nitrogen metabolism in the ectomycorrhizal fungus hebeloma crustuliniforme
TRANSCRIPT

Nev; Phytol. (1995), 131, 263-271
Nitrogen metabolism in the ectomycorrhizalfungus Hebeloma crustuliniforme
BY A L I M. Q U O R E S H P , I F T I K H A R AHMAD^*, DAVID MALLOCH^AND JOHAN A. HELLEBUST^
^Faculty of Forestry., University of Toronto, 33 Willcocks Street, Toronto, Ontario.,Canada M5S 3B3^Department of Botany, University of Toronto, 25 Willcocks Street, Toronto, Ontario,Canada M5S 3B2
{Received 2 February 1995 ; accepted 8 June 1995)
SUMMARY
With glucose as the carbon source, the ectomycorrhizal basidiomycete Hebeloma crustuliniforme (Bull. ex. Amans)Quel. utilizes nitrate, ammonium and amino acids as sources of nitrogen. Inoculated as a suspension of fine hyphaein such liquid media, the fungus expands rapidly, exhibiting a distinct exponential phase with a doubling time of1-2 d and a maximum content of mycelial protein during the first 7 d of growth. Its growth when amino acids serveas sole carbon and nitrogen source is, however, less rapid, and requires several days of induction under theseconditions. In H. crustuliniforme, assimilation of ammonium appears to be carried out primarily via the NADPH-glutamate dehydrogenase (GDH) pathway. Its NADPH-GDH levels in exponentially growing mycelia are severaltimes greater than the apparent nitrogen assimilation rates of the mycelia. Its levels of glutamine synthetaseactivity in comparison are very small during the exponential period of growth. The greatest levels of NADH-GDH are found in glucose-grown cultures during the post-exponential period, and in glucose-free amino acid-grown cultures during the active period of growth. This suggests a catabolic role for the NADH-GDH. Aspartateaminotransferase and alanine aminotransferase in H. crustuliniforme are highly active during the exponentialperiod of growth, and their concentrations are increased further in the presence of amino acids as sole nitrogensources, suggesting a key role for these enzymes in interactions between pathways of amino acid metabolism andcarbon metabolism in tbis fungus.
Key words: Ectomycorrhizal fungus, growth conditions, nitrogen assimilation, enzyme regulation, Hebelomacrustuliniforme.
hand, several studies of enzyme distribution, mutantlNTRODLCTiON analysis and ' ^ N H / labelling appear consistent withEctomycorrhiza! fungi have the enzymic potential to the hypothesis that GS and NADPH-GDH provideincorporate ammonium into glutamate either by the alternative ports for the entry of ammonium inreductive animation reaction of glutamate dehydro- ectomycorrhizal fungi belonging to both asco-genase (GDH), where ammonium is added directly mycetes and basidiomycetes (Genetet, Martin &to the a-amino position of 2-oxoglutarate, or via the Stewart, 1984; Martin et al., 1988; Ahmad et aL,glutamine synthetase (GS)/glutamate synthase 1990; Chalot et ai, 1991 ; Brun et al., 1992; Martin,(GOGAT) cycle, where ammonium is first added by Cote & Canet, 1994).GS to the amide position of glutamine and then The operation of both the GDH pathway and thetransferred by GOGAT to the a-amino position of GS/GOGAT cycle has been shown to be regulated2-oxoglutarate (Ahmad & Hellebust, 1991). How- by nutrient supply and growth conditions in fungi.ever, the role of GDH enzymes in ammonium The ^^NH4 * labelling patterns in the ectomycorrhizalassimilation of ectomycorrhizal fungi remains con- ascomycete Cenococcum geophilum suggest the com-troversial. The studies of Kershaw& Stewart (1992) bined operation of NADPH-GDH and GS inand Chalot e( a/. (1994) suggest that GS/GOGAT is rapidly growing mycelia (Martin et al, 1988),the primary pathway for ammonium assimilation in whereas at stationary phase in high ammoniumtwo ectomycorrhizal basidiomycetes. On tbe other media ammonium is assimilated primarily via the
NADPH-GDH pathway (Genetet et al., 1984). In* To whom correspondence should be addressed. comparison, the ectomycorrhizal basidiomycete

264 A. M. Quoreshi and others
Laccaria bicolor shows high levels of both GSand NADPH-GDH when ammonium is replaced bynitrate as the nitrogen source, and when both nitrateand ammonium are present the activities of GS andNADPH-GDH are greater in rapidly growinghyphae than in stationary cultures (Ahmad et al.,1990), A somewhat similar induction of GS andNADPH-GDH activities has been show n to occur inthe basidiomycete Hebeloma cylindrosporum whenammonium is replaced by glutamate (Chalot et al.,1991). It appears, therefore, that both the externalnitrogen source and the fungal growth status have amajor influence on the participation of GS andNADPH-GDH pathways in ectomycorrhizal fungi.
A marked induction of NADH-GDH and alanineaminotransferase (AlAT) has been reported in L.bicolor during post-exponential growth periods or inthe absence of glucose (Ahmad et al., 1990). In fungi,the activity of NADH-GDH is associated pritnarilywith the degradation of glutamate, and their AlATand aspartate aminotransferase (AsAT) activitiesplay a dominant role in the intermediar\' metabolismof amino acids (Ahmad & Hellebust, 1991), Thelevels of AsAT are also found to be stronglyregulated by growth conditions in mycorrhizal fungi(Chalot. Botton & Banvoy, 1989; Ahmad et al.,1990). These studies indicate an important linkbetween growth conditions and amino acid metab-olism in ectomycorrhizal fungi.
The present paper deals with the influence ofnitrogen sources on growth and nitrogen metabolismm the basidiomycete Hebeloma crustubniforme, anectomycorrhizal member of the family Corti-nariaceae. Ectomycorrhizal species oi Hebeloma grow-well in forest soils with a large input of ammoniumor other reduced sources of nitrogen (Sagara, 1992).The release of a highly active proteinase byH. crustuliniforme (Zhu, Guo & Dancik, 1990)indicates its acquisition of amino acids for itsnitrogen and carbon needs. The utilization of bothinorganic and organic nitrogen sources, therefore,appears to be a major feature of symbiotic re-lationships of this ectomycorrhizal fungus.
M A T E R I A L S AND M E T H O D SThe fungus
Hebeloma crustuliniforme (Bull. ex. Amans) Quel.was obtained from our own collection at theUniversity of Toronto. The fungus was maintainedfor 5 yr on MN (Melin-Norkrans) medium (seeHutchison, 1989) before initiating its culture underthe conditions described below.
Growth conditions and biomass measurements
The fungus was grown axenically in unstirred liquidmedium described by Ahmad & Hellebust (1991).The inoculum was prepared under axenic conditions
by blending a stationary phase culture and incu-bating the fine suspension produced for 3 d asdescribed by Ahmad et al. (1990), Four 5 mlsuspension aliquots were transferred to 100 mlculture solutions in 250 n:i] flasks plugged with foamstoppers, and incubated in the dark at 22 °C. Thecultures were shaken gently for 30 s daily to provideoxygenation during the exponential period of growthand at least every 2 d in the later period (Ahmad etal., 1990). Cultures were harvested by centrifugationand the packed cell volume was measured accordingto Ahmad et al. (1990).
Preparation of cell-free extracts and proteinfractionations
Fungal mycelia (0-3^ ml packed cell volume) wereground in 2-12 ml of extraction buffer (pH 7-6), andclarified cell-free preparations were obtained ac-cording to Ahmad et al. (1990). The preparation waseither used as a crude extract for enzyme assays, orpassed through a 0 22 //m filter assembly and loadedonto a Mono-Q* column attached to a fast liquidprotein chromatography (FPLC) system for ion-exchange fractionation of enzyme activities.
The FPLC media were prepared according toAhmad & Hellebust (1991). For GS isolation, theaddition of NH^Cl to a final concentration of 25 mMto these media was found to be necessary for thestability of GS activity during chromatographicprocedures. The salt gradient and fraction collec-tions were as described by Ahmad et al. (1990).
Enzyme assays and chemical analysis
The synthetase and transferase activities of glu-tamine synthetase, and aminating activities ofNADH-GDH and NADPH-GDH were determinedas described in Ahmad & Hellebust (1991). Oxoglu-tarate-dependent activities of aspartate amino-transferase (AsAT) and alanine aminotransferase(AlAT) were determined by coupled enzyme pro-cedures described by Ahmad & Hellebust (1991).Nitrate, ammonium, a-amino nitrogen and glucosein the growth medium and protein in cell-freeextract were determined as described in Ahmad et al.(1990).
RESULTS
Utilization of inorganic nitrogen sources
Because of the reported preference of ectomycor-rhizal Hebeloma species for the presence of largeconcentrations of ammonium (Sagara 1992), theinorganic N medium of Ahmad et al. (1990) wasmodified to contain 10 mM of either ammonium ornitrate as the nitrogen source. With glucose as thecarbon source, Hebeloma crustuliniforme grew rapidlywith either nitrate or ammonium as the nitrogensource, exhibiting a doubling time of 1-2 d during
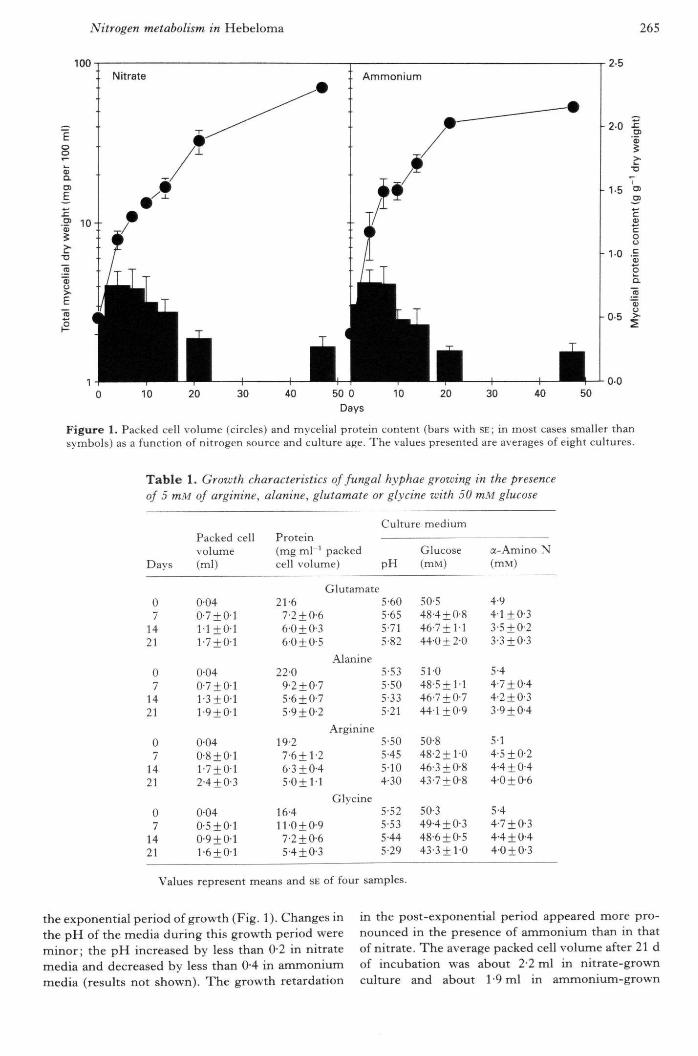
Nitrogen metabolism in Hebeloma 265
100
10 40 50
Figure 1. Packed cell volume (circles) and mycelial protein content (bars with SE; in most cases smaller thansymbols) as a function of nitrogen source and culture age. The values presetited are averages of eight cultures.
Table 1. Grotvth characteristics of fungal hyphae growing in the presenceof 5 mM of arginine, alanine, glutamate or glycine with 50 mM glucose
Days
07
1421
07
1421
07
1421
07
1421
Packed cellvolume(ml)
0'040 7 + 0'lM+0-117+0-1
0040-7 ±0-1l-3±011-9 + 0-1
0040-8+ 0-117 + 012-4 + 0-3
0-040-5 + 0-10-9 + 0-11-6 + 0-1
Protein{mg ml~^ packedcell volume)
Glutamate21-6
7-2 + 0-66-0 + 0-36-0 + 0-5
Alanine22-09-2 + 0-75-6 + 0-75-9 ±0-2
Arginine19-27-6+1-26-3 + 0-45-0+1-1
Glvcine16-4ll-0 + 0'97-2 + 0-65-4 ±0-3
Culture medium
p H
5-605'655-715'82
5'535'5O5-335-21
5-505-455-104-30
5-525-535-445-29
Glucose(mM)
50-548-4 ±0'846-7±M44-0 ±2'0
51-048-5 ±1-146'7±0744'l±0'9
50'848'2±l-046-3+0-8437±0-8
503494 + 0-348-6 + 0-543-3 + 1-0
a-Amino N(mM)
4-94-1+0-33-5 + 0-23-3 + 0-3
5'44-7 + 0-44'2±0'33'9±0-4
5'14-5 ±0-24-4 ±0-44-0 ±0-6
5 44-7 ±0-344 + 044-0 + 0-3
Values represent means and SE of four samples.
the exponential period of growth (Fig. I). Changes inthe pH of the media during this growth period wereminor; the pH increased by less than 0-2 in nitratemedia and decreased by less than 0-4 in ammoniummedia (results not shown). The growth retardation
in the post-exponential period appeared more pro-nounced in the presence of ammonium than in thatof nitrate. The average packed cell volume after 21 dof incubation was about 2-2 ml in nitrate-grownculture and about 1 -9 ml in ammonium-grown
266 A. M. Quoreshi and others
culture (Fig. 1). The mycelial protein content was inthe range of 10-11 mgml"' packed cell volumeduring the exponential phase of growth, and declinedgradually to about 5 mg ml"^ packed cell volume atthe end of 21 d growth period (Fig. 1). The growthretardation and lower mycelial protein contents inthe post-exponential period were not a result ofdeficiency of either nitrogen or carhon in themedium. With 10 mM of either nitrate or ammoniumand 50 mM glucose, f. 7-7 mM nitrate, 6-5 mMammonium and 35 mM glucose remained in themedia at the end of 21 d growth period (results notshown). The pH of the media, adjusted initially to5 5, rose to about 6 in nitrate media and declined toabout 3 in ammonium media (results not shown).These changes indicate that 2 mM 2-(A'^-morpholino)ethanesulphonic acid (MES) buffer (pK value ofc. 61 at 25 °C) in these fungal cultures was moreeffective in controlling the rise of pH in nitrate mediathan the decrease of pH in ammonium media thatresulted from the removal of the respective nitrogensources in these two media. It appears, therefore,that a more pronounced post-exponential growthretardation in the presence of ammonium comparedwith nitrate (Fig. 1) might at least in part beassociated with a large decline in the pH ofammonium media.
Utilization of amino acids as sole nitrogen sources
With 50 mM glucose as the carbon source, tbe growthof H. crustuliniforme was typically rapid in thepresence of 5 mM of alanine, glutamate, arginine orglycine as the sole nitrogen source (Table 1). Underthese conditions, the mycelial protein contentsduring exponential and post-exponential periods,and the biomass yield at the end of 21 d period weresimilar to those found in inorganic nitrogen media(Table 1, Fig. 1). About 30"o of glutamate andalanine, 25 "o of glycine and 20* 0 of arginine fromrespective nriedia, and about 14^0 of glucose from allfour media were removed by the fungus during thisperiod of growth (Table 1). The pH of the argininemedium dropped from 5 5 to 4 3 over 21 d, whereaspH showed little change in the other three media.
Amino acids as sole carbon and nitrogen sources
In the absence of glucose but with 50 mM of eitherglutamate, alanine, arginine or glycine providing thesole source of both carbon and nitrogen, H. crustu-liniforme suffered a considerable growth lag, and itsgrowth in the subsequent period was much less rapidthan that of cultures growing in the presence ofglucose (Tables 1, 2). Moreover, the onset ofthe stationary phase in these glucose-free amino acidmedia occurred with just over 1 ml packed cellvolume in the presence of glutannate and arginineand even less when alanine and glycine were presentas the amino acid source. Consistent with these
Table 2. Growth characteristics of fungal hyphaegrowing in the presence of 50 mM of glutamate, alanine,arginine or glycine as sources of carbon and nitrogen
Days
0102850
0102040
0101421
01028
Packed cellvolume(ml)
0'050-5+0-1O-8 + O'l0-9 + 01
0-50-8 ±0-1I-O + O-l1-3 + 0-1
0-05O'6 + OlM+0-11-7 + 0-1
0-040-4 ±0-10-5 + 0-1
Protein(mg ml"' packedcell volume)
Glutamate1158-6 + 0-97-5 ±0-58-6 ±0-8
Alanine11-55-6 + 0-65-4 ± 0-45-0 ±0-6
Arginine11-587 + 1-06-0 + 0-36-0+ 0-5
GIvcine21'6
9-2 + 0-69-3+0'3
Culture medium
a-AtninoNitrogen(mM)
50-848-7 + 0-947-2 + 0-746-2 + 1-0
51'347'3 + 0-644'5 + M42-1+l>0
50-347-0+1-146-7 + 1-144-0+2-0
50'549-8 ±0'648-1+0'4
N H /(mM)
O'OO0-030'200-60
0-000-060-200-50
0-000-100-300-60
0-000-020-05
Values represent means and SE of four samples.
smaller biomass yields, only about 9, 17, 14 and 7%of alanine, glutamate, arginine and glycine, re-spectively, were removed during the given grow^thperiods (Table 2). Under these catabolic conditions,a considerable amount of ammonium was releasedhy tbe fungus to the media, particularly in thepresence of alanine, glutamate and arginine.
Enzymology of nitrogen assimilation
Ion-exchange chromatography of a cell-free prep-aration of nitrate-grown H. crustuliniforme fraction-ated glutamine synthetase activity as an anionicprotein which eluted as a single peak at c. 180mMNaCl, and exhibited a transferase (GSt, non-physiological activity) to synthetase (GSs, physio-logical activity) ratio of c. 6-5 :1 (results not shown).Glutamate dehydrogenase fractionation of an am-monium-grown fungal extract showed the elution ofNADH-GDH at <r. 140mM NaCI and that ofNADPH-GDH at c. 260 mM NaCl. Both GDHenzymes were highly specific for their respectivecoenzyme. Aspartate aminotransferase activity inH. crustuliniforme is fractionated into two lon-exchangeable forms, a predominant neutral orcationic molecular form that did not bind to theMono-Q column, and a minor anionic form whichetuted at 120mM NaCl. Alanine aminotransferaseactivity was also fractionated into two isoforms, a pre-dominant form which eluted at 200 mM NaCl, anda minor form eluting at 280 mM NaCl.
Nitrogen metabolism in Hebeloma 267
15- •150
14 21 0 14
Days
21 0 14 21
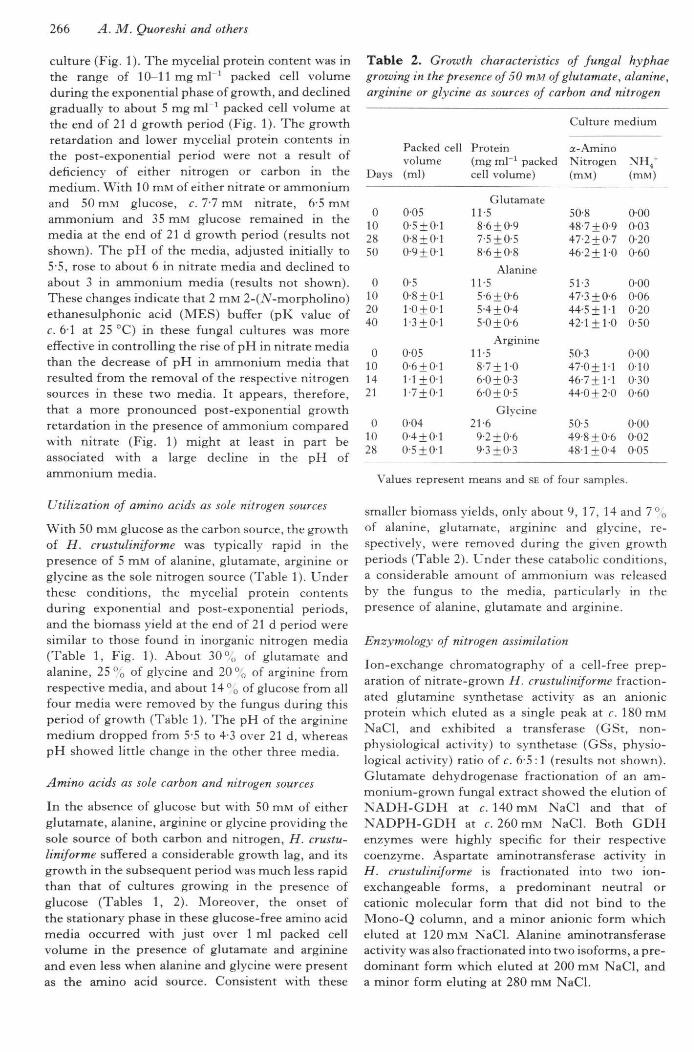
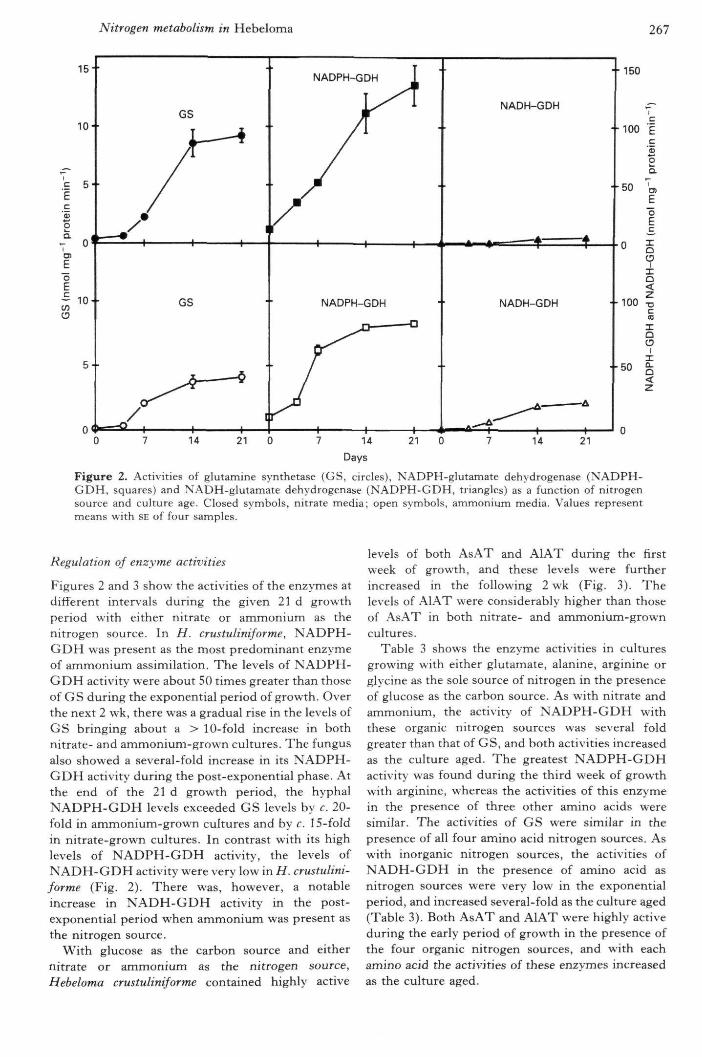
Figure 2. Activities of glutamine synthetase (GS, circles), NADPH-glutamate dehydrogenase (KADPH-GDH, squares) and NADH-glutamate dehydrogenase (NADPH-GDH, triangles) as a function of nitrogensource and culture age. Closed symbols, nitrate media; open symbols, ammonium media. Values representmeans with SE of four samples.
Regulation of enzyme activities
Figures 2 and 3 show the activities of the enzymes atdifferent intervals during the given 21 d growthperiod with either nitrate or ammonium as thenitrogen source. In H. crustuliniforme, NADPH-GDH was present as the most predominant enzymeof ammonium assimilation. The levels of NADPH-GDH activity were about 50 times greater than thoseof GS during tbe exponential period of growth. Overtbe next 2 wk, tbere was a gradual rise in the levels ofGS bringing about a > 10-fold increase in bothnitrate- and ammonium-grown cultures. Tbe fungusalso showed a several-fold increase in its NADPH-GDH activity during tbe post-exponential pbase. Attbe end of the 21 d growth period, the hypbalNADPH-GDH levels exceeded GS levels by c. 20-fold in ammonium-grown cultures and by c. 15-foidin nitrate-grown cultures. In contrast with its highlevels of NADPH-GDH activity, the levels ofNADH-GDH activity were very low in H. crustulini-forme (Fig. 2). There was, however, a notableincrease in NADH-GDH activity in the post-exponential period when ammonium was present asthe nitrogen source.
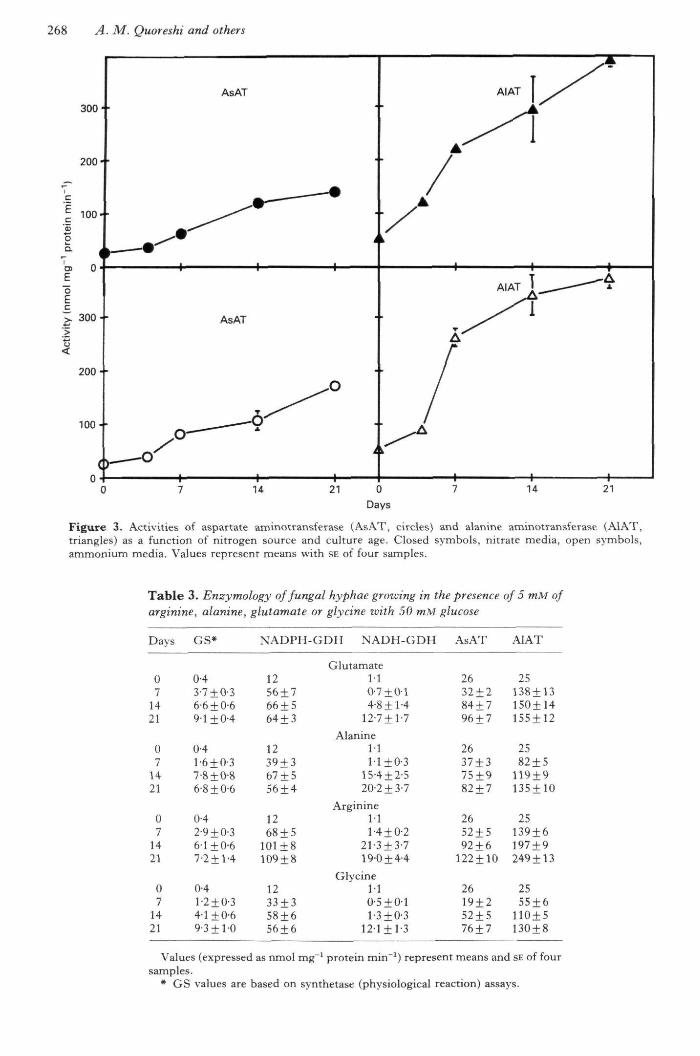
With glucose as the carbon source and eithernitrate or ammonium as the nitrogen source,Hebeloma crustuliniforme contained highly active
levels of both AsAT and AlAT during the firstweek of growth, and these levels were furtherincreased in the following 2 wk (Fig. 3). Thelevels of AlAT were considerably bigher tban thoseof AsAT in both nitrate- and ammonium-growncultures.
Table 3 shows the enzyme activities in culturesgrowing with either glutamate, alanine, arginine orglycine as tbe sole source of nitrogen in the presenceof glucose as the carbon source. As with nitrate andammonium, the activity of NADPH-GDH withthese organic nitrogen sources was several foldgreater than that of GS, and botb activities increasedas tbe culture aged. Tbe greatest NADPH-GDHactivity was found during the tbird week of growtbwith arginine, whereas the activities of this enzymein the presence of three otber aniino acids weresimilar. Tbe acrivities of GS were similar in thepresence of all four amino acid nitrogen sources. Aswith inorganic nitrogen sources, the activities ofNADH-GDH in tbe presence of amino acid asnitrogen sources were very low in tbe exponentialperiod, and increased several-fold as tbe culture aged(Table 3). Both AsAT and AlAT were highly activeduring the early period of growth in tbe presence ofthe four organic nitrogen sources, and with eachamino acid tbe activities of tbese enzymes increasedas tbe culture aged.
268 A. M. Quoreshi and others
AsAT
300' •
200-
100-
AIAT
AiAT
300-
200-
100-
AsAT
0
Days
21
Figure 3. Activities of aspartate aminotransferase (AsAT, circles) and alanine aminotransfeiase (AIAT,triangles) as a function of nitrogen source and culture age. Closed symbols, nitrate media, open symbols,ammonium media. Values represent means with SE of four samples.
Table 3. Enzymology of fungal hyphae growing in the presence of 5 mM ofarginine, alanine, glutamate or glycine with 50 mM glucose
Days
07
1421
07
21
07
1421
07
1421
G S *
0-43-7 + 0-36-6 + 0-69'l+0-4
0-41-6 + 0-37-8 ±0-86-8 ±0-6
0-42-9 + 0-36-1+0-67-2 + 1-4
0-41-2 +0-34-1 ±0-69'3±l-0
NADPH-GDH NADH-GDH
12S6±766 ±564 ±3
1239 + 367 + 556 ±4
1268 ±5
101 ±8109 + 8
1233 + 358 + 656 ±6
GlutamateM0-7 ±0-14-8 + 1-4
12-7±l-7Alanine
M1-1+0-3
15-4 + 2-520-2±3-7
ArginineMl-4±0'2
21-3±3-719-0 + 4-4
GlycineM0-5 + 0-11-3 + 0-3
121 + 1-3
AsAT
2632+284 + 796 ±7
2637 + 375+982±7
2652 + 592±6
122+10
2619 + 252 + 576 + 7
AIAT
25n8±13150+14155 ±12
2582 ±5
119 + 9135±1O
25139±6197±9249 ±13
2555±6
110 + 5130 + 8
Values (expressed as nmol mg"' protein min ) represent means and SE of foursamples.
* GS values are based on synthetase (physiological reaction) assays.
Nitrogen metabolism in Hebeloma 269
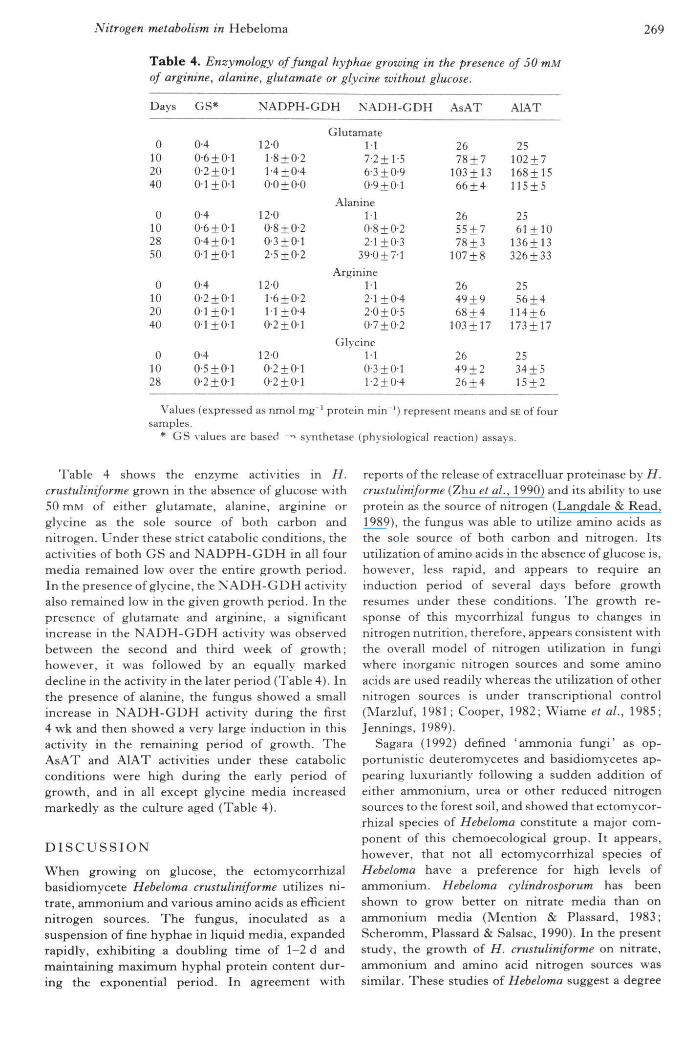
Table 4. Enzymology of fungal hyphae growing in the presence of 50 mMof arginine, alanine, glutamate or glycine without glucose.
Days
0102040
0102850
0102040
01028
GS*
0-40-6 + 010-2 + 010 1 + 0 1
0-40-6±0'l0'4±0'lO'l+O'l
0-4O'2 + O'lO-l+O'l0-1 ±0-1
0 40-5+ 0-10 2 + 01
NADPH-GDH NADH-GDH
12-0l-8 + 0'214 ±040'0±0-0
1200-8 ±0-20-3+0-12-5 + 0-2
12-01-6 + 0-21-1+040-2±0-l
12-0O'2 + O-lO'2 + O'l
GlutamateM7-2+1-56-3+0-90-9 + 0-1
AlanineM0-8 + 0-22-1+0-3
39-0 + 7-1
ArginineM2-1+042-0 ±0-50-7±0'2
GlycineM0'3±0-ll '2±04
AsAT
2678 + 7
103 + 1366±4
2655 + 778 + 3
]07±8
2649 + 968 + 4
1O3±]7
2649 + 226 + 4
AlAT
25102 + 7168+15115 ± 5
2561+10
136+13326 ±33
2556 + 4
114 + 6173±17
2534+515 + 2
Values (expressed as nmol mg ' protein min ') represent means and SE of foursamples.
* GS values are baseti -> synthetase (physiological reaction) assays.
Table 4 shows the enzyme activities in H.crustuliniforme grown in the absence of glucose with50 mM of either glutamate, alanine, arginine orglycine as the sole source of both carbon andnitrogen. Under these strict catabolic conditions, theactivities of both GS and NADPH-GDH in all fourmedia remained low over the entire growth period.In the presence of glycine, the NADH-GDH activity'also remained low in the given growth period, ln thepresence of glutamate and arginine, a significantincrease in the NADH-GDH activity was observedbetween the second and third week of growth;however, it was followed by an equally markeddecline in the activity in the later period (Table 4). Inthe presence of alanine, the fungus showed a smallincrease in NADH-GDH activity during the first4 wk and then showed a ver>' large induction in thisactivity in the remaining period of growth. TheAsAT and AlAT activities under these catabolicconditions were high during the early period ofgrowth, and in all except glycine media increasedmarkedly as the culture aged (Table 4).
D I S C U S S I O N
When growing on glucose, the ectomycorrhizalbasidiomycete Hebeloma crustuliniforme utilizes ni-trate, ammonium and various amino acids as efficientnitrogen sources. The fungus, inoculated as asuspension of fine hyphae in liquid media, expandedrapidly, exhibiting a doubling time of 1-2 d andmaintaining maximum hyphal protein content dur-ing the exponential period. In agreement with
reports of the release of extracelluar proteinase by H.crustuliniforme (Zhu et al., 1990) and its ability to useprotein as the source of nitrogen (Langdale & Read,1989). the fungus was able to utilize amino acids asthe sole source of both carbon and nitrogen. Itsutilization of amino acids in the absence of glucose is,however, less rapid, and appears to require aninduction period of several days before growthresumes under these conditions. The growth re-sponse of this mycorrhizal fungus to changes innitrogen nutrition, therefore, appears consistent withthe overall model of nitrogen utilization in fungiwhere inorganic nitrogen sources and some aminoacids are used readily whereas the utilization of othernitrogen sources is under transcriptional control(Marzluf, 1981; Cooper, 1982; Wiame et aL, 1985;Jennings, 1989),
Sagara (1992) defined 'ammonia fungi' as op-portunistic deuteromycetes and basidiomycetes ap-pearing luxuriantly following a sudden addition ofeither ammonium, urea or other reduced nitrogensources to the forest soil, and showed that ectomycor-rhizal species of Hebeloma constitute a major com-ponent of this chemoecological group. It appears,however, that not all ectomycorrhizal species ofHebeloma have a preference for high levels ofammonium, Hebeloma cylindrosporum has beenshown to grow better on nitrate media than onammonium media (Mention & Plassard, 1983;Scheromm, Plassard & Salsac, 1990). In the presentstudy, the growth of H. crustuliniforme on nitrate.ammonium and amino acid nitrogen sources wassimilar. These studies of Hebeloma suggest a degree

270 A. M. Quoreshi and others
of interspecific variation in the preference fornitrogen sources, and, by implication, in the operation of nitrogen metabolism in this largely myco-rrhizal genus of the Cortinariaceae.
According to Chalot et al. (1991), high activities ofboth GS and NADPH-GDH are present in rapidlygrowing mycelia of H. cylindrosporum. The presentstudy shows that a highly active NADPH-GDH isalso present in the mycelia of H. crustuliniformegrowing exponentially with different nitrogensources in the presence of glucose. However,its GS activities under these conditions arever\' low. Moreover, with each nitrogen source,H. crustuliniforme exhibits highest activities ofboth GS and NADPH-GDH in the post-exponentialperiod, whereas the reverse is true for amtnonium-and glutamate-grown H. cylindrosporum {Chalotet al., 1991). These results indicate a divergence inboth the expression and regulation of GS andNADPH-GDH in these two closely related speciesof ectomycorrhiza! fungi.
Our study of the relative role of GS and NADPH-GDH in H. crustuliniforme is based on in vitroenzyme activities. In Laccaria bicolor, the in vitrodata suggesting a simultaneous operation of GS andNADPH-GDH pathways in ammonium assimi-lation (Ahmad et al., 1990) were recently show^n to begenerally consistent with the ^^NH^-labelling pat-terns of this fungus (Martin et al., 1994). Obviously,the iti vitro data on GS and NADPH-GDH activitiesin H. crustuliniforme presented here also providelargely circumstantial evidence that needs to bevalidated by ^"NH^-labelling analysis as well as byenzyme inhibitors of this fungus. Barring this caveat,the ability of H. crustuliniforme to grow rapidly onammonium and other nitrogen sources, apparentlywith very low activity of GS in exponentially-growing mycelia, provides strong circumstantialevidence for the participation of NADPH-GDH inthe assimilation of ammonium by fungi. Our esti-mates of the rate of nitrate or ammonium as-similation, calculated as mycelial N content x growth(/f), for exponentially growing H. crustuliniformemycelia appear greater than the rate of GS activitiesfound in these mycelia. In H. crustuliniforme w itheither nitrate or ammonium as the nitrogen source,the smallest protein content in exponentially grow ingmycelia was about lOmg protein ml"' packed cellvolume, and a doubling time (/^) of about 2 d(2880 min) gave the most conservative estimate ofthe growth rates of these mycelia. Assuming aprotein:nitrogen ratio of 6 25 and calculating /j. aslog^ 2/t^ these values give a mycelial N content of115//niolml~' packed cell volume and a growthconstant (//) of 0-00024 min"\ respectively. Thus aconservative estimate of the rate of nitrate andammonium assimilation in exponentially growingmycelia is about 28 nmol ml"' packed cellvolume min"' or about 2 8 nmol mg"' protein min"'.
The activities of GS in exponentially growingmycelia in ammonium and nitrate media are belowlnmolmg"^ protein min"', whereas those ofNADPH-GDH exceed 20 nmol mg"' proteinObviously, NADPH-GDH is by far the predomi-nant enzyme of ammonium assimilation in H.crustuliniforme. It is interesting that although theammonium repression of GS was shown to be liftedin a NADPH-GDH deficient mutant oi H. cylindro-sporum, the increased activities of GS were unable tocompensate for the lack of NADPH-GDH in thisstrain (Chalot et al., 1991). Clearly, the role ofNADPH-GDH in the assimilation of ammoniumappears to be indispensable in all Hebeloma speciesstudied so far.
In the absence of glucose, the utilization of aminoacids as sole sources of both carbon and nitrogen byH. crustuliniforme appeared to generate an excess ofnitrogen that was evident by a considerable release ofammonium by this fungus to the external media.The activities of both GS and NADPH-GDH wereparticularly low in these cultures, suggesting that there-assimilation of ammonium by the two pathwaysconstituted only a minor part of the amino acidmetabolism under these conditions.
The present study has not addressed therole of GOGAT in glutamate biosynthesis inH. crustuhnifornie. In L. bicolor, the detection ofCiOGAT activity by Vezina, Margolis & Delaney(1989) at less than 1% of the measured GSactivities appears to raise questions concerning themethodology of GOGAT measurements in fungalextracts. A recent '"'"NH^ incorporation study ofL. bicolor indicates a much higher level of glutamateformation by the GS/GOGAT pathway in thisfungus (Martin et al., 1994).
In H. crustuliniforme, low activities of NADH-GDH during the exponential period of growth w ithdifferent nitrogen sources in the presence of glucose,and their increase daring the post-exponential periodappear consistent with the participation of thisenzyme primarily in the catabolic degradation ofglutamate. However, its operation in mycelia grow-ing w'ith amino acids in the absence of glucoseappears to be variable. Under these conditions, thefungus shows a inarked increase in NADH-GDHactivity during the mid-exponential period of growthwhen glutamate and arginine are present, but theonset of stationary phase in these media causes anequally marked decline in this activity. In thepresence of alanine, on the other hand, the fungusshows only a small increase of NADH-GDH activityduring the mid-exponential period, and a largeinduction in the stationary period. The reasons forsuch variation in the induction of NADH-GDH inH. crustuliniforme in the absence of glucose are notclear at present.
In ainino acid metabolism, a key link betweencarbohydrate and nitrogenous metabolites is pro-

Nitrogen metabolism in Hebeloma 271
vided by aminotransferase activities (.A,hmad &Hellebust, 1991). As with L. bicolor (Ahmad et aL,1990), H. crustuliniforme maintains high activities ofAsAT and AIAT during exponential growth. Al-though L. bicolor shows the induction of AIATduring the post-exponential period (Ahmad et al.,1990), in H. crustuliniforme the activities of bothAsAT and AIAT increase as the culture ages. Thatthese increases in aminotransferase activities inageing cultures are metabolic alterations for a moreefficient linkage between amino acid pools andcarbon metabolism is further evidenced by a largeinduction of AsAT and AIAT in mycelia growingactively in the absence of glucose with amino acids assole sources of both carbon and nitrogen.
The FPLC fractionation of enzyme activities in H.crustuliniforme separated GS as a single isoform,NADH-GDH and NADPH-GDH as two distinctmolecular forms with high nucleotide specificity,AsAT as a predominantly cationic but with aminor anionic isoform, and AIAT as two anionicisoforms where the predominant one is less neg-atively charged. These chromatographic profilesof GS, GDH and aminotransferase activities inH. crustuliniforme match well with those shown inL. bicolor by Ahmad et al. (1990), indicating a strongsimilarity in the molecular characteristics of theenzymes of nitrogen assimilation in these ectomycor-rhizal fungi.
In conclusion, the utilization of a wide variety ofnitrogen sources in the ectomycorrhizal basidio-mycete Hebeloma crustuliniforme is aided by theregulation of several metabolic pathways, whereNADPH-glutamate dehydrogenase together withglutamine synthetase participates in ammoniumassimilation, NADH-GDH is in\olved in the cata-bolic breakdown of glutamate, and both aspartateaminotransferase and alanine aminotransferase fa-cilitate an active linkage of both biosynthetic andcatabolic pathways with the fungal TCA cycle. Thepattern of alteration in these activities in response tochanges in the nutritional status of the externalmedia is highly distinct in Hebeloma crustuliniforme,and emphasizes the need to understand the complexnature of the regulatory diversity of the enzymologyof nitrogen assimilation in ectomycorrhizal fungi.
ACKNOWLEDGEMENTS
This work was supported by Grant A6032 from theNatural Sciences and Engineering Research Councilof Canada. We gratefully acknowledge Dr L. J.Hutchison for a gift of Hebeloma crustuliniformeaxenic culture and for helpful discussions.
REFERENCES
Ahmad I, Carleton TJ, Malloch DW, Hellebust JA. 1990.Nitrogen metabolism in the ectomycorrhizal fungus Laccariabicolor (R. Mre.) Orton. Neu: Phytologist 116: 431^41 .
Ahmad I, Hellebust JA. 1991. Enzymology of nitrogen as-similation in mycorrhiza. In: Norris JR. Read DJ, Verma AK,eds. Methods in Microbiology - Experiments with Mycorrhiza,Vol. 23. London: Academic Press. 181-202.
Bnm A, Chalot M, Botton B, Martin F. 1992. Purificationand characterization of glutanime synthetase and NADP-glutamate dehydrogenase from the ectomycorrhizal fungusLaccaria laicata. Plant Physiology 99; 938-y44.
Chalot M, Botton B, Banvoy J. 1989. Seasonal fluctuations ofgrowth and nitrogen metabolism in the ectomycorrhizalassociation Douglas f\r-Laccaria laccata. Agriculture, Eco-systems and Emironment 28: 5'j-64.
Chalot M, Brun A, Dehaud JC, Botton B. 1991. Ammonium-assimilating enzymes and their regulation in wild and NADP-glutamatt- dehydrogenase-deficient strains of the ectomycor-rhizal fungus Hebeloma cvlindrosporum. Phvsiologia Plantarunt83: 122-128.
Chalot M, Brun A, Finlay RD, Soderstrom B. 1994. Metabo-lism of ["CJ glutamate and ['^C] glutamine hy theectomycorrhizal fungus Paxillus involutus. Microbiology 140:
Cooper TJ. 1982. Nitrogen metabolism in Saccharomyces cere-visiae. In: Strathern JN. Jones EW. Broach JR, eds. TheMolecular Biology of the Yeast Saccharomvces : Metabolism andGene Expression. Nev\ York; Cold Spring Harbor Laboratory,
Genetet I, Martin F, Stewart GB. 1984. Nitrogen assimilationin mycorrhizas. Ammonium assimilation in the nitrogen starvedectomycorrhizal fungus Cenococcum grariiforme. Plant Physi-ology 76: 3^5-394.
Hutchison LJ. 1989. Studies on the cultural taxonomy ofectomycorrhisal basidiomycetes from temperate North America.Ph.D. thesis. University of Toronto, Canada.
Jennings DH. 1989. Some perspectives on nitrogen and phos-phorus metabolism in fungi. In: Boddy L, Merchant R. ReadDJ, eds. Nitrogen. Phosphorus and Sulphur Utilisation by Fungi.Cambridge: Cambridge University Press, 1-31.
Kershaw JL, Stewart JR. 1992. Metabolism of '^N-labelledammiinium by the ectomycorrhizal fungus Pisolithus tintorius(Pers.) Coker and Couch. Mycorrhiza 1: 71-77.
Langdale AR, Read DJ. 1989. Suhstrate decomposition andproduct release by ericoid and ectomycorrhizal fungi grown onprotein. Agriculture. Ecosystem a7id Etn-ironment 28: 285-291.
Martin F, Cote R, Canet D. 1994. NH,^ assimilation m theectomycorrhizal basidiomycete Laccaria bicolor (Maire) Orton,a ^^N-NMR study. A'pw Phytologist 128: 479-4-85,
Martin F, Stewart GR, Genetet I, Mourot B. 1988. Theinvolvement of glutamate dehydrogenase and glutaminesynthetase in ammonia assimilation hy the rapidly growingascomycete, Cenococcum geophilum Fr, A'e^' Phytologist 110:541-550.
Marzluf GA. 1981. Regulation of nitrogen metabolism and geneexpression in fungi. Microbiological Rerteu's 45: 437^61 ,
Mention M, Plassard C. 1983. Comparison de la nutritionnitrique et ammoniacale de quatre especes de basidiomycetesectomycorhiziens, Comptes Rendus de VAcademie des Sciences297: 489-492.
Sagara N. 1992. Experimental disturbances and epigeous fungi.In; Carroll GC, Wicklow DT, eds. The Eungal Community,New York: Marcel Dekker, Inc., 427^54.
Scheromm P, Plassard C, Salsac L. 1990. Effect of nitrate andammonium nutrition on the metabolism of an ectomycorrhizalbasidiomycete fungus Hebelcima cylindrosporum Roniagn. A'«tPhytologtst 114: 227-234,
Vezina, L-P, Margolis HA, Delaney S. 1989. Changes in theactivity of enzymes involved with primary nitrogen metabolismdue to ectomycorrhizal symbiosis on jack pine seedlings.Phvsiologia Plantarum 75: 55—62.
Wiame JM, Grenson M, Arts Jr. HN. 1985. Nitrogencataboiism repression in yeasts and filamentous fungi. Advancesin Microbial Physiology 26: 1-87.
Zhu H, Guo D, Dancik BP. 1990. Purification and characteriza-tion of an txtraceliuar acid proteinase from the ectomycorrhizalfungus Hebeloma crustuliniforme. Applied and EnvironmentalMicrobiology 56: 837-843.
