ectomycorrhizal communities associated with populus tremula growing on a heavy metal contaminated...
TRANSCRIPT

m y c o l o g i c a l r e s e a r c h 1 1 2 ( 2 0 0 8 ) 1 0 6 9 – 1 0 7 9
journa l homepage : www.e l sev i er . com/ loca te /mycres
Ectomycorrhizal communities associated with Populustremula growing on a heavy metal contaminated site
Doris KRPATAa,*, Ursula PEINTNERa, Ingrid LANGERb, Walter J. FITZb,Peter SCHWEIGERb
aInstitute of Microbiology, University Innsbruck, Technikerstrasse 25, 6020 Innsbruck, AustriabDepartment of Forest and Soil Sciences, University of Natural Resources and Applied Life Sciences,
Peter Jordan-Strasse 82, 1190 Vienna, Austria
a r t i c l e i n f o
Article history:
Received 30 October 2007
Received in revised form
30 January 2008
Accepted 14 February 2008
Corresponding Editor:
Derek T. Mitchell
Keywords:
Aspen
Ectomycorrhiza
ECM
Fungal community
Heavy metal
Populus tremula
Soil horizon
* Corresponding author.E-mail address: [email protected]
0953-7562/$ – see front matter ª 2008 The Bdoi:10.1016/j.mycres.2008.02.004
a b s t r a c t
European aspen is one of the most widely distributed trees in Central Europe and is a typical
early colonizer of poor and disturbed soils. However, little is known about ectomycorrhizal
(ECM) fungi in these ecosystems. We examined the ECM community of European aspen
growing on a heavily contaminated site in southern Austria by analysing ECM roots, sorting
them into morphotypes, subjecting them to DNA extraction, PCR, and DNA sequencing.
ECM root symbionts were sampled two times in 2004. During this time, the below-ground
community structure was relatively stable; we found no evidence of taxa adapted to sum-
mer or autumn conditions and only two species varied widely in occurrence between soil
horizons. The ECM fungal community was diverse (54 species), rich in Basidiomycota (43
species), and dominated by Cenococcum geophilum and fungi with corticoid basidiomes
(e.g. Thelephoraceae).
ª 2008 The British Mycological Society. Published by Elsevier Ltd. All rights reserved.
Introduction shifts in species composition (Smit et al. 1997; Turpeinen
Mining and smelting operations have resulted in a large
number of sites contaminated with heavy metals (some
100 K potentially contaminated sites in Western Europe)
(Quercia et al. 2006). Soil metal concentrations on these sites
frequently reach levels toxic to both plants (Cobbett 2003)
and microorganisms (Giller et al. 1998; Dai et al. 2004). As a con-
sequence, a reduced diversity of microorganisms (Moffett et al.
2003) has frequently been observed on such sites. Differences
in metal tolerance between organisms additionally result in
ritish Mycological Society
et al. 2004). Severe zinc (Zn) pollution can trigger the evolution
of an increased Zn tolerance in ECM fungi as was shown by
Colpaert et al. (2004, 2005) and Adriaensen et al. (2006) for Suil-
lus isolates. However, data concerning the effect of heavy
metal pollution on ectomycorrhizal (ECM) fungal communi-
ties are controversial: negative effects were reported by
Hartley et al. (1997), Hartley-Whitaker et al. (2000), and
Dunabeitia et al. (2004). Moreover, Ruhling & Soderstrom
(1990) reported a decrease of fruit body-producing species
with increased soil metal concentrations. This is in contrast
. Published by Elsevier Ltd. All rights reserved.

1070 D. Krpata et al.
to other data (Meharg 2003) reporting a high ECM fungal diver-
sity on metal contaminated sites.
Numerous studies have examined the above- as well as the
below-ground community structure of ECM fungi (Gardes &
Bruns 1996; Kernaghan & Harper 2001; Peter et al. 2001). Most
of these studies were conducted on coniferous trees with
only a few studies on broadleaf tree species. Of these, Populus
spp. have received some attention. Kaldorf et al. (2004) exam-
ined the ECM fungal community of transgenic hybrid aspen
(Populus tremula� P. tremuloides) clones planted on agricultural
land. They differentiated 23 ECM types by morphology, as well
as molecular methods, and found the community to be dom-
inated by a Tomentella sp. and Phialocephala fortinii. Other stud-
ies report slightly lower numbers of ECM types associated
with poplars (12–19) (Jakucs 2002; Kernaghan et al. 2003;
DeBellis et al. 2006). This compares with 43 putative mycorrhi-
zal fungi that produced fruit bodies in Populus tremuloides
dominated forests (Cripps & Miller 1993).
Poplars are of scientific interest because of the relatively
small size of their genome (Martin et al. 2004) and also because
of applied interest based on their use in short rotation forestry
(Dickmann 2006; French et al. 2006). They have further been
used for pollution monitoring (Madejon et al. 2004) and are
considered suitable for use in phytoremediation (Laureysens
et al. 2004). Screenings of Populus spp. for metal tolerance
and uptake revealed large differences between species and
clones (Djingova et al. 1999; Bittsanszky et al. 2005). Especially
a clone of European aspen (Populus tremula), collected from
a severely polluted site in southern Austria, was found to
accumulate and tolerate high concentrations of cadmium,
lead, and zinc in the leaves (Unterbrunner et al. 2006). Euro-
pean aspen is one of the most widely distributed trees in
Central Europe (Worrell 1995) and is a typical early colonizer
of poor and disturbed soils.
Few studies have investigated the ECM communities of
heavy metal disturbed sites: Mleczko (2004) investigated my-
corrhizal and saprobic macrofungi of two zinc wastes in
Southern Poland. He described 14 ECM types associated with
Pinus sylvestris and Betula pendula. Staudenrausch et al. (2005)
found 23 ECM types associated with Betula pendula at a ura-
nium mining heap.
ECM fungi are known to affect soil bioavailability and plant
uptake of heavy metals (Jentschke & Godbold 2000). Therefore,
they are predicted to play an important role in phytoremedia-
tion activities involving mycorrhizal hosts (Perotto & Martino
2001), such as, for example, European aspen. This conclusion
was reached based on the large amount of external hyphae
that stabilise soil structure and very effectively exploit soil
nutrients. Additionally, ECM fungi transfer elements very effi-
ciently directly to root cells. Tolerance to high soil metal con-
centrations and good functional compatibility with the plant
used in the phytoremediation activity are prerequisites for
effective ECM fungal symbionts (Khan et al. 2000). ECM fungi
associated with European aspen growing in metal-polluted
soil are expected to fit these characteristics.
In the wider context of a project aiming at testing the influ-
ence of mycorrhizal associations on metal uptake of poplars
suitable for phytoremediation purposes, the ECM fungal com-
munity on roots of European aspen growing on a heavily con-
taminated site in southern Austria was characterised by PCR
and ITS sequence analysis. The data were further used to eval-
uate the impact of soil factors (heavy metal content) on the
formation of the ECM community in organic and mineral
soil layers. The generated data are subsequently compared
with studies on diversity measures of ECM fungal communi-
ties from uncontaminated sites.
Materials and methods
Study site
The study site, which is next to the former lead smelter, is
characterised by a very rough relief formed by prehistoric
rockslide material from the mountain Dobratsch in 1348
(Krainer 1998). This area in Arnoldstein (Austria; N46 330 E13
420) has been affected by lead smelting from the end of the
15th century. In the 1950s smelting activities were extended
to the production of zinc and cadmium. Estimates of annual
emissions from 1989 to 1992, when the smelter was closed,
were 9–13 tonnes lead, 5 tonnes zinc and 10–100 kg cadmium
(Kasperowski 1993). Additionally, different sorts of wastes
such as slag materials were deposited on the site. All of these
activities have resulted in very high soil heavy metal concen-
trations (Kasperowski 1993). Smelting also resulted in severe
sulphur dioxide (SO2) emissions, which left the area mainly
deforested until 1979 when SO2 emissions were cut. From
then on, the barren land was partly re-vegetated by European
aspen (Populus tremula), goat willow (Salix caprea), and grami-
naceous species, such as Brachypodium pinnatum.
Three homogenous stands of aspen (A, B, C,) in the vicinity
of the former lead/zinc smelter were selected as sample plots
(approx. 250 m2 each). Aspen is clonal, and the stands could
well represent one individual with numerous ramets spring-
ing from a single root system. All ramets of aspen on the plots
were very similar in size and height. The estimated age of the
trees was about 20–25 y. In addition to aspen, few individuals
of other ECM species, such as Betula pendula, Salix caprea, and
Corylus avellana, and some saplings of Quercus robur and
Fraxinus excelsior grew on the plots. The soil of the sample plots
is a calcaric Cambisol [pH(H2O) 6.7–7.3] covered by a 5–8 cm
thick mor-like humus layer [pH(H2O) 5.8–6.2].
Soil sampling and physicochemical analysis
Each site was sampled following the same design. Samples
were taken from five randomly distributed spots using a soil
auger with an inner diameter of 7.5 cm. Auger cores were sep-
arated into the organic humus horizon (mor) and mineral soil
(10 cm) and stored in closed plastic bags. On site B a profile
was described and classified following the keys of the world
reference base for soil resources (FAO 1998). A representative
sample was taken from each horizon. Samples were trans-
ported in cooling boxes and stored thereafter in a cooling
chamber at 4 �C.
After homogenisation (mixing and, in the case of the
mineral horizons, sieving to <2 mm) soil subsamples were
air-dried for physicochemical characterisation following
standard procedures. Additionally, humus and mineral soils
were digested in acid mixtures of HClO4/HNO3 and HCl/HNO3,

Populus tremula growing on a heavy metal contaminated site 1071
respectively (Blum et al. 1996). NH4NO3 extracts of mineral
soils were carried out according to DIN 19730 (1995). Analysis
of lead, zinc, and cadmium was performed using inductively
coupled plasma mass spectrometry (ICPMS, Elan 9000 DRCe,
Perkin Elmer, Norwalk, USA).
Sampling
Fungal fruiting bodies were collected in 2004 and 2005. Identi-
fication of these was based on careful macro- and microscop-
ical analysis of morphological characters. Voucher material of
critical species is deposited in the herbarium IB (University of
Innsbruck).
In June 2004 (summer),a totalof15 soil blocks werecollected:
five blocks per plot (A, B, C), each block in close vicinity to one of
five randomly selected aspen trees. Soil blocks were 7.5 cm
square and 15–18 cm deep, including 10 cm deep mineral soil
and varying depths of the organic horizon. In September 2004
(autumn), soil blocks of the same size were taken adjacent to
the summer sampling patches. Each block was divided into
the mineral and the organic soil layer (Baier et al. 2006). These
soil samples were stored in plastic bags at 4 �C for 7 d at the
most until further processing. All roots of a soil sample were
washed out from the soil with tap water over a 2 mm sieve.
Roots of nontarget plants were excluded based on morphologi-
caldifferences. Roots collected in summer from thefive mineral
and organic samples of each plot were pooled, resulting in a to-
tal of three organic and three mineral root samples. Roots from
the autumn sampling were not pooled, resulting in a total of 15
organic and 15 mineral root samples.
For randomisation of selected root tips all roots of a sample
(pooled or not) were evenly distributed in a Petri dish of 14 cm
diam. The dish was subsequently placed on a 1� 1 cm grid
with randomly distributed 1� 1 cm squares marked in red.
All mycorrhizal systems positioned above these red-marked
squares, which represent 6.5 % of the total area of the dish,
were then selected for further analysis. Depending on the
quantity of mycorrhizal root tips in a sample, the procedure
of distributing the roots and selecting those on the red squares
was repeated up to three times to get a minimum number of
root tips (¼ 100 per sample). The number of selected root
tips was recorded for each soil sample. The selected mycorrhi-
zal systems were carefully cleaned under a binocular micro-
scope (SMZ-U, Nikon, Tokyo, Japan) by removing soil and
debris from the roots with tweezers. Individual root tips
were dissected and sorted according to their morphological
characteristics following Agerer (1987–2002): presence and
type of ramification, surface colour, texture, presence of
a mantle, emanating hyphae or rhizomorphs. Only mycorrhi-
zal root tips with the same morphology, from one sample (or
pooled sample) and from the same soil layer were joined,
resulting in a given morphotype sample. Within each mor-
photype sample several individual root tips were prepared
for DNA extraction, while others were subjected to compara-
tive morphological studies. In cases where DNA sequence
analysis gave several results for one presumed morphotype,
the total number of root tips of this morphotype was propor-
tioned according to the number of molecular results. Some
root samples were examined for arbuscular mycorrhizal root
colonisation at �50 magnification following standard clearing
and staining (Gazey et al. 1992). Thus, a total of 12 020 ECM root
tips of Populus tremula were randomly selected and sorted
from 30 soil cores (blocks) during summer and autumn 2004.
Molecular protocols
The rDNA ITS region, situated between the SSU (18S gene) and
the LSU (28S gene) of rRNA genes was amplified using the
primer pair ITS1� ITS4 for fruiting bodies and the primers
ITS1-F� ITS4 for mycorrhizal root tips (White et al. 1990; Gardes
& Bruns 1993). Unamplifiable samples, usually from brown
ECM root tips, were assumed to be dead. Also root tips with
multiple infections were excluded from subsequent analyses.
DNA extraction of fruiting bodies was carried out from
fresh material or dried herbarium specimen following stan-
dard protocols (Zolan & Pukkila 1986).
Of the 12 020 morphotyped ECM root tips, 209 were used for
molecular identification. Root tips of selected morphotypes
not exceeding 5 mm length (Gardes et al. 1991) were stored
in 1.5 ml Eppendorf tubes containing 50 ml CTAB buffer and
either processed immediately or stored at �20 �C. DNA was
extracted from individual root tips following Southworth (2000;
http://www.sou.edu/BIOLOGY/Faculty/Southworth/CTAB.htm).
Usually, only one tip was used for DNA extraction. Several tips
were extracted together only if they were part of one my-
corrhizal system, or if single mycorrhizal tips were very
small (less than 2 mm). The tips were ground in the
1.5 ml Eppendorf tubes containing 50 ml CTAB buffer using
a micropestle. After adding a further 550 ml CTAB buffer
(final concentration per sample: 12.5 mg hexadecyltrime-
thylammoniumbromide, 10 mM Tris–HCl (pH 8), 1.4 M
NaCl, 20 mM EDTA, 0.2 % b-mercaptoethanol) the samples
were incubated at 65 �C for 40–60 min. After centrifugation
for 7 min at 16 000 g the supernatant was precipitated us-
ing an equal volume of chloroform and centrifuged for
15 min at 16 000 g. The upper phase was transferred into
a new tube containing 750 ml cold (�20 �C) 98 % isopropyl
alcohol, which was put in the freezer for precipitation
(30 min to overnight). After centrifugation for 30 min at
16 000 g the pellet was washed with 200 ml cold (�20 �C)
70 % ethanol and centrifuged for 5 min at 7000 g. Superna-
tant was decanted and uncapped tubes invert until dry.
Dried DNA pellets were resuspended in 50 ml TE buffer
and directly used for PCR.
PCR was performed in a volume of 25 ml consisting of 5 ml
DNA and 20 ml PCR mix. The final concentrations were
0.6 mM Tris–HCL, 53.2 mM KCl, 1.5 mM MgCl2, 6.4 mM EDTA,
and 4 % glycerine, as well as 200 mM of each dNTP, 1.25 units
Taq polymerase (peqGOLD, all peqlab, Erlangen) and 0.4 mM
of each primer. PCR reactions were performed using a Techne
Unit Progene thermocycler (Techne, Cambridge) with the
following conditions: an initial step of 5 min at 94 �C was
followed by 40 cycles of denaturation at 94 �C for 1 min,
annealing at 50 �C for 55 s with annealing time increasing 3 s
each cycle, and extension at 72 �C for 45 s. Thermal cycling
was ended by a final extension at 72 �C for 6 min. Purified
PCR products (Amicon� Micron�-PCR Centrifugal filter de-
vices; Millipore, Billerica, MA) were sent to MWG-Biotech
(Ebersberg) for sequence analysis.

1072 D. Krpata et al.
DNA sequence analysis and taxonomic determinationof ECM fungi
After importing the sequences Populus ECM sequences and
closely related sequences obtained from public databases to
Sequencher�, the chromatograms were checked and all se-
quences were automatically aligned. Manual adjustment
was carried out with Se-Al alignment software (Rambaut
1996). Identification of mycorrhizas was based on our in-
house reference database (sequences created from voucher
material), BLAST searches (Altschul et al.1997), comparisons
with the most closely related sequences retrieved from the
public database UNITE (Koljalg et al. 2005), and the sequence
databases at the National Center for Biotechnology Informa-
tion (NCBI: http://www.ncbi.nlm.nih.gov/BLAST/).
ECM root sequences matching sequences from fruiting
bodies were designated by a genus and species epithet fol-
lowed by an IB voucher number. Sequences found on roots
but not as fruiting bodies were named based on data from
BLAST searches, sequence alignments, and advice from taxo-
nomic experts. We used a conservative approach to avoid
overestimation of species richness. Taxon categories were
assigned based on similarities to reference sequences as
follows: ECM fungal isolates were regarded as identified on
species level [1 operational taxonomic unit (OTU)], at se-
quence similarities �97 % with a reference sequence.
Diversity and cluster analyses
The ECM fungal communities associated with the roots were
described by the following five diversity measures: species
richness (S) is the total number of detected species. Simpson’s
Index (1-D) and Fisher’s alpha Index (Alpha Mean) were used
as measures of diversity, and Pielou’s J and the Shannon–
Wiener Index (H) were used as measures of evenness.
Abundance was defined as the total number of ECM root
tips of a species or group, and relative abundance was defined
as the total number of ECM root tips of a species or group
divided by the total number of all ECM root tips of all taxa.
We used relative abundances only and subsequent mentions
of abundance refer to these percentage values.
Frequency was defined as the number of soil cores (blocks)
in which a species or group was detected, and can, therefore,
be regarded as a measure of spatial heterogeneity. For fre-
quency calculations the five soil cores from each of the three
plots were pooled. Organic and mineral layers of pooled sam-
ples were treated separately. Sampling was carried out twice;
therefore, frequency calculations were based on a total of 12
pooled samples.
To estimate the number of undetected species, incidence-
based species richness estimators Jackknife1 (Burnham &
Overton 1979), ICE Mean, and Chao2 (Chao 1987) were calcu-
lated. These indices were performed using the program
EstimateS 8.0 (Colwell 2006).
Detrended correspondence analysis (DCA) was used to un-
ravel trends within the ECM fungal community. We tested
whether data grouped by each of the three categories (plot,
horizon, or season) were statistically different from random
associations. Multi-response permutation procedures (MRPP)
(Sørrenson) were applied to find separation between groups.
Species frequency was used with rare species down-weighted.
Cluster analyses and statistical tests were performed with PC-
ORD version 5.0 (McCune & Mefford 1999; McCune & Grace
2002).
The non-parametric Wilcoxon test for paired samples was
used to test for significant differences in the occurrence of se-
lected fungal taxa in organic and mineral soil layers. Only taxa
identified in five or more of the cores taken in the autumn
sampling were included in this analysis. Mann–Whitney U
tests were performed to test for homogeneity of seasons.
Only taxonomic groups represented by three or more species
were included in this analysis. P< 0.05 was considered as
statistically significant. These statistical tests were performed
using Statistica 6.0 software package (StatSoft Inc, Tulsa,
USA).
Results
Soils
Due to the long history of emissions, very high total heavy
metal concentrations were measured in the mineral soil
(2546–8068 mg kg�1 lead, 1957–3302 mg kg�1 zinc, and 23–
63 mg kg�1 cadmium) as well as in the organic horizon
(20 404–52 220 mg kg�1 lead, 8177–11 239 mg kg�1 zinc and 86–
111 mg kg�1 cadmium) of the three sample plots. Also
NH4NO3-extractable fractions in mineral soils were very high
(152–1335 mg kg�1 lead, 10 686–58 773 mg kg�1 zinc, 369–
2941 mg kg�1 cadmium).
Fungal community on ECM roots
In the summer, 5212 individual mycorrhizal root tips were
randomly selected. The mean number of selected ECM tips
per 100 ml soil amounted to 2529 (S.D.¼ 410; n¼ 3) for mineral
soils, and to 3459 (S.D.¼ 870; n¼ 3) for organic soils (range 542–
1202 root tips/sample; three pooled samples comprising five
cores each, organic and mineral layer separately). In autumn,
a total of 6808 individual mycorrhizal root tips were randomly
selected. The mean number of ECM tips per 100 ml soil
amounted to 1735 (S.D.¼ 318; n¼ 15) for mineral soils, and to
4263 (S.D.¼ 1895; n¼ 15) for organic soils (range 836–1952 root
tips/sample; organic and mineral layer separately).
The examination of Populus tremula root tips showed an
ECM mycorrhization degree of 95 %. Arbuscular mycorrhiza
was not observed. Due to the high abundance of brownish
black root tips a successful and reliable morphotyping was
limited. Sequences were obtained for an average of 53 % of
all processed root tip DNA extracts.
We recovered a total of 54 OTUs of ECM fungi on the roots
of P. tremula (Table 1). Basidiomycetes clearly dominated (43
OTUs) over ascomycetes (11 OTUs). Species richness of five
cores per plot and season (mineral and organic layer sepa-
rately) ranged from 7–15 OTUs in summer (mean 9 OTUs in
the organic horizon and 10 OTUs in the mineral horizon). In
autumn, we found 10–15 OTUs (mean 12 OTUs in the organic
horizon and 12 OTUs in the mineral horizon). The taxonomi-
cal groups with the highest below-ground species richness
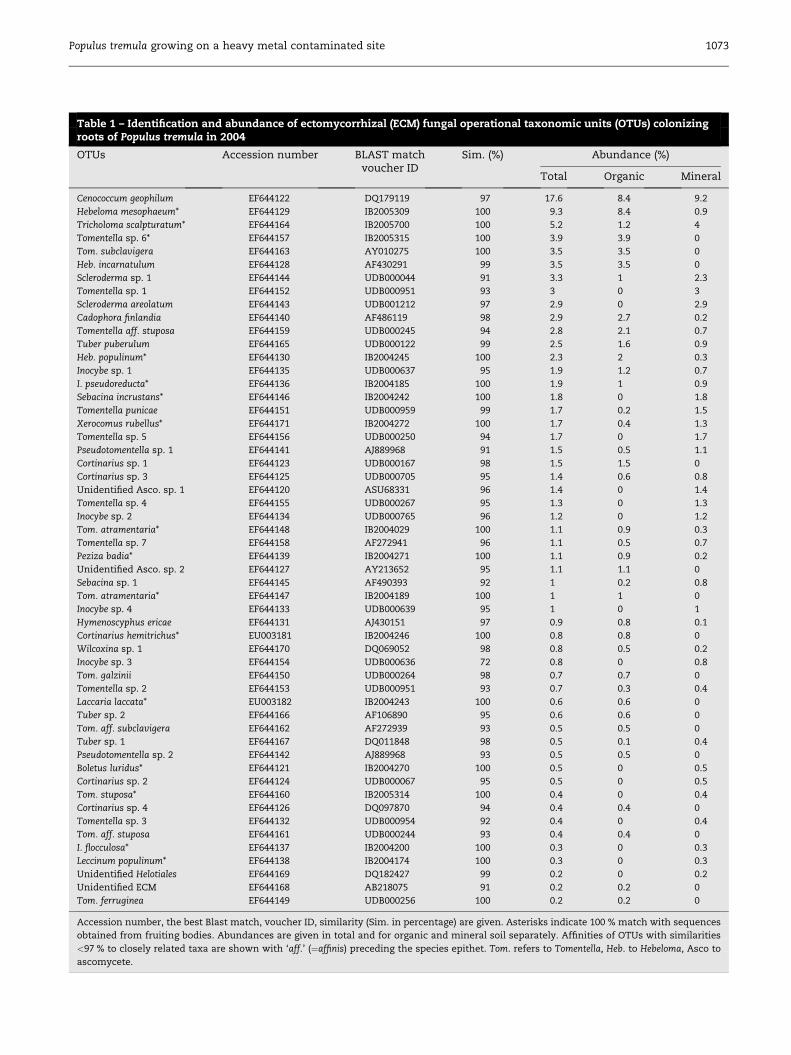
Table 1 – Identification and abundance of ectomycorrhizal (ECM) fungal operational taxonomic units (OTUs) colonizingroots of Populus tremula in 2004
OTUs Accession number BLAST matchvoucher ID
Sim. (%) Abundance (%)
Total Organic Mineral
Cenococcum geophilum EF644122 DQ179119 97 17.6 8.4 9.2
Hebeloma mesophaeum* EF644129 IB2005309 100 9.3 8.4 0.9
Tricholoma scalpturatum* EF644164 IB2005700 100 5.2 1.2 4
Tomentella sp. 6* EF644157 IB2005315 100 3.9 3.9 0
Tom. subclavigera EF644163 AY010275 100 3.5 3.5 0
Heb. incarnatulum EF644128 AF430291 99 3.5 3.5 0
Scleroderma sp. 1 EF644144 UDB000044 91 3.3 1 2.3
Tomentella sp. 1 EF644152 UDB000951 93 3 0 3
Scleroderma areolatum EF644143 UDB001212 97 2.9 0 2.9
Cadophora finlandia EF644140 AF486119 98 2.9 2.7 0.2
Tomentella aff. stuposa EF644159 UDB000245 94 2.8 2.1 0.7
Tuber puberulum EF644165 UDB000122 99 2.5 1.6 0.9
Heb. populinum* EF644130 IB2004245 100 2.3 2 0.3
Inocybe sp. 1 EF644135 UDB000637 95 1.9 1.2 0.7
I. pseudoreducta* EF644136 IB2004185 100 1.9 1 0.9
Sebacina incrustans* EF644146 IB2004242 100 1.8 0 1.8
Tomentella punicae EF644151 UDB000959 99 1.7 0.2 1.5
Xerocomus rubellus* EF644171 IB2004272 100 1.7 0.4 1.3
Tomentella sp. 5 EF644156 UDB000250 94 1.7 0 1.7
Pseudotomentella sp. 1 EF644141 AJ889968 91 1.5 0.5 1.1
Cortinarius sp. 1 EF644123 UDB000167 98 1.5 1.5 0
Cortinarius sp. 3 EF644125 UDB000705 95 1.4 0.6 0.8
Unidentified Asco. sp. 1 EF644120 ASU68331 96 1.4 0 1.4
Tomentella sp. 4 EF644155 UDB000267 95 1.3 0 1.3
Inocybe sp. 2 EF644134 UDB000765 96 1.2 0 1.2
Tom. atramentaria* EF644148 IB2004029 100 1.1 0.9 0.3
Tomentella sp. 7 EF644158 AF272941 96 1.1 0.5 0.7
Peziza badia* EF644139 IB2004271 100 1.1 0.9 0.2
Unidentified Asco. sp. 2 EF644127 AY213652 95 1.1 1.1 0
Sebacina sp. 1 EF644145 AF490393 92 1 0.2 0.8
Tom. atramentaria* EF644147 IB2004189 100 1 1 0
Inocybe sp. 4 EF644133 UDB000639 95 1 0 1
Hymenoscyphus ericae EF644131 AJ430151 97 0.9 0.8 0.1
Cortinarius hemitrichus* EU003181 IB2004246 100 0.8 0.8 0
Wilcoxina sp. 1 EF644170 DQ069052 98 0.8 0.5 0.2
Inocybe sp. 3 EF644154 UDB000636 72 0.8 0 0.8
Tom. galzinii EF644150 UDB000264 98 0.7 0.7 0
Tomentella sp. 2 EF644153 UDB000951 93 0.7 0.3 0.4
Laccaria laccata* EU003182 IB2004243 100 0.6 0.6 0
Tuber sp. 2 EF644166 AF106890 95 0.6 0.6 0
Tom. aff. subclavigera EF644162 AF272939 93 0.5 0.5 0
Tuber sp. 1 EF644167 DQ011848 98 0.5 0.1 0.4
Pseudotomentella sp. 2 EF644142 AJ889968 93 0.5 0.5 0
Boletus luridus* EF644121 IB2004270 100 0.5 0 0.5
Cortinarius sp. 2 EF644124 UDB000067 95 0.5 0 0.5
Tom. stuposa* EF644160 IB2005314 100 0.4 0 0.4
Cortinarius sp. 4 EF644126 DQ097870 94 0.4 0.4 0
Tomentella sp. 3 EF644132 UDB000954 92 0.4 0 0.4
Tom. aff. stuposa EF644161 UDB000244 93 0.4 0.4 0
I. flocculosa* EF644137 IB2004200 100 0.3 0 0.3
Leccinum populinum* EF644138 IB2004174 100 0.3 0 0.3
Unidentified Helotiales EF644169 DQ182427 99 0.2 0 0.2
Unidentified ECM EF644168 AB218075 91 0.2 0.2 0
Tom. ferruginea EF644149 UDB000256 100 0.2 0.2 0
Accession number, the best Blast match, voucher ID, similarity (Sim. in percentage) are given. Asterisks indicate 100 % match with sequences
obtained from fruiting bodies. Abundances are given in total and for organic and mineral soil separately. Affinities of OTUs with similarities
<97 % to closely related taxa are shown with ‘aff.’ (¼affinis) preceding the species epithet. Tom. refers to Tomentella, Heb. to Hebeloma, Asco to
ascomycete.
Populus tremula growing on a heavy metal contaminated site 1073
0
0,5
1
1,5
2
2,5
3
3,5
0 10 20 30 40 50 60OTUs
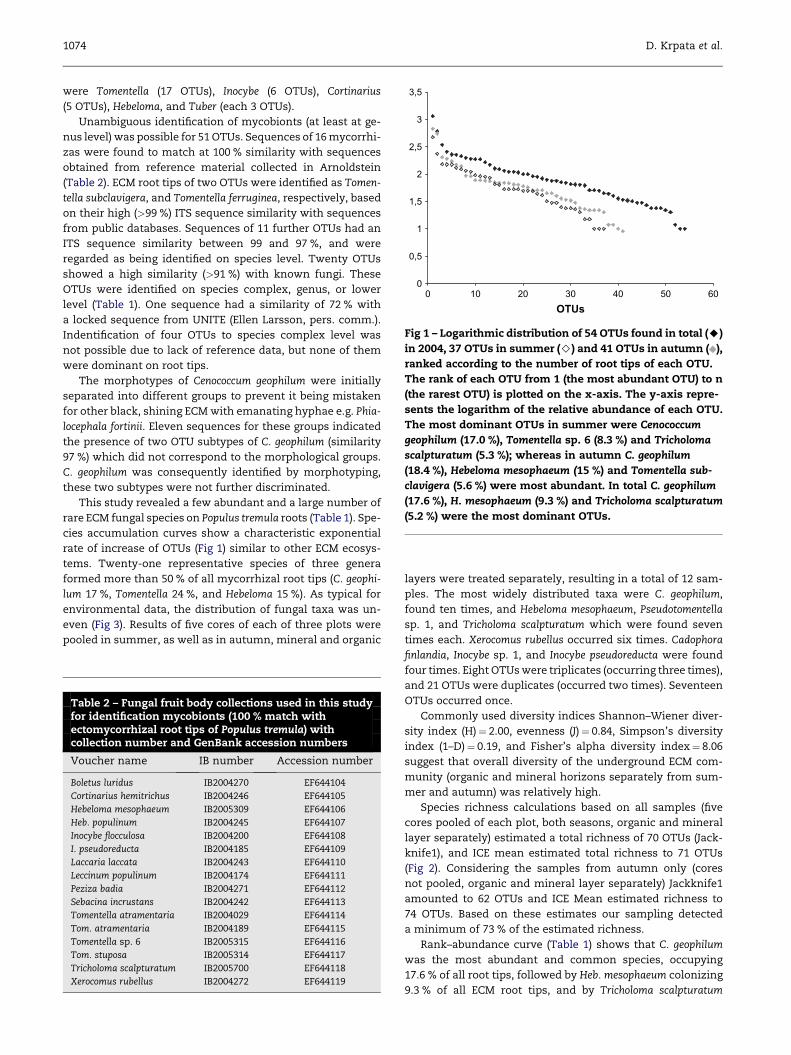
Fig 1 – Logarithmic distribution of 54 OTUs found in total (A)
in 2004, 37 OTUs in summer (>) and 41 OTUs in autumn ( ),
ranked according to the number of root tips of each OTU.
The rank of each OTU from 1 (the most abundant OTU) to n
(the rarest OTU) is plotted on the x-axis. The y-axis repre-
sents the logarithm of the relative abundance of each OTU.
The most dominant OTUs in summer were Cenococcum
geophilum (17.0 %), Tomentella sp. 6 (8.3 %) and Tricholoma
scalpturatum (5.3 %); whereas in autumn C. geophilum
(18.4 %), Hebeloma mesophaeum (15 %) and Tomentella sub-
clavigera (5.6 %) were most abundant. In total C. geophilum
(17.6 %), H. mesophaeum (9.3 %) and Tricholoma scalpturatum
(5.2 %) were the most dominant OTUs.
1074 D. Krpata et al.
were Tomentella (17 OTUs), Inocybe (6 OTUs), Cortinarius
(5 OTUs), Hebeloma, and Tuber (each 3 OTUs).
Unambiguous identification of mycobionts (at least at ge-
nus level) was possible for 51 OTUs. Sequences of 16 mycorrhi-
zas were found to match at 100 % similarity with sequences
obtained from reference material collected in Arnoldstein
(Table 2). ECM root tips of two OTUs were identified as Tomen-
tella subclavigera, and Tomentella ferruginea, respectively, based
on their high (>99 %) ITS sequence similarity with sequences
from public databases. Sequences of 11 further OTUs had an
ITS sequence similarity between 99 and 97 %, and were
regarded as being identified on species level. Twenty OTUs
showed a high similarity (>91 %) with known fungi. These
OTUs were identified on species complex, genus, or lower
level (Table 1). One sequence had a similarity of 72 % with
a locked sequence from UNITE (Ellen Larsson, pers. comm.).
Indentification of four OTUs to species complex level was
not possible due to lack of reference data, but none of them
were dominant on root tips.
The morphotypes of Cenococcum geophilum were initially
separated into different groups to prevent it being mistaken
for other black, shining ECM with emanating hyphae e.g. Phia-
locephala fortinii. Eleven sequences for these groups indicated
the presence of two OTU subtypes of C. geophilum (similarity
97 %) which did not correspond to the morphological groups.
C. geophilum was consequently identified by morphotyping,
these two subtypes were not further discriminated.
This study revealed a few abundant and a large number of
rare ECM fungal species on Populus tremula roots (Table 1). Spe-
cies accumulation curves show a characteristic exponential
rate of increase of OTUs (Fig 1) similar to other ECM ecosys-
tems. Twenty-one representative species of three genera
formed more than 50 % of all mycorrhizal root tips (C. geophi-
lum 17 %, Tomentella 24 %, and Hebeloma 15 %). As typical for
environmental data, the distribution of fungal taxa was un-
even (Fig 3). Results of five cores of each of three plots were
pooled in summer, as well as in autumn, mineral and organic
Table 2 – Fungal fruit body collections used in this studyfor identification mycobionts (100 % match withectomycorrhizal root tips of Populus tremula) withcollection number and GenBank accession numbers
Voucher name IB number Accession number
Boletus luridus IB2004270 EF644104
Cortinarius hemitrichus IB2004246 EF644105
Hebeloma mesophaeum IB2005309 EF644106
Heb. populinum IB2004245 EF644107
Inocybe flocculosa IB2004200 EF644108
I. pseudoreducta IB2004185 EF644109
Laccaria laccata IB2004243 EF644110
Leccinum populinum IB2004174 EF644111
Peziza badia IB2004271 EF644112
Sebacina incrustans IB2004242 EF644113
Tomentella atramentaria IB2004029 EF644114
Tom. atramentaria IB2004189 EF644115
Tomentella sp. 6 IB2005315 EF644116
Tom. stuposa IB2005314 EF644117
Tricholoma scalpturatum IB2005700 EF644118
Xerocomus rubellus IB2004272 EF644119
layers were treated separately, resulting in a total of 12 sam-
ples. The most widely distributed taxa were C. geophilum,
found ten times, and Hebeloma mesophaeum, Pseudotomentella
sp. 1, and Tricholoma scalpturatum which were found seven
times each. Xerocomus rubellus occurred six times. Cadophora
finlandia, Inocybe sp. 1, and Inocybe pseudoreducta were found
four times. Eight OTUs were triplicates (occurring three times),
and 21 OTUs were duplicates (occurred two times). Seventeen
OTUs occurred once.
Commonly used diversity indices Shannon–Wiener diver-
sity index (H)¼ 2.00, evenness (J)¼ 0.84, Simpson’s diversity
index (1–D)¼ 0.19, and Fisher’s alpha diversity index¼ 8.06
suggest that overall diversity of the underground ECM com-
munity (organic and mineral horizons separately from sum-
mer and autumn) was relatively high.
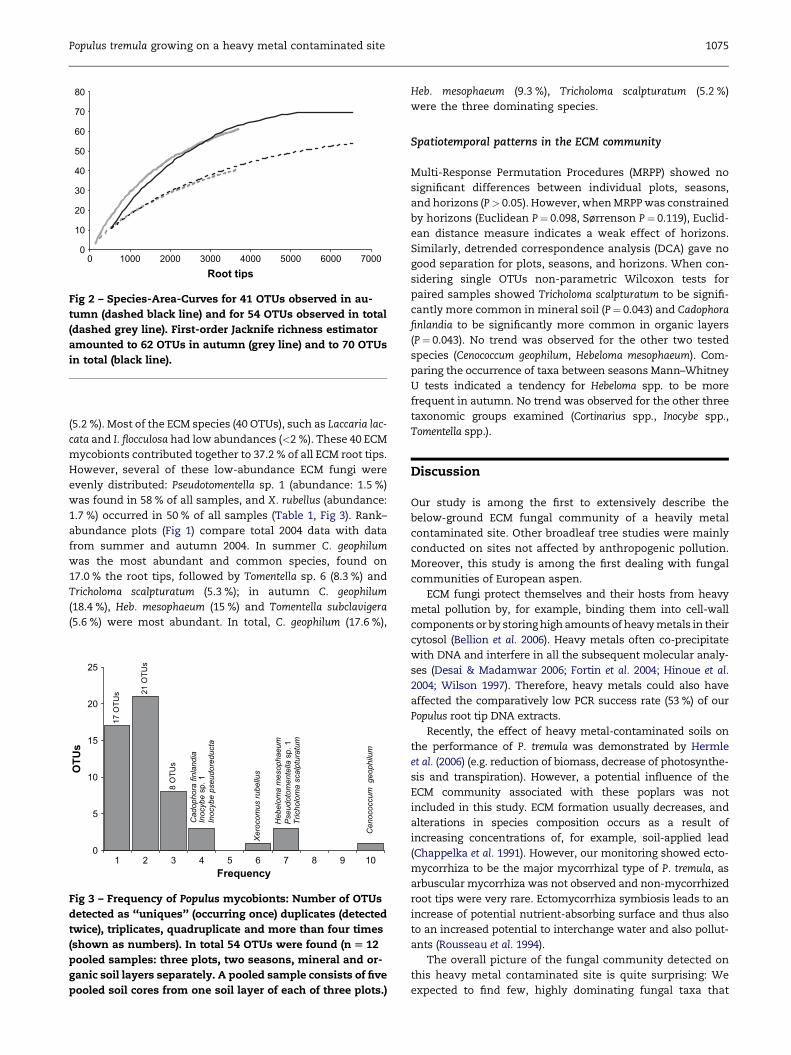
Species richness calculations based on all samples (five
cores pooled of each plot, both seasons, organic and mineral
layer separately) estimated a total richness of 70 OTUs (Jack-
knife1), and ICE mean estimated total richness to 71 OTUs
(Fig 2). Considering the samples from autumn only (cores
not pooled, organic and mineral layer separately) Jackknife1
amounted to 62 OTUs and ICE Mean estimated richness to
74 OTUs. Based on these estimates our sampling detected
a minimum of 73 % of the estimated richness.
Rank–abundance curve (Table 1) shows that C. geophilum
was the most abundant and common species, occupying
17.6 % of all root tips, followed by Heb. mesophaeum colonizing
9.3 % of all ECM root tips, and by Tricholoma scalpturatum
0
10
20
30
40
50
60
70
80
0 1000 2000 3000 4000 5000 6000 7000Root tips
Fig 2 – Species-Area-Curves for 41 OTUs observed in au-
tumn (dashed black line) and for 54 OTUs observed in total
(dashed grey line). First-order Jacknife richness estimator
amounted to 62 OTUs in autumn (grey line) and to 70 OTUs
in total (black line).
Populus tremula growing on a heavy metal contaminated site 1075
(5.2 %). Most of the ECM species (40 OTUs), such as Laccaria lac-
cata and I. flocculosa had low abundances (<2 %). These 40 ECM
mycobionts contributed together to 37.2 % of all ECM root tips.
However, several of these low-abundance ECM fungi were
evenly distributed: Pseudotomentella sp. 1 (abundance: 1.5 %)
was found in 58 % of all samples, and X. rubellus (abundance:
1.7 %) occurred in 50 % of all samples (Table 1, Fig 3). Rank–
abundance plots (Fig 1) compare total 2004 data with data
from summer and autumn 2004. In summer C. geophilum
was the most abundant and common species, found on
17.0 % the root tips, followed by Tomentella sp. 6 (8.3 %) and
Tricholoma scalpturatum (5.3 %); in autumn C. geophilum
(18.4 %), Heb. mesophaeum (15 %) and Tomentella subclavigera
(5.6 %) were most abundant. In total, C. geophilum (17.6 %),
Cenococcum
geophilum
Hebelo
ma m
esophaeum
Pseudotom
entella sp
. 1
Tric
holo
ma scalp
turatum
Xerocom
us rubellus
Cadophora fin
landia
Inocybe sp
. 1Inocybe pseudoreducta
8 O
TUs
21 O
TUs
17 O
TUs
0
5
10
15
20
25
1 2 3 4 5 6 7 8 9 10Frequency
OT
Us
Fig 3 – Frequency of Populus mycobionts: Number of OTUs
detected as ‘‘uniques’’ (occurring once) duplicates (detected
twice), triplicates, quadruplicate and more than four times
(shown as numbers). In total 54 OTUs were found (n [ 12
pooled samples: three plots, two seasons, mineral and or-
ganic soil layers separately. A pooled sample consists of five
pooled soil cores from one soil layer of each of three plots.)
Heb. mesophaeum (9.3 %), Tricholoma scalpturatum (5.2 %)
were the three dominating species.
Spatiotemporal patterns in the ECM community
Multi-Response Permutation Procedures (MRPP) showed no
significant differences between individual plots, seasons,
and horizons (P> 0.05). However, when MRPP was constrained
by horizons (Euclidean P¼ 0.098, Sørrenson P¼ 0.119), Euclid-
ean distance measure indicates a weak effect of horizons.
Similarly, detrended correspondence analysis (DCA) gave no
good separation for plots, seasons, and horizons. When con-
sidering single OTUs non-parametric Wilcoxon tests for
paired samples showed Tricholoma scalpturatum to be signifi-
cantly more common in mineral soil (P¼ 0.043) and Cadophora
finlandia to be significantly more common in organic layers
(P¼ 0.043). No trend was observed for the other two tested
species (Cenococcum geophilum, Hebeloma mesophaeum). Com-
paring the occurrence of taxa between seasons Mann–Whitney
U tests indicated a tendency for Hebeloma spp. to be more
frequent in autumn. No trend was observed for the other three
taxonomic groups examined (Cortinarius spp., Inocybe spp.,
Tomentella spp.).
Discussion
Our study is among the first to extensively describe the
below-ground ECM fungal community of a heavily metal
contaminated site. Other broadleaf tree studies were mainly
conducted on sites not affected by anthropogenic pollution.
Moreover, this study is among the first dealing with fungal
communities of European aspen.
ECM fungi protect themselves and their hosts from heavy
metal pollution by, for example, binding them into cell-wall
components or by storing high amounts of heavy metals in their
cytosol (Bellion et al. 2006). Heavy metals often co-precipitate
with DNA and interfere in all the subsequent molecular analy-
ses (Desai & Madamwar 2006; Fortin et al. 2004; Hinoue et al.
2004; Wilson 1997). Therefore, heavy metals could also have
affected the comparatively low PCR success rate (53 %) of our
Populus root tip DNA extracts.
Recently, the effect of heavy metal-contaminated soils on
the performance of P. tremula was demonstrated by Hermle
et al. (2006) (e.g. reduction of biomass, decrease of photosynthe-
sis and transpiration). However, a potential influence of the
ECM community associated with these poplars was not
included in this study. ECM formation usually decreases, and
alterations in species composition occurs as a result of
increasing concentrations of, for example, soil-applied lead
(Chappelka et al. 1991). However, our monitoring showed ecto-
mycorrhiza to be the major mycorrhizal type of P. tremula, as
arbuscular mycorrhiza was not observed and non-mycorrhized
root tips were very rare. Ectomycorrhiza symbiosis leads to an
increase of potential nutrient-absorbing surface and thus also
to an increased potential to interchange water and also pollut-
ants (Rousseau et al. 1994).
The overall picture of the fungal community detected on
this heavy metal contaminated site is quite surprising: We
expected to find few, highly dominating fungal taxa that

1076 D. Krpata et al.
were well adapted to this stressful environment. This initial
hypothesis was also supported by other studies of ECM com-
munities of zinc and uranium polluted soils: one morphotype
(an unidentified Pinirhiza arenosa) formed 70 % of all analysed
mycorrhizas followed by less abundant Tricholoma scalpturatum,
Hebeloma mesophaeum, and members of Thelephoraceae on roots
of Pinus sylvestris and Betula pendula growing in a zinc waste
(Mleczko 2004); three mycobiont species (Phialophora sp., Lactar-
ius decipiens, and Amanita muscaria) dominated the ECM commu-
nity of B. pendula growing at a uranium mining heap
(Staudenrausch et al. 2005). The high diversity and evenness of
the detected fungal communities in our study is contrary to
these findings. Our initial hypothesis of a low ECM diversity
could not be corroborated as 54 fungal taxa were detected as
mycobionts in this heavily contaminated site. A comparison
with other data indicates that diversity values of our contami-
nated Populus sites (Shannon-Wiener index H¼ 2; Simpson’s
diversity index 1-D¼ 0.189) lay within the range observed in
other studies on broadleaved trees, although data of similar
studies are not directly comparable due to different definitions
of OTUs: Smith et al. (2007) reported H¼ 1.077 on oak, DeBellis
et al. (2006) reported H¼ 3 on aspen, and H¼ 4.2 on birch.
Richard et al. (2005) found 1-D¼ 0.244 on saplings of Quercus,
while Smith et al. (2007) reported 0.525 on mature oaks. Also
studies of conifer-dominated forests reported similar values
[Ishida et al. (2007) reported H¼ 2.7–3.5 in conifer–broadleaf
mixed forests; Fransson et al. (2000) reported H¼ 1.96–2.44 in
a Norway spruce forest; Cline et al. (2005) reported 1.19 on
mature Douglas fir].
Basidiomycetes (43 OTUs forming 71 % of ECM root tips)
were the most numerous fungi associated with roots of
P. tremula. The species composition of our P. tremula stand
was comparable with those of mature forests (e.g. Horton
et al. 1999; Stendell et al. 1999; Jakucs 2002). However, many
of the taxa detected in Arnoldstein were typically observed
in early successional or disturbed habitats (Cenococcum, Hebe-
loma, Inocybe, Scleroderma, Tomentella, Tuber, Wilcoxina) (Nara
et al. 2003; Rudawska et al. 2006; Muhlmann et al. 2008) reflect-
ing the short and disturbed history of our Populus sites.
We detected a few species characteristic for aspen stands
(e.g. Hebeloma populinum, Leccinum populinum). As is typical
for communities of any taxonomic group (Putman 1994;
Horton & Bruns 2001), a few abundant and a large number of
rare ECM fungal species were found on P. tremula roots.
Dahlberg et al. (2001) compiled a review of 49 ECM fungal com-
munity studies of coniferous and deciduous forests. They
found Cenococcum geophilum, corticoid basidiomycetes and
Thelephoraceae to be the three most abundant or frequent
ECM fungal taxa, as recorded from mycorrhizas. It is now
widely known that species belonging to the Thelephoraceae
are among the most frequent and abundant ECM species in
Europe and Northern America (Horton & Bruns, 2001; Koljalg
et al. 2000), they are generalists and important mycorrhizal
partners of many deciduous trees and conifers. This is in
accordance with our study and a study on ECM of transgenic
aspen (Kaldorf et al. 2004). Conversely, the results obtained
in the latter study differ considerably regarding species com-
position and diversity. Different environmental conditions,
as well as different plant ages, may cause these discrepancies;
the estimated age of the naturally grown aspen trees in
Arnoldstein was about 20–25 y. In contrast, the aspen clones
investigated by Kaldorf et al. (2004) were planted in an exper-
imental field and did not exceed the age of six years. There-
fore, typical early-stage ECM fungi, such as Laccaria laccata,
were among the dominating mycobionts of young transgenic
plants, whereas this fungus was found with low abundance
(0.6 %) in our study.
Thirty-three fungal ECM taxa, which do not form conspic-
uous fruiting bodies (mainly Tomentella spp.), were detected on
the roots of P. tremula. These species colonized 59 % of all root
tips. Tomentella spp. form thin, resupinate fruiting bodies on
bare ground or on wooden substrates and are easily over-
looked. Thus, only sequences from fruiting bodies of four
Tomentella species (out of 17 on the root tips) were found to
match at 100 % similarity with sequences obtained from root
tips.
Cenococcum geophilum does not form fruit bodies but sclero-
tia of C. geophilum were found in the soil. However, it was not
possible to obtain cultures from C. geophilum. This anamorphic
ascomycete with worldwide distribution appeared to be the
best adapted mycobiont to the conditions in our plots, forming
17.6 % of all P. tremula mycorrhizal root tips. This agrees with
data from experiments with soil-applied lead, which was
found to lead to an increase in occurrence of C. geophilum
ECM on Pinus taeda up to 300 % compared with uncontami-
nated control soil (Chappelka et al. 1991). Similarly, Cripps
(2003) reported a prolific occurrence of C. geophilum on aspen
in areas of high pollution.
Obviously, all fungal lineages of this specific site are toler-
ant to the high heavy metal concentrations of the environ-
ment. The high degree of mycorrhization suggests that ECM
fungi play an important role in heavy metal uptake or avoid-
ance of the plants. Fungal immobilisation of heavy metals
could be another potential mechanism by which mycorrhizal
fungi alleviate metal toxicity to their hosts (Joner & Leyval
1997). However, interactions are very complex as they vary
in a fungal species- and metal-specific manner: e.g. cadmium
was almost exclusively localized in fungal cell walls of the
Hartig net, whereas zinc was accumulated in both cell walls
and the fungal cytoplasm (Frey et al. 2000); suilloid mycorrhi-
zas significantly enriched lead and zinc in their mantle,
although no such biofiltering effect was observed for
mycorrhizas formed by other taxa (Turnau et al. 2002). During
this study, cultures of several fungal species found in
Arnoldstein were obtained from fruit bodies and from
mycorrhizal root tips. Further studies on the heavy metal
concentrations of abundant mycobionts and of synthetic my-
corrhizae are planned to study a potential selective enrich-
ment of heavy metals at high concentrations within
mycorrhizal fungal structures (ECM mantle and Hartig net
hyphae).
Sequences of 16 fungal species detected as mycorrhizal
partners were found to match perfectly (>99 %) with se-
quences obtained from fruit bodies collected at the site. Thirty
percent of the ECM species found below-ground were also
found as fruiting bodies. This is higher than data reported in
other studies (e.g. Jonsson et al. 1999: 17 % in Scotch pine
stands; Peter et al. 2001: 22 % in a Norway spruce forest; Ishida
et al. 2007: 13 % in mixed conifer–broadleaf forests; Smith et al.
2007: 20 % when considering only epigeous taxa), but lower

Populus tremula growing on a heavy metal contaminated site 1077
than the 45 % detected in an extensively (8 years) sampled
xeric Quercus forest, where sampling included also hypogeous
taxa (Smith et al. 2007). Pritsch et al. (1997) observed fruiting
bodies from eight species out of a total of 16 ECM species asso-
ciated with black alder (Alnus glutinosa). In their study epige-
ous fruiting bodies were collected monthly over one growing
season. Similarly, Cripps (2004) found half of the mycorrhizal
fungi associated with P. tremuloides as fruiting bodies above
ground. The varying correspondence between detected fungal
fruiting bodies and ECM taxa often depends on the sampling
effort and the available taxonomical expertise.
The number of mycorrhizal Populus root tips per 100 ml soil
(1735 to 4263) was similar to or slightly higher than the num-
ber of Picea abies root tips per 100 ml soil (1117 to 2370)
reported from an Austrian site (Marth 2007).
Our results revealed trends of some species of the ECM
community to preferentially occur in different soil layers
(Genney et al. 2006), e.g. Tricholoma scalpturatum in the mineral,
and Cadophora finlandia in the organic horizon. This is in agree-
ment with Tedersoo et al. (2003), who studied a mixed, mostly
deciduous forest and found Cadophora finlandia mostly in the
A-horizon. In contrast, studies of boreal forests (Rosling et al.
2003) showed C. finlandia to preferentially colonise roots in
the mineral soil. Interestingly, in our study no preferences
for a certain soil layer could be observed of C. geophilum.
This contrasts with the studies of Goodman & Trofymow
(1998) and Fransson et al. (2000) who found that C. geophilum
was more common in the organic layer than in the mineral
soil. We speculate that the even distribution of C. geophilum
could be related to the high heavy metal concentrations which
generally lead to a considerable increase of C. geophilum abun-
dances (Chappelka et al. 1991).
Acknowledgements
We sincerely thank Friederike Gobl and Regina Kuhnert-
Finkernagel for assistance and helpful comments. This study
is part of the FWF Project P170120-B08 ‘Mycorrhizal associa-
tions of zinc/cadmium-accumulating poplars: perspectives
for phytoremediation?’. And last but not least the authors
wish to thank the two anonymous reviewers for their useful
comments.
r e f e r e n c e s
Adriaensen K, Vangronsveld J, Colpaert JV, 2006. Zinc-tolerantSuillus bovinus improves growth of Zn-exposed Pinus sylvestrisseedlings. Mycorrhiza 16: 553–558.
Agerer R, 1987–2002. Colour Atlas of Ectomycorrhizae. Einhorn-Verlag,Schwabisch Gmund.
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ, 1997. Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. Nucleic AcidsResearch 25: 3389–3402.
Baier R, Ingenhaag J, Blaschke H, Gottlein A, Agerer R, 2006.Vertical distribution of an ectomycorrhizal community inupper soil horizons of a young Norway spruce (Picea abies [L.]
Karst.) stand of the Bavarian Limestone Alps. Mycorrhiza 16:197–206.
Bellion M, Courbot M, Jacob C, Blaudez D, Chalot M, 2006. Extra-cellular and cellular mechanisms sustaining metal tolerancein ectomycorrhizal fungi. FEMS Microbiological Letters 254:173–181.
Bittsanszky A, Komives T, Gullner G, Gyulai G, Kiss J, Heszky L,Radimszkyc L, Rennenberg H, 2005. Ability of transgenic pop-lars with elevated glutathione content to tolerate zinc(2þ)stress. Environment International 31: 251–254.
Blum W, Spiegel H, Wenzel W, 1996. Bodenzustandsinventur, Kon-zeption und Durchfuhrung. Bundesminsterium fur Land undForstwirtschaft and Bundesministerium fur Wissenschaft,Verkehr und Kunst, Vienna.
Burnham KP, Overton WS, 1979. Robust estimation of populationsize when capture probabilities vary among animals. Ecology60: 927–936.
Chao A, 1987. Estimating the population size for capture–recap-ture data with unequal catchability. Biometrics 43: 783–791.
Chappelka AH, Kush JS, Runion GB, Meier S, Kelly WD, 1991. Effectof soil applied lead on seedling growth and ectomycorrhizalcolonisation of loblolly pine. Environmental Pollution 72:307–316.
Cline ET, Ammirati JF, Edmonds RL, 2005. Does proximity to ma-ture trees influence ectomycorrhizal fungus communities ofDouglas-fir seedlings? New Phytologist 166: 993–1009.
Cobbett C, 2003. Heavy metals and plants – model systems andhyperaccumulators. New Phytologist 159: 289–293.
Colpaert JV, Muller LAH, Lambaerts M, Adriaensen K,Vangronsveld J, 2004. Evolutionary adaptation to Zn toxicity inpopulations of Suilloid fungi. New Phytologist 162: 549–559.
Colpaert JV, Adriaensen K, Muller LAH, Lambaerts M, Faes C,Carleer R, Vangronsveld J, 2005. Element profiles and growthin Zn-sensitive and Zn-resistant suilloid fungi. Mycorrhiza 15:628–634.
Colwell RK, 2006. EstimateS. Version 8.0. Statistical Estimation ofSpecies Richness and Shared Species from Samples (Software and User’sGuide). Published at: http://viceroy.eeb.uconn.edu/estimates.
Cripps CL, Miller jr OK, 1993. Ectomycorrhizal fungi associatedwith aspen on three sites in the north-central Rocky Moun-tains. Canadian Journal of Botany 71: 1414–1420.
Cripps CL, 2003. Native mycorrhizal fungi with aspen on smelter-impacted sites in the Northern Rocky Mountains: occurrenceand potential use in reclamation. In: Conference Proceedings,National Meeting of the American Society of Mining and Reclama-tion, 3–6 June 2003, Lexington, KY.
Cripps CL, 2004. Ectomycorrhizal fungi above and below groundin a small, isolated aspen stand: a simple system revealsfungal fruiting strategies and an edge effect. In: Cripps CL (ed),Fungi in Forest Ecosystems: systematics, diversity and ecology. NewYork Botanical Garden Press, New York, pp. 249–265.
Dahlberg A, Schimmel J, Taylor AFS, Johannesson H, 2001. Post-fire legacy of ectomycorrhizal fungal communities in theSwedish boreal forest in relation to fire severity and loggingintensity. Biological Conservation 100: 151–161.
Dai J, Becquer T, Rouiller J, Reversat G, Bernhard-Reversat F,Lavelle P, 2004. Influence of heavy metals on C and N miner-alisation and microbial biomass in Zn-, Pb-, Cu-, and Cd-con-taminated soils. Applied Soil Ecology 25: 99–109.
DeBellis T, Kernaghan G, Bradley R, Widden P, 2006. Relationshipsbetween stand composition and ectomycorrhizal communitystructure in boreal mixed-wood forests. Microbial Ecology 52:114–126.
Desai C, Madamwar D, 2006. Extraction of inhibitor-freemetagenomic DNA from polluted sediments, compatiblewith molecular diversity analysis using adsorption andion-exchange treatments. Bioresource Technology 98:761–768.

1078 D. Krpata et al.
Dickmann DI, 2006. Silviculture and biology of short-rotationwoody crops in temperate regions: then and now. Biomass andBioenergy 30: 696–705.
DIN 19730, 1995. Bodenbeschaffenheit – Extraktion von Spurenele-menten mit Ammoniumnitratlosung. Beuth Verlag, Berlin,Germany.
Djingova R, Wagner G, Kuleff I, 1999. Screening of heavy metalpollution in Bulgaria using Populus nigra ‘Italica’. The Science ofthe Total Environment 234: 175–184.
Dunabeitia M, Hormilla S, Garcia-Plazaola J, Txarterina K,Arteche U, Becerril J, 2004. Differential responses of threefungal species to environmental factors and their role in themycorrhization of Pinus radiata D. Don. Mycorrhiza 14: 11–18.
FAO, 1998. World Reference Base for Soil Resources. FAO, ISRIC, ISSS,Rome.
Fortin N, Beaumier D, Lee K, Greer CW, 2004. Soil washing im-proves the recovery of total community DNA from pollutedand high organic content sediments. Journal of MicrobiologicalMethods 56: 181–191.
Fransson PMA, Taylor AFS, Finlay RD, 2000. Effects of continuousoptimal fertilization on belowground ectomycorrhizal com-munity structure in a Norway spruce forest. Tree Physiology 20:599–606.
French CJ, Dickinson NM, Putwain PD, 2006. Woody biomassphytoremediation of contaminated brownfield land. Environ-mental Pollution 141: 387–395.
Frey B, Zierold K, Brunner I, 2000. Extracellular complexation ofCd in the Hartig net and cytosolic Zn sequestration in thefungal mantle of Picea abies–Hebeloma crustuliniforme ectomy-corrhizas. Plant, Cell & Environment 23: 1257–1266.
Gardes M, White TJ, Fortin JA, Bruns TD, Taylor JW, 1991. Identi-fication of indigenous and introduced symbiotic fungi in ec-tomycorrhizae by amplification of nuclear and mitochondrialribosomal DNA. Canadian Journal of Botany 69: 180–190.
Gardes M, Bruns TD, 1993. ITS primers with enhanced specificityfor basidiomycetes – application to the identification of my-corrhizae and rusts. Molecular Ecology 2: 113–118.
Gardes M, Bruns TD, 1996. Community structure of ectomycor-rhizal fungi in a Pinus muricata forest: above- and below-ground views. Canadian Journal of Botany 74: 1572–1583.
Gazey C, Abbott L, Robson A, 1992. The rate of development ofmycorrhizas affects the onset of sporulation and productionof external hyphae by two species of Acaulospora. MycologicalResearch 96: 643–650.
Genney DR, Anderson IC, Alexander IJ, 2006. Fine-scale distribu-tion of pine ectomycorrhizas and their extramatrical myce-lium. New Phytologist 170: 381–390.
Giller KE, Witter E, McGrath SP, 1998. Toxicity of heavy metals tomicroorganisms and microbial processes in agricultural soils.Soil Biology and Biochemistry 30: 1389–1414.
Goodman DM, Trofymow JA, 1998. Comparison of communities ofectomycorrhizal fungi in old-growth and mature stands ofDouglas-fir at two sites on southern Vancouver Island. Cana-dian Journal of Forest Research 28: 574–581.
Hartley J, Cairney JWG, Meharg AA, 1997. Do ectomycorrhizalfungi exhibit adaptive tolerance to potentially toxic metals inthe environment? Plant and Soil 189: 303–319.
Hartley-Whitaker J, Cairney JWG, Meharg AA, 2000. Sensitivity toCd or Zn of host and symbiont of ectomycorrhizal Pinus syl-vestris L. (Scots pine) seedlings. Plant and Soil 218: 31–42.
Hermle S, Gunthardt-Goerg MS, Schulin R, 2006. Effects of metal-contaminated soil on the performance of young trees growingin model ecosystems under field conditions. EnvironmentalPollution 144: 703–714.
Hinoue M, Fukuda K, Wan Y, Yamauchi K, Ogawa H, Taniguchi H,2004. An effective method for extracting DNA from contami-nated soil due to industrial waste. Journal of the University ofOccupational and Environmental Health (Japan) 26: 13–21.
Horton TR, Bruns TD, Parker VT, 1999. Ectomycorrhizal fungi as-sociated with Arctostaphylos contribute to Pseudotsuga menziesiiestablishment. Canadian Journal of Botany 77: 93–102.
Horton TR, Bruns TD, 2001. The molecular revolution in ectomy-corrhizal ecology: peeking into the black-box. Molecular Ecology10: 1855–1871.
Ishida TA, Nara K, Hogetsu T, 2007. Host effects on ectomy-corrhizal fungal communities: insight from eight host spe-cies in mixed conifer-broadleaf forests. New Phytologist 174:430–440.
Jakucs E, 2002. Ectomycorrhizae of Populus alba L. in south Hun-gary. Phyton-Annales Rei Botanicae 42: 199–210.
Jentschke G, Godbold DL, 2000. Metal toxicity and ectomycorrhi-zas. Physiologia Plantarum 109: 107–116.
Joner EJ, Leyval C, 1997. Uptake of 109 Cd by roots and hyphae ofa Glomus mosseae/Trifolium subterraneum mycorrhiza from soilamended with high and low concentrations of cadmium.New Phytologist 135: 353–360.
Jonsson L, Dahlberg A, Nilsson MC, Zackrisson O, Karen O, 1999.Ectomycorrhizal fungal communities in late-successionalSwedish boreal forests, and their composition followingwildfire. Molecular Ecology 8: 205–215.
Kaldorf M, Renker C, Fladung M, Buscot F, 2004. Characterizationand spatial distribution of ectomycorrhizas colonizing aspenclones released in an experimental field. Mycorrhiza 14:295–306.
Kasperowski E, 1993. Schwermetalle in Boden im Raum Arnoldstein.Umweltbundesamt, Bundesministerium fur Umwelt, Jugendund Familie, Vienna.
Kernaghan G, Harper KA, 2001. Community structure of ectomy-corrhizal fungi across an alpine/subalpine ecotone. Ecography24: 181–188.
Kernaghan G, Widden P, Bergeron Y, Legare S, Pare D, 2003. Bioticand abiotic factors affecting ectomycorrhizal diversity in bo-real mixed-woods. Oikos 102: 497–504.
Khan AG, Kuek C, Chaudhry TM, Khoo CS, Hayes WJ, 2000. Role ofplants, mycorrhizae and phytochelators in heavy metal con-taminated land remediation. Chemosphere 41: 197–207.
Koljalg U, Dahlberg A, Taylor AFS, Larsson E, Hallenberg N,Stenlid J, Larsson KH, Fransson PM, Karen O, Jonsson L, 2000.Diversity and abundance of resupinate thelephoroid fungi asectomycorrhizal symbionts in Swedish boreal forests. Molec-ular Ecology 9: 1985–1996.
Koljalg U, Larsson K-H, Abarenkov K, Nilsson RH, Alexander IJ,Eberhardt U, Erland S, Høiland K, Kjøller R, Larsson E,Pennanen T, Sen R, Taylor AFS, Tedersoo L, Vralstad T,Ursing BM, 2005. UNITE: a database providing web-basedmethods for the molecular identification of ectomycorrhizalfungi. New Phytologist 166: 1063–1068.
Krainer K, 1998. Die Bergsturze des Dobratsch. Verlag des Natur-wissenschaftlichen Vereins fur Karnten, Klagenfurt.
Laureysens I, Bogaert J, Blust R, Ceulemans R, 2004. Biomassproduction of 17 poplar clones in a short-rotation coppiceculture on a waste disposal site and its relation to soil char-acteristics. Forest Ecology and Management 187: 295–309.
Madejon P, Maranon T, Murillo JM, Robinson B, 2004. White pop-lar (Populus alba) as a biomonitor of trace elements in con-taminated riparian forests. Environmental Pollution 132:145–155.
Marth A, 2007. Quantitative Analysen zur Architektur von Ekto-mykorrhizen von Picea abies. MSc thesis, University Inns-bruck, Innsbruck.
Martin F, Tuskan GA, DiFazio SP, Lammers P, Newcombe G,Podila GK, 2004. Symbiotic sequencing for the Populus meso-cosm. New Phytologist 161: 330–335.
McCune B, Mefford MJ, 1999. PC-ORD. Multivariate Analysis ofEcological Data. Version 5.0. MjM Software Design, GlendenBeach, OR.

Populus tremula growing on a heavy metal contaminated site 1079
McCune B, Grace JB, 2002. Analysis of Ecological Communities. MjMSoftware design, Glenden Beach, OR.
Meharg AA, 2003. The mechanistic basis of interactions betweenmycorrhizal associations and toxic metal cations. MycologicalResearch 107: 1253–1265.
Mleczko P, 2004. Mycorrhizal and saprobic macrofungi of two zincwastes in southern Poland. Acta Biologica Cracoviensia SeriesBotanica 46: 25–38.
Moffett BF, Nicholson FA, Uwakwe NC, Chambers BJ, Harris JA,Hill TCJ, 2003. Zinc contamination decreases the bacterial di-versity of agricultural soil. Fems Microbiology Ecology 43: 13–19.
Muhlmann O, Bacher M, Peintner U, 2008. Polygonum viviparummycobionts on an alpine primary successional glacier fore-front. Mycorrhiza 18: 87–95.
Nara K, Nakaya H, Wu BY, Zhou ZH, Hogetsu T, 2003. Under-ground primary succession of ectomycorrhizal fungi in a vol-canic desert on Mount Fuji. New Phytologist 159: 743–756.
Perotto S, Martino E, 2001. Molecular and cellular mechanisms ofheavy metal tolerance in mycorrhizal fungi: what perspectivesfor bioremediation? Minerva Biotechnologica 13: 55–63.
Peter M, Ayer F, Egli S, Honegger R, 2001. Above- and below-ground community structure of ectomycorrhizal fungi inthree Norway spruce (Picea abies) stands in Switzerland.Canadian Journal of Botany 79: 1134–1151.
Pritsch K, Boyle H, Munch JC, Buscot F, 1997. Characterization andidentification of black alder ectomycorrhizas by PCR/RFLPanalyses of the rDNA internal transcribed spacer (ITS). NewPhytologist 137: 357–369.
Putman RJ, 1994. Community Ecology. Chapman Hall, London.Quercia F, Gentile A, Falconi M, Vecchio A, Tarvainen T,
Schamann M, Fons Esteve J, 2006. Soil inventory and assessmentproject: the EEA experience towards an Europe-wide assessment ofareas under risk for soil contamination. European EnvironmentAgency, Athens.
Rambaut A, 1996. Se-Al: Sequence Alignment, version 1.0 alpha 1.University of Oxford, Oxford.
Richard F, Millot S, Gardes M, Selosse MA, 2005. Diversity andspecificity of ectomycorrhizal fungi retrieved from an old-growth Mediterranean forest dominated by Quercus ilex. NewPhytologist 166: 1011–1023.
Rosling A, Landeweert R, Lindahl BD, Larsson KH, Kuyper TW,Taylor AFS, Finlay RD, 2003. Vertical distribution of ectomy-corrhizal fungal taxa in a podzol soil profile. New Phytologist159: 775–783.
Rousseau J, Sylvia D, Fox A, 1994. Contribution of ectomycorrhizato the potential nutrient-absorbing surface of pine. NewPhytologist 128: 639–644.
Rudawska M, Leski T, Trocha LK, Gornowicz R, 2006. Ectomy-corrhizal status of Norway spruce seedlings from bare-root
forest nurseries. Forest Ecology and Management 236:375–384.
Ruhling A, Soderstrom B, 1990. Changes in fruit body productionof mycorrhizal and litter decomposing macromycetes inheavy metal polluted coniferous forests in north Sweden.Water Air and Soil Pollution 49: 375–388.
Smit E, Leeflang P, Wernars K, 1997. Detection of shifts in mi-crobial community structure and diversity in soil caused bycopper contamination using amplified ribosomal DNArestriction analysis. FEMS Microbiology Ecology 23: 249–261.
Smith ME, Douhan GW, Rizzo DM, 2007. Ectomycorrhizal com-munity structure in a xeric Quercus woodland based on rDNAsequence analysis of sporocarps and pooled roots. NewPhytologist 174: 847–863.
Staudenrausch S, Kaldorf M, Renker C, Luis P, Buscot F, 2005.Diversity of the ectomycorrhiza community at a uraniummining heap. Biology and Fertility of Soils 41: 439–446.
Stendell ER, Horton TR, Bruns TD, 1999. Early effects of prescribedfire on the structure of the ectomycorrhizal fungus commu-nity in a Sierra Nevada ponderosa pine forest. MycologicalResearch 103: 1353–1359.
Tedersoo L, Koljalg U, Hallenberg N, Larsson K-H, 2003. Fine scaledistribution of ectomycorrhizal fungi and roots acrosssubstrate layers including coarse woody debris in a mixedforest. New Phytologist 159: 153–165.
Turnau K, Mleczko P, Blaudez D, Chalot M, Botton B, 2002. Heavymetal binding properties of Pinus sylvestris mycorrhizas fromindustrial wastes. Acta Societatis Botanicorum Poloniae 71:253–261.
Turpeinen R, Kairesalo T, Haggblom MM, 2004. Microbial commu-nity structure and activity in arsenic-, chromium- and copper-contaminated soils. FEMS Microbiology Ecology 47: 39–50.
Unterbrunner R, Puschenreiter M, Sommer P, Wieshammer G,Tlustos P, Zupan M, Wenzel WW, 2006. Heavy metal accu-mulation in trees growing on contaminated sites in CentralEurope. Environmental Pollution 148: 107–114.
White TJ, Bruns T, Lee S, Taylor J, 1990. Amplification and directsequencing of fungal ribosomal RNA genes for phylogenetics.In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds), PCRProtocols: a guide to methods and applications, Academic Press,New York, pp. 315–322.
Wilson IG, 1997. Inhibition and facilitation of nucleic acid am-plification. Applied and Environmental Microbiology 63:3741–3751.
Worrell R, 1995. European aspen (Populus tremula L.): a review withparticular reference to Scotland II. Values, silviculture andutilization. Forestry 68: 231–244.
Zolan ME, Pukkila PJ, 1986. Inheritance of DNA methylation inCoprinus cinereus. Molecular and Cellular Biology 6: 195–200.