hippocalcin in rat retina. comparison with calbindin-d28k, calretinin and neurocalcin
TRANSCRIPT

Exp: Eye Res. (1995) 60, 257-266
Hippocalcin in Rat Retina. Comparison with Calbindin-D28k, Calretinin and Neurocalcin
E N R I C O B A S T I A N E L L I " , K E N T A K A M A T S U b , K A T S U O O K A Z A K I c , H I R O Y O S H I H I D A K A c AND R O L A N D P O C H E T a*
a Laboratoires d'Anatomie Pathologique et dd'Histologie, Facultd de Mddecine, Universitd Libre de Bruxelles, 808 route de Lennik, B-1070 Bruxelles, Belgium, b Department of Physiology, Toho University
School of Medicine, Ohmori-nishi, Ohta-ku, Tokyo 143, Japan and C Department of Pharmacology, Nagoya University School of Medicine, Tsurumai 65, Showa-ku. Nagoya 466, Japan
(Received Houston 4 August 1994 and accepted 31 October 1994)
The post-natal developmental expression in rat retina of four calcium-binding proteins belonging to the calmodulin-troponin-C family was investigated by immunohistochemistry using anti-calbindin-D28k, anti-calretinin, anti-hippocalcin and anti-neurocalcin polyclonal antibodies on paraffin sections from Wistar rat retinae aged from post-natal days 1 (P1), 5 (P5), 10 (PIO), 20 (P20) to adulthood (8 weeks). Immunoblot using anti-hippocalcin and homogenates proteins from retina, cerebellar cortex, hippocampus and cerebellum was also performed.
Hippocalcin immunoreactivity in adult rat retina was demonstrated by both immunohistochemistry and Western blot. During post-natal development, calbindin-D28k, calretinin and neurocalcin immunoreactivity were detected at P1 in ganglion cells, whereas hippocalcin immunoreactivity was seen later at P5 in this ceil layer. In the amacrine cell layer, neurocalcin immunoreactivity was detected at P5 and hippocalcin at PIO. Calbindin-D28k was labelling the immature horizontal cell, calretinin was detected in nearly all ganglion cells and in some amacrine cells since P1. These three calcium-binding proteins do not seem to play a role in synaptogenesis which takes place later. We confirmed that calbindin-D28k appeared to be a good marker for horizontal cells. The presence of hippocalcin, a myristoylated calcium-binding protein belonging to the recoverin subfamily and previously localized in few brain areas has been detected for the first time in retina.
Key words: calcium; calcium-binding proteins; amacrine cells; ganglion cells; horizontal cells.
1. Introduction
In rats, at birth, the retina is immature and will continue to develop for at least 3 weeks. Retinal neurons will differentiate and move inwards from the neuroblastic layer and first form the ganglion cells (Braekevelt et al. 1970). During the first 2 weeks, two phases of cell death occurs within the ganglion cell layer (Horsburgh et al. 1987). Synaptogenesis in the inner plexiform layer occurs on day 11 for amacrine cells and on day 13 for bipolar cells. Equally dramatic changes in functional properties occur during this period. For instance, the distribution of subclasses of amacrine cells undergoes a series of changes within the first 3 weeks (Zhang et al. 1990), during the first 2 weeks horizontal cells exhibit transient properties like acetylcholinesterase activity (Spira, 1974), ab- normal membrane permeability (Sarthy et al. 1983) or GABA expression (Versaux-Botteri et al. 1989).
Calcium ions which play a critical role in neural development (Kater et al. 1988; Cambray-Deakin et al. 1992) are probably involved at different stages of rat retina development. Intracellular calcium-binding proteins such as calbindin-D28k and calretinin may
* For correspondence.
0014 4835/95/030257+ 10 $08.00/0
play a role in this calcium control, although a precise physiological function, besides binding Ca z÷, has not yet been demonstrated. Calbindin-D28k and calretinin are expressed in retinal cells from different species (Rohrenbeck et al. 1987, 1989; Pasteels et al. 1990; Pochet et al. 1991; Hamano et al. 1990; Ellis et al. 1991; Uesugi et al. 1992). In rat, previous survey have shown that calbindin-D28k was expressed in horizontal cells (Rabi6 et al. 1985; Hamano et al. 1990; Pasteels et al. 1990; Chu et al. 1993) as early as embryonic day 17 (Uesugi et al. 1992) as well as in some amacrine and ganglion cells (Pasteels et al. 1990; Hamano et al. 1990; Uesugi et al. 1992) but not in photoreceptors or bipolar cells. Calretinin has not been detected in rat horizontal cells, only in amacrine and ganglion cells (Pasteels et al. 1990). Recently, two other calcium-binding proteins, hippo- calcin and neurocalcin [probably identical to CBP-18 (Lipp et al. 1993)], belonging to the recoverin subfamily have been discovered (Hidaka et al. 1993; Kobayashi et al. 1992). Neurocalcin has been localized in amacrine and ganglion cells (Nakano et al. 1992) whereas hippocalcin was found in hippocampus (Kobayashi et al. 1992), in the Purkinje cells of the cerebellum and moderately in the pyramidal cells of cerebral cortex (Saitoh et al. 1993). They both possess
@ 1995 Academic Press Limited

258 E. BASTIANELLI ET AL.
(A)
(B)
(c)
(D)
@
p i , , ,~
(E)
~:~ (H)
FIG. 1. Immunohistochemical controls used with rat retina sections from post-natal day 5 (P5) or post-natal day 20 (P20). (A) P20 with anti-calbindin-D28k; (B) P20 with anti-calbindin-D28k preincubated for 24 hr with calbindin (2-5/zg ml-1); (C) P5 with anti-calretinin; (D) P5 with anti-calretinin preincubated for 24 hr with calretinin (2'5/~g ml-]); (E) P5 with anti- neurocalcin; (F) P5 with anti-neurocalcin preincubated for 24 hr with neurocalcin (4/zg ml-1); (G) P20 with anti-hippocalcin and (H) P20 with anti-hippocalcin preincubated 24 hr with hippocalcin (4/~g ml 1). Scale bar = 20/~m.
a myristoylation consensus sequence. Their functions are still unknown but might be similar to that of recoverin. Insights into the ontogeny of calcium homeostasis can be gained by investigating the developmental expression of calcium-binding proteins. Considering the potential important calcium regu- latory activities of calbindin-D28k, calretinin, neuro- calcin and hippocalcin as well as their neuronal marker characteristics, we decided to look at their distribution in the retina both during post-natal development and adulthood an hopefully better under- stand their physiological importance.
2. Materials and Methods
Tissues
Experiments were performed on Wistar rats from post-natal day 1 (P1), 5 (P5), 10 (P10), 20 (P20) to adulthood (8 weeks). Rats were killed by an overdose of ether or sodium pentobarbitural. Two or more rat retinae were examined at each age point. For immunocytochemical studies, eyes were dissected out from rat head, retinae were removed and fixed for 4 hr in Helly's fixative (68 mM potassium dichromate, 70 mM sodium sulfate, 180 mM HgCL, and 4% for-

HIPPOCALCIN IN RETINA 259
maldehyde) rapidly after death. Fixed tissues were routinely dehydrated, embedded in paraffin, and sectioned at 7/zm.
Specificity of the Primary Antisera
Rabbit antiserum raised against chick duodenal calbindin-D28k was prepared as described (Spencer et al. 1976) and routinely used at 1/6000 dilution. The specificity of anti-calbindin-D28k has been described (Pasteels et al. 1990; Pochet et al. 1991). Rabbit anti- calretinin was obtained from Swant (Belinzona, Switzerland), previously characterized (Schwaller et al. 1993) and used at 1/1000 dilution. Rabbit polyclonal anti-hippocalcin antibodies were raised against the maltose-binding protein-hippocalcin fusion protein (Kobayashi et al. 1992) and used at 1/2000 dilution. Rabbit polyclonal anti-neurocalcin antibodies were previously characterized (Nakano et al. 1992) and used at 1 / 1000 dilution. The specificities of the antisera were also checked by absorption tests. Structures were not stained with antiserum pre- absorbed by its specific purified antigen. Anti- calbindin-D28k was preabsorbed with purified calbindin-D28k from rat cerebellum (Pasteels et al. 1987), anti-calretinin with purified calretinin from human recombinant calretinin expressed in E. coli (Schwaller et al. 1993), anti-hippocalcin with purified rat recombinant hippocalcin (LysHip) expressed in Escherichia coli (Kobayashi et al. 1993) and anti- neurocalcin by purified neurocalcin from bovine brain (see Fig. 1).
Immunohistochemical Staining and Immunoblot
Routinely dewaxed and hydrated sections were processed for immunohistochemistry according to a peroxidase-antiperoxidase (PAP) procedure modified from Vacca et al. (1980). Serum dilutions were made up in Coons Veronal Buffered Saline (CVBS: 10 mM 5,5-diethylbarbituric acid sodium salt (sodium ver- onal), pH 7-2, 150 mM NaC1) supplemented with 1% (v/v) normal sheep serum. The immunostaining sequence comprised the following steps: (1) rinse in CVBS; (2) preincubation in 5% (v/v) normal sheep serum; (3) incubation with rabbit polyclonal anti- bodies for 48 hr at 4°C in a moist chamber; (4) incubation (15 min) with sheep anti-rabbit immuno- globulin G (IgG) serum (1:100 dilution) (Laboratoire d'Hormonologie et Immunologie, Marloie, Belgium); (5) incubation (15 min) with soluble rabbit PAP complex 1:300 dilution (DAKO, Glostrup, Denmark). Between each step, sections were thoroughly rinsed for 10 min in CVBS. After the last rinse, staining was performed in 32 mM citrate phosphate buffer, pH 6.2, containing 16 mM 3,3'-diaminobenzidine-HC1 (DAB; Sigma, St Louis, MO, U.S.A.) and 0.01% H202. Each of these steps was performed at room temperature, except for step 3 which was performed at 4°C. For immuno-
blot analysis, rat retinae, cerebral cortex, hippocampus and cerebellum were homogenized in lOVol, of 20 mM Tris-HC1, pH 7"4, 2 mM BGTA, 1 mM DTT, 0"2 mM PMSF. The homogenates (100 Fg protein per row) were separated on 14% SDS-PAGB, and then subjected to immunostaining using anti-hippocalcin as described previously'(Takamatsu et al., 1992). MW markers were obtained from Biorad (Richmond, CA, U.S.A.). The relative area of each band was calculated using a Shimadzu TLC-Scanner.
3. Results
Figures are organized by age: Fig. 2 corresponds to post-natal day 1, Fig. 3 to post-natal day 5, Fig. 4 to post-natal day 10 and Fig. 5 to post-natal day 20.
Calbindin-D28k: [Figs 2(A), 3(A), 4 and 5(A)]
At post-natal day 1, in the neuroblastic layer, only calbindin-D28k immunoreactivity could be detected, in elongated cells [Fig. 2(A)]. They were localized on a single layer, in the outer third of the neuroblast layer, along the whole retina. They showed a spindle cytoplasm with two thin processes expanding across the neuroblastic layer from the pigment epithelial layer to the inner nuclear layer. Calbindin-D28k immunoreactivity was also detected in few amacrine cells of medium size with a round shaped cytoplasm from which rare thin processes expanded toward the inner plexiform layer. They were disposed on one row. Strong immunoreactivity was found in ganglion cells. As for amacrine cells, their number was smaller than the calretimn ones with a labelling intensity more variable. Ganglion ceils, distributed in two or three layers, were large with round cytoplasm. Immuno- labelled dendrites were ramifying inside the inner plexiform layer contrasting with the immnnonegative axons [Fig. 2(A)]. At post-natal day 5, identifiable horizontal cells were disposed on a single layer. Their oval cytoplasm and their thin processes running horizontally were calbindin-D28k immunoreactive [Fig. 3(A)]. the number of amacrine and ganglion cells positive for calbindin-D28k increased. Ganglion cell axons began to be immunoreactive [Fig. 3(A)]. Five days later, calbindin-D28k immunoreactivity could be detected in almost all horizontal ceils. They were disposed on a continuous, single regular row forming a thick network with their heavily labelled processes. Both their oval cytoplasm and nucleus were immuno- labelled. Amacrine cells displayed the same pattern of immunoreactivity as for P5, but processes were thicker and distributed more regularly in the IPL, beginning to segregate into layers [Fig. 4(A)]. Ganglion cells processes were easily identifiable at this time. At post- natal day 20, the calbindin-D28k immunoreactivity distribution was similar to adulthood. Almost all horizontal cells were strongly calbindin-D28k
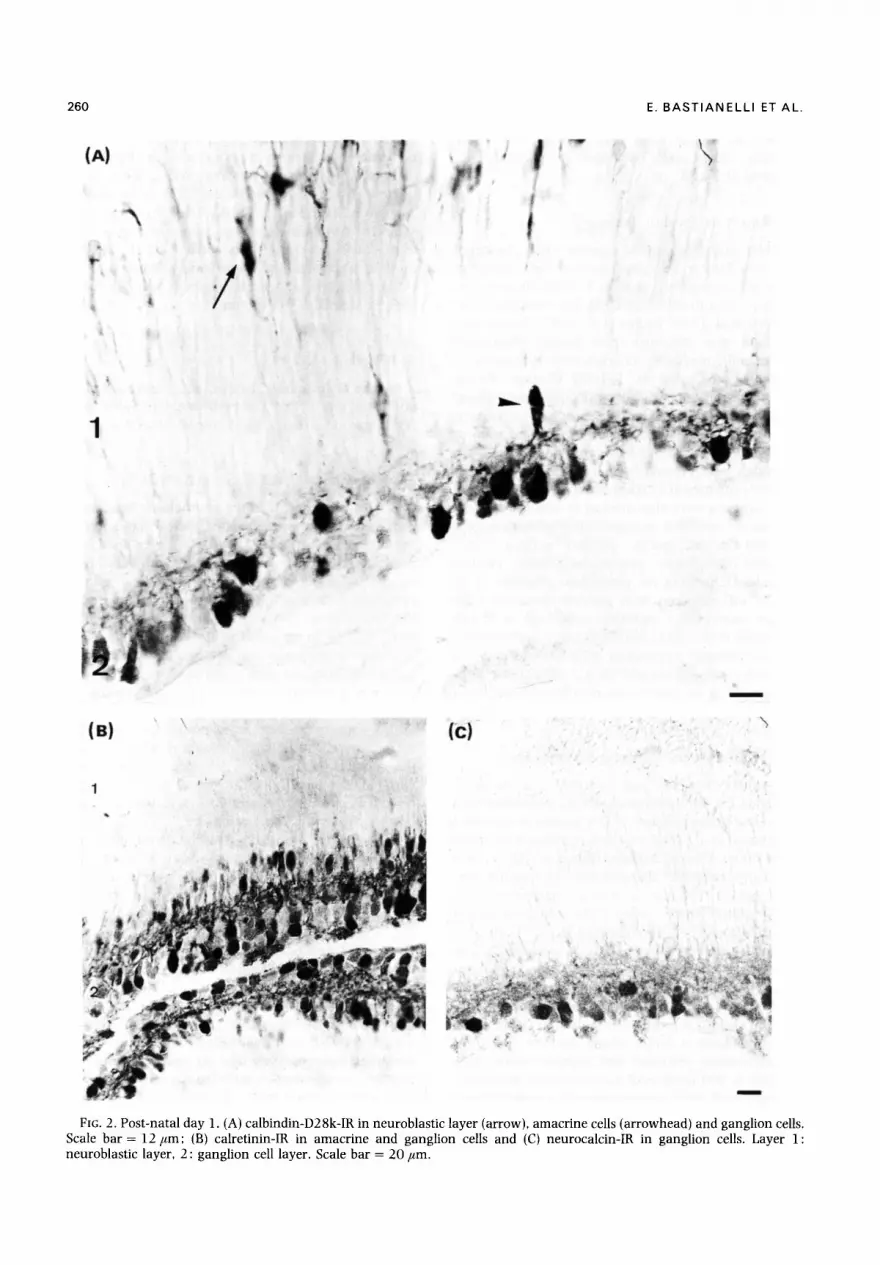
260 E. B A S T I A N E L L I ET AL.
(A)
1
i i ' ~
%
(u) (C:)
m
\
FIG. 2. Post-natal day l . (A) calbindin-D28k-IR in neuroblastic layer (arrow), amacrine ceils (arrowhead) and ganglion cells. Scale bar = 12/ ,m; (B) calretinin-IR in amacrine and ganglion cells and (C) neurocalcin-IR in ganglion cells. Layer 1: neuroblastic layer, 2 : ganglion cell layer. Scale bar = 20 #m.
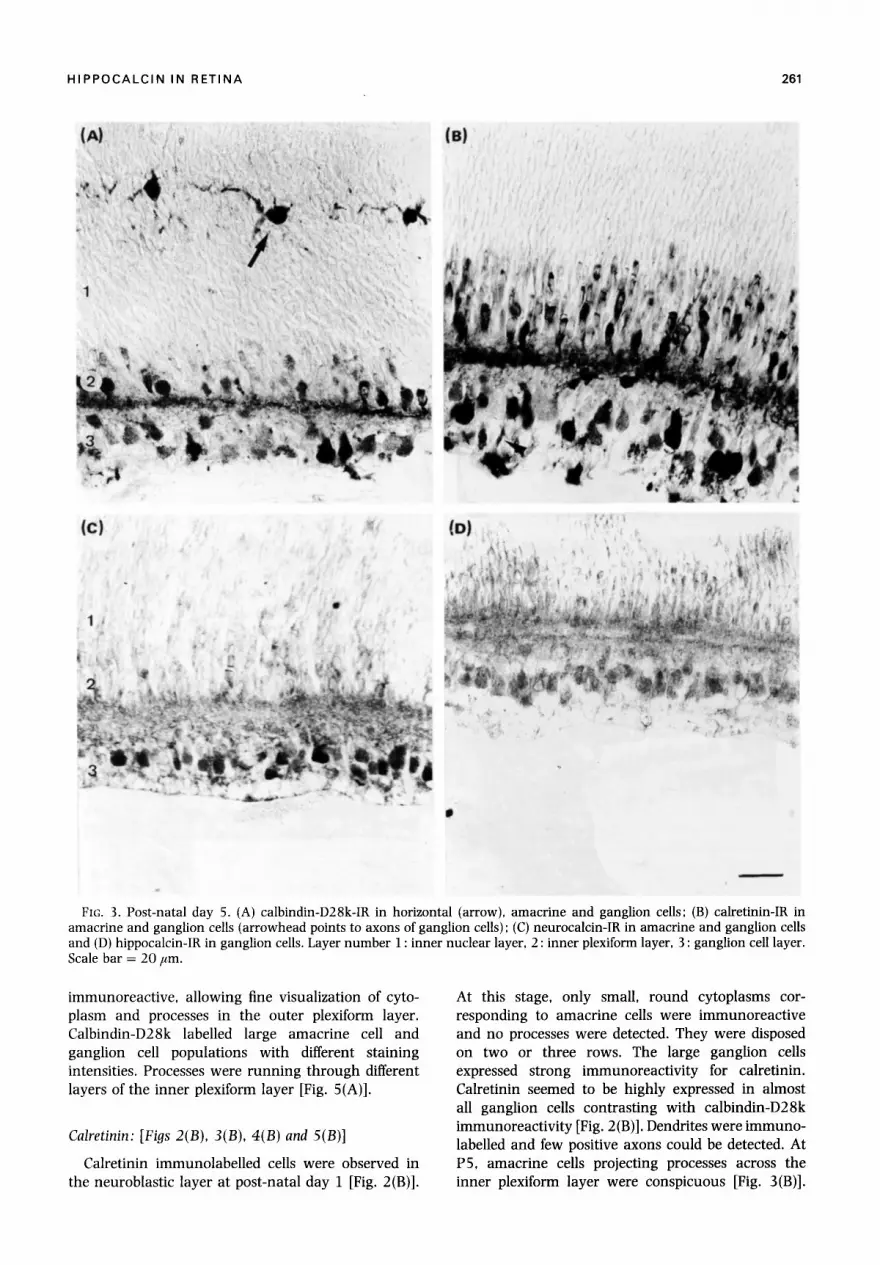
HIPPOCALCIN IN RETINA 261
(A) ' (B)
.,,
(c) ID|
0
FIG. 3. Post-natal day 5. (A) calbindin-D28k-IR in horizontal (arrow), amacrine and ganglion cells; (B) calretinin-IR in amacrine and ganglion cells (arrowhead points to axons of ganglion cells) ; (C) neurocalcin-IR in amacrine and ganglion cells and (D) hippocalcin-IR in ganglion cells. Layer number 1 : inner nuclear layer, 2 : inner plexiform layer, 3 : ganglion cell layer. Scale bar = 20 Fm.
immunoreactive, allowing fine visualization of cyto- plasm and processes in the outer plexiform layer. Calbindin-D28k labelled large amacrine cell and ganglion cell populations with different staining intensities. Processes were running through different layers of the inner plexiform layer [Fig. 5(A)].
Calretinin: [Figs 2(B), 3(B), 4(B) and 5(B)]
Calretinin immunolabelled cells were observed in the neuroblastic layer at post-natal day 1 [Fig. 2(B)].
At this stage, only small, round cytoplasms cor- responding to amacrine cells were immunoreact ive and no processes were detected. They were disposed on two or three rows. The large ganglion cells expressed strong immunoreactivi ty for calretinin. Calretinin seemed to be highly expressed in almost all ganglion cells contrasting with calbindin-D28k immunoreactivity [Fig. 2 (B)]. Dendrites were immuno- labelled and few positive axons could be detected. At P5, amacrine cells projecting processes across the inner plexiform layer were conspicuous [Fig. 3(B)].
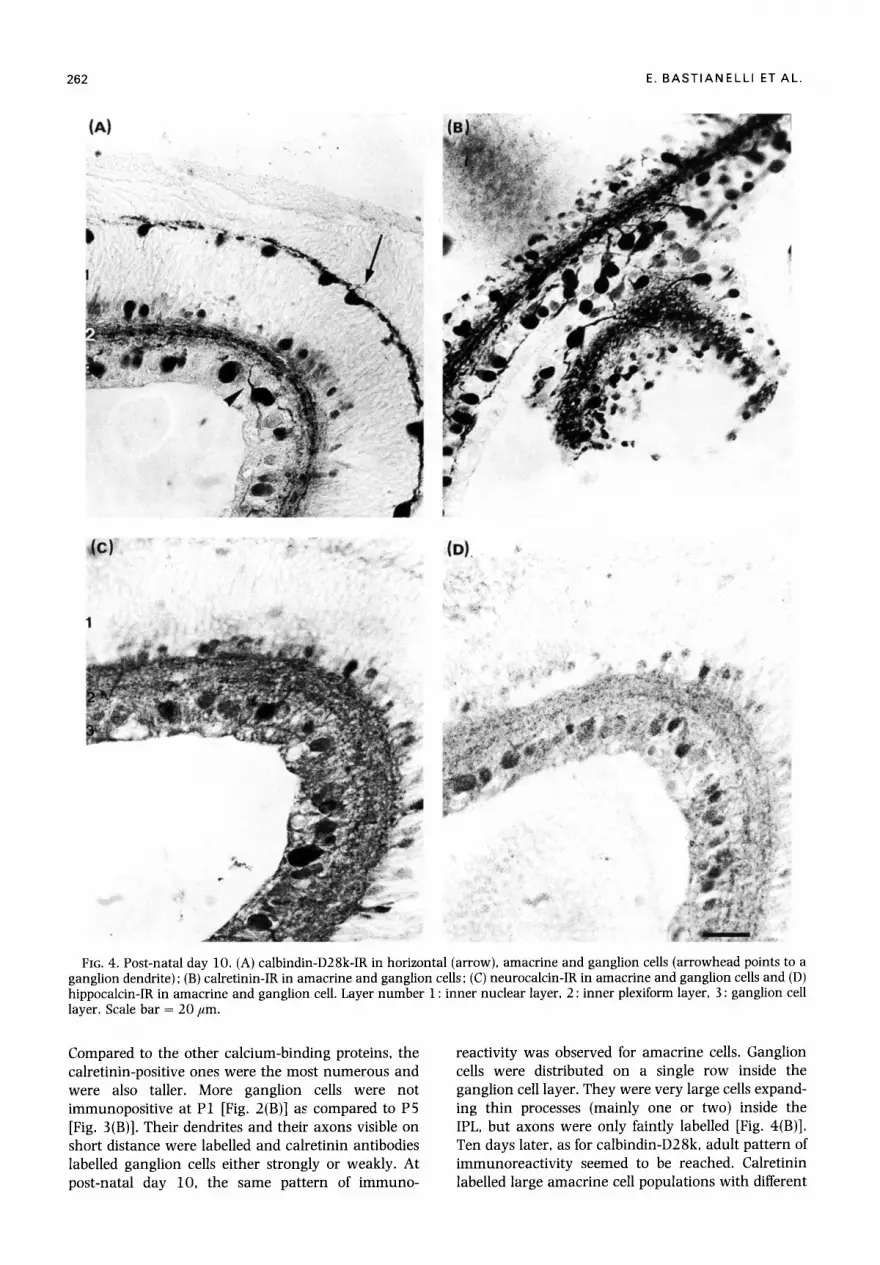
262 E. BASTIANELLI ETAL.
(A)
, . L
, ,
/ ,
r
~ ¢ ) : . . . . • ,~,~o~: ' ~ ~ - ~ . . . . . . ( D )
FIG. 4. Post-natal day 10. (A) calbindin-D28k-IR in horizontal (arrow), amacrine and ganglion cells (arrowhead points to a ganglion dendrite) ; (B) calretinin-IR in amacrine and ganglion cells; (C) neurocalcin-IR in amacrine and ganglion cells and (D) hippocalcin-IR in amacrine and ganglion cell. Layer number 1 : inner nuclear layer, 2 : inner plexiform layer, 3 : ganglion cell layer. Scale bar = 20/~m.
Compared to the other calcium-binding proteins, the calretinin-positive ones were the most numerous and were also taller. More ganglion cells were not immunopositive at P1 [Fig. 2(B)] as compared to P5 [Fig. 3(B)]. Their dendrites and their axons visible on short distance were labelled and calretinin antibodies labelled ganglion cells either strongly or weakly. At post-natal day 10, the same pattern of immuno-
reactivity was observed for amacrine cells. Ganglion cells were distributed on a single row inside the ganglion cell layer. They were very large cells expand- ing thin processes (mainly one or two) inside the IPL, but axons were only faintly labelled [Fig. 4(B)]. Ten days later, as for calbindin-D28k, adult pattern of immunoreactivi ty seemed to be reached. Calretinin labelled large amacrine cell populations with different

HIPPOCALCIN IN RETINA 263
l a l
I P
(el
! /' ,.
\
~ (D)
FIG. 5. Post-natal day 20. (A) calbindin-D28k-IR in horizontal, amacrine and ganglion ceils; (B) calretinin-IR in amacrine, inner plexiform layer and ganglion cells; (C) neurocalcin-IR in amacrine and ganglion cells and (D) hippocalcin-IR in amacrine and ganglion cells. Layer number 1 : inner nuclear layer, 2 : inner plexiform layer, 3 : ganglion cell layer. Scale bar = 20/zm.
staining intensities. Processes were running through different layers of the inner plexiform layer, three conspicuous calretinin-immunoreactive layers were detected [Fig. 5(B)] in accordance with previous work (Pasteels et al., 1990). Numerous ganglion cells were immunoreactive, especially large cells. Compared to the three other calcium-binding proteins investigated calretinin was very abundant in the cells as indicated
by the labelling intensity [Fig. 5(B)]. Both axons and dendrites were from this age immunopositive.
Neurocalcin: [Figs 2(C), 3(C), 4(C) and 5(C)]
Some large ganglion cells were found to be neurocalcin immunoreactive at post-natal day 1 [Fig. 2(C)]. Weak immunoreactivity in the inner plexiform

264 E, B A S T I A N E L L I ETAL.
1 2 3 4 ~
FIG. 6. Hippocalcin immunoblot: 100/zg of proteins from different tissue homogenates were loaded in each row. Lane 1: retina; lane 2: cerebral cortex; lane 3: hippocampus; lane 4: cerebellum. MW markers are bovine serum albumin (66200), hen egg white ovalbumin (45000), bovine carbonic anhydrase (31000), soybean trypsin inhibitor (21 500) and hen egg white lysosyme (14400).
layer could be already detected. At post-natal day 5, rare small round-shaped amacrine cells were immuno- reactive, positive processes were seen [Fig. 3(C)]. More ganglion cells do not appear to be labelled than at P1, they were either strongly or weakly labelled as for calbindin-D28k and calretinin [Fig. 3(C)]. Dendrites and axons do not appear labelled. At P10, small amacrine cells were strongly immunopositive, disposed on a single row with processes running inside the IPL. Large positive ganglion cells were detected with variable intensity. Only their dendrites were stained [Fig. 4(C)]. At P20, neurocalcin immuno- reactivity seemed to be restricted to a smaller cell population of amacrine cells. They were rare, localized on a single row near the inner plexiform layer in which no immunoreactive processes could be detected [Fig. 5(C)]. Ganglion cells were also of small number, but of very large size as observed for calretinin. Processes were only faintly visible.
Hippocalcin: [Figs 3(D), 4(D), 5(D)]
No immunoreactivity was detected at post-natal day 1. At post-natal day 5, rare amacrine cells, very
weakly labelled and positive ganglion cells were detected [Fig. 3(D)]. More strongly stained amacrine cells were seen five days later. They had small size and distributed only in a single layer. A differential staining among layers could be seen in the inner plexiform layer [Fig. 4(D)]. Immunopositive ganglion cells were of smaller size compared to the ganglion cells positive for the three other calcium-binding proteins investi- gated. [Fig. 4(D)]. No labelled processes could be seen. Although hippocalcin immunoreactivity seemed to be restricted to a small cell population, the number of positive cells was higher than for neurocalcin. Processes were easily visible inside the inner plexiform layer in which they discriminate in distinct layers. At P20, a large number of small ganglion cells were immunoreactive for hippocalcin. They were only weakly labelled. Their axons were immunopositive [Fig. 5(D)].
Western Blot
Figure 6 demonstrated that in rat retina homo- genate, anti-hippocalcin recognized a protein comi- grating with hippocalcin. The relative amount of hippocalcin in retina was estimated by measuring the area of the immunoreactive band. The relative protein concentration of hippocalcin in rat retina was only 1 / lO th of that in hippocampus. Cerebral cortex containing 2-4 times less hippocalcin than hippo- campus and cerebellum 3.4 times less.
4. Discussion
This study set out to describe throughout rat retinal postnatal development the cellular localization and expression of four EF-hand calcium-binding proteins namely calbindin-D28k, calretinin, hippocalcin and neurocalcin.
Although the rat retina is immature at birth, ganglion cells which differentiated at around em- bryonic day 15 form a layer made up of several rows of cells (Braekevelt et al. 1970; Raedler et al. 1975). In this study we have shown that (1) at birth ganglion cells already expressed calbindin-D28k, calretinin and neurocalcin: (2) hippocalcin could be detected from P5 in ganglion cells and from PIO in amacrine cells: (3) once expressed their presence will be maintained throughout development and during adulthood. In the neuroblastic layer (the inner ventricular zone) calbindin-D28k was detected in elongated cells with sparse perikaryal cytoplasm cell that extends into long, very slender processes. The location of their nucleus between the outer and the middle third of the ventricular zone correspond according to Raedler and Sievers (Raedler et al., 1975) to the first recognizable horizontal cell precursors. This result is in agreement with the recent work (Uesugi et al., 1992) showing that already at embryonic day 17 calbindin-D28k was present in the outer third of neuroblastic layer.

HIPPOCALCIN IN RETINA 265
Furthermore horizontal cells have consistently been labelled by calbindin-D28k in many species (Rohrenbeck et al., 1987; Rabid et al., 1985; Rohrenbeck et al., 1989 ; Vesaux-Botteri et al., 1989 ; Pochet et al., 1991; Hamano et al., 1990; Pasteels et al., 1990) including human (Verstappen et al., 1986). The presence of calbindin-D28k in immature hori- zontal cell provides a good marker for studies of the organization of horizontal cells during development and in pathology (Chu et al., 1993).
Calbindin-D28k and calretinin both binds calcium, but besides this physico-chemical properties little is known about its physiological roles. Some work has raised the possibility that calbindin-D28k may play a role in synaptogenesis during developing ch ick retina (Ellis et al., 1991) and developing rat hippocampus (Rami et al., 1987). Our results on amacrine cells do not support this hypothesis indeed, calbindin-D28k and calretinin were detected as soon as P1 whereas in rat, amacrine cells synaptogenesis does not occur before P l l (Horsburgh et al., 1987). Our results fit better with other work showing little correlation between calbindin-D28k and synaptogenesis in rat brain development (Enderlin et al., 1987), in mouse inner ear (Dechesne et al., 1988), in cat (Stichel et al., 1987) and monkey visual cortex (Hendrickson et al., 1991).
Neurocalcin and hippocalcin belongs to the recoverin subfamily (Terasawa et al., 1992 ; Kobayashi et al., 1992). Recoverin has been first purified from rod bovine retina (Dizhoor et al., 1991) and has been localized in bovine, mouse (McGinnis et al., 1992) and human (Wiechmann et al., 1993) cones and rods. Some cone bipolar cells from human, monkey and rat were also expressing recoverin (Milam et al., 1993). It seems that recoverin does not act directly on photoreceptor guanylate cyclase (Gray- Keller et al., 1993) but rather by inhibiting rhodopsin phosphorylation (Gorodovikova et al., 1994; Kawa- mura et al., 1994). Neurocalcin and hippocalcin possess like recoverin a myristoylation consensus sequence (Zozulya et al., 1992; Kobayashi et al., 1993). Their functions are still unknown and little is known about their distribution. Besides brain area, neurocalcin has previously been detected in amacrine and ganglion cells from bovine retina (Nakano et al., 1992) which is similar with our present findings. Recently, neurocalcin was also detected in tufted cells from the rat olfactory bulb (Bastianelli et al., 1993) and in neurons from the rat olfactory epithelium (Bastianelli et al., 1995). Hippocalcin seemed to be restricted to a few specific brain regions where its function is still unknown. This study demonstrated for the first time hippocalcin-like IR in retina. In rat retina, neurocalcin and hippocalcin distributions are quite similar however hippocalcin-IR could only be detected at P5 indicating a delay in its expression. Their probable myristoylation let us presume that either a calcium or a GTP myristoylation proteins
switch might play a role in cellular signalling as shown for recoverin (gozulya et al., 1992) and ADP- ribosylation factor respectively (Kahn et al., 1992). Further studies are needed to answer to questions about their role.
Acknowledgements
The authors thank Dr Edith Nemec for correcting the English, L~on Surardt for excellent technical assistance and Paulette Miroir for preparing the manuscript. This work w a s supported by a FRSM grant (no. 3.4517.92) to R.P. and by the Science Research Promotion Fund from the Japan Private School Promotion Foundation to K.T.
R e f e r e n c e s
Bastianelli, E., Okazaki, K., Hidaka, H. and Pochet, R. (1993). Neurocalcin immunoreactivity in rat olfactory bulb. Neurosci. Lett. 161, 165-8.
Bastianelli, E., Polans, A.S., Hidaka, H. and Pochet, R. (1995). Differential distribution of six calcium-binding proteins in the rat olfactory epithelium during postnatal development and adulthood. ]. Corap. Neurol. in press.
Braekevelt, C.R. and Hollenberg, M.J. (1970). The de- velopment of the retina of the albino rat. Am. ]. Anat. 127, 281-302.
Cambray-Deakin, M.A. and Burgoyne, R.D. (1992). In- tracellular calcium and N-methyl-D-aspartate- stimulated neuritogenesis in rat cerebelar granule cel cultures. Dev. Brain Res. 66, 25-32.
Chu, Y., Humphrey, M.F. and Constable, I.J. (1993). Horizontal cells of the normal and dystrophic rat retina: A wholemount study using immunolabelling for the 28-kDa calcium-binding protein. Exp. Ege Res. 57, 141-8.
Dechesne, C. J. and Thomasset, M. (1988). Calbindin (CaBP 28kDa) appearance and distribution during develop- ment of the mouse inner ear. Dev. Brain Res. 40, 233-42.
Dizhoor, A. M., Ray, S., Kumar, S., Niemi, G., Spencer, M., Brolley, D., Walsh, K. A., Philippov, P. P. Hurley, J. B. and Stryer, L. (1991). Recoverin: a calcium sensitive activator of retinal rod guanylate cyclase. Science 251, 915-18.
Ellis, J.H., Richards, D.E. and Rogers, J.H. (1991). Calretinin and calbindin in the retina of the developing chick. Cell Tissue Res. 264, 197-208.
Enderlin, S., Norman, A.W. and Celio, M.R. (1987). Ontogeny of the calcium binding protein calbindin D-28k in the rat nervous system. Anat. Embrgol. 177, 15-28.
Gorodovikova, t .N., Gimelbrant, A.A., Scnin, I.I. and Philippov, P. P. (1994). Recoverin mediates the calcium effect upon rhodopsin phosphorylation and cGMP hydrolysis in bovine retina rod cells. FIBS Lett. 349, 187-90.
Gray-Keller, M. P., Polaris, A. S., Palczewski, K. and Detwiler, P.B. (1993). The effect of recoverin-like calcium- binding proteins on the photoresponse of retinal rods. Neuron 10, 523-31.
Hamano, K., Kiyama, H., Emson, P.C., Manabe, R., Nakauchi, M. and Tohyama, M. (1990). Localization of two calcium binding proteins, calbindin (28kD) and parvalbumin (12kD), in the vertebrate retina. ]. Comp. Neurol. 302, 417-24.
Hendrickson, A. E., Van Brederode, J. F. M., Mulligan, K. A. and Colio, M. R. (1991). Development of the calcium-

266 E. BASTIANELLI ETAL.
binding proteins parvalbumin and calbindin in monkey striate cortex. ]. Comp. Neurol. 307, 626-46.
Hidaka, H. and Okazaki, K. (1993). Neurocalcin family: a novel calcinm-binding protein abundant in bovine central nervous system. Neurosci. Res. 16, 73-7.
Horsburgh, G.M. and Sefton, ]. (1987). Cellular degener- ation and synaptogenesis in the developing retina of the rat. ]. Comp. Neurol. 263, 553-66.
Kahn, R. A., Randazzo, P., Serafini, T., Weiss, O., Rulka, C., Clark, ]., Amherdt, M., Roffee, P., Orci, L., and Rothman, ]. E. (1992). The amino terminus of ADP-ribosylation factor (ARF) is a critical determinant of ARF activities and is a potent and specific inhibitor of protein transport. ]. Biol. Chem. 267, 13039-46.
Kater, S. B., Mattson, M. P., Cohan, C. S. and Connor, 1. A. (1988). Calcium regulation of the neuronal growth cone. Trends Neurosci. 11, 315-21.
Kawamura, S., Cox, ]. A. and Nef, P. (1994). Inhibition of rhodopsin phosphorylation by non-myristoylated recombinant recovedn. Biochem. Biophys. Res. Comm. 203, 121-7.
Kobayashi, M., Takamatsu, K., Saitoh, S., Miura, M. and Noguchi, T. ( 1992). Molecular cloning of hippocalcin, a novel calcinm-binding protein of the recoverin family exclusively expressed in hippocampus. Biochem. Biophys. Res. Commun. 189, 511-17.
Kobayashi, M., Takamatsu, K., Saitoh, S. and Noguchi, T. (1993). Myristoylation of hippocalcin is linked to its calcium-dependent membrane association properties. 1. Biol. Chem. 268, 18898-904.
Lipp, H.-P., Wolfer, D.P., Oin, W.X., Klee, C.B. and Heizmann, C.W. (1993). CBP-18, a Ca2+-binding protein in rat brain: tissue distribution and localization. ]. Neurochem. 60, 1639--49.
McGinnis, 1. F., Stepanik, P. L., Baehr, W., Subbaraya, I. and Lerious, V. (1992). Cloning and sequencing of the 23 kDa mouse photoreceptor cell-specific protein. FEBS Lett. 302, 172-6.
Milam, A. H, Dacey, D.M. and Dizhoor, A.M. (1993). Recoverin immunoreactivity in mammalian cone bi- polar cells. Visual Neurosci. 10, 1-12.
Nakano, A., Terasawa, M., Watanabe, M., Usuda, N., Morita, T. and Hidaka, H. (1992). Neurocalcin, a novel calcium binding protein with three EF-hand domains, expressed in retinal amacrine cells and ganglion cells. Biochem. Biophys. Res. Commun. 186, 1207-11.
Pasteels, B., Miki, M., Hatakenaka, S. and Pochet, R. (1987). Immunohistochemical cross-reactivity and electro- phoretic comigration between calbindin-D27kDa and visinin. Brain Res. 412, 107-13.
Pasteels, B., Rogers, ]. H., Blachier, F. and Pochet, R. (1990). Calbindin and calretinin localization in retina from different species. Visual Neurosci. 5, 1-16.
Pochet, R., Pasteels, B., Seto-Ohshima, A., Bastianelli, E., Kitaiima, S. and Van Eldik, L. ]. (1991). Calmodulin and Calbindin localization in retina from six vertebrate species. ]. Comp. Neurol. 314, 750-62.
Rabi6, A., Thomasset, M., Parkes, C.O. and Clavel, M. C. (1985). Immunocytochemical detection of 28000-MW calcium-binding protein in horizontal cells of the rat retina. Cell Tissue Res. 240, 493-6.
Raedler, A. and Sievers, I. (1975). The development of the visual system of the albino rat. Adv. Anat. Embryol. Cell Biol. 50, 1-87.
Rami, A., Brehier, A., Thomasset, M. and Rabi6, A. (1987). Cholecalcin (28-kDa Calcium-binding protein) in the rat hippocampus: development in normal animals and
in altered thyroid states. An immunocytochemical study. Dev. Biol. 124, 228-38.
Rohrenbeck, J., Wassle, H. and Heizmann, C.W. (1987). Immunocytochemical labelling of horizontal cells in mammalian retina using antibodies against calcium- binding proteins. Neurosci. Lett. 77, 255-60.
Rohrenbeck, J., Wassle, H. and Boycott, B.B. (1989). Horizontal cells in the monkey retina: Immuno- cytochemical staining with antibodies against calcium binding proteins. Eur. J. Neurosci. 1 ,407-20 .
Saitoh, S., Takamatsu, K., Kobayashi, M. and Noguchi, T. (1993). Distribution of hippocalcin mRNA and immuno- reactivity in rat brain. Neurosci. Lett. 157, 107-10.
Sarthy, P.V. and Hilbush, B.S. (1983). Lucifer yellow uptake in developing rat retina: selective staining of horizontal cells. Dev. Brain Res. 11 ,275-81 .
Schwaller, B., Buchwald, P., Blfimcke, I., Cello, M. R. and Hunziker, W. (1993). Characterization of a polyclonal antiserum against the purified human recombinant calcium binding protein calretinin. Cell Calcium 14, 6 3 9 4 8 .
Spencer, R., Charman, M., Emtage, J. S. and Lawson, D. E. M. (1976). Production and properties of vitamin-D- induced mRNA for chick calcium-binding protein. Eur.]. Biochem. 71, 399-409.
Spira, A.W. (1974). Ultrastructural localization of chol- inesterase activity in the developing rat retina. J. Histochem. Cytochem. 22, 868-73.
Stichel, C. C., Singer, W., Heizmann, C. W. and Norman, A. W. (1987). Immunochemical localization of calcium- binding proteins, parvalbumin and calbindin-D28k, in the adult and developing visual cortex of cats: a light and electron microscopic study. J. Comp. Neurol. 262, 563-77.
Takamatsu, K. and Uyemura, K. (1992). Identification of recoverin-like immunoreactivity in mouse brain. Brain Res. 571, 350-3.
Terasawa, M., Nakano, A., Kobayashi, R. and Hidaka, H. (1992). Neurocalcin: a novel calcium-binding protein from bovine brain. ]. Biol. Chem. 267, 19596-9.
Uesugi, R., Yamada, M., Mizuguchi, M., Bairnbridge, K. G. and Kim, S.U. (1992). Calbindin D-28k and parvalbumin immunohistochemistry in developing rat retina. Exp. Eye Res. 54, 491-9.
Vacca, L.L., Abrahams, S.I. and Naftchi, N. E. (1980). A modified peroxidase-antiperoxidase procedure for im- proved localization of substance P in rat spinal cord. ]. Histochem. Cytochem. 28, 297-304.
Versaux-Botteri, C., Pochet, R. and Nguyen-Legros, I. (1989). Immunohistochemical localization of GABA- containing neurons during postnatal development of the rat retina. Invest. Ophthalmol. Vis. Sci. 30, 652-9.
Verstappen, A., Parmentier, M., Chimoaga, M., Lawson, D. E. M., Pasteels, J. L. and Pochet, R. (1986). Vitamin D- dependent calcium binding protein immunoreactivity in human retina. Ophth. Res. 18, 209-214.
Wiechmann, A.F. and Hammarback, I.A. (1993). Mol- ecular cloning and nucleotide sequence of a cDNA encoding recoverin from human retina. Exp. Eye Res. 56, 463-70.
Zhang, D., Gallagher, M., Sladek, C.D. and Yeh, H.H. (1990). Postnatal development of corticotropin re- leasing factor-like immunoreactive amacrine cells in the rat retina. Dev. Brain Res. 51, 185-94.
Zozulya, S. and Stryer, L. (1992). Calcinm-myristoyl protein switch. Proc. Natl. Acad. Sci., USA 89, 11569-73.