parvalbumin- and calbindin-containing neurons express c-fos protein in primary and secondary...
TRANSCRIPT

Ž .Brain Research 761 1997 135–145
Research report
Parvalbumin- and calbindin-containing neurons express c-fos protein inŽ . 1,2primary and secondary mirror epileptic foci of the rat neocortex
Andras Mihaly a,), Magdolna Szente b, Zsolt Dubravcsik c, Bernadett Boda d, Erzsebet Kiraly c,Tunde Nagy e, Agota Domonkos e¨
a Department of Anatomy, Faculty of Medicine, UniÕersity of Kuwait, P.O. Box 24923 Safat, 13110 Kuwait, Kuwaitb Department of ComparatiÕe Physiology, Attila Jozsef UniÕersity, Szeged, Hungary
c Department of Anatomy, Albert Szent-Gyorgyi Medical UniÕersity, Szeged, Hungaryd Neurology Clinic, Albert Szent-Gyorgyi Medical UniÕersity, Szeged, Hungary
e Institute of Biochemistry, Biological Research Center of the Hungarian Academy of Science, Szeged, Hungary
Accepted 4 March 1997
Abstract
The present experiments aimed at the description and further immunocytochemical characterization of activated neocortical neuronsexpressing the c-fos gene. Focal seizures were induced by the topical application of isotonic, isohydric 4-aminopyridine solution to thefrontal neocortex of adult anesthetized Wistar rats. The EEG of both hemispheres was recorded from the surface of the skull. The animalswere perfused with fixative, coronal plane vibratome sections were cut and stained with cocktails containing polyclonal c-fos andmonoclonal calbindin or parvalbumin antibodies. The polyclonal c-fos antibody was tested with Western blotting and the diffusion of
w3 x4-aminopyridine investigated with autoradiography of H 4-aminopyridine. The c-fos protein was detected in every layer of theŽ .neocortex primary focus and in some allocortical areas of the treated hemisphere. Scattered immunostained nuclei were observed in
Ž .layers II, III, IV and VI of the contralateral neocortex mirror focus . Several parvalbumin- and calbindin-positive neurons contained thec-fos protein in both foci. The medium-sized non-pyramidal parvalbumin neurons were found in layers II–IV and VI of the neocortex andin stratum multiforme of the prepiriform cortex. The c-fos protein was colocalized with calbindin mainly in layers II and III in small andmedium-sized non-pyramidal neurons. The results prove that focal epileptiform activity of the neocortex activates diverse inhibitory
Ž .neuronal populations. As concluded, the inhibitory control is probably more effective in the contralateral hemisphere mirror focus thanŽ .on the side of 4-APY treatment primary focus . q 1997 Elsevier Science B.V.
Keywords: Immediate-early gene; Parvalbumin; Calbindin; Epilepsy; Neocortex; Rat
1. Introduction
The spread of epileptic activity has serious impacts inhuman neurology since grand mal convulsions are theresult of massive spread of the seizure over the whole
Žcerebrum. In some other seizure types e.g., in focal.rolandic epilepsy the intracortical propagation plays an
important role. The mechanism of spread is thought to be
) Corresponding author. Department of Zoology, JATE University, POBox 659, Szeged H-6701, Hungary.
1 Dedicated to the memory of Prof. Ferenc Joo.2 The results were presented in preliminary form of a congress abstract
at the LXI Cold Spring Harbor Symposium on Quantitative Biology, 29May–5 June 1996.
w xmainly the synaptic propagation 34 . Spread may lead tothe establishment of secondary epileptogenesis which mayappear in the region of callosal terminations of neocortical
w xneurons 32 . This mechanism is known as the mirror focusw x32,34 . The enormous complexity of the human cerebralcortex in terms of the intracortical and cortico-corticalconnections makes the investigation of these phenomenavery difficult. Small, lissencephalic mammals provide goodmodels for the study of the basic events underlying thespread of the seizure. There are literature data which
Ž .indicate the early development of secondary or mirrorfoci in rats subjected to neocortical convulsions through
w xthe local application of chemical convulsants 43,47,48 .These data prove that seizure activity may spread throughcommissural pathways and may induce electrographic
0006-8993r97r$17.00 q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 97 00317-X

( )A. Mihaly et al.rBrain Research 761 1997 135–145136
events in distant locations. There are some debates in theliterature as to the callosal origin of secondary epileptogen-
w xesis 7 . However, the rapid onset of the electrographicevents and the location of the activated cortical areasuggested the significant contribution of commissural ax-
w xons 32 . The other important question related to secondaryepileptogenesis is the characterization of neuronal assem-blies which become activated. There are electrophysiologi-cal data on the contribution of single inhibitory and excita-
w xtory neurons 36,47 . On the other hand, large neuronalpopulations have not been characterized yet in terms oftransmitter content, location and synaptic relationships.Therefore, one aim of the present study was to describe theintracortical location of the activated neuronal populationduring secondary epileptogenesis. The immunohistochemi-cal detection of the nuclear c-fos protein was used as a tool
w xfor the mapping of the activated cells 8 . The other aimwas to characterize some of the activated neurons on thebasis of their calcium-binding protein content. Parvalbu-min and calbindin antibodies were utilized in order toidentify some cells of the network showing neuronal hy-peractivity. There are literature data on parvalbumin- andcalbindin-containing cells of the central nervous systemŽ .CNS , which describe their chemical and functional prop-
w xerties 3,5 . These data indicate that most of the parvalbu-min- and calbindin-positive cells of the cerebral cortex are
w xinhibitory, GABAergic neurons 2,3,5,12,23 . On the basisof these data we suppose that these neurons are exception-ally important in the regulation and control of the seizureprocess.
2. Materials and methods
Experiments were carried out on 20 male Wistar ratsŽ . Ž180–220 g b.wt. , under general anesthesia sodium
.pentobarbital, 45 mgrkg, i.p. , and the animals were placedin a stereotaxic head holder. Their body temperature wasmaintained at 388C with a heating lamp. A 2=3 mm widerectangular craniectomy was made on the right side overly-
w xing the Prc 1 neocortical area 51 . The dura mater wascarefully removed, and the primary epileptic focus wasinduced by applying a piece of the artificial fibrin spongeSpongostan soaked in isotonic, isohydric 20 mM 4-
Ž . w x 2aminopyridine 4-APY solution 29 . Approx. 4 mm ofthe exposed cortical surface was covered with the drug.Silver ball electrodes were placed on the skull on both
w xsides to record the EEG 48 . The EEG was monitored asŽ .long as 1 h from the beginning of the seizure 12 animals .
Following the EEG recording, the electrodes were takenoff, the head of the animal was out from the stereotaxicframe and the head wound was closed. After that, theanimals were kept anesthetized for 1 h, then subjected totranscardial perfusion with cold 4% paraformaldehyde in
Ž .0.1 M phosphate buffer pH 7.4 . The heads were post-fixed for 4–6 h at 48C, then the brains were dissected and
immersed in the fixative for an additional 8–10 h. ShamŽ .operated controls three animals were treated and perfused
similarly, except for the 4-APY treatment: the corticalw xsurface was covered with the solvent of 4-APY 29 .
2.1. Immunohistochemistry
Coronal plane 40 mm thin, serial vibratome sectionsŽwere cut, washed in phosphate buffered saline PBS, pH
.7.4 and incubated according to three different methods:Ž .1. single primary antibody polyclonal anti-c-fos detected
with the peroxidase-anti-peroxidase method, using di-Ž .aminobenzidine tetrahydrochloride DAB chromogen;
Ž .2. primary antibody cocktails two primary antibodiesŽfollowed by fluorescent secondary antibodies anti-rab-
bit IgG-TRITC and anti-mouse IgG-FITC, final dilution.1:100 ;
Ž .3. primary antibody cocktails two primary antibodiesŽ .followed by biotinylated anti-mouse IgG 1:500 and
Ž .plain anti-rabbit IgG 1:50 , detected with streptavidin-Ž .peroxidase 1:800 and peroxidase-anti-peroxidase
Ž .1:500 and DAB and DAB-nickel-ammonium-sulfatew xsubstrate-chromogen solutions 15 ;
4. procedural control of the immunohistochemical stain-ings was performed by omitting the primary antibodiesand incubation with a cocktail containing normal mouseand rabbit sera diluted to 1:2000 and 1:1000, respec-tively.Some sections during method No. 1 were counter-
stained with cresyl violet in order to locate the neocorticallayers. Composite color photographs of the neocortex dis-playing every layer were used for counting and plotting.The cell counts and plottings were made at a final magnifi-cation of 300= . The neurons were counted in a 0.7 mm2
area. The neurons containing c-fos plus calcium-bindingŽ .protein CaBP , c-fos alone and CaBP alone were counted
separately. The ratio of the three different values has beencalculated as percentage. Countings were made from twoexperiments on 4-4 different double-stained sectionsŽ .method No. 3 and the arithmetical means of the individ-ual countings were used. Cell plottings were made manu-ally with the help of transparent paper.
The c-fos antibodies were purchased from Santa CruzŽ .Biotechnology USA , and diluted to 1:1000. Monoclonal
Ž . Ž .parvalbumin PV a235 and calbindin CB a300 antibod-ies were kindly donated by Prof. M. Celio. We dilutedthem to 1:2000. Secondary antibodies and other reagentswere purchased from Sigma, Vector and Dakopatts.
2.2. Western blotting
Ž .Anesthetized animals three rats were decapitated fol-lowing 1 h of registered seizure activity. The brains wereremoved quickly, the two hemispheres separated, andbrainstem and cerebellum thrown away. The tissue wasimmersed in liquid nitrogen, and stored at y808C. Brains

( )A. Mihaly et al.rBrain Research 761 1997 135–145 137
were homogenized, centrifuged, and the pellets subjectedto lipid extraction. The proteins in 10% sodium dodecylsulfate were separated by electrophoresis and transferred tonitrocellulose filters. The c-fos antibody was diluted to
Ž1:500 and the secondary antibody anti-rabbit IgG-per-.oxidase; Sigma to 1:10 000. The binding was visualized
w xwith the enhanced chemiluminescence method 49 .
2.3. Autoradiography
The preparation of tritiated 4-APY has been describedw x Žpreviously in detail 29 . During the experiments two
.animals 20 mmol ‘cold’ 4-APY was dissolved in 0.9%w3 xNaCl and 9 mCi of H 4-APY was added. The specific
activity of the solution was 46.8 mCirmmol. Spongostansoaked in this solution was placed on the exposed pialsurface. After exposure for 60 min the animals wereperfused with 2% paraformaldehyde and 4% glutaralde-
Ž .hyde in 0.1 M phosphate buffer pH 7.4 . Followingperfusion the brains were dissected and postfixed in thesame fixative for 12–24 h. Coronal plane sections 10 mmthick were cut with cryostat and covered with KodakNTB-2 emulsion. After exposure for 60 days, the sectionswere developed in Kodak D-19b solution. Dehydrated,unstained sections were mounted in entellan and examinedin a dark field.
The animal experiments and the treatment of the experi-mental animals in this study comply with the internationalprinciples of laboratory animal care. The experiments werepermitted by the Ethical Committee for the Protection of
ŽAnimals in Research Albert Szent-Gyorgyi Medical Uni-.versity .
3. Results
3.1. Electrophysiology
Soon after the local application of 4-APY the back-ground EEG activity diminished markedly and the first
Žhigh-frequency ictal activity developed in 3–5 min Fig..1 . The epileptic pattern consisted mainly of an initial
Ž .‘tonic’ phase with high frequency 15–20rs repetitivespike discharges followed by a ‘clonic’ phase with repeti-
Ž .tive multiple spike and wave bursts Fig. 1A . The dura-tion of the cortical ictal episodes varied from 30 to 100 s.The amplitude of the afterdischarges increased progres-sively and reached a maximum of 1.5–2.0 mV just beforethe end of the seizure. After six to eight repetitions of thehigh-frequency ictal episodes in the primary focus, an
Ž .epileptic mirror or secondary focus developed in theŽ .contralateral hemisphere Fig. 1B . The amplitudes of
paroxysmal discharges in the mirror focus were usuallylower than in the primary focus, although they showedprogressive increases in successive ictal periods. After15–20 repetitions of bilateral synchronous ictal events the
Fig. 1. The EEG appearance of focal 4-APY seizures in nembutalanesthesia. Traces 1 and 2 represent the contralateral, 3 and 4 the
Ž .ipsilateral stimulated sides. The black rectangles show the location ofthe primary and mirror foci, and the empty circles represent more
Ž .posterior indifferent electrode locations. The seizure begins in theŽ .primary focus A . The enlarged parts illustrate the characteristic pattern
Ž .of firing. The mirror focus is activated some minutes later B and at thisstage both foci are spiking synchronously. Note that the distant indifferentelectrodes do not record abnormal activity. Calibration: 1 s, 1 mV.
paroxysmal activity in primary and mirror focuses showeddissociation in some of the animals, as described earlierw x47 .
3.2. Western blotting
Protein fractions prepared from the primary focus andmirror focus homogenates after 1 h of continuous seizureactivity showed conspicuous immunoprecipitation at 62
Ž .kDa Fig. 2A,B . There was no difference as to the stain-ing intensity between the two hemispheres which indicatedthe widespread early expression of c-fos protein during
Ž . w3 xFig. 2. The results of Western blotting A, B and H 4-APY autoradiog-Ž .raphy C, D . A single protein band was visualized at 62 kDa both in
Ž . Ž .primary A and mirror foci B . The dark-field photographs of theŽ .primary focus C show the diffusion of the tritiated compound in the
Ž .whole depth of the cortex. The mirror focus D is devoid of radioactiv-ity. Roman numerals: cortical layers; bar: 100 mm.

( )A. Mihaly et al.rBrain Research 761 1997 135–145138

( )A. Mihaly et al.rBrain Research 761 1997 135–145 139
4-APY seizures. There were no other bands on the blotw xindicating the specificity of the antibody 31 .
3.3. Autoradiography
w3 xAfter 60 min of H 4-APY exposure, every layer of theŽ .primary focus contained silver grains Fig. 2C,D . The
silver grains were localized in the neuropil and capillariesŽ .not shown suggesting the transendothelial clearance ofthe drug. The labeling was confined to the area underlyingthe craniectomy – no lateral diffusion of the compound
Ž .has been observed not shown . No labeling was found inŽ .the contralateral neocortex Fig. 2D , indicating that the
drug did not diffuse to the area of the mirror focus.
3.4. Primary focus, c-fos labeling
The immunohistochemical experiments were performedon animals sacrificed 2 h after the beginning of the seizure.At this time the serial sections of the primary focusshowed the heavy labeling of layers II–VI. Densely packed,and strongly stained cell nuclei were seen in the investi-
Ž .gated areas of the neocortex Fig. 3A . Layer I contained afew c-fos-stained nuclei. Layers II, III and IV were stainedthe most heavily; seemingly every neuronal nucleus ex-pressed the c-fos protein. Layer V contained scattered, butstrongly stained nuclei and layer VI was somewhat moredensely populated with c-fos containing nuclei than layer
Ž .V Fig. 3A . The interhemispheric surface of Prc and themw x Ž .SmI areas 51 were also stained not shown . The areas
Žsurrounding the rhinal sulcus claustrocortex and regio. Ž .prepiriformis were also stained not shown . In the allo-
Ž . w xcortex layer II stratum densocellulare 44 contained theŽ .heaviest label Fig. 3B . Layer III, the endopiriform nu-
Ž .cleus Fig. 3B and in some animals the amygdaloid nucleicontained scattered fos immunoreactivity. As to the sub-cortical structures, the medial habenular nucleus and thereticular nucleus of the thalamus displayed conspicuous
Ž .c-fos immunoreactivity Mihaly et al., in preparation . Insham operated animals the neocortex contained rarely scat-tered c-fos labeling, which did not show any preference to
Ž .neocortical layers Fig. 3A . Faint nuclear staining wasŽ .also seen in layer II of the prepiriform cortex not shown .
( )3.5. Secondary mirror focus, c-fos labeling
The contralateral neocortical areas contained less stainednuclei, although the labeling was conspicuous and re-stricted to particular layers. Layers II, III and IV displayed
Fig. 4. Composite diagram of the mirror focus showing the density andŽ .location of c-fos-positive nuclei roman numerals indicate layers . Each
dot represents one immunostained nucleus. The arrow points towards theinterhemispheric fissure. Bar: 100 mm.
Ž .several c-fos-stained cell nuclei Fig. 4 . However, thestaining was stronger in layers II and III than in layer IVŽ .Fig. 4 . Layer V was almost empty, showing only one totwo stained nuclei. Layer VI contained more immunoreac-tive cells – this labeling was comparable to that of layer
Ž .IV not shown . The lateral parts of SmI contained lessnuclei, while the claustrocortex contained somewhat more
Ž .c-fos staining not shown . We observed the moderatestaining of layer II neurons in the prepiriform cortex of thecontralateral side. No c-fos staining was seen in the non-stimulated, contralateral neocortical and allocortical areasin sham operated animals.
3.6. ParÕalbumin and calbindin localization in the ratneocortex
Parvalbumin was found in neuronal cell bodies andprocesses in every layer of the rat neocortex. The cellbodies were uniformly distributed in layers II–VI with aslight dominance of layers II–IV. They were mostly largeand medium-sized non-pyramidal neurons. Characteristicneuropil staining was observed in layer V, where parvalbu-min-positive dot-like structures were abundant and sur-
Žrounded unstained neurons presumably pyramidal cells,.Fig. 6A . Dot-like parvalbumin-positive structures were
also seen in layers II–IV. In other layers fine, sometimesbeaded, immunostained fibers were seen, which built up a
Ž .dense network particularly in layer I not shown . Adifferent staining pattern was encountered with the cal-bindin serum. The cell bodies were distributed mainly inlayers II–IV, and they were medium-sized multipolar cellswith long and short dendrites. Long dendrites were some-times beaded in appearance. We observed some pyramidal
Ž .Fig. 3. c-fos immunoreactivity in the cerebral cortex on the side of the 4-APY treatment A, B , and the effect of cranial surgery in a sham operated animalŽ . Ž .C . A: c-fos staining is very strong in most of the layers layers are indicated by roman numerals , except for layer V. B: strong nuclear staining in the
Ž . Ž .allocortex Epd: dorsal endopiriform nucleus; arrowhead: claustrocortex . C: very few c-fos-positive nuclei small arrows are visible in the neocortexunderlying the cranial window. The large arrow points to the interhemispheric fissure, bars: 100 mm.
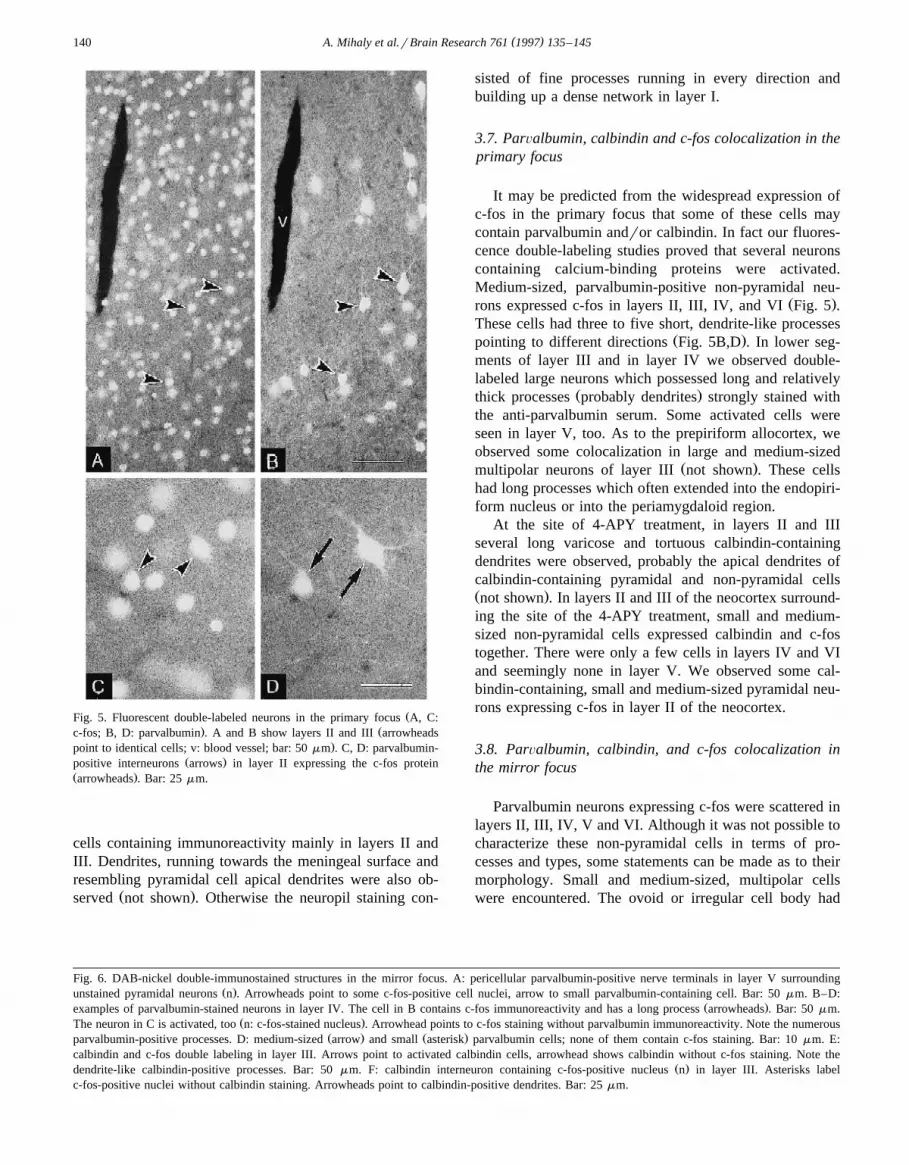
( )A. Mihaly et al.rBrain Research 761 1997 135–145140
ŽFig. 5. Fluorescent double-labeled neurons in the primary focus A, C:. Žc-fos; B, D: parvalbumin . A and B show layers II and III arrowheads
.point to identical cells; v: blood vessel; bar: 50 mm . C, D: parvalbumin-Ž .positive interneurons arrows in layer II expressing the c-fos protein
Ž .arrowheads . Bar: 25 mm.
cells containing immunoreactivity mainly in layers II andIII. Dendrites, running towards the meningeal surface andresembling pyramidal cell apical dendrites were also ob-
Ž .served not shown . Otherwise the neuropil staining con-
sisted of fine processes running in every direction andbuilding up a dense network in layer I.
3.7. ParÕalbumin, calbindin and c-fos colocalization in theprimary focus
It may be predicted from the widespread expression ofc-fos in the primary focus that some of these cells maycontain parvalbumin andror calbindin. In fact our fluores-cence double-labeling studies proved that several neuronscontaining calcium-binding proteins were activated.Medium-sized, parvalbumin-positive non-pyramidal neu-
Ž .rons expressed c-fos in layers II, III, IV, and VI Fig. 5 .These cells had three to five short, dendrite-like processes
Ž .pointing to different directions Fig. 5B,D . In lower seg-ments of layer III and in layer IV we observed double-labeled large neurons which possessed long and relatively
Ž .thick processes probably dendrites strongly stained withthe anti-parvalbumin serum. Some activated cells wereseen in layer V, too. As to the prepiriform allocortex, weobserved some colocalization in large and medium-sized
Ž .multipolar neurons of layer III not shown . These cellshad long processes which often extended into the endopiri-form nucleus or into the periamygdaloid region.
At the site of 4-APY treatment, in layers II and IIIseveral long varicose and tortuous calbindin-containingdendrites were observed, probably the apical dendrites ofcalbindin-containing pyramidal and non-pyramidal cellsŽ .not shown . In layers II and III of the neocortex surround-ing the site of the 4-APY treatment, small and medium-sized non-pyramidal cells expressed calbindin and c-fostogether. There were only a few cells in layers IV and VIand seemingly none in layer V. We observed some cal-bindin-containing, small and medium-sized pyramidal neu-rons expressing c-fos in layer II of the neocortex.
3.8. ParÕalbumin, calbindin, and c-fos colocalization inthe mirror focus
Parvalbumin neurons expressing c-fos were scattered inlayers II, III, IV, V and VI. Although it was not possible tocharacterize these non-pyramidal cells in terms of pro-cesses and types, some statements can be made as to theirmorphology. Small and medium-sized, multipolar cellswere encountered. The ovoid or irregular cell body had
Fig. 6. DAB-nickel double-immunostained structures in the mirror focus. A: pericellular parvalbumin-positive nerve terminals in layer V surroundingŽ .unstained pyramidal neurons n . Arrowheads point to some c-fos-positive cell nuclei, arrow to small parvalbumin-containing cell. Bar: 50 mm. B–D:
Ž .examples of parvalbumin-stained neurons in layer IV. The cell in B contains c-fos immunoreactivity and has a long process arrowheads . Bar: 50 mm.Ž .The neuron in C is activated, too n: c-fos-stained nucleus . Arrowhead points to c-fos staining without parvalbumin immunoreactivity. Note the numerous
Ž . Ž .parvalbumin-positive processes. D: medium-sized arrow and small asterisk parvalbumin cells; none of them contain c-fos staining. Bar: 10 mm. E:calbindin and c-fos double labeling in layer III. Arrows point to activated calbindin cells, arrowhead shows calbindin without c-fos staining. Note the
Ž .dendrite-like calbindin-positive processes. Bar: 50 mm. F: calbindin interneuron containing c-fos-positive nucleus n in layer III. Asterisks labelc-fos-positive nuclei without calbindin staining. Arrowheads point to calbindin-positive dendrites. Bar: 25 mm.

( )A. Mihaly et al.rBrain Research 761 1997 135–145 141

( )A. Mihaly et al.rBrain Research 761 1997 135–145142
two to four short processes which turned into further shortŽ .branches Fig. 6B,C,D . Some of the branches were longer,
Ž .thin and smooth Fig. 6B . Most of the calbindin neuronswhich expressed c-fos were located in layers II and IIIŽ .Fig. 6E . Small and medium-sized double-labeled non-pyramidal neurons and some pyramidal cells were seen in
Ž .layers II and III Fig. 6E,F,G .Counting of the immunostained neurons of the mirror
Ž .focus showed that 12% 325 cells of them containedŽ .parvalbumin and c-fos together, 8% 207 cells contained
Ž .parvalbumin and 80% 2103 cells c-fos alone. Altogether2635 cells were counted. Slightly different figures were
Ž .found with calbindin staining: 22% 726 of the immunos-tained cells contained calbindin and c-fos together, 9%Ž . Ž . Ž297 calbindin and 69% 2260 c-fos alone altogether
.3283 cells .No specific staining was detected in the sections incu-
bated with normal mouse and rabbit sera.
4. Discussion
Our previous experiments indicated that local applica-tion of 4-APY to the neocortex induced characteristicelectrophysiological changes which were comparable toseizures induced by other chemical agents and to clinical
w xconvulsions 29,47,48 . Our previous studies also provedthat, after carefully considering the possibility of artifacts,APY foci could be used for detailed neuropathological
w xstudies 27–29 . The present experiments proved that thefocal seizure spread ipsilaterally to other neocortical areasand the allocortex. This activation, which we called thelateral spread of epileptic activity, resulted in a broad-range and strong c-fos expression in every layer of thecerebral cortex, obviously equally affecting inhibitory andexcitatory neurons.
One remarkable feature of APY convulsions in cats andrats was the development of electrophysiologically de-tectable mirror foci in the homotopic, contralateral neocor-
w xtex 47,48 . These secondary events could be studied forhours and displayed the characteristic electrophysiologicalfeatures of convulsions both at population and cellular
w xlevels 47,48 . The present experiments clearly proved that4-APY seizures can also be used for the morphological
Ž .study of secondary or callosal epileptogenesis. We provedby means of autoradiography that 4-APY did not diffuse tothe contralateral hemisphere; therefore we could concludethat the immunohistochemical changes were the result ofsynaptic activation. The activation of the contralateral,
Žhomotopic neocortical areas the callosal or commissural.spread induced c-fos expression in diverse neuronal popu-
lations, including a considerable number of inhibitory cells.Our immunohistochemical findings on the localization ofc-fos were corroborated by the Western blotting experi-ments, which proved the presence of the c-fos protein inwhole rat brain homogenates. According to literature data
w x31 the 62 kDa band detected by the antibody shouldcorrespond to the c-fos protein. There were no otherprotein bands on the blot, which reflected the purity andspecificity of the polyclonal antibody.
4.1. The lateral spread of epileptic actiÕity
It was very clearly demonstrated in these experimentsthat neocortical and allocortical areas surrounding the pri-
Ž .mary focus the site of drug application are very suscepti-ble to the seizure activity. According to our previousneuropathological studies the local, mild traumatic effectsof the drug application did not involve the surrounding
w xcortical areas 28,29 . On the basis of our light microscopicautoradiography experiments, we can say that the diffusionof 4-APY to these areas should have been negligible, toow x25 . So, we have good reason to suppose that the expres-sion of c-fos in these areas was the consequence of neu-ronal hyperactivity, i.e., the lateral spread of the epilepticseizure.The immunohistochemical results showed that lam-inae II, III and IV contained the highest density of c-foslabeling. This observation corresponds to the electrophysi-ological analysis, which proved that layers IV and III are
w xthe most sensitive to epileptogenesis 9 . In the allocortexlayer II was labeled most heavily which corresponded to
w xprevious data on allocortical c-fos induction 16 . The mostplausible explanation for the pattern of activation is to
w xsuppose an important role of the horizontal plexus 14 .This plexus of intracortical pyramidal cell collaterals origi-
w xnates primarily from layer III neurons 45 , provides con-nections between excitatory and inhibitory cells and oper-
w xates presumably with excitatory amino acids 20 . TheŽphysiological properties of the plexus e.g., the possibility
.of potentiation make it a very powerful device of intracor-w xtical propagation of neuronal activity 14 .
4.2. Callosal or commissural spread
According to literature data, neocortical mirror focidevelop in the area of commissural or callosal connectionsw x32 . The scattered c-fos staining in the contralateral neo-cortex in our experiments could be explained on the basisof the termination pattern of commissural axons, whichdoes not prefer particular layers, but rather dissipates in thewhole depth of columnar slabs of the neocortexw x18,19,35,45 . The distribution of c-fos-stained cell nucleiwas very similar, except for the columnar appearance – thelatter having been obstructed presumably by the laterallyspread excitation of the horizontal plexus. It was also
Ž .demonstrated although not in mirror foci that focal neo-cortical seizures activate several thalamic nuclei and thesenuclei project back to the cortical site, thus creating closed
w xcircuits for the increased neuronal activity 6 . Since thethalamocortical afferents terminate in the close vicinity of
w xthe callosal axons 35 , we cannot exclude the possibilitythat thalamocortical afferents contribute to the activation

( )A. Mihaly et al.rBrain Research 761 1997 135–145 143
pattern of mirror foci. Moreover, it was shown that callosalfibers are necessary, but not sufficient for the triggering of
w xmirror foci 32 .
4.3. The role of inhibitory neurons in actiÕated corticalcircuits
Several lines of evidence suggest that parvalbumin andw xcalbindin are found in GABAergic cells 3,5,22,23 . There-
fore, on the basis of our double-labeling studies we canconclude that inhibitory, GABAergic interneurons play animportant role in the spread of focal convulsions. Suppos-ing that the number of c-fos immunoreactive nuclei re-
w xflected the number of the activated neurons 8,16 , we maysay that much less activation occurred in the mirror focusthan in the primary one, although both of them displayedcharacteristic EEG discharges. One plausible explanationof this is intracortical inhibition. Literature data analyzingthe changes of the inhibition following intrahippocampaltetanus toxin injections point out the existence of normalmonosynaptic inhibition in the sites of secondary epilepto-
w xgenesis 10 . In situ hybridization studies emphasized thatthere was no loss of GABAergic cells in epileptic hip-pocampi; moreover, the amount of glutamate decarboxyl-
w xase mRNA increased in the epileptogenic brain sites 33 .Our previous pharmacological studies pointed out the sig-nificance of inhibitory processes in the spread of acute
w xgeneralized seizures 25,26 . Analysis of the double-labeledcells proved that the seizure activated several differentinhibitory interneurons. Although these studies do notallow to draw final conclusions about the exact nature ofthese cells, some general statements may be done. Acti-vated inhibitory cells were present in layers II–IV and VI
w xwhich are sites of callosal terminations 19 , and thereforethe inhibitory cells could be activated by the incoming
Ž .callosal commissural stimuli. Electrophysiological stud-ies proved that GABA neurons contribute to the horizontal
w xplexus 1 , and also that these GABAergic axons convergew xon pyramidal cells causing inhibition 13 . Probably this
phenomenon could explain the paucity of c-fos-stainednuclei in layer V, where the pyramidal cell bodies are
Žsurrounded by inhibitory axon terminals parvalbumin-positive pericellular baskets, observed in our tissue sec-
.tions . On the basis of these data we may conclude thatcommissural stimulation in the mirror focus not only causedexcitation but also resulted in a feed-forward inhibitionwhich was strong enough to limit the number of theactivated neurons. However, in the primary focus thelateral excitation caused by the horizontal plexus wasmuch stronger, and therefore – although the activation ofinhibitory cells was obvious, too – the effect of the lateral
Ž .inhibition was minimal it was restricted to layer V . Thisconclusion involves the premise that the ratio of excitationand inhibition is very important and that excitation mayovercome easily if the ratio changes. However, the
widespread neuronal activation might also have been theresult of disinhibition.
4.4. The possible role of disinhibition in secondary epilep-togenesis
It is well known that in chronic alumina foci of themonkey the number of GABAergic nerve terminals de-
w xcreases significantly 39 . These studies suggested thatreduced inhibition might be linked to some forms ofepilepsy. On the other hand, immunohistochemical exami-nation of surgically resected human brain samples did notreveal a direct correlation between epilepsy and the de-
w xcrease of inhibitory neuron populations 11 . The roleattributed to disinhibition was also based on physiologicalobservations that reduction or blockade of recurrent androrfeed-forward inhibition mediated through A-type GABAreceptors leads to the appearance of epileptiform activityw x21 . However, studies on human brain tissue obtainedfrom epileptic patients indicated active GABA-mediated
w xinhibitory functions 4,24,41 . In addition, when epilepti-form activity was elicited by manipulations which did notreduce A-type receptor functions, inhibitory postsynaptic
Ž . w xpotentials IPSPs were normal 30,37,40 or even in-w xcreased in duration and amplitude 37,38 . Enhancement of
Žthe long-lasting late component of IPSPs presumably.mediated through B-type GABA receptors were also de-
tected in neurons of APY-induced primary and secondaryw xfoci in cat neocortex, in vivo 46,47 . According to litera-
ture data the convulsant properties of APY result, at leastin part, from the enhancement of both excitatory and
w xinhibitory synaptic transmission 40,46,47 .It is supposed that rebound excitation by long-lasting
hyperpolarizations can facilitate the generation of rhythmicactivity and contribute to the initiation and maintenance of
w xsynchronization in epileptic cell populations 50 . On theother hand an effective inhibition may reflect the existenceof protective mechanisms preventing the induction of ab-normal neuronal activity. The increase in duration andamplitude of the late IPSP may result from the activity of aselected population of inhibitory interneurons responsible
w xfor slow IPSP 17,42 . When the breakdown of inhibitionoccurs, it affects mainly the early, fast IPSP component
w xwhich is mediated through A-type receptors 47 . Thiscould happen only if the fast and slow IPSPs are mediated
w xby different interneuron populations 17,42 .We think that the results of the present study provide
immunohistochemical support for the physiological obser-vation that epileptiform activity could develop without theimpairment of inhibition. We did find two different in-terneuron populations which were activated by the seizureprocess.
Although there is some overlapping between calbindin-w xand parvalbumin-positive GABAergic cells 22 , they cer-
tainly represent different neuronal phenotypes. Furtherstudies are needed to characterize the receptor propertiesof their postsynaptic targets. Therefore, although it has

( )A. Mihaly et al.rBrain Research 761 1997 135–145144
been proven that epileptiform activity was induced by thew xblockade of A-type GABA receptors 21 , the relationship
between disinhibition and epileptogenesis must be evalu-ated cautiously.
Acknowledgements
The authors appreciate the technical help provided byŽ .Dr. Endre Dobo Dept. of Anatomy, Szeged, Hungary .
The radioactive 4-APY was prepared by Dr. Geza Toth,Ž .PhD Biological Research Center, Szeged, Hungary . This
work was supported by OTKA Grants T 017817r1994 andŽ .T 017572 Hungary and a grant from the Kuwait Founda-
Ž .tion for the Advancement of Sciences KFAS 96-07-14 .
References
w x1 K. Albus, P. Wahle, J. Lubke, C. Matute, The contribution ofGABA-ergic neurons to horizontal intrinsic connections in upper
Ž .layers of the cat’s striate cortex, Exp. Brain Res. 85 1991 235–239.w x2 S. Alcantara, I. Ferrer, Postnatal development of parvalbumin im-
munoreactivity in the cerebral cortex of the cat, J. Comp. Neurol.Ž .348 1994 133–149.
w x3 C. Andressen, I. Blumcke, M.R. Celio, Calcium-binding proteins:Ž .selective markers of nerve cells, Cell Tissue Res. 271 1993
181–208.w x4 M. Avoli, A. Olivier, Electrophysiological properties and synaptic
responses in the deep layers of the human epileptogenic neocortexŽ .maintained ‘in vitro’, J. Neurophysiol. 61 1989 589–606.
w x5 M.R. Celio, Calbindin D-28k and parvalbumin in the rat nervousŽ .system, Neuroscience 35 1990 375–475.
w x6 Collins, R.C., Olney, J.W. and Lothman, E.W., Metabolic andpathological consequences of focal seizures. In J.K. Penry and D.
Ž .Purpura Eds. , Epilepsy, Raven Press, New York, 1983, pp. 87–107.w x7 Creutzfeldt, O.D., Cortex Cerebri. Performance, Structural and Func-
tional Organization of the Cortex, Springer-Verlag, Berlin, 1993,615 pp.
w x8 M. Dragunow, R.W. Currie, R.L.M. Faull, H.A. Robertson, K.Jansen, Immediate-early genes, kindling and long-term potentiation,
Ž .Neurosci. Biobehav. Rev. 13 1989 301–313.w x9 J.S. Ebersole, A.B. Chatt, Laminar interactions during neocortical
Ž .epileptogenesis, Brain Res. 298 1984 253–271.w x10 R.M. Empson, J.G. Jefferys, Synaptic inhibition in primary and
secondary chronic epileptic foci induced by intrahippocampal tetanusŽ .toxin in the rat, J. Physiol. 465 1993 595–614.
w x11 I. Ferrer, B. Oliver, A. Russi, R. Casas, R. Rivera, Parvalbumin andcalbindin-D28k immunocytochemistry in human neocortical epilep-
Ž .tic foci, J. Neurol. Sci. 123 1994 18–25.w x12 I. Ferrer, T. Tunon, E. Soriano, A. del Rio, I. Iraizoz, M. Fonseca,
N. Guionnet, Calbindin immunoreactivity in normal human temporalŽ .neocortex, Brain Res. 572 1992 33–41.
w x13 R.J. Gellman, G.K. Aghajanian, Pyramidal cells in piriform cortexreceive a convergence of inputs from monoamine activated
Ž .GABAergic interneurons, Brain Res. 600 1993 63–73.w x14 C.D. Gilbert, Horizontal integration and cortical dynamics, Neuron 9
Ž .1992 1–13.w x15 M.B. Hancock, DAB-nickel substrate for the differential im-
munoperoxidase staining of nerve fibers and fiber terminals, J.Ž .Histochem. Cytochem. 30 1982 578–579.
w x16 T. Herdegen, J. Sandkuhler, P. Gass, M. Kiessling, R. Bravo, M.
Zimmermann, JUN, FOS, KROX, and CREB transcription factorproteins in the rat cortex: basal expression and induction by spread-
Ž .ing depression and epileptic seizures, J. Comp. Neurol. 333 1993271–288.
w x17 J.R. Howe, B. Sutor, W. Zieglgansberger, Characteristics of long-duration inhibitory postsynaptic potentials in rat neocortical neurons
Ž .in vitro, Cell. Mol. Neurobiol. 7 1987 1–18.w x18 S. Jacobson, E.M. Marcus, The laminar distribution of fibers of the
corpus callosum: a comparative study in the rat, cat, rhesus monkeyŽ .and chimpanzee, Brain Res. 24 1970 517–520.
w x19 Jones, E.G., Anatomy, development and physiology of the corpusŽ .callosum. In A.G. Reeves Ed. , Epilepsy and the Corpus Callosum,
Plenum Press, New York, 1985, pp. 3–20.w x20 Z.F. Kisvarday, A. Cowey, A.D. Smith, P. Somogyi, Interlaminar
and lateral excitatory amino acid connections in the striate cortex ofŽ .monkey, J. Neurosci. 9 1989 667–682.
w x21 Krnjevic, K., Significance of GABA in brain function. In G. Tunni-Ž .cliff and B.U. Raess Eds. , GABA Mechanisms in Epilepsy, Wiley,
New York, 1991, pp. 47–48.w x22 Y. Kubota, E.G. Jones, Co-localization of two calcium binding
Ž .proteins in GABA cells of rat piriform cortex, Brain Res. 600 1993339–344.
w x23 Y. Kubota, Y. Kawaguchi, Three classes of GABAergic interneu-Ž .rons in neocortex and neostriatum, Jpn. J. Physiol. 44 Suppl. 2
Ž .1994 145–148.w x24 D.A. McCormick, GABA as an inhibitory neurotransmitter in human
Ž .cerebral cortex, J. Neurophysiol. 62 1989 1018–1027.w x25 Mihaly, A., Morphological Alterations of Cerebral Cortical Struc-
tures in Experimental Aminopyridine Seizures. PhD Thesis, Bu-dapest, 1988, 132 pp.
w x26 A. Mihaly, K. Bencsik, T. Solymosi, Naltrexone potentiates 4-Ž .aminopyridine seizures in the rat, J. Neural Transm. Gen. Sect. 79
Ž .1990 59–67.w x27 A. Mihaly, F. Joo, M. Szente, Neuropathological alterations in the
neocortex of rats subjected to focal aminopyridine seizures, ActaŽ .Neuropathol. 61 1983 85–94.
w x28 A. Mihaly, F. Joo, M. Szente, Vasogenic brain edema in focal4-aminopyridine seizures: the role of neuronal hyperactivity, J.
Ž .Hirnforsch. 31 1990 77–86.w x29 A. Mihaly, G. Toth, M. Szente, F. Joo, Neocortical cytopathology in
focal aminopyridine seizures as related to the intracortical diffusionw 3 x Ž .of H 4-aminopyridine, Acta Neuropathol. 66 1985 145–154.
w x30 I. Mody, J.D. Lambert, U. Heinemann, Low extracellular magne-sium induces epileptiform activity and spreading depression in rat
Ž .hippocampal slices, J. Neurophysiol. 57 1987 869–888.w x31 J.I. Morgan, T. Curran, Stimulus-transcription coupling in the ner-
vous system: involvement of the inducible proto-oncogenes fos andŽ .jun, Annu. Rev. Neurosci. 14 1991 421–451.
w x32 Morrel, F., Callosal mechanisms in epileptogenesis. Identification oftwo distinct kinds of spread of epileptic activity. In A.G. ReevesŽ .Ed. , Epilepsy and the Corpus Callosum. Plenum Press, New York,1985, pp. 99–130.
w x33 A. Najlerahim, S.F. Williams, R.C. Pearson, J.G. Jefferys, Increasedexpression of GAD mRNA during the chronic epileptic syndrome
Ž .due to intrahippocampal tetanus toxin, Exp. Brain Res. 90 1992332–342.
w x34 Niedermeyer, E., The Epilepsies. Diagnosis and Management, Urbanand Schwarzenberg, Munich, 1990, pp. 9–34.
w x35 J. Olavarria, R.C. Van Sluyters, H.P. Killackey, Evidence for thecomplementary organization of callosal and thalamic connections
Ž .within rat somatosensory cortex, Brain Res. 291 1984 364–368.w x36 M.W. Oliver, J.J. Miller, Inhibitory processes of hippocampal CA1
pyramidal neurons following kindling-induced epilepsy in the rat,Ž .Can. J. Physiol. Pharmacol. 63 1985 872–878.
w x37 P. Perreault, A. Avoli, Effects of low concentrations of 4-amino-pyridine on CA1 pyramidal cells of the hippocampus, J. Neurophys-
Ž .iol. 61 1989 953–970.

( )A. Mihaly et al.rBrain Research 761 1997 135–145 145
w x38 P. Perreault, A. Avoli, Physiology and pharmacology of epileptiformactivity induced by 4-aminopyridine in rat hippocampal slices, J.
Ž .Neurophysiol. 65 1991 771–785.w x39 C.E. Ribak, B.A. Harris, J.E. Vaughn, E. Roberts, Inhibitory,
GABAergic nerve terminals decrease at sites of focal epilepsy,Ž .Science 205 1979 211–214.
w x40 P.A. Rutecki, F.J. Lebeda, D. Johnston, 4-Aminopyridine producesepileptiform activity in hippocampus and enhances synaptic excita-
Ž .tion and inhibition, J. Neurophysiol. 57 1987 1911–1924.w x41 P.A. Schwartzkroin, M.M. Hugland, Spontaneous rhythmic syn-
chronous activity in epileptic human and normal monkey temporalŽ .lobe, Epilepsia 27 1986 523–533.
w x42 M. Segal, A subset of local interneurons generate slow inhibitorypostsynaptic potentials in hippocampal neurons, Brain Res. 511Ž .1990 163–164.
w x43 I. Sherwin, Ictal-interictal unit firing pattern differences in peni-cillin-induced primary and secondary epileptogenic foci, Exp. Neu-
Ž .rol. 84 1984 463–477.w x44 Stephan, H., Allocortex, Springer-Verlag, Berlin, 1975, 998 pp.w x45 J. Szentagothai, The Ferrier Lecture, 1977. The neuron network of
the cerebral cortex: a functional interpretation, Proc. R. Soc. Lond.Ž .B 201 1978 219–248.
w x46 M. Szente, A. Baranyi, Mechanism of aminopyridine-induced ictalŽ .seizure activity in the cat neocortex, Brain Res. 41 1987 373–386.
w x47 M. Szente, B. Boda, Cellular mechanisms of neocortical secondaryŽ .epileptogenesis, Brain Res. 648 1994 203–214.
w x48 M. Szente, F. Pongracz, Comparative study of aminopyridine-in-duced seizure activities in primary and mirror foci of cat’s cortex,
Ž .Electroencephalogr. Clin. Neurophysiol. 52 1981 353–367.w x49 Vieira, A., Elkin, R.G. and Kuchler K., Western blotting and ligand
blotting using enhanced chemiluminescence and radioiodine detec-Ž .tion. In J.E. Celis Ed. , Cell Biology, A Laboratory Handbook, Vol.
2, Academic Press, San Diego, CA, 1994, pp. 314–321.w x50 J.X. Wang, J. Rinzel, Spindle rhythmicity in the reticular thalami
nucleus: synchronization among mutually inhibitory neurons, Neuro-Ž .science 53 1993 899–904.
w x51 K. Zilles, B. Zilles, A. Schleicher, A quantitative approach toc y t o a r c h i t e c t o n i c s .VI. The areal pattern of the cortex of the albino rat, Anat. Embryol.
Ž .159 1980 335–360.