porphyrin assembly formation on helical biopolymers
TRANSCRIPT

Journal of Inorganic and Organometallie Polymers, VoL 3, No. 1, I993
Porphyrin Assembly Formation on Helical Biopolymers
Robert F. Pasternack 1'2 and Esther J. Gibbs ~
Received July 14, t992
Studies on the interactions of trans-bis(N-methylpyridinium-4-yl)diphenyl- porphine with DNA and polyglutamate lead to a model for binding in which the porphyrin, under appropriate conditions of concentration and ionic strength, forms long-range assemblies on the polymer template. Large, con- servative induced circular dichroism signals are observed in the Soret region as a result of the assembly process. The phasing of the signal is a consequence of the helical sense of the porphyrin stacking, which, in turn, reflects the helicity of the polymer. The assembly organization can be inhibited by modifying solution conditions either to make porphyrin aggregation less favorable or to convert the polymer to a nonhelical conformation.
KEY WORDS: Self-assembly; porphyrins; aggregation; circular dichroism; nucleic acids; polypeptides.
I N T R O D U C T I O N
In add i t i on to te r r i tor ia l b ind ing observed for vi r tual ly any posi t ively charged species, ca t ionic meso- t e t r aa ry l po rphyr in s and me ta l t opo rphy r in s have been shown to in terca la te a n d / o r b ind external ly in the (minor ) groove of D N A [1 ]. A number of spect roscopic and kinet ic me thods have p roved useful in inves t igat ions of the re la t ionships a m o n g p o r p h y r i n structure, b ind ing mode, and base pa i r preference, none more so than circular d ichro i sm [ 2 - 5 ] . A l though po rphyr ins such as te t rakis (N-methyl - py r id in ium-4-y l )po rph ine (H2T4; Fig. 1) have i m p r o p e r axes of ro ta -
Department of Chemistry, Swarthmore College, Swarthmore, Pennsylvania 19081, Depart- ment of Chemistry, Goucher College, Towson, Maryland 21204.
2 To whom correspondence should be addressed at Department of Chemistry, Swarthmore College, Swarthmore, Pennsylvania 19081.
77
I053-0495/93/0300~)077507.00/0 © 1993 Plenum Publishing Corporation

78 Pasternack and Gibbs
H3C-N÷ N +~CHa
~ ~ ~ N H HN
H3C / ~CH 3
H~C.
NH
Fig. 1. Structures of several water-soluble meso-substituted porphyrins. Top: Tetrakis(N- methylpyridinium-4-yl)porphine, H2T4. Lower left: cis-bis(N-methylpyridinium-4-yl)diphenyl- porphine, cis-H2Pagg. Lower right: trans-bis(N-methylpyridinium-4-yl)diphenylporphine, trans-H2 P ~ .
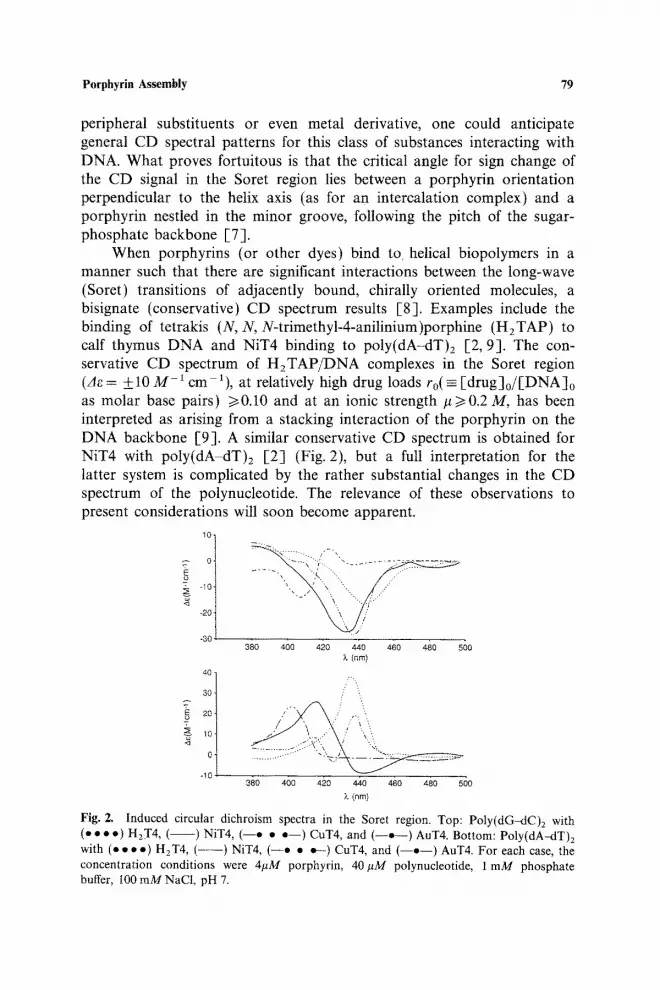
tion and are, therefore, not optically active, a circular dichroism (CD) spectrum is induced when the porphyrin binds to a chiral polymer like DNA. As we demonstrated in 1983 [2, 3], the sign of the induced signal (for monodispersed porphyrins) is a reliable indicator of binding mode. Negative induced CD signals in the Soret region correlate with porphyrin intercalation and positive induced CD signals are obtained in this spectral region when the porphyrin is bound externally in a groove. Several representative spectra are shown in Fig. 2 for poly(dG-dC)2, a synthetic DNA for which intercalation is favored with porphyrins having requisite structural features [2], and poly(dA-dT)2, which binds porphyrins in its minor groove without intercalation. In addition to the sign of the signal, the magnitude, 3e = eL-e~, is worth noticing. The percentage of the total absorption signal in the Soret region observed in this difference, I100 3e/~[, is less than 0.03% for the spectral features shown.
Interest was heightened in these measurements with synthetic polynucleotides by the finding that similar patterns, in which sign (and therefore binding mode) correlates with porphyrin structure, apply for natural DNAs as well [2]. A theoretical basis for this type of correlation was provided by Norden and co-workers [6]. The size and sign of induced circular dichroism spectra depend on a number of factors including proximity, magnitudes, and relative orientations of the transition moments of the chromophore and the chirally arranged bases. Since the orientation of the porphyrin electric dipole transition is relatively unaffected by
Porphyrin Assembly 79
peripheral substituents or even metal derivative, one could anticipate general CD spectral patterns for this class of substances interacting with DNA. What proves fortuitous is that the critical angle for sign change of the CD signal in the Soret region lies between a porphyrin orientation perpendicular to the helix axis (as for an intercalation complex) and a porphyrin nestled in the minor groove, following the pitch of the sugar- phosphate backbone E7].
When porphyrins (or other dyes) bind to helical biopolymers in a manner such that there are significant interactions between the long-wave (Soret) transitions of adjacently bound, chirally oriented molecules, a bisignate (conservative) CD spectrum results E8]. Examples include the binding of tetrakis (N, N, N-trimethyl-4-anilinium)porphine (H2TAP) to calf thymus DNA and NiT4 binding to poly(dA-dT)2 [2, 9]. The con- servative CD spectrum of H2TAP/DNA complexes in the Soret region (Ae = _+t0 M-1 cm 1), at relatively high drug loads ro(= [drug]o/[DNA]o as molar base pairs) >~0.10 and at an ionic strength #>0.2 M, has been interpreted as arising from a stacking interaction of the porphyrin on the DNA backbone [9]. A similar conservative CD spectrum is obtained for NiT4 with poly(dA-dT)2 [2] (Fig. 2), but a full interpretation for the latter system is complicated by the rather substantial changes in the CD spectrum of the polynucleotide. The relevance of these observations to present considerations will soon become apparent.
l° I -~o
-20
40
4° l 30
'~ 10
01 -10
...... ,. -)L .. . . . . . . . . . . .
• _, .\ - . . . / .
"\, /
380 4o0 420 440 4~0 4~0 s60 X (nm)
.--,
/ -- / \ /" :- ,.'".......
3§0 400 4;20 440 460 480 ~ 0 X (nm)
Fig. Z Induced circular dichroism spectra in the Soret region. Top: Poly(dG-dC)z with (o • • o) HzT4 , ( ) NIT4, ( - - e • o - - ) CUT4, and ( - - o - - ) AuT4. Bottom: Poly(dA~lT)2 with (e • • o) H2T4, ( - - - ) NIT4, ( - -e • o--) CuT& and ( - -o - - ) AuT4. For each case, the concentration conditions were 4/~M porphyrin, 40#M polynucleotide, 1 mM phosphate buffer, I00 mMNaCI, pH 7.

80 Pasternack and Gibbs
It was shown some years ago that the addition of certain polymers and salt to solutions of high molecular weight DNA causes a transition of the polynucleotide to a compact molecular configuration [10]. Because the changes are polymer and salt induced, these are referred to as psi or ~-condensates [11]. The formation of ¢-condensates produces some turbidity and alters the uv absorption spectrum of DNA [12], but the most dramatic change, and the one which has attracted the most attention, is the impact on the CD spectrum. The spectrum is characterized by very large extinction coefficient differences together with a significant scattering component. Methods have been developed to separate these two factors [13, 14] and theoretical models have been presented for the analysis of these C-type spectra [15-17]. The circular dichroism signal of the so-called ¢( + )-condensate is shown in Fig. 3; that of ordinary B-form DNA is also shown for comparison. A model for the 0-condensate has been proposed in which the molecules are oriented parallel to each other in planar arrays, packed into layers. The directions of the long axes of the molecules in adja- cent layers are displaced from one another, leading to a helical structure which may be right- or left-handed, producing either positive or negative CD signals [18]. What emerges from the theory of light interaction with these aggregates is that the primary mechanism for the anomalous optical properties involves a collective excitation of the entire aggregate, made possible by long-range induced dipole couplings [ 15 ]. The CD spectrum is not derivable from nearest-neighbor or next-to-nearest-neighbor inter- actions alone. Rather, it arises as a resonance phenomenon involving the long-range chiral packing.
200
/ 0 , L (rim)
& °
, y ' \
-200 I 210 350
I 330
(rim)
Fig. 3. C i rcu la r d ichro i sm spect ra of a ¢ ( + ) -condensate and of B-form D N A ; [ D N A ] = 96.8 #M, # = 0.1 M. ( . . . . ) ¢ ( + )-condensate; ( o . • • ) B-DNA. Inset : B - D N A spec t rum on a m o r e convenient scale.

Porphyrin Assembly 81
Tin•co, Maestre, and co-workers, in studies of drug interactions with these condensates, examined the binding of H2T4 and some of its metal derivatives [19]. They found that the metal-free porphyrin as well as its copper(II) and nicket(II) derivatives, all of which intercalate into DNA [2], provide extremely large 0-type spectra in the Soret region [e.g., Ae ,-~ 300-600 M 1 cm 1 with O( + )], the sign of which reflects the twist of the condensate. In contrast, MnT4, which does not intercalate [2], does not display anomalously large CD signals when bound to 0-aggregates. Although presumably still interacting with DNA, this metalloderivative apparently does not have a periodic repeat structure necessary to produce large CD effects.
Studies with these 0-type DNA condensates were extended by us [20] to include two mixed-periphery porphyrins, cis- and trans-bis(N-methyl- pyridinum-4-yl)diphenylporphine (cis- and trans-H2Pagg; Fig. 1). These porphyrins differ from H2T4 in their propensity to aggregate in aqueous solutions containing electrolyte, the self-association producing markedly altered Soret spectra. Figure 4 shows the influence on the absorption
1.50
c'-
"~,- 0.75 0 m <
0 350
i I
f" , . I .--.. / :.. ( "::..
I 50O
;L (nm)
Fig. 4. Absorp t ion spect ra of trans-H2Pagg in the Soret region, ( ) In water; (e • • • ) in 100 m M NaCI; ( - ) in 100 m M NaC1 con ta in ing 2 0 % glycerol. [trans-H~P,,8] = 5.1 ,uM.

82 Pasternack and Gibbs
spectrum of adding 100 mM NaC1 to an aqueous solution of trans-H2Pagg. When cis- or trans-H2Pagg is allowed to interact with O-condensates, large induced CD spectra having a conservative profile are obtained, the phasing of which reports the condensate twist (Fig. 5). The conservative profile implies that the porphyrin chromophores are strongly coupled to one another electronically and the size of the signal (llOOAe/e[~0.5 1%) suggests a periodic repeat over long distance.
When, under appropriate conditions of concentration and ionic strength, either of these two aggregating porphyrins is added to calf thymus DNA to which no peptide has been added, extremely large conservative CD signals are again obtained in the Soret region (Fig. 6), whereas CD signals of the DNA in the ultraviolet are almost totally unaffected. Light-scattering experiments further indicate that the DNA is not extensively aggregated, although the porphyrin chromophores appear to be so [-20]. The size of the induced CD signals is consistent with a long-range periodic repeat of porphyrins on a DNA template; no such signals are obtained under condi- tions in which the porphyrins are aggregated in solution (no DNA). In the presence of polynucleotides but under low-salt conditions, these porphyrins provide small negative CD features consistent with an intercalated species, but as salt is added to foster aggregation, large signals are produced [21].
-600
-800
J
%.
" . . . "
600
4OO
200
o
-200
-400
Fig. 5. of a O( + )-condensate (e • • •) and a ~,( - )-condensate (
i i t i | | i | i i i i
400 450 500
X (rim) Induced circular dichroism spectra in the Soret region of trans-H2 Pagg in the presence
).

Porphyrin Assembly 83
20O
0
-200
4 0 0 •
-600 '
-800
- t ,000 380
i
. . . :
4;0 &o 440 4;0 4;0 s;o ~, (rim)
Fig. 6. Induced C D spectrum in the Soret region of trans-H2P~gg when added to B-form DNA; [trans-H2Pagg ] = 5.1 ~M, [ D N A ] = 4 3 . 4 #M, [phosphate] = 1 raM, and [N aC I] = 100 mM. (e • • • ) no glycerol added; ( . . . . . . ) 20% glycerol/water.
Based upon these findings--as well as other spectroscopic and kinetic measurements [22J--we suggest that these large induced CD signals are signatures for the formation of long-range, helical porphyrin structures, with DNA serving as a template. The driving force for this assembly forma- tion is proposed to arise from the tendency for these porphyrins to form stacking-type aggregates, a property which can be tuned and modified by metal substitution. Therefore, whereas trans-CuPagg aggregates at least as extensively as does the parent free base porphyrin and, like trans-HzPagg, forms supramolecular assemblies on DNA, trans-AuPagg remains mono- meric in solution over the conditions considered and produces relatively small CD signals when binding to DNA, typical of a monodispersed porphyrin [21, 22].
'To test this model of porphyrin assembly formation we have con- sidered and reported on the interactions of H2T4 and trans-H2Pagg with a single-stranded DNA [23], but a more rigorous test is provided by a study of binding to a helical polyanion containing no purine or pyrimidine bases. We report here on findings obtained with polyglutamate. In addition, we examine the impact on the CD signals characteristic for assembly formation of using a glycerol/water mixed solvent system, a medium which promotes monomerization of the porphyrin.
EXPERIMENTAL
The two porphyrins used in these studies, tetrakis(N-methylpyridinium- 4-yl)porphine (H2T4) and trans-bis(N-methylpyridinium-4-yl)diphenyl-

84 Pasternack and Gibbs
porphine (trans-H2P,gg), were obtained from Midcentury as chloride salts. The identity and extent of peripheral methylation of these substances were checked by IH NMR in DMSO-d 6. Concentrations were determined in water using ~=2.26x10 s and 2.40x105M -1 cm -1, respectively, at the Soret maxima [24,25]. Calf thymus DNA (phenol extracted), poly(dA-dT)2, and poly(dG-dC)2 were obtained from Pharmacia and were used after extensive dialysis against a solution containing 1.0 mM phosphate buffer and 8.0 mM NaC1 (pH 7.0). Poly-D- and poly-L-glutamic acids were purchased from Sigma Chemical Co. The polypeptides were dialyzed against and stored in 2 mM acetate buffer, pH 4.5. All more standard reagents were obtained from Fisher Scientific and used without additional purification.
Since our earliest work with these prophyrins [20], we have found that to obtain quantitatively reproducible results, a strict protocol must be followed in the preparation of samples. In this protocol, DNA-containing solutions are first incubated with porphyrin under low-salt conditions; NaC1 is the last reagent added. Results are more precisely reproducible with polypeptides when the porphyrin is added last.
Spectral measurements were carried out on a Nicolet 9420 UV/VIS spectrophotometer and circular dichroism ~spectra were taken on Aviv 62DS and Jasco 600 spectrometers.
RESULTS AND DISCUSSION
Mixed Solvent Studies. To explore the proposed connection between assembly formation and the extraordinarily large CD signals obtained for the trans-HzPagg/DNA system, we considered the influence of a mixed solvent medium. It had been shown previously that the addition of one of a variety of water-miscible organic solvents can convert porphyrin aggregates not involving covalent links to monomers [24, 26], and indeed, glycerol proves effective in the deaggregation of trans-HzPagg in solution. Referring to Fig. 4, it can be seen that the addition of 100 mM NaC1 to an aqueous solution of trans-H2Pagg (no DNA added) leads to major changes in the absorption spectrum in the Soret region consistent with the conversion of a monomeric chromophore to an aggregate [27]. However, a 20% glycerol/water solvent restores the Soret band to that characteristic of porphyrin monomers.
A solution containing 5.1 #M trans-HzPagg, 43.4/~M DNA, and 100 mMNaC1 (added last) produced the CD spectrum shown in Fig. 6, consistent with assembly formation. However, under identical porphyrin, DNA, and salt conditions but in a solvent system containing 20% glycerol, the induced CD spectrum obtained is nearly two orders of magnitude

Porphyrin Assembly 85
smaller (see Fig. 6). The presence of glycerol, at a concentration level which markedly inhibits aggregation of trans-HzPagg in solution, also has a significant impact on the CD signal obtained for the trans-H2Pagg/DNA system, in a manner predicted by the assembly model. A lowered tendency for porphyrin aggregation leads to less extensive assembly formation and smaller circular dichroism features. This influence of solvent medium may help account for the conclusion reached by Munson and Fiel that cis- and trans-H2Pagg behave as standard DNA intercalators [28]. Their experiments on the relaxation of closed circular D N A by topoisomerase I were conducted in a 20% glycerol medium, and indeed, as is now apparent, under these conditions some of the more interesting and unusual aspects of D N A binding by these porphyrins are masked. In the highly cooperative processes we have described issues of medium and experi- mental protocol are crucial and must be carefully considered.
Polypeptide Studies. By raising the pH, polyglutamic acid can be converted from its highly ordered c~-helical form to a random coil form in which there are only limited regions of helicity. We were, therefore, able to consider the interaction of H2T4 and trans-H2Pagg with helical and random coil forms of a polypeptide by simply changing the pH from 4.5 to about 7 [29]. As shown in Fig. 7, trans-H2Pagg (but not H2T4) produces
1,0- ~ ~
~ 0.5- 7 ---. \
....
380 45O k (rim)
1,000 -~ ...
500 i "'... , "- 0 . . . . . . . . . . . . .
<3 -500 1 .... ""'" "",., y"
3'80 4'00 420 440 460 480 500 L (rim)
Fig. 7. Top: Absorption spectra of trans-H2P,g ~ in the Soret region; [trans- H2P,g~]=5-9pM- ( ) No protein; [NaC1]= 10raM; ( . . . . ) 50#M protein; [NaC1] = 10 raM; (o • • •) 50 pM protein; [NaCl] = 100 raM. Bottom: Induced circular dichroism spectrum of trans-H2P~gg/L-polyglutamate at pH 4.5; [NaC1]=100mM. At [NaC1] = t0 mM, the signal is at least 25x smaller. The CD spectrum of HzT4 under these same conditions ([NaCI] = 100raM) is so small as to appear as a horizontal line on this scale.
86 Pasternack and Gibbs
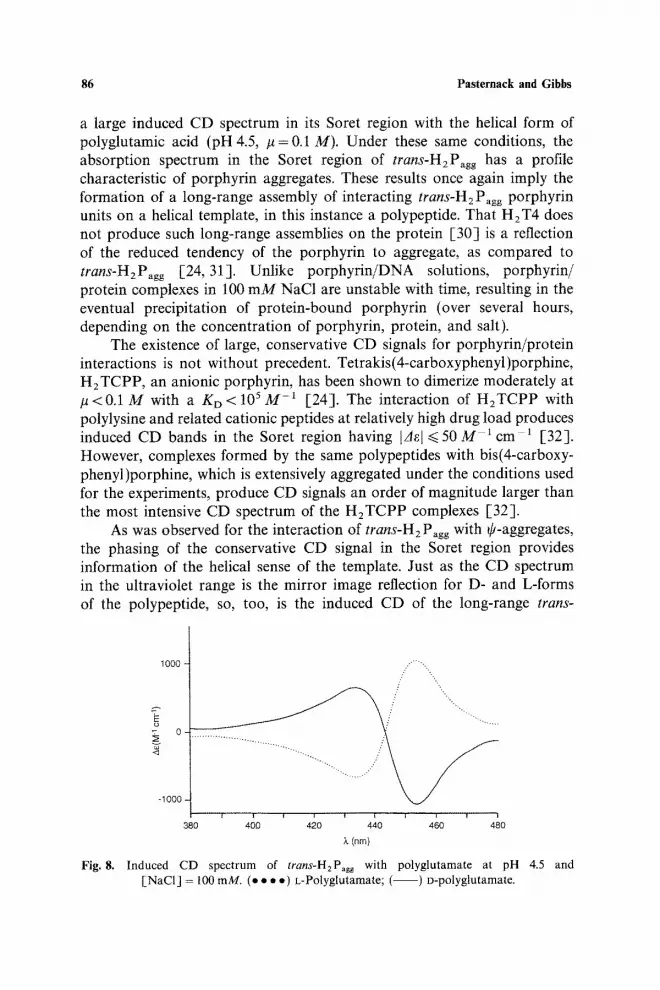
a large induced CD spectrum in its Soret region with the helical form of polyglutamic acid (pH 4.5, #=0.1 M). Under these same conditions, the absorption spectrum in the Soret region of trans-H2Pagg has a profile characteristic of porphyrin aggregates. These results once again imply the formation of a long-range assembly of interacting trans-H2Pagg porphyrin units on a helical template, in this instance a polypeptide. That H2T4 does not produce such long-range assemblies on the protein [30] is a reflection of the reduced tendency of the porphyrin to aggregate, as compared to trans-H2Pagg [24,31]. Unlike porphyrin/DNA solutions, porphyrin/ protein complexes in 100 mM NaC1 are unstable with time, resulting in the eventual precipitation of protein-bound porphyrin (over several hours, depending on the concentration of porphyrin, protein, and salt).
The existence of large, conservative CD signals for porphyrin/protein interactions is not without precedent. Tetrakis(4-carboxyphenyl)porphine, H2TCPP, an anionic porphyrin, has been shown to dimerize moderately at #<0.1 M with a K o < 105M -1 [24]. The interaction of H2TCPP with polylysine and related cationic peptides at relatively high drug load produces induced CD bands in the Soret region having IAel 450 M -1 cm ~ [32]. However, complexes formed by the same polypeptides with bis(4-carboxy- phenyl)porphine, which is extensively aggregated under the conditions used for the experiments, produce CD signals an order of magnitude larger than the most intensive CD spectrum of the H2TCPP complexes [32].
As was observed for the interaction of trans-H2 P~gg with ~-aggregates, the phasing of the conservative CD signal in the Soret region provides information of the helical sense of the template. Just as the CD spectrum in the ultraviolet range is the mirror image reflection for D- and L-forms of the polypeptide, so, too, is the induced CD of the long-range trans-
1000 ::""""...
E
-1000 -
1 i i i i i i i i
380 400 420 440 460 480
;',. (nm)
Fig. 8. Induced CD spectrum of trans-H2Pagg with polyglutamate at pH 4.5 and [NaCI] = 100 mM. (e • • . ) t,-Polyglutamate; ( ) o-polyglutamate.
Porphyrin Assembly 87
100"
~Eo 0 . . . .
400
-200"
I ' ~ ' ~ I J ' ' I
400 450 500 ~. (nrn)
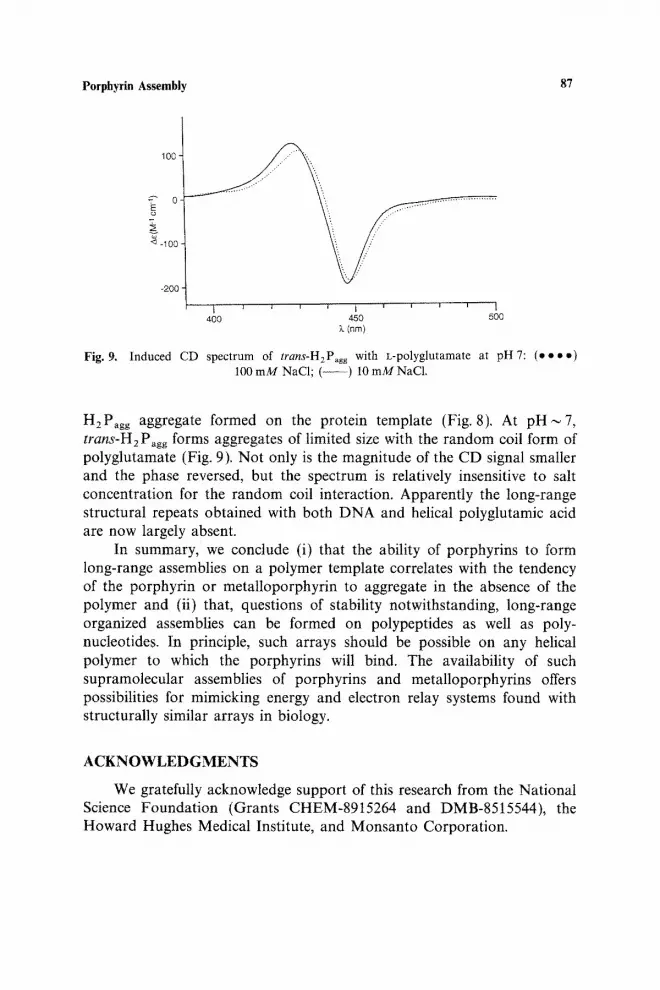
Fig, 9. Induced CD spectrum of trans-H2Pagg with L-polyglutamate at pH 7: (e • • o) 100 m M NaC1; ( - - ) 10 m M NaC1.
H2Pagg aggregate formed on the protein template (Fig. 8). At p H i 7 , trans-H2 Pagg forms aggregates of limited size with the random coil form of polyglutamate (Fig. 9). Not only is the magnitude of the CD signal smaller and the phase reversed, but the spectrum is relatively insensitive to salt concentration for the random coil interaction. Apparently the long-range structural repeats obtained with both DNA and helical polyglutamic acid are now largely absent.
In summary, we conclude (i) that the ability of porphyrins to form long-range assemblies on a polymer template correlates with the tendency of the porphyrin or metatloporphyrin to aggregate in the absence of the polymer and (ii) that, questions of stability notwithstanding, long-range organized assemblies can be formed on polypeptides as well as poly- nucleotides. In principle, such arrays should be possible on any helical polymer to which the porphyrins will bind. The availability of such supramolecular assemblies of porphyrins and metalloporphyrins offers possibilities for mimicking energy and electron relay systems found with structurally similar arrays in biology.
ACKNOWLEDGMENTS
We gratefully acknowledge support of this research from the National Science Foundation (Grants CHEM-8915264 and DMB-8515544), the Howard Hughes Medical Institute, and Monsanto Corporation.

88 Pasternack and Gibbs
R E F E R E N C E S
1. R. F. Pasternack and E. J. Gibbs, A C S Symp. Ser. 402, 59 (1989), and references therein. 2. R. F. Pasternack et al., Biochemistry 22, 2406 (1983). 3. R. F. Pasternack et al., Biochemistry 22, 5409 (1983). 4. R. F. Pasternack et al., Nucleic Acids Res. 14, 5919 (1986). 5. E. J. Gibbs etaL, J. Inorg. Biochem. 32, 39 (1988). 6. (a) R. Lyng etaL, Biopolymers 26, 1327 (1987). (b) M. Kubista etal., J. Phys. Chem. 92,
2352 (1988). 7. N. E. Geacintov et al., Biochemistry 28, 3087 (1987). 8. V. L. Makarov etal., Molekul. Biol. 11, 228 (1977). 9. M. J. Carvlin et aL, Biochem. Biophys. Res. Comm. 108, 66 (1982).
10. L. S. Lerman, U. S. Natl. Acad. Sci. 8, 1886 (1971). 11. C. F. Jordan etal., Nature New Biol. 236, 67 (1972). 12. D. Carroll, Biochemistry 11, 421 (1972). 13. C. Reich et aL, Biochemistry 19, 5208 (1980). 14. M. F. Maestre and C. Reich, Biochemistry 19, 5214 (1980). 15. D. Keller and C. Bustamante, J. Chem. Phys. 84, 2961 (1986). 16. D. Keller and C. Bustamante, J. Chem. Phys. 84, 2972 (1986). 17. M.-H. Kim et aL, J. Chem. Phys. 84, 2981 (1986). 18. Y. A. Shin et aL, Biopolymers 25, 2133 (1986), and references therein. 19. C. L. Phillips et al., Biochemistry 25, 7803 (1986). 20. E. J. Gibbs et al., Biochem Biophys. Res. Commun. 157, 350 (1988). 21. R. F. Pasternack etal., J. Am. Chem. Soc. 113, 7799 (1991). 22. R. F. Pasternack et al., in preparation. 23. R. F. Pasternack et al., Inorg. Chem. 29, 4483 (1990). 24. R. F. Pasternack etal., J. Am. Chem. Soc. 94, 4511 (1972). 25. M.-A. Sari et al., Biochemistry 29, 4215 (1990). 26. R. R. Das etaL, J. Am. Chem. Soc. 92, 3312 (1970). 27. C. R. Cantor and P.R. Schimmel, Biophys Chem. H (W. H. Freeman, San Francisco,
1980), pp. 390-398. 28. B. R. Munson and R. J. Fiel, Nucleic Acids Res. 20, 1315 (1992). 29. L. Stryer and E. R. Blout, J. Am. Chem. Soc. 83, 1411 (1961). 30. S. Ikeda etal., Biopolymers 31, 1257 (1991). (Our attempts to reproduce their results
produced comparably sized but nonconservative signals.) 31. R. F. Pasternack etaL, J. Am. Chem. Soc. 107, 8179 (1985). 32. P. Pancoska etal., Chem. Phys. 147, 401 (1990).
Prmted m Be~ium Verantwoordel~ke uitgever: Hubert Van Mae&. Altenastraat 20 B-8310 St.-Kruis