oral salbutamol decreases il-12 in patients with secondary progressive multiple sclerosis
TRANSCRIPT

Ž .Journal of Neuroimmunology 117 2001 156–165www.elsevier.comrlocaterjneuroin
Oral salbutamol decreases IL-12 in patients with secondary progressivemultiple sclerosis
Karim Makhlouf a, Manuel Comabella b, Jaime Imitola a, Howard L. Weiner a,Samia J. Khoury a,)
a Center for Neurologic Diseases, Brigham and Women’s Hospital, and HarÕard Medical School, 77 Louis Pasteur AÕenue, Boston, MA 02115, USAb Department of Clinical Neuroimmunology, Hospital Val d’Hebron, Barcelona, Spain
Received 14 February 2001; received in revised form 10 April 2001; accepted 13 April 2001
Abstract
Ž .IL-12 is a key cytokine for Th1 cell development and may be important in the pathogenesis of multiple sclerosis MS . The b -agonist2
salbutamol is known to decrease IL-12 production in monocytes of normal individuals through increased intracellular cAMP. In aprospective open-label study, we investigated by flow cytometry the effect of a 2-week long oral salbutamol treatment on monocyte IL-12production in 21 secondary progressive MS patients. Baseline IL-12 production was higher in patients than in healthy controls. Thetreatment induced a significant decrease in the percentage of IL-12-producing monocytes and dendritic cells that lasted up to 1 week aftertreatment interruption. This first report on the use of salbutamol in MS shows that this drug has immunomodulatory properties both invivo and in vitro, and may be beneficial in the treatment of MS. q 2001 Elsevier Science B.V. All rights reserved.
Keywords: Human; Multiple sclerosis; Salbutamol; b -agonists; Th1rTh2; Cytokines2
1. Introduction
Ž .Multiple sclerosis MS is generally considered as anŽ .immune-mediated disease Khoury et al., 1996 , character-
ized by a chronic inflammation in the central nervoussystem, leading to demyelination and ultimately to func-tional disability. Th1 cytokines are increased in MS andIFN-g was shown to induce exacerbations of the diseaseŽ .Panitch et al., 1987 .
IL-12 is a potent inducer of IFN-g secretion and drivesŽ .Th1 responses Trinchieri, 1995; Magram et al., 1996 . It
is a heterodimeric cytokine made of two disulfide-linkedchains, p35 and p40, and produced by antigen-presenting
Ž .cells APC like B cells, monocytesrmacrophages andŽ .dendritic cells DC . IL-12 is considered a key cytokine in
the pathogenesis of experimental autoimmune encephalo-Ž . Ž .myelitis EAE Segal et al., 1998; Leonard et al., 1995Ž .and MS Trembleau et al., 1995 , thus drugs that decrease
ŽIL-12 production are of potential therapeutic interest Karp.et al., 2000 . We have previously shown that IL-12 produc-
) Corresponding author. Tel.: q1-617-525-5370; fax: q1-617-525-5305.
Ž .E-mail address: [email protected] S.J. Khoury .
tion and the percentage of IL-12-producing monocytes areŽincreased in patients with progressive MS Balashov et al.,
.1997; Comabella et al., 1998 and that the increasedpercentage of IL-12-producing monocytes correlates withdisease activity and is normalized by pulse cyclophospha-
Ž .mide therapy Comabella et al., 1998 . Panina-BordignonŽ .et al. 1997 reported that b -agonists such as salbutamol2
Ž .SB, also called albuterol in the USA selectively inhibitthe production of IL-12 by human monocytes in vitro andin vivo in healthy subjects, through increased intracellularcAMP. In animal models of autoimmune disease, b -2
agonists were shown to suppress chronic-relapsing EAE inŽ .Lewis rats Wiegmann et al., 1995 , and to suppress
collagen-induced arthritis, a murine model for rheumatoidŽ .arthritis Malfait et al., 1999 . Furthermore, b -adrenergic2
receptor expression is increased on peripheral bloodŽ . Žmononuclear cells PBMC of patients with MS Arnason
.et al., 1988 , and is correlated with clinical and MRIŽ .disease activity Zoukos et al., 1994 . We now report the
results of a prospective open-label pilot study of theimmunological effects of oral SB, in particular on thepercentage of IL-12-producing peripheral blood monocytesof patients with secondary progressive multiple sclerosisŽ .SP MS .
0165-5728r01r$ - see front matter q 2001 Elsevier Science B.V. All rights reserved.Ž .PII: S0165-5728 01 00322-8

( )K. Makhlouf et al.rJournal of Neuroimmunology 117 2001 156–165 157
2. Patients and methods
2.1. Patients and study design
Sixteen MS patients followed at the outpatient MSclinic at the Brigham and Women’s Hospital in Bostonwere included in an open-labeled, longitudinal pilot studyto measure the effects of oral SB on IL-12 production.Five additional patients were enrolled to determine SBeffects on other cytokines and surface markers. The inclu-sion criteria were as follows:
Ø Clinically definite secondary progressive MS;Ø Age range:18–60 years;Ø Acceptable birth control method for women in repro-
ductive age.
The study was approved by the Institutional ReviewBoard and all patients gave informed consent. The mean
Ž .age was 49.5 years range: 32–57 . There were 11 menand 10 women. The average duration of the disease was
Ž11.7 years. The average EDSS score assessed within 6.months prior to inclusion was 4.6.
The exclusion criteria were:
Ø Pregnancy or lactation;Ø Concomitant therapy with any of the following drugs:
b -agonist or antagonist, diuretics, tricyclic antide-2
pressants, monoamine oxidase inhibitors, immuno-Žmodulatory agents such as methylprednisolone,
.glatiramer acetate or interferon b within 3 monthsprior to inclusion in the study;
Ø Abnormal ECG;Ø Diagnosis with any of the following diseases: chronic
hypertension, hyperthyroidism, diabetes, or seizuredisorder.
ŽTwenty-one healthy subjects 13 men, 8 women, mean.age: 37.9 years, range: 22–63 were used as normal con-
trols to assess baseline levels of IL-12 and IL-10. Twonormal individuals were monitored for 3 weeks at thesame schedule as the MS study patients.
2.2. Drug and treatment regimens
The MS patients received on the first day a single doseof 4 mg SB orally. Blood was obtained immediately prior
Žto the intake of the drug time point called pre-D1, or.Baseline, see Fig. 1 and after 2 h, corresponding to the
Ž .peak serum level of SB time point: Post D1 . Patientsreceived 4 mg SB daily in the morning for 1 week, then 4mg three times a day for 1 week. No medication was givenduring the last week of the study. Blood for IL-12 expres-sion was also obtained at the end of weeks 1 and 2, again
Žjust before and 2 h after the intake of the drug respec-.tively, pre-W1–post-W1 and pre-W2–post-W2 . Blood was
drawn before and after the morning dose at the end of thesecond week. Finally, a blood sample was obtained at the
Ž .end of the third week time point: W3 . The therapeutic SBserum level is between 3.8 and 14.8 ngrml with usual oral
Ž .doses in asthmatic patients Anonymous, 2000 .
2.3. Cell separation and culture conditions
PBMC were isolated by Ficoll–Hypaque density gradi-Žent centrifugation from Amersham Pharmacia Biotech,
. Ž 6 .Uppsala, Sweden and resuspended 2=10 rml in com-plete culture media consisting of RPMI 1640 mediumsupplemented with 10% fetal bovine serum, 4 mM L-glutamine, 25 mM Hepes buffer, 50 unitsrml penicillin,
Žand 50 mgrml streptomycin all from BioWhittaker,.Walkersville, MD . Since unstimulated monocytes produce
no detectable IL-12, in vitro stimulation was required.Optimal conditions for mononuclear cell stimulation weredetermined by previous kinetic studies using single stimu-
Žlating agents or combination of agents Comabella et al.,.1998 . Maximal stimulation of monocytes for IL-12 pro-
duction was obtained after sequential stimulation with
Fig. 1. Time schedule for blood sampling in relation to drug intake and dose: SBssalbutamol; D1sday 1; W1s last day of week 1; W2s last day ofweek 2; W3s last day of week 3.

( )K. Makhlouf et al.rJournal of Neuroimmunology 117 2001 156–165158
Ž . 6IFN-g and lipopolysaccharide LPS as follows: 2=10ŽPBMC were placed in polypropylene culture tubes Fisher
.Scientific, Pittsburgh, PA in a volume of 1 ml andŽ . Žstimulated with rhIFN-g 10 ngrml for 2 h and LPS 100
. Ž .ngrml for 22 h; monensin 3 mM was added to thePBMC culture tubes at the same time as LPS in order toinhibit cytokine secretion. Maximal T cell stimulation forIL-4, IL-5, IL-10 and IFN-g production was obtained as
Žfollows: PBMC were placed in 6-well plates Costar,.Cambridge, MA in a volume of 6 ml and cultured with a
Žcombination of phorbol 12-myristate 13-acetate PMA,. Ž .100 ngrml , ionomycin 250 ngrml , and phytohemagglu-
Ž .tinin PHA, 1 mgrml for 6 h, in the presence of monensinŽ .3 mM .
2.4. Antibodies and reagents
The following purified staining mouse anti-human anti-Ž .bodies were purchased from PharMingen San Diego, CA :
ŽFITC-labeled IgG1 and IgG2a both used as isotype con-.trols , FITC-labeled anti-CD3, anti-CD4 and anti-CD14,
PE-labeled anti-IFN-g, anti-IL-12, anti-IL-10, anti-IL-5 andanti-IL-4, FITC anti-CD40, PE anti-CD80, PE anti-CD54,PE-anti-CD86, and APC-anti CD33. The anti IL-12 anti-body recognized both IL-12p70 and p40. PMA, iono-
Žmycin, LPS, and saponin were obtained from Sigma St.. ŽLouis, MO ; PHA was from Murex Diagnostics Dartford,
. Ž w .England ; monensin GolgiStop and recombinant humanŽ .rh IFN-g were from PharMingen.
2.5. Staining and flow cytometry
The details of the staining procedure have been previ-Ž .ously described Comabella et al., 1998 . Briefly, PBMC
Žwere first stained using surface marker antibodies CD3 or.CD4 for T cells, CD14 for monocytes , fixed with 4%
Žparaformaldehyde solution from EMS, Ft. Washington,.PA , permeabilized with saponin, and then stained with
intracytoplasmic antibodies. All staining steps were carriedout at q48C in the dark. Cells were then analyzed using aFACSort flow cytometry system equipped with CellQuest
Ž .software both from Beckton Dickinson, San Jose, CA .40,000 events were acquired for monocytes and 10,000 forlymphocytes. The percentage of cytokine- or surfacemarker-positive cells was calculated by subtracting theisotype control antibody signal from the specific anti-cyto-kineranti-marker antibody signal.
2.6. In Õitro experiments
In order to investigate the in vitro effect of SB andcAMP on the expression of cytokines and of surfacemarkers in monocytes from normal controls, blood was
Žobtained from five healthy volunteers mean age: 26 years,.range: 22–36 and PBMC were isolated by gradient sepa-
Ž .ration Ficoll . Before staining, the cells were stimulatedŽ . Ž .by rhIFN-g 10 ngrml for 2 h and then LPS 100 ngrml
Ž .for 22 h, in the presence of monensin 3 mM at 378C. SBŽ .or dibutyric cAMP db-cAMP , both purchased from
Sigma, were added to the cultures from the beginning, atthe following concentrations: 0.1, 1, 10 and 100 nM forSB, and 10, 100 and 1000 mM for db-cAMP. Cell acquisi-
Žtion by flow cytometry was gated on the monocyte CD14.q cluster. Mouse anti-human antibodies against CD40,
Ž . Ž . Ž .CD54 or ICAM , CD80 or B7-1 , CD86 or B7-2 HLAŽ .DR2 and IL-12 Receptor b1 on T cells were purchased
Žfrom PharMingen. We used sodium butyrate also from.Sigma as control for db-cAMP, at the following concen-
trations: 20, 200 and 2000 mM.
2.7. Statistical methods
Out of 21 patients, 18 had a complete set of data. Thebaseline data was missing for one patient, although thispatient went through the entire treatment protocol. In
Žanother patient, the data at the end of the first week pre-.and post-W1 was missing. In a third patient, the data at
Ž .the end of week 3 W3 was missing. P values F0.05were considered as statistically significant.
A nonparametric repeated measures regression modelenabled us to evaluate the response using three predictorsŽ .or effects :
Ø The bolus predictor, assessing the impact of the bolusdose over the pre-dose measurement.
Ø The treatment predictor, assessing the daily treatmenteffect.
Ø The AwashoutB predictor, comparing the last mea-Ž . Žsurement W3 to the first measurement baseline, or
.pre-D1 , looking at the long-lasting effect of treat-ment, 1 week after its interruption.
We used the Mann–Whitney unpaired test to comparethe percentage of IL-12- or IL-10-producing monocytesbetween MS patients and controls in different settings. Wealso used the Wilcoxon test for paired data, and theKruskal–Wallis test for monocytes, dendritic cells andsurface marker data analysis when appropriate, and inorder to avoid repeated t-tests.
3. Results
3.1. MS patients haÕe a higher percentage of IL-12 pro-ducing monocytes
Ž .The secondary progressive MS patients ns20 had asignificantly higher percentage of IL-12-producing mono-
Ž .cytes at baseline mean"S.E.s28.3"2.4% when com-Žpared to 21 healthy controls mean"S.E.s6.92"1.01%,
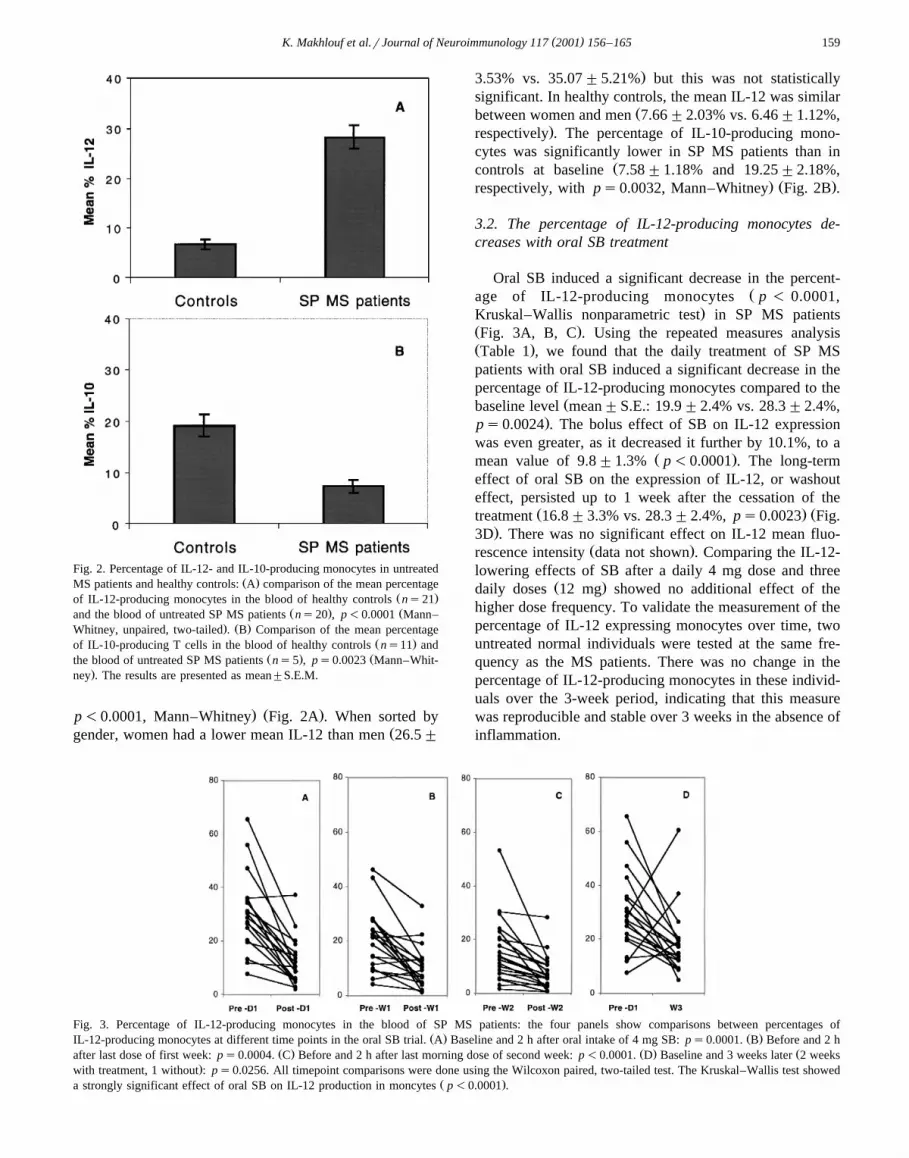
( )K. Makhlouf et al.rJournal of Neuroimmunology 117 2001 156–165 159
Fig. 2. Percentage of IL-12- and IL-10-producing monocytes in untreatedŽ .MS patients and healthy controls: A comparison of the mean percentage
Ž .of IL-12-producing monocytes in the blood of healthy controls ns21Ž . Žand the blood of untreated SP MS patients ns20 , p-0.0001 Mann–
. Ž .Whitney, unpaired, two-tailed . B Comparison of the mean percentageŽ .of IL-10-producing T cells in the blood of healthy controls ns11 and
Ž . Žthe blood of untreated SP MS patients ns5 , ps0.0023 Mann–Whit-.ney . The results are presented as mean"S.E.M.
. Ž .p-0.0001, Mann–Whitney Fig. 2A . When sorted byŽgender, women had a lower mean IL-12 than men 26.5"
.3.53% vs. 35.07"5.21% but this was not statisticallysignificant. In healthy controls, the mean IL-12 was similar
Žbetween women and men 7.66"2.03% vs. 6.46"1.12%,.respectively . The percentage of IL-10-producing mono-
cytes was significantly lower in SP MS patients than inŽcontrols at baseline 7.58"1.18% and 19.25"2.18%,
. Ž .respectively, with ps0.0032, Mann–Whitney Fig. 2B .
3.2. The percentage of IL-12-producing monocytes de-creases with oral SB treatment
Oral SB induced a significant decrease in the percent-Žage of IL-12-producing monocytes p - 0.0001,
.Kruskal–Wallis nonparametric test in SP MS patientsŽ .Fig. 3A, B, C . Using the repeated measures analysisŽ .Table 1 , we found that the daily treatment of SP MSpatients with oral SB induced a significant decrease in thepercentage of IL-12-producing monocytes compared to the
Žbaseline level mean"S.E.: 19.9"2.4% vs. 28.3"2.4%,.ps0.0024 . The bolus effect of SB on IL-12 expression
was even greater, as it decreased it further by 10.1%, to aŽ .mean value of 9.8"1.3% p-0.0001 . The long-term
effect of oral SB on the expression of IL-12, or washouteffect, persisted up to 1 week after the cessation of the
Ž . Žtreatment 16.8"3.3% vs. 28.3"2.4%, ps0.0023 Fig..3D . There was no significant effect on IL-12 mean fluo-
Ž .rescence intensity data not shown . Comparing the IL-12-lowering effects of SB after a daily 4 mg dose and three
Ž .daily doses 12 mg showed no additional effect of thehigher dose frequency. To validate the measurement of thepercentage of IL-12 expressing monocytes over time, twountreated normal individuals were tested at the same fre-quency as the MS patients. There was no change in thepercentage of IL-12-producing monocytes in these individ-uals over the 3-week period, indicating that this measurewas reproducible and stable over 3 weeks in the absence ofinflammation.
Fig. 3. Percentage of IL-12-producing monocytes in the blood of SP MS patients: the four panels show comparisons between percentages ofŽ . Ž .IL-12-producing monocytes at different time points in the oral SB trial. A Baseline and 2 h after oral intake of 4 mg SB: ps0.0001. B Before and 2 h
Ž . Ž . Žafter last dose of first week: ps0.0004. C Before and 2 h after last morning dose of second week: p-0.0001. D Baseline and 3 weeks later 2 weeks.with treatment, 1 without : ps0.0256. All timepoint comparisons were done using the Wilcoxon paired, two-tailed test. The Kruskal–Wallis test showed
Ž .a strongly significant effect of oral SB on IL-12 production in moncytes p-0.0001 .

( )K. Makhlouf et al.rJournal of Neuroimmunology 117 2001 156–165160
Table 1Effects of SB treatment on the percentage of IL-12-producing stimulatedmonocytes in SP MS patientsP values are in comparison to baseline.
Predictor Mean IL-12% P valueŽ ."S.E.
Ž .Baseline 28.3 "2.4 –Ž .Bolus 9.8 "1.3 -0.0001Ž .Daily treatment 19.9 "2.4 0.0024Ž .Wash-out 16.8 "3.3 0.0023
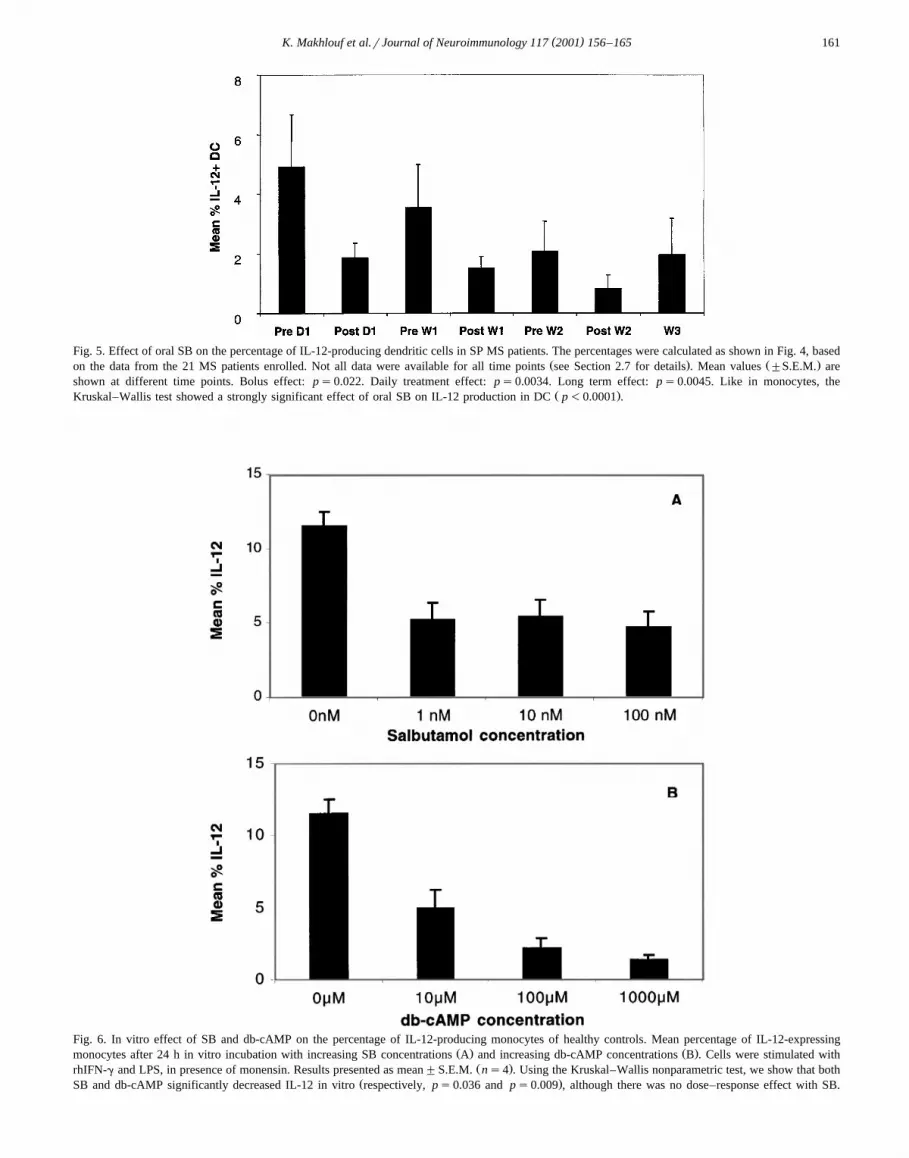
3.3. The percentage of IL-12-producing dendritic cells isalso reduced by oral SB
DC represent an important source of IL-12 in vivoŽ .Macatonia et al., 1995 , and SB was shown to inhibit
ŽIL-12 production in vitro in human DC Panina-Bordignon.et al., 1997 . Thus, we wanted to assess the in vivo effect
of oral SB on the DC-produced IL-12. There are two majorDC subsets in human peripheral blood: the plasmacytoidŽ . ŽCD123q , CD33y and the myeloid CD11cq , CD33
. Ž .q DC, which are both CD14y Reid et al., 2000 . TheCD33qrCD14y myeloid DC subset is responsible for
Ž .IL-12 secretion Reid et al., 2000 . In preliminary experi-Žments using a previously described gating strategy Thomas
.and Lipsky, 1994 , we stained the PBMC of three normalŽ .subjects and four untreated MS patients with CD14 FITC ,
Ž . Ž . Ž .CD33 APC and IL-12 PE, intracytoplasmic Fig. 4 .We showed that more than 95% of the IL-12qrCD14y
Ž . Ž .cells Region R2, Fig. 4B were also CD33q Fig. 4C .Consequently, in the data from the 21 patients followed inthis trial, we used the percentage of IL-12qrCD14ycells as a good approximation of the percentage of IL-12-producing DC. We show that oral SB also significantly
Ždecreased this percentage Kruskal–Wallis corrected for. Ž .ties: p-0.0001 Fig. 5 . Similarly, there was a signifi-
Ž . Žcant bolus effect ps0.022 , daily treatment effect ps. Ž .0.0034 and a long-term-effect ps0.0045 . The small
percentage values account for the very small size of theDC population in the total PBMC population.
3.4. The percentage of IL-12-producing monocytes de-creases with SB and db-cAMP in Õitro
To investigate the mechanisms of SB treatment effect,we used PBMC cultures from healthy donors that werestimulated in vitro, and we tested the effect of adding SBor db-cAMP to the cultures. db-cAMP is a membranepermeable cAMP analog that activates cAMP protein–kinases after intracytoplasmic release of two butyrategroups. The mean percentage of IL-12-producing mono-cytes after stimulation and 24 h incubation was 11.61"1.9%. In vitro addition of SB decreased this percentageŽ .Fig. 6A , with no apparent dose response. The percentageof IL-12q monocytes was 5.23"2.29% after 1 nM SBŽ . Žps0.052 , 5.41"2.35% after 10 nM SB ps0.048
.compared to baseline , and 4.76"1.99% after 100 nMŽ .ps0.038 . Treatment with db-cAMP induced a signifi-cant decrease in the percentage of IL-12-producing mono-cytes in a dose-dependent manner: 4.97"2.51% at 10mMŽ . Ž .ps0.012 , 2.14"1.5% at 100 mM ps0.0016 and
Ž . Ž .1.38"0.7% at 1000 mM ps0.002 Fig. 6B . Therewas no effect on the percentage of IL-12-producing mono-cytes after in vitro addition of increasing concentrations ofsodium butyrate, showing that it was cAMP, and not thetwo butyrate groups released at cell entry, that was respon-sible for the effect on IL-12.
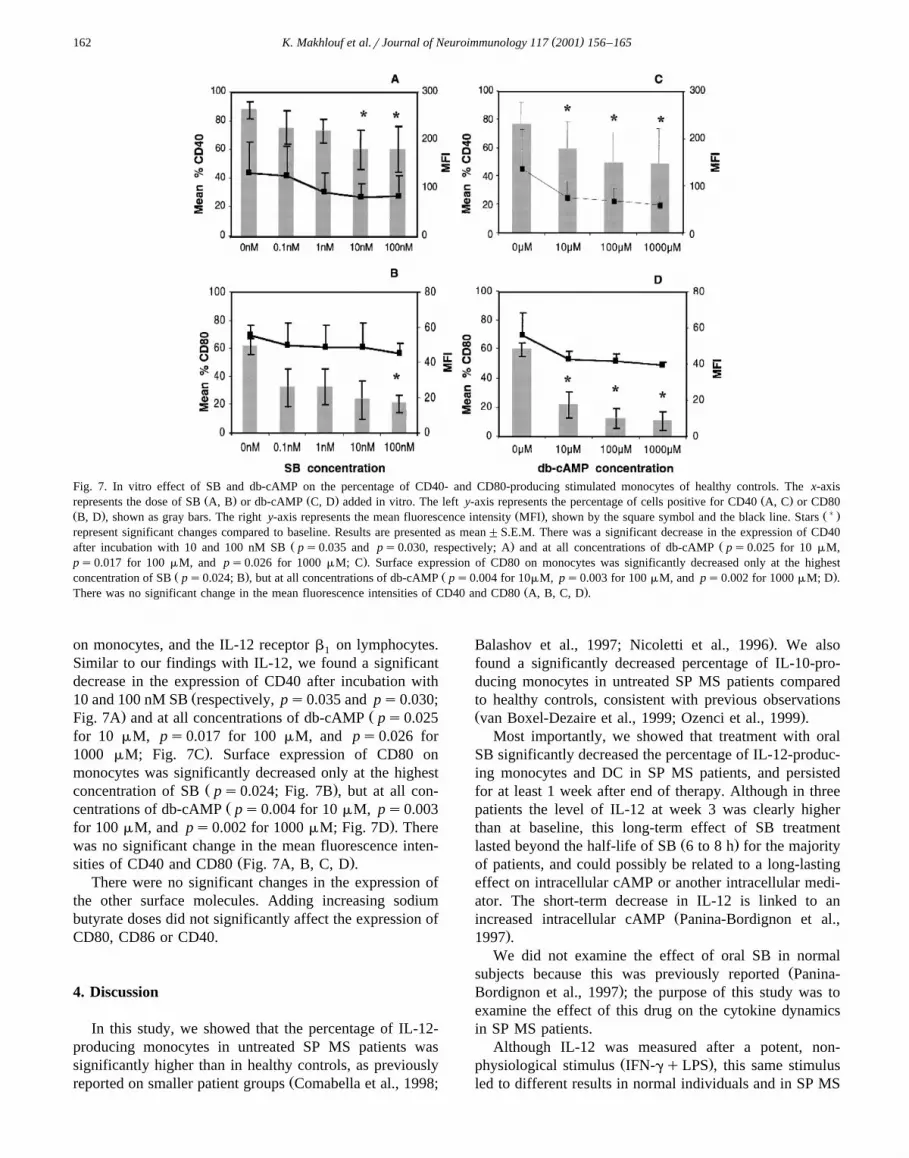
3.5. SB and db-cAMP in Õitro decrease the percentage ofCD40 and CD80-producing monocytes
We also studied the in vitro expression of the followingsurface markers: CD40, CD80, CD86, CD54, HLA class II
Ž .Fig. 4. Determination of the percentage of IL-12-producing dendritic cells: starting from the general monocyte cluster in PBMC region R1, A and thenŽ .gating on the upper left quadrant Region R2, B allows to determine the IL-12qrCD14y cell population. We show that this population is mostly
Ž .CD33q C , and thus corresponds to the myeloid DC population. The data shown in this figure is the representative of the seven subjects analyzed. Allsamples were stimulated by IFN-g and LPS for 24 h as described in Patients and methods.
( )K. Makhlouf et al.rJournal of Neuroimmunology 117 2001 156–165 161
Fig. 5. Effect of oral SB on the percentage of IL-12-producing dendritic cells in SP MS patients. The percentages were calculated as shown in Fig. 4, basedŽ . Ž .on the data from the 21 MS patients enrolled. Not all data were available for all time points see Section 2.7 for details . Mean values "S.E.M. are
shown at different time points. Bolus effect: ps0.022. Daily treatment effect: ps0.0034. Long term effect: ps0.0045. Like in monocytes, theŽ .Kruskal–Wallis test showed a strongly significant effect of oral SB on IL-12 production in DC p-0.0001 .
Fig. 6. In vitro effect of SB and db-cAMP on the percentage of IL-12-producing monocytes of healthy controls. Mean percentage of IL-12-expressingŽ . Ž .monocytes after 24 h in vitro incubation with increasing SB concentrations A and increasing db-cAMP concentrations B . Cells were stimulated with
Ž .rhIFN-g and LPS, in presence of monensin. Results presented as mean"S.E.M. ns4 . Using the Kruskal–Wallis nonparametric test, we show that bothŽ .SB and db-cAMP significantly decreased IL-12 in vitro respectively, ps0.036 and ps0.009 , although there was no dose–response effect with SB.
( )K. Makhlouf et al.rJournal of Neuroimmunology 117 2001 156–165162
Fig. 7. In vitro effect of SB and db-cAMP on the percentage of CD40- and CD80-producing stimulated monocytes of healthy controls. The x-axisŽ . Ž . Ž .represents the dose of SB A, B or db-cAMP C, D added in vitro. The left y-axis represents the percentage of cells positive for CD40 A, C or CD80
Ž . Ž . Ž) .B, D , shown as gray bars. The right y-axis represents the mean fluorescence intensity MFI , shown by the square symbol and the black line. Starsrepresent significant changes compared to baseline. Results are presented as mean"S.E.M. There was a significant decrease in the expression of CD40
Ž . Žafter incubation with 10 and 100 nM SB ps0.035 and ps0.030, respectively; A and at all concentrations of db-cAMP ps0.025 for 10 mM,.ps0.017 for 100 mM, and ps0.026 for 1000 mM; C . Surface expression of CD80 on monocytes was significantly decreased only at the highest
Ž . Ž .concentration of SB ps0.024; B , but at all concentrations of db-cAMP ps0.004 for 10mM, ps0.003 for 100 mM, and ps0.002 for 1000 mM; D .Ž .There was no significant change in the mean fluorescence intensities of CD40 and CD80 A, B, C, D .
on monocytes, and the IL-12 receptor b on lymphocytes.1
Similar to our findings with IL-12, we found a significantdecrease in the expression of CD40 after incubation with
Ž10 and 100 nM SB respectively, ps0.035 and ps0.030;. ŽFig. 7A and at all concentrations of db-cAMP ps0.025
for 10 mM, ps0.017 for 100 mM, and ps0.026 for.1000 mM; Fig. 7C . Surface expression of CD80 on
monocytes was significantly decreased only at the highestŽ .concentration of SB ps0.024; Fig. 7B , but at all con-
Žcentrations of db-cAMP ps0.004 for 10 mM, ps0.003.for 100 mM, and ps0.002 for 1000 mM; Fig. 7D . There
was no significant change in the mean fluorescence inten-Ž .sities of CD40 and CD80 Fig. 7A, B, C, D .
There were no significant changes in the expression ofthe other surface molecules. Adding increasing sodiumbutyrate doses did not significantly affect the expression ofCD80, CD86 or CD40.
4. Discussion
In this study, we showed that the percentage of IL-12-producing monocytes in untreated SP MS patients wassignificantly higher than in healthy controls, as previously
Žreported on smaller patient groups Comabella et al., 1998;
.Balashov et al., 1997; Nicoletti et al., 1996 . We alsofound a significantly decreased percentage of IL-10-pro-ducing monocytes in untreated SP MS patients comparedto healthy controls, consistent with previous observationsŽ .van Boxel-Dezaire et al., 1999; Ozenci et al., 1999 .
Most importantly, we showed that treatment with oralSB significantly decreased the percentage of IL-12-produc-ing monocytes and DC in SP MS patients, and persistedfor at least 1 week after end of therapy. Although in threepatients the level of IL-12 at week 3 was clearly higherthan at baseline, this long-term effect of SB treatment
Ž .lasted beyond the half-life of SB 6 to 8 h for the majorityof patients, and could possibly be related to a long-lastingeffect on intracellular cAMP or another intracellular medi-ator. The short-term decrease in IL-12 is linked to an
Žincreased intracellular cAMP Panina-Bordignon et al.,.1997 .
We did not examine the effect of oral SB in normalŽsubjects because this was previously reported Panina-
.Bordignon et al., 1997 ; the purpose of this study was toexamine the effect of this drug on the cytokine dynamicsin SP MS patients.
Although IL-12 was measured after a potent, non-Ž .physiological stimulus IFN-gqLPS , this same stimulus
led to different results in normal individuals and in SP MS

( )K. Makhlouf et al.rJournal of Neuroimmunology 117 2001 156–165 163
patients. In vivo, IL-12-secreting cells are activated at thesite of inflammation through T cell secreted cytokines orthrough CD40–CD154 ligation. The different response tostimulus-induced IL-12 production between monocytes ofnormal controls and SP MS patients in our study mayreflect a different state of AreadinessB of these cells, withincreased IL-12 mRNA level as previously reported in SP
Ž .MS patients van Boxel-Dezaire et al., 1999 .In a small subset of five patients, we also found that
treatment with oral SB induced an increase in the percent-age of IL-10-, IL-4- and IL-5-producing T cells, and a
Ž .decrease in IFN-g-producing T cells not shown . Giventhat SB increases cAMP, part of this preliminary finding isconsistent with the fact that cAMP-responsive elements areinvolved in the transcriptional activation of the human
Ž .IL-10 gene Platzer et al., 1999 . Cyclic AMP and othersubstances that increase cAMP upregulate IL-10 expres-
Žsion in human T cells Benbernou et al., 1997; Shin et al.,. Ž .1998 and monocytes Platzer et al., 1995, 1999 both in
vivo and in vitro. Moreover, IL-10 and IL-12 are part of animmunoregulatory circuit, as demonstrated in the animal
Ž .model EAE Segal et al., 1998 , where the disease-promo-ting effects of IL-12 are antagonized by the production ofIL-10, which in turn is regulated by the endogenous IL-12production. This is also in accordance with our data,showing an increased IL-12 and a decreased IL-10 produc-tion in monocytes of untreated SP MS patients. In anotherstudy, PBMC from MS patients had increased levels ofIL-12 p40 and decreased levels of IL-10 mRNA compared
Ž .with controls van Boxel-Dezaire et al., 1999 . Further-more, the IFN-b-1b-induced inhibition of IL-12 expression
Žin PBMC from MS patients is IL-10-dependent Wang et.al., 2000 .
The CD40–CD154 interaction is critical for the IL-12Žproduction by monocytes under Ag-driven onditions Bal-
.ashov et al., 1997; Cella et al., 1996 , but monocytes canproduce IL-12 via a CD40–CD154-independent pathway
Ž .in response to LPS DeKruyff et al., 1997 . We found thatSB decreased CD40 and to a lesser extent CD80 expres-sion by monocytes in vitro but not in vivo. This may be adose-related effect, since SB decreased CD40 and CD80expression in vitro at high concentrations only, which areunlikely to be reached in the serum with the doses usedduring our trial. It is known that CD80 expression is
Žincreased on B cells during MS relapses while CD86.expression remains stable , and that IFN-b-1b therapy
decreases CD80 expression and increases CD86 expressionŽ .Genc et al., 1997 .
Whereas in vitro treatment with db-cAMP induced asignificant decrease in the percentage of IL-12-producingmonocytes in a dose-dependent manner, in vitro treatmentwith increasing SB doses did not show the same effect, assignificantly decreased and comparable IL-12 levels werereached with all the doses. This is very similar to the factthat in vivo, a daily dose of 12 mg did not show anyadditional IL-12-lowering effect when compared to a daily
4 mg dose, suggesting a saturation of the IL-12 loweringeffect of SB.
Another class of drugs, the phosphodiesterase inhibitorsŽ .PDEI , are interesting to compare to SB and b -agonists2
in general, as they also target cAMP and increase itsintracytoplasmic level. While the b -agonists bind to a G2
protein-coupled transmembrane receptor that induces anincrease in cAMP upon activation of adenylate cyclase, thePDEI increase cAMP by inhibiting its degradation by
Ž .phosphodiesterases PDE . Among the 10 known differentfamilies of PDE, PDE3, PDE4 and to a lesser extendPDE7 are predominantly expressed by human immune
Ž .cells Ekholm et al., 1997; Bloom and Beavo, 1996 .Rolipram, a PDE4 inhibitor mainly known as antidepres-sant in humans, is the most extensively studied PDEI, and
Ž .is shown to prevent EAE in rats Sommer et al., 1995 andŽ .in non-human primates Genain et al., 1995 . It also
decreases in vitro the synovial cells IL-12 production inŽ .rheumatoid arthritis patients Kitagawa et al., 1999 . An
additional protective mechanism for rolipram in EAE inmice is its ability to reduce the blood–brain barrier perme-
Ž .ability Folcik et al., 1999 . An in vitro study using bloodof MS patients and healthy controls has recently shownthat PDE4 and PDE3 inhibitors have complex inhibitoryeffects on Th1-mediated immunity, leaving Th2-mediated
Žresponses mostly unaffected or enhanced Bielekova et al.,.2000 . Like SB, rolipram in vitro decreases CD80 expres-
sion on the surface of LPS-stimulated monocytes from MSpatients and healthy subjects when added for 12 h to the
Ž .culture media Bielekova et al., 2000 . In the same in vitroŽsettings, rolipram increases CD86 expression Bielekova et
.al., 2000 . We found a transient decrease in CD86 expres-sion in vivo after oral SB but not in vitro. This differencemay be an indirect effect of decreased IL-12 and IFN-gexpression, as suggested by Creery et al., 1996. The samegroup reported that LPS-stimulated monocytes down-regu-lated CD86 and up-regulated CD80 expression. Anotherlikely explanation could simply be a difference in theinduction kinetics of this marker between in vivo and invitro environments, or related to the experimental settingsŽ24 vs. 12 h stimulation, with and without IFN-g in addi-
.tion to LPS .Several reports have shown that the CD80rCD86 cos-
timulatory system is somehow altered in MS: high num-bers of CD80q B cells in the CSF of MS patientsŽ .Svenningsson et al., 1997 , and in the blood during
Ž .attacks Genc et al., 1997 , but also increased CD86 inŽ .blood monocytes Kouwenhoven et al., 2001 .
Our data suggest that SB induces a Th2 switch mainlythrough a cAMP-mediated decrease of IL-12 production inthe APC both in vitro and in vivo, and decreases CD80expression on the surface of monocytes in vitro.
It may seem paradoxical that SB, a drug mainly used inasthma which is a Th2 disease, induces a Th2 shift;however, this may explain the enhanced late asthmaticresponse to inhaled antigens in asthmatic subjects regularly

( )K. Makhlouf et al.rJournal of Neuroimmunology 117 2001 156–165164
Ž .treated with b -agonists Baghat et al., 1996 . It is also2
possible that the use of b -agonists during pregnancy2
andror in early life increases the risk of developing aller-gic diseases by skewing the immunologic response to
Žallergens towards a Th2 cytokine profile Panina-Bordig-.non et al., 1997 .
Thus, oral SB given at low therapeutic doses decreasesthe percentage of IL-12-producing monocytes and DC inSP MS patients, while it increases the percentage of IL-10-expressing T cells. This novel observation has impor-tant therapeutic implications for MS and other immunemediated diseases associated with increased IL-12 produc-tion. SB has a known safety profile, is well tolerated,inexpensive, and easy to administer and monitor. Our dataprovide support for the use of b -agonists in the treatment2
of MS alone or as adjunct therapy to improve the efficacyof current treatments.
Acknowledgements
This work was supported by the NIH grantsŽ . Ž .N01A105411 SJK and HLW , 1P01NS38037 HLW , the
NMSS pilot grant PP 0643 and by the Nancy Davis CenterWithout Walls.
References
Anonymous, 2000. Physician Desk Reference. Medical Economics Com-pany, Montvale, p. 2832.
Arnason, B.G., Brown, M., Maselli, R., Karaszewski, J., Reder, A., 1988.Blood lymphocyte beta-adrenergic receptors in multiple sclerosis.Ann. N. Y. Acad. Sci. 540, 585–588.
Baghat, R., Swystem, V., Cockroft, D., 1996. Salbutamol-induced in-crease of airway responsiveness to allergen and reduced protectionversus methacoline: dose response. J. Allergy Clin. Immunol. 97,47–52.
Balashov, K.E., Smith, D.R., Khoury, S.J., Hafler, D.A., Weiner, H.L.,1997. Increased interleukin 12 production in progressive multiplesclerosis: induction by activated CD4q T cells via CD40 ligand.Proc. Natl. Acad. Sci. U. S. A. 94, 599–603.
Benbernou, N., Esnault, S., Shin, H.C., Fekkar, H., Guenounou, M.,1997. Differential regulation of IFN-gamma, IL-10 and induciblenitric oxide synthase in human T cells by cyclic AMP-dependentsignal transduction pathway. Immunology 91, 361–368.
Bielekova, B., Lincoln, A., McFarland, H., Martin, R., 2000. Therapeuticpotential of phosphodiesterase-4 and -3 inhibitors in Th1-mediatedautoimmune diseases. J. Immunol. 164, 1117–1124.
Bloom, T.J., Beavo, J.A., 1996. Identification and tissue-specific expres-sion of PDE7 phosphodiesterase splice variants. Proc. Natl. Acad. Sci.U. S. A. 93, 14188–14192.
Cella, M., Scheidegger, D., Palmer, L.K., Lane, P., Lanzavecchia, A.,Alber, G., 1996. Ligation of CD40 on dendritic cells triggers produc-tion of high levels of interleukin-12 and enhances T cell stimulatorycapacity: T-T help via APC activation. J. Exp. Med. 184, 747–752.
Comabella, M., Balashov, K., Issazadeh, S., Smith, D., Weiner, H.L.,Khoury, S.J., 1998. Elevated interleukin-12 in progressive multiplesclerosis correlates with disease activity and is normalized by pulsecyclophosphamide therapy. J. Clin. Invest. 102, 671–678.
Creery, W., Diaz-Mitoma, F., Filion, L., Kumar, A., 1996. Differentialmodulation of B7-1 and B7-2 isoform expression on human mono-cytes by cytokines which influence the development of T helper cell
Ž .phenotype. Eur. J. Immunol. 26 6 , 1273–1277.DeKruyff, R.H., Gieni, R.S., Umetsu, D.T., 1997. Antigen-driven but not
lipopolysaccharide-driven IL-12 production in macrophages requirestriggering of CD40. J. Immunol. 158, 359–366.
Ekholm, D., Hemmer, B., Gao, G., Vergelli, M., Martin, R., Manganiello,V., 1997. Differential expression of cyclic nucleotide phosphodi-esterase 3 and 4 activities in human T cell clones specific for myelinbasic protein. J. Immunol. 159, 1520–1529.
Folcik, V.A., Smith, T., O’Bryant, S., Kawczak, J.A., Zhu, B., Sakurai,H., Kajiwara, A., Staddon, J.M., Glabinski, A., Chernosky, A.L.,Tani, M., Johnson, J.M., Tuohy, V.K., Rubin, L.L., Ransohoff, R.M.,1999. Treatment with BBB022A or rolipram stabilizes the blood-brainbarrier in experimental autoimmune encephalomyelitis: an additionalmechanism for the therapeutic effect of type IV phosphodiesteraseinhibitors. J. Neuroimmunol. 97, 119–128.
Genain, C.P., Roberts, T., Davis, R.L., Nguyen, M.H., Uccelli, A.,Faulds, D., Li, Y., Hedgpeth, J., Hauser, S.L., 1995. Prevention ofautoimmune demyelination in non-human primates by a cAMP-specific phosphodiesterase inhibitor. Proc. Natl. Acad. Sci. U. S. A.92, 3601–3605.
Ž .Genc, K., Dona, D.L., Reder, A.T., 1997. Increased CD80 q B cells inactive multiple sclerosis and reversal by interferon beta-1b therapy. J.Clin. Invest. 99, 2664–2671.
Karp, C., Biron, C., Irani, D., 2000. Interferon beta in multiple sclerosis:Ž .is IL-12 suppression the key? Immunol. Today 21 1 , 24–28.
Khoury, S.J., Weiner, H.L., Hafler, D.A., 1996. Immunological mecha-Ž .nisms in multiple sclerosis. In: Cook, S.D. Ed. , Handbook of
Multiple Sclerosis. Marcell Dekker, New York, pp. 145–155.Kitagawa, M., Mitsui, H., Nakamura, H., Yoshino, S., Miyakawa, S.,
Ochiai, N., Onobori, M., Suzuki, H., Sumida, T., 1999. Differentialregulation of rheumatoid synovial cell interleukin-12 production bytumor necrosis factor alpha and CD40 signals. Arthritis Rheum. 42,1917–1926.
Kouwenhoven, M., Teleshova, N., Ozenci, V., Press, R., Link, H., 2001.Monocytes in multiple sclerosis: phenotype and cytokine profile. J.Neuroimmunol. 112, 197–205.
Leonard, J.P., Waldburger, K.E., Goldman, S.J., 1995. Prevention ofexperimental autoimmune encephalomyelitis by antibodies againstinterleukin 12. J. Exp. Med. 181, 381–386.
Macatonia, S.E., Hosken, N.A., Litton, M., Vieira, P., Hsieh, C.S.,Culpepper, J.A., Wysocka, M., Trinchieri, G., Murphy, K.M., O’Garra,A., 1995. Dendritic cells produce IL-12 and direct the development ofTh1 cells from naive CD4q T cells. J. Immunol. 154, 5071–5079.
Magram, J., Connaughton, S.E., Warrier, R.R., Carvajal, D.M., Wu,C.Y., Ferrante, J., Stewart, C., Sarmiento, U., Faherty, D.A., Gately,M.K., 1996. IL-12-deficient mice are defective in IFN gamma produc-tion and type 1 cytokine responses. Immunity 4, 471–481.
Malfait, A.M., Malik, A.S., Marinova-Mutafchieva, L., Butler, D.M.,Maini, R.N., Feldmann, M., 1999. The beta2-adrenergic agonistsalbutamol is a potent suppressor of established collagen-inducedarthritis: mechanisms of action. J. Immunol. 162, 6278–6283.
Nicoletti, F., Patti, F., Cocuzza, C., Zaccone, P., Nicoletti, A., Di Marco,R., Reggio, A., 1996. Elevated serum levels of interleukin-12 inchronic progressive multiple sclerosis. J. Neuroimmunol. 70, 87–90.
Ozenci, V., Kouwenhoven, M., Huang, Y.M., Xiao, B., Kivisakk, P.,Fredrikson, S., Link, H., 1999. Multiple sclerosis: levels of inter-leukin-10-secreting blood mononuclear cells are low in untreatedpatients but augmented during interferon-beta-1b treatment. Scand. J.Immunol. 49, 554–561.
Panina-Bordignon, P., Mazzeo, D., Lucia, P.D., D’Ambrosio, D., Lang,R., Fabbri, L., Self, C., Sinigaglia, F., 1997. Beta2-agonists preventTh1 development by selective inhibition of interleukin 12. J. Clin.Invest. 100, 1513–1519.
Panitch, H.S., Hirsch, R.L., Haley, A.S., Johnson, K.P., 1987. Exacerba-

( )K. Makhlouf et al.rJournal of Neuroimmunology 117 2001 156–165 165
tions of multiple sclerosis in patients treated with gamma interferon.Lancet 1, 893–895.
Platzer, C., Meisel, C., Vogt, K., Platzer, M., Volk, H.D., 1995. Up-regu-lation of monocytic IL-10 by tumor necrosis factor-alpha and cAMPelevating drugs. Int. Immunol. 7, 517–523.
Platzer, C., Fritsch, E., Elsner, T., Lehmann, M.H., Volk, H.D., Prosch,S., 1999. Cyclic adenosine monophosphate-responsive elements areinvolved in the transcriptional activation of the human IL-10 gene inmonocytic cells. Eur. J. Immunol. 29, 3098–3104.
Reid, S.D., Penna, G., Adorini, L., 2000. The control of T cell responsesby dendritic cell subsets. Curr. Opin. Immunol. 12, 114–121.
Ž .Segal, B., Dwyer, B., Shevach, E., 1998. An interleukin IL -10rIL-12immunoregulatory circuit controls susceptibility to autoimmune dis-
Ž .ease. J. Exp. Med. 187 4 , 537–546.Shin, H.C., Benbernou, N., Fekkar, H., Esnault, S., Guenounou, M.,
Ž .1998. Regulation of IL-17, IFN-gamma and IL-10 in human CD8 qT cells by cyclic AMP-dependent signal transduction pathway. Cy-tokine 10, 841–850.
Sommer, N., Loschmann, P.A., Northoff, G.H., Weller, M., Steinbrecher,A., Steinbach, J.P., Lichtenfels, R., Meyermann, R., Riethmuller, A.,Fontana, A. et al., 1995. The antidepressant rolipram suppressescytokine production and prevents autoimmune encephalomyelitis. Nat.Med. 1, 244–248.
Svenningsson, A., Dotevall, L., Stemme, S., Andersen, O., 1997. In-creased expression of B7-1 costimulatory molecule on cerebrospinalfluid cells of patients with multiple sclerosis and infectious centralnervous system disease. J. Neuroimmunol. 75, 59–68.
Thomas, R., Lipsky, P.E., 1994. Human peripheral blood dendritic cell
subsets. Isolation and characterization of precursor and mature anti-gen-presenting cells. J. Immunol. 153, 4016–4028.
Trembleau, S., Germann, T., Gately, M.K., Adorini, L., 1995. The role ofIL-12 in the induction of organ-specific autoimmune diseases. Im-munol. Today 16, 383–386.
Trinchieri, G., 1995. Interleukin-12: a proinflammatory cytokine withimmunoregulatory functions that bridge innate resistance andantigen-specific adaptive immunity. Annu. Rev. Immunol. 13, 251–276.
van Boxel-Dezaire, A., Hoff, S., van Oosten, B., Verweij, C., Drager, A.,Ader, H., van Houwelingen, J., Barkhof, F., Polman, C., Nagelkerken,L., 1999. Decreased interleukin-10 and increased interleukin-12p40mRNA are associated with disease activity and characterize different
Ž .disease stages in multiple sclerosis. Ann. Neurol. 45 6 , 695–703.Wang, X., Chen, M., Wandinger, K.P., Williams, G., Dhib-Jalbut, S.,
2000. IFN-beta-1b inhibits IL-12 production in peripheral bloodmononuclear cells in an IL-10-dependent mechanism: relevance toIFN-beta-1b therapeutic effects in multiple sclerosis. J. Immunol. 165,548–557.
Wiegmann, K., Muthyala, S., Kim, D., Arnason, B., Chelmicka-Schorr,E., 1995. Beta-adrenergic agonists suppress chronicrrelapsing experi-
Ž .mental allergic encephalomyelitis CREAE in Lewis rats. J. Neu-roimmunol. 56, 201–206.
Zoukos, Y., Kidd, D., Woodroofe, M.N., Kendall, B.E., Thompson, A.J.,Cuzner, M.L., 1994. Increased expression of high affinity IL-2 recep-tors and beta-adrenoceptors on peripheral blood mononuclear cells isassociated with clinical and MRI activity in multiple sclerosis. Brain117, 307–315.