phylogenetic analysis of antimicrobial lactic acid bacteria from farmed seabass dicentrarchus labrax
TRANSCRIPT

Phylogenetic analysis of antimicrobial lactic acidbacteria from farmed seabass Dicentrarchuslabrax
Ouissal Chahad Bourouni, Monia El Bour, Pilar Calo-Mata, Radhia Mraouna,Boudabous Abedellatif, and Jorge Barros-Velàzquez
Abstract: The use of lactic acid bacteria (LAB) in the prevention or reduction of fish diseases is receiving increasing atten-tion. In the present study, 47 LAB strains were isolated from farmed seabass (Dicentrarchus labrax) and were phenotypi-cally and phylogenetically analysed by 16S rDNA and randomly amplified polymorphic DNA – polymerase chain reaction(RAPD–PCR). Their antimicrobial effect was tested in vitro against a wide variety of pathogenic and spoilage bacteria.Most of the strains isolated were enterococci belonging to the following species: Enterococcus faecium (59%), Enterococcusfaecalis (21%), Enterococcus sanguinicola (4 strains), Enterococcus mundtii (1 strain), Enterococcus pseudoavium (1 strain),and Lactococcus lactis (1 strain). An Aerococcus viridans strain was also isolated. The survey of their antimicrobial suscept-ibility showed that all isolates were sensitive to vancomycin and exhibited resistance to between 4 and 10 other antibioticsrelevant for therapy in human and animal medicine. Different patterns of resistance were noted for skin and intestines iso-lates. More than 69% (32 strains) of the isolates inhibited the growth of the majority of pathogenic and spoilage bacteriatested, including Listeria monocytogenes, Staphylococcus aureus, Aeromonas hydrophila, Aeromonas salmonicida, Vibrioanguillarum, and Carnobacterium sp. To our knowledge, this is the first report of bioactive enterococcal species isolatedfrom seabass that could potentially inhibit the undesirable bacteria found in food systems.
Key words: Enterococcus, lactic acid bacteria, aquaculture, probiotics, seabass.
Résumé : L’utilisation de bactéries lactiques (BL) dans la prévention ou la réduction de maladies chez les poissons suscitede plus en plus d’intérêt. Dans l’étude présente, 47 souches de BL ont été isolées du bar commun d’élevage (Dicentrarchuslabrax) et analysées sur les plans du phénotype et de la phylogénie par l’ADNr 16S et par PCR de l’ADN polymorphiqueamplifié au hasard (RAPD–PCR). Leur effet antimicrobien a été testé in vitro sur une large variété de bactéries pathogèneset de contamination. La plupart des souches isolées consistaient en entérocoques appartenant aux espèces suivantes : Entero-coccus faecium (59 %), Enterococcus faecalis (21 %), Enterococcus sanguinicola (4 souches), Enterococcus mundtii(1 souche), Enterococcus pseudoavium (1 souche) et Lactococcus lactis (1 souche). Une souche d’Aerococcus viridans aaussi été isolée. L‘évaluation de leur susceptibilité aux antimicrobiens a montré que tous les isolats étaient sensibles à lavancomycine et montraient une résistance à entre 4 et 10 autres antibiotiques d’importance thérapeutique en médecine et enmédecine vétérinaire. Différents patrons de résistance ont été notés chez les isolats prélevés de la peau et des intestins. Plusde 69 % (32 souches) des isolats inhibaient la croissance de la majorité des bactéries pathogènes et de contamination testées,dont Listeria monocytogenes, Staphylococcus aureus, Aeromonas hydrophila, Aeromonas salmonicida, Vibrio anguillarumet Carnobacterium sp. À notre connaissance, il s’agit ici du premier rapport portant sur des espèces d’entérocoques bioactifsisolés du bar commun qui pourraient potentiellement inhiber les bactéries indésirables trouvées dans les systèmes alimentaires.
Mots‐clés : Enterococcus, bactéries lactiques, aquaculture, probiotiques, bar commun.
[Traduit par la Rédaction]
Introduction
In aquaculture, prophylactic or therapeutic antibiotic abusehas rapidly led to antibiotic resistance within the bacterialcommunities found in the fish farming areas (Hjelm et al.
2004). However, the introduction of probiotic bacteria in fishfeed has been proposed as an alternative to antimicrobial usefor treatment of farmed fish, shellfish, and penaeid shrimpaquaculture (Irianto and Austin 2002; Riquelme et al. 2001;Ajitha et al. 2004; Farzanfar 2006). Thus, in recent years
Received 11 November 2011. Revision received 10 January 2012. Accepted 11 January 2012. Published at www.nrcresearchpress.com/cjmon 23 March 2012.
O.C. Bourouni, M. El Bour, and R. Mraouna. Institut national des sciences et technologies de la mer (INSTM), rue 2 Mars 1934, 2025Salammbô; Tunis, Tunisia.P. Calo-Mata and J. Barros-Velàzquez. Department of Analytical Chemistry, Nutrition and Food Science, LHICA, School of VeterinarySciences, University of Santiago de Compostela, E-27002 Lugo, Spain.B. Abedellatif. Laboratoire de microorganismes et biomolécules actives, Faculté des Sciences, Tunis, Tunisia.
Corresponding author: Monia El Bour (e-mail: [email protected]).
463
Can. J. Microbiol. 58: 463–474 (2012) doi:10.1139/W2012-014 Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.

there is a noted increase in studies involving isolation of bac-terial strains exhibiting antimicrobial activities (Huys et al.2006). The majority of probiotic bacteria are from the lacticacid bacteria (LAB) group, which includes species belongingto the genera Lactobacillus, Leuconostoc, Pediococcus,Lactococcus, Streptococcus, Aerococcus, Carnobacterium,Enterococcus, Oenococcus, Teragenococcus, Vagococcus,and Weisella (Stiles and Holzapfel 1997; Makras et al. 2006).Recently, LAB that synthesize and secrete Gram-positive
and Gram-negative sensitive bacteriocins were used to im-prove the aquatic environment in the shrimp farming areasand for pathogenic bacteria removal (Røssland et al. 2003;Calo-Mata et al. 2008). Although these LAB are a part ofthe normal intestinal fish flora in Arctic char (Salvelinus alpi-nus), Atlantic salmon (Salmo salar), and marine culturedfish, they do not belong to the aquatic environments (Ringøet al. 1995, 2002; Ringø 2008; Stiles and Holzapfel 1997;González et al. 2000; Mareková et al. 2007; Calo-Mata et al.2008; Vijayabaskar and Somasundaram 2008).Seabass (Dicentrarchus labrax) is the main fish species
cultured around the Mediterranean basin, and it has high eco-nomic value for the aquaculture industry. However, the ef-fects of antibiotic use in aquaculture on marine organisms orin selection of antibiotic-resistant bacteria in the natural eco-system are currently unknown (Sarmah et al. 2006).Microbial infections causing high seabass larva mortalities
has led to abusive antibiotic use and has induced ecologicalproblems in the Tunisian coastal ecosystem (El Bour et al.2001; Bouamama 2001; Dellali 2001). Although it is knownthat LAB intestinal microflora may influence fish growth andhealth and prevent pathogenic infection, there is no study thatshows their use in aquatic farming. In the present study, us-ing 16S rRNA sequencing and randomly amplified polymor-phic DNA – polymerase chain reaction (RAPD–PCR)analysis, we typed the LAB found mainly in the gastrointesti-nal tract of the farmed seabass. To perform the preliminaryprobiotic LAB screening, we established the antimicrobialsusceptibility pattern of each of the strain. In addition, theability of the LAB strains to produce antibacterial compoundsagainst spoiling and pathogenic bacteria was investigated.
Materials and methods
Fish and experimental conditionsFrom February to October 2007, European seabass (Dicen-
trarchus labrax) were sampled from a fish farm in Hergla(Monastir, Tunisia). The fish were sacrificed in a water–icemixture and then kept on ice for 3 h until they reached thelaboratory. Thirty fish specimens weighing 180–220 g wereexamined. Skin patches (2 cm × 1 cm) were aseptically ex-cised, the fish was dissected, and the intestinal content up tothe pyloric caeca was removed. All of the samples wereweighed and homogenized for 1 min in sterile plastic bagsplaced in a Stomacher (Seward, London, UK). The skin andgut homogenates were diluted in 0.9% saline, then 0.1 mL ofthe diluted homogenates was spread on the surface of the deMan – Rogosa Sharpe (MRS) and M17 plates (Oxoid, Ltd.,London, UK). All of the plates were incubated for 48–72 hat 30 °C, and the different colonies were then picked fromeach plate and transferred to MRS or M17 broth (Oxoid) forexperimental use.
Phenotypic characterization of the bacterial strainsAll purified bacterial isolates were subjected to the follow-
ing standard tests: colony morphology, cell morphology, mo-tility, Gram staining, cytochrome oxidase and catalaseproduction, glucose and lactose fermentation tests, H2S gasproduction, and the ability to grow at 10 and 45 °C in media(Brain Heart Infusion) containing 6.5% NaCl at pH 9.6(Schleifer and Kilpper-Bälz 1984). All of the Gram-positiveLAB strains were further tested by miniaturized API 50 CHbiochemical tests (BioMérieux, Marcy L’Etoile, France). Theidentification test data were analysed using the APILABPLUS software, version 4.0 (BioMérieux).
Genetic identification and phylogenetic analysis of LABstrainsAfter overnight culture of the LAB strains in MRS broth,
1 mL of the bacterial cultures was centrifuged at 10 000g for10 min. The resulting pellets were resuspended in 180 µL oflysis buffer (20 mmol/L Tris–Cl, pH 8, 2 mmol/L EDTA,1.2% Triton X-100, 20 mg/mL lysozyme). Ten millilitres oflysis buffer was prepared by mixing 4 mL of lysozyme(10 mg/mL, in double-distilled water), 4 mL of 50 mmol/LTris–HCl, 200 µL of 100 mmol/L EDTA, 120 µL of TritonX-100, and 1.68 mL of Milli-Q water. All of the reagentswere purchased from the Sigma Chemical Co. (St. Louis,Missouri, USA). After 2 h of incubation at 37 °C, 25 µL ofproteinase K (10 mg/mL) (Sigma) was added to the resus-pended pellets. Following 30 min of incubation at 70 °C,DNeasy tissue minikits (QIAGEN Inc., Valencia, California,USA) were used to purify and extract the bacterial DNA. AQuanti-iT kit and Qubit fluorimeter (Invitrogen) were used todetermine the concentration of the purified DNA.The LAB strains were genotypically characterized by a
PCR that targeted the 16S rRNA gene. A universal set of pri-mers was used in the PCR reaction and resulted in a 800 bpproduct: p8FPL (forward: 5′-AGTTTGATCCTGGCTCAG-3′) and p806R (reverse: 5′-GGACTACCAGGGTATCTAAT-3′) (McCabe et al. 1995). The following amplification condi-tions were used: a 7 min denaturing step at 94 °C, followedby 30 denaturation cycles (94 °C for 1 min), annealing(55 °C for 1 min), and extension (72 °C for 1 min), and afinal extension (72 °C for 15 min). All of the amplificationassays contained 100 ng of template DNA, 25 µL of mastermix (BioMix, Bioline Ltd., London, UK), which included re-action buffer, dNTPs, magnesium chloride, Taq DNA poly-merase and PCR water (Genaxis, Montigny le Bretonneaux,France), and 5 µL of each oligonucleotide primer to achieve afinal volume of 50 µL. All of the PCR assays were carried outin a MyCycler Thermal Cycler (BioRad Laboratories, Her-cules, California, USA). PCR products were analysed on 2.5%horizontal agarose (MS-8, Pronadisa, Madrid, Spain) gels.Prior to sequencing, PCR products were purified using an
ExoSAP-IT kit (GE Healthcare, Uppsala, Sweden). The Big-Dye Terminator version 3.1 Cycle Sequencing kit (AppliedBiosystems) and the PCR primers were used in the sequenc-ing reaction for the PCR products. The sequencing reactionswere analysed automatically by the ABI 3730 XL DNA Ana-lyser (Applied Bio systems) with a POP-7 system. Thesingle-nucleotide polymorphisms (SNP) in the DNA sequen-ces were visualized using the Chromas software (GriffithUniversity, Queensland, Australia). The BLAST tool (Na-
464 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.

tional Centre for Biotechnology Information) was used tosearch for sequence homologies. Sequence alignment was ac-complished using the ClustalX software (Larkin et al. 2007).Phylogenetic and molecular evolutionary analyses were con-ducted with the MEGA software (Kumar et al. 2008), usingthe neighbour-joining method (Saitou and Nei 1987) and Ki-mura 2-parameter model with 1000 bootstrap replicates toconstruct distance-based trees.
RAPD-PCR analysisRAPD–PCR was performed using a 200 ng of template
DNA, 25 µL of a master mix (BioMix, Bioline Ltd., London,UK), which contained reaction buffer, dNTPs, magnesiumchloride, Taq DNA polymerase and PCR water (Genaxis,Montigny le Bretonneaux, France), and 14 pmol of M13 (5′-GAGGGTGGCGGTTCT-3′) (Andrighetto et al. 2001) toachieve a final volume of 50 µL. The amplification reactionswere performed using an Applied Biosystems thermal cycler(GeneAmp-PCR System 2700). The following reaction con-ditions were used: an initial 5 min denaturation step at94 °C, followed by 33 cycles of denaturing (94 °C for1 min), annealing (45 °C for 1 min), extension (72 °C for1 min), and final extension (72 °C for 15 min). The PCRproducts (10 µL) were separated and visualized on horizontal1.5% agarose gels (MS-8, Pronadisa, Madrid, Spain) electro-phoresed at 80 V in a 1× TAE buffer solution (Tris–acetate–EDTA) that contained ethidium bromide (10 mg/mL). The123 bp DNA ladder (DNA LADDERS D 5042, Sigma) wasused as a size marker. All of the PCR reactions were per-formed in triplicate to check for reproducibility. For each ofthe reactions, we included a tube without template DNA toserve as a negative control.
Antimicrobial susceptibility testingAntimicrobial agent sensitivity was determined by an agar
diffusion method developed by Chabbert (1982). The follow-ing 16 antibiotics that are relevant in human and animal med-icine were used to test the bacterial strains: vancomycin(30 µg), penicillin G (10 mg), amoxicillin (25 µg), oxacilin(5 µg), cefoxitin (30 µg), ceftriaxone (30 µg), streptomycin(10 mg), tobramycin (10 µg), neomycin (30 mg), chloramphe-nicol (30 µg), tetracycline (30 mg), oleandomycin (15 mg),nitrofurantoin (300 mg), trimethoprim–sulphamide (25 µg),rifampicin (30 µg), and oxolinic acid (30 µg). Overnight cul-tures in MRS broth of the LAB strains (5 mL) were spreadout on the surface of a Mueller–Hinton agar plate (Oxoid).Subsequently, sterile disks were impregnated with the antimicro-bial agents and placed on the plates. After an overnight incuba-tion at 30 °C, the diameter of the zone of inhibition around eachdisk was measured. Based on the zones of inhibition, the straintested was characterized as sensitive, intermediate, or resistantaccording to French national guidelines (Comité de l’Antibio-gramme de la Société Française de Microbiologie 1996).
Antibacterial activityThe potential bacteriocin-producing bacterial strains were
screened against a range of 39 pathogenic and spoilage mi-croorganisms (Table 1). The bacteriocin activity of the LABstrains was initially screened by the standardized agar diskdiffusion test. Briefly, Mueller–Hinton (Oxoid) agar plateswere seeded with a bacterial lawn of each indicator strain at
a 105 CFU/mL concentration. Then, extracellular extractswere prepared by centrifugation, at 7000 r/min (11 000g) for15 min, of 48 h cultures in MRS of each strain, and the cell-free extract was sterilized by filtration through a 0.22 µm fil-ter (Millex GS, Millipore, St. Quentin, France). Twenty mi-crolitres of each LAB strain extracellular extract was placedon the 6 mm sterile disks (Oxoid), which were previouslyplaced on surface of the agar plates. The plates were incu-bated overnight at 37 °C, and then the antimicrobial activitywas detected as translucent halos in the bacterial lawn sur-rounding the disks. A nisin-producing Lactococcus lactisstrain was used as a positive control for antimicrobial activity.
Results
Identification of LAB strainsForty-seven LAB strains were isolated from seabass: 26
strains from the intestinal tract and 21 strains from the skin.All of the isolates were Gram-positive, catalase-negative,nonmotile, nonsporeforming and chain-forming cocci thatwere able to ferment glucose and grow in media containing6.5% NaCl at 10 and 45 °C. All of the strains produced acidfrom glucose, lactose, fructose, mannose, esculin, salicin, mal-tose, and trehalose but not from rhamnose, dulcitol, inuline,xylitol, turanose, lyscose, fructose, arabitol, ceto-gluconate, er-ythriol, xylose, and adonitol. Phenotypic analysis revealedthe species names of 37 strains: 19 strains were Enterococ-cus faecium, 10 strains were Enterococcus faecalis, 4 strainswere Enterococcus sanguinicola, 1 strain was Enterococcusmundtii, 1 strain was Enterococcus pseudoavium, 1 strainwas Lactococcus lactis, and 1 strain was Aerococcus viri-dans. The remaining 10 isolates that could not be identifiedby phenotypic analysis could be identified to the genuslevel as Enterococcus spp.The total DNA of all of the isolates was purified, and a
16S rRNA fragment was PCR amplified and sequenced. Thealignment of the 16S rRNA sequences showed that all of thestrains exhibited a very high homology (≥95%) to other En-terococcus strains deposited in the GenBank. A 16S rRNAgene sequence dendrogram constructed with GenBank data-base reference sequences is shown in Fig. 1. The dendrogramanalysis revealed a close relatedness between the isolates andthe type strains. According to this classification scheme, the10 isolates characterized at the genus level could be assignedto E. faecium (9 strains) and E. sanguinicola (1 strain).
RAPD–PCR analysisUsing the M13 primers, further genetic intraspecific
Enterococcus isolate characterization was performed byRAPD–PCR. The dendrogram derived from the amplificationprofiles of the isolates is shown in Fig. 2. This analysis clas-sified and discriminated the isolated Enterococcus spp. intodifferent clusters for each species.The 3 main clusters that were detected had an arbitrarily
chosen 40% similarity level that was used to define the spe-cies: Cluster 1 grouped isolates belonging to the E. faeciumspp., Cluster 2 grouped isolates belonging to the E. sanguini-cola spp., and Cluster 3 grouped isolates belonging to theE. faecalis spp. In addition, 4 independent strains were de-tected. Interestingly, both the RAPD and the 16S RNA-basedphylogenetic analysis data correlated well with each other.
Bourouni et al. 465
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.

Following the comparison of the amplification profiles ofthe 10 isolates identified at the genus level with those gener-ated for the other strains, it was possible to characterize 9 ofthe isolates as E. faecium and the remaining isolate as E. san-guinicola. These results confirmed the data resulting from thedendrogram generated by the 16S RNA phylogenetic analysis.As shown in Fig. 2, E. faecium isolates were divided into
several RAPD patterns. Three of the isolates were predomi-nant and closely related, and their RAPD patterns were foundin 22 of the 28 E. faecium isolates. These patterns were alsofound in the enterococci isolated from the skin and the intes-tines. The other 6 E. faecium spp. generated 3 different pat-terns with additional amplification bands. An identical RAPD
pattern was observed in all 9 of the E. faecalis strains iso-lated from the skin and the intestine, and only 1 E. faecalisstrain (UPAA3) exhibited a different RAPD pattern. Finally,the E. sanguinicola spp. profiles were quite similar and hadminor differences in their amplification patterns.
Antimicrobial susceptibility testingAll of the strains tested were resistant to 4 or more differ-
ent antimicrobial agents. Bacterial resistance to neomycin,oxacillin, cephalosporins (cefoxitin, ceftriaxon), aminogluco-sids (streptomycin, tobramycin, and neomycin), macrolids(oleandomycin), and oxolinic acid was common (Fig. 3). Incontrast, penicillin, chloramphenicol, tetracycline, rifampicin,
Table 1. Pathogenic and spoilage indicator microorganisms used to test the antibacterial activitiesof lactic acid bacteria isolates.
Code Species OriginAmH01 Aeromonas hydrophila ATCC 7966BaC23 Bacillus cereus ATCC 14893BaP31 Bacillus pumilus ATCC 7061BaS05 Bacillus subtilis subsp. spizizenii ATCC 6633BxT01 Brochotrix thermosphacta ATCC 11509CbD21 Carnobacterium divergens ATCC 35677CbM01 Carnobacterium maltaromaticum LHICA collectionEbA01 Enterobacter aerogenes ATCC 13048EbC11 Enterobacter cloacae ATCC 13047HaA02 Hafnia alvei ATCC 9760KlOx11 Klebsiella oxytoca ATCC 13182KlP02 Klebsiella planticola ATCC 33531KlPn21 Klebsiella pneumoniae subsp. pneumoniae ATCC 10031Lb30A Lactobacillus saerimneri LHICA collectionMoM02 Morganella morganii subsp. morganii ATCC 8076HPhD11 Photobacterium damselae ATCC 33539PrM01 Proteus mirabilis ATCC 14153PrP11 Proteus penneri ATCC 33519PrV21 Proteus vulgaris ATCC 9484PsF12 Pseudomonas fluorescens ATCC 13525PsFr51 Pseudomonas fragi ATCC 4973PsG21 Pseudomonas gessardii LHICA collectionSrM53 Serratia marcescens subsp. marcescens ATCC 274SyE21 Staphylococcus epidermidis ATCC 35983SyX11 Staphylococcus xylosus ATCC 29971StM03 Stenotrophomonas maltophilia ATCC 1363759 Staphylococcus aureus ATCC 91444521 Staphylococcus aureus ATCC 358454032 Listeria monocytogenes NCTC 119941112 Listeria monocytogenes 1112 LHICA collectionCI34.1 Pseudomonas anguilliseptica Seabreama
ACR5.1(AS) Aeromonas salmonicida Turbota
CI52.1(VCI) Vibrio anguillarum Seabreama
ACC30.1 Photobacterium damselae subsp. piscida Solea
V62 Vibrio anguillarum Seabreamb
VF Vibrio anguillarum Seabassc
AF Aeromonas salmonicida Seabassc
V90.11.287(V287) Vibrio anguillarum Seabassd
AH2 Pseudomonas fluorescens Lates niloticusd
aProvided by Professor J.L. Romalde (Spain).bProvided by Professor G. Breuil (France).cProvided by Professor J.C. Raymond (France).dProvided by Professor L. Gram (Denmark).
466 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.

Fig. 1. Phylogenetic relationships according to the partial sequencing of the 16S rRNA gene of lactic acid bacteria (LAB) isolates and refer-ence strains from GenBank by means of the neighbour-joining method. GenBank accession numbers of the LAB isolates are in parentheses.
Bourouni et al. 467
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.

UPAA15 Enterococcus sp. GU460385
UPAA59 Enterococcus GU460407
UPAA105 Enterococcus GU460417
UPAA24 Enterococcus GU460389
UPAA27 Enterococcus GU460391
UPAA30 Enterococcus GU460393
UPAA25 Enterococcus faecium GU460390
UPAA114 Enterococcus faecium GU460422
UPAA58 Enterococcus faecium GU460406
UPAA54 Enterococcus GU460403
UPAA111 Enterococcus GU460420
UPAA64 Enterococcus GU460410
UPAA80 Enterococcus faecium GU460414
UPAA16 Enterococcus faecium GU460386
UPAA108 Enterococcus sp. GU460419
UPAA7 Enterococcus faecium GU460383
UPAA31 Enterococcus sp. GU460391
UPAA32 Enterococcus GU460395
UPAA37 Enterococcus GU460398
UPAA53 Enterococcus GU460402
UPAA83 Enterococcus GU460415
UPAA4 Enterococcus sp. GU460381
UPAA56 Enterococcus sp. GU460404
UPAA63 Enterococcus sp. GU460409
UPAA78 Enterococcus sp. GU460413
UPAA120 Enterococcus sp. GU460425
UPAA88 Enterococcus sp. GU460416
UPAA118 Enterococcus sp. GU460424
UPAA113 Enterococcus sanguinicola GU460421
UPAA116 Enterococcus sanguinicola GU46423
UPAA1 Enterococcus sanguinicola GU460379
UPAA57 Enterococcus sanguinicola GU460405
UPAA33 Enterococcus sp. GU460396
UPAA3 Enterococcus GU460380
UPAA6 Enterococcus GU460382
UPAA10 Enterococcus GU460384
UPAA20 Enterococcus GU460387
UPAA29 Enterococcus GU460392
UPAA36 Enterococcus GU460397
UPAA48 Enterococcus GU460401
UPAA65 Enterococcus GU460411
UPAA107 Enterococcus GU460418
UPAA62 Enterococcus GU460408
UPAA42 Enterococcus mundtii GU460399
UPAA69 Aerococcus viridans GU460412
UPAA47 Lactococcus lactis GU460400
UPAA23 Enterococcus faecium GU460388
1009080706050403020100 1009080706050403020100
1
2
3
faecium
faecium
faecium
faecium
faecium
faecium
faecium
faecium
faecium
faecium
faecium
faecium
faecalis
faecalis
faecalis
faecalis
faecalis
faecalis
faecalis
faecalis
faecalis
faecalis
Fig. 2. RAPD–PCR patterns of the isolates obtained by using the primers M13, and dendrogram obtained by UPGMA of correlation value ofmerged normalized RAPD–PCR patterns.
468 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.
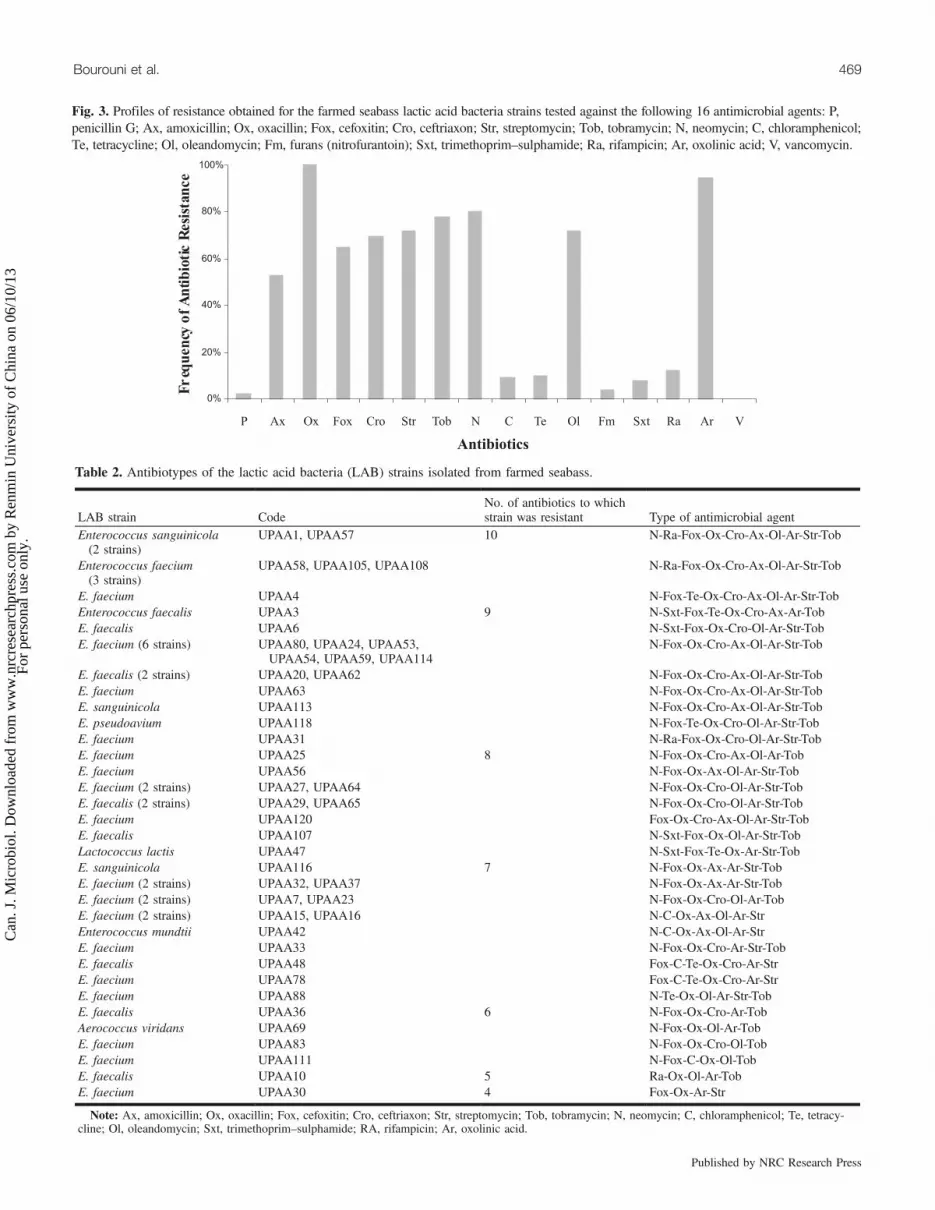
Fig. 3. Profiles of resistance obtained for the farmed seabass lactic acid bacteria strains tested against the following 16 antimicrobial agents: P,penicillin G; Ax, amoxicillin; Ox, oxacillin; Fox, cefoxitin; Cro, ceftriaxon; Str, streptomycin; Tob, tobramycin; N, neomycin; C, chloramphenicol;Te, tetracycline; Ol, oleandomycin; Fm, furans (nitrofurantoin); Sxt, trimethoprim–sulphamide; Ra, rifampicin; Ar, oxolinic acid; V, vancomycin.
Table 2. Antibiotypes of the lactic acid bacteria (LAB) strains isolated from farmed seabass.
LAB strain CodeNo. of antibiotics to whichstrain was resistant Type of antimicrobial agent
Enterococcus sanguinicola(2 strains)
UPAA1, UPAA57 10 N-Ra-Fox-Ox-Cro-Ax-Ol-Ar-Str-Tob
Enterococcus faecium(3 strains)
UPAA58, UPAA105, UPAA108 N-Ra-Fox-Ox-Cro-Ax-Ol-Ar-Str-Tob
E. faecium UPAA4 N-Fox-Te-Ox-Cro-Ax-Ol-Ar-Str-TobEnterococcus faecalis UPAA3 9 N-Sxt-Fox-Te-Ox-Cro-Ax-Ar-TobE. faecalis UPAA6 N-Sxt-Fox-Ox-Cro-Ol-Ar-Str-TobE. faecium (6 strains) UPAA80, UPAA24, UPAA53,
UPAA54, UPAA59, UPAA114N-Fox-Ox-Cro-Ax-Ol-Ar-Str-Tob
E. faecalis (2 strains) UPAA20, UPAA62 N-Fox-Ox-Cro-Ax-Ol-Ar-Str-TobE. faecium UPAA63 N-Fox-Ox-Cro-Ax-Ol-Ar-Str-TobE. sanguinicola UPAA113 N-Fox-Ox-Cro-Ax-Ol-Ar-Str-TobE. pseudoavium UPAA118 N-Fox-Te-Ox-Cro-Ol-Ar-Str-TobE. faecium UPAA31 N-Ra-Fox-Ox-Cro-Ol-Ar-Str-TobE. faecium UPAA25 8 N-Fox-Ox-Cro-Ax-Ol-Ar-TobE. faecium UPAA56 N-Fox-Ox-Ax-Ol-Ar-Str-TobE. faecium (2 strains) UPAA27, UPAA64 N-Fox-Ox-Cro-Ol-Ar-Str-TobE. faecalis (2 strains) UPAA29, UPAA65 N-Fox-Ox-Cro-Ol-Ar-Str-TobE. faecium UPAA120 Fox-Ox-Cro-Ax-Ol-Ar-Str-TobE. faecalis UPAA107 N-Sxt-Fox-Ox-Ol-Ar-Str-TobLactococcus lactis UPAA47 N-Sxt-Fox-Te-Ox-Ar-Str-TobE. sanguinicola UPAA116 7 N-Fox-Ox-Ax-Ar-Str-TobE. faecium (2 strains) UPAA32, UPAA37 N-Fox-Ox-Ax-Ar-Str-TobE. faecium (2 strains) UPAA7, UPAA23 N-Fox-Ox-Cro-Ol-Ar-TobE. faecium (2 strains) UPAA15, UPAA16 N-C-Ox-Ax-Ol-Ar-StrEnterococcus mundtii UPAA42 N-C-Ox-Ax-Ol-Ar-StrE. faecium UPAA33 N-Fox-Ox-Cro-Ar-Str-TobE. faecalis UPAA48 Fox-C-Te-Ox-Cro-Ar-StrE. faecium UPAA78 Fox-C-Te-Ox-Cro-Ar-StrE. faecium UPAA88 N-Te-Ox-Ol-Ar-Str-TobE. faecalis UPAA36 6 N-Fox-Ox-Cro-Ar-TobAerococcus viridans UPAA69 N-Fox-Ox-Ol-Ar-TobE. faecium UPAA83 N-Fox-Ox-Cro-Ol-TobE. faecium UPAA111 N-Fox-C-Ox-Ol-TobE. faecalis UPAA10 5 Ra-Ox-Ol-Ar-TobE. faecium UPAA30 4 Fox-Ox-Ar-Str
Note: Ax, amoxicillin; Ox, oxacillin; Fox, cefoxitin; Cro, ceftriaxon; Str, streptomycin; Tob, tobramycin; N, neomycin; C, chloramphenicol; Te, tetracy-cline; Ol, oleandomycin; Sxt, trimethoprim–sulphamide; RA, rifampicin; Ar, oxolinic acid.
Bourouni et al. 469
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.
trimethoprim–sulphamide, and nitrofurantoin were the mosteffective antibiotics against the majority of the bacterial iso-lates, since the strains showed only 2%, 9%, 10%, 12%, 8%,and 4% resistance to these antibiotics, respectively. Interest-ingly, all of the strains were vancomycin sensitive.The antimicrobial resistance patterns of the enterococci
isolates indicated a considerable diversity of antibiotypes, aswe noted 25 different antibiotypes that exhibited resistanceto 4 to 10 antibiotics (Table 2).The skin isolate antimicrobial resistance patterns frequently
coincided with those patterns revealed for the Enterococcusspp. recovered from the fish intestines. However, 9 skin iso-late antibiotype patterns were not observed in intestinal iso-lates. On the other hand, the other 10 patterns assigned tothe intestinal isolates were not observed in the skin isolates.
Antibacterial activityEnterococcal isolates were assayed for inhibitory activities
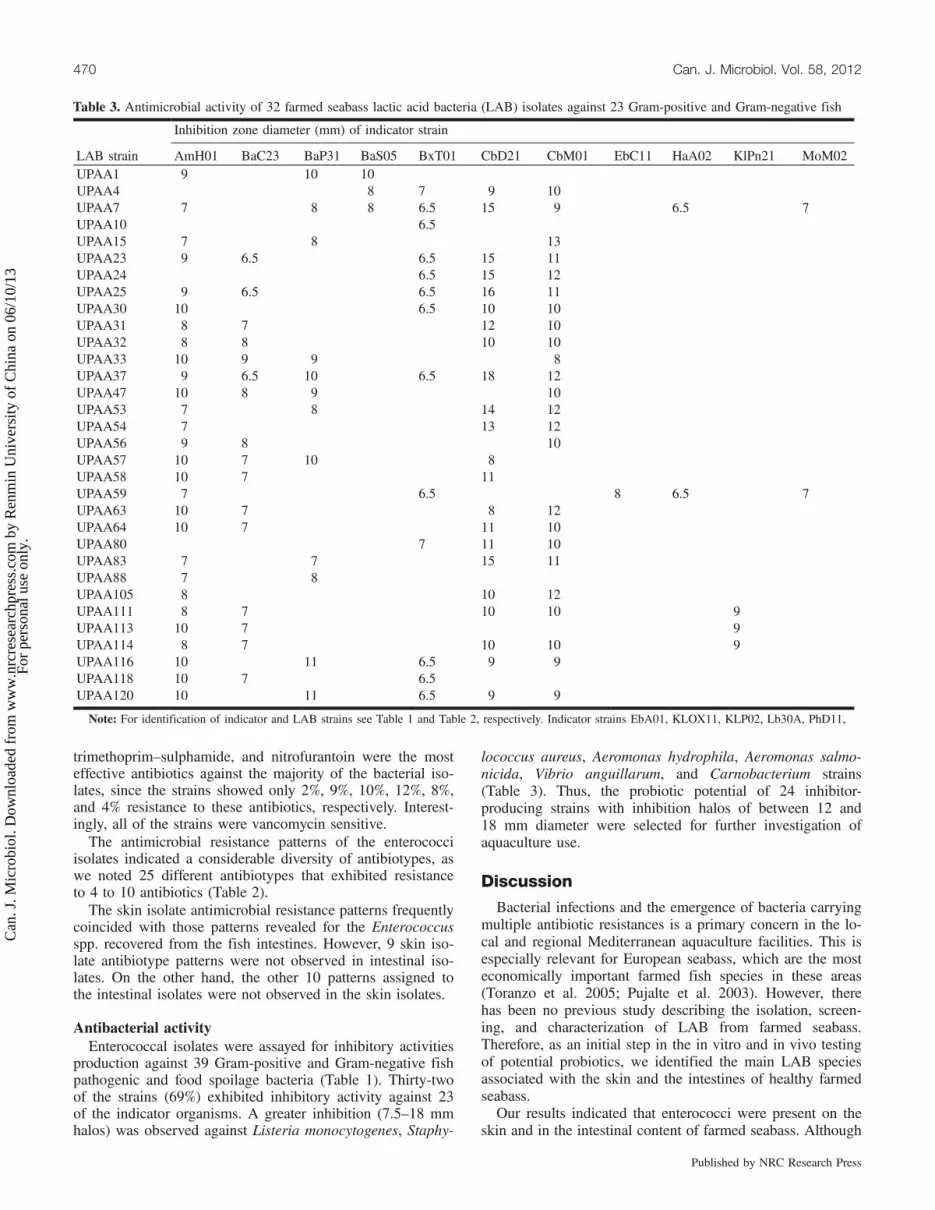
production against 39 Gram-positive and Gram-negative fishpathogenic and food spoilage bacteria (Table 1). Thirty-twoof the strains (69%) exhibited inhibitory activity against 23of the indicator organisms. A greater inhibition (7.5–18 mmhalos) was observed against Listeria monocytogenes, Staphy-
lococcus aureus, Aeromonas hydrophila, Aeromonas salmo-nicida, Vibrio anguillarum, and Carnobacterium strains(Table 3). Thus, the probiotic potential of 24 inhibitor-producing strains with inhibition halos of between 12 and18 mm diameter were selected for further investigation ofaquaculture use.
Discussion
Bacterial infections and the emergence of bacteria carryingmultiple antibiotic resistances is a primary concern in the lo-cal and regional Mediterranean aquaculture facilities. This isespecially relevant for European seabass, which are the mosteconomically important farmed fish species in these areas(Toranzo et al. 2005; Pujalte et al. 2003). However, therehas been no previous study describing the isolation, screen-ing, and characterization of LAB from farmed seabass.Therefore, as an initial step in the in vitro and in vivo testingof potential probiotics, we identified the main LAB speciesassociated with the skin and the intestines of healthy farmedseabass.Our results indicated that enterococci were present on the
skin and in the intestinal content of farmed seabass. Although
470 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Table 3. Antimicrobial activity of 32 farmed seabass lactic acid bacteria (LAB) isolates against 23 Gram-positive and Gram-negative fish
Inhibition zone diameter (mm) of indicator strain
LAB strain AmH01 BaC23 BaP31 BaS05 BxT01 CbD21 CbM01 EbC11 HaA02 KlPn21 MoM02UPAA1 9 10 10UPAA4 8 7 9 10UPAA7 7 8 8 6.5 15 9 6.5 7UPAA10 6.5UPAA15 7 8 13UPAA23 9 6.5 6.5 15 11UPAA24 6.5 15 12UPAA25 9 6.5 6.5 16 11UPAA30 10 6.5 10 10UPAA31 8 7 12 10UPAA32 8 8 10 10UPAA33 10 9 9 8UPAA37 9 6.5 10 6.5 18 12UPAA47 10 8 9 10UPAA53 7 8 14 12UPAA54 7 13 12UPAA56 9 8 10UPAA57 10 7 10 8UPAA58 10 7 11UPAA59 7 6.5 8 6.5 7UPAA63 10 7 8 12UPAA64 10 7 11 10UPAA80 7 11 10UPAA83 7 7 15 11UPAA88 7 8UPAA105 8 10 12UPAA111 8 7 10 10 9UPAA113 10 7 9UPAA114 8 7 10 10 9UPAA116 10 11 6.5 9 9UPAA118 10 7 6.5UPAA120 10 11 6.5 9 9
Note: For identification of indicator and LAB strains see Table 1 and Table 2, respectively. Indicator strains EbA01, KLOX11, KLP02, Lb30A, PhD11,
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.

enterococci are intestinal Gram-positive bacteria that are typ-ically found in mammals and birds (Baele et al. 2002; Linajeet al. 2004), the occurrence of enterococci in fish or fish en-vironments has been mentioned in rare microflora analysesand surveys (Cai et al. 1999; Kanoe and Abe 1988; Petersenand Dalsgaard 2003; Michel et al. 2007). The classicalidentification of enterococcal species requires numerousphysiological tests, and the commercially available kits arefrequently insufficient for an accurate identification(Angeletti et al. 2001). In the present study, phenotypic anal-ysis was complemented with partial 16S rRNA sequencingand phylogenetic analysis as well as with RAPD cluster anal-ysis. Remarkably, a high correlation between the RAPD pro-files and 16S rRNA phylogenetic data was observed. Ourresults were similar to previous reports (Vancanneyt et al.2002; Linaje et al. 2004).The most commonly isolated LAB species from the Euro-
pean seabass in our study was E. faecium (59%), followed byE. faecalis (21%). In addition, E. sanguinicola (4 strains),E. mundtii (1 strain), E. pseudoavium (1 strain), Lactococcuslactis (1 strain), and Aerococcus viridans (1 strain) were alsoisolated. Nevertheless, E. faecalis, E. faecium, and other en-terococci were not previously considered as indigenous flora
of the fish gut (Ringø and Gatesoupe 1998). However, thehigh prevalence of E. faecium and faecalis isolates may indi-cate changes in the bacterial diversities of farmed fish. In ad-dition, Kanoe and Abe (1988) found high counts ofE. faecalis and E. faecium in the intestinal samples of marinefish, and Petersen and Dalsgaard (2003) reported the predom-inance of these species among the enterococci isolated fromintegrated and traditional fish farms. Both studies suggestthat enterococci are normal intestinal microflora of fish.RAPD–PCR proved to be a useful genotypic characteriza-
tion method to detect intraspecific differences among the bac-terial isolates, and the results obtained were in agreementwith the 16S rRNA phylogenetic analysis. In the presentstudy, amplification profiles generated with the primer M13displayed conserved patterns at the intraspecific level. As re-ported by several authors (Andrighetto et al. 2001; Suzzi etal. 2000; Vancanneyt et al. 2002), RAPD–PCR was a validand accurate method for enterococcal identification and de-tecting genetic diversity at the species level.Antimicrobial typing results revealed that Enterococcus
spp. isolated from seabass were vancomycin sensitive andwere resistant to 4 or more of the antimicrobial agents tested.According to previous studies (Bhattacherjee et al. 1988;
pathogenic and food spoilage microorganism indicator strains.
SrM53 SyX11 59 4521 4032 1112 AS AF V62 Vcl V287 VF9 9 12 12 12 9 10
13 12 11 10 9 96.5 6.5 6.5 16 16 10 10
10 8 817 18 11 10 9 11 13 1016 15 11 11 13 10 11 1018 15 10 13 10 10 816 15 11 10 13 11 11 1010 10 11 10 10 11 10 1010 14 10 10 12 10 12 1013 15 9 10 14 11 10 1310 10 8 10 10 10 10 1016 16 9 11 13 10 12 12
10 11 11 10 10 10 10 10 916 17 10 9 10 10 12 1015 16 10 9 13 10 13 1110 9 12 10 12 10 10 10
10 11 11 12 12 10 10 10 1010 11 11 12 10
6.5 6.5 6.510 10 10 12 10 12 1010 11 11 1115 15 10 10 9 916 16 10 9 13 11 11 11
8 9 9 8 915 15 10 10 10
14 10 12 10 11 1010 10 12 12 11 9 910 15 10 11 10 10 109 9 12 12 10 10 10 11
9 9 11 11 10 10 10 11
ACC30.1, CI34.1, PrM01, PsF12, PrP11, Pr21, PsFr51, PsG21, SyE21, and StM03 were not inhibited by any LAB strain.
Bourouni et al. 471
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.

Pathak et al. 1993; Goñi-Urriza et al. 2000; Rhodes et al.2000), the increase in antibiotic use can facilitate the emer-gence of resistant bacteria, which is amplified by the hori-zontal transfer of transposons and plasmids harbouringresistance genes. In the present study, Enterococcus isolatesdemonstrated a variety of antimicrobial resistance characteris-tics, including high resistance to neomycin, oxacillin,cephalosporins (cefoxitin, ceftriaxon), aminoglucosids (strep-tomycin, tobramycin, and neomycin), macrolids (oleandomy-cin), and oxolinic acid. The enterococci group have beenknown to be resistant to most antibiotics used in clinicalpractice. They are naturally resistant to cephalosporins, ami-noglucosids, and clindamycin and may also be resistant totetracyclines and erythromycin (Lukàsova and Sustàckovà2003). Enterococci are known to acquire antibiotic resistancewith relative ease and to be able to spread these resistancegenes to other species (Kühn et al. 2000).Interestingly, all of the isolates characterized in the present
study were vancomycin sensitive, which is relevant in termsof their potential use as probiotics and (or) food additives.Furthermore, enterococci isolates producing inhibitory ac-
tivities were isolated from the skin and intestines of seabass.It has been reported previously that LAB found on skin andin intestines of other fish species can produce antibacterialcompounds that are able to inhibit the growth of severalspoiling or pathogen microorganisms, thereby preventingfish alterations or infections (Ringø 1999, 2008; Spanggaardet al. 2001; Rengpipat et al. 2008; Vijayabaskar andSomasundaram 2008). Enterococcus spp. have a large antimi-crobial spectra that is effective against a broad range ofGram-positive and Gram-negative pathogenic and spoilagemicroorganisms. They are well known to produce variousantibacterial peptides (bacteriocins), ribosomally synthesized,generally called enterocins (Klaenhammer 1993; Strompfováet al. 2008). Previous studies have reported that the inhibitoryspectrum of LAB was restricted to other Gram-positive bac-teria (Abee et al. 1995), although other studies have inferredthat LAB can inhibit Gram-negative fish pathogens (Gildbergand Mikkelsen 1998; Jöborn et al. 1997; Ringø 2008;Robertson et al. 2000). Earlier studies have described severalmarine bacteria that can produce inhibitory substancesagainst pathogens found mainly in aquaculture systems (No-gami and Maeda 1992; Austin et al. 1995; Rengpipat et al.1998; Gram et al. 1999; Campos et al. 2006; Chahad et al.2007). The use of these pathogens for their antimicrobialsubstances has gained importance in fish farming areas as amore efficient antibiotic alternative (Vijayan et al. 2006).Likewise, Spanggaard et al. (2001) reported that use of thesebacteria to prevent the establishment of exogenous bacteriashould offer a significant contribution to the control of unde-sirable pathogenic bacteria.In conclusion, this study highlighted the presence of
enterococcal LAB dominating the skin and intestinal micro-flora of farmed European seabass. These findings demon-strated a variety of antimicrobial resistance characteristics ofthe isolates and several inhibitor-producing strains on theskin and in the intestinal content of farmed seabass. To ex-ploit such enterococcal strains to prevent infections in farm-ing areas, we will continue investigations to elucidate theirpotential probiotic uses.
AcknowledgementsThe authors sincerely thank Dr. Francisco Barros (Unidad
de Medicina Molecular, Fundación Públicia Galega de Me-dicina Xenómica, Santiago de Compostela) for his excellenttechnical assistance with DNA sequencing. We are gratefulalso to all the staff of the Department of Analytical Chemis-try, Nutrition and Food Science, LHICA, School of Veteri-nary Sciences, University of Santiago de Compostela, Lugo,for their technical help, especially Karola B., Inmaculada F.and Arlindo S. The authors are grateful to Dr. Lone Gramfrom the Danish Institute of Piscicultural Research; Dr. J.C.Raymond from Comité national des pêches maritimes et desélevages marins, France; Dr. J. L. Romalde from Departa-mento de Microbiología y Parasitología, Facultad de Biolo-gía, Universidad de Santiago de Compostela, Spain; and Dr.G. Breuil from the IFREMER Palavas, France; for the provi-sion of the microbial strains.
ReferencesAbee, T., Krockel, L., and Hill, C. 1995. Bacteriocins: modes of
action and potentials in food preservation and control of foodpoisoning. Int. J. Food Microbiol. 28(2): 169–185. doi:10.1016/0168-1605(95)00055-0. PMID:8750665.
Ajitha, S., Sridhar, M., Sridhar, N., Singh, I.S.B., and Carghese, V.2004. Probiotic effects of lactic acid bacteria against Vibrioalginolyticus in Penaeus (Fenneropenaeus) Indicus (H. MilneEdwards). Asian Fish. Sci. 17: 71–80.
Andrighetto, C., Knijff, E., Lombardi, A., Torriani, S., Vancanneyt,M., Kersters, K., et al. 2001. Phenotypic and genetic diversity ofenterococci isolated from Italian cheeses. J. Dairy Res. 68(2): 303–316. doi:10.1017/S0022029901004800. PMID:11504393.
Angeletti, S., Lorino, G., Gherardi, G., Battistoni, F., De Cesaris, M.,and Dicuonzo, G. 2001. Routine molecular identification ofenterococci by gene-specific PCR and 16S ribosomal DNAsequencing. J. Clin. Microbiol. 39(2): 794–797. doi:10.1128/JCM.39.2.794-797.2001. PMID:11158155.
Austin, B., Stuckey, L.F., Robertson, P.A.W., Effendi, I., and Griffith,D.R.W. 1995. A probiotic strain of Vibrio alginolyticus effective inreducing diseases caused by Aeromonas salmonicida, Vibrioanguillarum and Vibrio ordalii. J. Fish Dis. 18(1): 93–96. doi:10.1111/j.1365-2761.1995.tb01271.x.
Baele, M., Devriese, L.A., Butaye, P., and Haesebrouck, F. 2002.Composition of enterococcal and streptococcal flora from pigeonintestines. J. Appl. Microbiol. 92(2): 348–351. doi:10.1046/j.1365-2672.2002.01537.x. PMID:11849364.
Bhattacherjee, J., Pathak, S., and Gaur, A. 1988. Antibiotic resistanceand metal tolerance of coliform bacteria isolated from GomatiRiver water at Lucknow city. J. Gen. Appl. Microbiol. 34(5): 391–399. doi:10.2323/jgam.34.391.
Bouamama, K. 2001. Mytilus galloprovincialis de la lagune deBizerte: populations bactériennes et Biomarqueurs non spécifi-ques. Diplôme des études approfondies (DEA). Faculté desSciences de Tunis, Institut des Sciences et Technologies de la Mer.
Bouroni Chahad, O., El Bour, M., Mraouna, R., Abdennaceur, H.,and Boudabous, A. 2007. Preliminary selection study of potentialprobiotic bacteria from aquacultural area in Tunisia. Ann.Microbiol. 57(2): 185–190. doi:10.1007/BF03175205.
Cai, Y., Suyanandana, P., Saman, P., and Benno, Y. 1999.Classification and characterization of lactic acid bacteria isolatedfrom the intestine of common car and freshwater prawns. J. Gen.Appl. Microbiol. 45(4): 177–184. doi:10.2323/jgam.45.177.PMID:12501375.
472 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.

Calo-Mata, P., Arlindo, S., Boehm, K., de Miguel, T., Pascoal, A.,and Barros-Velazquez, J. 2008. Current application and futuretrends of lactic acid bacteria and their bacteriocins for thebiopreservation of aquatic food products. Food BioprocessTechnol. 1(1): 43–63. doi:10.1007/s11947-007-0021-2.
Campos, C.A., Rodríguez, Ó., Calo-Mata, P., Prado, M., and Barros-Velázquez, J. 2006. Preliminary characterization of bacteriocinsfrom Lactococcus lactis, Enterococcus faecium and Enterococcusmundtii strains isolated from turbot (Psetta maxima). Food Res.Int. 39(3): 356–364. doi:10.1016/j.foodres.2005.08.008.
Chabbert, Y.A. 1982. L’antibiogramme. In Bactériologie médicale.Edited by L. Le Miror and M. Véron. Flammarion. MedecineScience, Paris. pp. 205–212.
Comité de l’Antibiogramme de la Société Française de Microbiologie.1996. Statement 1996 CA-SFM. Zone sizes and MIC breakpointsfor non-fastidious organisms. Clin. Microbiol. Infect. 2(Suppl. 1):S46–S49. doi:10.1111/j.1469-0691.1996.tb00875.x.
Dellali, M. 2001. Utilisation d’indicateurs microbiologiques etbiochimiques chez Ruditapes decussatus et Mytilus galloprovin-cialis dans la biosurveillance de la lagune de Bizerte: Validation decertains biomarqueurs. Thèse de doctorat, Faculté des Sciences deBizerte.
El Bour, M., Attia El Hilli, H., Mraouna, R., and Ayari, W. 2001.Bacterial study of mesophilic aeromonads distribution in shellfish.In Proceedings of the 5th International Conference on theMediterranean Coastal Environment, MEDCOAST, pp. 557–565.
Farzanfar, A. 2006. The use of probiotics in shrimp aquaculture.FEMS Immunol. Med. Microbiol. 48(2): 149–158. doi:10.1111/j.1574-695X.2006.00116.x. PMID:17064272.
Gildberg, A., and Mikkelsen, H. 1998. Effects of supplementing thefeed to Atlantic cod (Gadus morhua) fry with lactic acid bacteriaand immuno-stimulating peptides during a challenge trial withVibrio anguillarum. Aquaculture, 167(1–2): 103–113. doi:10.1016/S0044-8486(98)00296-8.
Goñi-Urriza, M., Capdepuy, M., Arpin, C., Raymond, N., Caumette,P., and Quentin, C. 2000. Impact of an urban effluent on antibioticresistance of the riverine Enterobacteriaceae and Aeromonas ssp.Appl. Environ. Microbiol. 66(1): 125–132. doi:10.1128/AEM.66.1.125-132.2000. PMID:10618213.
González, C.J., Encinas, J.P., García-López, M.L., and Otero, A.2000. Characterization and identification of lactic acid bacteriafrom freshwater fishes. Food Microbiol. 17(4): 383–391. doi:10.1006/fmic.1999.0330.
Gram, L., Melchiorsen, J., Spanggaard, B., Huber, I., and Nielsen, T.F. 1999. Inhibition of Vibrio anguillarum by Pseudomonasfluorescens AH2, a possible probiotic treatement of fish. Appl.Environ. Microbiol. 65(3): 969–973. PMID:10049849.
Hjelm, M., Bergh, Ø., Riaza, A., Nielsen, J., Melchiorsen, J., Jensen,S., et al. 2004. Selection and identification of autochthonouspotential probiotic bacteria from turbot larvae (Scophthalmusmaximus) rearing units. Syst. Appl. Microbiol. 27(3): 360–371.doi:10.1078/0723-2020-00256. PMID:15214642.
Huys, G., Vancanneyt, M., D'Haene, K., Vankerckhoven, V.,Goossens, H., and Swings, J. 2006. Accuracy of species identityof commercial bacterial cultures intended for probiotic ornutritional use. Res. Microbiol. 157(9): 803–810. doi:10.1016/j.resmic.2006.06.006. PMID:16919915.
Irianto, A., and Austin, B. 2002. Probiotics in aquaculture. J. FishDis. 25(11): 633–642. doi:10.1046/j.1365-2761.2002.00422.x.
Jöborn, A., Olsson, J.C., Westerdahl, A., Conway, P.L., andKjelleberg, S. 1997. Colonization in the fish intestinal tract andproduction of inhibitory substances in intestinal mucus and faecalextracts by Carnobacterium sp. strain K1. J. Fish Dis. 20(5): 383–392. doi:10.1046/j.1365-2761.1997.00316.x.
Kanoe, M., and Abe, T. 1988. Enterococcal isolates from environ-mental sources. Microbios Lett. 38: 15–20.
Klaenhammer, T.R. 1993. Genetics of bacteriocins produced by lacticacid bacteria. FEMS Microbiol. Rev. 12(1–3): 39–85. PMID:8398217.
Kühn, S., Iversen, A., Burman, L.G., Olsson-Liljequist, B., Franklin,A., Finn, M., et al. 2000. Epidemiology and ecology ofenterococci, with special reference to antibiotic resistant strains,in animals, humans and the environment. Example of an ongoingproject within the European research programme. Int. J.Antimicrob. Agents, 14(4): 337–342. doi:10.1016/S0924-8579(00)00146-1. PMID:10794956.
Kumar, S., Nei, M., Dudley, J., and Tamura, K. 2008. MEGA: abiologist-centric software for evolutionary analysis of DNA andprotein sequences. Brief. Bioinform. 9(4): 299–306. doi:10.1093/bib/bbn017. PMID:18417537.
Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGetti-gan, P.A., McWilliam, H., et al. 2007. Clustal W and Clustal Xversion 2.0. Bioinformatics, 23(21): 2947–2948. doi:10.1093/bioinformatics/btm404. PMID:17846036.
Linaje, R., Coloma, M.D., Pérez-Martínez, G., and Zúñiga, M. 2004.Characterization of faecal enterococci from rabbits for theselection of probiotic strains. J. Appl. Microbiol. 96(4): 761–771. doi:10.1111/j.1365-2672.2004.02191.x. PMID:15012814.
Lukàsova, J., and Sustàckovà, A. 2003. Enterococci and antibioticresistance: review article. Acta Vet. (Brno), 72: 315–323.
Makras, L., Triantafyllou, V., Fayol-Messaoudi, D., Adriany, T.,Zoumpopoulou, G., Tsakalidou, E., et al. 2006. Kinetic analysis ofthe antibacterial activity of probiotic lactobacilli towards Salmo-nella enterica serovar Typhimurium reveals a role for lactic acidand other inhibitory compounds. Res. Microbiol. 157(3): 241–247.doi:10.1016/j.resmic.2005.09.002. PMID:16266797.
Mareková, M., Lauková, A., Skaugen, M., and Nes, I. 2007. Isolationand characterization of a new bacteriocin, termed enterocin M,produced by environmental isolate Enterococcus faecium AL41. J.Ind. Microbiol. Biotechnol. 34(8): 533–537. doi:10.1007/s10295-007-0226-4. PMID:17551760.
McCabe, K.M., Khan, G., Zhang, Y.H., Mason, E.O., and McCabe,E.R.B. 1995. Amplification of bacterial DNA using highlyconserved sequences: automated analysis and potential formolecular triage of sepsis. Pediatrics, 95(2): 165–169. PMID:7838630.
Michel, C., Pelletier, C., Boussaha, M., Douet, D.-G., Lautraite, A.,and Tailliez, P. 2007. Diversity of lactic acid bacteria associatedwith fish and the fish farm environment, established by amplifiedrRNA gene restriction analysis. Appl. Environ. Microbiol. 73(9):2947–2955. doi:10.1128/AEM.01852-06. PMID:17337536.
Nogami, K., and Maeda, M. 1992. Bacteria as biocontrol agents forrearing larvae of the Crab Portunus trituberculatus. Can. J. Fish.Aquat. Sci. 49(11): 2373–2376. doi:10.1139/f92-261.
Pathak, S., Bhattacherjee, J., and Ray, P. 1993. Seasonal variation insurvival and antibiotic resistance among various bacterial popula-tions in a tropical river. J. Gen. Appl. Microbiol. 39(1): 47–56.doi:10.2323/jgam.39.47.
Petersen, A., and Dalsgaard, A. 2003. Species composition andantimicrobial resistance genes of Enterococcus spp., isolated fromintegrated and traditional fish farms in Thailand. Environ.Microbiol. 5(5): 395–402. doi:10.1046/j.1462-2920.2003.00430.x. PMID:12713465.
Pujalte, M.J., Sitjà-Bobadilla, A., Macián, M.C., Belloch, C., Álvarez-Pellitero, P., Pérez-Sanchez, J., et al. 2003. Virulence and moleculartyping of Vibrio harveyi strains isolated from cultured dentex,gilthead sea bream and European sea bass. Syst. Appl. Microbiol.
Bourouni et al. 473
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.

26(2): 284–292. doi:10.1078/072320203322346146. PMID:12866856.
Rengpipat, S., Phianphak, W., Piyatiratitivorakul, S., and Menasveta,P. 1998. Effects of a probiotic bacterium on black tiger shrimpPenaeus monodon survival and growth. Aquaculture, 167(3–4):301–313. doi:10.1016/S0044-8486(98)00305-6.
Rengpipat, S., Rueangruklikhit, T., and Piyatiratitivorakul, S. 2008.Evaluations of lactic acid bacteria as probiotics for juvenileseabass Lates calcarifer. Aquacult. Res. 39(2): 134–143. doi:10.1111/j.1365-2109.2007.01864.x.
Rhodes, G., Huys, G., Swings, J., McGann, P., Hiney, M., Smith, P.,and Pickup, R.W. 2000. Distribution of oxytetracyline resistanceplasmids between Aeromonas in hospital and aquacultureenvironments: Implication of Tn 1721 in dissemination of thetetracycline resistance determinat Tet A. Appl. Environ. Microbiol.66(9): 3883–3890. doi:10.1128/AEM.66.9.3883-3890.2000.PMID:10966404.
Ringø, E. 1999. Lactic acid bacteria in fish: antibacterial effectagainst fish pathogens. In Effects of antinutrients on the nutritionalvalue of legume diets. Vol. 8. Edited by A. Krogdahl, S.D.Mathiesen, and I. Pryme. COST 98. EEC Publication, Luxem-bourg. pp. 70–75.
Ringø, E. 2008. The ability of Carnobacteria isolated from fishintestine to inhibit growth of fish pathogenic bacteria: a screeningstudy. Aquacult. Res. 39(2): 171–180. doi:10.1111/j.1365-2109.2007.01876.x.
Ringø, E., and Gatesoupe, F.J. 1998. Lactic acid bacteria in fish: areview. Aquaculture, 160(3–4): 177–203. doi:10.1016/S0044-8486(97)00299-8.
Ringø, E., Strom, E., and Tabachek, J.A. 1995. Intestinal microfloraof salmonids: a review. Aquacult. Res. 26(10): 773–789. doi:10.1111/j.1365-2109.1995.tb00870.x.
Ringø, E., Seppola, M., Berg, A., Olsen, R.E., Shillinger, U., andHolzapfel, W. 2002. Characterization of Carnobacterium diver-gens strain 6251 isolated from intestine of Artic Charr (Salvelinusalpines L.). Syst. Appl. Microbiol. 25(1): 120–129. doi:10.1078/0723-2020-00080. PMID:12086178.
Riquelme, C.E., Jorquera, M.A., Rojas, A.I., Avendaño, R.E., andReyes, N. 2001. Addition of inhibitor-producing bacteria to masscultures of Argopecten purpuratus larvae (Lamarck, 1819).Aquaculture, 192(2–4): 111–119. doi:10.1016/S0044-8486(00)00461-0.
Robertson, P.A.W., O’Dowd, C., Burells, C., Williams, P., andAustin, B. 2000. Use of Carnobacterium sp. as a probiotic forAtlantic salmon (Salmon solar L.) and rainbow trout (Oncor-hynchus mykiss,Walbaum). Aquaculture, 185(3–4): 235–243.doi:10.1016/S0044-8486(99)00349-X.
Røssland, E., Andersen Borge, G.I., Langsrud, T., and Sørhaug, T.2003. Inhibition of Bacillus cereus by strains of Lactobacillus andLactococcus in milk. Int. J. Food Microbiol. 89(2–3): 205–212.doi:10.1016/S0168-1605(03)00149-1. PMID:14623386.
Saitou, N., and Nei, M. 1987. The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Mol. Biol. Evol. 4(4): 406–425. PMID:3447015.
Sarmah, A.K., Meyer, M.T., and Boxall, A.B. 2006. A globalperspective on the use, sales, exposure pathways, occurrence, fateand effects of veterinary antibiotics (Vas) in the environment.Chemosphere, 65(5): 725–759. doi:10.1016/j.chemosphere.2006.03.026. PMID:16677683.
Schleifer, K., and Kilpper-Bälz, R. 1984. Transfer of Streptococcusfaecalis and Streptococcus faecium to the genus Enterococcusnom. rev. as Enterococcus faecalis comb. nov. and Enterococcusfaecalis comb. nov. Int. J. Syst. Bacteriol. 34(1): 31–34. doi:10.1099/00207713-34-1-31.
Spanggaard, B., Huber, I., Nielsen, J., Sick, E.B., Pipper, C.B.,Martinussen, T., et al. 2001. The probiotic potential againstvibriosis of the indigenous microflora of rainbow trout. Environ.Microbiol. 3(12): 755–765. doi:10.1046/j.1462-2920.2001.00240.x. PMID:11846769.
Stiles, M.E., and Holzapfel, W.H. 1997. Lactic acid bacteria of foodsand their current taxonomy. Int. J. Food Microbiol. 36(1): 1–29.doi:10.1016/S0168-1605(96)01233-0. PMID:9168311.
Strompfová, V., Lauková, A., Simonová, M., and Marciňáková, M.2008. Occurrence of the structural enterocin A, P, B, L50B genesin enterococci of different origins. Vet. Microbiol. 132(3–4): 293–301. doi:10.1016/j.vetmic.2008.05.001. PMID:18547750.
Suzzi, G., Caruso, M., Gardini, F., Lombardi, A., Vannini, L.,Guerzoni, M.E., et al. 2000. A survey of the enterococci isolatedfrom an artisanal Italian goat’scheese (semicotto caprino). J. Appl.Microbiol. 89(2): 267–274. doi:10.1046/j.1365-2672.2000.01120.x. PMID:10971758.
Toranzo, A.E., Magariňos, B., and Romalde, J.L. 2005. A review ofthe main bacterial fish diseases in mariculture systems. Aqua-culture, 246(1–4): 37–61. doi:10.1016/j.aquaculture.2005.01.002.
Vancanneyt, M., Lombardi, A., Andrighetto, C., Knijff, E., Torriani,S., Björkroth, K.J., et al. 2002. Intraspecies genomic groups inEnterococcus faecium and their correlation with origin andpathogenicity. Appl. Environ. Microbiol. 68(3): 1381–1391.doi:10.1128/AEM.68.3.1381-1391.2002. PMID:11872491.
Vijayabaskar, P., and Somasundaram, S.T. 2008. Isolation ofbacteriocin producing lactic acid bacteria from fish gut andprobiotic activity against common fresh water fish pathogenAeromonas hydrophila. Biotechnology, 7(1): 124–128. doi:10.3923/biotech.2008.124.128.
Vijayan, K.K., Bright Singh, I.S., Jayaprakash, N.S., Alavandi, S.V.,Somnath Pai, S., Preetha, R., et al. 2006. A brackishwater isolateof Pseudomonas PS-102, a potential antagonistic bacteriumagainst pathogenic vibrios in penaeid and non-penaeid rearingsystems. Aquaculture, 251(2–4): 192–200. doi:10.1016/j.aquaculture.2005.10.010.
474 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
06/1
0/13
For
pers
onal
use
onl
y.