oral administration of squid lecithin-transphosphatidylated phosphatidylserine improves memory...
TRANSCRIPT

Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
Contents lists available at ScienceDirect
Progress in Neuro-Psychopharmacology & BiologicalPsychiatry
j ourna l homepage: www.e lsev ie r .com/ locate /pnp
Oral administration of squid lecithin-transphosphatidylatedphosphatidylserine improves memory impairment in aged rats
Bombi Lee a, Bong-Jun Sur a,b, Jeong-Jun Han c, Insop Shim a,b, Song Her d, Yang-Seok Lee e,Hye-Jung Lee a,b, Dae-Hyun Hahm a,b,⁎a Acupuncture and Meridian Science Research Center, Kyung Hee University, Seoul, 130-701, Republic of Koreab BK21 PLUS Korean Medicine Science Center, College of Korean Medicine, Kyung Hee University, Seoul 130-701, Republic of Koreac Glonet BU, Doosan Co., Yongin-si, Gyeonggi-do 448-795, Republic of Koread Korea Basic Science Institute Chuncheon Branch, Gangwon-do 200-701, Republic of Koreae Dept. of Meridian Medical Science, Graduate School of Kyung Hee University, Gyeonggi-do 448-795, Republic of Korea
Abbreviations: PS, lecithin-derived phosphatidylsericholine-transphosphatidylated PS; cAMP, cyclic adenosinnervous system; BC-PS, bovine cerebral cortex PS; BSE, bothies; Soy-PS, soybean-derived PS; PC, phosphatidylcholinEPA, eicosapentaenoic acid; AchE, acetylcholinesterase; CMWM, Morris water maze; PA, phosphatidic acid; PBS, pavidin–biotin-peroxidase; DAB, 3,3′-diaminobenzidine;polymerase chain reaction; CHT, choline transporter; mAline receptor type 1; GAPDH, glyceraldehyde-3-phosphateysis of variance.⁎ Corresponding author at: Acupuncture and Merid
Department of Korean Medicine, Kyung Hee University, 1Seoul 130-701, Republic of Korea. Tel.: +82 2 961 0366;
E-mail address: [email protected] (D.-H. Hahm).
http://dx.doi.org/10.1016/j.pnpbp.2014.07.0040278-5846/© 2014 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 3 May 2011Received in revised form 9 July 2014Accepted 9 July 2014Available online 22 July 2014
Keywords:AgingCholinergic systemMemoryPhosphatidylserineSquid
Recently, lecithin-derived phosphatidylserine (PS), which originates frommarine life, has receivedmuch attentionas a viable alternative to bovine cerebral cortex PS. In this study, the use of squid phosphatidylcholine-transphosphatidylated PS (SQ-PS) was evaluated through examination of its ameliorating effects on age-associated learning and memory deficits in rats. Aged rats were orally administered SQ-PS (10, 20, or 50 mg/kgper day) once a day for seven days 30 min prior to behavioral assessment in a Morris water maze. SQ-PS adminis-tration produced significant dose-dependent improvements in escape latency forfinding the platform in theMorriswatermaze in the aged rats even though Soy-PS administration also exhibited comparable improvementswith SQ-PS. Biochemical alterations in the hippocampal cholinergic system, including changes in choline acetyltransferaseand acetylcholinesterase immunoreactivity, were consistent with the behavioral results. In addition, SQ-PS treat-ment significantly restored age-associated decreases of choline transporter and muscarinic acetylcholine receptortype 1 mRNA expression in the hippocampus. These results demonstrate that orally administered SQ-PS dose-dependently aids in the improvement of memory deficits that occur during normal aging in rats. This suggeststhat SQ-PS may be a useful therapeutic agent in the treatment of diminished memory function in elderly people.
© 2014 Elsevier Inc. All rights reserved.
1. Introduction
Lecithin-derived phosphatidylserine (PS) is a constituent of mem-brane phospholipids that are especially abundant in the brain. It elicitsa variety of neurochemical activities including the release and turnoverof neurotransmitters such as acetylcholine, norepinephrine, and dopa-mine (Casamenti et al., 1979). PS also results in increased levels of
ne; SQ-PS, squid phosphatidyl-e monophosphate; CNS, centralvine spongiform encephalopa-e; DHA, docosahexaenoic acid;hAT, choline acetyltransferase;hosphate-buffered saline; ABC,RT-PCR, reverse transcription-ChR-M1, muscarinic acetylcho-dehydrogenase; ANOVA, anal-
ian Science Research Center,Hoegi-dong, Dongdaemun-gu,
fax: +82 2 963 2175.
glucose and cAMP in the CNS (Bruni and Toffano, 1982), particularlyin amnesiac patients (Pepeu et al., 1986).
A number of clinical studies report that the oral administration ofbovine cerebral cortex PS (BC-PS) improves the cognitive function of el-derly people suffering from age-associated memory impairments,Alzheimer's disease, and senile dementia with few side effects (Engelet al., 1992). However, the use of BC-PS asmedicine or a dietary supple-ment is now discouraged due to the risk of contamination by the prionthat is likely to cause bovine spongiform encephalopathies (BSE) suchas mad cow disease and Creutzfeldt–Jakob disease (Prusiner, 1991). Inaddition, only about 3 g of PS can be obtained from one bovine cortexwhich is too small an amount to adequately maintain a PS supply forthe market at the best price (Kato-Kataoka et al., 2010).
To solve these problems, soybean-derived PS (Soy-PS), which is aBSE risk-free PS, was enzymatically produced from soybean phosphati-dylcholine (PC) by phospholipase D-catalyzed transphosphatidylationof soybean PC (Jorissen et al., 2002). Some studies have demonstratedthat the oral administration of Soy-PS to aged rat results in a significantimprovement of memory and other cognitive functions (Jorissen et al.,2010; Suzuki et al., 2001). However, compared with BC-PS which con-tains approximately 10% DHA (Chen et al., 1989), Soy-PS, which does

Table 1Fatty acid composition of lipids in standard chow diet used in thisstudy.
Fatty acid Content (%)
Palmitic acid (C16:0) 20.51Stearic acid (C18:0) 8.37Oleic acid (C18:1) 32.04Linoleic acid (C18:2) 28.76Linolenic acid (C18:3) 2.45EPA (C20:5) 0.41DHA (C22:6) 0.48Other fatty acids 6.98SUM 100
The values were expressed in % of total fatty acid in chow diet.
2 B. Lee et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
not contain DHA species, provides little benefit with regard to age-associated memory impairment (Schreiber et al., 2000). Because ofsome clinical reports indicating a little insufficient activity of soybeanPS in improving memory and other cognitive functions in elderlyhuman, much researcher is still investigating to find a new alternativeof BS-PS (Schreiber et al., 2000). Therefore, in recent years, a mixtureof squid skin PC-derived PS has received attention as a potential alterna-tive to BC-PS since it has substantial amounts of docosahexaenoic(DHA) and eicosapentaenoic acid (EPA; Hosokawa et al., 2000; Leeet al., 2010), active components in the beneficial effects of PS. Further-more, many reports show that DHA plays an important role in brainfunction and neurodevelopment (Kim et al., 2011; Mills et al., 2011).Therefore, it may be postulated that phospholipids sourced from ani-mals improve learning and memory ability to a greater extent thanthose sourced from soybeans, which is probably due to DHA action atthe sn-2 position (Sommer Hartvigsen et al., 2004). DHA is released asa result of pancreatic phospholipase action in the body and seems toexert a synergistic effect with the residual PS moiety on the improve-ment of brain function (Vakhapova et al., 2010).
However, the direct action of orally administered marine-derived PSin the CNS is still in question and its mechanisms remain poorly under-stood (Drago et al., 1981; Lee et al., 2010). The current study investigatesthe effect of squid derived PS (SQ-PS) on age-related spatial memory inaged rats in the Morris water maze (MWM) by means of assessing im-provement of SQ-PS as a nutritional supplement for age-associatedmemory impairment. To evaluate the relationship between orally ad-ministered SQ-PS and cholinergic function, we examined the neuropro-tective effects of SQ-PS on the central acetylcholine system by assessingChAT and AchE immunohistochemistry of hippocampal neurons assamemethods described in our previous study (Lee et al., 2010). The ef-fect of a Soy-PS treatment was also analyzed on the same parameters inyoung and aged rats in order to compare their effects with those obtain-ed with the SQ-PS treatment.
2. Materials and methods
2.1. Animals
Adult male Sprague-Dawley (SD) rats weighing 260–280 g (sixweeks old) and 500–530 g (15 months old) were obtained from CharlesRiver Laboratories (Yokohama, Japan). The rats were housed in alimited-access rodent facility with up to five rats per polycarbonatecage. The room controls were set to maintain temperature at 22 ± 2 °Cwith relative humidity at 55 ± 15%. Cages were lit using artificial lightfor 12 h each day. Sterilized drinking water and standard chow dietwere supplied ad libitum to each cage throughout the experiments. Theanimal experiments were conducted in accordance with the Guide forthe Care and Use of Laboratory Animals (NIH Publications No. 80-23, re-vised in 1996) and were approved by the Kyung Hee University Institu-tional Animal Care and Use Committee. All animal experiments began atleast seven days after the animals arrived.
2.2. Experimental groups
The rats were randomly divided into six groups as follows: 1) saline-treated normal adult group (younggroup; sixweek old subjects, n=8);2) saline-treated aged group (Aged group; sham control; 15 month oldsubjects, n=10); 3) 10 mg/kg SQ-PS-treated aged group (AG-SQ-PS10,n = 7); 4) 20 mg/kg SQ-PS-treated aged group (AG-SQ-PS20, n = 7);and 5) 50 mg/kg SQ-PS-treated aged group (AG-SQ-PS50, n = 7) and6) 50 mg/kg Soy-PS-treated aged group (AG-Soy-PS50, n = 7). Thedaily doses were determined based on previous studies by Lee et al.(2010). The SQ-PS and Soy-PS used in this study were manufacturedand kindly provided by Doosan Co. Glonet BU (Youngin-si, Korea). Theapplied SQ-PS formula contains 92% PS, 1% PC, and 5% phosphatidicacid (PA). In the case of Soy-PS, it contained 90% PS, 2% PC, and 6% PA.
Fatty acid compositions of the standard chow diets for the rats andphospholipids such as SQ-PS and Soy-PS are shown in Table 1 and 2, re-spectively. Because total fat content of the standard chow diet is 5% per100 g diet, and DHA and EPA contents are 0.48% and 0.41% of total fattyacids in chow diet (Table 1.), 0.024 g DHA and 0.021 g EPA in 100 g dietare daily supplied. In the case of the AG-SQ-PS50 (50 mg PS/kg bodyweight) group, orally administrated amount of squid PS is 10 mg perday per rat weighing 200 g. Because 40.7 g DHA and 13.5 g EPA are in-cluded in 100 g squid PS (Table 2), 4.07 mg DHA and 1.35 mg EPA aredaily administered to each rat weighing 200 g. In summary, less than0.024 g DHA and 0.21 g EPA might be ingested in the standard dietgroup as a control, respectively, and 4.07 mg DHA and 1.35 mg EPAwere orally administered to the rats in the SQ-PS diet groups. The PSwas homogenously suspended in saline and orally administrated tothe subjects once a day for seven consecutive days. MWM testing wasperformed 30 min subsequent to every PS treatment on each of theseven days.
2.3. Morris water maze test
2.3.1. Morris water maze apparatusThe MWM test was performed using a polypropylene circular pool
(painted white internally, 2.0 m in diameter and 0.35 m high). The poolcontained water, which was maintained at a temperature of 22 ± 2 °C.During MWM testing, a platform 15 cm in diameter was located 1.5 cmbelow thewater in one of four sections of the pool andwas approximate-ly 50 cm from the sidewalls. The pool was divided into four quadrants ofequal area andwas surrounded by several external cues. A digital cameramounted to the ceiling above the pool was connected to a computerizedrecording system equipped with a tracking program (S-MART: PanLabCo., Barcelona, Spain) which permitted on-and off-line automated track-ing of the paths taken by the rat.
2.3.2. Hidden platform trial for acquisition testThe animals received three trials per day. The rats were trained to
find the hidden platform, which remained in a fixed location through-out the test. The trials lasted for a maximum of 180 s, with the time ittook to find the submerged platform recorded each time. The animalswere tested in this way three trails per day for six days, and received a60 s-probe trial on the seventh day. Finding the platform was definedas staying on it for at least 4 s before the acquisition time of 180 sended. Because of rats that crossed the platform without stopping(jumping immediately into the water) were left to swim. After stayingon the platform for 10 s, the ratswas gently picked up using a steel spat-ula, returned to its home cage, and allowed to warm up and dry offunder a 125-W heat lamp. If the rat failed to find the platform in the al-lotted time itwas placed onto the platform for 20 s and assigned a laten-cy of 180 s. The water was stirred in between each trial to removeolfactory traces of previous swim patterns. The entire procedure tookplace over seven consecutive days and each animal had three trainingtrials per day with a 30–40 min interval.

Table 2Fatty acid composition of phosphatidylserine originated from squid (SQ-PS) and soybean (Soy-PS).
Myristic(14:0)
Palmitic(16:0)
Palmitoleic(16:1)
Stearic(18:0)
Oleic(18:1)
Linoleic(18:2)
Linolenic(18:3)
Eicosanoic(20:1)
EPAa
(20:5)DHAb
(22:6)Others
Soy-PS – 17.7 1.3 1.3 14.4 61.2 1.4 – – – 2.7SQ-PS 1.0 23.3 0.8 5.5 2.7 0.2 – 4.0 13.5 40.7 8.3
a EPA: Eicosapentaenoic acid.b DHA: Docosahexaenoic acid. The values were expressed in g/100 g SQ-PS or Soy-PS.
3B. Lee et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
2.3.3. Visible platform (cued trial) testThe cued trial (three trials per rat) was performed on the first day to
assess the rats' motivation to escape from the water and to evaluatetheir sensorimotor integrity. The platform was placed in the fixed loca-tion of quadrant and had a visible black cue on it. The animalwas placedin the pool and given 90 s to reach the platform which could be identi-fied by the visible black platform above the surface of thewater. Latencyto reach the visible platform was automatically calculated.
2.3.4. Probe trial for retention testFor the probe trial, each rat was placed into the water diagonally
from the target quadrant and, for 60 s, was allowed to search thewater from which the platform had been removed. The time (% oftotal time) spent searching for the platform in the former platformquadrant versus the other three quadrants was measured for each rat.
2.4. Open field test
Prior to water maze testing the rats were individually housed in arectangular container made of dark polyethylene (45 × 45 × 35 cm) ina dimly lit room equipped with a video camera above the center ofthe floor and locomotor activity was measured. Locomotor activitywas monitored by a computerized video-tracking system using the S-MART program (PanLab Co., Barcelona, Spain). The animals wereallowed to adapt for 10 min in the container before the distance theytraveled (in meters) was recorded during a 5 min test. They also ana-lyzed the total distance crossed by rats in the open field test (in additionto the number of lines crossed between the squares area). The Plexiglassquare arenawas divided into nine equal-sized squares on the floor. Thenumber of lines crossed (with all four paws) between the square areawas recorded for 5 min.
2.5. Choline acetyltransferase (ChAT) immunohistochemistry
For immunohistochemical study, each animal was deeply anesthe-tizedwith sodiumpentobarbital (80 mg/kg, by intraperitoneal injection),and thenperfused through the ascending aorta using normal saline (0.9%)followed by 300ml of 4% paraformaldehyde in 0.1M phosphate-bufferedsaline (PBS). The brainwas removed and cut. The sectionswere immuno-stained for ChAT expression using the avidin–biotin–peroxidase (ABC)complexmethod. Briefly, the sectionswere incubatedwithprimary rabbitanti-ChAT antibody (1:2000 dilution; Cambridge Research BiochemicalsCo., Bellingham, UK) in 0.1 M PBS with 0.3% Triton X-100 (PBST) for 72h at 4 °C. The sections were incubated for 120 min at room temperaturewith biotinylated anti-rabbit goat IgG (for the anti-ChAT antibody).Both secondary antibodies were obtained from Vector Laboratories Co.(Burlingame, CA, USA) and diluted at 1:200 in PBST containing 2%normal goat serum. To visualize immunoreactivity the sections were in-cubated for 90 min in the ABC reagent (Vectastain Elite ABC kit; VectorLabs. Co., Burlingame, CA, USA) and then incubated in a solution contain-ing 3′-diaminobenzidine (DAB; Sigma-AldrichChemical Co., St. Louis,MO,USA). The sections were viewed at a 200× magnification with the num-ber of cells within 100 × 100 mm2 grids counted by observers blind tothe experimental groups. Hippocampal area cells were obtained
according to the stereotactic atlas of Paxinos and Watson (1986). Thecells were counted in three sections per rat within the hippocampal area.
2.6. Acetylcholinesterase (AchE) immunohistochemistry
For AchE immunohistochemistry, the sections were washed inPBS and incubated in a solution with 25 mg of acetylthiocholine io-dine for 1 h. The solution was composed of 32.5 ml of 0.1 M sodiumhydrogen phosphate buffer (NaH2PO4·H2O, pH 6.0), 2.5 ml of 0.1 Msodium citrate, 5 ml of 30 mM copper sulfate, 5 ml of 5 mM potassi-um ferricyanide, and 5 ml of distilled water. The color of the mixingsolution was green. The densities of stained nuclei of the hippocam-pal cells were measured using a Scion image program (Scion Co.,Frederick, MD, USA). The sections were viewed at 200× magnifica-tion and the number of cells within 100 × 100 mm2 grids was count-ed by observers blind to the experimental groups. Hippocampal areacells were obtained according to the stereotactic atlas of Paxinos andWatson (1986). The cells were counted in three sections per ratwithin the hippocampal area.
2.7. Total RNA preparation and RT-PCR analysis
For RT-PCR analysis, the hippocampus from each of four rats in eachgroup were just randomly selected after behavioral test and isolated.After decapitation, the brain was quickly removed and stored at−80 °C until use. The total RNA was isolated from the brain sampleusing a TRIzol® reagent (Invitrogen Co., Carlsbad, CA, USA) and RNAwas extracted according to the supplier's instruction. ComplementaryDNA was synthesized from total RNA with a reverse transcriptase(Takara Co., Shiga, Japan). Choline transporter (CHT) and muscarinicacetylcholine receptor type 1 (mAChR-M1) mRNA expression levelswere determined by the reverse transcription-polymerase chain reac-tion (RT-PCR). RT-PCR was performed using a PTC-100 programmablethermal controller (MJ Research, Inc.,Watertown,MA, USA). All primerswere designed using published mRNA sequences and primer designsoftware (Primer 3; The Whitehead Institute for Biomedical Research,Cambridge, MA, USA; www.genome.wi.mit.edu) offered through theirwebsite. The following sequences were used: for GAPDH (409 bp), (for-ward) 5′-ATC CCA TCA CCA TCT TCC AG-3′ and (reverse) 5′-CCT GCTTCA CCA CCT TCT TG-3′; for CHT1 (90 bp), (forward) 5′-CCC ATG CGCTTT CCA TAG AT-3′ and (reverse) 5′-AGG TGG CCT GTT TTT TGC AA-3′; for mAChR-M1 (331 bp), (forward) 5′-CCT CCC AAA AGC TCC CCA-3′ and (reverse) 5′-TGT CCC GGA AGG CTT TGT-3′. The PCR productswere separated on 1.2% agarose gels and stainedwith ethidiumbromideafter which the density of each band was evaluated using an image-analyzing system (i-Max™, CoreBio System Co., Seoul, Korea). Comple-mentary DNA expression levels were determined by calculating the rel-ative density of each CHT1 or mAChR-M1 band to GAPDH.
2.8. Sample preparation and measurement of fatty acid profile
After behavioral tests for 1 week, the rat were anesthetized with so-dium pentobarbital (50 mg/kg, i.p.), the hippocampus was separated aspreviously described (Hashimoto et al., 2002). The tissues were storedat −80 °C by flash-freezing in liquid N2 until use. Total lipid was

4 B. Lee et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
extracted fromhippocampuswithmixture solvent (chloroform,metha-nol, Distilled water, 65/35/4, v/v/v) according to previously described(Folch et al., 1957), and fatty acid compositionwas analyzed by gas–liq-uid chromatography (Ohkubo and Tanaka, 2010). The esters preparedby transmethylation with BF3/methanol. An Agilent 7890A series gaschromatograph equipped with flame ionization detector (FID) andDB-225 capillary column (30m×0.25mm i.d.) (Agilent J &WScientific,CA, USA) was used. Column temperature was raised from 175 to 220 °Cat 2.5 °C/min. Injector and detector temperatures were 250 °C and260 °C, respectively. Carrier gas was helium, and hydrogen and airwere supplied to the FID. Fatty acids were identified by comparing re-tention times with the lipid standard (Sigma-Aldrich). The same proto-col as above was used for analyzing fatty acid composition of the ratdiet.
2.9. Statistical analyses
The experimental results are expressed as the mean ± standarderror (SE). The behavioral data were calculated and analyzed by re-peated measures analysis of variance (ANOVA) using SPSS (Version13.0; SPSS Inc., Chicago, IL, USA). The statistical significance amonggroups was further analyzed using Tukey's post-hoc test. Immuno-histochemical data and PCR analysis were also analyzed by a one-way ANOVA followed by Tukey's post-hoc test. Statistical significancewas set at p b 0.05.
3. Results
3.1. Effect of SQ-PS during the visible platform trial ofMorriswatermaze test
To exclude the possibility of impairing the vision of the subjects andaltering the motivation to escape the water due to SQ-PS administra-tion, a cued version of theMWM testwas performed and the swimmingtime to reach the visible platformwasmeasured (Fig. 1C).When trainedto a visible platform there were no significant differences betweengroups [F(5,44) = 0.291; p = 0.881]. The latency to find the visibleplatform did not differ among groups on the first, second, or third trials(Fig. 1C). On the second and third trials, the latency was markedly re-duced in all groups, as compared to the first trial. All rats, irrespectiveof grouping, were able to locate the visible platform more rapidly asthe trials progressed.
3.2. Effect of SQ-PS during the hidden platform trial of the Morris watermaze test
Rats in the Young group rapidly learned the location of the sub-merged hidden platform and reached it within 20 s onDay 6 of the trials(Fig. 1C). The Aged group showed amarked retardation in escape laten-cy, probably due tomemory deficits resulting from age-induced impair-ments of learning and memory. Analysis of the training data byrepeated-measures ANOVA showed that escape latency differed signif-icantly among the groups when the times were averaged over all ses-sions [F(5,40) = 9.216, p b 0.001]. During the experiment, escapelatency decreased over time [F(5,200) = 142.076, p b 0.001], andthere was a significant interaction between experimental groups andtime [F(25,200) = 2.939, p b 0.05]. Tukey's post-hoc test revealed thatrats in both the AG-SQ-PS20 and AG-SQ-PS50 groups had significantlyreduced swimming latency compared with subjects in the controlgroup (AG-SQ-PS20 group: p b 0.01 on Days 5 and 6; AG-SQ-PS50group: p b 0.01 on Days 5 and 6; Fig. 1C). The Aged groupwas not signif-icantly different from other groups in terms of mean swimming speed,as calculated by dividing the total swim distance by the latency[F(5,44) = 0.860, p = 0.517] (Fig. 1E). Total distance traveled in eachgroup was closely associated with escape latency in this task (data notshown). Based on these results, the SQ-PS-treated aged rats showed
greater improvements in acquisition during the hidden-platform trialand reached the platform quicker than the saline-treated aged rats.
3.3. Effect of SQ-PS during the probe trial of the Morris water maze test
To examine the spatial memory of rats, performance in the probe testonDay 7was analyzed by comparing the percentage of time spent swim-ming to the expected position of the platform (Fig. 1D). The time spentswimming around was significantly reduced in the rats that swam tothe target area where the platform had been located [F(5,44) = 5.213,p b 0.01]. The aged rats displayed severely impaired spatial performancein the MWM (p b 0.01). Rats in the 20 mg/kg (p b 0.05) and 50 mg/kgSQ-PS-treated (p b 0.01) groups spent more time around the platformarea than did those in the Aged group. SQ-PS treatment significantly at-tenuated the age-induced deficit of learning and memory demonstratedin thewatermaze. Thus, SQ-PS-treated rats showed a significant amelio-ration in thememory retention test because they spentmore time in thequadrant where the platform was formerly located and swam over theformer location of the platform more frequently. Our results haveshowed that swimming latency time in the AG-SQ-PS20 group was sim-ilar to that of the AG-Soy-PS50 group.
3.4. Effect of SQ-PS during the open field test
Analysis of performance in the open-field test by a parametric one-way ANOVA showed no significant differences between groups interms of memory deficit-related locomotor activity [F(2,24) = 0.870,p = 0.439] or the total number of line crossings [F(2,24) = 1.857, p =0.190] (Fig. 2). This indicates that SQ-PS administration did not affectpsychomotor function as measured by performance in the MWM test.
3.5. Effect of SQ-PS on hippocampal choline acetyltransferase
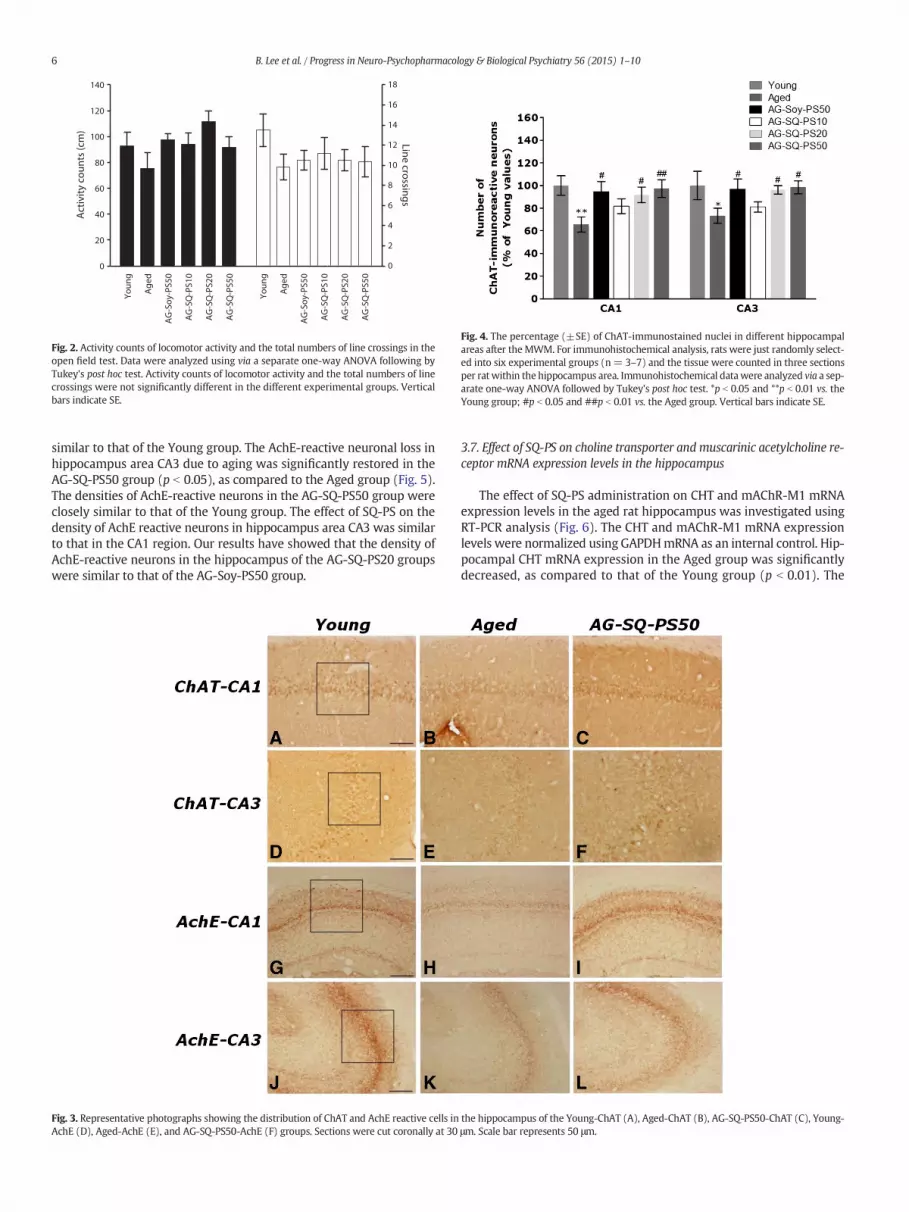
Following the behavioral tasks, brain tissue samples from the subjectswere analyzed using immunohistochemistry to investigate the effect ofSQ-PS administration onneuronal cell loss due to aging. ChAT immunore-activity analysis in the CA1 and CA3 areas of the hippocampus are shownin Fig. 3. Comparison of the numbers of ChAT-immunoreactive neuronsusing a one-way ANOVA revealed a significant difference among thegroups [F(5,71) = 3.957, p b 0.01; CA1 and F(5,71) = 4.104, p b 0.01;CA3]. The brains of the Aged group showed significant neuronal cell lossin the CA1 area of the hippocampus, as compared to the Young group(p b 0.01). The number of ChAT-immunoreactive neurons significantlyincreased in hippocampal region CA1 in the AG-SQ-PS20 (p b 0.05) andthe AG-SQ-PS50 groups (p b 0.01), as compared to the Aged group(Fig. 4). The number of ChAT-immunoreactive neurons significantly in-creased in hippocampal area CA3 in the AG-SQ-PS20 (p b 0.05) and theAG-SQ-PS50 groups (p b 0.05), as compared to the Aged group (Fig. 4).Losses of ChAT-immunoreactivity in aged rats were significantly restoredby the SQ-PS administration, and the number of ChAT-immunopositiveneurons was closely similar to that of the Young group. Our results haveshowed that ChAT activity in the hippocampus of the AG-SQ-PS20group was similar to that of AG-Soy-PS50 group.
3.6. Effect of SQ-PS on hippocampal acetylcholinesterase
The density of AchE-immunopositive fibers in the CA1 and CA3 areaof the rat hippocampus was significantly reduced in the Aged group, ascompared to the Young group (Fig. 3). Comparison of the numbers ofAchE-positive neurons density using a one-way ANOVA revealed a sig-nificant difference among the groups [(5,71) = 6.019, p b 0.001; CA1and F(5,71) = 2.721, p b 0.05; CA3]. The AchE-reactive neuronal lossin hippocampus area CA1 due to aging was significantly restored inthe AG-SQ-PS20 (p b 0.05) and the AG-SQ-PS50 groups (p b 0.01), ascompared to the Aged group (Fig. 5). The density of AchE-reactive neu-rons in the AG-SQ-PS20 group and the AG-SQ-PS50 group was closely
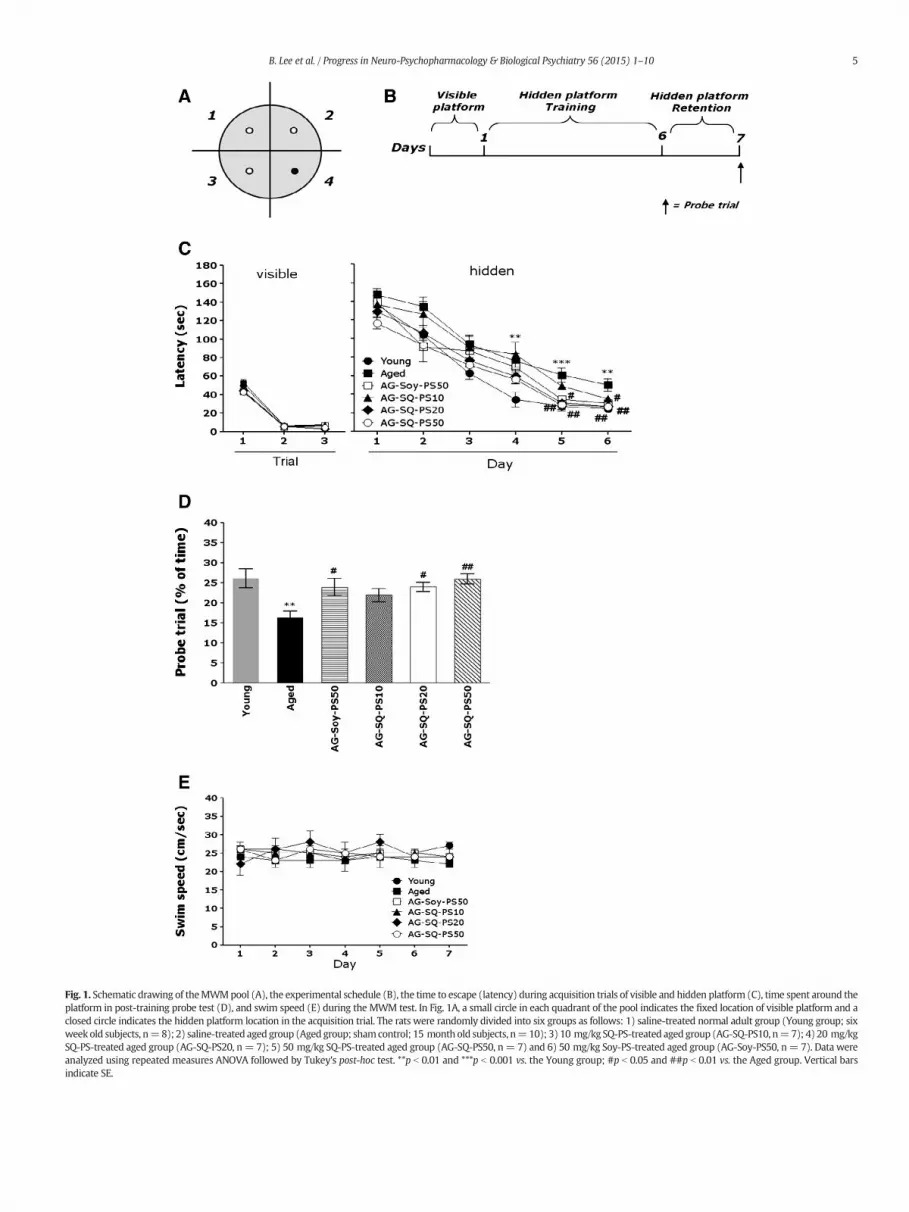
Fig. 1. Schematic drawing of theMWMpool (A), the experimental schedule (B), the time to escape (latency) during acquisition trials of visible and hidden platform (C), time spent around theplatform in post-training probe test (D), and swim speed (E) during the MWM test. In Fig. 1A, a small circle in each quadrant of the pool indicates the fixed location of visible platform and aclosed circle indicates the hidden platform location in the acquisition trial. The rats were randomly divided into six groups as follows: 1) saline-treated normal adult group (Young group; sixweek old subjects, n=8); 2) saline-treated aged group (Aged group; shamcontrol; 15 month old subjects, n=10); 3) 10 mg/kg SQ-PS-treated aged group (AG-SQ-PS10, n=7); 4) 20 mg/kgSQ-PS-treated aged group (AG-SQ-PS20, n= 7); 5) 50 mg/kg SQ-PS-treated aged group (AG-SQ-PS50, n= 7) and 6) 50 mg/kg Soy-PS-treated aged group (AG-Soy-PS50, n= 7). Data wereanalyzed using repeated measures ANOVA followed by Tukey's post-hoc test. **p b 0.01 and ***p b 0.001 vs. the Young group; #p b 0.05 and ##p b 0.01 vs. the Aged group. Vertical barsindicate SE.
5B. Lee et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
Youn
g
Age
d
AG
-Soy
-PS5
0
AG
-SQ
-PS1
0
AG
-SQ
-PS2
0
AG
-SQ
-PS5
0
Youn
g
Age
d
AG
-Soy
-PS5
0
AG
-SQ
-PS1
0
AG
-SQ
-PS2
0
AG
-SQ
-PS5
0
0
20
40
60
80
100
120
140
0
2
4
6
8
10
12
14
16
18A
ctiv
ity c
ount
s (c
m)
Line crossings
Fig. 2. Activity counts of locomotor activity and the total numbers of line crossings in theopen field test. Data were analyzed using via a separate one-way ANOVA following byTukey's post hoc test. Activity counts of locomotor activity and the total numbers of linecrossings were not significantly different in the different experimental groups. Verticalbars indicate SE.
Fig. 4. The percentage (±SE) of ChAT-immunostained nuclei in different hippocampalareas after the MWM. For immunohistochemical analysis, rats were just randomly select-ed into six experimental groups (n= 3–7) and the tissue were counted in three sectionsper ratwithin the hippocampus area. Immunohistochemical datawere analyzed via a sep-arate one-way ANOVA followed by Tukey's post hoc test. *p b 0.05 and **p b 0.01 vs. theYoung group; #p b 0.05 and ##p b 0.01 vs. the Aged group. Vertical bars indicate SE.
6 B. Lee et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
similar to that of the Young group. The AchE-reactive neuronal loss inhippocampus area CA3 due to aging was significantly restored in theAG-SQ-PS50 group (p b 0.05), as compared to the Aged group (Fig. 5).The densities of AchE-reactive neurons in the AG-SQ-PS50 group wereclosely similar to that of the Young group. The effect of SQ-PS on thedensity of AchE reactive neurons in hippocampus area CA3 was similarto that in the CA1 region. Our results have showed that the density ofAchE-reactive neurons in the hippocampus of the AG-SQ-PS20 groupswere similar to that of the AG-Soy-PS50 group.
Fig. 3. Representative photographs showing the distribution of ChAT and AchE reactive cells inAchE (D), Aged-AchE (E), and AG-SQ-PS50-AchE (F) groups. Sections were cut coronally at 30
3.7. Effect of SQ-PS on choline transporter and muscarinic acetylcholine re-ceptor mRNA expression levels in the hippocampus
The effect of SQ-PS administration on CHT and mAChR-M1 mRNAexpression levels in the aged rat hippocampus was investigated usingRT-PCR analysis (Fig. 6). The CHT and mAChR-M1 mRNA expressionlevels were normalized using GAPDHmRNA as an internal control. Hip-pocampal CHT mRNA expression in the Aged group was significantlydecreased, as compared to that of the Young group (p b 0.01). The
the hippocampus of the Young-ChAT (A), Aged-ChAT (B), AG-SQ-PS50-ChAT (C), Young-μm. Scale bar represents 50 μm.
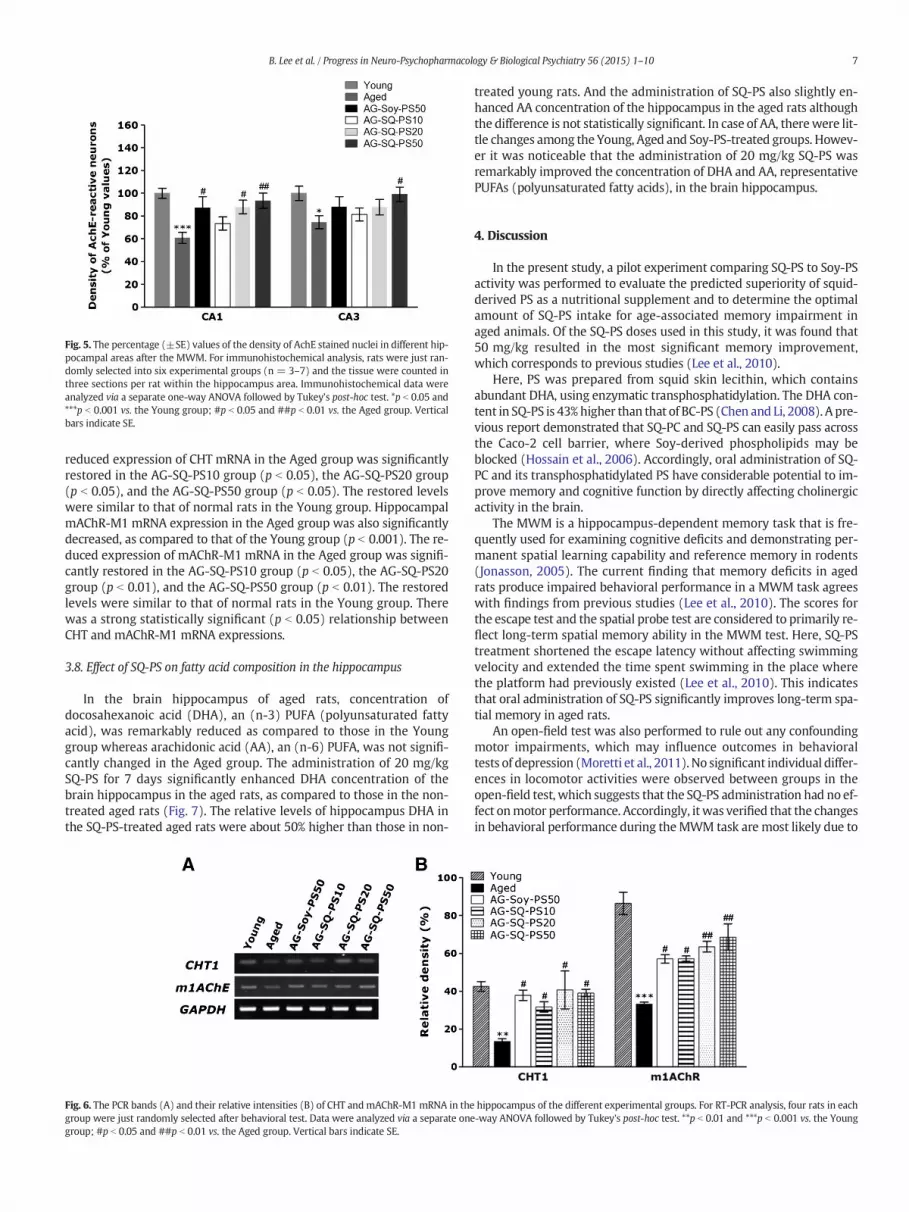
Fig. 5. The percentage (±SE) values of the density of AchE stained nuclei in different hip-pocampal areas after the MWM. For immunohistochemical analysis, rats were just ran-domly selected into six experimental groups (n = 3–7) and the tissue were counted inthree sections per rat within the hippocampus area. Immunohistochemical data wereanalyzed via a separate one-way ANOVA followed by Tukey's post-hoc test. *p b 0.05 and***p b 0.001 vs. the Young group; #p b 0.05 and ##p b 0.01 vs. the Aged group. Verticalbars indicate SE.
7B. Lee et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
reduced expression of CHT mRNA in the Aged group was significantlyrestored in the AG-SQ-PS10 group (p b 0.05), the AG-SQ-PS20 group(p b 0.05), and the AG-SQ-PS50 group (p b 0.05). The restored levelswere similar to that of normal rats in the Young group. HippocampalmAChR-M1 mRNA expression in the Aged group was also significantlydecreased, as compared to that of the Young group (p b 0.001). The re-duced expression of mAChR-M1 mRNA in the Aged group was signifi-cantly restored in the AG-SQ-PS10 group (p b 0.05), the AG-SQ-PS20group (p b 0.01), and the AG-SQ-PS50 group (p b 0.01). The restoredlevels were similar to that of normal rats in the Young group. Therewas a strong statistically significant (p b 0.05) relationship betweenCHT and mAChR-M1 mRNA expressions.
3.8. Effect of SQ-PS on fatty acid composition in the hippocampus
In the brain hippocampus of aged rats, concentration ofdocosahexanoic acid (DHA), an (n-3) PUFA (polyunsaturated fattyacid), was remarkably reduced as compared to those in the Younggroup whereas arachidonic acid (AA), an (n-6) PUFA, was not signifi-cantly changed in the Aged group. The administration of 20 mg/kgSQ-PS for 7 days significantly enhanced DHA concentration of thebrain hippocampus in the aged rats, as compared to those in the non-treated aged rats (Fig. 7). The relative levels of hippocampus DHA inthe SQ-PS-treated aged rats were about 50% higher than those in non-
Fig. 6. The PCR bands (A) and their relative intensities (B) of CHT and mAChR-M1 mRNA in thegroup were just randomly selected after behavioral test. Data were analyzed via a separate onegroup; #p b 0.05 and ##p b 0.01 vs. the Aged group. Vertical bars indicate SE.
treated young rats. And the administration of SQ-PS also slightly en-hanced AA concentration of the hippocampus in the aged rats althoughthe difference is not statistically significant. In case of AA, therewere lit-tle changes among the Young, Aged and Soy-PS-treated groups. Howev-er it was noticeable that the administration of 20 mg/kg SQ-PS wasremarkably improved the concentration of DHA and AA, representativePUFAs (polyunsaturated fatty acids), in the brain hippocampus.
4. Discussion
In the present study, a pilot experiment comparing SQ-PS to Soy-PSactivity was performed to evaluate the predicted superiority of squid-derived PS as a nutritional supplement and to determine the optimalamount of SQ-PS intake for age-associated memory impairment inaged animals. Of the SQ-PS doses used in this study, it was found that50 mg/kg resulted in the most significant memory improvement,which corresponds to previous studies (Lee et al., 2010).
Here, PS was prepared from squid skin lecithin, which containsabundant DHA, using enzymatic transphosphatidylation. The DHA con-tent in SQ-PS is 43%higher than that of BC-PS (Chen and Li, 2008). A pre-vious report demonstrated that SQ-PC and SQ-PS can easily pass acrossthe Caco-2 cell barrier, where Soy-derived phospholipids may beblocked (Hossain et al., 2006). Accordingly, oral administration of SQ-PC and its transphosphatidylated PS have considerable potential to im-prove memory and cognitive function by directly affecting cholinergicactivity in the brain.
The MWM is a hippocampus-dependent memory task that is fre-quently used for examining cognitive deficits and demonstrating per-manent spatial learning capability and reference memory in rodents(Jonasson, 2005). The current finding that memory deficits in agedrats produce impaired behavioral performance in a MWM task agreeswith findings from previous studies (Lee et al., 2010). The scores forthe escape test and the spatial probe test are considered to primarily re-flect long-term spatial memory ability in the MWM test. Here, SQ-PStreatment shortened the escape latency without affecting swimmingvelocity and extended the time spent swimming in the place wherethe platform had previously existed (Lee et al., 2010). This indicatesthat oral administration of SQ-PS significantly improves long-term spa-tial memory in aged rats.
An open-field test was also performed to rule out any confoundingmotor impairments, which may influence outcomes in behavioraltests of depression (Moretti et al., 2011). No significant individual differ-ences in locomotor activities were observed between groups in theopen-field test, which suggests that the SQ-PS administration had no ef-fect onmotor performance. Accordingly, it was verified that the changesin behavioral performance during theMWM task are most likely due to
hippocampus of the different experimental groups. For RT-PCR analysis, four rats in each-way ANOVA followed by Tukey's post-hoc test. **p b 0.01 and ***p b 0.001 vs. the Young

Fig. 7. Concentration of fatty acids, such as palmitic (C16:0, A), stearic (C18:0, B), oleic (C18:1), linoleic (C18:2), linolenic (C18:3), arachidonic acid (C20:4n6) and DHA (C22:6n3) in thebrain hippocampus of SQ-PS-treated or Soy-PS-treated rats. **p b 0.01 vs. the Young group; ##p b 0.01 vs. the Aged group. Vertical bars indicate SE. The numbers of animals for analyzingeach fatty acid composition are 3 as indicated in the bracket in the figure.
8 B. Lee et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
improved memory function instead of changes in sensorimotor func-tion, such as motor output and limb flexibility.
The hippocampus is a medial temporal lobe structure that has beenimplicated in the consolidation of declarative memory in humans andspatial memory in rodents (Burger et al., 2007; Miyagawa et al., 1998).Impairments in the hippocampus generally result in diminished spatiallearning ability. Thus, most age-related psychosomatic disorders are as-sociatedwith decreasedworkingmemory function that is directly relat-ed to alterations of hippocampal function (Casamenti et al., 1991). Here,it is demonstrated that SQ-PS treatment protects aged rats from spatialworking memory deficits and attenuates the decrease in AchE andChAT-immunoreactivity in the hippocampus. It is likely that the ob-served learning- and memory-related improvements of SQ-PS-treatedanimals in the MWM are associated with the attenuation of hippocam-pal cell loss, the increase in central cholinergic function, and the preven-tion of degeneration in the cholinergic neuronal population of the basalforebrain (Millan et al., 1988).
On the other hand, the current results show that the altered expres-sion levels of CHT andmAChR-M1mRNA in the hippocampus are asso-ciated with the memory deficits due to normal aging. Thus, this might
also contribute to reduced changes in cholinergic markers in the hippo-campus as well. It was also demonstrated that SQ-PS treatment signifi-cantly attenuates decreases in CHT andmAChR-M1mRNA expression inthe hippocampus of aged rats. Several studies have suggested an associ-ation between hippocampal CHT andmAChR-M1 expression andmem-ory performance, particularly in water maze tests (Falkenberg et al.,1992; Schaaf et al., 2000). These findings suggest that the memory-improving activity of SQ-PS can be verified by using several behavioralparadigms, and that this type of amelioration is exerted throughmodu-lation of cholinergic neurons in the brain.
The expression and activation of AchE and ChAT regulate the dy-namic concentration of ACh in cholinergic synapses in the brain. Thus,the hippocampal expression of AchE and ChAT and its correlation withthe memory performance of aged rats was examined. Previous bio-chemical and behavioral evidence indicates that central cholinergictransmission declines with age (Gage et al., 1988) and with dementiaof the Alzheimer's type (Uabundit et al., 2010). This type of decline isclosely associated with cognitive disturbances. PS enhances cognitiveperformance not only in aged rats but also in elderly human subjectsshowing varied levels of age-dependent or disease-related memory

9B. Lee et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
impairment (Crook et al., 1991; Kato-Kataoka et al., 2010; Lee et al.,2010). The beneficial effects of PS could be related to the increases inthe central cholinergic function (Lee et al., 2000).
High proportions of PUFAs in the brain are crucial for maintaining thestructure and physiological function of the central nervous system(Chang et al., 2001). A deficit of dietary DHA is known to contribute to in-flammatory signaling, apoptosis and neuronal dysfunction, and is closelyassociatedwith a cognitive decline in the elderly people and in thosewithage-related neurological disorders (Babenko and Semenova, 2010). Also,animal studies using experimental diets lacking (n-3) PUFA demonstrat-ed that those diets elicited a significant reduction of DHA level in cerebralmembranes, which means that it is eventually associated with distur-bances of neural functions such as visual acuity, attention, learning, andmemory (Bourre et al., 1989). Dietary supplementation of DHA resultedin the restoration of age-related declines of cognitive functions by stimu-lating synthesis of new polyunsaturated phosphatidylserine species inthe brain cerebral cortex and hippocampus (Babenko and Semenova,2010). It was also reported that AA supplementation also amelioratedcognitive dysfunction caused by either organic brain damage or agingnot neurodegenerative diseases like Alzheimer's disease (Kotani et al.,2006). The purpose of this experiment was to determinewhether hippo-campus FA composition can be altered as a result of the repeated admin-istration of SQ-PS, and whether the changes of FA composition can affectthe age-related monoaminergic and cholinergic neurotransmission sys-tems in the brain tissues. Our results clearly showed that the administra-tion of SQ-PS significantly increased the concentration of DHA and AA inthe hippocampal regions of the rat brains. Accordingly, it can be sug-gested that a positive correlation was observed between percentage oftime spent swimming to the expected position of the platform in theMWM and the concentration of DHA and AA in the brain. Because exog-enous phospholipids may serve as an extra supply for endogenous phos-pholipid turnover in the cellmembranes (Claro et al., 2006), thememory-improving activity of SQ-PS can be attributed to a biochemical role playedby PS. PS may elicit the fast and stale turnover of phospholipids in themembranes of brain tissues. In addition, PS is an important activator ofseveral enzymes including protein kinase C, which plays a crucial rolein synaptic plasticity and information storage (Orr et al., 1992; Van derZee andDouma, 1997; Vance, 2003). It has also been suggested that PS in-fluences the regulation of glutamate receptor-mediated signaling (Gagnéet al., 1996). Glutamate receptors are thought to be involved in the regu-lation of long-lasting potentiation of synaptic transmission believed tounderlie information storage in the brain (Bliss and Collingridge, 1993).Accordingly, it is possible that PS directly acts on downstream cascadesof intracellular signaling in memory consolidation processes.
In the present study, administration of Soy-PS exhibited a compara-ble effect of improving memory ability with that of SQ-PS in the agedrats, although SQ-PS has plenty of (n-3) PUFA such as DHA and EPAthat are not detected in Soy-PS. It indicates that the contribution ofDHA and EPA in SQ-PS to the maintenance of structural and physiolog-ical function of the brain was not remarkable in the aged rats. Becausehigh proportion of (n-3) PUFA in the brain membranes is importantfor maintaining normal metabolism and function of the brain, DHAand EPA in SQ-PS administeredmay charge a certain role ofmaintainingbrain integrity if those fatty acids are released individually from PS afterintake and can be moved to the brain tissues in their intact forms. Also,since the contents of (n-3) PUFA in the brains gradually decreased withaging, those lipids should be supplied to the brain either directly fromfood supplements such as maternal milk and fish oil or by internal syn-thesis in small amounts from α-linolenic acid. In this respect, a dietary-induced depletion of (n-3) PUFA is important to investigate the effect ofdiet including (n-3) PUFA such as DHA and EPA on brain integrity andmemory function in animal study. Accordingly, a long-term (n-3)PUFA deficient feeding over two generations was often exploited in an-imal studies even though severe deficiency of (n-3) PUFA is scarcelycaused in human (Aïd et al., 2005; Innis and de La Presa, 2001; Kimuraet al., 2011). Unfortunately, we did not use (n-3) PUFA-deficient diet
for feeding the rats in this study. The standard chow diet used in thisstudy includes small but substantial amounts of (n-3) PUFA as shownin Table 1. It seems that (n-3) PUFA in the chow diet played a certainrole of maintaining the basal level of brain function in the aged ratseven though DHA contents in the brain hippocampus in the aged ratswas noticeably reduced as compared with those in young rats in thepresent study.
5. Conclusion
The present study demonstrates that memory and cognitive deficitsinduced by age-induced hippocampal lesions are closely related to thedegeneration of cholinergic neurons in the rat hippocampus. Moreover,orally administered SQ-PS significantly ameliorates learning and mem-ory deficits through the recovery of cholinergic activity in the brain. SQ-PS treatment improved performance in the spatial memory test andprotected hippocampal cholinergic neurons from age-associated de-struction. The attenuation of impairments of memory and cognitionby SQ-PS administration may be due to the restoration of cholinergicneurochemical activity. It is likely that SQ-PS is strongly effective inprotecting against memory-related neuronal degeneration in the brainand slowing the progression of memory deficits associated with variousneurodegenerative diseases.
Acknowledgments
This research was supported by a grant of the Korea HealthcareTechnology R&D Project, Ministry for Health, Welfare & Family Affairs(Grant No. A091037) and the National Research Foundation of Koreafunded by the Korean government (MEST) (2013R1A1A2008487 and2013R1A1A2063051), Republic of Korea.
References
Aïd S, Vancassel S, LinardA, LavialleM,Guesnet P. Dietary docosahexaenoic acid [22: 6(n-3)]as a phospholipid or a triglyceride enhances the potassium chloride-evoked release ofacetylcholine in rat hippocampus. J Nutr 2005;135(5):1008–113.
Babenko NA, Semenova YA. Effects of long-term fish oil-enriched diet on the sphingolipidmetabolism in brain of old rats. Exp Gerontol 2010;45(5):375–80.
Bliss TV, Collingridge GL. A synaptic model of memory: Long term potentiation in the hip-pocampus. Nature 1993;361(6407):31–9.
Bourre JM, Francois M, Youyou A, Dumont O, Piciotti M, Pascal G, Durand G. The effects ofdietary alpha-linolenic acid on the composition of nerve membranes, enzymatic ac-tivity, amplitude of electrophysiological parameters, resistance to poisons and perfor-mance of learning tasks in rats. J Nutr 1989;119(12):1880–92.
Bruni A, Toffano G. Lysophosphatidylserine, a short-lived intermediatewith plasmamem-brane regulatory properties. Pharmacol Res Commun 1982;14(6):469–82.
Burger C, Lopez MC, Feller JA, Baker HV, Muzyczka N, Mandel RJ. Changes in transcriptionwithin the CA1 field of the hippocampus are associated with age-related spatiallearning impairments. Neurobiol Learn Mem 2007;87(1):21–41.
Casamenti F, Mantovani P, Amaducci L, Pepeu G. Effect of phosphatidylserine on acetyl-choline output from the cerebral cortex of the rat. J Neurochem 1979;32(2):529–33.
Casamenti F, Scali C, Pepeu G. Phosphatidylserine reverses the age-dependent decrease incortical acetylcholine release: amicrodialysis study. Eur J Pharmacol 1991;194(1):11–6.
Chang MC, Contreras MA, Rosenberger TA, Rintala JJ, Bell JM, Rapoport SI. Chronicvalproate treatment decreases the in vivo turnover of arachidonic acid in brainphospholipids: a possible common effect of mood stabilizers. J Neurochem 2001;77(3):796–803.
Chen S, Li KW. Comparison of molecular species of various transphosphatidylatedphosphatidylserine (PS) with bovine cortex PS by mass spectrometry. Chem PhysLipids 2008;152(1):46–56.
Chen S, Benfenati E, Fanelli R, Kirschner G, Pregnolato F. Molecular species analysis ofphospholipids by negative ion fast atom bombardment mass spectrometry: applica-tion of surface precipitation technique. Biomed EnvironMass Spectrom 1989;18(12):1051–6.
Claro FT, Patti CL, Abílio VC, Frussa-Filho R, Silva RH. Bovine brain phosphatidylserine at-tenuates scopolamine induced amnesia in mice. Prog Neuropsychopharmacol BiolPsychiatry 2006;30(5):881–6.
Crook TH, Tinklenberg J, Yesavage J, Petrie W, Nunzi MG, Massari DC. Effects ofphosphatidylserine in age-associated memory impairment. Neurology 1991;41(5):644–9.
Drago F, Canonico PL, Scapagnini U. Behavioral effects of phosphatidylserine in aged rats.Neurobiol Aging 1981;2(3):209–13.
Engel RR, Satzger W, Günther W, Kathmann N, Bove D, Gerke S, et al. Double-blind cross-over study of phosphatidyl-serine vs. placebo in patients with early dementia ofAlzheimer type. Eur Neuropsychopharmacol 1992;2(2):149–55.

10 B. Lee et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 56 (2015) 1–10
Falkenberg T, Mohammed AK, Henriksson B, Persson H,Winblad B, Lindefors N. Increasedexpression of brain-derived neurotrophic factormRAN in rat hippocampus is associat-ed with improved spatial memory and enriched environment. Neurosci Lett 1992;138(1):153–6.
Folch J, Lees M, Sloane Stanley GH. A simple method for the isolation and purification oftotal lipides from animal tissues. J Biol Chem 1957;226(1):497–509.
Gage FH, Chen KS, Buzsaki G, Armstrong D. Experimental approaches to age-related cog-nitive impairments. Neurobiol Aging 1988;9(5-6):645–55. [Review].
Gagné J, Giguére C, Tocco G, Ohayon M, Thompson RF, Baudry M, et al. Effect ofphosphatidylserine on the binding properties of glutamate receptors in brain sectionsfrom adult and neonatal rats. Brain Res 1996;740(1–2):337–45.
Hashimoto M, Hossain S, Shimada T, Sugioka K, Yamasaki H, Fujii Y, Shido O, et al.Docosahexaenoic acid provides protection from impairment of learning ability inAlzheimer's disease model rats. J Neurochem 2002;81(5):1084–91.
HosokawaW, Shimatani T, KanadaT, InoueY, Takahashi K. Conversion to docosahexeanoicacid-containing phosphatidylserine from squid skin lecithin by phospholipase D-mediated transphosphatidylation. J Agric Food Chem 2000;48(10):4550–4.
Hossain Z, Kurihara M, Hosokawa M, Takahashi K. Docosahexeanoic acid andeicosapentaenoic acid-enriched phosphatidylcholine liposomes enhance thepermeability, transportation and uptake of phospholipids in Caco-2 cells. MolCell Biochem 2006;285(1–2):155–63.
Innis SM, de La Presa Owens S. Dietary fatty acid composition in pregnancy alters neuritemembrane fatty acids and dopamine in newborn rat brain. J Nutr 2001;13(1):118–22.
Jonasson Z. Meta-analysis of sex differences in rodent models of learning and memory: areview of behavioral and biological data. Neurosci Biobehav Rev 2005;28(8):811–25.
Jorissen BL, Brouns F, Van Boxtel MP, Riedel WJ. Safety of soy-derived phosphatidylserinein elderly people. Nutr Neurosci 2002;5(5):337–43.
Jorissen BL, Brouns F, Van Boxtel MP, Ponds RW, Verhey FR, Jolles J, et al. The influence ofsoy-derived phosphatidylserine on cognition in age-associated memory impairment.Nutr Neurosci 2010;4(2):121–34.
Kato-Kataoka A, Sakai M, Ebina R, Nonaka C, Asano T, Miyamori T. Soybean-derivedphosphatidylserine improves memory function of the elderly Japanese subjectswith memory complaints. J Clin Biochem Nutr 2010;47(3):246–55.
Kim HY, Moon HS, Cao D, Lee J, Kevala K, Jun S, et al. N-docosahexaenoylethanolamidepromotes development of hippocampal neurons. Biochem J 2011;435(2):327–36.
Kimura F, Ito S, Endo Y, Doisaki N, Koriyama T, Miyazawa T, Fujimoto K. Supplementationof DHA-rich microalgal oil or fish oil during the suckling period in mildly n−3 fattyacid-deficient rat pups. Lipids 2011;46(12):1101–10.
Kotani S, Sakaguchi E, Warashina S, Matsukawa N, Ishikura Y, Kiso Y, Yamashima T, et al.Dietary supplementation of arachidonic acid and docosahexaenoic acid improvescognitive dysfunction. Neurosci Res 2006;56(2):159–64.
Lee D, Hirashima N, Kirino Y. Rapid transbilayer phospholipid redistribution associatedwith exocytotic release of neurotransmitters from cholinergic nerve terminals isolat-ed from electric ray Narke japonica. Neurosci Lett 2000;291(1):21–4.
Lee B, Sur BJ, Han JJ, Shim I, Her S, Lee HJ, et al. Krill phosphatidylserine improves learningand memory in morris water maze in aged rats. Prog Neuropsychopharmacol BiolPsychiatry 2010;34(6):1085–93.
Millan F, Guidolin D, Polato P, Nunzi MG, Toffano G. Structural changes of basal forebraincholinergic neurons in the aged rats. Effect of phosphatidylserine administration. In:Pepeu G, Tomlinson B, Wischik CM, editors. New trends in aging research. Padova:Liviana Press; 1988. p. 221–31.
Mills JD, Hadley K, Bailes JE. Dietary supplementation with the omega-3 fatty aciddocosahexaenoic acid in traumatic brain injury. Neurosurgery 2011;68(2):474–81.
Miyagawa H, Hasegawa M, Fukuta T, Amano M, Yamada K, Nabeshima T. Dissociation ofimpairment between spatial memory, and motor function and emotional behavior inaged rats. Behav Brain Res 1998;91(1-2):73–81.
Moretti M, de Souza AG, de Chaves G, de Andrade VM, Romao PR, Gavioli EC, et al. Emo-tional behavior in middle-aged rats: implications for geriatric psychopathologies.Physiol Behav 2011;102(1):115–20.
Ohkubo T, Tanaka Y. Administration of DHA-PS to aged mice was suitable for increasinghippocampal PS and DHA ratio. J Oleo Sci 2010;59(5):247–53.
Orr JW, Keranen LM, Newton AC. Reversible exposure of the pseudosubstrate domain ofprotein kinase C by phosphatidylserine and diacylgliberol. J Biol Chem 1992;267(22):15263–6.
Paxinos G, Watson C. The rat brain in stereotaxic coordinates. New York: Academic Press;1986.
Pepeu G, Giovannelli L, Giovannini MG, Pedata F. Effect of phosphatidylserine on corticalacetylcholine release and calcium in adult and aging rats. In: Horrocks LA, Freysz I,Toffano G, editors. Biochem Mol PharmacolPadova: Liviana Press; 1986. p. 265–71.
Prusiner SB. Molecular biology of prion disease. Science 1991;252(5012):1515–21.[Review].
Schaaf MJ, De Kloet ER, Vreugdenhil E. Corticosterone effects on BDNF expression in thehippocampus. Implications for memory formation. Stress 2000;3(3):201–8.
Schreiber S, Kampf-Sherf O, Gorfine M, Kelly D, Oppenheim Y, Lerer B. An open trial ofplant-source derived phosphatidylserine for treatment of age-related cognitive de-cline. Isr J Psychiatry Relat Sci 2000;37(4):302–7.
Sommer Hartvigsen M, Mu H, Sørig Hougaard K, Lund SP, Xu X, Høy CE. Influence of die-tary triacylglycerol structure and level of n−3 fatty acids administered during devel-opment on brain phospholipids and memory and learning ability of rats. Ann NutrMetab 2004;48(1):16–27.
Suzuki S, Yamatoya H, Sakai M, Kataoka A, Furushiro M, Kudo S. Oral administration ofsoybean lecithin transphosphatidylated phosphatidylserine improves memory im-pairment in aged rats. J Nutr 2001;131(11):2951–6.
UabunditN,Wattanathorn J,Mucimapura S, IngkaninanK. Cognitive enhancement andneu-roprotective effects of Bacopamonnieri in Alzheimer's diseasemodel. J Ethnopharmacol2010;127(1):26–31.
Vakhapova V, Cohen T, Richter Y, Herzog Y, Korczyn AD. Phosphatidylserine containingomega-3 fatty acids may improve memory abilities in non-demented elderly withmemory complaints: a double-blind placebo-controlled trial. Dement Geriatr CognDisord 2010;29(5):467–74.
Van der Zee EA, Douma BR. Historical review of research on protein kinase C in learningand memory. Prog Neuropsychopharmacol Biol Psychiatry 1997;21(3):379–406.
Vance JE. Molecular and cell biology of phosphatidylserine and phosphatidylethanol-amine metabolism. Prog Nucleic Acid Res Mol Biol 2003;75(1):69–111.