 ndash1280
Rossi L (1985) La nicchia ecologica ndash Scuola e Cultura 93 265 ndash 278Rybak J I (2002) The release of phosphorus and nitrogen by living and decompos-
ing snails ndash Pol J Ecol 50 17ndash24Sand-Jensen K amp Borum J (1991) Interactions among phytoplankton periphyton
and macrophytes in temperate freshwater and estuaries ndash Aquat Bot 41 137 ndash175Sand-Jensen K amp Madsen T-V (1991) Minimum light requirements of submerged
freshwater macrophytes in laboratory growth experiments ndash J Ecol 79 749 ndash764Schmitt R J (1996) Exploitation competition in mobile grazers trade-offs in use of
a limited resource ndash Ecology 77 408 ndash425Sheldon S P (1987) The effects of herbivorous snails on submerged macrophyte
communities in Minnesota lakes ndash Ecology 68 1920 ndash1931ndash (1990) More on freshwater snail herbivory a reply to Broumlnmark ndash Ecology 71
1215 ndash1216Slobodchikoff C N amp Schulz W C (1980) Measures of niche overlap ndash Ecol-
ogy 61 1051 ndash1055Stoner A W Ray M amp Waite J M (1995) Effects of a large herbivorous gastro-
pod on macrofauna communities in tropical seagrass meadows ndash Mar EcolProgr Ser 121 125 ndash131
Thomas J D Nwanko D I amp Sterry P R (1985) The feeding strategies of juve-nile and adult Biomphalaria glabrata (Say) under simulated natural conditions andtheir relevance to ecological theory and snail control ndash Proc Royal Soc Lond B266 177 ndash209
Turner A M Fetterolf S A amp Bernot S J (1999) Predator identity and con-sumer behavior differential effects of fish and crayfish on the habitat use of afreshwater snail ndash Oecologia 118 242 ndash247
Underwood G J C (1991) Growth enhancement of the macrophyte Ceratophyllumdemersum in the presence of the snail Planorbis planorbis the effect of grazingand chemical conditioning ndash Freshwat Biol 26 325 ndash334
Zar J H (1984) Biostatistical Analysis 2nd ed ndash Prentice Hall Englewood CliffsNJ
Submitted 11 April 2003 accepted 27 October 2003