optimasi produksi bioetanol dari sirup sorgum
TRANSCRIPT

OPTIMASI PRODUKSI BIOETANOL DARI SIRUP SORGUM (Sorghum bicolor) MENGGUNAKAN KHAMIR POTENSIAL
DARI BATANG DAN BIJI SORGUM
SKRIPSI
oleh SITI RODIYAH 145090101111004
JURUSAN BIOLOGI FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS BRAWIJAYA MALANG
2018

OPTIMASI PRODUKSI BIOETANOL DARI SIRUP SORGUM (Sorghum bicolor) MENGGUNAKAN KHAMIR POTENSIAL
DARI BATANG DAN BIJI SORGUM
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains dalam Bidang Biologi
oleh SITI RODIYAH 145090101111004
JURUSAN BIOLOGI FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS BRAWIJAYA MALANG
2018

ii
HALAMAN PENGESAHAN SKRIPSI
OPTIMASI PRODUKSI BIOETANOL DARI SIRUP SORGUM
(Sorghum bicolor) MENGGUNAKAN KHAMIR POTENSIAL
DARI BATANG DAN BIJI SORGUM
SITI RODIYAH
145090101111004
Telah dipertahankan di depan Majelis Penguji
pada tanggal 20 Juli 2018 dan dinyatakan memenuhi syarat untuk
memperoleh gelar Sarjana Sains dalam bidang Biologi
Menyetujui
Pembimbing
Tri Ardyati, M.Agr., Ph.D
NIP 196712131991032001
Mengetahui
Ketua Program Studi S-1 Biologi
Fakultas MIPA Universitas Brawijaya
Rodiyati Azrianingsih, S.Si., M.Sc., Ph.D
NIP. 197001281994122001

iii
HALAMAN PERNYATAAN SKRIPSI
Saya yang bertanda tangan di bawah ini:
Nama : Siti Rodiyah
NIM : 145090101111004
Jurusan : Biologi
Penulis Skripsi Berjudul : Optimasi Produksi Bioetanol dari Sirup
Sorgum (Sorghum Bicolor) menggunakan
Khamir Potensial dari Batang dan Biji Sorgum
Dengan ini menyatakan bahwa:
1. Skripsi ini adalah benar-benar karya saya sendiri dan bukan hasil
plagiat dari karya orang lain. Karya-karya yang tercantum dalam
Daftar Pustaka Skripsi ini semata-mata digunakan sebagai acuan
atau referensi
2. Apabila kemudian hari diketahui bahwa isi Skripsi saya
merupakan hasil plagiat, maka saya bersedia menanggung akibat
hukum dari keadaan tersebut
Demikian pernyataan ini dibuat dengan segala kesadaran
Malang, 24 Juli 2018
Yang menyatakan,
Siti Rodiyah
145090101111004

iv
PEDOMAN PENGGUNAAN SKRIPSI
Skripsi ini tidak dipublikasikan namun terbuka untuk umum dengan
ketentuan bahwa hak cipta ada pada penulis. Daftar pustaka
diperkenankan untuk dicatat, tetapi pengutipan hanya dapat dilakukan
seizin penulis dan harus disertai kebiasaan ilmiah untuk
menyebutkannya.

v
Optimasi Produksi Bioetanol dari Sirup Sorgum (Sorghum bicolor)
Menggunakan Khamir Potensial dari Batang dan Biji Sorgum
Siti Rodiyah, Tri Ardyati
Jurusan Biologi,Fakultas Matematika dan Ilmu Pengetahuan Alam,
Universitas Brawijaya, Malang
2018
ABSTRAK
Tanaman sorgum (Sorghum bicolor) memiliki kadar gula tinggi pada
bagian batang yang dapat dimanfaatkan untuk produksi bioetanol sebagai
bahan bakar alternatif. Penelitian ini bertujuan untuk mengeksplorasi
kemampuan khamir indigenous batang dan biji sorgum dalam
memproduksi etanol, mengetahui pengaruh penambahan kadar nitrogen
pada media fermentasi untuk produksi etanol dari sirup sorgum dan
mengidentifikasi khamir potensial berdasarkan daerah Internal
Transcribed Spacer (ITS) rDNA. Isolat khamir diisolasi dari batang dan
biji sorgum menggunakan teknik cawan tuang menggunakan media
YMEA dan dimurnikan menggunakan metode dilusi bertingkat. Isolat
yang diperoleh diuji toleransi terhadap kadar etanol menggunakan media
YME cair yang mengandung etanol dengan konsentrasi 0%, 5%, 10% dan
15%, selanjutnya diuji kemampuan isolat dalam menghasilkan etanol.
Isolat unggul digunakan untuk memfermentasi sirup sorgum dengan
penambahan 0,2 % dan 0,5 % ammonium sulfat ((NH4)2SO4). Isolat
unggul diidentifikasi spesies berdasarkan daerah ITS menggunakan
primer ITS 4 dan ITS 5. Hasil isolasi didapatkan delapan isolat khamir
dengan morfologi yang berbeda dan diperoleh dua isolat unggul yang
toleran terhadap kadar etanol 10 – 15% yaitu J1 dan J3 dan mampu
menghasilkan etanol sebesar 7,17 % dan 7,93 %. Penambahan kadar
nitrogen 0,5% menghasilkan etanol lebih tinggi dibandingkan
penambahan kadar nitrogen 0,2%. Isolat J1 dan J3 mampu menghasilkan
etanol dengan kadar berturut-turut 14,05 % dan 10,73 %. Isolat J1
teridentifikasi sebagai Candida tropicalis dengan similaritas 99 %
sedangkan isolat J3 teridentifikasi sebagai Wickerhamomyces anomalus
dengan similaritas 99%.
Kata kunci: fermentasi etanol, kadar nitrogen, khamir, sirup sorgum

vi
Optimization of Bioethanol Production from Soghum Syrup
(Sorghum bicolor) using Potential Ethanol-Producing Yeasts from
Sorghum Stalk and Grains
Siti Rodiyah, Tri Ardyati
Biology Department, Faculty of Mathematic and Natural Sciences,
University of Brawijaya, Malang
2018
ABSTRACT
Sorghum (Sorghum bicolor) is a plant contains high sugar
content that can be used as substrate for bioethanol production, an
alternative biofuel. This study aimed to explore the ability of indigenous
yeast from sorghum stalk and grain in ethanol production, to evaluate the
effect of nitrogen source on fermentation medium for ethanol production
from sorghum syrup and to identify the potential yeasts based on Internal
Transcribed Spacer (ITS) rDNA region. Yeasts were isolated from
sorgum stalk and grains using pour plate methods and serial dilution
method for purification of isolates. The selected isolate then used for
sorghum syrup fermentation with addition of 0,2 % and 0,5 % ammonium
sulfate. Identification of isolates were based on ITS region using primers
ITS 4 and ITS 5. Isolation resulted eight yeast isolates with different
morphology, two isolates were tolerant to ethanol concentration of 0 to
15 %, names ada J1 and J3 isolates and capable to produce ethanol. The
addition of 0.5% nitrogen source showed better production of ethanol
than 0.2 %. Isolate J1 and J3 produced ethanol with concentration of
14.05 % and 10.73 % respectively. Isolate J1 was identified as Candida
tropicalis with 99 % similarity and isolate J3 was identified as
Wickerhamomyces anomalus with 99 % similarity.
Keywords: ethanol fermentation, nitrogen content, sorghum syrup, yeast

vii
KATA PENGANTAR
Alhamdulillahi Rabbil Alamiin, dengan ungkapan rasa syukur
kehadirat Allah Yang Maha Kuasa, akhirnya penulis dapat
menyelesaikan penyusunan skripsi yang merupakan syarat untuk
memperoleh gelar Sarjana Sains dalam bidang Biologi di Fakultas
Matematika dan Ilmu Pengetahuan Alam Universitas Brawijaya Malang.
Pada kesempatan ini penulis ingin menyampaikan ucapan terimakasih
kepada:
1. Ibu Tri Ardyati, M.Agr., Ph.D selaku dosen pembimbing yang
telah dengan sabar mendampingi, memberi pengarahan, serta
tambahan ilmu yang berguna bagi penulis.
2. Bapak Dr. Suharjono, MS dan bapak Yoga Dwi Jatmiko, S.Si.,
M.App.Sc selaku dosen penguji yang telah memberi saran yang
bermanfaat demi perbaikan penyusunan skripsi.
3. Ibu Nanik Dwi Rahayu selaku laboran Laboratorium Mikrobiologi
yang telah memfasilitasi peminjaman alat dan media sehingga
penelitian ini dapat terlaksana.
4. Orang tua penulis atas segala doa, dukungan dan motivasi yang
tidak terkira.
5. Eli Wulandari, Fatimatul Isnaeni, Ameta, Naura dan Azka selaku
saudara penulis yang telah memberikan dukungan tidak terkira
pada penulis.
6. Hani Khairina, Violita Yulpha, Wiaam Raniah dan Sayyidatul
Awalia sebagai rekan seperjuangan, Alifia Issabella yang
memberikan bimbingan dan dukungan.
7. M. Rifqi Ramadhan yang telah menemani, mendukung dan
memberikan motivasi.
8. Rekan-rekan peneliti Laboratorium Mikrobiologi dan rekan-rekan
Biologi Angkatan 2014, serta seluruh civitas akademik Jurusan
Biologi Fakultas MIPA Universitas Brawijaya.
Penulisan skripsi ini merupakan upaya optimal penulis sebagai sarana
terbaik dalam pengembangan ilmu pengetahuan. Saran dan kritik yang
membangun sangat diharapkan untuk menjadikan karya ini semakin
bermanfaat.
Malang, 24 Juli 2018
Penulis

viii
DAFTAR ISI
Halaman
ABSTRAK .................................................................................... v
ABSTRACT ................................................................................. vi
KATA PENGANTAR ................................................................. vii
DAFTAR ISI ................................................................................ viii
DAFTAR TABEL ........................................................................ x
DAFTAR GAMBAR ................................................................... xi
DAFTAR LAMPIRAN ................................................................ xii
DAFTAR LAMBANG/SINGKATAN ....................................... xiii
BAB I PENDAHULUAN
1.1 Latar Belakang ............................................................
1.2 Rumusan Masalah .......................................................
1.3 Tujuan Penelitian ........................................................
1.4 Manfaat Penelitian ......................................................
1
2
2
2
BAB II TINJAUAN PUSTAKA
2.1Tanaman Sorgum (Sorghum bicolor) dan
Keunggulannya untuk Produksi Etanol ......................
2.2 Khamir untuk Produksi Etanol ..................................
2.3 Jalur Fermentasi Etanol .............................................
2.4 Faktor yang Mempengaruhi Produksi Etanol oleh
Khamir ........................................................................
2.5 Identifikasi Molekular Khamir berdasarkan
Internal Transcribed Spacer (ITS) .............................
4
6
6
9
11
BAB III METODE PENELITIAN
3.1 Waktu dan Tempat Penelitian ....................................
3.2 Isolasi dan Skrining Khamir ......................................
3.3 Fermentasi Nira Sorgum ............................................
3.4 Identifikasi Khamir Potensial ....................................
13
13
14
16
BAB IV HASIL DAN PEMBAHASAN
4.1 Jumlah Sel Khamir yang Diisolasi dan Skrining
Khamir dari Batang dan Biji Sorgum .........................
4.2 Produksi Etanol dari Fermentasi Sirup Sorgum ........
4.2.1 Isolat J1 ............................................................
4.2.2 Isolat J3 ............................................................
18
21
22
25

ix
4.2.3 Saccharomyces cerevisiae ...............................
4.3 Hasil Identifikasi Isolat Potensial ..............................
27
29
BAB V KESIMPULAN DAN SARAN
5.1 Kesimpulan ................................................................
5.2 Saran ..........................................................................
34
34
DAFTAR PUSTAKA .................................................................. 35
LAMPIRAN ................................................................................. 39

x
DAFTAR TABEL
Nomor Halaman
1 Kandungan Nutrisi Biji Sorgum Dibandingkan Bahan
Pokok Lain .....................................................................
4
2 Kandungan Nira Sorgum Dibandingkan Nira Tebu ........ 5
3 Faktor Fisika dan Non Nutrisi yang Memengaruhi
Proses Fermentasi Etanol ...............................................
10
4 Komposisi Master Mix untuk Amplifikasi PCR ............. 16
5 Kondisi Siklus Amplifikasi PCR .................................... 17

xi
DAFTAR GAMBAR
Nomor Halaman
1 Proses konversi atom karbon dari satu molekul gula
menjadi karbondioksida dan etanol.................................
7
2 Jalur fermentasi etanol pada S. cerevisiae dan Z. mobilis 8
3 Konversi asam piruvat menjadi etanol ............................ 9
4 Daerah ITS pada gen rRNA kapang ................................ 12
5 Morfologi sel khamir hasil isolasi ................................... 18
6 Toleransi isolat terhadap kadar etanol.............................. 19
7 Kadar etanol yang dihasilkan oleh masing-masing isolat
menggunakan media standar dan media sirup sorgum ....
20
8 Kadar etanol yang dihasilkan masing-masing isolat
dengan penambahan kadar nitrogen 0,2 % dan 0,5 % .....
22
9 Kadar etanol, jumlah sel dan gula reduksi pada fermentasi
sirup sorgum oleh isolat J1 ..............................................
24
10 Kadar etanol, jumlah sel dan gula reduksi pada fermentasi
nira sorgum oleh isolat J3 ................................................
26
11 Kadar etanol, jumlah sel dan gula reduksi pada fermentasi
nira sorgum oleh Saccharomyces cerevisiae .....................
28
12 Diagram lokasi primer yang digunakan untuk amplifikasi
daerah ITS1-5,8S-ITS2 .....................................................
29
13 Pohon filogeni dari isolat J1 yang teridentifikasi sebagai
Candida tropicalis ...........................................................
31
14 Pohon filogeni dari isolat J3 yang teridentifikasi sebagai
Wickerhamomyces anomalus ...........................................
32

xii
DAFTAR LAMPIRAN
Nomor Halaman
1 Hasil BLAST isolat J1 ........................................................ 37
2 Hasil BLAST isolat J3 ........................................................ 38
3 Matriks jarak isolat J1 dengan sekuen pembanding ........... 39
4 Matriks jarak isolat J3 dengan sekuen pembanding ........... 39
5 Hasil elektroforesis DNA hasil PCR ................................... 39
6 Hasil Nano Drop DNA ........................................................ 40
7 Sekuen isolat hasil sekuensing ............................................ 40
8 Kurva standar glukosa ......................................................... 41
9 Analisis statistik toleransi kadar etanol ............................... 42
10 Hasil analisis deskriptif kadar etanol menggunakan media
standar dan media sirup sorgum ..........................................
43
11 Hasil uji Tukey kadar etanol menggunakan media standar
dan sirup sorgum .................................................................
44
12 Analisis statistik kadar etanol dengan penambahan nitrogen
0,2 % dan 0,5 % ....................................................................
45

xiii
DAFTAR LAMBANG DAN SINGKATAN
Simbol/singkatan Keterangan
°C derajat celcius
ATP Adenosin Tri Pospat
bp basepair
cfu Colony forming unit
CO2 Karbon dioksida
g gram
gL-1 Gram per liter
mL mililiter
nm Nano meter
pH Power of Hydrogen
α alfa
λ Panjang gelombang

1
BAB I
PENDAHULUAN
1.1 Latar Belakang
Produksi etanol dari sumber terbarukan mendapatkan banyak
perhatian dalam beberapa tahun terakhir karena persediaan bahan bakar
minyak yang terus berkurang. Bahan bakar terbarukan dapat berasal dari
berbagai jenis tanaman pangan dengan kadar gula tinggi seperti tebu,
jagung, singkong, beras, kentang dan sebagainya. Terdapat alternatif
tanaman lain yang tidak dimanfaatkan sebagai bahan pangan dan
memiliki kandungan gula yang tinggi yaitu sorgum. Sorgum merupakan
salah satu sumber energi terbarukan yang mempunyai potensi besar untuk
mengatasi persediaan bahan bakar dunia dan membantu mengurangi
emisi gas yang dihasilkan oleh minyak bumi (Subagio & Aqil, 2013;
Iticha, 2016).
Sorgum merupakan salah satu jenis tanaman serealia yang tumbuh di
daerah tropis. Sorgum dapat mempertahankan kondisi fisiologisnya pada
kondisi kelembaban tanah yang rendah sehingga dapat dengan mudah
dibudidayakan walaupun di daerah kering. Sorgum juga diketahui
memiliki toleransi yang luas terhadap daerah yang tergenang air maupun
lahan marginal. Sorgum juga relatif lebih tahan terhadap gangguan hama
dan penyakit tanaman pertanian. Biji sorgum dapat digunakan sebagai
bahan pangan pengganti beras dan jagung serta dapat digunakan sebagai
bahan baku industri pangan seperti gula, monosodium glutamat (MSG),
asam amino dan industri minuman (Sirappa, 2003). Pemanenan biji
sorgum menyisakan hasil samping berupa daun dan batang sorgum. Daun
sorgum dapat digunakan untuk pakan ternak sapi dan batang sorgum
seringkali dibuang percuma. Batang sorgum tersebut dapat diperas untuk
mendapatkan nira sorgum yang banyak mengandung gula (Rutto dkk.,
2013).
Nira dari batang sorgum dapat digunakan sebagai bahan baku
pembuatan gula, sirup, dan etanol. Sorgum menghasilkan 23 %
karbohidrat yang dapat difermentasi, membutuhkan pupuk nitrogen 37 %
lebih sedikit, dan irigasi 17 % lebih sedikit dibandingkan dengan jagung.
Sorgum juga dapat menghasilkan etanol lebih banyak dibandingkan
jagung pada musim kemarau (Rutto dkk., 2013). Sebagai sumber energi
alternatif untuk produksi etanol, bahan baku dari sorgum tergolong murah
dan dapat dengan mudah didapatkan.

2
Produksi etanol menggunakan bahan baku berupa substrat alami
membutuhkan agen untuk fermentasi. Beberapa jenis mikroorganisme
khususnya kelompok khamir diketahui dapat mengubah gula menjadi
etanol melalui fermentasi anaerobik. Jenis khamir yang diketahui dapat
menghasilkan etanol yaitu Saccharomyces cerevisiae (Naser, 2014).
Karakteristik tempat tumbuh khamir di alam yaitu pada lingkungan yang
mengandung gula, kelembaban tinggi dan cenderung berair seperti pada
sayur dan buah-buahan busuk (Rao dkk., 2008). Produksi bioetanol
dipengaruhi oleh beberapa faktor, antara lain mikroorganisme dan media
pertumbuhan. Kandungan karbon dan nitrogen dalam media
pertumbuhan khamir diketahui sebagai komponen utama bagi
metabolisme sel khamir (Cruz dkk., 2002). Oleh karena itu perlu
dilakukan eksplorasi kemampuan khamir indigenous untuk
memfermentasi nira sorgum dan diketahui komposisi media pertumbuhan
yang optimal untuk produksi bioetanol.
1.2 Rumusan Masalah
Rumusan masalah dari penelitian ini yaitu:
1. Bagaimana kemampuan khamir indigenous batang dan biji
sorgum dalam produksi etanol?
2. Bagaimana pengaruh kadar nitrogen pada media fermentasi
untuk produksi etanol dari sirup sorgum?
3. Khamir jenis apakah yang potensial untuk produksi etanol?
1.3 Tujuan Penelitian
Tujuan dari penelitian ini yaitu
1. Mengeksplorasi kemampuan khamir indigenous batang dan biji
sorgum dalam produksi etanol.
2. Mengetahui pengaruh kadar nitrogen pada media fermentasi
untuk produksi etanol dari sirup sorgum.
3. Mengidentifikasi khamir potensial berdasarkan daerah Internal
Transcribed Spacer (ITS) rDNA.
1.4 Manfaat Penelitian
Manfaat dari penelitian ini yaitu mendapatkan khamir potensial yang
dapat dikembangkan untuk produksi etanol dan mengetahui komposisi

3
media yang optimal terutama komposisi karbon dan nitrogen dalam
media fermentasi sirup sorgum untuk produksi etanol sehingga dapat
meningkatkan produksi bioetanol dengan harapan dapat dimanfaatkan
sebagai bahan bakar alternatif untuk mengurangi penggunaan minyak
bumi.

4
BAB II
TINJAUAN PUSTAKA
2.1 Tanaman Sorgum (Sorghum bicolor) dan Kenggulannya untuk
Produksi Bioetanol
Sorgum (Sorghum bicolor L.) merupakan salah satu jenis tanaman
serealia yang memiliki potensi besar di bidang pangan dan dikembangkan
di Indonesia. Tanaman sorgum merupakan tanaman C4 yang adaptif
terhadap stress lingkungan dan memiliki efisiensi fotosintesis tinggi
(Almodares & Hadi, 2009). Tanaman ini memiliki toleransi yang luas
terhadap daerah kering maupun tergenang sehingga cocok ditanam di
berbagai tipe tanah dan daerah, serta relatif tahan terhadap gangguan
hama dan penyakit tanaman. Biji sorgum dapat digunakan sebagai bahan
pangan dan berbagai kebutuhan industri sebagai bahan baku untuk
pembuatan gula, monosodium glutamat (MSG), asam amino dan industri
minuman. Prospek terbesar penggunaan biji sorgum yaitu sebagai bahan
pangan pengganti beras, jagung dan berbagai makanan pokok lainnya
karena diketahui memiliki kandungan nutrisi yang hampir sama (Tabel 1)
(Sirappa, 2003).
Tabel 1. Kandungan nutrisi biji sorgum dibandingkan bahan pokok lain
Bahan
pangan
Kalori
(kal)
Protein
(g)
Le-
mak
(g)
Karbo-
hidrat
(g)
Air
(%)
Serat
(%)
Ca
(mg)
P
(mg)
Fe
(mg)
Sorgum 332 11 3,3 73 11,2 2,3 28 287 4,4
Beras 360 7 0,7 79 9,8 1 6 147 0,8
Jagung 361 9 4,5 72 13,5 2,7 9 380 4,6
Kentang 83 2 0,1 19 - - 11 56 0,7
Ubi kayu 157 1,2 0,3 35 63 - 33 40 0,7
Ubi jalar 123 1,8 0,7 28 - - 30 49 0,7
Terigu 365 8,9 1,3 77 - - 16 106 1,2
Selain biji, bagian lain dari tanaman sorgum juga banyak
dimanfaatkan, seperti batang sorgum yang diketahui memiliki kandungan
gula tinggi hampir sama dengan tebu (Tabel 2). Kandungan amilum pada
sorgum lebih tinggi daripada tebu yang menjadi masalah pada proses
pengkristalan gula sorgum sehingga gula berbentuk cair (Sirappa, 2003).
Kandungan gula yang tinggi tersebut dapat dijadikan sebagai bahan baku

5
pembuatan bioetanol. Batang sorgum diperas untuk mendapatkan nira
yang mengandung glukosa, sukrosa dan bentuk gula lainnya yang dapat
dikonversi menjadi etanol. Tanaman sorgum yang dijadikan sebagai
bahan baku etanol dicirikan dengan adanya akumulasi karbohidrat
terfermentasi dalam batang yang mencapai 15 – 25 % sehingga untuk
produksi bioetanol akan lebih menguntungkan apabila pengembangan
tanaman dilakukan di daerah kering (Subagio & Aqil, 2013). Sukrosa,
glukosa dan fruktosa merupakan gula yang dominan pada sirup sorgum
dengan konsentrasi berturut-turut 42,38 gL-1, 23,17 gL-1 dan 28,95 gL-1.
Total konsentrasi dari ketiga gula tersebut mencapai 94,5 gL-1 dengan
nilai rata-rata antara 70 – 110 gL-1 (Luo dkk., 2014).
Tabel 2. Kandungan nira sorgum dibandingkan nira tebu
Komposisi Nira Sorgum Nira Tebu
Brix (%) 13,60 – 18,40 12 – 19
Sukrosa (%) 10 – 14,40 9 – 17
Gula reduksi (%) 0,75 – 1,35 0,48 – 1,52
Gula total (%) 11 - 16 10 – 18
Amilum (ppm) 209 -1.764 1,50 – 95
Asam akonitat (%) 0,56 0,25
Abu (%) 1,28 – 1,57 0,40 – 0,70
Tanaman lain yang biasa digunakan untuk produksi bioetanol yaitu
tebu, jagung, sagu, singkong, ketela dan sebagainya. Namun diantara
tanaman tersebut, sorgum memiliki beberapa karakteristik yang lebih
unggul diantaranya efisien dalam mengubah energi matahari dan
karbohidrat yang dihasilkan tinggi, toleran terhadap berbagai jenis
lingkungan dengan berbagai fungsi, memiliki konsentrasi gula tinggi
yang dapat terfermentasi langsung, semua komponen dari tanaman
sorgum memiliki nilai ekonomi mulai dari biji, batang dan serat yang
dihasilkan dari hasil pemerasan batang. Waktu tumbuh sorgum lebih
pendek yaitu antara 3-5 bulan dibandingkan tebu antara 10 – 12 bulan dan
kuantitas air yang dibutuhkan hanya 1/3 dari tebu (Almodares & Hadi,
2009).

6
2.2 Khamir untuk Produksi Bioetanol
Khamir merupakan jamur ascomycota atau basidiomycota yang dapat
bereproduksi dengan tunas atau membelah dan menghasilkan spora yang
tidak diselubungi badan buah. Diversitas spesies khamir di niche khusus
ditentukan berdasarkan kemampuannya memanfaatkan sumber karbon
dan nutrisi selektif yang sesuai dengan habitatnya. Khamir dapat diisolasi
dari daerah terestrial, akuatik atau aerial, namun tanaman merupakan
habitat terbesar dari khamir. Sebagian besar niche khamir pada
lingkungan berair dari tanaman yang mengandung gula dan
persebarannya ke tanaman lain dibantu oleh serangga (Azhar dkk., 2017).
Berbagai jenis khamir dikomersialisasi dan digunakan dalam produksi
etanol atau CO2, termasuk juga strain spesifik yang digunakan untuk
pengembang roti, wine, beer dan etanol untuk bahan bakar. Khamir
Saccharomyces cerevisiae telah dikenal secara luas digunakan dalam
produksi etanol dan keperluan lain dengan strain yang spesifik (Naser,
2014). S. cerevisiae banyak digunakan di industri etanol karena memiliki
toleransi pH yang luas. Khamir dalam pembuatan roti juga digunakan
sebagai starter untuk produksi etanol karena biaya yang murah dan dapat
dengan mudah diperoleh (Azhar dkk., 2017).
Hambatan utama dalam fermentasi gula yaitu peningkatan suhu antara
35 – 45 °C dan konsentrasi etanol yang lebih dar 20 %. Laju pertumbuhan
dan metabolisme khamir meningkat dengan meningkatnya suhu sampai
pada titik optimum. Peningkatan konsentrasi etanol selama fermentasi
dapat menghambat pertumbuhan dan viabilitas sel khamir. Terhambatnya
pertumbuhan khamir pada media yang mengandung etanol tinggi juga
menghambat produksi etanol. Strain khamir etanol toleran dan
termotoleran yang dapat bertahan pada kondisi stress medium dapat
diisolasi dari sumber alam seperti tanah, air, tanaman dan hewan. Hal
tersebut dikarenakan sel khamir beradaptasi dengan lingkungan dalam
waktu yang lama dan melewati seleksi alam (Azhar dkk., 2017).
2.3 Jalur Fermentasi Etanol
Produksi bioetanol sebagian besar berasal dari proses fermentasi gula.
Gula seperti glukosa pada tanaman diperoleh sebagai hasil dari proses
fotosintesis. Melalui proses fermentasi oleh mikroorganisme, 1 molekul
gula diubah menjadi 2 molekul etanol dan CO2 (persamaan 1; Gambar 1)
(Barnett, 2003). Fermentasi merupakan dekomposisi lambat oleh
mikroorganisme dari molekul organik kompleks seperti pati menjadi

7
molekul lebih sederhana seperti etanol. Fermentasi gula untuk produksi
etanol terjadi dalam kondisi anaerobik (Naser, 2014).
C6H12O6 → 2 C2H5OH + 2 CO2 + panas .............................................. (1)
(Barnett, 2003)
Gambar 1. Proses konversi atom karbon dari satu molekul gula
menjadi karbondioksida dan etanol
Etanol diproduksi dari glukosa melalui konsumsi fermentatif piruvat.
Glikolisis merupakan proses yang mengubah glukosa menjadi produk
teroksidasi, piruvat, yang mensuplai ATP untuk produksi biomassa.
Piruvat dapat difermentasi menjadi etanol dengan reaksi piruvat
dekarboksilase (PDC) dan alkohol dehidrogenase (ADH) dengan
melepaskan satu karbon sebagai karbon dioksida (CO2) di bawah kondisi
anaerobik. Proses fermentasi etanol dapat dipelajari secara intensif pada
S. cerevisiae dan E. coli, dikarenakan kemajuan teknologi dalam bidang
rekayasa genetik. Spesies lain yang juga dapat dijadikan sebagai alternatif
yaitu Zymomonas mobilis yang memiliki kemampuan memproduksi
etanol melalui jalur Entner-Doudoroff (ED) tanpa melaui jalur Embden-
Meyerhof-Parnas (EMP) untuk glikolisis (Gambar 2). Meskipun jalur
EMP merupakan jalur utama glikolisis di sebagian besar eukariotik dan
prokariotik, jalur glikolisis lebih beragam pada prokariotik. Di antara
jenis-jenis jalur glikolisis, jalur ED merupakan jalur yang banyak
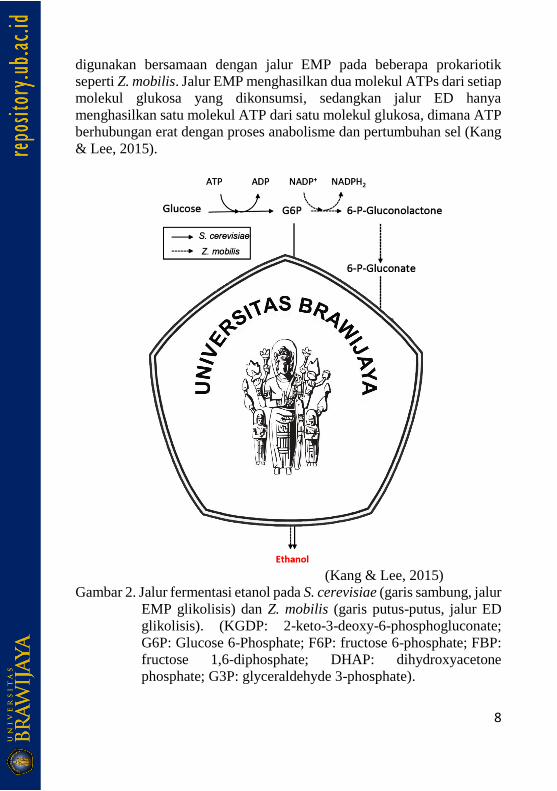
8
digunakan bersamaan dengan jalur EMP pada beberapa prokariotik
seperti Z. mobilis. Jalur EMP menghasilkan dua molekul ATPs dari setiap
molekul glukosa yang dikonsumsi, sedangkan jalur ED hanya
menghasilkan satu molekul ATP dari satu molekul glukosa, dimana ATP
berhubungan erat dengan proses anabolisme dan pertumbuhan sel (Kang
& Lee, 2015).
(Kang & Lee, 2015)
Gambar 2. Jalur fermentasi etanol pada S. cerevisiae (garis sambung, jalur
EMP glikolisis) dan Z. mobilis (garis putus-putus, jalur ED
glikolisis). (KGDP: 2-keto-3-deoxy-6-phosphogluconate;
G6P: Glucose 6-Phosphate; F6P: fructose 6-phosphate; FBP:
fructose 1,6-diphosphate; DHAP: dihydroxyacetone
phosphate; G3P: glyceraldehyde 3-phosphate).

9
Reaksi pertama yang dilakukan dalam fermentasi yaitu glikolisis.
Proses katabolisme tersebut merombak D-glucose menjadi piruvat untuk
menghasilkan 2 molekul ATP setiap molekul glukosa. Khamir merubah
piruvat menjadi etanol dan karbon dioksida pada fermentasi alkohol dan
keseluruhan proses tersebut memberikan energi kimiawi bagi sel yeast
yang disimpan pada ikatan fosfat dari ATP (Gambar 3). Asam piruvat
akan dirubah menjadi asetaldehid melalui reaksi enzimatik piruvat
dekarboksilase. Asetaldehid kemudian dirubah menjadi etanol melalui
reaksi enzim alkohol dehidrogenase (Barnett, 2003).
(Barnett, 2003)
Gambar 3. Konversi asam piruvat menjadi etanol
2.4 Faktor yang Mempengaruhi Produksi Etanol oleh Khamir
Faktor yang menentukan laju fermentasi diantaranya kelimpahan sel
khamir dalam medium, viabilitas sel khamir, tingkat oksigen terlarut,
konsentrasi nitrogen terlarut, konsentrasi karbon terlarut dan faktor
mineral lain yang dibutuhkan sel khamir untuk tumbuh. Selain itu juga
terdapat beberapa faktor fisika dan non nutrisi yang memengaruhi proses
fermentasi, diantaranya adalah suhu, pH, agitasi atau pengadukan, aerasi,
tekanan udara, volume inokulum, umur inokulum dan waktu fermentasi
(tabel 3) (Strobel & Sullivan, 1999).

10
Tabel 3. Faktor fisika dan non nutrisi yang memengaruhi proses
fermentasi etanol
Faktor Range setting
Suhu 10 – 40 °C
pH 3 – 9
Agitasi 50 – 500 rpm
Aerasi 0,1 – 2 vvm
Tekanan udara 1 – 15 lb/in2
Volume inokulum 1 – 15 % (v/v)
Umur inokulum 0,5 – 5 hari
Waktu fermentasi 0,8 – 14 hari
Menurut Waites dkk. (2005) faktor kunci yang berpengaruh pada hasil
fermentasi diantaranya adalah kualitas dan kuantitas inokulum, pemilihan
media, penentuan dan pengembangan nutrisi, suhu, pH dan gradien
oksigen. Produksi bioetanol dapat dipengaruhi oleh beberapa faktor
antara lain media. Media dalam pembuatan bioetanol harus memenuhi
kebutuhan dasar untuk pembentukan biomassa dan produk fermentasi
(Widayanti dkk., 2013). Karbohidrat dan nitrogen merupakan unsur
utama yang dibutuhkan untuk pertumbuhan suatu organisme dan interaksi
dari kedua nutrisi tersebut memegang peran penting dalam metabolisme
(Cruz dkk., 2002).
Sel khamir tumbuh pada kondisi dengan berbagai sumber energi. Sel
khamir lebih sering mengonsumsi sumber karbon berupa glukosa dan
fruktosa dibandingkan dengan sukrosa, rafinosa, gliserol, etanol atau
asetat (Broach, 2012). Konsentrasi gula dalam medium pertumbuhan
khamir harus seimbang. Kadar gula yang terlalu tinggi dapat menghambat
pertumbuhan sel khamir dan kadar etanol yang terlalu tinggi dapat
mematikan sel khamir. Glukosa yang melebihi 15 % (v/v) medium dapat
menghambat kinerja berbagai enzim yang dihasilkan khamir (Wiratno
dkk., 2014). Kandungan gula dalam medium dimanfaatkan oleh sel
khamir untuk pembentukan ATP yang diperlukan untuk penambahan
biomassa sel khamir. Glukosa akan dipecah dirubah menjadi produk
teroksidasi melalui glikolisis dengan hasil samping berupa etanol dalam
kondisi anaerobik (Kang & Lee, 2015).
Selain sumber karbon, ketersediaan nitrogen dalam media juga
merupakan faktor penting untuk pertumbuhan sel khamir. Berbagai jenis

11
sumber nitrogen dapat ditambahkan ke dalam medium pertumbuhan
seperti amonium sulfat, urea, ekstrak khamir, pepton dan sumber lainnya.
Sumber nitrogen yang umum digunakan yaitu amonium sulfat dan urea
karena harga yang terjangkau untuk produksi etanol (Li dkk., 2017). Nira
sorgum sudah mengandung NH4+ sebanyak 21,4 ppm dan NO3
- sebanyak
4,4 ppm sebagai sumber nitrogen (Laopaiboon dkk., 2009). Namun untuk
produksi etanol diperlukan penambahan sumber nitrogen tambahan untuk
mendukung pertumbuhan sel khamir. Kandungan nitrogen dalam
medium digunakan sel khamir untuk melangsungkan proses transkripsi,
metabolisme, dan kemampuan biosintesis lainnya. Terbatasnya
kandungan nitrogen dalam medium akan memperlambat laju
pertumbuhan sel khamir, terutama pada proses reduksi di biogenesis
ribosomal dan translasi (Broach, 2012).
2.5 Identifikasi Molekular Khamir berdasarkan Internal Transcribed
Spacer (ITS)
Sistron rRNA eukariotik terdiri atas gen 18S, 5,8S dan 28S rRNA yang
ditranskripsi menjadi satu unit oleh RNA polimerase I. Proses post-
transkripsional membagi sistron dan memisahkan dua celah atau internal
transcribed spacer (ITS). Kedua celah tersebut mengapit gen 5,8S dan
umumnya disebut daerah ITS (Scoch dkk., 2012). Daerah ITS umum
digunakan untuk mengidentifikasi kapang dan khamir hingga tingkat
spesies. Daerah ITS dipilih untuk identifikasi khamir karena merupakan
bagian kompleks gen rRNA, primer kapang universal yang bersifat
conserve, mudah diamplifikasi meskipun menggunakan jumlah DNA
sedikit karena memiliki banyak salinan pada gen RNA dan tingkat
variabilitas sekuen ITS yang tinggi (Calderone & San-Blas, 2008).
Daerah ITS yang umum diamplifikasi terdiri dari ITS1 – 5,8S – ITS2.
Primer ITS1 dan ITS4 digunakan untuk mengamplifikasi daerah ITS1
dan ITS2 (Hesham dkk., 2017). Identifikasi molekular kapang dan khamir
menghasilkan lebih dari 100.000 sekuen yang disimpan dalam database
yang dapat membantu keperluan identifikasi khamir berdasarkan penanda
ITS (Bellemain dkk., 2010).
Daerah ITS dapat diamplifikasi dengan Polymerase Chain Reaction
(PCR) dengan amplikon yang dihasilkan sekitar 527-700 bp (Das & Deb,
2015). Prinsip dasar PCR yaitu amplifikasi urutan DNA target dan
meminimalkan amplifikasi DNA non-target. Oleh karena itu PCR
memerlukan primer spesifik untuk urutan basa target, dan komponen lain

12
yang dibutuhkan untuk menjalankan PCR. Ketika PCR selesai, dihasilkan
sekuen spesifik dalam jumlah banyak. Untai tunggal DNA dapat
dipisahkan melalui gel electrophoresis (Fatchiyah dkk., 2011).
(Korabecna, 2007)
Gambar 4. Daerah ITS pada gen rRNA kapang
Elektroforesis merupakan proses bergeraknya molekul bermuatan
pada suatu medan listrik. Kecepatan gerak suatu molekul bergantung
pada muatan, bentuk dan ukuran molekul. Molekul seperti asam nukleat
dan protein dapat dipisahkan melalui elektroforesis. Posisi molekul yang
terseparasi dapat dideteksi dengan pewarnaan. Elektroforesis suatu
molekul dibutuhkan suatu matriks penyangga untuk mencegah difusi
karena arus listrik. Penyangga yang dapat digunakan yaitu gel agarosa.
DNA yang terseparasi divisualisasi dengan penambahan ethidium
bromide yang dapat berikatan dengan DNA secara kuat melalui interaksi
di antara basa DNA (Fatchiyah dkk., 2011).

13
BAB III
METODE PENELITIAN
3.1 Waktu dan Tempat
Penelitian ini dilaksanakan pada Bulan Desember 2017 – Mei 2018 di
Laboratorium Mikrobiologi, Fakultas Matematika dan Ilmu Pengetahuan
Alam, Universitas Brawijaya, Malang.
3.2 Isolasi dan Seleksi Khamir Potensial
Isolat khamir yang digunakan diisolasi dari batang dan biji sorgum.
Sampel batang dan biji sorgum diperoleh dari Peternakan Sapi TAPOS
Bogor. Sel khamir diisolasi menggunakan metode cawan tuang dengan
media YMEA (3 gL-1 yeast extract, 3 gL-1 malt extract, 3 gL-1 pepton, 10
gL-1 glukosa dan 1,5 % agar) yang telah disterilisasi pada suhu 121°C
selama 15 menit (Nasreen dkk., 2014). Sampel batang dan biji sorgum
dibiarkan selama 48 jam di kebun tebu sebagai habitat alami untuk
menumbuhkan khamir alami dari batang dan biji sorgum. Sampel
ditimbang sebanyak 25 g dan dimasukkan ke dalam 225 mL NaCl 0,85
% dan dilakukan pengenceran berseri hingga 10-6. Sebanyak 0,1 mL
sampel dari setiap seri pengenceran dimasukkan ke dalam cawan Petri
dan dituang dengan media YMEA yang mengandung Streptomycin 50
µg/mL untuk menghambat pertumbuhan bakteri (Lee dkk., 2011; Ebabhi
dkk., 2013). Cawan Petri diinkubasi pada suhu 30 °C selama 48 jam.
Koloni yang tumbuh dengan karakter morfologi berbeda dimurnikan
menggunakan metode kuadran streak dan disubkultur pada YMEA
miring. Kultur isolat disimpan pada suhu 4 °C hingga perlakuan
selanjutnya.
Sel khamir yang telah murni diuji kemampuan tumbuh pada kadar
etanol tinggi. Kemampuan pertumbuhan sel khamir pada kadar etanol
tinggi dilakukan menggunakan media YME cair steril yang mengandung
etanol dengan konsentrasi 0 %, 5 %, 10 % dan 15 %. Volume medium
yang digunakan yaitu sembilan mL kemudian ditambahkan kultur cair
isolat berumur 24 jam sebanyak satu mililiter. Kultur diinkubasi pada
suhu ruang selama 48 jam (Ali & Khan, 2014). Kemampuan pertumbuhan
sel khamir pada kadar etanol tinggi dilakukan secara bertahap mulai dari
0%, kemudian isolat yan mampu bertahan diinokulasi sebanyak 10 %
pada kadar 5% dan seterusnya hingga kadar etanol 15%. Viabilitas sel

14
khamir pada medium diukur menggunakan spektrofotometer dengan λ
600 nm, kekeruhan medium pada jam ke-0 dibandingkan dengan
kekeruhan pada jam ke-48 untuk mengetahui adanya pertumbuhan sel
khamir. Hasil kemampuan pertumbuhan sel khamir pada beberapa kadar
etanol yang digunakan kemudian dianalisis menggunakan software SPSS
V16.0 for Windows dengan metode one-way ANOVA dan uji Tukey
dengan α=0,05.
Isolat khamir yang diperoleh diuji pula kemampuan fermentasinya
menggunakan media standar dengan glukosa 5 % dan media sirup
sorgum. Komposisi media standar yang digunakan yaitu glukosa 50 gL-1,
yeast extract 1 gL-1, KH2PO4 5 gL-1, (NH4)2SO4 2 gL-1 dan MgSO4 0,4 gL-
1 (Cheng dkk., 2009). Isolat khamir diuji kemampuannya dalam
memfermentasi sirup sorgum dengan kondisi fermentasi Brix medium 20,
inokulum 10 % (v/v) dengan densitas 106 cfu/mL. Kondisi tersebut
digunakan karena sebagian besar khamir mampu memfermentasi nira
sorgum pada Brix 20 dan jumlah inokulum tersebut merupakan jumlah
yang standar untuk fermentasi sirup sorgum (Luo dkk., 2014; Phutela &
Kaur, 2014). Inkubasi dilakukan selama tujuh hari dengan masing-masing
isolat tiga ulangan. Sampel fermentasi didestilasi menggunakan destilator
pada suhu ± 90 °C, kemudian destilat yang diperoleh diukur kadar
etanolnya menggunakan alkohol meter. Hasil skrining kemampuan
fermentasi etanol kemudian dianalisis menggunakan software SPSS
V16.0 for Windows dengan metode one-way ANOVA dan uji Tukey
dengan α=0,05.
3.3 Fermentasi Nira Sorgum
Dua isolat dengan toleransi etanol dan kemampuan menghasilkan
etanol dalam kadar tertinggi dilanjutkan untuk fermentasi sirup sorgum
dengan perlakuan kadar nitrogen yang berbeda. Fermentasi sirup sorgum
dilakukan melalui dua tahapan yaitu pre-fermentasi dan fermentasi.
Tahap pre-fermentasi dilakukan dengan menumbuhkan 10 % (v/v,
densitas 106 cfu/mL) khamir hasil skrining ke dalam media sirup sorgum
volume 250 mL dengan Brix 25 selama 12 jam untuk mengadaptasi isolat
khamir dan melewati fase lag pertumbuhan sel khamir. Khamir yang
diinokulasikan ke dalam medium pre-fermentasi sebelumnya
ditumbuhkan dalam media YME cair selama 48 jam dan disetarakan
densitas selnya. Hasil pre-fermentasi kemudian digunakan sebagai starter

15
fermentasi dengan diinokulasikan sebanyak 10% (v/v) ke dalam medium
fermentasi.
Fermentasi dilakukan dengan dua perlakuan penambahan kadar
nitrogen yaitu 0,2% dan 0,5%. Sumber nitrogen yang digunakan yaitu
Ammonium Sulfat ((NH4)2SO4). Brix medium diatur menjadi 25 dengan
cara melarutkan sirup sorgum dengan akuades dan diukur menggunakan
refraktometer. Khamir Saccharomyces cerevisiae digunakan sebagai
kontrol positif penghasil etanol menggunakan sirup sorgum. Kadar
etanol, gula reduksi dan jumlah sel dianalisis pada hari ke-1, 3, 5 dan 7
dengan masing-masing isolat tiga ulangan.
Sampel fermentasi sebanyak 80 mL didestilasi menggunakan
destilator dengan suhu ± 90 °C. Destilat yang didapatkan diukur kadar
etanolnya menggunakan alkohol meter dan dihitung kadar etanol dari
sampel menggunakan persamaan (2). Gula pereduksi diukur
menggunakan metode 3,5-Dinitrosalicylic (DNS) (Miller, 1959). Reagen
DNS dibuat dengan melarutkan (w/v) 1% DNS, 0,05% sodium sulfat, 1%
NaOH dan Potassium sodium tartrate (Garam Rochelle) 40 % secara
terpisah. Sebanyak tiga mL sampel fermentasi dimasukkan ke dalam
tabung reaksi dan ditambahkan reagen DNS tiga mL. Sampel kemudian
dipanaskan dalam waterbath suhu 100 °C selama 15 menit. Sampel
didinginkan pada suhu ruang dan ditambahkan Garam rochelle 40 %
sebanyak satu mL. Absorbansi sampel diukur menggunakan
spektrofotometer dengan λ 575 nm dan konsentrasi gula pereduksi
dibandingkan dengan kurva standar glukosa. Kurva standar dibuat
dengan menghubungkan konsentrasi glukosa (sumbu x) dan OD (sumbu
y).
M1V1 = M2V2 ......................................................................................... (2)
Viabilitas khamir dihitung dengan cara menumbuhkan sel khamir
pada media YMEA. Sampel fermentasi diambil sebanyak satu mL
kemudian dilakukan pengenceran berseri hingga 106 dan 0,1 mL dari hasil
pengenceran ditumbuhkan pada media YMEA dengan metode cawan
sebar. Cawan Petri kemudian diinkubasi pada suhu 30 °C selama 48 jam
dan dihitung jumlah koloni yang tumbuh. Hasil pengukuran kadar etanol,
gula reduksi dan jumlah sel khamir dianalisis menggunakan software
SPSS V16.0 for Windows dengan metode one-way ANOVA dan uji
Tukey dengan α=0,05.

16
3.4 Identifikasi Khamir Potensial Berdasarkan Internal Transcribed
Spacer (ITS)
Dua isolat dengan kemampuan fermentasi tinggi dipilih untuk
identifikasi yaitu isolat J1 dan J3. Isolat ditumbuhkan pada media YME
cair pada suhu 30 °C selama 48 jam. Kultur isolat kemudian ditumbuhkan
pada media YMEA dengan metode cawan sebar dan diinkubasi pada
suhu 30 °C selama 48 jam. Koloni yang tumbuh diambil untuk ekstraksi
DNA. DNA isolat khamir diekstraksi menggunakan I-Genomic Soil DNA
Extraction Mini Kit INTRON sesuai dengan prosedur yang ditetapkan.
DNA hasil ekstraksi diuji kuantitatif untuk mengetahui konsentrasi DNA
dengan menggunakan Nano Drop. Uji kualitatif DNA dilakukan
menggunakan elektroforesis (Fatchiyah dkk., 2011).
DNA hasil ekstraksi diamplifikasi menggunakan Polymerase Chain
Reaction (PCR) dengan primer ITS4 (5’-
TCCTCCGCTTATTGATATGC-3’) dan ITS 5 (5’-
GGAAGTAAAAGTCGTAACAAGG-3’). Komposisi Master mix untuk
PCR (Tabel 3) dihomogenasi dan dimasukkan ke dalam mesin PCR
dengan kondisi reaksi ditunjukkan pada tabel 5 (Hesham dkk., 2017).
Tabel 3. Komposisi master mix untuk amplifikasi PCR
Komponen Volume (uL) Konsentrasi
Nuclease free water 19 -
PCR mix 25 -
Primer 1 f (ITS 4) 2 10 pmol/uL
Primer 2 f (ITS 5) 2 10 pmol/uL
DNA Template
2
J1 13,8 ng/uL
J3 13,4 ng/uL
Total 50 uL -
Hasil amplifikasi DNA divisualisai dengan elektroforesis
menggunakan agarosa 1,5 % pH 8. Pewarna Ethidium Bromide (EtBr)
ditambahkan ke dalam agarosa sebanyak satu µL dan dihomogenkan.
Campuran agarosa dituang ke dalam cetakan gel hingga memadat.
Cetakan gel dimasukkan ke dalam chamber elektroforesis dan diisi
dengan buffer TBE hingga cetakan terendam. Marker DNA TriDyeTM 1

17
kb DNA Ladder dan sampel DNA sebanyak 2 µL dimasukkan ke dalam
sumuran gel. Sampel DNA dielektroforesis pada tegangan 100 V selama
30 – 45 menit. Hasil elektroforesis didokumentasikan menggunakan Gel-
Doc UV transiluminator (Fatchiyah dkk., 2011). Hasil PCR dianalisis
menggunakan sequencing analysis di 1st Base, Malaysia. Sekuen hasil
sequencing digabungkan menggunakan program BioEdit Sequence
Alignment Editor ver. 7.2.5 dan disejajarkan menggunakan ClustalW.
Sekuen homolog dicari menggunakan database National Center for
Biotechnology Information (NCBI) menggunakan BLASTN (Lee dkk.,
2011). Hasil perbandingan dari BLAST dibentuk pohon filogeni
menggunakan program MEGA 6 dengan algoritma Neighboor-Joining
dan estimasi jarak menggunakan Jukes-Cantor dengan analisis bootstrap
1.000 pengulangan (Hesham dkk., 2017).
Tabel 4. Kondisi siklus amplifikasi PCR
Langkah Suhu (°C) Waktu (detik)
Denaturasi awal 95 300
40 siklus:
Denaturasi
94
60
Annealing 55 60
Ekstensi 72 60
Ekstensi akhir 72 600
(Hesham dkk., 2017)

18
BAB IV
HASIL DAN PEMBAHASAN
4.1 Jumlah Sel Khamir yang Diisolasi dan Skrining Khamir dari
Batang dan Biji Sorgum
Dari biji sorgum didapatkan jumlah sel sebanyak 4,6 x 105 cfu/g
sedangkan dari batang sorgum sebanyak 3,8 x 107 cfu/g. Jumlah total
isolat yang didapatkan deapan isolat, terdapat empat isolat yang berasal
dari batang sorgum (T1, T2, T4 dan T5) dan empat isolat dari biji sorgum
(J1, J2, J3 dan J4). Isolat dipilih berdasarkan karakter morfologi koloni
yang berbeda. Delapan isolat tersebut kemudian diamati morfologi selnya
menggunakan mikroskop perbesaran 1000 X dengan pewarnaan
Methylen Blue 0,02 % (Gambar 5).
a. b. c. d.
e. f. g. h.
Gambar 5. Morfologi sel khamir hasil isolasi, a.) isolat T1, b.) isolat T2,
c.) isolat T4, d.) isolat T5, e.) isolat J1, f.) isolat J2, g.) isolat J3,
h.) isolat J4 (M 1000 X, ─ = 10 µm)
Isolat khamir yang telah murni diuji toleransinya pada kadar etanol
tinggi. Pertumbuhan sel khamir tersebut diketahui melalui nilai
absorbansi yang diukur mengunakan spektrofotometer dengan panjang
gelombang 600 nm. Semua isolat diketahui mampu bertahan pada
konsentrasi etanol 5% dan 10 % pada medium dan hanya isolat J2 dan J3
yang mampu tumbuh pada konsentrasi etanol 15 % (Gambar 6). Khamir

19
yang mampu tumbuh pada kadar etanol tinggi menunjukkan toleransinya
terhadap etanol yang merupakan salah satu ciri spesifik khamir yang
dapat menghasilkan etanol melalui fermentasi. Khamir yang dapat
melakukan fermentasi etanol dan menghasilkan etanol dalam jumlah
tinggi umumnya dapat bertahan pada medium dengan konsentrasi etanol
antara 5 – 15 % ketika ditumbuhkan pada medium cair (Lee dkk., 2011;
Tikka dkk., 2013). Kadar etanol yang tinggi dalam medium secara umum
berpengaruh terhadap penyerapan glukosa, maltosa, amonium dan asam
amino menyebabkan sel mengalami lisis kehilangan nukleotida, asam
amino dan potasium. Khamir yang memfermentasi karbohidrat menjadi
etanol memiliki toleransi terhadap kadar etanol tinggi disebabkan adanya
beberapa peningkatan regulasi gen yang berada pada membran sel khamir
sehingga sel khamir mampu toleran terhadap etanol. Peningkatan regulasi
gen yang berhubungan dengan struktur dinding sel yang berhubungan
dengan toleransi terhadap etanol meliputi gen TIP1 untuk metabolisme
mannoprotein, SED1 untuk metabolisme glikoprotein, SPI1 untuk
resistensi terhadap asam, dan HSP150 untuk organisasi dinding sel (Ma
& Liu, 2010).
Gambar 6. Toleransi isolat terhadap kadar etanol
Kadar etanol
0%
5%
10%
15%
20
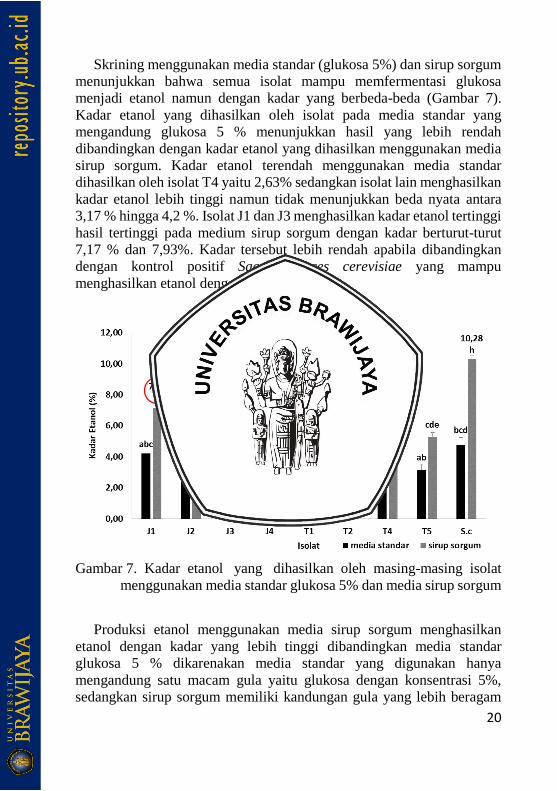
Skrining menggunakan media standar (glukosa 5%) dan sirup sorgum
menunjukkan bahwa semua isolat mampu memfermentasi glukosa
menjadi etanol namun dengan kadar yang berbeda-beda (Gambar 7).
Kadar etanol yang dihasilkan oleh isolat pada media standar yang
mengandung glukosa 5 % menunjukkan hasil yang lebih rendah
dibandingkan dengan kadar etanol yang dihasilkan menggunakan media
sirup sorgum. Kadar etanol terendah menggunakan media standar
dihasilkan oleh isolat T4 yaitu 2,63% sedangkan isolat lain menghasilkan
kadar etanol lebih tinggi namun tidak menunjukkan beda nyata antara
3,17 % hingga 4,2 %. Isolat J1 dan J3 menghasilkan kadar etanol tertinggi
hasil tertinggi pada medium sirup sorgum dengan kadar berturut-turut
7,17 % dan 7,93%. Kadar tersebut lebih rendah apabila dibandingkan
dengan kontrol positif Saccharomyces cerevisiae yang mampu
menghasilkan etanol dengan kadar 10,28 %.
Gambar 7. Kadar etanol yang dihasilkan oleh masing-masing isolat
menggunakan media standar glukosa 5% dan media sirup sorgum
Produksi etanol menggunakan media sirup sorgum menghasilkan
etanol dengan kadar yang lebih tinggi dibandingkan media standar
glukosa 5 % dikarenakan media standar yang digunakan hanya
mengandung satu macam gula yaitu glukosa dengan konsentrasi 5%,
sedangkan sirup sorgum memiliki kandungan gula yang lebih beragam

21
yaitu sukrosa, glukosa, dan fruktosa dengan konsentrasi berturut-turut 4,2
%, 2,3 % dan 2,8 % (Luo dkk., 2014). Kandungan gula yang lebih
beragam menyediakan sumber karbon yang lebih banyak untuk
melangsungkan proses glikolisis dan fermentasi etanol. Kandungan gula
yang beragam serta kandungan amilum yang tinggi menyediakan karbon
yang cukup banyak sehingga dapat dimanfaatkan oleh sel khamir untuk
pembentukan ATP dan menghasilkan etanol (Sirappa, 2003).
Isolat J1 dan J3 digunakan untuk optimasi fermentasi sirup sorgum
dengan kadar nitrogen yang berbeda karena kedua isolat tersebut
menunjukkan toleransi yang tinggi terhadap etanol dan mampu
memproduksi etanol dari substrat sirup sorgum dalam jumlah yang tinggi
dibandingkan dengan isolat yang lain.
4.2 Produksi Etanol dari Fermentasi Sirup Sorgum
Fermentasi sirup sorgum oleh isolat J1 dan J3 dengan penambahan
kadar nitrogen berupa (NH4)2SO4 0,2 % dan 0,5 % ditunjukkan pada
gambar 8. Penambahan kadar nitrogen 0,5 % menghasilkan kadar etanol
yang lebih tinggi dibandingkan penambahan kadar nitrogen 0,2 %. Isolat
J1 dengan waktu inkubasi tujuh hari menghasilkan etanol dengan kadar
8,67 % pada penambahan 0,2 % (NH4)2SO4 sedangkan penambahan 0,5
% menghasilkan etanol dengan kadar 14,05 %. Isolat J3 menghasilkan
etanol dengan kadar 8,33 % pada penambahan (NH4)2SO4 0,2 % dan
10,80 % pada penambahan (NH4)2SO4 dengan kadar 0,5 %. Isolat kontrol
Saccharomyces cerevisiae menghasilkan etanol lebih tinggi
dibandingkan isolat J1 dan J3 dengan masing-masing kadar 20,83 % dan
22,10 % dengan penambahan (NH4)2SO4 0,2 % dan 0,5 %. Kedua isolat
J1 dan J3 mampu menghasilkan etanol, namun peningkatan kadar yang
dihasilkan tidak begitu tinggi apabila dibandingkan dengan S. cerevisiae
yang menunjukkan peningkatan kadar dari hari ke-1 menuju hari ke-3.
Kadar etanol yang dihasilkan S. cerevisiae pada hari ke-3 hingga hari ke-
7 tidak menunjukkan perbedaan kadar yang signifikan.
Perbedaan kadar etanol yang dihasilkan dengan penambahan nitrogen
yang berbeda menunjukkan pengaruh terhadap produksi etanol.
Peningkatan kadar nitrogen di dalam medium akan meningkatkan laju
transkripsi dan pembelahan sel sehingga laju pertumbuhan sel khamir
lebih cepat dengan penambahan kadar nitrogen 0,5%. Jumlah sel khamir
yang lebih tinggi berpengaruh pada proses glikolisis yang terjadi
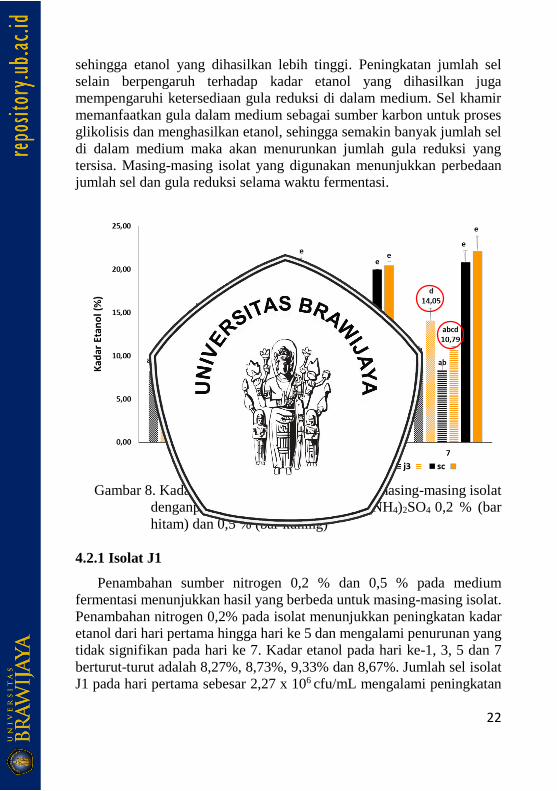
22
sehingga etanol yang dihasilkan lebih tinggi. Peningkatan jumlah sel
selain berpengaruh terhadap kadar etanol yang dihasilkan juga
mempengaruhi ketersediaan gula reduksi di dalam medium. Sel khamir
memanfaatkan gula dalam medium sebagai sumber karbon untuk proses
glikolisis dan menghasilkan etanol, sehingga semakin banyak jumlah sel
di dalam medium maka akan menurunkan jumlah gula reduksi yang
tersisa. Masing-masing isolat yang digunakan menunjukkan perbedaan
jumlah sel dan gula reduksi selama waktu fermentasi.
Gambar 8. Kadar etanol yang dihasilkan oleh masing-masing isolat
denganpenambahan kadar nitrogen (NH4)2SO4 0,2 % (bar
hitam) dan 0,5 % (bar kuning)
4.2.1 Isolat J1
Penambahan sumber nitrogen 0,2 % dan 0,5 % pada medium
fermentasi menunjukkan hasil yang berbeda untuk masing-masing isolat.
Penambahan nitrogen 0,2% pada isolat menunjukkan peningkatan kadar
etanol dari hari pertama hingga hari ke 5 dan mengalami penurunan yang
tidak signifikan pada hari ke 7. Kadar etanol pada hari ke-1, 3, 5 dan 7
berturut-turut adalah 8,27%, 8,73%, 9,33% dan 8,67%. Jumlah sel isolat
J1 pada hari pertama sebesar 2,27 x 106 cfu/mL mengalami peningkatan

23
pada hari ke-3 menjadi 1,05 x 107 cfu/mL. Jumlah sel pada hari ke-5
menunjukkan jumlah yang tidak berbeda dengan hari ke-3 yaitu sebesar
1,03 x 107 cfu/mL, dan mengalami penurunan pada hari ke-7 menjadi
7,57 x 106 cfu/mL. Kadar gula reduksi pada hari pertama sebesar 45,96
g/L mengalami peningkatan pada hari ke-3 menjadi 88,5 g/L. Selanjutnya
kadar gula reduksi menurun pada hari ke-5 menjadi 38,47 g/L dan pada
hari ke-7 menjadi 23,44 g/L (Gambar 9).
Penambahan nitrogen 0,5% pada media fermentasi menunjukkan
peningkatan kadar etanol dari hari ke-1, 3, 5 dan 7 berturut-turut 9,79%,
10,43%, 12,87% dan 14,05%. Peningkatan kadar etanol berbanding
terbalik dengan gula reduksi yang semakin menurun setiap harinya yaitu
berturut-turut 45,35 g/L, 44,66 g/L, 23,55 g/L dan 14,88 g/L. Jumlah sel
isolat J1 pada hari ke-1 sejumlah 2,27 x 106 cfu/mL dan mengalami
peningkatan pada hari ke-3 menjadi 1,6 x 107 cfu/mL. Jumlah sel pada
hari ke-5 dan 7 mengalami penurunan yaitu berturut-turut sebesar 1,4 x
107 cfu/mL dan 9,7 x 106 cfu/mL.
a.
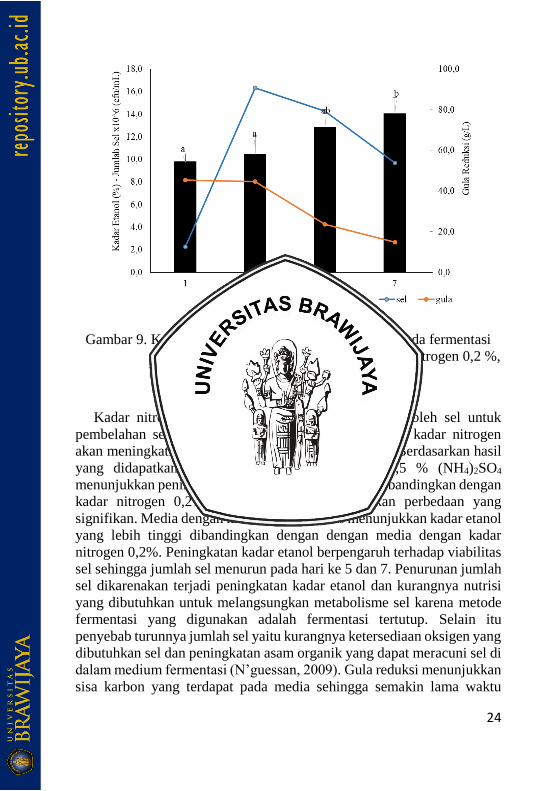
24
b.
Gambar 9. Kadar etanol, jumlah sel dan gula reduksi pada fermentasi
sirup sorgum oleh isolat J1, a.) penambahan nitrogen 0,2 %,
b.) 0,5%
Kadar nitrogen pada media fermentasi dibutuhkan oleh sel untuk
pembelahan sel (Broach, 2012), sehingga peningkatan kadar nitrogen
akan meningkatkan laju pertumbuhan dari sel khamir. Berdasarkan hasil
yang didapatkan, jumlah sel pada penambahan 0,5 % (NH4)2SO4
menunjukkan peningkatan jumlah yang lebih tinggi dibandingkan dengan
kadar nitrogen 0,2% meskipun tidak menunjukkan perbedaan yang
signifikan. Media dengan kadar nitrogen 0,5% menunjukkan kadar etanol
yang lebih tinggi dibandingkan dengan dengan media dengan kadar
nitrogen 0,2%. Peningkatan kadar etanol berpengaruh terhadap viabilitas
sel sehingga jumlah sel menurun pada hari ke 5 dan 7. Penurunan jumlah
sel dikarenakan terjadi peningkatan kadar etanol dan kurangnya nutrisi
yang dibutuhkan untuk melangsungkan metabolisme sel karena metode
fermentasi yang digunakan adalah fermentasi tertutup. Selain itu
penyebab turunnya jumlah sel yaitu kurangnya ketersediaan oksigen yang
dibutuhkan sel dan peningkatan asam organik yang dapat meracuni sel di
dalam medium fermentasi (N’guessan, 2009). Gula reduksi menunjukkan
sisa karbon yang terdapat pada media sehingga semakin lama waktu
25
fermentasi, jumlah gula reduksi akan semakin menurun. Beberapa isolat
khamir diketahui menghasilkan α-amylase dan glukoamilase yang dapat
menghidrolisis amilum atau pati menjadi gula sederhana (Jamai dk.,
2006). Peningkatan gula reduksi pada penambahan kadar (NH4)2SO4
0,2% dapat terjadi akibat hidrolisis pati di dalam medium menjadi gula
sederhana, sehingga meningkatkan kadar gula reduksi pada hari ke-3 dan
kembali menurun karena digunakan untuk metabolisme sel pada hari ke-
5 dan 7.
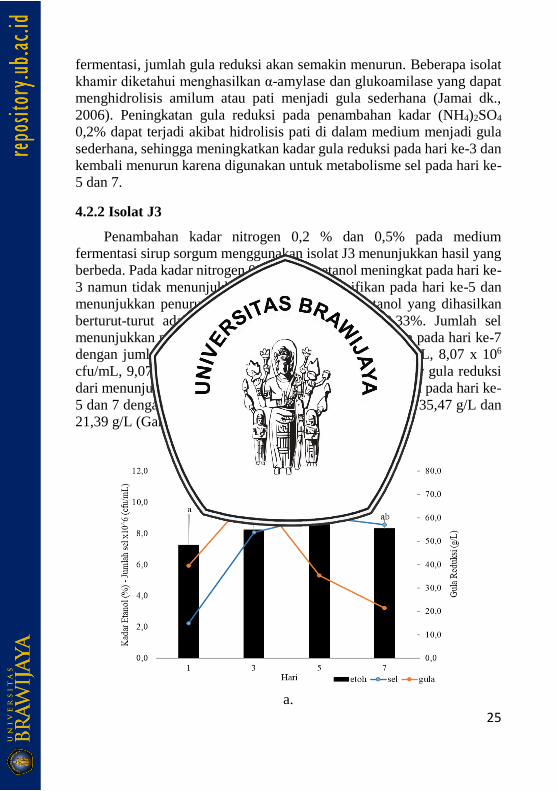
4.2.2 Isolat J3
Penambahan kadar nitrogen 0,2 % dan 0,5% pada medium
fermentasi sirup sorgum menggunakan isolat J3 menunjukkan hasil yang
berbeda. Pada kadar nitrogen 0,2%, kadar etanol meningkat pada hari ke-
3 namun tidak menunjukkan peningkatan signifikan pada hari ke-5 dan
menunjukkan penurunan pada hari ke-7. Kadar etanol yang dihasilkan
berturut-turut adalah 7,27%, 8,23%, 8,67% dan 8,33%. Jumlah sel
menunjukkan peningkatan hingga hari ke-5 dan menurun pada hari ke-7
dengan jumlah sel berturut-turut adalah 2,23 x 106 cfu/mL, 8,07 x 106
cfu/mL, 9,07 x 106 cfu/mL, dan 8,57 x 106 cfu/mL. Kadar gula reduksi
dari menunjukkan peningkatan pada hari ke 3 dan menurun pada hari ke-
5 dan 7 dengan kadar berturut-turut 39,49 g/L, 71,25 g/L, 35,47 g/L dan
21,39 g/L (Gambar 10).
a.

26
b.
Gambar 10. Kadar etanol, jumlah sel dan gula reduksi pada fermentasi
sirup sorgum oleh isolat J3, a.) penambahan nitrogen 0,2 %,
b.) 0,5%
Sedangkan pada penambahan nitrogen 0,5%, kadar etanol meningkat
dengan kadar berturut-turut 8,33%, 9,25%, 10,38% dan 10,79%. Jumlah
sel juga menunjukkan peningkatan hingga hari ke-7 yaitu berturut-turut
2,46 x 106 cfu/mL, 9,46 x 106 cfu/mL, 1,0 x 107 cfu/mL dan 1,13 x 107
cfu/mL. Kadar gula reduksi menunjukkan peningkatan pada hari ke-3 dan
menurun pada hari ke-5 dan 7 dengan nilai berturur-turut 42,66 g/L, 44,42
g/L, 22,82 g/L, 17,05 g/L.
Jumlah sel cenderung tetap pada kadar nitrogen 0,2% sedangkan pada
kadar (NH4)2SO4 0,5 % pada hari ke-7 jumlah sel meningkat
kemungkinan dikarenakan nutrisi di dalam medium masih mencukupi
untuk pertumbuhan isolat dan kadar etanol yang dihasilkan belum mampu
menghambat pertumbuhan dari isolat J3. Gula reduksi semakin menurun
pada hari ke-7 karena digunakan oleh sel untuk melangsungkan proses
metabolisme. Peningkatan kadar gula reduksi pada hari ke-3 dapat
dikarenakan adanya proses perombakan sumber karbon kompleks seperti
pati menjadi gula sederhana sehingga meningkatkan jumlah gula reduksi
dalam medium. Beberapa isolat khamir diketahui menghasilkan α-

27
amylase dan glukoamilase yang dapat menghidrolisis amilum atau pati
menjadi gula sederhana yang dibutuhkan oleh sel (Jamai dk., 2006).
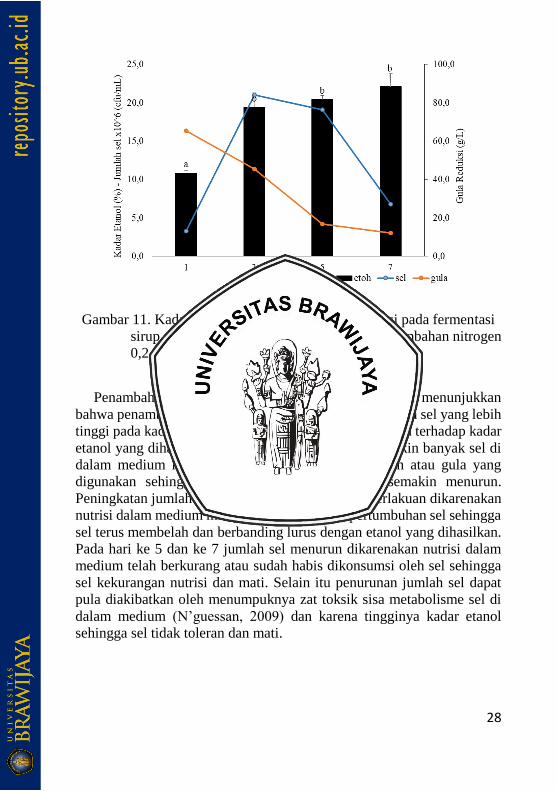
4.2.3 Saccharomyces cerevisiae
Penambahan kadar nitrogen 0,2 % dan 0,5 % pada medium fermentasi
menunjukkan hasil yang berbeda. Pada kadar nitrogen 0,2%, kadar etanol
meningkat pada hari ke 3 dan relatif konstan pada ke-5 dan 7 dengan
kadar berturut-turut 11,63%, 19,33%, 20% dan 20,83%. Kadar gula
reduksi menurun hingga hari ke 7 dengan nilai berturut-turut 93,65 g/L,
49,18 g/L, 34,38 g/L dan 17,16 g/L. Jumlah sel mengalami kenaikan pada
hari 3 dan mengalami penurunan pada hari ke 5 dan 7 dengan jumlah
berturut-turut 3,03 x 106 cfu/mL, 1,7 x 107 cfu/mL, 1,6 x 107 cfu/mL dan
5,6 x 106 cfu/mL. Sedangkan pada penambahan nitrogen 0,5%
menunjukkan hasil kadar etanol yang meningkat hingga hari ke-7 dengan
kadar berturut-turut 10,85%, 19,40%, 20,42% dan 22,1 %. Kadar gula
reduksi menurun hingga hari ke-7 dengan nilai berturut-turut 65,77 g/L,
45,37 g/L, 16,77 g/L dan 12,17 g/L. Jumlah sel mengalami kenaikan pada
hari ke-3 dari 3,27 x 106 cfu/mL menjadi 2,1 x 107 cfu/mL, sedangkan
pada hari ke 5 dan 7 mengalami penurunan menjadi 1,9 x 107 cfu/mL dan
6,77 x 106 cfu/mL.
a.
28
b.
Gambar 11. Kadar etanol, jumlah sel dan gula reduksi pada fermentasi
sirup sorgum oleh isolat S. cerevisiae, a.) penambahan nitrogen
0,2 %, b.) 0,5%
Penambahan kadar nitrogen dari kedua perlakuan menunjukkan
bahwa penambahan nitrogen berpengaruh terhadap jumlah sel yang lebih
tinggi pada kadar (NH4)2SO4 0,5 % sehingga berpengaruh terhadap kadar
etanol yang dihasilkan menjadi lebih tinggi pula. Semakin banyak sel di
dalam medium maka semakin banyak sumber karbon atau gula yang
digunakan sehingga semakin lama gula reduksi semakin menurun.
Peningkatan jumlah sel pada hari ke-3 dari kedua perlakuan dikarenakan
nutrisi dalam medium masih mencukupi untuk pertumbuhan sel sehingga
sel terus membelah dan berbanding lurus dengan etanol yang dihasilkan.
Pada hari ke 5 dan ke 7 jumlah sel menurun dikarenakan nutrisi dalam
medium telah berkurang atau sudah habis dikonsumsi oleh sel sehingga
sel kekurangan nutrisi dan mati. Selain itu penurunan jumlah sel dapat
pula diakibatkan oleh menumpuknya zat toksik sisa metabolisme sel di
dalam medium (N’guessan, 2009) dan karena tingginya kadar etanol
sehingga sel tidak toleran dan mati.

29
4.3 Hasil Identifikasi Isolat Potensial
Hasil identifikasi berdasarkan daerah ITS menunjukkan bahwa isolat
J1 teridentifiksai sebagai Candida tropicalis dengan similaritas 99% dan
isolat J3 teridentifikasi sebagai Wickerhamomyces anomalus dengan
similaritas 99%. Kedua isolat tersebut dalam pohon filogeni
menunjukkan berada pada cabang yang berbeda dengan isolat yang
teridentifikasi. Hal tersebut dikarenakan primer yang digunakan untuk
amplifikasi DNA adalah primer ITS 4 dan 5 sedangkan sekuen
pembanding yang digunakan merupakan sekuen dari daerah ITS 1 dan 2
yang secara umum diamplifikasi menggunakan primer ITS 1 dan ITS 4.
Primer ITS 1 sebagai forward primer terletak pada ujung 3’ pada daerah
Small Sub Unit (SSU) dan ITS4 sebagai primer referse pada bagian 5’
dari Large Sub Unit (LSU) sehingga daerah ITS yang terletak di antara
SSU dan LSU akan teramplifikasi secara utuh (gambar 12) (Martin &
Rygiewicz, 205). Sedangkan jika digunakan primer ITS4 dan ITS 5,
primer terletak lebih jauh dari daerah ITS dibandingkan primer ITS1 dan
ITS 4 sehingga daerah yang teramplifikasi lebih luas.
(Martin & Rygiewicz, 205)

30
Gambar 12. Diagram lokasi primer yang digunakan untuk amplifikasi
daerah ITS1-5,8S-ITS2
Hasil Blast menunjukkan bahwa isolat J1 memiliki similaritas degan
Candida tropicalis hingga 99% (Lampiran 1). Namun pada hasil pohon
filogeni isolat J1 berada diluar cabang kelompok Candida tropicalis dan
berada dalam satu cabang dengan kelompok Candida albicans.
Perbedaan hasil tersebut mengakibatkan isolat J1 belum diketahui secara
pasti spesienya. Berdasarkan sampel yang digunakan dimungkinkan
isolat J1 adalah Candida trpicalis yang memiliki habitat alami pada
permukaan buah-buahan, tanah dan sebagian pada sampel darah mamalia
yang terinfeksi Candida tropicalis. Sedangkan Candida albicans
sebagian besar diisolasi dari tubuh mamalia yang terjangkit Candidiasis
karena khamir ini merupakan khamir patogen.
Candida tropicalis memiliki koloni berwarna krem, berbentuk bulat
dengan tekstur halus, bentuk sel bulat hingga oval dengan ukuran sel
antara 2 – 10 µm (Shariq & Sohail, 2018). Meskipun beberapa strain
merupakan khamir patogen, C. tropicalis banyak digunakan untuk
fermentasi etanol dan xylitol. Candida tropicalis juga memiliki
kemampuan untuk memetabolisme senyawa fenolik dan mendegradasi
beberapa senyawa lignin yang tidak berpengaruh terhadap proses
fermentasi (Martin dkk., 2010). Candida tropicalis sering digunakan
untuk fermentasi etanol dari pati meskipun dalam laju yang lambat karena
C. tropicalis menghasilkan enzim glukoamilase yang dapat
menghidrolisis pati menjadi polisakarida yang lebih pendek dengan hasil
akhir berupa glukosa terlarut (Jamai dk., 2006). Sirup sorgum diketahui
mengandung amilum dengan jumlah 209 – 1.764 ppm (Sirappa, 2003)
sehingga dimungkinkan isolat J1 atau C. tropicalis menghidrolisis
amilum dalam medium sehingga kadar gula reduksi meningkat pada hari
ke-3.
Fermentasi sirup sorgum menggunakan C. tropicalis dibandingkan
dengan S. cerevisiae (N’guessan, 2009) menunjukkan bahwa pada 8 jam
awal, kadar etanol yang dihasilkan C. tropicalis lebih tinggi namun
setelah 12 jam fermentasi kadar etanol yang dihasilkan oleh S. cerevisiae
lebih tinggi. Dalam fermentasi sirup sorgum, C. tropicalis aktif pada awal
fermentasi dan semakin menurun pada akhir fermentasi. Penurunan
jumlah sel pada C. tropicalis diakibatkan oleh ketersediaan oksigen yang
menurun sehingga sel tidak dapat tumbuh. Selain itu Candida tropicalis
diketahui dapat tumbuh dengan baik pada medium dengan kadar etanol 6

31
– 8% (N’guessan, 2009; Shariq & Sohail, 2018), sehingga pada
konsentrasi etanol yang lebih tinggi jumlah sel semakin berkurang.
Gambar 13. Pohon filogeni dari isolat J1 yang teridentifikasi sebagai
Candida tropicalis
Isolat J3 yang teridentifikasi sebagai Wickerhamomyces anomalus
menunjukkan berada pada kelompok yang berbeda pada pohon filogeni.
Isolat J3 pada pohon filogeni menunjukkan kekerabatan yan dekat dengan
Wilia javanica dan berada diluar kelompok Wickerhamomyces meskipun
Wilia javanica merupakan nama sinonim dari Wickerhamomyces
anomalus. Wickerhamomyces anomalus atau dikenal sebagai Pichia
anomala merupakan organisme yang sering ditemukan pada proses
pembusukan makanan dan produk biji-bijian. Wickerhamomyces
anomalus memiliki kemampuan tumbuh pada berbagai jenis karbon, suhu
antara 3 – 37 °C, pH antara 2-12 dan dibawah tekanan osmotik dan
dengan sedikit atau tidak adanya oksigen (Passoth, dkk., 2006). Toleransi
yang tinggi terhadap kondisi lingkungan menyebabkan khamir ini dapat
diisolasi dari berbagai sumber seperti tumbuhan berbunga, permukaan
buah, produk susu dan roti, kontaminasi minyak, makanan asinan, air
Candida tropicalis CTR11
Candida tropicalis CTR145
Candida tropicalis CTR770
Candida tropicalis ZA029
Candida tropicalis SY5-3
Candida tropicalis ATCC 750
Candida tropicalis ATCC:66029
Candida tropicalis 1920
Candida tropicalis CBS 94T
Candida chaulioides CBS 10157T
Candida heliconiae CBS 10000T
Candida riodocensis CBS 10087T
Candida oslonensis CBS 10146T
Candida tolerans CBS 8613
Bj1
Candida albicans CBS 1893
Candida albicans CBS 2702T
Candida tropicalis CTR 11
Candida tropicalis CTR145
Candida tropicalis CTR770
Candida tropicalis ZA029
Candida tropicalis SY5-3
Candida tropicalis ATCC 750
Candida tropicalis ATCC 66029
Candida tropicalis 1920
Candida tropicalis CBS 94T
Candida chaulioides CBS 10157T
Candida heliconiae CBS 10000T
Candida riodocensis CBS 10087T
Candida oslonensis CBS 10146T
Candida tolerans CBS 8613
J1
Candida chaulioides CBS 10157T
Candida albicans CBS 1893
Candida albicans CBS 2702T

32
limbah, lingkungan laut, jaringan manusia dan dapat pula ditemukan
dalam saluran pencernaan serangga (lalat, kumbang dan nyamuk)
(Capelli dkk., 2014).
Khamir ini banyak digunakan dalam pembuatan produk makanan,
sebagai agen biopreservatif yang tersertifikasi oleh European Food Safety
Authority (EFSA) dengan kualifikasi status kemanan pada level 1 (QPS-
1). Aplikasi lain dari khamir ini yaitu pada bidang biokontrol, fermentasi
makanan, dan produksi therapeutic molecules yang digunakan dalam
pembuatan obat. Wickerhamomyces anomalus juga diketahui memiliki
kemampuan antimikroba dengan spektrum luas yang dapat melawan
berbagai jenis mikroorganisme meliputi khamir, kapang dan bakteri.
Beberapa mekanisme yang dilakukan dapat beupa kompetisi nutrisi,
produksi etil asetat dan dibunuh langsung menggunakan Killer Toxins
(KTs). KTs merupakan kelompok glikoprotein dengan berbagai berat
molekul yang aktivitasnya optimal dalam pH dan suhu yang luas (Capelli
dkk., 2014).
Gambar 14. Pohon filogeni isolat J3 yang teridentifikasi sebagai
Wickerhamomyces anomalus
Wickerhamomyces anomalus 09TJ067
Wickerhamomyces anomalus IWG-DR-j
Wickerhamomyces anomalus F17.12
Pichia anomala NRRL Y-336T
Wickerhamomyces anomalus CBS 5759T
Wickerhamomyces anomalus CBS 110
Wickerhamomyces anomalus CBS 5759
Wickerhamomyces anomalus UFLA CWFY47
Monilia javanica CBS 261T
Wickerhamomyces anomalus 183
Wickerhamomyces anomalus M297B
Wickerhamomyces anomalus 11HX213
Wickerhamomyces siamensis CBS 12570T
Wickerhamomyces edaphicus CBS 10408T
Willia javanica CBS 251T
J3
Wickerhamomyces anomalus 09TJ067
Wickerhamomyces anomalus IWG-DR-j
Wickerhamomyces anomalus F17.12
Pichia anomala NRRL Y-336T
Wickerhamomyces anomalus CBS 5759T
Wickerhamomyces anomalus CBS 110
Wickerhamomyces anomalus CBS 5759
Wickerhamomyces anomalus UFLA CWFY 47
Monilia javanica CBS 261T
Wickerhamomyces anomalus 183
Wickerhamomyces anomalus M297B
Wickerhamomyces anomalus 11HX213
Wickerhamomyces siamensis CBS 12570T
Wickerhamomyces edaphicus CBS 10408T
Wilia javanica CBS 251T
J3

33
Wickerhamomyces anomalus merupakan khamir non-Saccharomyces
yang sering digunakan untuk pembuatan wine. Pada pembuatan wine
menggunakan campuran isolat khamir, W. anomalus aktif di awal
fermentasi dan memulai pembusukan buah anggur ketika produksi asam
asetat dan etil asetat tinggi. Pada fermentasi monokultur menggunakan
W. anomalus diketahui bahwa populasi sel dapat mencapai 107 cfu/mL
selama proses fermentasi, sedangkan dibandingkan dengan S. cerevisiae
yang akan mengalami penurunan jumlah sel pada hari ke 3 (Lombard,
2016). Hal tersebut sesuai dengan hasil yang didapatkan pada penelitian
ini bahwa isolat J3 yang teridentifikasi sebagai W. anomalus
menghasilkan etanol yang lebih rendah tetapi menghasilkan aroma yang
lebih kuat dibandingkan dengan S. cerevisiae, jumlah sel isolat J3 tidak
mengalami penurunan signifikan setelah melewati 3 hari proses
fermentasi jika dibandingkan dengan S. cerevisiae yang mengalami
penurunan jumlah sel pada hari ke 5.

34
BAB V
KESIMPULAN DAN SARAN
5.1 Kesimpulan
Kesimpulan dari penelitian ini adalah:
1. Delapan isolat khamir diperoleh dari sampel batang dan biji
sorgum, isolat J1 dapat tumbuh dalam medium dengan kadar
etanol 10 % sedangkan isolat J3 mampu tumbuh dengan kadar
medium hingga 15%. Isolat J1 dan J3 merupakan isolat unggul
yang mampu memfermentasi sirup sorgum dan menghasilkan
etanol dengan kadar yang tinggi.
2. Penambahan kadar nitrogen 0,5% pada medium sirup sorgum
menunjukkan produksi etanol yang lebih tinggi daripada
penambahan kadar nitrogen 0,2 %. Pada penambahan kadar
nitrogen 0,5 %, isolat J1 mampu menghasilkan etanol dengan
kadar 14,05% sedangkan isolat J3 mampu menghasilkan etanol
dengan kadar 10,73%.
3. Isolat J1 berdasarkan hasil Blast teridentifikasi sebagai Candida
tropicalis dengan similaritas 99% dan isolat J3 teridentifikasi
sebagai Wickerhamomyces anomalus dengan similaritas 99%.
5.2 Saran
Disarankan untuk mengamplifikasi DNA khamir menggunakan
primer universal untuk khamir yaitu ITS 1 dan ITS 4 sehingga sekuen
hasil amplifikasi sesuai dengan sekuen pembanding dari database sekuen
khamir untuk pembuatan pohon filogeni. Disarankan pula untuk
meningkatkan jumlah karbon untuk produksi etanol menggunakan isolat
J1 sehingga diketahui rasio C/N yang optimal untuk produksi etanol
menggunakan sirup sorgum dan menguji fermentasi sirup sorgum
menggunakan campuran isolat J1, J3 dan Saccharomyces cerevisiae.

35
DAFTAR PUSTAKA
Ali, M.N. & M.M. Khan. 2014. Screening, identification and
characterization of alcohol tolerant potential bioethanol producing
yeast. Current Research in Microbiology and Biotechnology,
2(1):316-324 Almodares, A. & M.R. Hadi. 2009. Production of bioethanol from sweet
sorghum: A review. African Journal of Agricultural Research,
4(9): 772-780. Azhar, S.H.M., R. Abdulla, S.A. Jambo, H. Marbawi, J.A. Gansau,
A.A.M. Faik, K.F. Rodrigues. 2017. Yeast in sustainable
bioethanol production: A review. Biochemistry and Biophysics
Reports, 10(2017): 52-61. Barnett, J.A. 2003. A history of research on yeast 5: the fermentation
pathway. Yeast, 20: 509-543.
Bellemain, E., T. Carlsen, C. Brochman, E. Coissac, P. Taberlet, H.
Kauserud. 2010. ITS as an environmental DNA barcode for fungi:
an in silico approach reveals potential PCR biases. BMC
Microbiology, 10:189. Broach, J.R. 2012. Nutritional control of growth and development in
yeast. Genetics, 192:73-105. Calderone, R.A. & G. San-Blas. 2008. Pathogenic fungi: Insight in
molecular biology. Expert Rev. Anti Infect. Ther, 6(5):591-592. Capelli, A., U. Ulissi, M. Valzano, C. Damiani, S. Epis, M.G. Gabrielli,
S. Conti, L. Polonelli, C. Bandi, G. Favia, I. Ricci. 2014. A
Wickerhamomyces anomalus killer strain in the malaria vector
Anopheles stephensi. PloS ONE 9(5): e95988. Cheng, N.G., M. Hasan, A.C. Kumoro, C.F. Ling, M. Tham. 2009.
Production of ethanol by fed-batch fermentation. Pertanika
Journal Science & Technology, 17(2):399-408. Cruz, S.H., E.M. Cilli, J.R. Ernandes. 2002. Structural complexity of the
nitrogen source and influence on yeast growth and fermentation. J.
Inst. Brew., 108(1):54-61. Das, S. & B. Deb. 2015. DNA barcoding of fungi using Ribosomal ITS
Marker for genetic diversity analysis: A Review. Int. J. Pure App.
Biosci., 3(3):160-167.

36
Ebabhi, A.M., A.A. Adekunle, W.O. Okunowo, A.A. Osuntoki. 2013.
Isolation and characterization of yeast strain from local food crops.
Journal of Yeast and Fungal Research, 4(4):38-43. Fatchiyah, E.L. Arumingtyas, S. Widiarti, S. Rahayu. 2011. Biologi
molekuler: prinsip dasar analisis. Erlangga. Jakarta.
Hesham, A.E, E. A. Mohamed, A.M.M. Mawad, A. Elfarash, B.S.A. El-
Fattah, M.A. El-Rawy. 2017. Molecular characterization of
fusarium solani degrades a mixture of low and high molecular
weight polycyclic aromatic hydrocarbons. The Open
Biotechnology Journal, 2017(11):27-35. Iticha, T.N. 2016. Isolation and screening of ethanol tolerant yeast for
bio-ethanol production in ethiopia. Global Journal of Life Sciences
and Biological Research, 2(2):1-7. Jamai, L., K. Ettayebi, J.E. Yamani, M. Ettayebi. 2006. Production of
ethanol from starch by free and immobilized Candida tropicalis in
the presence of α-amylase. Bioresource Technology 98(2007):
2765-2770. Kang, A. & T.S. Lee. 2015. Converting sugars to biofuels: Ethanol and
beyond. Bioengineering, 2:184-203; doi:
10.3390/bioengineering2040184.
Korabecna, M. 2007. The variability in the fungal ribosomal DNA (ITS1,
ITS2, and 5,8 S rRNA Gene): its biological meaning and
application in medical mycology. Formatex, A. Mendez-Vilas Ed.
783-787. Laopaiboon, L., S. Nuanpeng, P. Srinophakun, P. Klanrit, P. Laopaiboon.
2009. Ethanol production from sweet sorghum juice using very
high gravity technology: Effects of carbon and nitrogen
supplementations. Bioresource Technology, 100(2009):4176-
4182. Lee, Y.J., Y.R. Choi, S.Y. Lee, J.T. Park, J.H. Shim, K.H. Park, J.W. Kim.
2011. Screening wild yeast strains for alcohol fermentation from
various fruits. Mycobiology, 39(1):33-39.
Li, Z., D. Wang, Y. Shi. 2017. Effect of nitrogen source on ethanol
production in very high gravity fermentation of corn starch.
Journal of the Taiwan Institute of Chemical Engineers,
70(2017):229-235. Lombard, J. 2016. Characterisation of Wickerhamomyces anomalus and
Kazachstania aerobia: Investigating fermentation kinetics and

37
aroma production. Institute for Wine Biotechnology, Faculty of
AgriSciences, Stellenbosch University. Luo, Z., L. Wang, A. Shahbazi. 2014. Optimization of ethanol production
from sweet sorghum (Sorghum bicolor) juice using response
surface methodology. Biomass and Bioenergy, 67(2014):53-59.
Ma, M. & Z.L. Liu. 2010. Mechanisms of ethanol tolerance in
Saccharomyces cerevisiae. Appl. Microbiol Biotechnol (2010)87:
829-845.
Martin, K.J. & P.T. Rygiewicz. 2005. Fungal-spesific PCR primers
developed for analysis of the ITS region of environmental DNA
extracts. BMC Microbiology 5:28. Martin, J.F.G., M. Cuevas, V. Bravo, S. Sanchez. 2010. Ethanol
production from olive prunings by autohydrolysis and fermentation
with Candida tropicalis. Renewable Energy 35(2010): 1602-1608. Miller, G.L. 1959. Use of dinitrosalicylic acid reagent for determination
of reducing sugar. Analytical Chemistry, 3(31):426-428. N’guessan, F.K., D.Y. N’Dri, F. Camara, M.K. Dje. 2009.
Saccharomyces cerevisiae and Candida tropicalis as starter
cultures for the alcoholic fermentation of tchapalo, a traditional
sorghum beer. World Journal Microbiol Biotechnol 26(2010):693-
699. Naser, A. 2014. Isolation and characterization of yeast for bioethanol
production, using sugarcane molasses. Brac University. Dhaka.
Disertation. Nasreen, Z., S. Jabeen, M. Shafique, S. Usman, T. Yaseen, A. Yasmeen,
S. Nazir. 2014. Production of alcohol by yeast isolated from apple,
orange and banana. International Journal of Food and Nutrition
Sciences, 1(2):16-19. Passoth, V., E. Fredlund, U.A. Druvefors, J. Schnurer. 2006.
Biotechnology, physiology and genetics of the yeast Pichia
anomala. FEMS Yeast Research 6: 3-13. Phutela, U.G. & J. Kaur. 2014. Process optimization for ethanol
production from sweet sorghum juice using Saccharomyces
cerevisiae strain NRRL Y-2034 by response surface methodology.
Sugar Tech, 16(4):411-421. Rao, R.S., B. Bhadra, S. Shivaji. 2008. Isolation and characterization of
ethanol-producing yeast from fruits and tree barks. Letters in
Applied Microbiology, 47:19-24.

38
Rutto, L.K., Y. Xu, M. Brandt, S. Ren, M.K. Kering. 2013. Juice, ethanol,
and grain yield potential of five sweet sorghum Sorghum bicolor
[L.] Moench) Cultivars. Journal of Sustainable Bioenergy Systems,
3:113-118.
Scoch, C.L., K.A. Seifert, S. Huhndorf, V. Robert, J.L. Spouge, C.A.
Levesque, W. Chen. 2012. Nuclear ribosomal internal transcribed
spacer (ITS) region as a universal DNA barcode marker for fungi.
PNAS, 109(16):6241-6246. Shariq, M. & M. Sohail. 2018. Application of Candida tropicalis MK-
160 for the production of xylanase and ethanol. Journal of King
Saud University-Science 2018.04.009. Sirappa, M.P. 2003. Prospek pengembangan sorgum di indonesia sebagai
komoditas alternatif untuk pangan, pakan dan industri. Jurnal
Litbang Pertanian, 22(4): 133-140.
Strobel, R.J & G.R. Sullivan. 1990. Experimental design for
improvement of fermentation. ASM Press. Washingon. Subagio H. & M. Aqil. 2013. Pengembangan produksi sorgum di
indonesia. Seminar Nasional Inovasi Teknologi Pertanian, 2013.
Tikka, C., H.P. Osuru, N. Atluri, P.C.V. Raghavulu, N. Kumaryellapu,
I.S. Mannur, U.V. Prasad, S. Aluru, N. Varma, M. Bhaskar. 2013.
Isolation and characterization of ethanol tolerant yeast strains.
Bioinformation 9(8): 421-425.
Waites, M.J., N.L. Morgan, J.S. Rockey & G. Higton. 2005. Industrial
microbiology, an introduction. Blackwell Publishing. London. Widayanti, N.P., W.S. Rita, Y. Ciawi. 2013. Pengaruh konsentrasi
ammonium sulfat ((NH4)2SO4) sebagai sumber nitrogen terhadap
produksi bioetanol berbahan baku Glacilaria sp. Jurnal Kimia,
7(1):1-10.
Wiratno, E.N., T. Ardyati, A.K. Wardani. 2014. Effect of reducing sugar
and total nitrogen to ethanol production from molasses by
Saccharomyces cerevisiae. J. Exp. Life Sci., 4(2):50-55.