biology of the world's largest invertebrate, the colossal squid
TRANSCRIPT

1
1
Biologyoftheworld’slargestinvertebrate,thecolossalsquid2
(Mesonychoteuthishamiltoni):ashortreview3
4
5
RuiRosa1*,VanessaM.Lopes1,MiguelGuerreiro1,KathrinBolstad2,JoséC.6
Xavier3,47
8
9
10
1MARE –Marine Environmental Sciences Centre, LaboratórioMarítimo da Guia,11
FaculdadedeCiênciasdaUniversidadedeLisboa,Portugal.12
2 Institute forApplied EcologyNewZealand,AucklandUniversity of Technology,13
Auckland,NewZealand.14
3MARE–MarineandEnvironmentalSciencesCentre(MARE),DepartmentofLife15
Sciences,UniversityofCoimbra,3004‐517Coimbra,Portugal.16
4 British Antarctic Survey, Natural Environment Research Council, High Cross,17
MadingleyRoad,CambridgeCB30ET,UnitedKingdom.18
19
20
*Correspondingauthor:[email protected]
22
23
24
25
26
27
28
29
30
31
32
33

2
Abstract34
Thepresentstudyaimstoreviewthecurrentbiologicalknowledgeonthelargest35
(heaviest) living invertebrate, the colossal squid Mesonychoteuthis hamiltoni36
(Robson,1925).Thissquid isknowntobeendemicoff theSouthernOcean(SO),37
withacircumpolardistributionspreading fromtheAntarcticcontinentup to the38
Sub‐AntarcticFront.Smalljuveniles(<40mmmantlelength)aremainlyfoundfrom39
the surface to 500 m, and the late juvenile stages are assumed to undergo40
ontogeneticdescenttodepthsreaching2000m.Thus,thisgiantspendsmostpartof41
itslifeinthemeso‐andbathypelagicrealms,whereitcanreachatotallengthof642
meters. The maximum weight recorded so far was 495 kg. Mesonychoteuthis43
hamiltoni is presently reported from the diets of 17 different predator species,44
comprising penguins, sea birds, fishes and marine mammals, and feeds on45
myctophids,Patagoniantootfish,sleepersharkandothersquids.Isotopicanalysis46
placesthecolossalsquidasoneofthetoppredatorsintheSO.Basedonphylogenetic47
inferences,itisassumedthatthissquidisnotcapableofhigh‐speedpredator–prey48
interactions,butitisratheranambushpredator.Theireyes,thelargestintheplanet,49
seemedtohaveevolvedtodetectverylargepredators(e.g.spermwhales)rather50
thantodetectmatesorpreysatlongdistances.Ontheotherhand,andlikemany51
otherdeep‐seaanimals,itpossesseslight‐emittingorgansontheeyes,whichmay52
provide ventral camouflage and counter‐illumination or some sort of deceptive53
techniquetocapturetheirprey.AlthoughM.hamiltoniisquiteabundantintheSO,54
its life existence remains one of the ocean’s great mysteries. This unique55
invertebrategiantcontinuestoattractconsiderableattentionfrommediaandthe56
publicingeneral,butitsstudyalsoconstitutesavaluablesourceofinsightintothe57
biophysicalprinciplesbehindbody‐sizeevolution.58
59
Keywords:colossalsquid,Mesonychoteuthishamiltoni,cranchiids,SouthernOcean;60
gigantism61
62

3
Introduction63
TheSouthernOceanconsistsofasystemofdeep‐seabasinsseparatedbythe64
ScotiaRidge,theKerguelenplateau(Indiansection)andtheMacquarieRidge(South65
ofNewZealand).WhilethesouthislimitedbytheAntarcticcontinent,thenorthern66
part is surrounded by theAntarctic Polar Frontal Zone (APFZ), beginning at the67
Antarcticconvergence/AntarcticPolarFront(APF).Themainsurfacecurrentisthe68
AntarcticCircumpolarCurrent(ACC),whichrunseastwardsaroundthecontinent69
(Orsietal.1995);Mooreetal.(1997);(Mooreetal.1999).Theseassociatedfronts70
createanobstacletonorth‐south“meridional”circulation,andthediscontinuities71
of temperature and salinity in the APFZ constitute an important barrier in72
biogeographicalinteractions(Barnesetal.2006).Nonetheless,Antarcticlifeisrich73
anddiversewithhighlevelsofendemism(Arntzetal.1994;ClarkeandJohnston74
2003).75
Withinthisuniqueregion,cephalopodsareknowntoplayakeyroleinthe76
marinefoodwebs,becausetheyconstituteavitalalternatefoodsourcetoAntarctic77
krillEuphausiasuperba(CollinsandRodhouse2006).Theyareimportantprey,and78
thussustainlargepopulationsofpenguins,procellariformbirds,sealsandtoothed79
whales(Clarke1980;Xavieretal.2002a;CherelandDuhamel2004;Chereletal.80
2004;CherelandHobson2005;CherelandHobson2007;Chereletal.2007;Xavier81
etal.2011).Yet,despitetheirimportantecologicalrole,thebasicknowledgeofthe82
bio‐ecologyofSouthernOceancephalopodsisstillpoorlyknown(Xavieretal.1999;83
Xavier et al. 2014; Alvito et al. 2015; Guerreiro et al. 2015), probably due to an84
absenceofafisheryindustry.Thismayberelatedtothelownumberofexploitable85
species(Xavieretal.2007),lackofbasicknowledgeofthespecies’biology,stocks’86

4
unpredictability,andtherelativeinaccessibilityofthesestocksforexplorationand87
study(Rodhouseetal.1996;Rodhouse1998;CollinsandRodhouse2006).88
Themajorityof the resident SouthernOcean cephalopod fauna comprises89
endemic species of octopods (cirrate and incirrate), and oegopsid squids,which90
differgreatlyfromthetaxafoundatlowerlatitudes.Thereisalsoatotalabsenceof91
myopsid squids and cuttlefish, and normally abundant families are rare. Unique92
Southern Ocean taxa include the squid families Psychroteuthidae and93
Batoteuthidae, and the genera Psychroteuthis, Kondakovia, Alluroteuthis,94
Slosarczykovia, Batoteuthis and Mesonychoteuthis. Moreover, several of these95
cephalopods occupy similar niches to (and compete with) some pelagic top96
predators,namelysharksandlargefishes(RodhouseandWhite1995;Rodhouseet97
al.1996;CherelandDuhamel2004;CherelandHobson2005).Oneoftheseisthe98
mostmassive living cephalopod ‐ the colossal squidMesonychoteuthis hamiltoni99
(Robson1925)(Fig.1).ThissquidisknowntobeaSouthernOceanendemic,and100
althoughitispreyedbymanytoppredators,itsbasicbiologyandecologyremain101
oneof theocean’sgreatmysteries.This reviewaims to summarize thecurrently102
scattered bio‐ecological knowledge of the elusive, largest living invertebrate; an103
overviewofthespecies’taxonomyanddetailedtreatmentofthemorphologywillbe104
treatedinaseparateforthcomingpublication(Bolstadetal.inprep).105
106
Polargigantismandsomekeymorphologicalfeaturesofthecolossalsquid107
Foralongtimepolarscientistshaveobservedthatorganismsofsuchareas108
canreachremarkablylargesizes.Polargigantismisnotabletoolforunderstanding109
the biophysical principles and ecological theories (e.g. temperature‐size rule,110
Bergmannrule,oxygenavailabilitycoupledwith lowmetabolicrateshypothesis)111

5
behindbodysizeevolution(Atkinson1994;ChapelleandPeck1999;Chapelleand112
Peck 2004; Vermeij 2016). Although not consensual, some argue that this113
phenomenon may be linked to the fact that polar giant taxa share common114
evolutionary histories with deep‐sea taxa. In other words, polar gigantism is115
associated with abyssal gigantism. Yet, it is worth noting that the evolutionary116
historyof(benthic)SouthernOceanfaunaiscomplexandtherespective“invasions”117
workedbothways(i.e.shelftodeepandviceversa)(Brandtetal.2007;Strugnellet118
al.2011).OneextremecaseofpolargigantismintheSouthernOcean(whereother119
examples of this phenomenon include foraminiferans, sponges, ctenophores,120
isopods,copepods,amphipods,pycnogonids,pteropods,annelids,echinoderms)is121
the colossal squid (Fig. 1), which was first described by Robson (1925) from122
fragments from two squids obtained from sperm whale stomach contents.123
Posteriorly,thedescriptionofitsbeakswasdonebyKlumovandYukhov(1975),124
McSweeny (1970), and then reviewed by Clarke (1980) and Xavier and Cherel125
(2009).126
ThecolossalsquidbelongstothefamilyCranchiidae,or“glass”squids,which127
are generally small‐to‐medium sized species; M. hamiltoni is several orders of128
magnitudelargerthansomeothercranchiidspecies,attainingamaximummantle129
length(ML)of2.5metersandtotallength(TL)of6meters.Themaximumweight130
recordedtodatewas495kg(Fig.1)(JerebandRoper2010).Apartfromthecoelom131
(fluid‐filledbuoyancychamber),whilemostcranchiidshavetranslucentbodies(see132
examplesinFig.2A,BandC),M.hamiltonihasmuchmoremuscledensity(Fig.2D133
andE).Afurthernotabledifferencebetweenthecolossalsquidandothercranchiids134
isthepresenceofswivellinghooksonitstentacleclubs(Voss1980;Vossetal.1992)135
(Fig.2G).Thus,thecolossalsquidisaclear“outlier”inthecranchidgroup.136

6
137
Earlystages138
Recordsofeggmassesandhatchlingsareessentiallynon‐existent.Between139
1962and1985,only33juvenileswerereported,allcaughtusingRMTandIKMT140
nets (Fig. 3). Their mantle length (ML) ranged from 3 to 146mm, with a clear141
relationshipbetweenjuvenilesizeanddepthofcapture.Infact,individualsbelow142
ML40mmhavebeenmostly foundbetweenthesurfaceand500m(Fig.3) (see143
more juvenile morphometric information in McSweeny 1970; Fillipova 1972;144
Filippova 1979; Rodhouse and Clarke 1985; Filippova 1991; Filippova and145
Pakhomov1994;LuandWilliams1994;AndersonandRodhouse2002;Filippova146
2002).Nonetheless,themajorityofthespecimensknownsofarhavebeencollected147
bytrawls(N=155;Fig.3)butbesidestheirlocationnootherrelevantdata(e.g.ML,148
lifestage)areavailable.149
150
Age,growthandreproduction151
Besidesthecharacteristiclowtemperatures,theSouthernOceanisalsowell152
knownbythedrasticannualcyclesofproductivity.Theoscillationsfromabundance153
to famine are thought to be associated with increased resistance to starvation,154
reducedcompetitionandunusuallyslowgrowthrates(LindstedtandBoyce1985;155
ArnettandGotelli2003;Clarke2003).Deep‐seaandpolarenvironmentsarealso156
usuallylinkedwithlowmetabolicratesandlongerlongevity.Forinstance,Robison157
et al. (2014) recently reported a female deep‐sea benthic octopus Graneledone158
boreopacifica undertaking an astonishing 53‐month brooding period (by far the159
longestegg‐broodingperiodeverreportedforanyanimal).Ifbroodingcomprises160
aboutaquarterofthisoctopodlifespan,thisspeciesmaybeamongthelongest‐lived161

7
ofallcephalopods.Theirfindingsareunparalleledbythosefromthewell‐studied162
shallow‐waterspeciesandhighlightsthenotionofhowlittleweknowaboutlifein163
marineextremeenvironments(deep‐seaandpoles).Withinthiscontext,onemight164
imaginethatM.hamiltonimaylivemuchlongerthantheirtemperateandtropical165
counterparts, i.e. far longerthan12to18months.Themostcommonmethodfor166
measuringgrowthincephalopods,andconsequentagedetermination,iscounting167
growth increments on statoliths (Lipinski 1986; Rodhouse and Hatfield 1990);168
however,M. hamiltoni material reported to date has not yet been sufficient to169
validatetheperiodicityofstatolith‐ringformationforthisspecies.170
The colossal squid takes longer to mature and reproduce than its more171
northernrelatives,becomingmatureatlengthsofatleast1mandweightabove30172
kg(JerebandRoper2010).Althoughincolderwaterscephalopodstendtoproduce173
a low number of large eggs (Collins and Rodhouse 2006), the colossal squid’s174
potentialfecundityhasbeenestimatedatamaximumof4.2millionoocytes(Jereb175
andRoper2010),whichmakes itoneofthemost fecundcephalopods,especially176
whencomparedtootherpolarcephalopodfauna(CollinsandRodhouse2006).177
178
Distribution179
Part of the known distribution of the colossal squid is based on the rare180
encounterswithliveorrecentlydeadspecimensaboard(mainly)toothfishfishing181
(e.g. Fig. 1 D and E) vessels and the stomach contents of the squid’s predators.182
Mesonychoteuthis hamiltoni has a circumpolar distribution, ranging from the183
Antarcticcontinent(excludingrespectiveshelvesandKerguelenPlateau)uptothe184
Sub‐AntarcticFront(SAF)(Fig.4).BasedonXavieretal.(2015b),thehighestvalues185
ofthespecies’predictedhabitatsuitabilityarefound:i)betweentheWeddellSea186

8
(Atlanticsector)andthewestoftheKerguelenarchipelago(~60°E),andii)between187
180°Eand120°WintheRoss/Amundsenareas.188
189
Vertical(ontogenetic)movements190
Available information on colossal squid’s verticalmigrating habits is very191
scarce.AsshowninFigure3,smalljuveniles(<40mmML)aremainlyfoundfrom192
thesurfaceto500m.Thehigherproductivitynearthesurfacemayconfertheyoung193
higherchancesoffeedingbetweenhatchingandtheirprobablemigrationtodeeper194
waters.Atlatejuvenilestages,M.hamiltoni isassumedtoconductanontogenetic195
descenttodepthsaround2000m(LuandWilliams1994).Thus,thecolossalsquid196
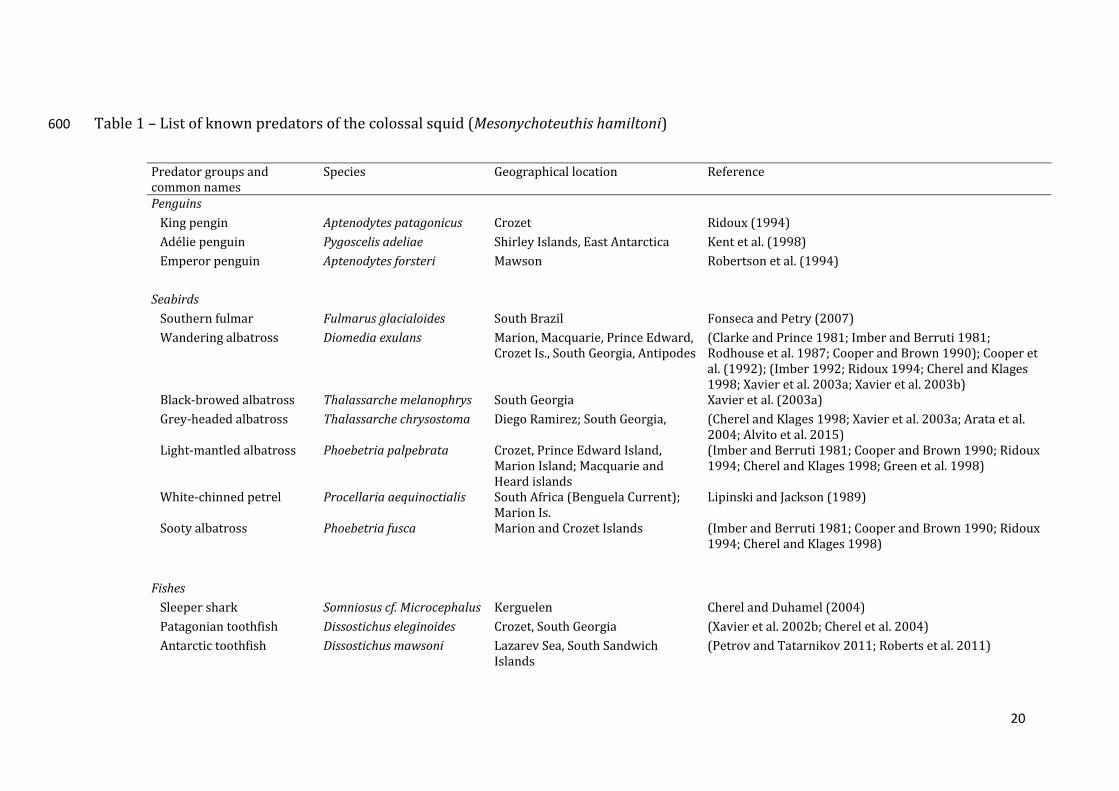
likelyspendsthemajorityofitslife(latejuvenile,subadultandadultstages)inthe197
meso‐andbathypelagiczonesoftheSouthernOcean.Nonetheless,itisworthnoting198
that femalespecimenshavebeencapturedbyfishingvesselsatdepthsshallower199
thantheabovementionedrange,suggestingthatfemalesmaymigratetoshallower200
waterstospawn,asdoothercranchiids.201
202
Predatorandpreyinteractions203
Theundigestedremainsofcolossalsquid(beaks,tentacularhooksorsucker204
rings)havebeenfoundinagreatvarietyofpredators’stomachcontents.Presently,205
M.hamiltoni isknownfromthedietsof17differentpredatorspecies,comprising206
penguins,seabirds,fishesandmarinemammals(Table1).Thelargerpredatorsof207
the colossal squids are the spermwhales (Clarke 1980) and the sleeper sharks208
(CherelandDuhamel2004) (Fig.5),but in contrast to the former, the latter isa209
bottom scavenger and ambush predator. One of the more remarkable predator210

9
observations has beenmade at the Kerguelen Islands, where 66% of the shark211
stomachscontainedcolossalsquidremains(CherelandDuhamel2004).212
ColossalsquidbeakshavealsobeenfoundquitefrequentlyinthePatagonian213
toothfish (Dissostichus eleginoides) stomach contents (Fig. 5), and in turn, beak‐214
shapedbitesandscratches fromclubhooks (matching thecolossal squid’s)have215
alsobeenfoundalongthebodiesofcapturedPatagoniantoothfish(Remesloetal.216
2015).Thismaysuggestreciprocalpredator‐preydynamics.Patagoniantoothfish217
may be able to forage squids larger themselves because they attack dead or218
moribundcolossalsquids.Remesloetal.(2015)alsoarguethat,duetoanatomical219
constraints(namelytoothshape),thetoothfishmayonlyscavengetheleftoversof220
otherpredators,includingthecolossalsquiditself.Cannibalismisalsoverycommon221
amongcephalopods,includinggiantsquids(BolstadandO'Shea2004;Ibáñezand222
Keyl2010).223
Seabirds,namelyalbatrosses,arealsoknowtoscavengeM.hamiltoni(Fig.5).224
Albatrossstomachsoftencontaintheremainsofcephalopodpreythatwouldhave225
been several times heavier that their ownmean size (Croxall and Prince 1994;226
Xavier et al. 2003b; Xavier and Croxall 2007; Alvito et al. 2015; Guerreiro et al.227
2015).Infact, incertainperiods(e.g.endofinterbreeding/beginningofbreeding228
period)scavengingplaysacrucialroleinthedietsofwandering(Diomedeaexulans),229
black‐browed (Thalassarche melanophrys) and grey‐headed (Thalassarche230
chrysostoma)albatrosses,asmorethan60%ofthecephalopodremainsrecovered231
had potentially been scavenged (more than 95% of the total estimatedmass of232
cephalopodsconsumed)(Alvitoetal.2015).233
Regarding the squid’s own feeding ecology,M. hamiltoni specimens with234
intactorundigestedstomachcontentsremainunknown.Moreover,dietarystudies235

10
incephalopodsaredifficulttoconductasgutcontentsareusuallyinanadvanced236
state of digestion, making identification of prey items difficult to impossible.237
Cephalopodsalsomaceratetheirpreysfinelybeforeingestion,usingtheirbeaksand238
radula. It hasbeen suggested thatM.hamiltoni feedsonmyctophids, Patagonian239
toothfish, sleeper sharks andother squids (Jereb andRoper2010;Roberts et al.240
2011). Based on phylogenetic (metabolic) inferences and polar gigantism241
constraints,RosaandSeibel(2010)arguedthatthecolossalsquidisprobablynota242
predatorcapableofhigh‐speedpredator–prey interactions (seealsosection3.5).243
Therefore, the very large but immobilized (or weakened) toothfish specimens244
captured by longline would present easy, opportunistic prey for M. hamiltoni245
(Yukhov2012;Remesloetal.2015).246
Anumberofstudieshavealsousedadifferentapproachtodeterminethe247
trophicpositionofthecolossalsquid,namelytheuseofisotopicanalysis(e.g.Cherel248
andHobson2005).Nitrogenisotopesratio(δ15N)indicatetheconsumer’strophic249
position,whereascarbonisotopesratio(δ13C)canprovideinformationonhabitat250
occupation. Assessment of the nitrogen signatures have demonstrated that M.251
hamiltoniisdefinitelyoneofthetoppredatorsintheSouthernOcean(Fig.6).252
253
Vision254
Giant(Architeuthisdux)andcolossalsquidspossessthelargesteyesinthe255
Worldoceans(Fig.7),andontheplanet(Walls1942;LandandNilsson2002).Since256
bothofthesesquidspeciesarepelagicorganismsthatshare(orcompetefor)similar257
ecologicalnicheswithother(vertebrate)toppredators,thetremendousdifferences258
ineyesizemayindicateadifferentpurposefortheuseofeyes.Nilssonetal.(2012)259

11
suggestedthattheverylargeeyesmayhaveconferredanevolutionaryadvantagein260
increased ability to detect large predators (especially spermwhales), rather for261
utilityindetectingmatesorpreyatlongdistances.Theseauthorsmodelledlarge262
squids’visualrangeandproposedthatthegianteyesenablethemtodetectsperm263
whales as they trigger plankton bioluminescence while swimming through the264
water column. In other words, as whales conduct hunting dives, they attain265
considerable speed and agitate the deep waters where a great variety of266
bioluminescentorganismsthrive.Thecolossalsquid’seyes,duetotheirlargeretina,267
areabletodetectsuchsmallsourcesoflightfromadistance,althoughtheymaynot268
necessarilyregisterthepredatorsbeforebeingdetectedthemselvesbythewhale’s269
sonar(therangeofwhichcanexceed120m).Thesquidmay,however,bevisually270
warnedinsufficienttimetoattemptanescape(Nilssonetal.2012).Nonetheless,it271
isworthnotingthatSchmitzetal.(2013)alsoarguedthatM.hamiltoni’seyesare272
withintheexpectedallometricrangeforsquids,andthusmaybeasimpleresultof273
aphylogeneticallyconservedtrait.274
275
Metabolism276
Rosa and Seibel (2010) calculated the metabolic rates and energy277
requirements of the colossal squid through a depth‐temperature gradient278
representative of the Southern Ocean (Fig. 8). This phylogenetic inference was279
conducted based on other deep‐sea cranchiid data and followed the expected280
allometricrelationshipbetweenbodysizeandmetabolism(Seibel2007;Seibeland281
Drazen2007;Rosaetal.2009).Basedontheirfindings,theauthorsarguedthatthe282
colossal squid “is not a voracious predator capable of high‐speed predator–prey283
interactions.Itis,rather,anambushorsit‐and‐floatpredatorthatusesthehookson284

12
itsarmsandtentaclestoensnarepreythatunwittinglyapproach”.Thecolossalsquid285
waslongthoughttobeanintrepidpredator,duetothe“kraken”‐relatedmythsand286
storiesassociatedwithitslargesize,butafterthisstudy,ithasbeenpostulatedthat287
thecolossalsquidperhapsdoesnotdeservesuchareputation.Thequestionthatwe288
nowraiseis:doesanorganismneedtobeanactivepursuitpredatortobefearsome?289
Forinstance,whitesharks,leopards,crocodiles,largemonkfishandpythonsareall290
ambushpredators(manywithlowenergydemands),anditistheircapabilitytohide291
andstrikewithoutwarningthatmakestheminherentlyaggressive.292
293
Bioluminescence294
Likethegreatmajorityofdeepseainhabitants,M.hamiltonipossesseslight‐emitting295
organs(photophores),intheformoftwoelongatestructuresontheventralsurface296
ofeacheye(Fig.9)(Herringetal.2002).Thesmallerislocatedanteriorlywhilethe297
largeriscrescent‐shapedandlocatedposteriorlyaroundtheoutsiderimoftheiris298
(Voss1980).M.hamiltoniphotocytescontaincrystalloidswhoseprofilesappearas299
rectangularblocksorelongateneedles,andthereflectorplateletsareunusual,being300
associated with a microtubular array that confers their edges a comb‐like301
appearance (Fig. 9). Interestingly,withinCranchiidae,members of the subfamily302
Cranchiinaeobliteratethesilhouetteoftheeyeballbyilluminatingnumeroussmall303
ventrallydirectedphotophores,whereasthetaoniines(includingM.hamiltoni)use304
fewer,moreelaborate,elongatereflectorstospreadthelightfromthephotophores305
overtheentireprojectedventralsurfaceoftheeyeball(Herringetal.2002).Likeall306
taoniines, the two photophores of the M. hamiltoni have a mirror‐image307
arrangementinwhichtheilluminatedsurfaceofeachlight‐emittingorganextends308
towards the other, across the main ventral area of the eyeball. The subocular309

13
photophores of M. hamiltoni may serve as ventral camouflage and counter‐310
illumination, to blend in with downwelling light when seen from below (Young311
1975;Young1977;Youngetal.1980).Yet,thisseemstobenotusefulinlargesquid312
individuals since their main predators are not visually‐orientated hunters.313
Photophoresmayalsobeusedasadeceptivetechnique–whenilluminated,other314
organismsmaydisregardthecolossalsquidasathreat,allowingittoapproachits315
prey.Alternatively,thephotophoresmaybeusedas“flashlights”tomoreaccurately316
predictthedistanceneededtostretchitshookedtentaclesandsuccessfullycapture317
unwittingprey.318
319
Futuredirections320
Furthereffortsshouldbemadetounderstandmoreaboutthelifeofthisgiant.Ithas321
beenassumedthatM.hamiltoniisquiteabundantintheSouthernOcean(biomass322
ofaround90milliontonnes)andtherehavebeensomereportsonitsinterestto323
fisheries(JerebandRoper2010).Everysample,fragmentorsightinghasprovedto324
beahighlyvaluablesourceofinformationandhelpedscientiststounveilmoreand325
moreaboutM.hamiltoni.Baitedcamerasandremotelyoperatedvehiclescouldbe326
deployedinthesearchoflivefootage,ashashappenedrecentlyforotherdeepsea327
“giant”squids,e.g.Architeuthisdux(KuboderaandMori2005)andTaningiadanae328
(Kuboderaetal.2006).M.hamiltoniandotherAntarcticorganismshasbeenwidely329
usedforeducationandoutreachactivitiesworldwide(Xavieretal.2015a).Besides330
the fact that these unique invertebrate giants continue to attract considerable331
attentionfrommediaandthepublicingeneral,itisimportanttohighlight,inthe332
words of Vermeij (2016), that: “Gigantism is (… ) a functionally distinct and333
ecologicallyimportantconditionthatisbothenabledbyresourcesandcompelledby334

14
naturalselection.Thedistributionofmaximumsizeintimeandspacecantherefore335
informourunderstandingofmajorpatternsinthehistoryoflife.”336
337
Acknowledgements338
ThisstudybenefitedfromthestrategicprogramofMARE,financedbyFCT(MARE‐339
UID/MAR/04292/2013). JX is supported by the Investigator FCT program340
(IF/00616/2013)andbytheFoundationforScienceandTechnology(Portugal)and341
isassociatedtoSCARAnT‐ERA,SCAREGBAMM,ICED,BAS‐CEPHprograms.RRis342
supportedbytheInvestigatorFCTprogram(IF/01373/2013).343
344
References345
346
Alvito PM, Rosa R, Phillips RA, Cherel Y, Ceia F, Guerreiro M, Seco J, Baeta A, Vieira RP, Xavier 347 JC (2015) Cephalopods in the diet of nonbreeding black‐browed and grey‐headed 348 albatrosses from South Georgia. Polar Biology 38:631‐641 349
Anderson CIH, Rodhouse PG (2002) Distribution of juvenile squid in the Scotia Sea in relation to 350 regional oceanography. Bulletin of Marine Science 71:97‐108 351
Arata J, Robertson G, Valencia J, Xavier J, Eacute C, Moreno CA (2004) Diet of grey‐headed 352 albatrosses at the Diego Ramírez Islands, Chile: ecological implications. Antarctic 353 Science 16:263‐275 doi doi:10.1017/S095410200400207X 354
Arnett AE, Gotelli NJ (2003) Bergmannʼs rule in larval ant lions: tes ng the starva on resistance 355 hypothesis. Ecol Entomol 6:645‐650 356
Arntz WE, Brey T, A. GV (1994) Antarctic zoobenthos. Oceanogr Mar Biol Ann Rev 32:241‐304 357 Atkinson D (1994) Temperature and organism size ‐ a biological law for ectotherms? . Adv Ecol 358
Res 25:1‐58 359 Barnes DKA, Hodgson DA, Convey P, Allen CS, Clarke A (2006) Incursion and excursion of 360
Antarctic biota: past, present and future Global Ecology and Biogeography 15:121‐142 361 Bolstad KS, O'Shea S (2004) Gut contents of a giant squid Architeuthis dux (Cephalopoda: 362
Oegopsida) from New Zealand waters. New Zealand Journal of Zoology 31:15‐21 363 Brandt A, Gooday AJ, Brandão SN, Brix S, Brökeland W, Cedhagen T, Choudhury M, Cornelius 364
N, Danis B, De Mesel I (2007) First insights into the biodiversity and biogeography of 365 the Southern Ocean deep sea. Nature 447:307‐311 366
Chapelle G, Peck LS (1999) Polar gigantism dictated by oxygen availability. Nature 399:114‐115 367 Chapelle G, Peck LS (2004) Amphipod crustacean size spectra: new insights in the relationship 368
between size and oxygen. Oikos 106:167‐175 369 Cherel Y, Duhamel G (2004) Antarctic jaws: cephalopod prey of sharks in Kerguelen waters. 370
Deep‐Sea Research Part I‐Oceanographic Research Papers 51:17‐31 doi 371 10.1016/j.dsr.2003.09.009 372

15
Cherel Y, Duhamel G, Gasco N (2004) Cephalopod fauna of subantarctic islands: new 373 information from predators. Marine Ecology Progress Series 266:143‐156 doi 374 10.3354/meps266143 375
Cherel Y, Hobson KA (2005) Stable isotopes, beaks and predators: a new tool to study the 376 trophic ecology of cephalopods, including giant and colossal squids. Proceedings of 377 Royal Society B 272:1601‐1607 378
Cherel Y, Hobson KA (2007) Geographical variation in carbon stable isotope signatures of 379 marine predators: a tool to investigate their foraging areas in the Southern Ocean. 380 Marine Ecology Progress Series 329:281‐287 doi 10.3354/meps329281 381
Cherel Y, Hobson KA, Guinet C, Vanpe C (2007) Stable isotopes document seasonal changes in 382 trophic niches and winter foraging individual specialization in diving predators from 383 the Southern Ocean. Journal of Animal Ecology 76:826‐836 doi 10.1111/j.1365‐384 2656.2007.01238.x 385
Cherel Y, Klages N (1998) A review of the food of albatrosses. In: Robertson G, Gales R (eds) 386 Albatross Biology and Conservation. Surrey Beatty & Sons, Chipping Norton, Australia, 387 pp 113‐136 388
Clarke A (2003) Evolution, adaptation, and diversity: global ecology in an Antarctic context. In: 389 Huiskes AHL, Gieskes WWC, Rozema J, Schorno RML, van der Vies SM, Wolff WJ (eds) 390 Antarctic Biology in a Global Context. Int. Biol. Symp. Backhuys Publishers, Leiden, pp 391 3‐17 392
Clarke A, Johnston NM (2003) Antarctic marine benthic diversity. Oceanogr Mar Biol Ann Rev 393 41:47‐114 394
Clarke MR (1980) Cephalopoda in the diet of sperm whales of the southern hemisphere and 395 their bearing on sperm whale biology. Discovery Reports 37:324 396
Clarke MR, Goodall N (1994) Cephalopods in the diets of three Odontoceti cetacean species 397 stranded at Tierra del Fuego, Globicephala melaena (Traill, 1809), Hyperoodon 398 planifrons Flower, 1882 and Cephalorhynchus commersonii (Lacepede, 1804). Antarctic 399 Science 6:149‐154 400
Clarke MR, MacLeod N (1982) Cephalopod remains from the stomachs of sperm whales caught 401 in the Tasman sea. Memoirs of the National Museum Victoria 43:25‐42 402
Clarke MR, MacLeod N, Paliza O (1976) Cephalopod remains from the stomachs of Sperm 403 whales caught off Peru and Chile. Journal of Zoology 180:477‐493 404
Clarke MR, Prince PA (1981) Cephalopod remains in regurgitations of black‐browed and grey‐405 headed albatrosses at South Georgia. British Antarctic Survey Bulletin 54:1‐8 406
Collins MA, Rodhouse PG (2006) Southern Ocean cephalopods. Advances in Marine Biology 407 50:191‐265 408
Cooper J, Brown CR (1990) Ornithological research at the sub‐Antarctic Prince Edward Islands: 409 a review of achievements. South Africa Trans Nav Antarkt 20:40‐57 410
Cooper J, Henley S, Klages N (1992) The diet of the wandering albatross Diomedea exulans at 411 subantarctic Marion Island. Polar Biology 12:477‐484 412
Croxall JP, Prince PA (1994) Dead or alive, night or day ‐ how do albatrosses catch squid. 413 Antarctic Science 6:155‐162 414
Filippova J (1991) Morpho‐ecological aspects of the study of Antarctic squids. Bull Mar Sci 415 49:662 416
Filippova JA (2002) Review of Soviet/Russian studies on squids in the Antarctic Ocean. Bulletin 417 of Marine Science 71:255‐267 418
Filippova JA, Pakhomov EA (1994) Young squid in the plankton of Prydz Bay, Antarctica. 419 Antarctic Science 6:171‐173 doi doi:10.1017/S095410209400026X 420
Filippova JAY, V. L. (1979) Specific composition and distribution of cephalopod molluscs in 421 meso‐ and bathypelagic Antarctic waters. Antarktika Doklady Komissi:175‐187 422
Fillipova JA (1972) New data on the squids (Cephalopoda: Oegopsida) from the Scotia Sea 423 (Antarctic). Malacologia 11:391‐406 424

16
Fiscus CH, Rice DW, Wolman AA (1989) Cephalopods from the stomachs of sperm whales 425 taken off California. NOAA Technical Report NMFS 83:1‐12 426
Fonseca Vd, Petry M (2007) Evidence of food items used by Fulmarus glacialoides (Smith 1840) 427 (Procellariiformes: Procellariidae) in Southern Brazil. Polar Biology 30:317‐320 doi 428 10.1007/s00300‐006‐0185‐7 429
Green K, Kerry KR, Disney T, Clarke MR (1998) Dietary studies of light‐mantled sooty 430 albatrosses Phoebetria palpebrata from Macquarie and Heard Islands. Marine 431 Ornithology 26:19‐26 432
Guerreiro M, Phillips RA, Cherel Y, Ceia FR, Alvito P, Rosa R, Xavier JC (2015) Habitat and 433 trophic ecology of Southern Ocean cephalopods from stable isotope analyses. Marine 434 Ecology Progress Series 530:119‐134 doi 10.3354/meps11266 435
Herring PJ, Dilly PN, Cope C (2002) The photophores of the squid family Cranchiidae 436 (Cephalopoda: Oegopsida). Journal of Zoology 258:73‐90 437
Ibáñez CM, Keyl F (2010) Cannibalism in cephalopods. Rev Fish Biol Fish 20:123‐136 438 Imber MJ (1992) Cephalopods eaten by wandering albatrosses Diomedea exulans L. breeding 439
at six circumpolar localities. Journal of the Royal Society of New Zealand 22:243‐263 440 Imber MJ, Berruti A (1981) Proccellariiform seabirds as squid predators. In: Cooper J (ed) 441
Proceedings of the Symposium on Birds of the Sea and Shore. African Seabird Group, 442 Cape Town, pp 43‐61 443
Jereb P, Roper CFE (2010) Cephalopods of the world. An annotated and illustrated catalogue of 444 cephalopod species known to date. Myopsidae and Oegopsidae. FAO, Rome 445
Jereb P, Roper CFE, Norman MD, Finn JK (2014) Cephalopods of the world. An annotated and 446 illustrated catalogue of cephalopod species known to date. Volume 3. Octopods and 447 Vampire Squids. . FAO, Rome 448
Kent S, Seddon J, Roberston G, Wienecke BC (1998) Diet of Adélie penguins Pygoscelis adeliae 449 at Shirley Island, East Antarctica, January 1992. Marine Ornithology 26:7‐10 450
Klumov SK, Yukhov VL (1975) Mesonychoteuthis hamiltoni Robson, 1925 (Cephalopoda: 451 Oegopsida). Antarktika Doklady Komission 14:159‐189 452
Korabelnikov L (1959) The diet of sperm whales in the Antarctic seas. Priroda 3:103‐104 453 Kubodera T, Koyama Y, Mori K (2006) Observations of wild hunting behaviour and 454
bioluminescence of a large deep‐sea, eight‐armed squid, Taningia danae. Proceedings 455 of Royal Society B doi:10.1098/rspb.2006.0236 456
Kubodera T, Mori K (2005) First‐ever observations of a live giant squid in the wild. Proceedings 457 of Royal Society B in press 458
Land MF, Nilsson D‐E (2002) Animal Eyes. Oxford University Press, Oxford 459 Lindstedt SL, Boyce MS (1985) Seasonality, fasting endurance, and body size in mammals. Am 460
Nat 125:873‐878 461 Lipinski MR (1986) Methods for the validation of squid age from statoliths. Journal of the 462
Marine Biological Association of the United Kingdom 66:505‐526 doi 463 doi:10.1017/S0025315400043095 464
Lipinski MR, Jackson S (1989) Surface‐feeding on cephalopods by procellariiform seabirds in 465 the southern Benguela region, South Africa. Journal of Zoology 218:549‐563 466
Lu CC, Williams R (1994) Contribution to the biology of squid in the Prydz Bay region, 467 Antarctica. Antarctic Science 6:223‐229 doi doi:10.1017/S0954102094000349 468
McClain CR, Balk MA, Benfield MC, Branch TA, Chen C, Cosgrove J, Dove AD, Gaskins LC, Helm 469 RR, Hochberg FG (2015) Sizing ocean giants: patterns of intraspecific size variation in 470 marine megafauna. PeerJ 3:e715 471
McSweeny ES (1970) Description of the juvenile form of the Antarctic squid Mesonychoteuthis 472 hamiltoni Robson. Malacologia:323‐332 473
Moore JK, Abbott MR, Richman JG (1997) Variability in the location of the Antarctic Polar Front 474 (90 degrees‐20 degrees W) from satellite sea surface temperature data. Journal of 475 Geophysical Research‐Oceans 102:27825‐27833 doi 10.1029/97jc01705 476

17
Moore JK, Abbott MR, Richman JG (1999) Location and dynamics of the Antarctic Polar Front 477 from satellite sea surface temperature data. Journal of Geophysical Research‐Oceans 478 104:3059‐3073 479
Nilsson DE, Warrant EJ, Johnsen S, Hanlon R, Shashar N (2012) A unique advantage for giant 480 eyes in giant squid. Current biology : CB 22:683‐688 doi 10.1016/j.cub.2012.02.031 481
Orsi AH, Whitworth T, Nowlin WD (1995) On the meridional extent and fronts of the Antarctic 482 Circumpolar Current Deep‐Sea Research Part I‐Oceanographic Research Papers 483 42:641‐673 doi 10.1016/0967‐0637(95)00021‐w 484
Petrov AF, Tatarnikov VA (2011) Results of investigation of the diet of antarctic toothfish 485 Dissostichus mawsoni (Nototheniidae) in the Lazarev Sea. Journal of Ichthyology 486 51:131‐135 doi 10.1134/S0032945210051017 487
Remeslo AV, Yakushev MR, Laptikhovsky V (2015) Alien vs. Predator: interactions between the 488 colossal squid (Mesonychoteuthis hamiltoni) and the Antarctic toothfish (Dissostichus 489 mawsoni). Journal of Natural History 49:2483‐2491 doi 490 10.1080/00222933.2015.1040477 491
Ridoux V (1994) The diets and dietary segregation of seabirds at the Subantarctic Crozet 492 Islands. Marine Ornithology 22:1‐192 493
Roberts J, Xavier JC, Agnew DJ (2011) The diet of toothfish species Dissostichus eleginoides and 494 Dissostichus mawsoni with overlapping distributions. J Fish Biol 79:138‐154 doi 495 10.1111/j.1095‐8649.2011.03005.x 496
Robertson G, Williams R, Green K, Robertson L (1994) Diet composition of emperor penguin 497 chicks Aptenodytes forsteri at two Mawson Coast colonies, Antarctica. Ibis 136:19‐31 498
Robison BH, Seibel BA, Drazen JC (2014) Deep‐Sea Octopus (Graneledone boreopacifica) 499 conducts the longest‐known egg‐brooding period of any animal. Plos One 9:e103437 500
Robson GC (1925) On Mesonychoteuthis, a new genus of oegopsid Cephalopoda. Annals and 501 Magazine of Natural History:272‐277 502
Rodhouse P, Hatfield E (1990) Age determination in squid using statolith growth increments. 503 Fisheries Research 8:323‐334 504
Rodhouse PG (1998) Large and meso‐scale distribution of the ommastrephid squid Martialia 505 hyadesi in the Southern Ocean: a synthesis of information relevant to fishery 506 forecasting and management. Korean Journal of Polar Research 8:145‐154 507
Rodhouse PG, Arnbom T, Fedak MA, Yeatman J, Murray AWA (1992) Cephalopod prey of the 508 southern elephant seal, Mirounga leonina L. Canadian Journal of Zoology 70:1007‐509 1015 510
Rodhouse PG, Clarke MR (1985) Growth and distribution of young Mesonychoteuthis hamiltoni 511 Robson (Mollusca: Cephalopoda): An Antarctic squid. Vie et Milieu 35:223‐230 512
Rodhouse PG, Clarke MR, Murray AWA (1987) Cephalopod prey of the wandering albatross 513 Diomedea exulans. Marine Biology 96:1‐10 514
Rodhouse PG, Prince PA, Trathan PN, Hatfield EMC, Watkins JL, Bone DG, Murphy EJ, White 515 MG (1996) Cephalopods and mesoscale oceanography at the Antarctic Polar Front: 516 Satellite tracked predators locate pelagic trophic interactions. Marine Ecology Progress 517 Series 136:37‐50 doi 10.3354/meps136037 518
Rodhouse PG, White MG (1995) Cephalopods occupy the ecological niche of epipelagic fish in 519 the Antactic Polar Frontal zone. Biological Bulletin 189:77‐80 doi 10.2307/1542457 520
Rosa R, Seibel BA (2010) Slow pace of life of the Antarctic colossal squid. Journal of the Marine 521 Biological Association of the United Kingdom 90:1375‐1378 doi 522 10.1017/s0025315409991494 523
Rosa R, Trueblood L, Seibel BA (2009) Ecophysiological influence on scaling of aerobic and 524 anaerobic metabolism of pelagic gonatid squids. Physiological and Biochemical Zoology 525 82:419‐429 526

18
Schmitz L, Motani R, Oufiero C, Martin C, McGee M, Gamarra A, Lee J, Wainwright P (2013) 527 Allometry indicates giant eyes of giant squid are not exceptional. BMC Evolutionary 528 Biology 13:45 529
Seibel BA (2007) On the depth and scale of metabolic rate variation: scaling of oxygen 530 consumption rates and enzymatic activity in the Class Cephalopoda (Mollusca). Journal 531 of Experimental Biology 210:1‐11 532
Seibel BA, Drazen JC (2007) The rate of metabolism in marine animals: environmental 533 constraints, ecological demands and energetic opportunities. Philosophical 534 Transactions of the Royal Society B 362:2061‐2078 535
Seibel BA, Thuesen EV, Childress JJ (2000) Light‐limitation on predator‐prey interactions: 536 consequences for metabolism and locomotion of deep‐sea cephalopods. Biological 537 Bulletin 198:284‐298 538
Stowasser G, Atkinson A, McGill RAR, Phillips RA, Collins MA, Pond DW (2012) Food web 539 dynamics in the Scotia Sea in summer: A stable isotope study. Deep Sea Research Part 540 II: Topical Studies in Oceanography 59‐60:208‐221 doi 10.1016/j.dsr2.2011.08.004 541
Strugnell JM, Cherel Y, Cooke IR, Gleadall IG, Hochberg FG, Ibáñez CM, Jorgensen E, 542 Laptikhovsky VV, Linse K, Norman Mea (2011) The Southern Ocean: Source and sink? 543 Deep Sea Res Part II 58:196‐204 544
Vermeij GJ (2016) Gigantism and Its Implications for the History of Life. Plos One 11:e0146092 545 doi 10.1371/journal.pone.0146092 546
Voss N, Stephen SJ, Dong Z (1992) Family Cranchiidae. Smithsonian Contributions to Zoology 547 513:187‐210 548
Voss NA (1980) A generic revision of the Cranchiidae (Cephalopoda, Oegopsida). Bulletin of 549 Marine Science 30:365‐412 550
Walls GL (1942) The vertebrate eye and its adaptive radiation. Bloomfield Hills, Cranbrook 551 Xavier JC, Allcock L, Cherel Y, Hoving H‐J, Lipinski M, Gomes‐Pereira JN, Piatkowski U, Pierce G, 552
Rodhouse PG, Rosa R, Liz Shea L, Strugnell J, Vidal E, Villanueva R, Ziegler A (2015a) 553 The future challenges in cephalopod research for the next decade. Journal of the 554 Marine Biological Association of the United Kingdom 95:999‐1015 555
Xavier JC, Cherel Y (2009) Cephalopod beak guide for the Southern Ocean. British Antarctic 556 Survey, Cambridge, UK 557
Xavier JC, Croxall JP (2007) Predator‐prey interactions: why do larger albatrosses eat bigger 558 squid? Journal of Zoology 271:408‐417 doi 10.1111/j.1469‐7998.2006.00224.x 559
Xavier JC, Croxall JP, Reid K (2003a) Inter‐annual variation in the diet of two albatross species 560 breeding at South Georgia: implications for breeding performance. Ibis 145:593‐610 561
Xavier JC, Croxall JP, Trathan PN, Rodhouse PG (2003b) Inter‐annual variation in the 562 cephalopod component of the diet of wandering albatrosses Diomedea exulans 563 breeding at Bird Island, South Georgia. Marine Biology 142:611‐622 564
Xavier JC, Phillips RA, Cherel Y (2011) Cephalopods in marine predator diet assessments: why 565 identifying upper and lower beaks is important. ICES Journal of Marine Science 566 68:1857‐1864 doi 10.1093/icesjms/fsr103 567
Xavier JC, Raymond B, Jones DC, Griffiths H (2015b) Biogeography of cephalopods in the 568 Southern Ocean using habitat suitability prediction models. Ecosystems 19:220‐247 doi 569 10.1007/s10021‐015‐9926‐1 570
Xavier JC, Rodhouse PG, Purves MG, Daw TM, Arata J, Pilling GM (2002a) Distribution of 571 cephalopods recorded in the diet of the Patagonian toothfish (Dissostichus 572 eleginoides) around South Georgia. Polar Biology 25:323‐330 573
Xavier JC, Rodhouse PG, Purves MG, Daw TM, Arata J, Pilling GM (2002b) Distribution of 574 cephalopods recorded in the diet of the Patagonian toothfish (Dissostichus 575 eleginoides) around South Georgia. Polar Biology 25:323‐330 doi 10.1007/s00300‐001‐576 0343‐x 577

19
Xavier JC, Rodhouse PG, Trathan PN, Wood AG (1999) A Geographical Information System (GIS) 578 atlas of cephalopod distribution in the southern ocean. Antarctic Science 11:61‐62 579
Xavier JC, Walker K, Elliott G, Cherel Y, Thompson D (2014) Cephalopod fauna of South Pacific 580 waters: new information from breeding New Zealand wandering albatrosses. Marine 581 Ecology Progress Series 513:131‐142 doi 10.3354/meps10957 582
Xavier JC, Wood AG, Rodhouse PG, Croxall JP (2007) Interannual variations in cephalopod 583 consumption by albatrosses at South Georgia: implications for future commercial 584 exploitation of cephalopods. Marine and Freshwater Research 58:1136‐1143 doi 585 10.1071/mf06237 586
Young RE (1975) Transitory eye shapes and the vertical distribution of two midwater squids. 587 Pacific Science 29:243‐255 588
Young RE (1977) Ventral bioluminescence countershading in midwater cephalopods. In: Nixon 589 M, Messenger J (eds) Biology of cephalopods Symposia of the Zoological Society of 590 London Nº38 Academic Press, London, pp 161‐190 591
Young RE, Kampa EM, Maynard SD, Mencher FM, Roper CFE (1980) Counterillumination and 592 the upper depth limits of midwater animals. Deep‐Sea Research Part A 27:671‐691 593
Yukhov VL (2012) Antarctic deep‐water squid (АDS) Mesonychoteuthis hamiltoni, Robson, 594 1925. Distribution and borders of the geographic range. Main results of complex 595 research in the Azov and Black Seas Basin and the World Ocean. Kerch: YugNIRO 596 50:241‐248. Russian 597
598
599
20
Table1–Listofknownpredatorsofthecolossalsquid(Mesonychoteuthishamiltoni)600
Predatorgroupsandcommonnames
Species Geographicallocation Reference
Penguins
Kingpengin Aptenodytespatagonicus Crozet Ridoux(1994)Adéliepenguin Pygoscelisadeliae ShirleyIslands,EastAntarctica Kentetal.(1998)Emperorpenguin Aptenodytesforsteri Mawson Robertsonetal.(1994) Seabirds Southernfulmar Fulmarusglacialoides SouthBrazil FonsecaandPetry(2007)Wanderingalbatross Diomediaexulans Marion,Macquarie,PrinceEdward,
CrozetIs.,SouthGeorgia,Antipodes(ClarkeandPrince1981;ImberandBerruti1981;Rodhouseetal.1987;CooperandBrown1990);Cooperetal.(1992);(Imber1992;Ridoux1994;CherelandKlages1998;Xavieretal.2003a;Xavieretal.2003b)
Black‐browedalbatross Thalassarchemelanophrys SouthGeorgia Xavieretal.(2003a)Grey‐headedalbatross Thalassarchechrysostoma DiegoRamirez;SouthGeorgia, (CherelandKlages1998;Xavieretal.2003a;Arataetal.
2004;Alvitoetal.2015)Light‐mantledalbatross Phoebetriapalpebrata Crozet,PrinceEdwardIsland,
MarionIsland;MacquarieandHeardislands
(ImberandBerruti1981;CooperandBrown1990;Ridoux1994;CherelandKlages1998;Greenetal.1998)
White‐chinnedpetrel Procellariaaequinoctialis SouthAfrica(BenguelaCurrent);MarionIs.
LipinskiandJackson(1989)
Sootyalbatross Phoebetriafusca MarionandCrozetIslands (ImberandBerruti1981;CooperandBrown1990;Ridoux1994;CherelandKlages1998)
Fishes Sleepershark Somniosuscf.Microcephalus Kerguelen CherelandDuhamel(2004)Patagoniantoothfish Dissostichuseleginoides Crozet,SouthGeorgia (Xavieretal.2002b;Chereletal.2004)Antarctictoothfish Dissostichusmawsoni LazarevSea,SouthSandwich
Islands(PetrovandTatarnikov2011;Robertsetal.2011)
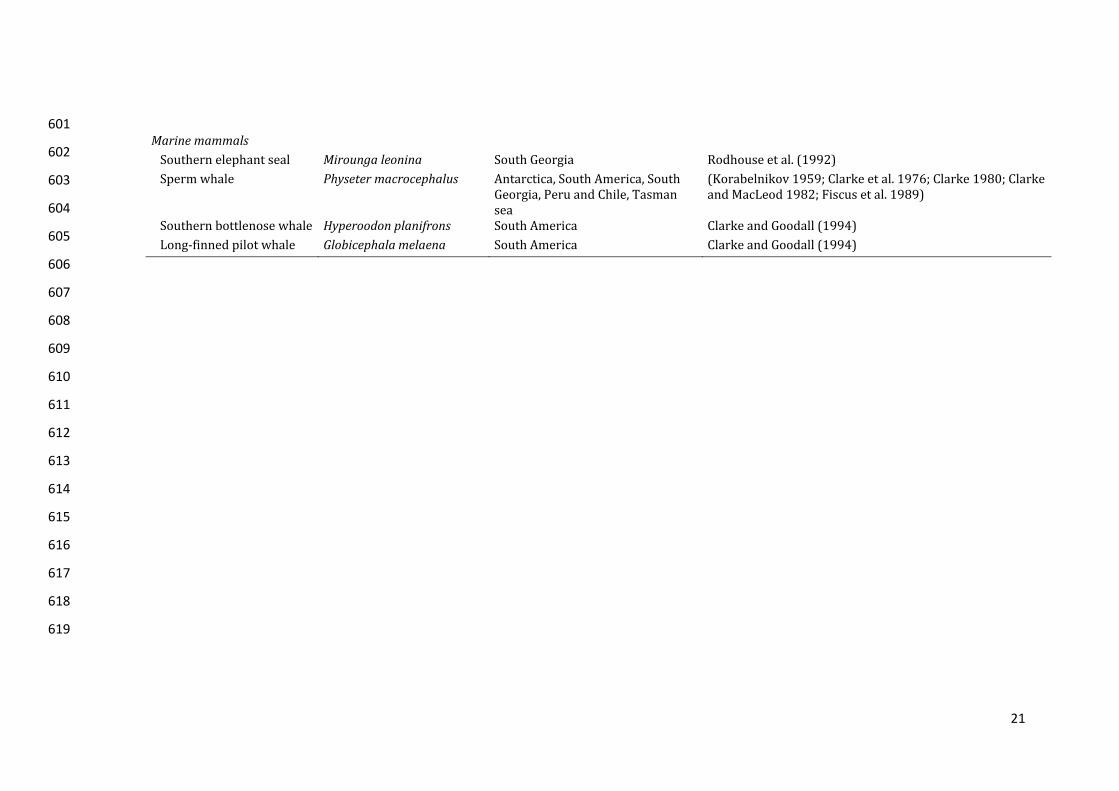
21
601
602
603
604
605
606
607
608
609
610
611
612
613
614
615
616
617
618
619
Marinemammals Southernelephantseal Miroungaleonina SouthGeorgia Rodhouseetal.(1992)Spermwhale Physetermacrocephalus Antarctica,SouthAmerica,South
Georgia,PeruandChile,Tasmansea
(Korabelnikov1959;Clarkeetal.1976;Clarke1980;ClarkeandMacLeod1982;Fiscusetal.1989)
Southernbottlenosewhale Hyperoodonplanifrons SouthAmerica ClarkeandGoodall(1994)Long‐finnedpilotwhale Globicephalamelaena SouthAmerica ClarkeandGoodall(1994)

22
620
Figurecaptions621
622
Figure1–Lengthandweight(logarithmicscale)ofthemarineinvertebrate(grey623
bars) and vertebrate giants (blue bars), with a special emphasis on the624
cephalopods:giantoctopus(Enteroctopusdofleini),giantsquid(Architeuthis625
dux) and colossal squidMesonychoteuthis hamiltoni (red bar). Cephalopod626
datafromJerebandRoper(2010)andJerebetal.(2014).Othergroups’data627
fromMcClainetal.(2015)andreferenceswithin.628
629
Figure 2 – Morphological dissimilarities between the colossal squid630
(Mesonychoteuthishamiltoni)andothercranchiidspecies.PanelsA,BandC631
showthetranslucentbodiesofdeep‐seaglasssquidsTeuthoweniapellucida632
and Taonius borealis (copyright: MBARI). Panels D and E show the much633
densermusculatureofthecolossalsquid(photocredits:APSanAspringcrew634
ofSanfordCompany).PanelsF,G,Hhighlightthebeaksize,thepresenceof635
rotating hooks on its tentacles, and eye lens size of the colossal squid636
(copyright:MuseumofNewZealandTePapaTongarewa).637
638
Figure3–Numberofcolossalsquid(Mesonychoteuthishamiltoni)specimenscaught639
in nets (RMT‐ Rectangular midwater trawl, IKMT – Isaacs‐Kidd Midwater640
trawl, and other trawls) within the Southern Ocean, and the relationship641
betweenjuvenilemantlelength(mm)anddepthofcapture.642
643
Figure 4 – Map of the locations of all known colossal squid (Mesonychoteuthis644
hamiltoni) occurrences. “Squid” symbol represents locations obtained from645
nets and the other symbols represent predator diet studies (penguins,646
seabirds,fishandsharks)[modifiedfromXavieretal.(2015b)].647
648
Figure5–Predictedmantle length(mm)of thecolossalsquid(Mesonychoteuthis649
hamiltoni)foundinthestomachsremains(namelybeaks)ofitspredators.The650
estimatedvaluesarebasedonbeaks’morphometrics(namely lowerrostral651
length)andtherespectiveallometricequation(seeXavierandCherel2009).652

23
Abbreviations: A.p. – Aptenodytes patagonicus; Dio.e. – Diomedia exulans;653
Diss.e. ‐ Dissostichus eleginoides; G.m. ‐ Globicephala melaena; H.p. ‐654
Hyperoodonplanifrons;P.f.–Phoebetriafusca;P.m.–Physetermacrocephalus;655
P.p. ‐ Phoebetria palpebrata; S.m. – Somniosus cf Microcephalus; T.c. ‐656
Thalassarche chrysostoma. The references for beak data (for the different657
predatorspecies)canbefoundinTable1.658
659
Figure6–δ15Nvalues(trophiclevel)ofsquid(greybars)andothermarineanimals660
collected in the Southern Ocean. * ‐ corrected (beak) values. Data from661
Guerreiroetal.(2015),Stowasseretal.(2012)andreferenceswithin.662
663
Figure7–Biggesteyes(mm)intheocean.DatafromWalls(1942),LandandNilsson664
(2002)andNilssonetal.(2012).665
666
Figure8–Metabolicecologyofthecolossalsquid(Mesonychoteuthishamiltoni)in667
the Southern Ocean. Panel A: Effect of size in the routine mass‐specific668
metabolic rate of deep‐sea cranchiid squids (blue circles, from a 0.017g669
Liocranchia valdiviae to 500 kg M. hamiltoni) and of other cephalopod670
counterparts, including loliginids(green line), coastalommastrephids (dark671
red line), coastal benthic octopods (red line), pelagic gelatinous octopods672
(lightblue line)andvampiresquid(purple line).Valueof20kggiantsquid673
(Architeuthisspp.;solidtriangle)wasbasedontheactivityofcitratesynthase674
measuredinmantlemuscle(Seibeletal.2000).Rateswerestandardizedto675
1.5ºC assuming a Q10 of 2. Data from Rosa and Seibel (2010) and Seibel676
(2007).PanelsBandCrepresentdepth‐relatedchangesintemperatureinthe677
circum‐AntarcticSouthernOcean.Dashedlinesrepresentsub‐Antarcticareas678
(50.58S0.58E;50.58S90.58E;50.58S179.58W;50.58S90.58W)andsolidlines679
represent Antarctic areas (60.58S0.58E; 60.58S90.58E; 60.58S179.58W;680
60.58S90.58W);(B)projectedenergyconsumption(kcalday‐1;basedon4.7681
kcal/lO2)asafunctionofdepth(temperaturedependenceof2)inthecircum‐682
AntarcticSouthernOceanregion.Thicksolidlinerepresentsthemeanvalues683
obtainedfromthedifferenttemperatureprofiles.Thindashedlinerepresents684

24
the95%confidence interval for themeans (modified fromRosaandSeibel685
2010).686
687
Figure9–Photophoresofthecolossalsquid(Mesonychoteuthishamiltoni).PanelA688
showsthelightorganattherearoftheeyeball,whilePanelBshowstheeyelid689
beingpulledbacktorevealpartofthelightorgan(attherearoftheeyeball)690
(copyright: Museum of New Zealand Te Papa Tongarewa). Panels C and D691
show the colossal squid’s photocytes containing paracrystalline aggregates692
(needle‐likeorrectanglestructures)andPanelEshowsthereflectorplatelets693
with edges of comb‐like appearance (modified from Herring et al. 2002;694
copyright:JohnWiley&Sons,Inc.).695
696