normalization of tumor-induced increases in hepatic amino acid transport after surgical resection
TRANSCRIPT

ANNALS OF SURGERYVol. 221, No. 1, 50-58© 1995 J. B. Lippincott Company
Normalization of Tumor-InducedIncreases in Hepatic Amino AcidTransport After Surgical ResectionN. Joseph Espat, M.D.,* Barrie P. Bode, Ph.D.,t D. Scott Lind, M.D.,*Edward M. Copeland, M.D.,* and Wiley W. Souba, M.D., Sc.D.t
From the Department of Surgery, University of Florida College of Medicine,* Gainesville, Florida;and the Division of Surgical Oncology, Department of Surgery, Massachusetts General Hospitaland Harvard Medical School,t Boston, Massachusetts
BackgroundThe liver of the host with cancer requires increased amounts of amino acids to support thesynthesis of glucose and key defense proteins. To study the effect of the growing tumor onhepatic amino acid uptake, the authors measured hepatic transport activity in tumor-bearing ratsand in rats at various times after tumor resection.
MethodsFischer-344 rats were implanted subcutaneously with methylcholanthrene-induced fibrosarcomacells (MCA sarcoma). When the tumors reached 10% of body weight, hepatic amino acidtransport activity was assayed or the animals underwent surgical removal of the tumor. In animalsthat underwent tumor excision, livers were removed at 1, 3, or 5 days post-resection, and hepaticplasma membrane vesicles (HPMVs) were prepared. Nontumor-bearing pair-fed rats undergoingsham implantation or sham resection served as controls. System N (glutamine), System A(MeAIB), and System y+ (arginine) transport activity were assayed, which allowed the authors tocompare differences in tumor-induced rates of transport and the influence of resection ontransport activity.
ResultsSystem A transport activity was unaltered by tumor growth. In contrast, the presence of thegrowing tumor increased arginine and glutamine uptake by the liver. Hepatic glutamine transportremained elevated for 5 days after tumor resection, although by postoperative day 5 there was atrend toward normalization. In contrast, arginine transport remained increased by twofold on post-resection day 1 and had normalized by postoperative day 3. The enhanced arginine transport wasa result of an increase in maximal transport velocity (Vm,,) rather than a change in carrier affinity.
ConclusionsIncreases in hepatic amino acid transport normalize within several days of tumor resection,indicating a key role for the tumor in the induction of this response. The observation that hepaticglutamine transport activity remains augmented after tumor resection longer than any othertransporter studied suggests a key role for this amino acid in overall hepatic nitrogen metabolismand may partially explain the persistent glutamine depletion that is characteristic of the tumor-bearing host.
50

Hepatic Amino Acid Transport After Tumor Resection 51
Progressive malignant disease is associated with atranslocation of amino acids from peripheral musclestores to the liver. The liver uses these amino acids forgluconeogenesis and the synthesis of key defense pro-teins. It has been suggested that the growing tumor "elab-orates" substances that result in the production of host-derived cytokines, which initiate or perpetuate this cata-bolic response.",2 We previously demonstrated thathepatic plasma membrane amino acid transport activityis increased in the tumor-bearing rat and that this in-crease is mediated by circulating factors.3 These adapta-tions in hepatocyte amino acid transport appear to bea component of the complex syndrome termed cancercachexia. It is reasonable to predict, based on this work-ing hypothesis and on clinical experiences with malnour-ished patients with cancer, that removal ofthe tumor willresult in the restoration of host metabolism to normal.Indeed, the failure of patients with cancer to gain weightand strength after major "curative" extirpative surgeryshould raise the suspicion of residual disease.
In previous studies, we have shown that the rate ofglu-tamine and arginine transport across the hepatic plasmamembrane is tumor size dependent.4'5 Increases in he-patic amino acid transport activity are apparent evenwhen the tumor is small and when the host does not yetexhibit anorexia or cachexia. This accelerated transportincreases the availability of precursors that support met-abolic pathways involved in de novo protein synthesisand gluconeogenesis, ureagenesis, polyamine biosynthe-sis, and nitric oxide production.68 These metabolic ad-aptations in the liver may be essential for the host to re-spond to the growing tumor, and, as observed in otherkinds ofcatabolic patients,9 10 augmented hepatic aminoacid uptake during critical illness may a confer a survivaladvantage to the host. In the current studies, we havetested the hypothesis that this increase in hepatic aminoacid transport is the result of the presence of the tumoritself and would, therefore, decay to basal values aftertumor resection. The purpose of the current investiga-tions was to measure hepatic amino acid transport aftertumor resection to determine if and when transport ac-tivity normalized and whether these changes were selec-tive or specific for a particular amino acid transporter.
Presented at the 47th Annual Meeting of the Society for Surgical On-cology; March 17-20, 1994; Houston, Texas.
Supported by NIH grant 56790 (Dr. Souba).Address correspondence to Wiley W. Souba, M.D., Sc.D., Chief, Divi-
sion of Surgical Oncology, Department of Surgery, MassachusettsGeneral Hospital, Cox Building, 100 Blossom Street, Boston, MA02114.
Accepted for publication May 23, 1994.
METHODSImplantation/Resection of TumorsAdult male Fischer-344 rats (175-200 g, Harlan In-
dustries, Indianapolis, IN) were used for the studies. Allexperiments were approved by the Committee for theUse and Care ofLaboratory Animals at the University ofFlorida. The rats were housed individually in wire cagesunder standard conditions (12-hour light-dark cyclesand access to rat chow and water ad libitum) and wereallowed at least 5 days to acclimate to the animal carefacility. Before study, rats were weighed and separatedrandomly into two groups. Tumor-bearing rats un-derwent bilateral subcutaneous flank implantation of 2X 2 X 2-mm cubes of methylcholanthrene-induced fi-brosarcoma cells (MCA tumor), as described pre-viously." The incision was closed with surgical staples.Control rats underwent sham tumor implantation andwere pair fed to match carcass weight. When the tumorscomprised 10% ofbody weight (14-16 days post-implan-tation) control and tumor-bearing animals were dividedfurther into subgroups. In some animals, the livers wereexcised and hepatic plasma membrane vesicles were pre-pared as described previously. In the remaining rats ineach group, the tumors were excised under anesthesia af-ter the flanks were prepared in a sterile fashion. After tu-mor resection, the wound was irrigated with normal sa-line, and the skin was closed primarily with a continuous3-0 nylon suture. Control sham-implanted rats were pre-pared similarly, and a sham resection was performed. Af-ter making the flank skin incision, a subcutaneouspocket similar to the size of that present in the tumor-bearing rats was created and closed with staples. The tu-mor-resected rats and sham-resected controls then weredivided into three additional subgroups. Livers were ex-cised, and hepatic membrane vesicles were prepared onpostoperative days 1, 3, or 5.
Preparation of Liver Plasma MembraneVesicles
Hepatic plasma membrane vesicles (HPMVs) fromthe livers of control and tumor-bearing rats were pre-pared as described previously.'2 Vesicle aliquots werestored at -70 C in Nunc cryotubes (InterMed, Copenha-gen, Denmark) until studied. Relative enrichment or im-poverishment in the specific activity of the marker en-zymes 5'-nucleotidase'3 and glucose-6-phosphatase'4were assayed to evaluate vesicle purity biochemically. In-organic phosphate was measured by the method de-scribed by Fiske. 5
Vesicle Amino Acid Transport AssayHepatic plasma membrane vesicle transport of gluta-
mine, arginine, and the nonmetabolizable amino acid
Vol. 221 - No. 1

52 Espat and Others
analog 2-(Methylamino)isobutyric acid (MeAIB) wasevaluated by a rapid mixing/filtration technique de-scribed previously.'2 All transport assays were carriedout at 22 C. With the exception of uptake time coursesand osmolarity plots, transport was terminated at the 10-second time point to ensure measurement of initial ratesunder these assay conditions. L-[3H]-arginine and L-[G-3H]-glutamine were purchased from Amersham (Arling-ton Heights, IL). L-[3H]-MeAIB was obtained fromAmerican Radiolabeled Chemicals (St. Louis, MO).
System A and System N activity were defined as theNa+-dependent transport of MeAIB and glutamine, re-spectively. The Na+-dependent component of aminoacid transport was calculated by subtracting uptake inthe presence of K+ (Na+-independent uptake, triplicateor quadruplicate determinations) from that observed inthe presence of Na+ (total uptake, quadruplicate deter-minations). Because arginine transport by HPMVs pre-viously has been shown to be primarily Na+-dependent(System y+),'6 saturable carrier-mediated argininetransport was determined by the evaluation ofthe uptakeof [3H]-arginine in K+ buffer in either the presence orabsence of at 10 mmol/L excess of unlabeled arginine.Similarly, saturable Na+-independent glutamine uptakewas determined by subtracting uptake values in the pres-ence of excess unlabeled glutamine (nonsaturable up-take, quadruplicate samples) from that in the absence ofexcess (10 mmol/L) glutamine (total Na+-independentuptake, quadruplicate samples). Kinetic parameterswere evaluated by varying the arginine concentration invarious experiments over the range of 25 ,uM to 8.0 mMand subtracting the nonsaturable component obtainedin the presence of excess unlabeled arginine. Kinetic ex-periments also were performed at the 10-second initialrate conditions.Osmotic adjustment for varying concentrations of all
three amino acids studied were made with sucrose whereappropriate. In all transport experiments, blank values(no vesicles present) were determined and subtractedfrom the corresponding amino acid uptakes. Membraneprotein was determined by a modified Lowry method,'7and all uptakes were normalized to protein. Transportvelocities and kinetic parameters were determined fromscintillation counts and protein content of vesicle sam-ples using a computer software program. Data are ex-pressed as transport velocity in pmol/mg protein/unittime.
Statistical AnalysisData were analyzed by either Student's t test or by
analysis of variance, where appropriate, followed byFischer LSD test if a significant F value was obtained,using a computer (Macintosh LC, Apple Computer, Inc.,
Glucose6-phosphatase _~UCruae LiverGlucse_-phsp a* Homogenate
U Vesicles
5' Nucleotidase
0 15 30 45 60Specific Activity(pmollhr/mg protein)
Figure 1. Specific activity of marker enzymes in plasma membrane andendoplasmic reticulum. The specific activities of the marker enzymes 5'-nucleotidase and glucose-6-phosphatase in vesicles were compared withthose in the crude liver homogenates. The activities in HPMVs were ex-pressed relative to those in the homogenates. The plasma membranemarker 5'-nucleotidase was enriched, whereas the cytosolic (endoplasmicreticulum) enzyme glucose-6-phosphatase was impoverished, indicatingvesicle purity. *p < 0.05, **p < 0.01 vs. crude liver homogenate.
Cupertino, CA) and commercial software (Statview SE+ Graphics, Abacus Concepts, Inc., Berkeley, CA). Thelevel of statistical significance was set at p < 0.05.
RESULTSAll skin incisions after implantation or resection
healed without evidence of dehiscence or infection.Mean tumor size at the time of HPMV preparation ortumor resection was 10 ± 1% of total body weight.
Assessment of HPMV IntegrityWe have previously documented the plasma mem-
brane vesicular nature and functional reliability of ourvesicles' preparations in terms of their capacity for time-and concentration-dependent carrier-mediated accumu-lation of amino acids, enrichment in plasma membraneenzymatic markers, and electron microscopic appear-ance.18 In the current report, the specific activity of se-lected marker enzymes were measured to evaluate vesi-cle enrichment in plasma membrane and impoverish-ment in endoplasmic reticulum. Vesicles from controland tumor-resected animals rats were enriched 14- to 15-fold in the specific activity of the plasma membraneenzyme marker 5'-nucleotidase, over the crude liverhomogenates, (2.7 ± 0.20 ,tmoles/mg protein/hr in ho-mogenate vs. 39.5 ± 2.9 in vesicles, p < 0.01, Fig. 1) Con-comitant with the marked enrichment in plasma mem-brane, preparation of vesicles from both control and tu-mor-resected animals did not result in an enrichment ofthe microsomal membrane fraction, as evidenced by
Ann. Surg. *-January 1995
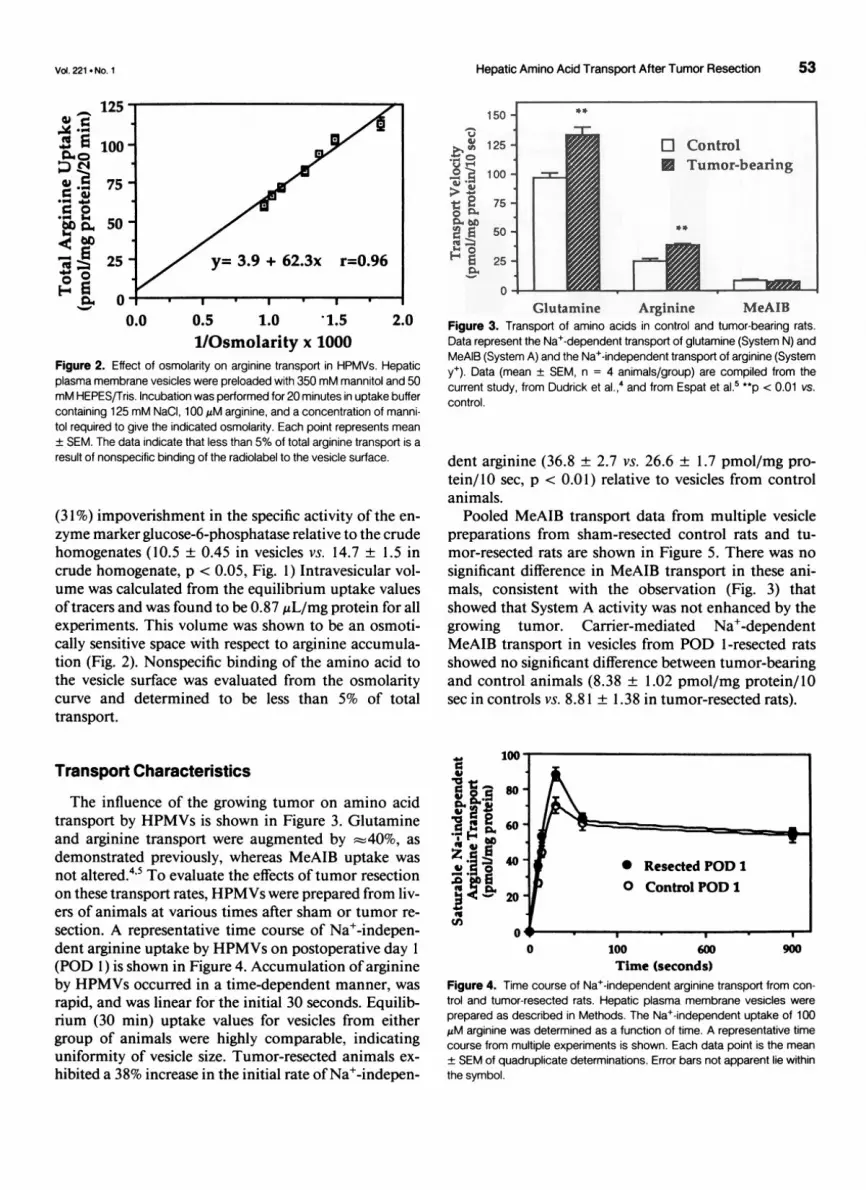
Hepatic Amino Acid Transport After Tumor Resection 53
1000
75
; 50
25 y= 3.9 + 62.3x r=0.96
0.0 0.5 1.0 1.5 2.01/Osmolarity x 1000
Figure 2. Effect of osmolarity on arginine transport in HPMVs. Hepaticplasma membrane vesicles were preloaded with 350 mM mannitol and 50mM HEPES/Tris. Incubation was performed for 20 minutes in uptake buffercontaining 125 mM NaCI, 100 AM arginine, and a concentration of manni-tol required to give the indicated osmolarity. Each point represents mean± SEM. The data indicate that less than 5% of total arginine transport is aresult of nonspecific binding of the radiolabel to the vesicle surface.
(31 %) impoverishment in the specific activity of the en-
zyme marker glucose-6-phosphatase relative to the crudehomogenates (10.5 ± 0.45 in vesicles vs. 14.7 ± 1.5 incrude homogenate, p < 0.05, Fig. 1) Intravesicular vol-ume was calculated from the equilibrium uptake valuesof tracers and was found to be 0.87 uL/mg protein for allexperiments. This volume was shown to be an osmoti-cally sensitive space with respect to arginine accumula-tion (Fig. 2). Nonspecific binding of the amino acid tothe vesicle surface was evaluated from the osmolaritycurve and determined to be less than 5% of totaltransport.
Transport Characteristics
The influence of the growing tumor on amino acidtransport by HPMVs is shown in Figure 3. Glutamineand arginine transport were augmented by :40%, as
demonstrated previously, whereas MeAIB uptake was
not altered.4'5 To evaluate the effects of tumor resectionon these transport rates, HPMVs were prepared from liv-ers of animals at various times after sham or tumor re-
section. A representative time course of Na+-indepen-dent arginine uptake by HPMVs on postoperative day 1
(POD 1) is shown in Figure 4. Accumulation of arginineby HPMVs occurred in a time-dependent manner, was
rapid, and was linear for the initial 30 seconds. Equilib-rium (30 min) uptake values for vesicles from eithergroup of animals were highly comparable, indicatinguniformity of vesicle size. Tumor-resected animals ex-
hibited a 38% increase in the initial rate ofNa+-indepen-
150 -
U
t,' 125-
. Co
_ 100-
t: 2 75.o vw
2 ; 50-o0
O Control* Tumor-bearing
AE 25
Glutamine Arginine MeAIBFigure 3. Transport of amino acids in control and tumor-bearing rats.Data represent the Na+-dependent transport of glutamine (System N) andMeAIB (System A) and the Na+-independent transport of arginine (Systemy+). Data (mean ± SEM, n = 4 animals/group) are compiled from thecurrent study, from Dudrick et al.,4 and from Espat et al.5 **p < 0.01 vs.control.
dent arginine (36.8 ± 2.7 vs. 26.6 ± 1.7 pmol/mg pro-tein/10 sec, p < 0.01) relative to vesicles from controlanimals.
Pooled MeAIB transport data from multiple vesiclepreparations from sham-resected control rats and tu-mor-resected rats are shown in Figure 5. There was nosignificant difference in MeAIB transport in these ani-mals, consistent with the observation (Fig. 3) thatshowed that System A activity was not enhanced by thegrowing tumor. Carrier-mediated Na+-dependentMeAIB transport in vesicles from POD 1-resected ratsshowed no significant difference between tumor-bearingand control animals (8.38 ± 1.02 pmol/mg protein/10sec in controls vs. 8.81 ± 1.38 in tumor-resected rats).
11Sso1h=so
I 60
::i.4 0RscePO
40,$°ve20 "r0 Control POD 1
0 100 600 900Time (seconds)
Figure 4. Time course of Na+-independent arginine transport from con-trol and tumor-resected rats. Hepatic plasma membrane vesicles wereprepared as described in Methods. The Na+-independent uptake of 1001AM arginine was determined as a function of time. A representative timecourse from multiple experiments is shown. Each data point is the mean± SEM of quadruplicate determinations. Error bars not apparent lie withinthe symbol.
Vol. 221 * No. I
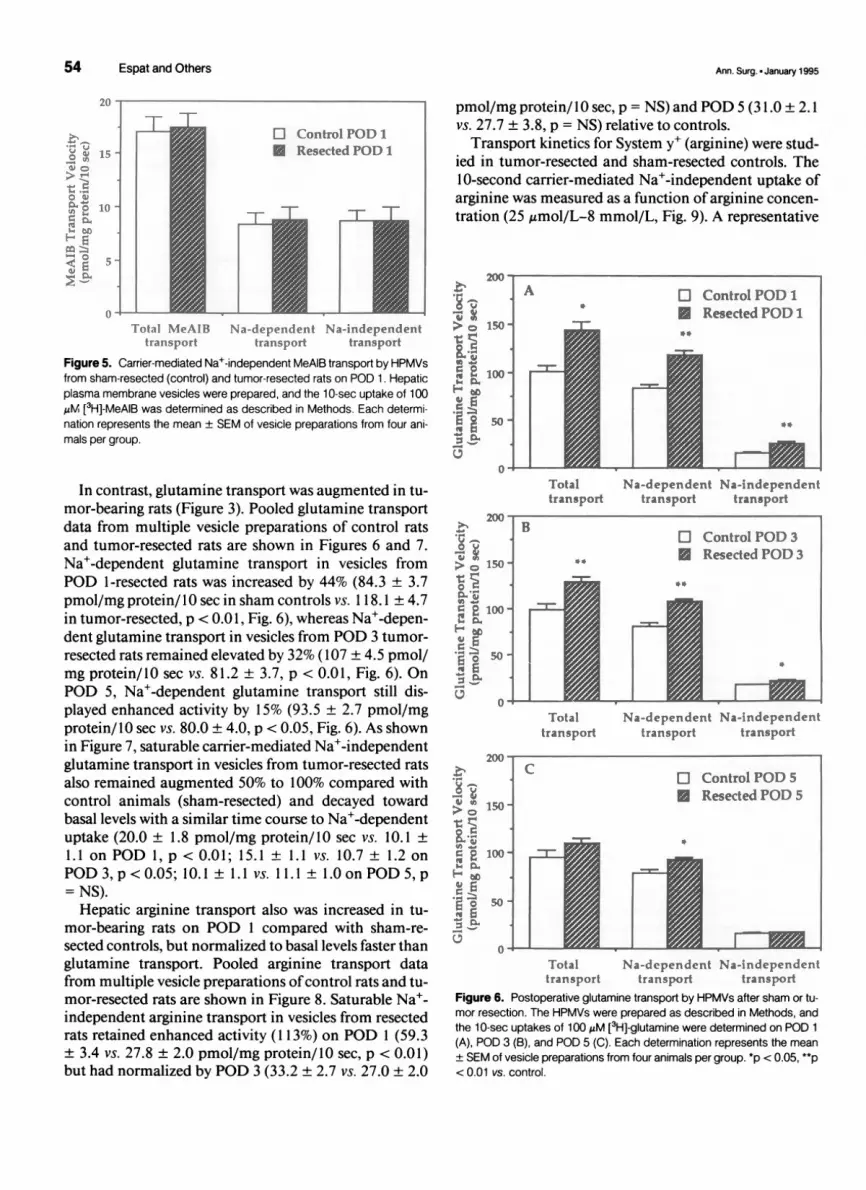
54 Espat and Others
20 -
o-
.-,'-J 0u 1
OQJn
-I El Control POD 1U Resected PODl
0' 10
1 E 0
Total MeAIB Na-dependent Na-independenttransport tranisport transport
Figure 5. Carrier-mediated Na+-independent MeAIB transport by HPMVsfrom sham-resected (control) and tumor-resected rats on POD 1. Hepaticplasma membrane vesicles were prepared, and the 1 0-sec uptake of 100,M [3H]-MeAIB was determined as described in Methods. Each determi-nation represents the mean ± SEM of vesicle preparations from four ani-mals per group.
In contrast, glutamine transport was augmented in tu-mor-bearing rats (Figure 3). Pooled glutamine transportdata from multiple vesicle preparations of control ratsand tumor-resected rats are shown in Figures 6 and 7.Na+-dependent glutamine transport in vesicles fromPOD 1 -resected rats was increased by 44% (84.3 ± 3.7pmol/mg protein/ 10 sec in sham controls vs. I 1 8.1 ± 4.7in tumor-resected, p < 0.01, Fig. 6), whereas Na+-depen-dent glutamine transport in vesicles from POD 3 tumor-resected rats remained elevated by 32% (107 ± 4.5 pmol/mg protein/10 sec vs. 81.2 ± 3.7, p < 0.01, Fig. 6). OnPOD 5, Na+-dependent glutamine transport still dis-played enhanced activity by 15% (93.5 ± 2.7 pmol/mgprotein/ IO sec vs. 80.0 ± 4.0, p < 0.05, Fig. 6). As shownin Figure 7, saturable carrier-mediated Na+-independentglutamine transport in vesicles from tumor-resected ratsalso remained augmented 50% to 100% compared withcontrol animals (sham-resected) and decayed towardbasal levels with a similar time course to Na+-dependentuptake (20.0 ± 1.8 pmol/mg protein/0 sec vs. 10.1 +1.1 on POD 1, p < 0.01; 15.1 + 1.1 vs. 10.7 ± 1.2 onPOD 3, p <0.05; 10.1 + 1.1 vs. 11.1 ± 1.0on POD 5, p= NS).Hepatic arginine transport also was increased in tu-
mor-bearing rats on POD 1 compared with sham-re-sected controls, but normalized to basal levels faster thanglutamine transport. Pooled arginine transport datafrom multiple vesicle preparations ofcontrol rats and tu-mor-resected rats are shown in Figure 8. Saturable Na+-independent arginine transport in vesicles from resectedrats retained enhanced activity (113%) on POD 1 (59.3+ 3.4 vs. 27.8 ± 2.0 pmol/mg protein/10 sec, p < 0.01)but had normalized by POD 3 (33.2 ± 2.7 vs. 27.0 ± 2.0
pmol/mg protein/ 10 sec, p = NS) and POD 5 (31.0 ± 2.1vs. 27.7 ± 3.8, p = NS) relative to controls.Transport kinetics for System y+ (arginine) were stud-
ied in tumor-resected and sham-resected controls. The10-second carrier-mediated Na+-independent uptake ofarginine was measured as a function of arginine concen-tration (25 ,mol/L-8 mmol/L, Fig. 9). A representative
200
-q
> o 150
r, 0
{<m
-t
00 tLL7~~~-idmTotal Na-dependent Na-independenttransport transport transport
IITF)Total Na-dependent Na-independent
transport transport transport
5;
01Total Na-dependent Na-independenttransport transport transport
Figure 6. Postoperative glutamine transport by HPMVs after sham or tu-mor resection. The HPMVs were prepared as described in Methods, andthe 1 0-sec uptakes of 100 ,uM [3H]-glutamine were determined on POD 1(A), POD 3 (B), and POD 5 (C). Each determination represents the mean± SEM of vesicle preparations from four animals per group. *p < 0.05, **p< 0.01 vs. control.
Ann. Surg. *-January 1995
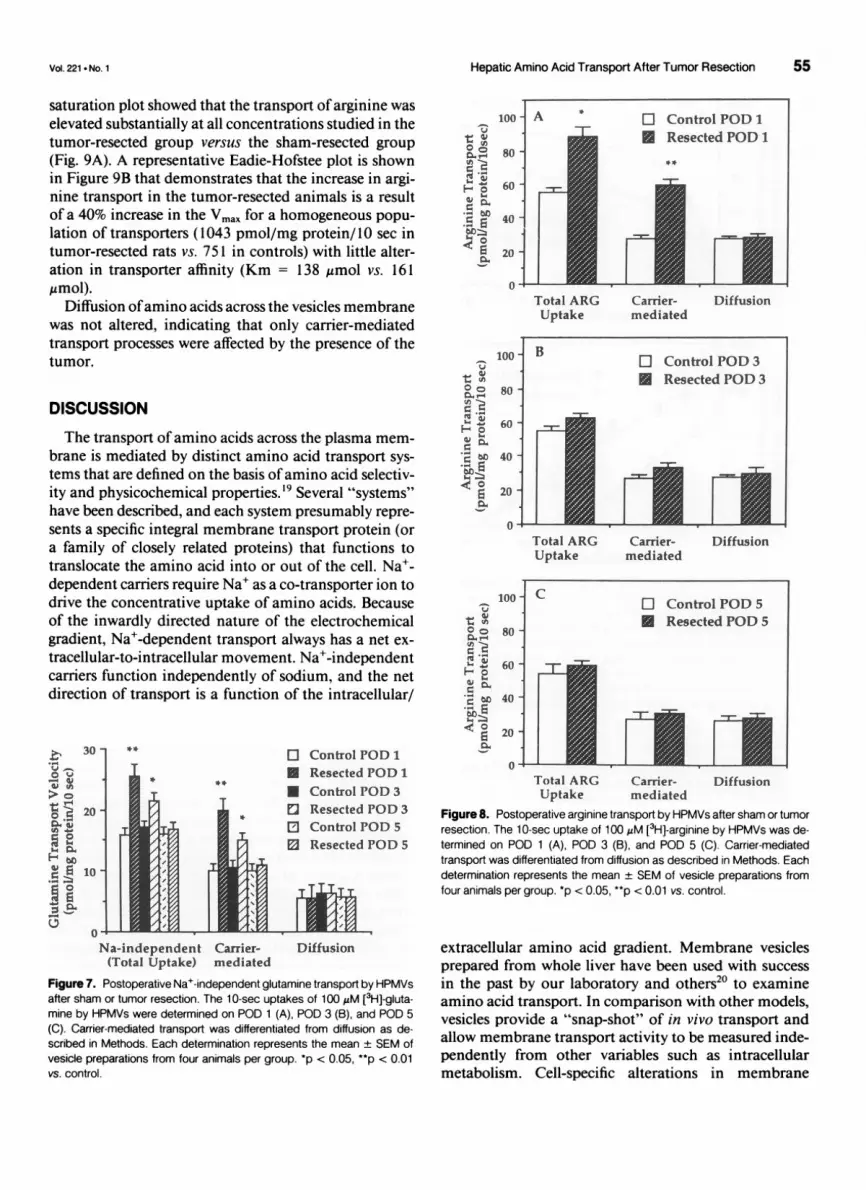
Hepatic Amino Acid Transport After Tumor Resection 55
saturation plot showed that the transport of arginine waselevated substantially at all concentrations studied in thetumor-resected group versus the sham-resected group(Fig. 9A). A representative Eadie-Hofstee plot is shownin Figure 9B that demonstrates that the increase in argi-nine transport in the tumor-resected animals is a resultof a 40% increase in the Vmax for a homogeneous popu-lation of transporters (1043 pmol/mg protein/10 sec intumor-resected rats vs. 751 in controls) with little alter-ation in transporter affinity (Km = 138 qrmol vs. 161,urmol).
Diffusion ofamino acids across the vesicles membranewas not altered, indicating that only carrier-mediatedtransport processes were affected by the presence of thetumor.
DISCUSSIONThe transport ofamino acids across the plasma mem-
brane is mediated by distinct amino acid transport sys-tems that are defined on the basis ofamino acid selectiv-ity and physicochemical properties.'9 Several "systems"have been described, and each system presumably repre-sents a specific integral membrane transport protein (ora family of closely related proteins) that functions totranslocate the amino acid into or out of the cell. Na+-dependent carriers require Na+ as a co-transporter ion todrive the concentrative uptake of amino acids. Becauseof the inwardly directed nature of the electrochemicalgradient, Na+-dependent transport always has a net ex-
tracellular-to-intracellular movement. Na+-independentcarriers function independently of sodium, and the netdirection of transport is a function of the intracellular/
t, 30 El Control POD1, %J 0 Resected POD
> o Control PlOD 3
t 20 Resected POD 3El Control PODS
54E ResectedPOD5
0
Na-independent Carrier- Diffusion(Total Uptake) mediated
Figure 7. Postoperative Na+-independent glutamine transport by HPMVsafter sham or tumor resection. The 1 0-sec uptakes of 100 OM [3H]-gluta-mine by HPMVs were determined on POD 1 (A), POD 3 (B), and POD 5(C). Carrier-mediated transport was differentiated from diffusion as de-scribed in Methods. Each determination represents the mean ± SEM ofvesicle preparations from four animals per group. *p < 0.05, **p < 0.01vs. control.
~u
Hov
(A
be0
Total ARG Carrier-Uptake mediated
Diffusion
_100 B E Control POD3t U Resected POD 3, C 80
60
bet 40
E 20
0Total ARG Carrier- DiffusionUptake mediated
100 C C1 Control POD 5t U Resected POD 5
80-
= 60 -
F-o
to 40-
20 -
0Total ARG Carrier- DiffusionUptake mediated
Figure 8. Postoperative arginine transport by HPMVs after sham or tumorresection. The 1 0-sec uptake of 100 1IM [3H]-arginine by HPMVs was de-termined on POD 1 (A), POD 3 (B), and POD 5 (C). Carrier-mediatedtransport was differentiated from diffusion as described in Methods. Eachdetermination represents the mean ± SEM of vesicle preparations fromfour animals per group. *p < 0.05, **p < 0.01 vs. control.
extracellular amino acid gradient. Membrane vesiclesprepared from whole liver have been used with successin the past by our laboratory and others20 to examineamino acid transport. In comparison with other models,vesicles provide a "snap-shot" of in vivo transport andallow membrane transport activity to be measured inde-pendently from other variables such as intracellularmetabolism. Cell-specific alterations in membrane
Vol. 221 * No. 1

56 Espat and Others
0.0 2.0 4.0 6.0 8.0
Arginine (mM)
12000 B
400w \* Resected POD1
0 Control POD
200
{0_SS
0 200 400 600 800Velocity/[Argininel
Figure 9. Kinetic analysis of saturable carrier-mediated Na+-independentarginine transport by HPMVs on POD 1 after sham or tumor resection.
(A) Saturation plot. The 10-sec Na+-independent uptake of arginine as
a function of extravesicular glutamine concentration was determined as
described in Methods. (B) Eadie-Hofstee linear transformation of Na+-in-dependent arginine transport. The negative slope of each line is the Km
(affinity of the transporter for arginine); the y-intercept is the V,W,. Actual
kinetic values are provided in the text. Each data point is the mean
SEM of quadruplicate determinations. Error bars not apparent lie within the
symbol.
transport activity that occur in intact hepatocytes are re-
tained during the preparation of vesicles.2'
We chose to study three amino acids that are
transported across the hepatic plasma membrane by dis-
tinctly different carrier proteins. In the case ofthe amino
acid glutamine, its transport into the hepatocyte is medi-
ated by the Na+-dependent system "N," 22 whereas its
transport out of the liver is mediated by the Na+-inde-pendent system "n." 23 The presence of these two gluta-
mine transporters working simultaneously but in oppo-
site directions allows the liver to control the uptake and
release of glutamine through differential expression of
each carrier.24'25 Arginine uptake by the liver, in contrast
to glutamine, is mediated by the Na+-independentagency, System y+.26 The net direction of uptake is into
the hepatocyte because the intracellular arginine concen-
tration is ten times lower than the plasma concentration.
System A mediates the uptake of small neutral amino
acids, such as alanine, and its transport commonly isevaluated using the selective nonmetabolizable aminoacid analog MeAIB.'8
Similar to previous investigations,4'5 the present stud-ies demonstrate that glutamine and arginine transportwere enhanced in the liver of rats bearing the subcutane-ously implanted methylcholanthrene-induced fibrosar-coma tumor. The tumor is locally aggressive, but rarelymetastasizes and results in death approximately 5 to 6weeks after implantation secondary to the complicationsof cancer cachexia. We chose to study rats with tumorsthat comprised 10% of body weight because previousstudies have shown that transport activity is enhancedwhen the tumor reaches this size. Although transportrates for glutamine and arginine are increased further asa function of tumor size, the high mortality associatedwith resection of larger tumors precluded us from study-ing animals with a greater than 10% tumor volume.Our results demonstrate that tumor-induced increases
in hepatic amino acid transport return to pretumor im-plantation levels after surgical resection of the malig-nancy. The rate of normalization was different for gluta-mine and arginine. Arginine transport had normalizedby the third day after tumor resection, whereas normal-ization of System "N" activity (Na+-dependent gluta-mine transport) required 5 days. The significance ofthese differences is unclear but may be consistent withthe important role ofglutamine in overall hepatic nitro-gen metabolism. Moreover, persistent hepatic glutamineuptake helps explain the marked glutamine depletionthat is characteristic of the host with cancer. Na+-inde-pendent glutamine transport (system n) also was in-creased in the postoperative period and normalized in asimilar temporal fashion to System N. These alterationsin transport activity were not caused by the surgical stressoftumor resection, as evidenced by lack ofenhanced ac-tivity in livers from sham-resected pair-fed controls.
Consistent with previous studies that have evaluatedtransport kinetic in the livers of tumor-bearing animals,kinetic analysis of System y+ (arginine transport) indi-cated that the persistent increase in transport activity af-ter surgical resection was the result of a Vmax effect. Thetransport activity of a specific carrier can increase or de-crease in two principal ways-either the affinity (Km =Michaelis constant) of the transporter for its substratechanges or the number of active transporters in the cellmembrane changes (Vmax = maximal transport velocity).An increase in transporter affinity occurs when the con-formation ofthe carrier protein is altered so that the sub-strate binds more readily to its catalytic site on the trans-porter. In contrast to a Km effect, an increase in thenumber of active transporter proteins in the cell mem-brane usually involves de novo protein synthesis, pre-sumably ofthe carrier itself. The increase in hepatic argi-
Ann. Surg. * January 1995

Vol. 221 * No. 1
nine transport that we recently observed in tumor-bear-ing animals was caused by changes in Vmax of the y+carrier5; our current results demonstrate that the increasein the Vmax of System y+ persists (POD 1).The liver of the host with cancer requires an increase
in amino acids to support hepatic metabolism, in partic-ular the synthesis of key defense proteins. Warren et al.demonstrated that synthetic rates oftotal cellular hepaticprotein in hepatocytes from tumor-bearing animals havebeen shown to be twice that of control hepatocytes, withthe increase proportional to tumor burden.27'28 Synthesisof both secretory and structural proteins increases, butno net accrual of protein in hepatocytes occurs, consis-tent with an increase in the release ofde novo synthesizedacute phase proteins into the bloodstream and a simulta-neous increase in the degradation rate of structural pro-teins. Thus, in rats with advanced cancers, the liver ap-pears to increase and re-prioritize protein synthesis rates,events that require an increase in available amino acidprecursors. Both glutamine and arginine are required forprotein synthesis, but they also play special roles in he-patic nitrogen metabolism. Glutamine plays a centralrole in ammonia metabolism in the liver and also canserve as a gluconeogenic precursor. Arginine is the exclu-sive precursor for the biosynthesis ofthe multifunctionalbioregulatory molecule nitric oxide, which plays a keyrole in the lysis of tumor cells.29 Augmented hepatic ar-ginine transport may support nitric oxide biosynthesis inthe tumor-bearing host.Amino acid transport and its relevance to substrate use
and interorgan metabolic flow has been an area of greatinterest. We have been interested in the role of the liverin the host with cancer and the regulation of this re-sponse. Previous studies have demonstrated that the in-crease in hepatic amino acid transport activity that oc-curs in the liver of tumor-bearing rats is mediated by se-rum factors4'5 and can be attenuated by administrationofantibody to tumor necrosis factor (Souba et al., manu-script in review). It has been hypothesized that the grow-ing tumor elaborates factors that stimulate host cells torelease cytokines. Our data are consistent with this no-tion because removal of the tumor resulted in normal-ization ofhepatic transport rates. The return oftransportactivity over a several-day period (rather than hours) sug-gests that these signals persist for a period of time afterthe tumor has been extirpated. It also is apparent thatthere are differences in normalization rates for the trans-porters we evaluated. Moreover, the observation that he-patic glutamine transport activity remains augmentedafter tumor resection longer than the other transportersstudied suggests a key role for this amino acid in overallhepatic metabolism and may partially explain the persis-tent glutamine depletion that is characteristic ofthe hostwith cancer after surgical resection of the malignancy.
Hepatic Amino Acid Transport After Tumor Resection 57
References1. Kern KA, Norton JA. Cancer cachexia. JPEN J Parenter Enteral
Nutr 1988; 12:286-298.2. Souba WW. Glutamine and cancer. Ann Surg 1993; 218:715-728.3. Dudrick PS, Copeland EM, Souba WW. Hepatocyte glutamine
transport in advanced malignant disease. Surg Forum 1992; 43:13-15.
4. Dudrick PS, Inoue Y, Espat NJ, Souba WW. Na+-dependent glu-tamine transport in the liver of tumor-bearing rats. Surg Oncol1993; 2:205-215.
5. Espat NJ, Lind DS, Copeland EM, Souba WW. Accelerated argi-nine transport in the liver of tumor-bearing rats. Ann Surg Oncol1994; 1:147-156.
6. Bode BP, Tamarappoo BK, Mailliard M, et al. Characteristics andregulation ofhepatic glutamine transport. JPEN J Parenter EnteralNutr 1990; 14:5 lS-SSS (suppl).
7. Schertel B, Eichler W. Polyamine biosynthesis in arginine-starvedand refed rats. Biol Chem Hoppe-Seyler 1991; 372:27-33.
8. Curran RD, Billiar TR, Stuehr DJ, et al. Hepatocytes produce ni-trogen oxides from L-arginine in response to inflammatory prod-ucts from Kupffer cells. J Exp Med 1989; 170:1769-1774.
9. Rosenblatt SR, Clowes GHA, George B, et al. Exchange of aminoacid by muscle and liver in sepsis: comparative studies in vivo andin vitro. Arch Surg 1983; 118:167-175.
10. Clowes GHA, Hirsch E, George B, et al. Survival from sepsis: thesignificance of altered protein metabolism regulated by proteolysisinducing factor, the circulating cleavage product of interleukin- 1.Ann Surg 1985; 202:446-458.
11. Austgen TR, Dudrick PS, Sitren HS, et al. The effects ofglutamine-enriched total parenteral nutrition on tumor growth and host tis-sues. Ann Surg 1992; 215:107-113.
12. Pacitti AJ, Inuoe Y, Souba WW. Tumor necrosis factor stimulatesNa+-dependent amino acid transport in plasma membrane vesiclesfrom rat liver. J Clin Invest 1993; 91:474-483.
13. Moore DJ. Enzyme purification and related techniques. MethodsEnzymol 1971; 22:130-148.
14. Swanson MA. Glucose-6-phosphatase from liver. Methods Enzy-mol 1955; 2:541-543.
15. Fiske CH, Subbarow Y. The colorimetric determination of phos-phorus. J Biol Chem 1925; 66:375-400.
16. Inoue Y, Bode BP, Beck D, et al. Arginine transport in humanliver: characterization and effects of nitric oxide synthase inhibi-tors. Ann Surg 1993; 218:350-363.
17. Bensadoun A, Weinstein D. Assay of proteins in the presence ofinterfering materials. Anal Biochem 1976; 70:241-250.
18. Pacitti AJ, Austgen TR, Souba WW. Adaptive regulation of ala-nine transport in hepatic membrane vesicles (HPMVs) from theendotoxin-treated rat. J Surg Res 1991; 51:46-53.
19. Souba WW, Pacitti AJ. How amino acids get into cells: mecha-nisms, models, menus, and mediators. JPEN J Parenter EnteralNutr 1992; 16:569-578.
20. Jacob R, Rosenthal N, Barrett EJ. Characterization of glutaminetransport by liver plasma membrane vesicles. Am J Physiol 1986;25 l:E509-E514.
21. Schenerman MA, Kilberg MS. Maintenance of glucagon-stimu-lated System A amino acid transport activity in rat liver plasmamembrane vesicles. Biochim Biophys Acta 1986; 856:428-436.
22. Kilberg MS, Handlogten ME, Christensen HN. Characteristics ofan amino acid transport system in rat liver for glutamine, aspara-gine, histidine, and closely related analogs. J Biol Chem 1980; 255:4011-4019.
23. Pacitti AJ, Inoue Y, Souba WW. Characterization ofNa+-indepen-dent glutamine transport in rat liver. Am J Physiol 1993; 265:G90-G98.

58 Espat and Others
24. Fafournoux P, Demigne C, Remesy C, et al. Bidirectionaltransport of glutamine across the cell membrane in rat liver. Bio-chem J 1983; 216:401-408.
25. Pacitti AJ, Copeland EM III, Souba WW. Mechanisms of net he-patic glutamine release in the tumor-bearing rat. Surg Oncol 1992;1:173-182.
26. Pacitti AJ, Copeland EM, Souba WW. Stimulation of hepatic sys-tem y+-mediated L-arginine trasport by an inflammatory agent.Surgery 1992; 1 12:403-41 1.
Ann. Surg. * January 1995
27. Warren RS, Jeevanandam M, Brennan MF. Comparison of he-patic protein synthesis in vivo versus in vitro in the tumor-bearingrat. J Surg Res 1987; 42:43-50.
28. Warren RS, Jeevanandam M, Brennan MF. Protein synthesisin the tumor-influenced hepatocyte. Surgery 1985; 98:275-282.
29. Li I, Kilbourn RG, Fidler IJ. Role of nitric oxide in the lysis oftumor cells by cytokine activated endothelial cells. Cancer Res199 1; 5 1:2531-2535.