novel type of interstitial cell (cajal-like) in human fallopian tube
TRANSCRIPT

JqM,lU ffiiliiil" J. Cell. Mol. Med. Vol 9, No 2, 2005 pp. 479-523
Special Anicle
Novel type of interstitial cell (Cajal-Iike)
in human fallopian tube
L.M. Popescu&c*o Sanda M. Cionteaa, D. Cretoiuqc, M.E. Hinescu&., E. RaduccoN. Ionescu ao M. Ceausu c, Mihaela Gherghiceanu c,
R.I. Bragab, Florina Vasilescuc, L. Zagretnb, Carmen Ardeleanuc
a Department of Cellular and Molecular Medicine,b Department of Plrysiolog,
"Carol Davila" University of Medicine and Pharmacy, Bucharest, Rotnaniac "Wctor Babes" National Institute of Patholog, Bucharest, Romania
Received: March 21,2005; Accepted: trIay 6, 2005
AbstractWe describe here - presumably for the first time - a Cajal-like tlpe of tubal interstitial cells (I-ICC), resembling thearchetypal enteric ICC. t-ICC were demonstrated in situ and in vitro onfresh preparatiozs (tissue cryosections and pri-mary cell cultwes) using methylene-blue, crystal-violeg Janus-Green B or MitoTracker-Green FM Probe vital stainings.Also, I-ICC were identified n faed specimens by light microscopy (methylene-blue, Giemsa, trichrome stainings,Gomori silver-impregnation) or fansmission electrcn rnicrcscopy (TEM). The positive diagnosis of I-ICC was strength-enedby immunohistochemistry (HC; CDll7/c-kit+ andother 14 antigens) and immunofluorescence (IF; CDllT/c-kit+and other 7 antigens). The spatial densrty of I-ICC (ampullar-segment cryosections) was l0G-150 cells/mm2. Non-con-ventional light microscopy (NCLM) of Epon semithin-sections revealed a network-like distribution of t-ICC in laminapropria and smooth muscle meshwort. t-ICC appeared located beneath of epithelium, in a l0-15pm thick 'belt', wherelStZyo of cells were I-ICC. In the whole lamina propria, I-ICC were about 9Yo, and in muscularis 4o/o. In toto,I-ICCrepresent -8% of subepithelial cells, as counted by NCLM. In vitro, t-ICC were 9.9!0.9% of total cell population.
TEM showed that the diagnostic ' gold standardl (Huizinga et al., 1997) is fulfilled by 'our' t-ICC. However, weslggest a'platinum standart, adding a new defining criterion - characteristic cytoplasmic processes (number:l-5; length: tens of pm; thickness: S0.5pm; aspect: moniliform; branching: dichotomous; organization: network,labyrinthic-system). Quantitatively, the ultrastructural architecture of t-ICC is: nucleus,23.5!3.2% of cell volume,with heterochromatin 49.1+3.8yo; mitochondria,4.S!1.7%o; rough and smooth endoplasmic-reticulum (l.1110.6%0,1.0!0.2yo, respectively); caveolae,3.4!0.5o/o. We found more caveolae on the surface of cell processes versus cellbody, as confirmed by IF for caveolins. Occasionally, the so-called 'Ca2+-release units'(subplasmalemmal closeassociations of caveolasl-endoplasmic reticulunrtmitochondria) were detected in the dilations of cell processes.Electrophysiological single unit recordings of I-ICC in primary cultures indicated sustained spontaneous electricalactivity (amplitude of field potentials: 57.26!6.56mY).
Besides the CDllT/c-kit marker, t-ICC expressed variously CD34, caveolins 1&2, c,-SMA, 3-100, vimentin,nestin, desmin, NK-l. t-ICC were negative for: CD68, CDla, CD62R NSE, GFAR chromogranin-A, PGP9.5, butIHC showed the possible existence of (neuro)endocrine cells in tubal interstitium. We call them'JF cells'.
In conclusion, the idenfification of t-ICC might open the door for understanding some tubal functions,e.g. pace-making/peristaltismn secretion (auto-, juxta- and/or paracrine), regulation of neurotransmission(nitrergic/purinergic) and intercellular signaling, via the very long processes. Furthermore, I-ICC might even beuncommitted bipolential progenitor cells.
Keywords: interstitial cells of Cajal - uncommitted progenitor cells - CDll7/c-kit - intercellular signaling -caveolae/caveolins - vimentin - calcium release units - uterine tube - oviduct - tubal endocrine cells
* Correspondence to: L.M. POPESCU, M.D., Ph.D.Department of Cellular and Molecular Medicine,"Carol Davila" University of Medicine and Pharmacy,P.O. Box 35-29, Bucharest 35, Romania.Tel: +40(0)7214 535 298; Fax: +40(0)21-312 4885E-mail : [email protected]
Reprinted from:Journal of Cellular and Molecular Medicine
Motto:
'As long as there is a hunger for knowledge anda deep desire to uncover the truth, microscopywill continue to unveil Mother Nature's deepestand most beautiful secrets.tL Orci & M.S Peppo, Miuoscopy: an afl?, 2002 (ref. tl}
Published bv:the CMM Foundati<in

Introduction
Santiago Ramon y Cajal reported at the end of thenineteenth century a special type of cell ('nervousinterstitial cell') in the gastrointestinal tract and inother mammalian tissues, using methylene-blue vitalstaining and/or Golgi impregnation method [2,3].Although Cajal could not discern an uxon among thecytoplasmic processes of these cells, he consideredthem possible accessory neurons (a short review inref. [4]). Nowadays, habitually, such cells are calledinterstitial cells of CajalQCC) t5l.
ICC were described along the digestive tube [6-12]and several TEM ground nrles for in situICC identifi-cation were formulated [13-15], eve,n an 8-criteria'goldstandard' was proposed by Hvtznga et al. [15].ICC arenow considered to play a key role in pacemaking andneurotansmission in gastrointestinal tact [1G20].Moreover, CDI l7lc-kit became a widely acceptd reli-able marker for ICC at the microscopic level 12.1141.
In a series of recent papers, ICC-like cells have beenclaimed to occur in other organs: vas deferens [25], ure-thrd,B6-28l,b1afulzg-33l,ureter[34,35],uterus[36,371, blood vessels [38-40], lymphatics l4ll, pro{ate f42,431, penis f44l,mammary eland I45l andpancreas [46].
Apparently, the fallopian tube was unexplored forthe occurrence of interstitial (Cajal-like) cells, inspite of the fact that this hollow organ is a tube (hkethe digestive tube) andexibits peristaltic waves (likethe digestive tube, again !). We were unable to findout any reference in the international biomedical lit-erature conceming the possible existence of ICC orCajal-like cells in human fallopian tube.
From a microscopical point of view, the fallopiantube appears disappointingly simple: an epithelium +lamina propria (stroma) and two interwined smoothmuscle layers covered by serosa. Moreover, histologicfindings in fallopian tubes have been only sporadicallydescribed 147 ,481, although fallopian tubes have a vital(subtle) role in promoting human fertility [49]. Albeitin mammals fertilization and early embryo develop-ment take place in the fallopian tube [50], a mysteryremains: how this organ manages to orchestate thetransfer [47] so that sperrnatozoa moving towards theovary and the egg moving away from the ovaqy meetin the right place, at the right time, for fertilization. Apresumptive IICC network might hence be the micro-scopic missing-link (conductor ?) of the tubal players:the epithelial'conveyel, multidirectional smooth-mus-cle bundles, nerve endings, and small blood vessels.
480
Fig. I Human fallopian tube; mucosal layer; 100x, oilimmersion. Routine light microscopy, hematorylin &eosin staining. Note the possible t-ICC, withavery long(over 25 ynl, monilifom (uneven caliber) cytoplasnicprolongation. 'F' marks the euchromatic nucleus of afibroblast-like cell; cap = capillary; ven : venule;RBCs = red blood cells.
We present here morphological, irnmunochemicaland electophysiologcal evidence for the occurence ofCajal-like int€rstitial cells in the human fallopian tube,in the non-epithelial space.
Material and methods
Human tissue specimens
Histologically normal portions of fallopian tube ampullar regionwere obtained from 1l healthy women of non-reproductive age,after surgical procedures for leiomyoma (total abdominal hys-terectomy with bilateral salpingo-oophorectomy). Sampleswere taken after informed consent, using a protocol approved bythe local Bioethics Committee, in accordance to generallyaccepted intemational practice. Tissue samples have been divid-ed into appropriate-sized slices for cell culture, conventionalhistology and TEM. For immunohistochemistry, tissue sampleswere obtained from 14 archived paraftin-embedded specimens
ofpatients that had undergone surgical procedures (total abdom-inal hysterectomy with bilateral anexectomy) for a non-neo-plastic disease, as well as after salpingectomy or tubal ligation.

Cell culture
Human fallopian tube fragments were collected into sterile tubescontaining Dulbecco's Modified Eagle's Medium @MEM),supplemented with 100 UI/ml penicillin, 0.1 mglml strepto-mycin, and 0.25 ltglml amphotericin (Sigma Chemical, St.Louis, MO, USA), placed on ice, and transported to the cellculture laboratory. Samples were processed within 30 min fromsurgery rinsing them with sterile DMEM. Then fragments weredissected in Petri dishes along the longitudinal axis, exposingthe epithelium, and the mucosal layer was gently peeled off.The experimental protocol was an adaptation of recentlydescribed methods [51, 52]. The remaining muscular tissue wasminced in fragments of about I mm3 and incubated on anorbital shaker for 4 h, at 37oC, with l0 mg/ml collagenase typeIa and 2000 U/ml deoxyribonuclease I (Sigma Chemical, St.Louis, MO, USA) in PBS, without C*+ and Mg2+. Dispersedcells were separated from non-digested tissue by filtrationthrough a 40 pm diameter cell strainer (BD Labware, San Jose,
CA, USA), collected by centrifugation at 3009, and resuspend-ed in a I : I DMEM: Ham's Nutrient F- 12 mixture, supplement-ed with l0% fetal calf serum, 1.5 mM HEPES, 100 UI/ml peni-cillin, 0.1 mg/ml sheptomycin and 0.25 pg/ml amphotericin(Sigma Chemical, St. Louis, MO, USA). Cell density wascounted in a haemocytometer and viability was assessed usingthe Trypan blue dye exclusion test.
J. Cell. MoL Med. Vol9, No 2, 2005
t-rcc
1,, .,':
rlI.:.ll
1l:1. l-.f'*;:
;* a -__t'-, *%.ry,].. :i:l:.'. ':. r',
..
-+
t.lcc
K:r,,1..'
}.?i.'r', ,.
,t'::.1'lr\''t
i ''li'!..:t:,t'.'
Cells were distributed in 25 cm2 plastic culture flasks, or onglass coverslips into 24-well plates (BD Labware, San Jose, , 'l
cA'UsA)atadensityof50,000cel1s/cm2,andmaintainedat37'C, in a humidified atmosphere (5%o CO2 in air) until becom-ing semi-confluent (usually 4 days after plating). Culhre medi-um was changed every 48 h. Cell cultures were examined by l":
phase contrast microscopy, under an inverted Nikon TE200microscope.
Fig. 2 A-C Human fallopian tube; muscularis; lightmicroscopy. Wtal methylene-blue staining, before fixa-rlon (cryosections). A. Original magrification, 10x.Note the very selective staining of only two I-ICC in afield of 13,000 p*2, which normally would display tensof cells, when routine methodology is used (see Fig. Ifor comparison). B, C. Higher magnification (100x,oil immersion), photographic reconstructions. Note thevery long (over 100 pm), moniliform cytoplasmic pro-cesses (anows) emerging from I-ICC body.
-'l
- i'jix:.f,:ea{s*' ..r r:a€*-'39!6x'
i*.&&*P
481

'.. "#
Phasd
;cohtrastM.a, .,-t& I+
bt
Fig. 3 Human fallopian tube, preconfluent primary cellcultures. A, B. Cells with t-ICC-like morphology can berecognized among smooth muscle cells. Observe the verylong, monilifofin processes of I-ICC and the close con-tacts with smooth muscle cells (green dashed lines).Original magnihcation 40x. C-E. Conventional lightmicroscopy. C. Giemsa staining, original magnification100x, oil immersion. D, E. Crystal-violetvital staining;original magnification 40x. 'Beads' of the moniliformcytoplasmic process are indicated by arrowheads. A net-work-like t-ICC disposition is shown in E.
,. Crystal violet
.\\
\ ut!
'octmpus-like'\r-&cc
'to .
-t*
,t&
*.*i
Itl
1
-t*Fp'*
4
6r *tA
d
# .,:
GIEMSASTAIN
t'l ,t,A'=i;lg. i'F i*o
?*
,:f,tt #it
j1,iq
I,:G
*::
fu! .d,
{
!t
/.
r-$ffifl d
\ li:dt'.
s*1
tr482
.*

J. Cell. Mol. Med. Yol9, No 2, 2005
Fig.4 t-ICC in primary cul-ture, original magnification40x. A. Crystal-violet vitalstaining makes the I-ICC visi-ble, using conventional lightmicroscopy; B. The same cellafter tlte dye was washed-out(30 min with DMEM); thet-ICC was visible only in phasecontrast. C. The same cellafter Trypan blue staining;(this viability test shows thatthe cell is still alive, since thedye was excluded). 'Invisible'prolongations may be onlypresumed using conventionallight microscopy.
Cytogenetic analysis was performed in order to evaluate thecell genome status. The cells were grown until 70-80% conflu-ency at which point colchicines was added to a final concentra-tion of 2pglml for 3 h. Cells were then trypsinized, pelleted bycentrifugation, resuspended in 0.075 M KCI solution and incu-bated at 37oC, for 30 min. Cells were fixed in a 3:lmethanol:acetic acid fixative solution for 30 min, at 4oC, pelletedby centifugation, supematant was removed and fresh fixativewas added. Karyotyping was assessed on GTG-banded slides.50 metaphases were counted with a Nikon Microphot SA micro-scope and karyotyping was performed using a Lucia Cytogenetics1.2.9 system. A normal 46,M karyotype was revealed.
Vital stainings
Fresh tissue samples. For the vital methylene blue staining weused Shabadash's method, modified by Niculescu [53] in 1958.In brief, fallopian tube samples were immersed in a bath ofwarm (37'C) methylene blue dye (0.025%) and incubated for30 min in the presence of resorcine (0.015%), glucose (0.02%),magnesium chloride (0.15%) and sodium chloride (0.85%).Staining procedure was stopped, under light microscope con-trol, by fixation with 8% ammonium molibdate solution. Afterextensive washing, tissue samples were frozen (minus 25oC)and cut with a cryomicrotome (Leica; Bensheim, Germany).Sections (4-5 pm thickness) were dehydrated in absoluteethanol, cleared in toluene and mounted on slides, in syntheticresin. Slides were examined under Nikon 600E microscope.
Trypanblue
cell
Primary cell cultures. Methylene blue vital staining: cel\swere washed in pre-warmed phenol red-free DMEM (SigmaChemical, St. Louis, MO, USA), and incubated for 20 min in a0.02o/o methylene blue solution, 37.C (Merck KGaA,Darmstadt, Germany). After a rapid wash with phenol red-freeDMEM medium, pictures were taken within 10 min to avoidthe spontaneous color loss. A Nikon inverted TE200 micro-scope equipped with a Nikon DN-5 digital camera was used toexamine the samples.
Crystal-violet vital staining. Cells grown on coverslips in2 cm round wells (the {ttt day of culture), in DMEM with 10%fetal bovine serum (FBS), HEPES 1.5 mM, were washed threetimes in pre-warmed (37"C) culture medium. The cells wereimmersed in 0.01% crystal-violet (E.Letz Zweiggeschiift,Berlin NW, Germany) in culture medium for 2-5 min, at37oC(in a humidified atmosphere, 5% CO2 in air), and afterwardsrapidly washed with phenol red-free DMEM, and examined.Cells were then incubated at 37oC in DMEM, and gently agi-tated periodically to remove the dye. After 30 min, cells viabil-ity was tested with Trypan bllue 0.2%o,5 min, at 37oC, washedthree times with DMEM, and examined under microscope.
Janus green B vital staining. Cultured cells were immersedfor 30 min in 0.O2oh Janus green B (Schering Kahlbaum AGBerlin, Germany) [5a] in DMEM and kept at 37oC,ina humid-ified atrnosphere, 5%o CO2 in air. After repeated washes inDMEM, cells were examined under inverted Nikon TE200microscope with heated object-stage, and photographed.
Mitochondria labeling with MitoTracker GreenFM Cultures were labeled with MitoTracker Green FM(Molecular Probes, Eugene, OR, USA), the lipophylic, selective
483

. ti.I!
- ,! :'
-a
Fig. 5 A-D Human fallopian tube. Methylene-bfue vinl snining of cells having t-ICC morpholory. Notn selectivelystained cells with long, moniliform, prolongations (red dashed lines). Day 4 of primary culflre. Arrows indicate closecontacts between t-ICC and smooth muscle cells (green dashed lines). Original magnification 4Ox, Nikon TE 300.
dye, which is concentrated by active mitochondria [55,56].Cells grown on coverslips were removed from culture and incu-bated in phenol red-free DMEM completed with l0% fetal calfserum and antibiotic-antimycotic solution (Sigma Chemical, St.Louis, MO, USA) in the presence of 80nM MitoTracker GreenFM. Cells were incubated for 30 min, at37"C, in a humidifiedatmosphere (5% COz in air), subsequently washed, and exam-ined by fluorescence microscopy (450-490 nm excitation light,520 nm barrier filter; Nikon TE200 microscope).
Giemsa staining. This procedure is not areal vital staining,since the dye and the fixation agent (methanol) are acting con-comitantly on Live cells. Giemsa stain was performed afterremoving the medium from culture wells and adding a llI (vlv)mixture of 0.4% Giemsa solution in methanol and distilledwater (pH 6.9) and incubating, for 30 min, at room tempera-
484
hue. Coverslips were then washed three times in distilled waterand examined.
Conventional light microscopy
Classic techniques. Parafiin embedded sections (4 pm thick-ness) of formalin-fixed material were examined after usualstaining (hematoxylin&eosin, HE), trichrome stainings (hema-toxylin&eosin/methylene-blue, van Gieson) and silver impreg-nations, according to generally accepted procedures [57],adapted to our laboratory.
Gomori silver impregnation. After de-waxing, sectionswere treated with l% potassium permanganate for 3 min,rinsed in tap water and then bleached in l% oxalic acid. After

:-i:-
tr.1 ":1 ;"--' ::
"'.,.r ,r i1:. +.' .''..':
,""
I.
t
lrt
,r.
'.-t-
J. Cell. MoL Med. Vol9, No 2, 2005
another series of rinsing, the sections were treated wi th2%o ironalumn for 15 min. Foliowing repeated washes in deionisedwater, sections were immersed in ammoniacal silver nitrate, forI min, rinsed again, dehydrated, cleared in xylene and counter-stained with eosin.
Giemsa stain was applied on fallopian tube paraffin-embedded sections, after deparaffination and rehydrationthrough graded alcohols to water. Working Giemsa solution(Giemsa stock diluted with distilled water) was maintained onsections for about 10 min, under microscopic control. Slideswere then washed in distilled water, followed by dehydrationin alcohol 960/o and,two changes ofisopropilic alcohol, clearedin benzene, and mounted in Entellan (Merck KGaA,Darmstadt, Germany).
Vital dyeson fixed and paraffin-embedded tissueJanus green B stain. Sections were flooded with freshly pre-pared Janus green B (aqueous solution 0.10%) under micro-scopic control, and then rinsed in acidulated (acetic acid) dis-tilled water. Samples were then dehydrated through alcohols,cleared in xylen and mounted in a synthetic resin.
Crystal violet stain. Hydrated sections were maintained incrystal violet solution (1%) under microscopic control. Afterdehydration in 96%o and absolute alcohols, samples were dif-ferentiated. Differentiation was stopped in water and repeateduntil good contrast was obtained and mounted in a syntheticresin.
Fig. 6 A-E Human t-ICCin primary culture.4,,8. Wtal staining withJanus green B, a marker formitochondria in living cells.Note the location ofmitochon-dria at the cell-body level, as
well as along the long, monil-iform processes. In fact, thedilations ofprocesses are filledwith Janr"s green B positivemitochondria. Original mag-nification,40x.C. Phase contrast microsco-py of a living riCC. Thesame t-ICC after loading (D)with MitoTracker Green FM.D, E. Fluorescence images ofmitochondria with MitoTrackerGreen FM. Green fluorescentmitochondria can be seen
around the cell nuclei and
within expansions locatedeither along or at the endings
of long, moniliform, cell pro-cesses (ori ginal magnifi cation:
40x objective).
e
.tJanUS t
green B. l
,o,it
Janusgreen B
4
**fl&
,:i/
48s

GIEMSASTAIN
Fig. 7 Human fallopian tube; 100x, oil immersion. Routine light microscopy; different staining methods, afterfix-ation, paraffrn embedding and sectioning. Note the spindle-shaped I-ICC (dashed lines), with very long, monili-form, cytoplasmic processes. F : nuclei of fibroblast-like cells; L : lymphocyte.
486
*'l*tFtigI,
I
dH\

Safranin O (Basic Red 2). Paraffin-embedded sectionswere de-waxed, hydrated and stained with safranin O(Dr. Griibler & Co., Leipzig, Germany) aqueous solution(l%), under microscopic control. After a brief wash and dehy-dration in absolute alcohol, sections were cleared in xyleneand mounted in a synthetic resin.
hctoria blue (Basic Blue 26). Sections mounted ontoslides were stained with Victoria-blue @r. Gr0bler & Co.,Leipzig, Germany) alcoholic solution (l%), afiprocessed asabove.
Tlansmission electron microscopy (TEM)
Small fragments (about I mm3) were fixed in 4%o glu-taraldehyde (in 0.1 M cacodylate buffer), pH7.3, for 4 h, at4oC. After a brief wash in 0.1 M cacodylate buffer (CB),tissue samples were post-fixed with l% osmium tetroxidein 0.1 M cacodylate buffeq pH 7.3, at 4oC, followed bydehydration in a graded series of ethanols. After impregna-tion in propylene oxide, the samples were immersedovernight in a mixture ofpropylene oxide and Epon resin,and embedded in Epon 812, as usually. Ultrathin sectionswere cut using an MT-7000 ultramicrotome (ResearchManufacturing Company Inc., Tucson, AZ, USA). Thesesections (50 nm) were collected on Formvar-coated coppergrids, stained with uranyl acetate and lead citrate, andobserved in a CM 12 Philips electron microscope, at anacceleration voltage of60 kV.
J. Cell. Mol. Med. Vol9, No 2, 2005
The quantitative evaluation of t-ICC ultrastructural char-acteristics, (e.g. relative volumes occupied by nucleus, mito-chondria etc.) was performed using the classical test-systemdescribed by Weibel [58].
Non-conventional light microscopy(NCLM)
Control semi-thin sections (less than I pm) were stained with0.25% Toluidine blue and examined by light microscopy(Nikon Eclipse E600). Representative photomicrographswere taken using Nikon Plan 40x and Nikon plan Fluor100x/1.30 oil.
Because semi-thin sections are about 100 times thinner,the spatial resolving power in NCLM is much better thanusing paraffrn-embedded sections. Therefore, NCLM waspreferentially used for quantitative and morphometric analy-sis of FICC (e.g. total and relative number of cells, numberand length ofprocesses etc.).
Immunohistochemistry (IHC)
The human fallopian-tube specimens were tested using the fol-lowing antibodies: CDll7/c-kit, polyclonal, I:I00 @AKO,Glostrup, Denmark), CD34, monoclonal, l:100, cloneQBEND10 (Biogenex, San Ramon, CA, USA), 5-100, poly-
Fig. 8 A, B Human fallopian tube; trichrome staining(hematoxylino eosin, methylene-blue) after fixation,paraffin embedding and sectioning. Cells witl long,thin, blue-stained processes (red dashed lines), sugges-tive for I-ICC morphology, can be identified in laminapropria. Original magnification 100x, oil immersion.
487

clonal, l:500 (DAKO, Glostrup, Denmark), d-smooth muscleactin (o-SMA), monoclonal, 1:1500, clone 1A4 (SigmaChemical, St. Louis, MO), CD57, monoclonal, l:50, cloneNKl, (DAKO, Glostrup, Denmark), nestin, monoclonal,1:100, clone 5326 (Santa Cruz, CA, USA), desmin, mono-clonal, 1:50, clone D33, (DAKO, Glostrup, Denmark),vimentin, monoclonal, l:50, clone V9, (DAKO, Glostrup,Denmark), NSE, monoclonal, l:50, clone BBS,A{CNl-H14,(DAKO, Glostrup, Denmark), GFAP, monoclonal, 1:50, clone6F2, (DAKO, Glostrup, Denmark), CD68, monoclonal, 1:50,clone PG-M1, (DAKO, Glostrup, Denmark), CD6ZP(selectin), monoclonal, 1:25, clone 1E3, (DAKO, Glostrup,Denmark), CD1a, monoclonal, 1:30, clone CDla-235,(Novocastra, Newcastle upon Tyne, UK), chromogranin A,monoclonal, 1:50, clone LK2HIO (Novocastra, Newcastleupon Tyne, IJ'K), PGP 9.5, monoclonal, l:40, clone 1041,(Novocastra, Newcastle upon Tyne, UK). A11 specimens werecounterstained with Mayer's hematoxylin, examined and pho-tographed on a Nikon Eclipse 600 microscope.
Immunohistochemistry was performed on 3 pm thick sec-tions from formalin-fixed paraffin-embedded specimens,according to the Avidin-Biotin-Complex method of Hsu [59],modified by Bussolati and Gugliotta [60]. Briefly, the proce-dure comprised: deparaffination in xylene and alcohol series,rehydration, washing in phosphate buffer saline (PBS), block-ing with normal serum, for 20 min, incubation with primaryantibody overnight then with standard labeled streptavidin-antibody biotin (LSAB kit, DAKO, clostrup, Denmark),washing in carbonate buffer and developing in 3,3'-DABhydrochloride/H2O2. Microwave antigen retrieval (in 10 mMcitrate buffeq pH 6.0) was performed for CDllT/c-kit, chro-mogranin A, S-100 and CD68.
For CDl17 and CD34 double immunostaining in humanfallopian tube an ABC indirect tristadial method was used:
Lumen
DAB (diaminobenzidine), HRP (horse radish peroxidase)chromogenic substrate developing for CD I I 7 polyclonal anti-body (brown stain) and fast red alpha naphtol Ap (alkalinephosphatase) chromogenic substrate in 0.2M Tris buffer, pH10, for CD34 monoclonal antibody (red stain).
Negative controls were done using an irrelevant primaryantibody or replacing the secondary antibody with phos-
t-tcc
Fig. 10 A,B Gomori silver impregnation of hrrnut fallopian fimbriae (A) and additional eosin counterstaining of thesame (B). t-ICC with one, two and four processes can be seen underlying the epithelium. Original magnification 100x.
488
Fig. 9 Human fal-lopian tube; 100x, oilimmersion; van Giesonstaining. t-ICC pro-cesses appear in dark-brown ys. the red con-nective tissue.
traa l'1
\I
*.,I
*t
,l
Lamina

J. Cell. MoL Med. Vol9, No 2, 2005
Fig. 11 A-D Non-conventional light microscopy (NCLM). Semi-thin sections (0.5-l pm thick) of fallopian-rube wall;Epon-block cross sections were stained with toluidine blue, and photographed under a light microscope. The absoluteand relative numbers of I-ICC were counted. White dashed lines mark areas where cormts were done: superficial an4subepithelialregionsoflaminapropri4equivalentto 50o/oand100%ofepitheliumheight,respectively(arrows).Atleast20 |-ICC are shown by red dashed lines in lamina propria. * : epithelial peg cells. Original magnification, 4Ox.
phate buffered saline. A positive control (a tissue known toexpress the marker in question) was used for every specificimmunocytochemical stain. To ensure the reliability ofexperimental study, internal quality control of immunocyto-chemical techniques was performed as a part of an imple-mented and certified quality assurance system (ISO9001/2001).
Immunofluorescence (IF)
Primary cultured cells grown on coverslips were fixed in2%o paraforrnaldehyde for 10 min, then washed in PBS and
permeabilized in PBS containing 0.5o/o bovine serum albu-
min (BSA) and 0.075%o saponin (PBSSA) for 15 min (a11
from Sigma Chemical, St. Louis, MO, USA). Cells were

!Xi-! i
&':l 'ja. "l
r. 'i.*|**4'
* :{*.,.1i._ 1
&.3:.
*€a
€ 3,,
{i\**qf;
* Fl:'.4.*...*.:
incubated with primary antibodies, diluted in PBSSA, for4 h at room temperature on an orbital shaker. After threeserial rinses, primary antibodies were detected using sec-
ondary monoclonal rat anti-mouse biotinylated antibodies,clone A85-1, 1:200 (BD Pharmingen, San Jose, CA, USA)and streptavidin - Alexa Fluor 546 (Invitrogen MolecularProbes, Eugene, OR, USA). A second primary antibody
was then applied and, subsequently, a secondary FITC-conjugated polyclonal antibody. Finally, nuclei were
counterstained with I pg/ml Hoechst 33342 (Sigma
Chemical, St. Louis, MO, USA). Cultured cells were test-
ed using the following primary antibodies: mouse anti-human CD117lc-kit, monoclonal, clone Ab 81, 1:100
490
-.€*"u:
l]':!\..\
Fig. 12 A, B Non-conven-tional light microscopy (tolui-dine blue staining; originalmagnification, 100x, oil im-mersion) of human fallopiantube. Note the very long,moniliform processes of I-ICCin muscularis (red dashedlines). The area within theellipsoid, encompassing acapillary (B), is also shown inthe electron micrograph inFig.18A.
(Santa Cruz Biotech); mouse anti-human CD34-FITC,monoclonal, clone 8G12, l:10 (BD Immunocytometrysystems San Jose, CA, USA); mouse anti-vimentin, mon-oclonal, clone V9, l:100 (DakoCytomation, Glostrup,Denmark); mouse anti-g-smooth muscle actin, FITC-conjugated, monoclonal, clone lA4, l:200 (SigmaChemical, St. Louis, MO, USA); mouse anti-desmin,monoclonal, clone RD30, 1:50 (BD Pharmingen, San Jose,
CA, USA); mouse anti-caveolin 1, monoclonal, clone2297,1:100 (BD Pharmingen, San Jose, CA, USA); mouse
anti-caveolin 2, monoclonal, clone 65, 1:100 (BDPharmingen, San Jose, CA, USA). Negative controls used
the same technique, without primary antibodies. Pictures
*;'"6*.3* ,.., '' '{"si*''%.,'..*:r'
:"q;i\- * -

J. Cell. Mol. Med. Vol9, No 2, 2005
&:":. **
Fig. 13 A-D Non-conventional light microscopy. Toluidine blue stained semi-thin sections: longitudinal (A), oblique(B&C) or cross-sections (D). The red dashed lines indicate at least 10 I-ICC (cell bodies and the emergent long, monil-iform processes) amongst the smooth muscle fascicles. Original magnification, 100x, oil immersion.
were taken using a Nikon DXI camera mounted on a
Nikon TE300 microscope and Nikon PlanApo 40x and 60xobjectives.
Paraffin-embedded formalin-fixed sections of human
oviducts were also used to study CDla expression using IF.
Standard dewaxed sections were rinsed in PBS, blockedusing PBS supplemented with 0.5% bovine serumalbumin(BSA), and incubated overnight at room temperature with a
FlTC-conjugated monoclonal CDla antibody (clone HI149,working dilution 1:10 in PBS supplemented with 0.5% BSAand 2mM sodium azide, BD Pharmingen, San Jose, CA,USA). The slides were subsequently rinsed in PBS, cover-slipped using a fluorescence mounting medium(DakoCytomation, Glostrup, Denmark) and examined.
Non-specific fluorescence was evaluated using an irrele-vant isotype-matched FITC conjugated antibody as nega-
tive control.
Electrophysiology
Extracellular single unit recordings were performed on t-ICC inprimary cultures (the 3.0 day of culture), grown on coverslipsin 2 cm round wells, in DMEM supplemented with l0% fetalbovine serum (FBS), HEPES I .5mM. A Nikon inverted TE200microscope equipped with a Nikon DN-5 digital camera and an
Eppendorf (Eppendorf AG, Hamburg, Germany) NK I micro-manipulator were used to position the microelectrodes. On-sitemade glass microelectrodes (WPI 1B150F-4) were filled with3 moL{L potassium acetate and had resistances befween 4 and 8
MOhms. The field potential variation was measured using a
WPI Duo 773 elechometer (World Precision Instruments,Sarasota, FL, USA), and outputs were displayed on an oscillo-scope. Electrical signals were digitized using aDigidata 1322Adata processor (Axon Instruments, Union CiE CA, USA) and
digitally stored. The Clampex 8.2 software was used for data
acquisition and analysis.
491

0123456Number of cyoplasmic processes of I-ICC
Fig. 14 The number of cytoplasmic proc,esses
fromt-ICC body) plottedagainstthe fiequenry in rhe wholepopulation of I-ICC (amina propria + muscularis); NCLM:232 FICC were counted, on 52 photographs. The curvelooks like the right 'portion of a typical Gaussian curve'.
Results
Interstitial cells resembling the archetypal entericICC, if present in human fallopian tube (t-ICC),might be found in:
- mucosa: in the 'unremarkable' lamina pro-pria - the interstitium containing fibroblasts,mast cells, lymphoid cells, collagen and retic-ular fibers, etc.;
- muscularis, in the loose connective tissuewhich fills interstitial spaces between thebundles of circular and longitudinal smoothmuscle fibers, or in
- subserosa, the loose connective tissuebetween serosa and the muscularis.
The traditional, routine staining with hema-toxylin and eosin (Fig. 1) cannot be succesfully usedfor identification of I-ICC. The main bias remainsthe difficulty to differentiate between the cell pro-cesses and collagen fibres.
Supravital staining of fallopian tube frag-ments with methylene-blue resulted in the visual-ization of Cajal-like interstitial cells (Fig. 2).Interpreting in visual terms, Fig. 2 seems the first(strong) indication that I-ICC do exist. Additionalproof was provided by growing I-ICC in vitro.
We estimated the spatial density of I-ICC in theampullary region of the fallopian tube since this is
492
pynform spindle(l) Q)
I-ICC shape
Fig. 15 Histogram showing the average length (pm) oft-ICC processes as a function of cell-body shape; numberof processes in brackets; NCLM: 216 cells, on 50 pho-tographs were counted (p < 0.001).
the location of fertilization and early embryo cleav-age. Using the methylene-blue vital staining fol-lowed by cryosectioning, wo found a value of100-150 cells/mm2 (Fig. 2). This density is of thesame order of magnitude as that reported byAlbertiet al. [61], who estimated the distribution of c-kitpositive ICC in rat colon.
t-ICC in primary culture
t-ICC were successfully maintained in primary cul-ture and can easily be identified (distinguished fromthe smooth muscle cells) before reaching confluence(Fig. 3.{, B). Starting with the 3rd day in culture,|-ICC appear with long, monilifurm processes actend-ing from the cell body (Figs.3-{). Concomitantly,some cells begin to contact each other or establish'connections'with smooth muscle cells, creating anetwork appeaftmce. A rough quantitative estimationof in vitro cell population (n : 32$ reveals 9.9+0.9%t-ICC and 69+2.5yo smooth muscle cells. Theremaining were fibroblasts (17.8t1.8olo) or cells ofother (difficult to establish) phenotypes.
We used Giemsa stain to recognize culturedI-ICC, since this stain is a mixture containingmethylene-blue eosinate and methylene-blue chlo-ride, dissolved in methyl alcohol, with glycerol as a

stabiliser. Fig.2C displays a spectacular octopus-liket-ICC. Since Giemsa stain is acting concomitantly asdye and fixative, we searched for the characteristicmoniliform aspect of I-ICC processes :using vitalstains: crystal violet (Fig. 3D, E; Fig. 4A{),methylene blue (Fig. 5A-D), Janus green B (Fig.6,4', B), MitoTracker Green FM (Fig. 6D, E).
Vital stainings with crystal violet and methy-lene blue revealed that in living cells, the I-ICC pro-cesses are very long (several tens of pm), have anuneven caliber, with dilated portions, resembling'beads on a string'.
Previous TEM studies on pancreatic Cajal-likecells in our laboratory showed that the dilated por-tions of the long processes, frequently accomodatemitochondria [46]. Janus green B, a well-knownvitol stain, with high affinity for mitochondria [62]was used to assess I-ICC viability and localize mito-chondria. The initial dark green-blue color, due tomitochondria stained with Janus green B, became abrownish gray one, and finally decolorized (leuco-compound). Fig. 6.{, B illustrate the staining oft-ICCbody and dilations of the processes. Color changesare obvious. To double-check these results, we used a
fluorescent probe for mitochondria, MitoTrackerGreen FM [55,56]. Figs. 6C & E indicate that fluo-rescent mitochondria are located in both cell body anddilations of long, moniliform cell processes.
J. Cell. Mol. Med. Vol9, No 2, 2005
Other tested vital dyes (orange G, methyl greenand safranin O) failed to stain I-ICC in primary cul-ture, under our experimental conditions.
Usual light microscopy
Since I-ICC were successfully identified in primarycell cultures with various staining procedures, wefurther tried to figure out, whether is possible tostain 'selectively' Cajal-type cells in fixed, paraf-fin-embedded sections, using vital dyes.
Vital stains (methylene blue, crystal violet,Victoriablue, Janus green B and safranin O), as wellas Giemsa stain are effective in pointing out theI-ICC processes (Fig. 7A-F). Similar results(Figs. 8, 9) were obtained using trichrome stainingsmethods (hematoxylin/eosin/methylene-blue, andvan Gieson). Taking into account that Ramon yCajal discovered the 'interstitial neurons' usingGolgi silver impregnation, besides methylene-bluevital staining, it appeared attractive to test silverimpregnation too. Modified Gomori silver impreg-nation (Fig. 10) particularly reveals more and longercell processes than the other mentioned techniques.For instance, 'octopus'-1ike IICC depicted in Fig.l0B seems similar to those found in cell cultwe(compare with Figs. 3C,24D and 4l).
Fig. 16 Bundles of unmyeli-nated nerve fibres in themuscular layer of a humanfallopian hrbe. Photographicreconstruction of a semi-thin(less than 1 pm) sectionstained with toluidine blue.Higher magnification image(TEM) of the squared dottedarea is presented in Fig. 188.Red arrow indicates the pos-sible location of a t-ICC-process in the connective tis-sue surrounding nerve fibresfascicles.
493

Non-conventional light microscopy (NCLM):
Topography, relative density anddistinctive processes of t-ICC
NCLM consists essentially in processing tissuespecimens as for TEM, but (semi)thin sections,less than 1 pm thickness, were stained with tolui-dine blue, and finally examined under a lightmicroscope. This approach enables a better struc-tural discrimination of I-ICC; compare Figs. I and7-10 with Figs. 11-13. NCLM seems the electionmethodology to easily establish the location andthe number of t-ICC. However, the limitations ofNCLM versus TEM are obvious (compare Fig.12B with Fig. 18A, and Fig. 16 with Fig. tSB).
Localization: I-ICC are residents in thenon-epithelial interstitial domains, mainly in theconnective tissue of mucosal lamina propria and,in musculans, among the smooth muscle cells(Figs. 11-13, 49). In our opinion, the absolutenumber of IICC seems irrelevant for the presenttime, since t-ICC functions are still subjects ofspeculations. However, we thought that it could beuseful to identi$r some microscopic areas wheret-ICC might be located preferentially. Therefore,we chose five zones of interest across the tubalwall (from lumen to serosa) for estimating sepa-rately the relative numeric density (RND) of FICC(percentage oft-ICC out ofall cells found in thatarea). The results were:
l. the border epithelium/lamina propria, a'belt' 10 pm thick underneath the basementmembrane of the endosalpinx epithelium(RND: 18 + 2Vo, the highest local densityacross the tubal wall);
2. the subepithelial portion of lamina propria(- 20pm thick) (RND : 11.7 + 0.9%o\;
3. the whole territory of lamina propria(RND : - 8%);
4. the muscularis (RND: 7.8 + I.2Vo), and,apparently the less important,
5. submesothelial space (RND : not assessed).These data, also illustrated in Fig. 49, show
clearly a decreasing gradient from epithelium toserosa.It might not be incidental that t-ICC repre-sent 9.9 + 0.9yo of cell population in primary cul-ture (p. 492) and the average numeric density ofI-ICC in lamina propria is about 8o/o or in muscu-laris 7.8 + 1.2yo.
494
Cytoplasmic processes of t-ICC: Fig. 14shows that -50Yo of I-ICC observable by NCLMexhibit only one long process, -30%o and, *15ol.o
have two and three processes, respectively.Therefore, cell-body shape frequency in the caseof t-ICC is: pyriform, fusiform, triangular, tetra-gonal or pentagonal. When a I-ICC exhibits onlyone process, this prolongation is about 30pm (ormore) long. However, when two or three process-es emerge from the I-ICC body, their length isquite equal, around 20pm (Fig. l5).
Close relationships of I-ICC processes withvarious tissue components: l3% of lamina pro-pria I-ICC processes 'look for' the coveringepithelium, while, as expected, none of the mus-cularis I-ICC processes reaches it. Small bloodvessels (capillaries and postcapillary venules)within the mucosal and muscular layers werecontacted by 6% of all I-ICC processes, withalmost equal contribution from t-ICC in both his-tological locations. Close relationships are alsoestablished with nerve fibers (Fig. 16) andbetween I-ICC themselves.
Like in other organs, cytoplasmic processes,which define Cajal-like cells, are long (tens of pm),thin (50.51tm\ and monilifurm. A characteristicI-ICC process network-Iike layout is noteworthy.
TEM:
Ultrastructural characterization of t-ICC
There is no question that TEM is essential toestablish that a given extradigestive interstitialcell-type becomes (or not) a member of theCajal-like cells' selective club.
During the last ten years, several authors madeefforts to systematize the ultrastructural features inorder to facilitate the electron microscope diagno-sis of ICC n3-15, 631 or interstitial Cajal-likecells [64]. In our opinion, the most ellaborate setof criteria is the so-called 'gold standard' pro-posed by Huizinga, Thuneberg, Vanderwinden,Rumessen, 1997 [15] and composed of eight crite-ria. Actually, this standard includes the six criteriasuggested by Faussone-Pellegrini and Thuneberg,1999 ll4l and the three criteria formulated byKomuro et al., 1999 [13]. We emphasize that allthe eight criteria required by the 'gold standard'

J. Cell. Mol. Med. Yol9, No 2, 2005
Table I Semiquantitative data a) conceming the ultrastructure (TEM) of t-ICC (this study) in comparison witltle gancrelic [12] and archetypal enteric ICC (Auerbach's plexus from snallintestinerana,Ciitod; as,,su_tnma,'rized in ref. [0, 14].
Fallopiantube
Pancreas
(exocrine)
Smallintestine
Colon
++ +r
,,iiTT
+++
+
SMC IC
t*+ ++
^')Symbols;O = absent; t: occasionally present; + =present; the relative richness is marked by + + or + + +;Mitochondria as percent of cytoplasmic volume: i: 4.8+1.7o/o; ii = 8.7+0.8%
b) Cav. = caveolae; BL : basal lamina; sER : smooti endoplasmic reticulum; rER : rough endoplasmic reticulum;Mi : mitochondria; IF : intermediate filaments; mT: microtubules; d: thin (actin) filaments.
c) CC : close contacts; GJ : gap junction; SMC : smooth muscle cells; IC : ICC or interstitial Cajal-like cells.* contacts with ductal smooth muscle cells.
[15] are easily recognized in our TEM images(Figs. 17-23):
1. numerous, large mitochondria (Figs. 17, 23;Table I - above & Table 2, p. 503);
2. bundles of intermediate filaments (Figs. 19;Table l);
3. absence of thick/ilamenrs (Figs. 17-23);4. presence ofsurface caveolae (Figs. 19-21,23;
Tables l&2);5. variable developed bosal lomina (Fig. 17;
Table l);6. contacts between ICC and nerve bundles
(Figs. 18A,20; Table 1);7. presence of a well-developed smooth and
rough endoplasmic reticulum (Figs. 17-21,238,C; Tables l&2);
8. close apposition or gap-junction contact withsmooth muscle cells (Fig. 22B; Table 1) orwith each other (Figs. 19,21; Table 1).
Table I shows that tubal interstitial cells areindeed Cajal-like cells, according to the 'gold stan-dard'criteria. In our opinion, the 'gold standard'[15]is instrumental for the morphological identification ofICC and can serve to recognize interstitial Cajal-like
cells outside the musculature of the gastrointestinaltract. However, we suggest that two other criteriacan be added, thus obtaining a decalogue (8+2), a'platinum standard'. Tlhe gh criterion: which ofthe following microscopic (ultra)structures - epithe-lium, capillaries, smooth muscle cells or nerve fibers- is (are) the targe(s) in a given organ (e.g. for fal-lopian tube, all four). The rationale for the 10th crite-rionis, in our opinion, imposedby a syllogism: (l) tobe Cajal-like, a cell has to have cytoplasmic process-es; (2) not any branching interstitial cell is aCajal(-like) cell; (3) only cells with cerrain charac-teristic processes could be considered Cajal-like:
-number (1-5, but 21rrr most cases);
-length (usualy several tens of gm [45], up to100 pm 114,461, or even more [39]);
- thickness (around or less than 0.5 pm; fre-quently < 0.2 pm);
- aspect (moniliform, with dilations accomodat-ing usually mitochondria);
- branching (dichotomous pattern);
- organizatian in network - labyrinthic sl,stem(processes overlapping with processes ofother similar cells).
495

, edifitbiftii,::.,:,'bdu, :, :: bd:coriitddr lif:tlie, l,\re:,oo.atitlil ,
:W1Npq!,1@* r,,, :,,.',,,'..
2 pnr
496
t-lcc

#
16r
J. Cell. MoL Med. Vol9, No 2, 2005
'\: t"-.t'- 1 '."
\.{
Fig. f 8 A. Human fallopian tube; TEM; relationships of t-ICC with a blood capillary. Note at least two t-ICC (marked
! UZ),bu.t, Probably, there are other two similar cells (marked 3 & 4),and a very long and thin cytoplasmic process (inbetween dashed lines). All these cells and processes surround a blood capillary (RBC: red blood iell; doible anow-heads indicate transcytotic vesicles or endothelial caveolae). The distance between t-ICC processes and the abluminalface of the endothelium is frequently around or less than I pm. Red arrowheads show dilations of the cytoplasmic pro-cess, giving the moniliform aspect. The length of the process in between the dashed lines is at least 40 [m, traving verythin portions, less than 0.2 pm, and, therefore, not visible in light microscopy. B. TEM of unmielinated nerve f,rberi. Theaxons sectioned longitudinally (AX), obliqually (Ax) or tansversally (ax) are engulfed by the cytoplasm of Schwanncells. Note the long moniliform process of a I-ICC in between dashed lines which develops a labyrinthic system (upperpart). N = nucleus; V: vesicle; rER: rough endoplasmic reticulum, sER: smooth endoplasmic reticulum.
1.4\ \ttr"
ffi-ff\ti
\r\\.\**td**
b
b ,t \ \{h -\
\"t\
!:,tw*ti:#+&q)'*tr-'i:{ffi:''dJVr'
'' r*ig|g}S,,."'-*'
,:$,
{t,
497

'1*"
Collagen .** "$
s
q.
Gapjunction
, ,:trq{rr
Gollagen
q "4lt*'
!1
,t$
/ or.3* ff@.'- ;;'- * ; - * *; '*r
_.{ :' .*r *"og Fe{1t* L-W,&#.
-'.. Y.-'*Y
flu:''i$*t :.,:- V
",",rn tsi.l'* 'ig,&-.::+:-aw- i! 'a l:'-..\
l. 'tt
+gB

si:i'sE5s:H ai€:e-tE5.>?9=,.A.= Z+E=E;15i lgi IsE
' 5E€ g
iEi,r;E:I;E EE::liE-"= c E: E:;, E : =; iigiigii;BEa€€EEiEFtsE3;ia=iE:€FEi:"9:sE3;;agE t;E-E
sEiEgii;3i g griE€
I.
";":*i":"\4
n
:itt
t{Et{
f$orot
cu
".,O' Ot
'i4S;
'^O"(G'igE
othoc)
>III l:.;ii oE "rtI' i: 6 !'-{
-a t!, lI<z t!! 9L-- -<'f=\1=t=-rFUt-t:ICI6
Jl:I:r l- l-,n\D5
t
is
co,ctt
soo
.:l
1.GY
,a;c-dw+'.da'.-:,r"Srlfi"
-,{
llEr
.',
a,#tjFY, i
Eooo.9
c
.ctctJ
ffi#lrtrBflxt''F*n#trfr$lfg&q*3
***,L
T<*&
,.,&,\:
o.(E
o
iirl t.'-l ,t'tlll r
1;* Et.i' g
r" "j' , U,',3;' *' i o
. o
-*, . .rt*; .g"{-." ; "."'f -C'"
"tP 'r
't E-.' r1 t a..
,'' fi ,,.-1 , i,7 ll;t -5r: ffi
dHtdd/_*4f
il, [* ']1..\ 'i"-.i.l1
\uu2 r"i i''"il i+;
in''' | .,-'"*l '
.. rf' t'Flr
iTt jrt g€.'>
t-.,'' ' ; 1>T
'' \"- t""-\ -.'*&a
L*,@d*

'-ti>'lE€5: iE; ?:EEEr=€-.124 >': {'4= 7 ;i =lElzzE ;a:,; E ;: p :: i;?iE :EE*i?z:z;7
-.c E1::it? *4 A 'rEl e . r€-i"rZe,:EEi;3EE i € =iatZ7?a9 " ai i ? zEa * zi ^-.?E?24^=EzPEi"*:e€iH; j ;:'i€ 5 ?ryi72i_z=
=Zzii.z,Ze *a67.it tE
_eiteE :ii{E *E iz ErE jizEn':azi1!,E?l:F a9?=59 -:,,! t- it i? zE:= i' 4E 2= 7 e=, 724
I ,'i'
lLo-o'
otoz
o"'ooo
aO, troo,o..Eo.
oOoC)
' cct)>'=.,-o i'oXJ1g
lto
I*-;:
''.i:i,;
ir{;:,.1 .[ ".t,,
ql *",-1i
-''" Ll
.:.
{fr.)-
3,J
'4 t/'i .. at+€.'t
'. tla lt: -!-+'tt'
(E(l,=;o.'=(u(l)o1-Eco-;., ;
:'"il' .
t jj lt'.*.+ I
- '*t ':

J. Cell. Mol. Med. Vol9, No 2, 2005
Fig. 2l Cross section (higher magnification) showing a small sector in the wall of an arteriole, corresponding to theblue inset from Fig. 20, p. 500. Note the endothelial cells which lie on a basement membrane and have numerouscaveolae (transcytotic vesicles) on both sides (luminal and abluminal; double arrowheads). Beyond the intima, twotypical smooth muscle cells with caveolae, dense plaques, dense bodies and myohlaments, can be seen. t-ICC pro-cesses (red dashed lines) are present in the peri-arteriolar space. m: mitochondria; sEft: smooth endoplasmic retic-ulum; rER: rough endoplasmic reticulum; arrowheads : caveolae.
A striking ultrastructural hnding, observable in the green inset, is shown in Fig.23C, at higher magnification.
501

Ca2* storage/release unit
process
Fig. 23A-C A. Note the presence of numerous microtubules and caveolae (arrowheads) in the t-ICC process. B. Atypical so called 'Ca2+ storage/release unit'is shown in the t-ICC process. Such units were described inthe peripheryof smooth muscle cells: caveolae associated with endo(sarco)plasmic reticulum (ER) and mitochondria (m). Caveolamarked 1 shows clearly a diaphragm at the neck level. C. The inset presents an unexpected caveolar conformation:'reversed caveola'(green arrowhead). To the best ofour knowledge, no such image has ever been published before.
502
Fig. 22 Ao B Human fallopiantube; TEM. A. Close apposi-
:., tion (x.{,) befween a I_ICCprocess and a macro-
phage. B. Gap junc-tion (dotted inset)
between a smoothmuscle cell and
,. a I-ICC pro-longation.
,*t .
i.fuJff
. "Ji:
':.-'
,,,,lffi;r i':: l]l!
ju
"i
,lhGap
* =i microtubulGs ,:.t ' 't , r . .
\J_

J. Cell. Mol. Med. Vol9, No 2, 2005
}tf.?. Cgmnariyl o!-ug y,lgtne volumes (%o) ocupied by various subcellular eomponents of human rubalcajal-like interstitial cells (t-ICC) and tubal fibroblasts (of the same specimens).
Nucleus
Heterochromatin **
Cyoplasm
Mitochondria ***
Rough endoplasmic reticulum ***
Smooth endoplasmic reticulum ***
Caveolae(surface vesicles) ***
23.6 + 3.2(n= 15)
49.1 + 3.8(n= 12)
4.8 + 1.7(n= 12)
1.1 + 0.6(n:8)
1.0 + 0.2(n:8)
3.4 + 0.5(n: 12)
56.4'+ 42(n:7) '
28.2+ 3.1(n=7)
4.9 + 1.5(n= 10)
ll.2 + 1.8(n:7)
0,7 + 0.2(n:7)
virtuallyabsent *'e*:*
Organelles considered as characteristic featuresfor ICC or Cajal-like cells were not ordinarilydescribed in quantitative terrns. In our opinion, elu-sive descriptions (e.g.'numerous' mitochondria,'well-developed smooth and rough endoplasmicreticulum or 'a large number' of caveolae etc.)
[1]15, 63, &l should be replaced by precise numer-ical information.Table 2 shows that the relative vol-ume of mitochondria is -5% in I-ICC. This is similarto intestinal smooth muscle cells [65], tubal fibrob-lasts (Table 2) or sheep endometrial stomal cells[66]. However, -5% is only a half of the mitochon-drial content in pancreatic Cajal-like cells [45] andmuch less than in other cell types (20-35%o): cardrasmyocytes, parietal gastic cells, nephrocytes, hepato-cytes etc. Finally, what does 'numerous'mean ?!
The relative caveolar volume ?3.5%; Table 2) iscomparable to values in intestinal smooth muscle(21%; [67]) or bladder smooth muscle (3.66%;M. Gherghiceanu & L.M. Popescu, to be published). Inaddition, we found in I-ICC, more caveolae alongcytoplasmic processes (1 caveola I I.73 pm ofmembrane length) versus cell body (1 caveo-Ia I 2.66 pm), further confirmed by IF results (vdeinfra,Fig.24A,B).
Noteworthy, we found at the level of IICC pro-cesses (Fig. 21B) ultastructural complexes called'Ca2+-release units' , cnmposed of endoplasmic reticu-
:*'** % of cytoplasmic volume;**** less than 0.1%. NB.: In vitro, human cultured fibro-blasts may present caveolae.
lum + caveolae + mitochondri4 adjacent to plas-malemma. Such unis were recently described insmooth muscle [68]. To the best of our knowledge, nosttch'Ca2+-release units' were previously described inICC or ICC-like processes.
Differential diagnosis. Fig. 17A,B and Table 2show unequivocally that the cells that we describe byTEM as intentitial (Cajal-like) cells in human fallopi-an tube are not (morphologically) fibroblasts. In sittt,fibroblasts ofadulthumans have an euchromatic nucle-us and a rough endoplasmic reticulurn, with dilated cis-temae, about l0 times more developed than I-ICC.Moreover, basal lamina, caveolae and intercellularjunctions are absent in human fibroblasts in tissue [69].Caveolae could be found in human [70] or mouse [71]fibroblasts, but only in vitro, in cultured cells.
The original definition of the myolibroblastwas based on electron microscopy [69], and nowa-days myofibroblasts are still definable essentiallyby ultrastructure as highly differentiated cells, hav-ing features in common with smooth muscle cellsand fibroblasts [72, 73]. The three essential mor-phological elements ofmyofibroblasts are: (i) stress
fibers, (ii) cell+o-stroma attachment sites(fibronexuses), and (iii) intercellular intermediate(adherens) and gap junctions. But, in the t-ICC,there are no stress fibers, fibronexuses ot adherensjunctions in situ, by TEM, despite some cells in pri-
%o of cell volume;o% of nuclear volume;
503

Fig.24 A-E Human fallopian tube. I-ICC, primary culture (day 4). Immunofluorescence labeling for caveolin-l(A), caveolin-2 (B), desmin (C), vimentin (D) and smooth muscle actin (E). FlTC-conjugated secondary antibodies(green) were used to visualise the reactions; Hoechst 33342 (blue) for nuclear counterstaining. Antibodies againstcaveolin-l (A) and caveolin-2 (B) stain preferentially I-ICC processes, similar to intermediate filaments (desmin -C and vimentin - D). Original magnification, 60x. All the cells shown above coexpressed CDll7/c-kit, as seen inFigs. 26, 35, 39 and 41.
mary cultures (Figs. 24E and 35). Against myofi-broblast morphological phenotype of I-ICC standalso: absence of dense bodies and attachmentplaques, as well as a poor developed rough endo-plasmic reticulum (Table 2), and Golgi area. Thedifferential diagnosis seems obligatory since some
504
authors [74] considered gastric and intestinal ICCto be myofibroblasts. Although, Walter [75] sug-gested the presence of myofibroblasts in the mucos-al layer of uterine tube of eight mammalian species,this assumption was based only on IHC, not byTEM, as required [69,72,73).

J. Cell. MoL Med. Vol9, No 2, 2005
?
;ri.Nf, ;
,:M&qr,*'.alt
/c-kit
Fig. 25 A-E Human fallopian tube.Note the strong CDll7lc-l,tt im-munoreactivity of the spindle-shapedI-ICC (anows) in lamina propria(A{) and the isolated rounded c-kitpositive mast cells (magenta encir-cled areas in B, D and E). Counter-staining with Mayer's hematoxylin.Original magnification,40xA, B and100x C-E.
;.rtr'd*$
d,:]W
trjkll
q. ...
,l:
fu:,, S
s,&,,.
&
"ry117|c-ktt-
ffi
ffik4,
*&ilCD117lc-kit
c&.il :
. ....,*1*
CD117lc-kit
Immunophenotyping
t-ICC were further characterized by correlationsbetween the cell morphology and the cell immuno-phenotype: fifteen different antibodies were usedfor single- or double-immunolabeling. Stainingintensities were considered, according to a scorefrom (-) to (+ + + +) (see Table 3; [76]).
CDll7/c-kit is a transmembrane receptor forstem cell factor, usually considered as a quite spe-cific immunochemical marker for ICC orCajal-like cells [4,21]. However, Torihashi et al.[77] reported that even some typical ICC in humangut do not appear to express c-kit. Moreover,Rumessen and Vanderwinden [63] proposed theclassification of mesenchimal cells: either
Fig.26 A, B Human fallopian tube primary cultures. I-ICC assessed by IF for CDll7/c-kit (red); nuclei counter-stained with Hoechst 33342 (blue). Original magnification, 60x.
"\-*#M
W-ffi
,w,
., ",:.
I{
505
a.s
*'qjr'
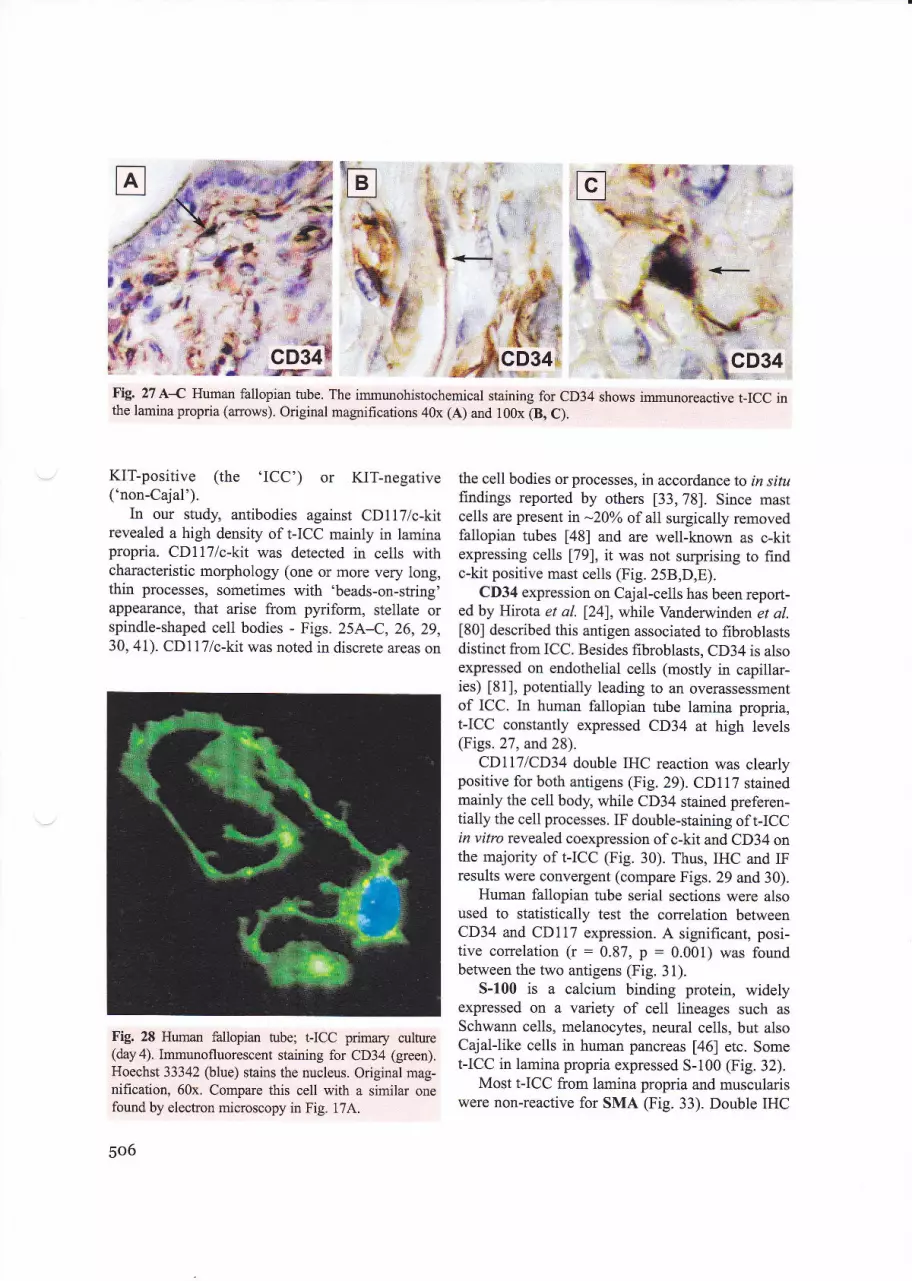
Fig. 27 A{ Human fallopian tube. The immunohistochemical staining for CD34 shows immunoreactive I-ICC inthe lamina propria (arrows). Original magrifications 40x (A) and 100x (8, C).
g
t'
%T
i;;1
't
t'f'
KlT-positive (the 'ICC') or KlT-negative('non-Cajal').
In our study, antibodies against CD1l7/c-kitrevealed a high density of t-ICC mainly in laminapropria. CDll7/c-kit was detected in cells withcharacteristic morphology (one or more very long,thin processes, sometimes with'beads-on-string'appearance, that arise from pyriform, stellate orspindle-shaped cell bodies - Figs. 25A1,26,29,30, 41). CD1l7lc-kit was noted in discrete areas on
Fig. 28 Human fallopian tube; t-ICC primary culture(daya). Immunofluorescent staining for CD34 (green).Hoechst 33342 (blue) stains the nucleus. Original mag-nification, 60x. Compare this cell with a similar onefound by electron microscopy in Fig. l7A.
506
:: ..' ;s?. ._t: le:'-.,o **l'$ry&r 9 , ,:€, :::,, ', ,:4 !,1,,?;t f ,i$': .. $r'+4
the cell bodies or processes, in accordance to in situfindings reported by others [33,78]. Since mastcells are present in -20oh of all surgically removedfallopian tubes [48] and are well-known as c-kitexpressing cells [79], it was not surprising to findc-kit positive mast cells (Fig. 25B,D,E).
CD34 expression on Cajal-cells has been report-ed by Hirota et al. [24], while Vanderwinden et al.[80] described this antigen associated to fibroblastsdistinct from ICC. Besides fibroblasts, CD34 is alsoexpressed on endothelial cells (mostly in capillar-ies) 181], potentially leading to an overassessmentof ICC. In human fallopian tube lamina propria,t-ICC constantly expressed CD34 at high levels(Figs.27, and 28).
CDILT|CD34 double IHC reaction was clearlypositive for both antigens (Fig. 29). CDl17 stainedmainly the cell body, while CD34 stained preferen-tially the cell processes. IF double-staining of I-ICCin vitro revealed coexpression of c-kit and CD34 onthe majority of I-ICC (Fig. 30). Thus, IHC and IFresults were convergent (compare Figs. 29 and 30).
Human fallopian tube serial sections were alsoused to statistically test the correlation betweenCD34 and CDl17 expression. A significant, posi-tive correlation (r : 0.87, p : 0.001) was foundbetween the two antigens (Fig. 3l).
5-100 is a calcium binding protein, widelyexpressed on a variety of cell lineages such as
Schwann cells, melanocytes, neural cells, but alsoCajal-like cells in human pancreas [46] etc. SomeI-ICC in lamina propria expressed 5-100 (Fig. 32).
Most t-ICC from lamina propria and musculariswere non-reactive for SMA (Fig. 33). Double IHC
,ffi:ryiRg4t

J. Cell. Mol. Med. Vol9, No 2, 2005
*!F
CD117lc-kit,,.fr
,,if,
!
& cD34te.?... ' gi
gqffG ::'::# *
Fig. 29 A-D Human fallopian tube; lamina propria of mucosal layer. Double IHC staining. Brown staining (DAB)indicates CD117/c-kit immunoreactivity and red staining (Fast red) shows CD34 positive t-ICC (arrows). Moniliformcell processes have a stronger reactivity for CD34 than for CD1l7/c-kit, the second marker being preferentially dis-tributed on cell bodies (8, D). Original magnihcation 100x.
for 5-100 and SMA displayed two differentimmunoreactive pattems of t-ICC: cells expressingboth 5-100 & SMA in lamina propria, or expressingeither S-100 or SMA in muscularis (Fig. 34). In pri-mary cultured cells we found I-ICC expressing bothc-kit and sMA (Fig. 35).
NK-l is a monoclonal antibody directed againstCD57 also present in neural cells. Fig. 36 showsscattered immunoreactive cells weakly stained forNKl in lamina propria.
Nestin is a component of the intermediate fila-ment network, expressed in gastrointestinal stromal
Fig. 30 Human fallopian tube; I-ICC primary culture (day 5). Double immunofluorescent labeling for CDl17lc-kit(red) and CD34 (green). This cell is positive for both markers, and co-localisation appears as yellow areas. Hoechst33342 nuclear counterstaining (blue). Original magnification, 60x.
507

CDl17lc-ht
CD34
s-r00
a-SMA
CD57
nestin
desmin
vimentin
NSE
GFAP
CD68
CD62P
CDla
Chromo A
PGP9.5
l:100
1:100
1:500
l:1 500
1:50
l:100
l:50
1:50
l:50
1:50
1:50
1:25
l:30
l:50
l:40
DAKO
Biogenex
DAKO
Sigma
DAKO
Santa Cruz
DAKO
DAKO
DAKO
DAKO
DAKO
DAKO
Novocastra
Novocastra
Novocastra
++++++++++
+
+
+
+
t
polyclonal
QBEndl0
polyclonal
1A4
NKl
5326
D33
V9
BBSAiCM-H14
6F2
PG.Ml
rE3
CDla-235
LK2HIO
10Al
Table3 Summary of immuno-histochemical results for I-ICCfrom human fallopian tube.The intensity of t-ICC reactiv-ity was assessed semiquantita-tively using an adaptation ofQuick score method 1761.Intensity:o Negative (no staining of any
oellular part at high magnifi-cation); -r OcssionaW weakpositive +
r Low (only visible at highmagnification): +
o Medium (readily visible atlow magnification): + +
r lligh (shikingly positive athigh magnification): + + +
e Strong (strikingly positiveeven at low magnification);++++
tumors and intestinal cells of Cajal [82]. Some of t-ICC in our experiment expressed nestin (Fig. 37).
IHC for desmin showed the lack of expressionat the level of t-ICC in lamina propria (Fig. 38A),concomitantly with immunopositive reactions insmooth muscle cells of either the circular layer (Fig.38B) or longitudinal layer (Fig.38C). Conversely,in vitro, in primary culture, IICC (identified byc-kit positivity) expressed desmin (Fig. 39).
IHC for vimentin labeled I-ICC (Fie. 40A). Doubleimmunostaining vimentin+ CDll7/c-kit (Fig. aOB)
demonstates that, indeed I-ICC express vimentin. IFimages like in Fig. 4l strongly support such findings.Torihashret ol. [83] andDuquette et al.l36lconsideredthat vimentin could be a marker for Cajal-like cells (inthe absence of a positive reaction for c-kit).
IF double-labeling of CDll7/c-kit and cave-olin-l and caveolin-2 in human fallopian tube pri-mary cultures showed both types of caveolar pro-teins expressed by IICC (superimpose Fig.24A,Bon Fig. 26). Cho and Daniel [78] reported coexpres-sion of caveolin-l and c-kit in ICC and intestinalsmooth muscle, but our data add some detail oncaveolin-l and caveolin-2 cellular distribution:caveolae were more abundant at the level of cellprocesses of t-ICC, as seen in both TEM (e.g. Fig.17A) and fluorescence microscopy.
IHC with antibodies against some additionalantigens showed negative immunostaining forCDla (Fig.42A), CD62P (Fig. 42B), CD6S (Fig.
Fig. 31 Correlation between CDllT/c-kit and CD34expression on t-ICC. Serial fallopian tube sections wereassessed for either CD34 or CDllT/c-kit expression byIHC. Positive cells within lamina propria and musculariswere counted and results expressed as percentage ofthetotal number of cells with t-ICC morphology.
y:O.7914x+0.1793
R2 : O.76a2, r : 0.87, p - 0.001, n : 9
508

M#
s-100
Fig. 32 A, B Human fallopian tube. Some t-ICC are5-100 immunoreactive (arrows). DAB (brown) wasused to visualise the immune reaction. Counterstainingwith Mayer's hematoxylin. Original magnification,100x.
43), GFAP (Fig.44), NSE (Fig.45), chromograninA (Fig. 464), PGP 9.5 (Fig. 47A). These results sup-port distinction between I-ICC and other cell types(pericytes, macrophages, mast cells, smooth musclefibers, neural cells, neuroendocrine cells, etc.) [84].
Collateral findings:
Neuroendocrine scattered cells ('JF')
We describe here cells located among smooth mus-cle cells, identified as positive for chromo-
J. Cell. MoL Med. Vol9, No 2, 2005
Fig. 33 d B Human fallopian tube. A. a SMA immuno-staining shows non-reactive t-ICC in the lamina propria(a:row). B. Note the two cr SMA-negative spindle-shaped interstitial cells lining the circular muscle layer(CM) (arows). Counterstaining with Mayels hema-toxylin. Original magnification, 100x.
granin A (Fig.46B,C) or for PGP9.5 (Fig. a7B).In situ, such cells have a different morphology thant-ICC. As expected, I-ICC were not immunoreac-tive for both chromogranin A and PGP9.5 (Figs.46A and,47 A).
Because of chromogranin A and PGP9.5 positiv-iry we suggest that these cells scattered amongsmooth muscle cells of fallopian tube wall couldbelong to a diffirse neuroendocrine system [85, 86].We refer to these possible neuroendocrine cells as
'JF cells'.
|#,'aa
:i*i:::mi.:
'.t... .
'. sth "I irr
"1i'rrl' ' -r-
k' s:1*9Q
509

" ;,widlWW'
s-100 & sMA
. . .. fifo,
,r rlil '"ile
Fig. 34 A{ Dual immunohistochemical staining for5-100, brown (DAB) and SMA, red (Fast red) of thehuman fallopian tube lamina propria (A) and muscu-laris (B, C). Note the presence of t-ICC expressing both5-100 & SMA in lamina propria (black arrow), whilemuscularis t-ICC expressed either 5-100 (black anow-heads) or SMA (green arrows). The longitudinal musclelayer (LM) stains for SMA. The image shown in B is aphotographic reconstruction. Original magnification,100x.
Very recently, Dursun et al. reported a case ofprimary neuroendocrine carcinoma of the humanfallopian tube, apparently for the first time [87].Although they noted Chromogranin A focally posi-
#
S-100 & SMA;rr# #,
'* *."#l,dl,4fin
tive tumoral cells, by IHC, no explanation for theorigin of this carcinoma was provided. We suggestthat 'JF' neuroendocrine cells identified hereincould represent the starting point for this tumor.
*S tt:''' c
.rJ \ "'
Pffidlil'*1e

J. Cell. Mol. Med. Vol9, No 2, 2005
Fig. 35 Human fallopian tube; primary cultures (day 5). Double immunostaining for smooth muscle cr-actin (green)and CDllT lc-kit (red). The cell body of an ICC in the lower part of the image, shows SMA positivity, resemblingstress fibers. The cell nucleus stains blue with Hoechst 33342. Original magnification, 60x.
Fig.36 NK-l immunostaining of normal human fal-lopian tube. Arrows indicate triangular cells with thinprocesses, immunoreactive for NK-l; the double arrowpoints out long cell processes. Counterstaining withMayer's hematoxylin. Original magnification, 100x.
Electrophysiology
The aim of the extracellular single unit recordings wasto determine whether t-ICC in primary culture exhibitor not electrical activity. ln our experiments, sustainedspontaneous electrical activity was recorded in most ofthe investigated cells. Field potentials averaged57 .26 + 6.56 mV in amplitude (n : 30), without show-ing a rhythmical pattem (Fig. a8). The potentials wererather short (0.98 + 0.07 ms) when compared to theelectrophysiological properlies of smooth muscle cells[88]. These preliminary results show that t-ICC in sub-confluent primaly culture have spontaneous electricalactivity. Since the single unit recording were done inprimary cell cultures, derived from muscularis andlamina propria, it was not possible to establish theexact origin of the examined I-ICC.
Discussion
Identification of Cajal-like interstitial cells:methods and criteria
We report here the identification of a novel Cajal-like interstitial cell (t-ICC) in the non-epithelialspace of human fallopian-tube wall (Fig. a9).These cells are similar to archetypal enteric cellsof Cajal (ICC).
#
:*!
'.., :':l'
'-- "- *
**-s=B€.8*
;# {E E*-" / ,u *
:b€*F %- *: &'*-t-r3+*
,/ #h..
q&e -: ***ry'* ti*,'k ;rla}t
)/^/"9..& &''%d NK'l
&
511

d€i;a* '
-.:;:,i*t:i9,:a1 '
: t.?i:rnestin
Fig. 37 A-C Human fallopian tube, immunostainingfor nestin. A, B. In lamina propria, I-ICC show acytoplasmic pattern of nestin expression (arrows).C. A t-ICC, located in the loose connective tissueundemeath of serosao exhibits long, nestin immuno-positive processes. Original magnifications 40x (A)and l00x (B, C).
To establish the 'affiliation' to the (heteroge-nous) group of Cajal-like interstitial cells in a giventissue, we suggest the following algorithm:
I. IdentificationA. Optical microscopy
a. (Supra)vital methylene-blue stainingb. Toluidine-blue staining of semi-thin
sections of Epon-embedded tissue (afterglutaraldehyde/osmium fixation)NCLM
c. CDllTlc-kit immunodetection - IHCand/or IF
B. Transmission electron microscopy (TEM).II. Characterization
A. Cell culturesB. Cellular electrophysiologyC. Biochemical and molecular analyses
A.a. The supravital methylene blue staining('en bloque') and/or vital staining (cryosections)offer an indication on the possible existence ofCajal-like cells in the respective tissue. We have toremind here that, at the end of the 19th centuryRamon y Cajal described the cells which eventual-ly received his name, right by using methylene-blue staining.
5t2
nestin
A.b. NCLM points out the topography, the rel-ative density and the relationships of Cajal-likeinterstitial cells with other microscopic structures(e.g. Fig.49). Moreover, NCLM discloses the lim-ited diversity of cell-body shapes, and, especially,the morphology of the characteristic cytoplasmicprocesses. Briefly, NCLM gives presumptivediagnosis.
A.c. Positive IHC and/or IF for CDllTlc-kit,together with the two above-criteria, give an accu-rate diagnosis.
B. TEM, when available, gives certitudediagnosis.
*For the identification of Cajal-like interstitial cells
we considered the following set of criteria, relying ontopographical, morphological and antigenic atffibutesthat exclude, quite accurately, other cell types:
:-i:

J. Cell. Mol. Med. Vol9, No 2, 2005
Fig. 38 A-C Human fallopian tube stained for desmin and counterstained with Mayer's hematoxylin(cross-sectioned). Putative t-ICC in lamina propria (A, arrow) lacks desmin expression, while smooth musclecells in the circular (B) and longitudinal (C) muscle layers are desmin-immunoreactive. Original magnification100x. CM : circular muscle layer; LM : longitudinal muscle layer.
r' #I' $- ^$o":P'n
Location (Fig. a9) in interstitium (non-epithelial space); frequently join adjacenthistologic structures (smooth muscle cells,nerves, small blood vessels or epithelia) vlatheirCharacteristic processes, andCDll7/c-kit positivity (although we cannotexclude that sometimes Cajal and Cajal-likecells, that respect the first couple of criteria,might test negative for this marker).
Obviously, vital staining with methylene blue(relying on poorly understood molecular mecha-nisms) has more than historical significance, since,for instance, it reveals Cajal-like interstitial cells inmyometrium [37] and mlmmary gland [45].
To facilitate the certitude diagnosis by electronmicroscopy, we propose the following set of ultra-structural criteria ('platinum standard') for Cajal-like interstitial cells:
desmin
Fig. 39 Human fallopian tube; preconfluent t-ICC primary culture; immunostaining for desmin and CD ll7lc-kir.The cell is positive for both desmin (green) and CD 117 lc-kit (red). Nuclear counterstaining with Hoech st 33342(blue), original magnification 60x. Compare with the same cell inFig.24 C.
513

#.,jtru
ffi*'sM*, rT*$I.aqilry- lt
* *{s.]n*l
'.*.r,r. ::.:.
"**
' 'k..., i'I&f 'g!.
vimentin' $tq': *r
Fig. 40 d B Human fallopian tube; IHC. Immunostaining for vimentin (A) and double immurrostaining for vimentinplusCDll7/c-kit(B). A.Notethevimentinimmunoreactivet-ICCprocesses(anows).B.Cellswithmorphologysug-gestive for I-ICC stain both for CDl17/c-kit (brown - DAB) and vimentin (red - Fast red), mainly at the level of theirlong, moniliform processes (anows). Counterstaining with Mayer's hematoxylin. Original magnification 100x.
Fig.41 Human fallopian tube; subconfluent primaryculture. Double immunofluorescent labeling of an'octopus'-like t-ICC: vimentin (green) and CD117/c-kit (red). Vimentin reactivify is mainly localized with-in the cell processes, and CDllTlc-kit has a patchypattern. Compare with Figs. 3C and 24D. The cellnucleus is shown in blue (Hoechst 33342); originalmagnification 60x.
5L4
4fi
1
1t;;
1. Location in the non-epithelial space;2. Close contacts with targets: nerve bun-
dles, and./or epithelia, and/or smooth mus-cle cells, and/or capillaries;
3. Characteristic cytoplasmic processes:i. Number (1-5, frequently: 2-3);ii. Length (tens up to hundreds of pm);iii. Thickness (uneven caliber, < 0.5 pm);iv. Aspect: monilform, usually with mito-
chondria in dilations;v. Presence of'Cz2+ release units';vi. Branching: dichotomous pattern;vii. Organizttion in network - labyrinth-
ic system: overlapping cytoplasmic pro-CCSSES;
4. Gap junctions: with smooth muscle cellsor with each other;
5. Basal lamina: occasionally present;6. Caveolae: 2-4Yo of cytoplasmic volume;
-0.5 caveolae / pm of cell membranelength;
7. Mitochondria: 5-I0o/o of cytoplasmic vol-ume;
8. Endoplasmic reticulum: about l-2o ,
either smooth or rough;9. Cytoskeleton: intermediate and thin fila-
ments, as well as microtubules, present;10. Myosin thick filaments: undetectable.
" .**;:r
<.;,. 'vimentin & c-kit

J. Cell. Mol. Med. Vol9, No 2, 2005
Hffi
&.xFig. 42 A-C Human fallopian tube A, B. No positiveimmune reaction of cells in lamina propria for CDla andCD62P. C. IF using FlTC-conjugated anti-CDla mono-clonal antibodies. Arrows indicate cells CDla-immunoreactive in the human fallopian tube lamina pro-pria. HoweveE the lack of processes and an overallround shape suggest a cell type different from t-ICC.Original magnification: 40x (A, B); 20x (C).
ffiry {r' s
ry& ....
**''..%.@ %cD68
Fig. 43 IHC for CD68 distribution in human fallopian-tttbe lamina propria. The only immunoreactive cells (*)are subepithelial macrophages (mast cells ?), while therecognizable t-ICC do not bind the antibody. Mayer,shematoxylin counterstaining; original magnification 100x.
3-,*a:q -
Fig. 44 Human fallopian tube lamina propria stainedfor GFAP. Cells with morphology suggestive for t-ICCare negative for GFAP. Mayer's hematoxylin counter-staining; original magnification 100x.
g:..*
tMsL*,
,. ,?q,L_""q! -.a
_ + -ai- *d^-+t
NSE
Fig. 45 Human fallopian tube labeled for NSE (photographic reconstruction). Larnina propria I-ICC lack NSEimmunoreactivify. Original magnification 1 00x.
"-*'.ry.{
x.d&
.i'- {
,LA'% * ,"*&
515

.iie
*{'h ?
tr5 *tr *'.dt
s ''Y"*tr
F.
-,,$l#qs;r l-amina:
propr-{a:r
&
f# *
chromogranin A^&:.il
#
Museularis
nh*.*-
chromogranin A
Fig. 46 A{ Human fallopian tube immunostained for chromogranin A. A. No chromogranin A positive ce1ls areseen in lamina propria. B, C. Several chromogranin A-positive granules can be seen (arrowheads) in the muscu-lar layer. Original magnihcation, 100x.
i:
%"*"uiff
ii
s'UE *:*s*.:5a
:u ' 'a-i"tr',lc g
f,@,&lc#.s
Fig. 47 A, B. Immu-nohistochemical de-tection of PGP9.5 inhuman fallopian tube.A. No staining is seenin the lamina pro-pria. B. Immunore-active cells are detect-ed in the muscularlayer, among the smo-oth muscle hbers.Original magnifica-tion 100x.
,*# Mu,
;{b.^r"'
n a*5-
"tr'Fr"l #
Itlt"' ';', ,il11:
'- *1$:''.' {..ch*romogranin ij &._*.,*ffi .
-g*r.:.,::;
516
PGP 9.5

J. Cell. MoL Med. Vol9, No 2, 2005
Fig. 48 A-D. A' B. Extracellular single unit recordings display sustained spontaneous eleckical activity in t-ICC.C. Spontaneous field potential pattem. D. Phase conhast microscopy (40x) showing the recording glass electrode inthe proximity of the I-ICC body.
ln our opinion, the first six criteria of the 'plat-inum standord' could be sufficient to diagnose aninterstitial (Cajal-like) cell.
t.ICC:Uncommitted (bipotential) progenitor cell
Markers traditionally assigned to somatic stem cells(hematopoietic, neural, etc) have been found in ICCsubpopulations: CD1l7 122,231, CD34 [24], nestin
[89], vimentin [90], the embryonic form of myosinheavy chain [91]. This could rise the questionwhether interstitial Cajal-like cells are terminally dif-ferentiated or not.
Cells retaining their self-renewal capacrty and abilityto give rise to qpecializedprogenywithin a specific tissue
can be considered somatic stem cells [92]. Apparently,ICC and Cajal-like cells match this description.
It seems that ICC are (not post-) mitotic cells: rnvilro studies showed that they undergo stem cell fac-tor-dependent proliferation 193, 941; preventing c-kitsignaling leads to their depletion in experimental [95]and pathologic [96] circumstances; GIST are linkedto monoclonal, abnorrnal ICC proliferation [97].
Cajal cells are able to switch phenotype, since they(trans?)differentiate into smooth muscle cells, when c-kit signaling is blocked [98]. Moreover, their malig-nant counterparts in GIST occasionally express eithersmooth muscle [99] orneuroendocrine [100] marken.
Canonical ICC and smooth muscle cells arisefrom a common c-kit-positive mesenchymal progen-itor. The precursor cell expresses vimentin and shares
ultrastructural features with the *adulf' cell type [95,
5r7

981. I-ICC described here might belong to two (possi-bly overlapping) subpopulations: one endowed withspontaneous electrical activity, closer to enteric ICC,and the other, less similar to archetypal ICC, closer toprogenitor cells. It remains to be seen whether func-tionally dissimilar Cajal-like subpopulations havedistinct locations. Moreover, no experimental evi-dence excluding the possibility that CajalJike cellsmight become a typical ICC. From this point of view,Cajal-like cells could mean more than simply ICC.
To summarize, Cajal-like cells are an amazinglyheterogenous population that could retain differentia-tion potential or, at least, be able to switch phenotypeunder norrnal or pathologic circumstances.
t.ICC:An immune surveillance role?
Yamazaki and Eyden described in human fallopiantube a network of CD34-positive interstitial cells withaputative immune surveillance role [101]. CDll7/c-kitexpression was not investigated, making it difficult todetermine the relationship between the CD34-posi-tive cell network and CDI 17 positive I-ICC.
One may expect a partial overlap of the two celltJpes, as some enteric ICC U02l and some Cajal-likecells [46, 90, 103] are positive for CD34 and, to a cer-tain extent, look similar. However, it seems reason-able to assume important differences between the twocell types, as their proportions in the whole cell pop-ulation are significantly dissimilar [101]. Differencescan be also found when looking at the origins of thesecells: CD34 positive fibroblasts might have a blood-bome precursor [104] tlrat crosses endothelia, whereasICC and Cajal-like cells are more likely to arise fromtissue-resident mesenchymal progenitor cells [105].
Arguments that apparently favor a role for any ofthe two cell types in immune surveillance are thepresence of close contacts that might play a role intransfering antigenic peptides to antigen presentingcells for cross-priming [06] (seen here in Fig.22A),and the shategical location of t-ICC at the cross-roadsof various histologic compartments, some in directcontact with the environment. Interactions between t-ICC and cells of the innate immune system (macro-phages -Fig.221\) could represent another indicationof their involvment in organism defense.
On the other hand, the I-ICC lack of markers usu-ally associated with immune functions (e.g. CDla,CD62P, CD68), as well as the lack of structures that
518
have a role in antigen processing and presentation,apparently question the immune surveillance view-point. Nevertheless, further studies are required todescribe the cellular and molecular mechanisms thatmight b€ involved.
As shown in Fig. 49,the stroma of lamina propriain the human fallopian tube is mainly composed of aCajal-like cell reticulum. A subepithelial meshworkof Cajal-like cells (c-kit positive, CD34 ?) and CD34-positive (c-kit ?) undifferentiated cells seems to markthe epithelial-stromal border. However, the sole pres-ence of such a network do not ascribe it an immunesurveillance role.
t.ICC:Networks for pacemaking
Apacemaker role is now generally accepted for enter-ic ICC, which generate electrical rhyttrmicity in thegastrointestinal f::act 122,23,107). Why should notIICC be responsible for the human fallopian tubeperistalsis ? The more so as, Yoneda et al. [708]reported that, in mouse proximal colon, the submu-cosal c-kit-positive bipolar cells spontaneously gener-ate plateau potentials with rhythms different fromthose generated by smooth muscle cells.
In tenns of physiological evidence, our electro-physiological results suggest that I-ICC could possesselectrical properties that do not rule out a presumptivepacemaker role. Two lmportant prerequisites are firl-filled, as concems t-ICC as pacemaking network: a)the existence of numerous gap junctions (cf. [109]and Figs. 19, 20 and 228\, and b) integrated Ca2+handling by endoplasmic reticulum and mitochondria[107], in other words, the finding of 'C*+ releaseunits'(Fig 23B,C), which include caveolae too. Thus,our data correlate well with recent studies, on intesti-nal smooth muscle, desiped to examine the relation-shiF between caveolin-l, Ca2+-handling and ICC pac-ing [78]. Nevertheless, in vitro studies may not beexactly applicable to intact ICC networks [110].
t.ICC:Possible regulators of neurotransmission andintercellular signaling
By analogy with the role ascribed to enteric ICC177,201, there is no reason to overlook the possibleparticipation of t-ICC in regulating neurotransmis-

*t'l:t
*;a
f. t,',
*,:::::.
*.::, l'
*:tt'*
re-tI-
-?tIII
tlA-f!I
J. Cell. Mol. Med. Vol9, No 2, 2005
NO}{.EPITIIELIAL SPACT-8?n
tubal-ICC{like}
lo 20S!IJSCULA*I5
Fig. 49 Schematic drawing (not to scale): cross section of the human fallopian tube wall, ampular region. Numbersrepresent the relative density of I-ICC in a given area (percentage of t-ICC out of all cells seen in that area). Datawere obtained mainly by NCLM (semi-thin sections stained with toluidine-blue): 4903 cells - from 82 photographs,randomly taken, were counted. t-ICC are located between the mucosal epithelium (left) and the serosal mesotheli-um (right). In fact, t-ICC are residents in lamina propria and in muscularis (circular myocytes are cut longitudinal-ly and longitudinal myocytes are cut transversally). Note the gradient of \-ICC relative densir): from -l8o^, imme-diately undemeath of mucosal epithelium, towards -7.5Yo inmuscularis. Close spatial relationships of I-ICC meshwith nerve bundles and./or capillaries are also depicted.
'tt .
100Fm
20
sion. At the level of human fallopian tube, besidesthe well-known contractile effect of acetylcholine[111], the possible modulation of adrenergic neuro-transmission by endogenous purines was reported
[112], and purinergic receptors described [113].Moreover, the involvement of NO/cGMPpathway incontrol of human fallopian tube contactility wasestablished [114,115]. The strategical location oft-ICC, across the fallopian tube wall, shown in Fig.49, suggests the speculation that vla secretory mech-anisms, these Cajal-like cells are players in a subtleand redundant game. I-ICC have at least tlree ways
to regulate the activity of neighboring cells: auto-,juxta- and para-crine. Anyway, the juxtacrine mech-anisms are supported by TEM findings (Figs. 19, 20and22).
ln our opinion, a special place in fallopian tubephysiology should be occupied by the close relation-ships between the subepithelial I-ICC network andthe adjacent epithelium. Presumably, such relation-ships are established by secretory paracrine mecha-nisms, and we can imagine one-way or 'round trip'molecular haffic. The regulation of oviductin secre-tion [116] might be an example.
r8%
519

t.ICC:Possible players in tissue remodeling
Extacellular manix and certain cell types (e.g. myo-fibroblasg,'distant relatives' of interstitial Cajal-like)are incriminated recently in tissue remodeling tl lT-1191.Moreover, vascularremodeling seems to be, at least inpart, due to a phenotypic conversion, reminiscent of afibroblast developmental program [120]). The noveltype of Cajat-like cells described here might partici-pate in remodeling human fallopian tube tissues,where inflamrnation or infection recrurently occur.One may speculate that the differential expression ofcertain antigens on t-ICC found in our study could bethe result of such a phenotypic versatility.
Instead of conclusions
The gap between the clinical progress supportingreproductive technologies U}l,l2zl and the almostobsolete information available on molecular histologyand physiology of human fallopian tube is surprisingand challenging.
Acknowledgements
This study was partially supported by tlre grant VIASAN03911200/. from the National Ministry of Education andResearch. The authors gratefirlly thank to the staff ofDiagnostic Immunohistochemisty Uni!'Victor Babes'
National Institfie of Patholory, Nfis. Maria Buzatr , N{rs. A.Bungardean" Mr. T Regalia for expert technical assisance, Dr.A.C. Popescu for his help with Fig. 49, and Dr. Mircea kabUfor useful suggestions and critical rrading of the manuscript.
References
l. Orci L, Pepper MS. Microscopy: an arf!. Nat Rev Mol CellBiol.20O2;3: 133-7 .
2. Cajat SR Sur les ganglions et plexus nerverx de I'intestin.C.R. Soc Biol (Paris). 1893; 45:217-23.
3. Cajal SR Les nouvelles idees sur la structure du systdmeneryeux chez I'homme et chez les vert6bres. Pmis: C.Reinwald & Cie; 1895.
4. Min Kg Seo IS.Interstitial cells ofCajal in the human smallintestine: immunochemical and ultrastructural study.U tras truct Pat ho l. 2003 ; 27 : 67 -7 8.
520
ThunebergL. One hundredyean ofinterstitial cells of CajalJuficrosc Res Tech. 1999;47: 223-38.Vajda J, Feher E. Diskibution and fine structwe of the inter-stitial cells of Cajal. Acta Morphol Acad Sci Hung.1980; 28:251-8.Faussone Pellegrini MS, Cortesini C. Ultrastuctural fea-tures and localization of the interstitial cells of Cajal in thesmooth muscle coat of human esophagus, J SubmicroscCytol. 1985; 17: 187-97.Faussone-Pellegrini MS, Pantalone D, Cortesini C.Smooth muscle cells, intorstitial cells of Cajal and myentericplexus interrelationships in the human colon. Acta Anat(B as el). 1990; 139: 3 | 44.Rumessen JJ, Peters S, Thuneberg L. Light- and elechonmicroscopical studies of interstitial cells of Cajal (ICC) andmuscle cells at the submucosal border of human colot. LabInves t. 1993 ; 68: 48 I -95Rumessen JJ. Identification of interstitial cells of Cajal.Significance for studies of human small intestine and colon.Dan Med Bull. 1994; 4l:275-93.Hagger \ Gharaie S, Finlayson C, Kumar D. Dstibutionof the interstitial cells of Cajal in the human anorectum. ./Auton Nerv Syst. 1998; 73: 75-9.Thuneberg L, Rumessen JJ, Mikkelsen HB, peters Sn
Jessen H. Structuml aspects of intemtital cells of Cajal as
intestinal pacemaker cells, In: Huizinga JD, editor.Pacemaker activity and intercellular communication. BocaRaton: CRC Press; 1995. p.193-222.Komuro T, Seki K Horiguchi tr( Ulhasfirctural character-ization of the interstitial cells of Cajal. Arch Histol Cytol.1999;62:295-316.Faussone Pellegrini MS, Thuneberg L. Guide to the identi-fication of interstitial cells of Cajal. Mieosc Res Tech. 1999;47:248-66.Iluizinga JD, Thuneberg L, Vanderwinden JM,Rumessen il. Interstitial cells of Cajal as targets for phar-macological intervention in gashointostinal motor disorders.Trends Pharmacol fti. 1997;18: 393-403Daniel EE, Berezin I. lnterstitial cells of Cajal are they majorplayer in conhol of gashointestinal motility? J GastrointestMotil. 1992;4: l-24.Sanders KM. A case for interstitial cells of Cajal irs p.rce-
makers and mediators of neurotansmission in the gashoin-testinal ract. Gastromterol. 1996; 111: 492-515Koh SD, Sanders KM, Ward SM. Spontaneous electricalrhythmicity in cultured interstitial cells of Cajal, isolatedfrom the murine small intestine. J Physiol (Lond). 1998;513:203-13.Thomsen L, Robinson TL, Lee JCF, Farraway LdHughes MJG, Andrews DW, Huizinga JD. Interstitial cellsof Cajal generate a rhythmic pacem*er clrrent. Nature Med.1998;4: 848-51.
Ilirst GD, Ward SM. Interstitial cells: involvement in rhyth-micity and neural control of gut smooth mvscle. J Pltysiol.2@3:550:33746.Maeda II, Yamagata d Nishikawa S, Yoshinaga ItKobayashi S, Nishi tri Nishikawa SI. Requirement of c-kitfor development of intestinal pacemaker system.D evelopment. 1992; | | 6: 369 -7 5
Ward SM, Burns AJ, Torihashi S, Sanders KM. Mutationof the proto-oncogene c-kit blocks development of interstitial
5.
6.
7,
10.
11.
12.
9.
13.
15.
16.
t7.
18.
t9.
2r.

cells and elechical rhythmicity in murine intestine . J physiot.
1994; 480l. 9l-7.Huizinga JD, Thuneberg L, Kluppel M, Malysz J,Mckkelsen HB, Bernstein A. W&it gene required for inter-stitial cells of Cajal and for intestinal p:rcemaker activity.Nanre. 1995 ; 37 3 : 347 -9.Hirota S, Isozaki tri Moriyama Y, Hashimoto lt NishidaT, Ishiguro S, Kawano li Hanada N, Kurata An TakedaM, Muhammad TG, Matsuzawa Y, Kanakura y,Shinomura Y, Kitamura Y. Gain-of-function mutations ofc-kit in human gasfointestinal stromal t.ttnors. Scimce.1998;279: 577-80Burton LD, Ifousley GD, Salih SG, Jaeenwood D.P2X2receptor expression by interstitial cells ofCajal in vasdeferens implicated in semen emission. Auton Neurosci BasicClin. 2O00; M: 147 -61 .
Sergeant GP, Hollywood MA, McCloskey KD,Thornbury KD, Mclfale NG. Specialized pacemaking cellsin the mbbit urethra- J Physiol. 2000; 526l.35946.Sergeant GP, Ilollywood MA, McCloskey KD, McHaleNG, Thornbury KD. Role of IP(3) in modulation of sponta-neous activity in pacemaker cells of nbbit wethru Am JPhysial Cell Physiol. 2001; 280: C1349-56.Johnston L, Sergeant GP, Ilollywood MA, ThornburyKD, McHale NG. Calcium oscillations in interstitial cells ofthe rabbit urethra. J Plrysiol. 2005; 565: 449-61.McCloskey KD, Gurney AM. Kit positive cells in theguinea-pig bladder. J Urol. 2002;168:832-6.Blyweert W Aa F, Ost D, Stagnaro M Ridder D.Interstitial cells of the bladder: the missing link? 8,,/OG.20M;1 I l: 57-60.Ilashitani II, Yanai Y, Suzuld H. Role of interstitial cellsand gap junctions in the hansmission of spontaneous Ca2+signals in detrusor smooth muscles of the guinea-pig urinarybladder. J Pfusiol. 2004; 559: 567-8 l.Gillespie JI, Markerink-van Ittersum M, de Vente J.cGMP-generating cells in the bladder wall: identification ofdistinct networks of interstitial cells. BJtl Int. 2004; 94ttt4-24.Davidson RAo McCloskey KD. Morphology and IocaLiza-tion of interstitial cells in the guinea pig bladder: structuralrelationships with smooth muscle and neurons. ./ fJrol.2005;173: 1385-90
PezoneMA, Watkins SC, AlberSM, KingWEnde GroatC[ Chancellor MB Fraser MO. Identification of c-kit-positive cells in the mouse ureter: the interstitial cells ofCajalofthe urinary tract. Am J Physiol Renal Plrysiol.2O03;284:925-9.Metger & Schuster T, Till H, Franke FE, Dietz HG.Cajal-like cells in the upper urinary tract: comparative studyin various species. Pediatr Surg Int. 2005;21: 169-74Duquette Rd Shmygol d Vaillant C, Mobasheri A, PopeM, Burdyga T, Wray S. Vimentin-positive, c-KlT-negativeinterstitial cells in human and rat uterus: A role in pacemak-ing? Biol Reprod. 2005; 72: 27 6-83.Ciontea MS, Radu E, Regalia T, Ceafalan L, Cretoiu D,Gherghiceanu M, Braga Rf, Malincenco M,Z,agreanL,Hinescu ME, Popescu LM. C-kit immunoposifive intersti-tial cells (Cajal-type) in human myometrium. J Cell MolMed.2005;9:407-20.
J. Cell. Mol. Med. Vol9, No 2, 2005
Pucovsky V, Moss RF, Bolton TB. Non-contactile cellswith thin processes resembling interstitial cells of Cajal foundin the wall of guinea-pig mesenteri c afteies. J physiol . 2003;552:11943.Harhun MI, Gordienko DV, Povstyan OV, Moss RF,Bolton TB. Function of interstitial cells of Cajal in the rabbitportal vein. Circ Res. 20M; 95: 619-26.Harhun MI, Pucovsky V, Povstyan OV, Gordienko DV,Bolton TB. Interstitial cells in the vasculature. J Cell MotMed. 2005; 9: 23243.McCloskey KD, Hollywood Md Thornbury KD, WardSM, McIIale NG. Kit-like immunopositive cells in sheepmesenteric lymphatic vessels. Cell Tissue Res.2002; 310: 77 -84.
Exintaris B, Klemm MF, Lang RJ. Spontaneous slow waveand conhactile activity of the guinea pig prostate. JUrol.2002;168:315-22.Van der Aa F, Rmkams T, Blyweert W, De Ridder D.Interstitial cells in the human prostate: a new therapeutic tar-gel? Prostate. 2@3; 56: 250-5.Ilashitani II, Suzuki H. Identification ofinterstitial cells ofCajal in corporal tissues of the guinea-pig penis. Br JPharmacol. 2004: I 4 I : 199 -204.Popescu LM, Andrei F, Ilinescu ME. Snapshots of mam-mary gland interstitial cells: methylene blue vital staining andc-kit immunopositlity. J Cell Mol Med. 2005;9: 476-7 .
Popescu LM, Ilinescu ME, Ionescu N, Ciontea MS,Cretoiu D, Ardeleanu C. Interstitial cells of Cajal in pan-creas. J Cell Mol Med.2005;9: 169-90.Hendrickson MR, Kempson RL. Normal histology, of theuterus and fallopian tubes. In: Stemberg SS, editor. Histologyfor pathologists, 2nd edition. Philadelphia: Lippincott-Raven;1997.p.879-928.Hunt JL, Lynn A.d Histologic features of surgicallyremoved fallopian flfres. Arch Pathol Lab Med.2002; 126:951-5.
Hunter RIL Have the Fallopian tubes a vital role in promot-ngfertrhty? Acta Obstet Gynecol Scand. l99B;'77:475-86.Leere HJ, Tay JI, Reischl J, Downing SJ. Formation ofFallopian tubal fluid: role of a neglected epithelium.Reproduction. 2001 ; 12 I : 33946.Arbab F, Goldsby J, Matijevic-Aleksic N, Huang G, RuanKII lluang JC. Prostacyclin is an autocrine regulator in theconbaction ofoviductal smooth muscle. Hum Reprod.2002;l7: 3053-9.
Morimoto T, Head JR, MacDonald PC, Casey ML.Thrombospondin-l expression in human myomehium beforeand during pregnancy, before and during labor, and in humanmyometial cells in culture. Biol Reprod. 19981,59:862-70.Niculescu I. An atlas conceming morphological aspects ofvisceral nerve endings. lst ed. Bucharest: Editura Medicala;1958.
Yack JE. Janus Green B as a rapid, vital stain for pheripher-al nerves and chordotonal organs in insects. "/ NanrosciMethods. 193; 49: 17 -22.
Keij JF, Bell-Prince C, Steinkamp JA. Staining of mito-chondrial membranes with l0-nonyl acridine orange,MitoFluor Greerl and MitoTracker Green is affected bymito-chondrial membrane potential altering drugs. Cytomew.2000:39:203-10.
38.
39.
40.
4t.
23.
24.
42.
43.
44.
45.
46.
47.
25.
26.
27.
28.
29.
30.
31.
34.
48.
49.
50.
51.
32.
33.
52.
53.
54.
35.
36.
3l- 55_
52r

74.
75.
76.
77.
78.
79.
80.
56.
57.
58.
59.
60.
61.
81.
82.
83.
84.
85.
62.
63.
64.
65.
66.
67.
68.
86.
87.
88.
89.
69.
70.
71.
Vanden Berghe P, Ilennig GW, Smith TI( Characteristicsof intermittent mitochondrial transport in guinea pig entericnerve fibers. Am J Pltysiol Gastrointest Liver Physiol.2OO4;286: G67l-82.Bancroft JD, Gamble M, editors. Theory and practice ofhistological techniques. 5th ed. London: ChurchillLivingstone; 2002.Weibel ER Practical methods for biological morphomehy.Stereological methods. London: Academic hess; 1979.Hsu SM, Raine L, Fanger H. Use of avidin-biotin-perox!dase complex (ABC) in immwroperoxidase techniques: acomparison between ABC and unlabeled antibody @AP) pro-cedures. -/I/rstochem Cytochem. l98L; 29: 577 -80.Bussolati G, Gugliotta P. Nonspecific staining of mast cellsby avidin-biotin-peroxidase complexes (ABC). J HistochemCytochem. 1983; 31: l4l9-21.Alberti E, Mikkelsen HB, Lar.sen JO, Jimenez M. Motilitypattems and distribution of intentitial cells of Cajal and nih-ergic neurons in the proximal, mid- and distal-colon ofthe rat.Neurogastromterol Motil. 2005; l7 : 13347.Lazarow d Cooperstein SJ. Studies on the enzymatic basisfor the Janus green B staining reaction. J HistochemCytochem. 1953; l: 23441.Rumessen JJ, Vanderwinden JM. Intentitial cells in themusculature ofthe gastrointestinal hact: Cajal and beyond.Int Rev Cytol.2003;229:1 15-208.Ifuizinga JD, Faussone-Pellegrini MS. About the presenceofinterstitial cells ofCajal outside the musculahne ofthe gas-hointestinal tract. J Cell Mol Med.2005;9:468-73.Popescu LM, Diculescu I, Zelck U, Ionescu N.Ultastructural distibution of calcium in smooth muscle cellsof guinea-pig taenia coli. A conelated electron microscopicand quantitative stuly. Cell Tissue Res. 1974;154:357-78.Brinsfeld TH, Fisher M, Hawk IfW. Ultrastructure ofendometial stromal cells in the ewe during the estous cycleand early pregnancy . J Anim Sci. 1974;39: 565-74.Popescu LM. A conceptual model of the excitation-conhac-tion coupling in smooth muscle: the possible role of the sur-face microvesicles. Studia Biophys. 1974;44: l4l-53.Moore ED, Voigt T, Kobayashi YM, Isenberg G, Fay FgGallitelli MF, Franzini-Armstrong C. Organization of Ca2+release units in excitable smooth muscle of the guinea-pigurinary bladder. Bioplrys J. 20M; 87: 1836-47 .
Schurch W' Seemayer TA, Gabbiani G. Myofibroblast. In:Sternberg SS, editor. Histology for pathologists, 2nd edition.Philadelphia: Lippincott-Raven; 1997 . p. 129-65.Wheaton tri Sampsel tri Boisvert Fl\d, Davy d Robbins S,Riabowol Ii Loss of functional caveolae during senescence
of human fibroblasts. J Cell Physiol.2Dl; 187:226-35.Crpona F, Cohen AW, Cheung MW, Sotgia F, Schubert
I Battista M, Lee II, FrankPG, Lisanti MP. Muscle-spe-cific interaction of caveolin isoforms: differential complexformation between caveolins in fibroblastic vs. muscle cells.Am J Plrysiol Cell Physiol.2005; 288: C677-91.Schurch W' Seemayer Td Gabbiani G. The myofibroblast:a quarter century after its discovery. Am J Surg Pathol. 1998:22: l4l-7.Eyden B. The myofibroblast: an assessment of conhovenialissues and a definition usefirl in diagnosis and research.
Wtrastruct Pathol. 2001; 25: 39-50.
Powell D[ Mifrlin RC, Valentich JD, Crowe SE, SaadaJI, West AB. Myofibroblasts. I. Paracrine cells important inhealth and disease. Am J Physiol. 1999;277: Cl-9.Walter I. Myofibroblasts in the mucosal layer of the uterinettbe. Ial J Anat Embryol. 19981'103:259-66.Lee II, DouglasJones AG, Morgan JM, Jasani B. Theeffect of fixation and processing on the sensitivity of oesto-gen receptor assay by immunohistochemistry in breast carci-noma- J Clin Pathol.2@2;55:236-8.Torihashi S, Horisawa l\4, Watanabe Y. c-Kit immunoreac-tive interstitial cells in the human gashointestinal nact. "/AutonNerv Sysl. 1999; 75: 38-50.Cho WJ, Daniel EE. Proteins of interstitial cells of Cajal andintestinal smooth muscle, colocalized with caveoLtn-!. Am JPhysiol Gastrointest Liver Physiol. 2005; 288: G57l-85.Puxeddu I, Piliponsky AM, Bachelet I, Levi-schaffer F.Mast cells in allergr and beyond- Int J Biochem Cell Biol.2003;35: 160l-7.Vanderwinden JM, Rumessen JJ, De Laet MH,Vanderhaeghen JJn Schifrmann SN. CD34+ cells in humanintestine are fibroblasts adjacent to, but distinct from, intersti-tial cells of Cajal. Lab lwest. 19991'79 59-65.Fina L, Molgaard IfV, Robertson D, Bradley NJ,Monaghan P, Delia D, Sutherland DR, Baker MA,Greaves MF. Expression of the CD34 gene in vasculaf,endothelial ce'L\s. Blood. l99O;75: 2417-26.Tsujimura T, Makiishi-Shimobayashi C, Lundkvist J,Lendahl U, Nakasho K Sugihara A, Iwasaki T,Mano M,Yamada N, Yamashita I( Toyosaka A, Terada N.Expression of the intermediate filament nestin in gashoin-testinal shomal tumon and interstitial cells of Cajal, Am JPathol. 2001; 158: 817-23.Torihashi $ Ward S1\rI, Sanders KM. Developmant ofc-Kit-positive cells and the onset of electrical rhythmicity inmurine small intestine. Gastroenterobgt. 1997 ; ll2: 144-55.Grider JR Focus on "molecular markers expressed in cul-h.red and freshly isolated interstitial cells of Cajal,,. Am JPhysiol Cell Physiol. 2000; 279: C2M-5.Vento P, Soinila S. Quantitative comparison of growth -associated protein GAP-43, neuron specific enolase and pro-tein gene product 9.5 as neuronal markers in mafure humanintestine. ./-Flrstochem Cytochem. 1999; 47 ; 1405-15.An S, Zenisek D. Regulation of exoclosis in neurons andneuroendocrine cells. Cun Opin Neurobiol.2004; 14: 522-30.
Dursun P, Selman MC, Taskiran C, Usubutun d AyhanA. Primary neuroendocrine carcinoma of the fallopian tube: acase rqorl. Am J Obstet Gynecol. 20M; 190: 568-71.Kuriyama II, Kitamura I! Itoh T, Inoue R Physiologicalfeafures of visceral smooth muscle cells, with special refer-ence to recepton and ion channels. Physiol Rev. 1998; 78:8 I l-920.Vanderwinden JM, Gillard tr! De Laet MIf, Messam CA,Schifrmann SN. Dstribution of the intermediate filamentnestin in the muscularis propria of the human gashointestinallract. Cell Tusue Res. 2002; 309: 261 -8.Ortiz-Hidalgo C, de Leon B, Albores-Saavedra J. Stromaltumor of the gallbladder with phenotype of interstitial cells ofCajal: a previously unrecognized neoplasm. Am J Surg.P atho I, 2000; 24 : I 420-3.
72.
73.
522
90.

9l . Sakurai S, Fukasawa T, Chong JM, Tanaka d FukayamaM. Embryonic form of smooth muscle myosin healy chain(SMemb/1r4HC-B) in gastrointestinal stromal tumor andinterstitial cells of Cajal. I m J Pathol. 1999; 154: 23-8.
92. RaffM. Adult stern cell plasticity: fact or artifact? Annu RevCell Dev Biol.2003; l9:l-22.
93. Epperson d llatton WJ, Callaghan B, Doherty P, WalkerRL, Sanders KM, Ward SM, Horowib B. Molecularmarkers expressed in cultured and freshly isolated interstitialcells of Cajal. Am J Physiol Cell Physiol.2000;279: C529-39.
94. Li CX, Liu BII, Tong WD, Zhtng LY, Jiang YP.Dissociation, culhre and morphologic changes of interstitialcells of Cajal rz vitro, World J Gastroenterol.2005; 1 1: 2838-40.
95. Torihashi $ Ward SM, Nishikawa S, Nishi K KobayashiS, Sanders KM. c-kit-dependent development of interstitialcells and electical activity in the murine gastrointestinal tract.Cell Tissue Res. 19951'280:97-lll.
96. Tong WD, Liu BII, Zhang LY, Xiong RP, Liu P, ZhangSB. Expression ofc-kit messenger ribonucleic acid and c-kitprotein in sigmoid colon of patients with slow transit consfi-panon. Int J Colorectal Dis.2005;20:363-7.
97. Isozaki li Terris B, Belghiti J, Schifrmann S, Hirota S,Vanderwinden JM. Germline-activating mutation in thekinase domain of KIT gene in familial gashointestinal stro-mal tumors. lz J Pathol.2000; 157: 1581-5.
98. Torihashi S, Nishi tri Tokutomi Y, Nishi T, Ward S,Sanders KM. Blockade of kit signaling induces tansdiffer-entiation of interstitial cells of Cajal to a smooth muscle phe-no$e. Gastroenterologt. 1999; l 17 : 140-8.
99. Sarlomo-Rikala M, Kovatich AJ, Barusevicius A,Mettinen M. CDI l7: a sensitive marker for gashointestinalsfromal tumors that is more specificthanCD34. Mod. Pathol.1998, ll 728-34.
100. Dhimes P, Lopez-Carreira M, Ortega-Serrano MP,Garcia-Munoz II, Martinez-Gonzalez MA" Ballestin C.Gashointestinal autonomic nerve tumows and their separa-
tion from other gastrointestinal stromal hrnours: an ultra-shuchral and immunohistochemical study of seven cases.Virchows Arch. 1995; 426:27-35.
101. Yamazaki I( Eyden BP. Ultrashuchml and immunohisto-chemical studies of stromal cells in lamina propria of humanfallopian hrbe ampullar mucosa: the recogrrition of 'CD34positive reticular network' and its putative function forimmune surveillance. J Submicrosc Cynl Pathol. 1995;28:325-37.
102. Sircar K Hewlett BR, Huizinga JD, Chorneyko triBerezin I, Riddell RH.Interstitial cells of Cajal as prccursors
of gashointestinal shomal tumors. Am J Surg Pmhol. 1999;23:377-89.
103. Piaseczna Pd Rolle U, Solari V, Puri P, Interstitial cells ofCajal in the human normal urinary bladder and in the bladderof patients with megacystis-microcolon intestinal hypoperi-stalsis syndrome. BJU Int.20M;94: 143-6.
104. Bucala R, Spiegel LA., Chesney J, Hogan M, Cerami A.Circulating fibrorytes define a new leukocyte subpopulationthat mediates tissue repair. Mol Med. 1994; 1: 7 l-81.
J. Cell. MoL Med. Vol9, No 2, 2005
105. Young IIM. Embryological origin of interstitial cells ofCajal |uficrosc Res Tech. 1999;47: 303-8.
106. Neijssen J, Iferberts C, Drijflrout JW, Reits E, Janssen L,Neelies J. Cross-presentation by intercellular peptide transferthrough gap jtnctions. Nanre. 2005, 434: 83-8.
107. Ward SM, Ordog T, Koh SD, Baker Sd Jun JY, AmbergG, Monaghan K Sanders KM. Pacemaking in interstitialcells ofCajal depends upon calcium handling by endoplasmicreticulum and mitochondriu J Plrysiol.2000; 525: 355-61.
108. Yoneda S, Takano II, Takaki M, Suzuh IL Properties ofspontaneowly active cells distributed in the submucosal layerof mouse proximal colon. J Physiol.2002; 542: 887-97.
109. Ilanani M, Farmgia G, Komuro T. Intercellular couplingofinterstitial cells of Cajal in the digestive tract. .Inr.R ev Cytol.2005;242:249-82.
110. Daniel EE, Willis A, Cho WJ, Boddy G. Comparisons ofneural and pacing activities in intestinal segments fromWAil++ a6 tgy67y1y; mice. Neurogastroenterol Motil.2005;l7:355-65.
1l l. Jankovic SM, Protic BA, Jankovic SV. Contractile effect ofacetylcholine on isolated ampullar segment of Fallopiantubes. Pharmacol Res. 2004; 49: 3 1 -5.
112. Samuelson [JE, Wiklund NP, Gustafsson LE. Endogenouspurines may modulate adrenergic neurotransmission in thehuman fallopian ttbe. Neurosci Lett. 1988; 86: 5l-5.
113. Zganshin AU, Vafna ZR" Fatkullin IF. Pharmacologicalcharacterization of P2-receptors in human fallopian tubes.Bull Exp Biol Med.2M4;137:242-5.
114. Ekerhovd E, Brannstrom M, Alexandersson M,Norstrom A. Evidence for nibic oxide mediation of conhac-tile activity in isolated sbips of the human Fallopian tube.Hum Reprod. 1997 ; 12:. 301 -5.
115. Ekerhovd E, Norstrom A. lnvolvement of a nitic oxide-cyclic guanosine monophosphate pathway in conhol offal-lopian tube conhactihty. Glnecol Endocrinal. 20M; 19 239-46.
ll6.BritonJones C, Hung Lok I, Yuen PM, Chiu TTY,Cheung LP, Haines C, Regulation of human oviductinmRNA expression n vivo. Fertil Steril. 20011. 7 5: 942-6.
I17. Streuli C. Extracellular matrix remodelling and cellular dif-ferentiation. Curr Opin Cell Biol. 1999,ll 63440.
I18. Tomasek JT, Gabbiani G Hinz B, Chaponnier C, BrownRA. Myofibroblasts and mechanoregulalion of connectivetissue remodelling . Nat Rev Mol Cell Biol. 2002;3: 349-63.
119. Demouillere d Darby IdGabbiani G. Normal and patho-logic soft tissue remodeling: role of the myofibroblast, withspecial emphasis on liver and kidney fibrosis. Lab Invest.2003;83: 1689-707.
120. Sartore S, Chiavegato A, Faggin E, Franch & Puato MAusoni S, Pauletto P. Conhibution ofadventitial fibroblaststo neointima formation and vascular remodeling. From inno-cent bystander to active participant. Circ Res.200l; 89: I I I l-21.
121. Watrelot d lfamilton J, Grudzinskas JG. Advances in theassessment of the uterus and fallopian tube function. BeJrPract Res Clin Obst Gynaecol.2003; l7: 187-209.
122.T^levi \ Gualtieri R" In vivo versus lll vitro fertil.zahon,Eur J Obst Gynecol Reprod Biol. 2004; 1 I 55: 568-571.
523

;\ }d $ ) 1.l i:.rq. [-- \.']-: \ \ *." [ {lw.itt$nrt taii 1.. lnpmttu -ttudtt:t(}PEItA () MN IA
\ \ q $"c b \ft i IX \ h g q. "S .\& g t- .,
8{
{,' $&n }}}:Kg {l$ [qL]Y$fr p$ { t.ju.t: 't:.: " t''.t.i. t 'f y _'. , ,... . .t q:s {... l: t:
(a.:""x4C.i'.1.-,,:,' i: ., lor;c.';i,;fi;ji;r.r Ni'l'ari !!jr r\.Jrr'!l, i !
s'\ *'\s&" &; htOMh{IA
3 "s; -;\1"{.,}5.$.fl{:t{"} - h jifl}g{;f}CHIRURGICA"
l/ i.{ C l.r .t t. {, tl "F s 'i 'it".:1"
Quurur:r
Utrcnchns prgin:r ilqtruri erhilxri:r ,
A !t s'!'lt L o s.{ }l I,*.;:,: J :. :,i :: - r '
h{" D" C. C" XX}\'t},
{"ffi.{"$&eLi#&ffir{'*nCu r*
,i l }' iq \J .'1 ld :\,1 { ii 'i.} I li ii ."r .'. ', I-{lcJrrryss, "8wurlt4, C'd/rgir ,llrzfJar., cF fd'rac rr Jriu,?'ua
,&r{,*re- Jfet,trd frr,frffoir ,
llN:.g""l l"ii .\i,. l-ii 5iir,Iii'l ii.r.f ., j, i...4jetcKj e- a'6r 6.9r* i;t ealmt At'zt*sxir
Fralif,ir:r.
i i ,;. i . | : .i .: i | : ' I'
f I C) A Ii I{ E I! D c V I v r E. JAp"al Er t( J()AN r< rIER&r. VgRBtiEK. )
:,.
r-r.ltL: ,nl{
Fallopian tubes (oviducts) as seen by ancient anatomists.Reproductions from Andrea Vesalius, Opera omnia anatomica & chirurgica (1725) andFredericus Ruysch, Operaomnia anatomico-medico-chirurgica (1737). Courtesy of the Medical Library of the 'Carol Davila' University ofMedicine and Pharmacy, Bucharest (library catalogue #CS XVIII V10 and #CS XVIII iV15).