nitrogen addition does not change amf colonization ... - mdpi
TRANSCRIPT

Citation Wu F Zheng X Cao M
Guan X Jiang J Nitrogen Addition
Does Not Change AMF Colonization
but Alters AMF Composition in a
Chinese Fir (Cunninghamia lanceolata)
Plantation Forests 2022 13 979
httpsdoiorg103390f13070979
Academic Editor Choonsig Kim
Received 21 April 2022
Accepted 21 June 2022
Published 22 June 2022
Publisherrsquos Note MDPI stays neutral
with regard to jurisdictional claims in
published maps and institutional affil-
iations
Copyright copy 2022 by the authors
Licensee MDPI Basel Switzerland
This article is an open access article
distributed under the terms and
conditions of the Creative Commons
Attribution (CC BY) license (https
creativecommonsorglicensesby
40)
Article
Nitrogen Addition Does Not Change AMF Colonization butAlters AMF Composition in a Chinese Fir (Cunninghamialanceolata) PlantationFan Wu dagger Xiang Zheng dagger Minmin Cao Xin Guan and Jiang Jiang
Collaborative Innovation Center of Sustainable Forestry in Southern China of Jiangsu Province Key Labortory ofSoil and Water Conservation and Ecological Restoration in Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China fanwu98njfueducn (FW) xiang-zhengoutlookcom (XZ)healer_caooutlookcom (MC) klein_bluefoxmailcom (XG) Correspondence jiangjiangnjfueducn Tel +86-159-5197-2417dagger These authors contributed equally to this work
Abstract Aims Our aim was to investigate how N addition affects arbuscular mycorrhizal fungal(AMF) growth in Chinese fir plantations Methods A Chinese fir plantation was treated withfour different N addition treatments for one and half years starting in April 2019 AMF colonizationhyphal length density community composition and soil properties were under measurement ResultsN addition caused inapparent effects on AMF colonization hyphal length density and functionalguilds (rhizophilic edaphophilic and ancestral) The predominant AMF species in the soil wasSeptoglomus viscosum N addition altered AMF community and some rare species (eg Entrophosporainfrequens) disappeared with N addition Conclusion AMF community structure was more sensitiveto short-time N deposition than the symbiotic relationship between AMF and host plants
Keywords N addition arbuscular mycorrhizal fungi community composition colonizationChinese fir
1 Introduction
Atmospheric N deposition has increased strongly over the last decades due to N usein agriculture fossil fuels and land conservation [12] and ammonium nitrogen (NH4
+-N)nitrate nitrogen (NO3
minus-N) and dissolved organic nitrogen deposition fluxes were up to873 873 and 998 kg N haminus1 yrminus1 respectively [3] Increased N deposition raises carbonsinks increases N availability and NPP in some ecosystems [4] and typically acidifiessoil [5] Meanwhile N deposition due to its increasing rate and duration possibly has anegative impact with on the growth composition and function of soil microbes [6]
Arbuscular mycorrhizal fungi (AMF) are ubiquitous components belonging to thephylum Glomeromycota They can form mycorrhizal symbionts with approximately 80of land plants [7] AMF gain a significant presence in increasing the uptake of N and Pto their host plants [89] and improving disease resistance which determines the stabilityand versatility of ecosystem [10] It should be noted that AMF have intra- and extra-radical structures [11] Based on their patterns at the family level AMF can be classified asedaphophilic rhizophilic or ancestral Rhizophilic AMF help plants reduce root pathogeninfection with fine root and edaphophilic AMF promote plant nutrients with extraradicalmycelium [12]
AMF structure and community composition can be influenced by climate changebut the factors that affact AMF parameters are not well understood Decreasing pH andP availability caused by N addition can make a big difference in changing soil proper-ties [1314] so plants need AMF to overcome resource limitations However the impact ofN application on AMF remains controversial AMF colonization might be decreased [1516]
Forests 2022 13 979 httpsdoiorg103390f13070979 httpswwwmdpicomjournalforests
Forests 2022 13 979 2 of 14
promoted [17] or have no significant effect [18] by N addition N addition can also en-hance [1819] suppress [20] or have no significant effect [21] on AMF spore density andhyphae Some investigations showed that N addition treatments caused significant sup-pressive impacts on AMF abundance richness and diversity [2223] whereas others foundno significant impact [1924] Meanwhile how these environmental factors shape the AMFcommunity under N addition is not fully understood A study observed that the relativeabundance of AMF decreased the reduction of plant demand for N [25] Importantly themorphological modification of AMF caused by soil nutrient changes may be relevant totheir composition owing to the differences of morphological internal responses to nutrientmanipulations [2627]
Chinese fir (Cunninghamia lanceolata) is a quick-growing timber tree species in southChina [28] On account of its high wood quality and termite resistance it is frequentlyplanted [29] However the productivity of the Chinese fir is declining due to soil acidi-fication soil aluminum poisoning and unreasonable planting methods Lu [30] showedthat the number of AMF operational taxonomic units (OTUs) had an inverted V-shapedchange with the age of the Chinese fir Liu [14] found that the Chinese fir under N additionfor 2 years would increase AMF colonization and diversity Cao [31] observed that high Naddition exerted a significant negative effect on AMF community However few studiesshowed the interactions of Chinese fir and AMF in terms of N deposition and nutrientavailability Therefore we purposed to address these questions (1) how does N additionaffect AMF colonization and the diversity and composition of AMF in Chinese fir (2) wouldthe change of soil nutrients availability affect the responses of AMF groups
2 Materials and Methods21 Study Sites and Sampling
The soil experiment was carried out in the Fengyang Mountain Nature Reserve(2853prime56primeprime N 19010prime56primeprime E) Zhejiang Province China The mean annual precipitationand annual temperature are 2400 mm and 123 C Soil properties (0ndash10 cm) are totalnitrogen (TN) 411 g kgminus1 total carbon (TC) 5873 g kgminus1 NH4
+-N 1571 mg gminus1 NO3minus-
N 1466 mg gminus1 total phosphorus (TP) 03 g kgminus1 and bulk density 076 g cmminus3 TheChinese fir was 39 years old with an average diameter at breast height of 2175 cm and132 m height
The local N deposition was estimated to be 34 kg N haminus1 yrminus1 The N addition treat-ments included four different levels (four plots per treatment were considered as four inde-pendent repetitions sixteen 10 m times 10 m plots in total) to simulate future climate changescenarios no fertilization 50 kg N haminus1 yrminus1 100 kg N haminus1 yrminus1 and 200 kg N haminus1 yrminus1which were regarded as control (CK) low-nitrogen (LN) medium-nitrogen (MN) andhigh-nitrogen (HN) respectively Separated adjacent plots were separated by a 10-m-widebuffer strip From April 2019 N was added monthly by spraying urea (CO (NH2)2) solutionon the forest floor as N addition treatments Meanwhile CK plots were sprayed with thesame volume of water
Samples were collected in November 2020 Along the ldquoSrdquo route six standard treeswere selected in each plot Weeds and litter around the sample trees were removed beforecollecting Roots and rhizosphere soil around the average standard tree were collected inthe 0ndash20 cm soil layer from four directions The fine roots and about 1000 g of rhizospheresoil (soils adhering to roots by shaking off root surface) from six standard trees werecollected and mixed as one root and one soil sample in each plot and sixteen root andsoil samples were collected The soil samples were used for soil physicochemical analysisand the measurement of AMF colonization spore isolation studies and measured hyphallength density (HLD) The soil samples were air-dried and the properties were measuredwhile the other soil samples were stored with root samples (washed using tap water) in therefrigerator at 4 C until used
Forests 2022 13 979 3 of 14
22 Soil Properties Analysis
All the soil samples were sieved through a 2 mm sieve Soil pH was measured in asoil-to-water ratio of 125 (vv) Soil NH4
+-N and NO3minus-N concentration were estimated
by colorimetric analysis [32] Soil available nitrogen (AN) is soil NH4+-N plus NO3
minus-N Soil available phosphorus (AP) and total phosphorus (TP) were extracted by sodiumbicarbonate first and their contents were followed by molybdenum blue colorimetry [33]Soil total organic carbon was measured by TOC-L CPHCPN [34] Soil total carbon (C) andtotal nitrogen (N) were measured by an elemental analyzer (Vario MACRO cube ElementarTrading Shanghai Shanghai China)
23 AMF Colonization and Extraradical Hphae Length Density
To highlight AMF structures roots samples were strained following the methodsdescribed by Philips and Hayman [35] and the percentage of mycorrhizal colonization wasestimated in the light of McGonigle et al [36] Extraradical HLD was determined on thebasis of the description by Jakoben et al [37]
24 AMF Spore Separation and Identification
The AMF spores used for identification were obtained from the 20 g air-dried rhizo-sphere soil samples following the wet sieving and decanting techniques as described byBrundrett [38] In order to determine the spore density the isolated intact healthy sporeswere extracted and placed on filter paper We separated them by color spore size orna-mentations hyphal attachments and wall layers and then used a dissecting microscopeat 45timesmagnification to observe them Spores were mounted on slides in polyvinylndashlacticacidndashglycerine (PVLG) and a mixture of PVLG with Melzerrsquos (11 vv) The spores wereidentified according to the International Culture Collection of Vesicular Arbuscular Mycor-rhizal fungi (INVAM) (httpinvamcafwvuedu last accessed in 21 May 2022) and recentadvances in Glomeromycota taxonomy [3940]
25 AMF Community Parameters and Statistical Analysis
The spore density (SD) frequency of occurrence (FO) relative abundance (RA) impor-tance value (IA) ShannonndashWiener index (H) Simpson index (D) species richness (SR) andpercentage were used to estimate the AMF community structure [41ndash43]
Correlation analysis was performed using SPSS 260 to test for relationship betweenthe AMF community composition parameters and soil properties Venn was pictured forVenny 20 Graphical presentation was carried out using Origin 2022 Redundancy analysis(RDA) was conducted with Canoco 50 to reveal the influence of soil properties on theAMF communities
3 Results31 Effects of N Addition on Soil AMF Colonization Spore Density and External HyphalLength Density
AMF colonization (p = 0824) spore density (p = 0229) and external hyphal lengthdensity (p = 0435) were not significantly influenced by the nitrogen treatment (Table 1) TheAMF colonization ranged from 74 to 88 Spore density was highest in MN treatmentwhich averaged up to 365 spores per 1g air-dried soil and spore density was lowest value inthe control treatment External HLD was highest in the control treatment which averagedup to 510 m gminus1 and was the lowest in the LN treatment
Forests 2022 13 979 4 of 14
Table 1 Responses of arbuscular mycorrhizal fungi under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
Colonization () 8175 plusmn 225 8475 plusmn 160 8300 plusmn 310 8200 plusmn 270 030 082SD (spore gminus1 air-dried soil) 25569 plusmn 3984 29769 plusmn 3233 36463 plusmn 3785 31894 plusmn 4176 165 023
HLD (mg) 510 plusmn 095 353 plusmn 077 373 plusmn 046 453 plusmn 068 098 044SR 21 21 20 19 - -H 189 plusmn 011 196 plusmn 011 167 plusmn 021 194 plusmn 001 102 042D 079 plusmn 002 080 plusmn 003 069 plusmn 007 081 plusmn 001 192 018
SD spore density HLD external hyphal length density SR species richness H ShannonndashWiener index DSimpson index CK no fertilization LN 50 kg Nmiddothaminus1middotyrminus1 MN 100 kg Nmiddothaminus1middotyrminus1 HN 200 kg Nmiddothaminus1middotyrminus1Error bars represent plusmn SE of mean (n = 4)
32 Effects of N Addition on Soil AMF Community Composition
Although there was no significant effect of nitrogen treatment on the Shannon indexand Simpson index nitrogen addition decreased total spore richness (Table 1) The numberof unique AMF species decreased gradually with N addition (Figure 1)
Forests 2022 13 x FOR PEER REVIEW 4 of 16
Table 1 Responses of arbuscular mycorrhizal fungi under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
Coloniza-
tion () 8175 plusmn 225 8475 plusmn 160 8300 plusmn 310 8200 plusmn 270 030 082
SD (spore
gminus1 air-dried
soil)
25569 plusmn
3984
29769 plusmn
3233
36463 plusmn
3785
31894 plusmn
4176 165 023
HLD (mg) 510 plusmn 095 353 plusmn 077 373 plusmn 046 453 plusmn 068 098 044
SR 21 21 20 19 - -
H 189 plusmn 011 196 plusmn 011 167 plusmn 021 194 plusmn 001 102 042
D 079 plusmn 002 080 plusmn 003 069 plusmn 007 081 plusmn 001 192 018
SD spore density HLD external hyphal length density SR species richness H ShannonndashWiener
index D Simpson index CK no fertilization LN 50 kg Nmiddothaminus1middotyrminus1 MN 100kg Nmiddothaminus1middotyrminus1 HN
200kg Nmiddothaminus1middotyrminus1 Error bars represent plusmn SE of mean (n = 4)
32 Effects of N Addition on Soil AMF Community Composition
Although there was no significant effect of nitrogen treatment on the Shannon index
and Simpson index nitrogen addition decreased total spore richness (Table 1) The num-
ber of unique AMF species decreased gradually with N addition (Figure 1)
Figure 1 Venn diagram showing the unique and shared arbuscular mycorrhizal fungi species under
different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN
200kg N haminus1 yrminus1
In total 26 AMF species of spore were detected in four nitrogen treatments (Figure
2 Table 2 Figure A1) These AMF belonged to 12 genera (Acaulospora Archaeospora
Claroideoglomus Dentiscutata Diversispora Entrophospora Funneliformis Gigaspora
Glomus Rhizophagus Sclerocystis and Septoglomus) within six families (Acaulospo-
raceae Ambisporaceae Claroideoglomus Diversisporaceae Gigasporaceae and Glomer-
aceae)
Figure 1 Venn diagram showing the unique and shared arbuscular mycorrhizal fungi species underdifferent nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1HN 200 kg N haminus1 yrminus1
In total 26 AMF species of spore were detected in four nitrogen treatments (Figure 2Table 2 Figure A1) These AMF belonged to 12 genera (Acaulospora ArchaeosporaClaroideoglomus Dentiscutata Diversispora Entrophospora Funneliformis GigasporaGlomus Rhizophagus Sclerocystis and Septoglomus) within six families (AcaulosporaceaeAmbisporaceae Claroideoglomus Diversisporaceae Gigasporaceae and Glomeraceae)
Forests 2022 13 x FOR PEER REVIEW 5 of 16
(a) (b)
Figure 2 The frequency of occurrences (a) and relative abundance (b) among arbuscular mycor-
rhizal fungi under different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN
100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Table 2 Occurrence frequency relative abundance and important values of arbuscular mycorrhizal
fungi species in Chinese fir under nitrogen treatment
Species
Number AMF
Nitrogen
Treatment FO () RA () IV ()
CK LN MN HN
1 Acaulospora bierticulata + + + + 8125 1179 4652
2 Acaulospora denticulata minus + + minus 1250 159 705
3 Acaulospora excavat + + + + 9375 8163 8769
4 Acaulospora foveata + + + + 2500 499 1500
5 Archaeospora schenckii + minus minus minus 625 091 358
6 Claroideoglomus claroideum + + minus minus 1250 1500 1375
7 Claroideoglomus ethunicatum + + + + 4375 8500 6438
8 Dentiscutata heterogama minus minus + + 1250 10000 5625
9 Diversispora etunicatum minus + + + 2500 10000 6250
10 Entrophospora infrequens + minus minus minus 625 10000 5313
11 Funneliformis mosseae + + + + 3750 10000 6875
12 Funneliformis geosporum + + + + 3125 5441 4283
13 Gigaspora albida minus + + + 2500 4559 3530
14 Glomus aggregatum + minus + minus 625 10000 5313
15 Glomus ambisporum + minus minus minus 625 221 423
16 Glomus clarum + + + + 9375 012 4694
17 Glomus melanosporum + + + + 6875 639 3757
18 Glomus microaggregatum + + minus + 1875 314 1095
19 Glomus multicaule + + + + 6250 372 3311
20 Glomus multiforum + + + + 9375 2753 6064
21 Glomus reticulatum + + + + 10000 3461 6731
22 Rhizophagus aggregatus + + + + 10000 9739 9870
23 Rhizophagus intraradices minus + minus minus 625 261 443
24 Sclerocystis liquidambaris + + + + 3750 4098 3924
25 Sclerocystis sinuosum + + + + 8750 5902 7326
26 Septoglomus viscosum + + + + 10000 10000 10000
FO RA and IV are the frequency of occurrences relative abundance and importance value respec-
tively CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Figure 2 The frequency of occurrences (a) and relative abundance (b) among arbuscular mycor-rhizal fungi under different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Forests 2022 13 979 5 of 14
Table 2 Occurrence frequency relative abundance and important values of arbuscular mycorrhizalfungi species in Chinese fir under nitrogen treatment
Species Number AMFNitrogen Treatment
FO () RA () IV ()CK LN MN HN
1 Acaulospora bierticulata + + + + 8125 1179 46522 Acaulospora denticulata minus + + minus 1250 159 7053 Acaulospora excavat + + + + 9375 8163 87694 Acaulospora foveata + + + + 2500 499 15005 Archaeospora schenckii + minus minus minus 625 091 3586 Claroideoglomus claroideum + + minus minus 1250 1500 13757 Claroideoglomus ethunicatum + + + + 4375 8500 64388 Dentiscutata heterogama minus minus + + 1250 10000 56259 Diversispora etunicatum minus + + + 2500 10000 625010 Entrophospora infrequens + minus minus minus 625 10000 531311 Funneliformis mosseae + + + + 3750 10000 687512 Funneliformis geosporum + + + + 3125 5441 428313 Gigaspora albida minus + + + 2500 4559 353014 Glomus aggregatum + minus + minus 625 10000 531315 Glomus ambisporum + minus minus minus 625 221 42316 Glomus clarum + + + + 9375 012 469417 Glomus melanosporum + + + + 6875 639 375718 Glomus microaggregatum + + minus + 1875 314 109519 Glomus multicaule + + + + 6250 372 331120 Glomus multiforum + + + + 9375 2753 606421 Glomus reticulatum + + + + 10000 3461 673122 Rhizophagus aggregatus + + + + 10000 9739 987023 Rhizophagus intraradices minus + minus minus 625 261 44324 Sclerocystis liquidambaris + + + + 3750 4098 392425 Sclerocystis sinuosum + + + + 8750 5902 732626 Septoglomus viscosum + + + + 10000 10000 10000
FO RA and IV are the frequency of occurrences relative abundance and importance value respectively CK nofertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris andSeptoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum(100) gt Rhizophagus aggregatus (9870) gt Acaulospora excavat (8769) gt Sclerocystis sinuo-sum (7326) gt Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followedby Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomusetunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglomusviscosum had the highest IV followed by Acaulospora excavat (9042) Rhizophagus ag-gregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum (6956)(Figure 3)
Forests 2022 13 979 6 of 14
Forests 2022 13 x FOR PEER REVIEW 6 of 16
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-
mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris and
Septoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum
(100) Rhizophagus aggregatus (9870) Acaulospora excavat (8769) Sclerocystis sin-
uosum (7326) Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)
had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus
(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-
cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)
Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-
tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followed
by Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomus
etunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglo-
mus viscosum had the highest IV followed by Acaulospora excavat (9042) Rhizopha-
gus aggregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum
(6956) (Figure 3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitrogen treat-
ments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soil
pH However N addition increased soil N under LN treatment (+1505) Under LN treat-
ment soil AP and TP were the highest Soil NP increased slightly with N addition (Table
3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitro-gen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soilpH However N addition increased soil N under LN treatment (+1505) Under LNtreatment soil AP and TP were the highest Soil NP increased slightly with N addition(Table 3)
Table 3 Soil properties under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
pH 400 plusmn 008 402 plusmn 018 383 plusmn 008 393 plusmn 008 057 064TOC (gkg) 139 plusmn 009 124 plusmn 026 136 plusmn 014 150 plusmn 011 135 030
NH4+-N (mgkg) 2807 plusmn 104 2948 plusmn 192 3248 plusmn 291 3357 plusmn 305 116 037
AN (mgkg) 6143 plusmn 054 6090 plusmn 260 6452 plusmn 384 6465 plusmn 276 054 067AP (mgkg) 782 plusmn 096 898 plusmn 052 803 plusmn 073 805 plusmn 059 051 069
C (gkg) 8978 plusmn 406 9875 plusmn035 9310 plusmn 525 9168 plusmn 320 070 058N (gkg) 578 plusmn 025 665 plusmn 028 600 plusmn 028 565 plusmn 013 337 006
P (mgkg) 23464 plusmn 2257 23981 plusmn 2102 22852 plusmn 1505 22315 plusmn 2556 012 095CN 1554 plusmn 005 1567 plusmn 059 1655 plusmn 068 1622 plusmn 033 103 042NP 2531 plusmn 260 2562 plusmn 278 2673 plusmn 250 2681 plusmn 170 010 096
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus
C total carbon N total nitrogen P total phosphorus CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1 One-way ANOVA and LSD were used to analyze the differencesError bars indicate plusmn SE of mean (n = 4)
34 Relationship between Soil Properties and AMF
AMF colonization was negatively correlated with soil TOC (p lt 005) (Table 4) externalHLD was negatively connected with soil P (p lt 005) spore density was positively relatedto soil C (p lt 005) and species richness was in a positive correlation with soil TOC and C(p lt 005) The Shannon indices presented a positive correlation with soil pH (p lt 005)
Forests 2022 13 979 7 of 14
Table 4 Spearman correlation coefficients between soil properties and parameters of arbuscularmycorrhizal fungi
pH TOC NH4+-N AN AP C N P CN NP
Colonization 0012 minus0643 minus0437 minus0571 minus0245 minus014 minus0126 0142 0025 minus0112HLD minus0062 0267 0245 0288 minus0531 minus026 minus0197 minus0701 minus0154 0564SD minus0338 0214 0008 0251 0161 0679 0477 0283 0489 0002SR 0101 0604 0488 0551 0114 063 0478 0039 033 0117H 0623 0478 0341 0142 0081 0306 0328 minus0023 minus0098 0117D 0560 0408 0264 0105 012 0193 0209 0002 minus0066 007
Acaulospora minus0378 minus0575 minus0506 minus052 0004 minus0332 minus0418 0575 0332 minus0708 Archaeospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025
Claroideoglomus 0464 minus0303 0302 minus0029 minus0047 0077 0175 0071 minus0268 minus0045Dentiscutata 0014 0244 0434 0451 017 0453 0349 0165 0203 minus0047Diversispora 0057 0482 minus0205 minus0057 minus0051 minus0173 minus0304 0384 0399 minus0447
Entrophospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025Funneliformis 005 0764 0175 0266 minus0045 minus0163 minus0201 minus0093 0113 minus0033
Gigaspora minus0644 minus0385 minus0672 minus0466 0042 minus024 minus0419 0636 0581 minus0711 Glomus 0341 minus0133 0242 0033 minus0086 0199 0279 minus0363 minus0258 0393
Rhizophagus 0275 minus0152 minus0376 minus0438 minus0378 0023 0081 minus0056 minus0124 0208Septogolomus minus0138 0252 023 0404 021 0179 0193 minus0194 minus0104 0288
Ancestral minus0367 minus0508 minus0508 minus0524 minus0036 minus0425 minus0513 0575 0347 minus0757 Edaphophilic minus0083 0381 minus0341 minus0154 minus004 minus0218 minus0381 0505 0509 minus0582 Rhizophilic 0361 0431 053 052 004 0435 0541 minus0617 minus04 0801
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C
total carbon N total nitrogen P total phosphorus HLD external hyphal length density SD spore density SRspecies richness H ShannonndashWiener index D Simpson index p lt 005 p lt 001
Archaeospora was in a negative correlation with soil NP (p lt 005) Funneliformis waspositively correlated with soil TOC (p lt 001) and Gigaspora was negatively connectedwith soil pH NH4
+-N (p lt 005) and soil NP (p lt 001) but positively related to soil Pand CN (p lt 005) Ancestral AMF was in a negative correlation with soil NP (p lt 001)edaphophilic AMF was negatively correlated with soil NP (p lt 005) and rhizophilic AMFwas positively connected with soil NP (p lt 001) but negatively related to soil P (p lt 005)
RDA was conducted to evaluate the relationship between the soil chemical properties and theAMF community (Figure 4) RDA1 accounted for 6699 of the variations while RDA2 accountedfor 2189 Of the evaluated attributes the NP had significant conditional effects (p lt 001) whilethe soil TOC (p = 0076) and AN (p = 0092) caused referential effects to the AMF communities
Forests 2022 13 x FOR PEER REVIEW 8 of 16
density SD spore density SR species richness H ShannonndashWiener index D Simpson index p lt
005 p lt 001
Archaeospora was in a negative correlation with soil NP (p 005) Funneliformis was
positively correlated with soil TOC (p 001) and Gigaspora was negatively connected
with soil pH NH4+-N (p 005) and soil NP (p 001) but positively related to soil P and
CN (p 005) Ancestral AMF was in a negative correlation with soil NP (p 001)
edaphophilic AMF was negatively correlated with soil NP (p 005) and rhizophilic AMF
was positively connected with soil NP (p 001) but negatively related to soil P (p 005)
RDA was conducted to evaluate the relationship between the soil chemical properties
and the AMF community (Figure 4) RDA1 accounted for 6699 of the variations while
RDA2 accounted for 2189 Of the evaluated attributes the NP had significant condi-
tional effects (p 001) while the soil TOC (p = 0076) and AN (p = 0092) caused referential
effects to the AMF communities
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular
mycorrhizal fungi communities under different nitrogen treatments TOC total organic carbon
NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C total carbon
N total nitrogen P total phosphorus The red arrows represent soil properties the blue arrows
represent arbuscular mycorrhizal fungi communities
4 Discussion
N addition can make a difference on AMF diversity and community composition
because with the increase of soil nutrients plants would have a strategy to acquire nutri-
ents by fine roots or AMF [2644] Our results showed that NP is a factor that has a sig-
nificant effect on AMF communities (Figure 4) AMF community structure might have a
connection with the dynamics of soil conditions and C-to-P trade between AMF and
plants It has been agreed that AMF exchanges P derived from soil for plant carbon [1744ndash
47] In our study N addition had insignificant influence on AMF colonization external
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular mycorrhizalfungi communities under different nitrogen treatments TOC total organic carbon NH4
+-N ammoniumnitrogen AN available nitrogen AP available phosphorus C total carbon N total nitrogen P total phosphorusThe red arrows represent soil properties the blue arrows represent arbuscular mycorrhizal fungi communities
Forests 2022 13 979 8 of 14
4 Discussion
N addition can make a difference on AMF diversity and community compositionbecause with the increase of soil nutrients plants would have a strategy to acquire nutrientsby fine roots or AMF [2644] Our results showed that NP is a factor that has a significanteffect on AMF communities (Figure 4) AMF community structure might have a connectionwith the dynamics of soil conditions and C-to-P trade between AMF and plants It has beenagreed that AMF exchanges P derived from soil for plant carbon [1744ndash47] In our studyN addition had insignificant influence on AMF colonization external HLD spore densityor diversity This finding may be explained as the symbiotic relationship between AMFand root system being stable [48]
Generally there is little specificity between AMF and the host plant and severalAMF species could colonize one AM plant at the same time [4950] However differentpatterns of colonization with N addition suggest a preference for specific AMF species [51]In our study we observed that Glomeraceae was the most predominant family amongall treatments Glomus Septoglomus and Acaulospora were the predominant genera andSeptoglomus viscosum and Acaulospora foveata were the predominant species Our resultsslightly disagree with previous studies which presented that Glomus and Acaulospora asthe predominant species due to their smaller size of spore which helped them producemore spores in less time [485253] This discrepancy can be answered by the diversespecies and age of the host plants The high isolation frequencies of Septoglomus viscosumindicated that it has a strong ability to adapt to the environment (N deposition) Septoglomusviscosum can improve plant growth and production [54] which is mainly related to rootand aboveground biomass [55]
Archaeospora schenckii Entrophospora infrequens and Glomus ambisporum disappearedwith N treatments (Figure 3) which was in keeping with Eom [19] who also found thatEntrophospora infrequens decreased with N addition In P-rich (low NP) soils nitrogenapplication reduces underground carbon allocation and rare species decrease as N in-creases A previous study showed that the influence of N addition on AMF communityappeared different with added N promoting Glomeraceae in A Sieversiana while lesseningthem in L chinensis [16] Some Glomus species were sensitive to N application whichindicated that certain species of AMF play an indispensable role in host plant response toenvironmental change
As identified in most surveys conducted in other fields [56ndash58] we also discoveredFunneliformis mosseae presenting in our study which was reported to promote plant nutri-ents [59] GmosAAP1 an amino acid permease has been characterized from the extraradicalstructure of Funneliformis mosseae which indicates a potential for amino acid uptake [60]Our study showed that with N addition its importance decreased This decrease maybe explained by a decrease in the dependency on Funneliformis mosseae for N nutrientacquisition by the host plant
Above all our results showed that AMF participating in and manage the soil nutrientcycle could be sensitive to small changes in edatopes Therefore studying mycorrhizalsymbionts increases the knowledge of climatic variation [61ndash63]
AMF were grouped into three guilds namely edaphophilic rhizophilc and ancestralIn our study the percentage followed the order rhizophilic gt edaphophilic gt ancestral(Figure 5c) N addition altered AMF species but did not change functional groups Despitethe addition of nitrogen trees may reduce dependence on AMF Meanwhile a large groupof rhizophilic guilds function to reduce root pathogen infection and primarily benefit plantswith fine roots prone to pathogen infections
Forests 2022 13 979 9 of 14Forests 2022 13 x FOR PEER REVIEW 10 of 16
Figure 5 Cont
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-

Forests 2022 13 979 2 of 14
promoted [17] or have no significant effect [18] by N addition N addition can also en-hance [1819] suppress [20] or have no significant effect [21] on AMF spore density andhyphae Some investigations showed that N addition treatments caused significant sup-pressive impacts on AMF abundance richness and diversity [2223] whereas others foundno significant impact [1924] Meanwhile how these environmental factors shape the AMFcommunity under N addition is not fully understood A study observed that the relativeabundance of AMF decreased the reduction of plant demand for N [25] Importantly themorphological modification of AMF caused by soil nutrient changes may be relevant totheir composition owing to the differences of morphological internal responses to nutrientmanipulations [2627]
Chinese fir (Cunninghamia lanceolata) is a quick-growing timber tree species in southChina [28] On account of its high wood quality and termite resistance it is frequentlyplanted [29] However the productivity of the Chinese fir is declining due to soil acidi-fication soil aluminum poisoning and unreasonable planting methods Lu [30] showedthat the number of AMF operational taxonomic units (OTUs) had an inverted V-shapedchange with the age of the Chinese fir Liu [14] found that the Chinese fir under N additionfor 2 years would increase AMF colonization and diversity Cao [31] observed that high Naddition exerted a significant negative effect on AMF community However few studiesshowed the interactions of Chinese fir and AMF in terms of N deposition and nutrientavailability Therefore we purposed to address these questions (1) how does N additionaffect AMF colonization and the diversity and composition of AMF in Chinese fir (2) wouldthe change of soil nutrients availability affect the responses of AMF groups
2 Materials and Methods21 Study Sites and Sampling
The soil experiment was carried out in the Fengyang Mountain Nature Reserve(2853prime56primeprime N 19010prime56primeprime E) Zhejiang Province China The mean annual precipitationand annual temperature are 2400 mm and 123 C Soil properties (0ndash10 cm) are totalnitrogen (TN) 411 g kgminus1 total carbon (TC) 5873 g kgminus1 NH4
+-N 1571 mg gminus1 NO3minus-
N 1466 mg gminus1 total phosphorus (TP) 03 g kgminus1 and bulk density 076 g cmminus3 TheChinese fir was 39 years old with an average diameter at breast height of 2175 cm and132 m height
The local N deposition was estimated to be 34 kg N haminus1 yrminus1 The N addition treat-ments included four different levels (four plots per treatment were considered as four inde-pendent repetitions sixteen 10 m times 10 m plots in total) to simulate future climate changescenarios no fertilization 50 kg N haminus1 yrminus1 100 kg N haminus1 yrminus1 and 200 kg N haminus1 yrminus1which were regarded as control (CK) low-nitrogen (LN) medium-nitrogen (MN) andhigh-nitrogen (HN) respectively Separated adjacent plots were separated by a 10-m-widebuffer strip From April 2019 N was added monthly by spraying urea (CO (NH2)2) solutionon the forest floor as N addition treatments Meanwhile CK plots were sprayed with thesame volume of water
Samples were collected in November 2020 Along the ldquoSrdquo route six standard treeswere selected in each plot Weeds and litter around the sample trees were removed beforecollecting Roots and rhizosphere soil around the average standard tree were collected inthe 0ndash20 cm soil layer from four directions The fine roots and about 1000 g of rhizospheresoil (soils adhering to roots by shaking off root surface) from six standard trees werecollected and mixed as one root and one soil sample in each plot and sixteen root andsoil samples were collected The soil samples were used for soil physicochemical analysisand the measurement of AMF colonization spore isolation studies and measured hyphallength density (HLD) The soil samples were air-dried and the properties were measuredwhile the other soil samples were stored with root samples (washed using tap water) in therefrigerator at 4 C until used
Forests 2022 13 979 3 of 14
22 Soil Properties Analysis
All the soil samples were sieved through a 2 mm sieve Soil pH was measured in asoil-to-water ratio of 125 (vv) Soil NH4
+-N and NO3minus-N concentration were estimated
by colorimetric analysis [32] Soil available nitrogen (AN) is soil NH4+-N plus NO3
minus-N Soil available phosphorus (AP) and total phosphorus (TP) were extracted by sodiumbicarbonate first and their contents were followed by molybdenum blue colorimetry [33]Soil total organic carbon was measured by TOC-L CPHCPN [34] Soil total carbon (C) andtotal nitrogen (N) were measured by an elemental analyzer (Vario MACRO cube ElementarTrading Shanghai Shanghai China)
23 AMF Colonization and Extraradical Hphae Length Density
To highlight AMF structures roots samples were strained following the methodsdescribed by Philips and Hayman [35] and the percentage of mycorrhizal colonization wasestimated in the light of McGonigle et al [36] Extraradical HLD was determined on thebasis of the description by Jakoben et al [37]
24 AMF Spore Separation and Identification
The AMF spores used for identification were obtained from the 20 g air-dried rhizo-sphere soil samples following the wet sieving and decanting techniques as described byBrundrett [38] In order to determine the spore density the isolated intact healthy sporeswere extracted and placed on filter paper We separated them by color spore size orna-mentations hyphal attachments and wall layers and then used a dissecting microscopeat 45timesmagnification to observe them Spores were mounted on slides in polyvinylndashlacticacidndashglycerine (PVLG) and a mixture of PVLG with Melzerrsquos (11 vv) The spores wereidentified according to the International Culture Collection of Vesicular Arbuscular Mycor-rhizal fungi (INVAM) (httpinvamcafwvuedu last accessed in 21 May 2022) and recentadvances in Glomeromycota taxonomy [3940]
25 AMF Community Parameters and Statistical Analysis
The spore density (SD) frequency of occurrence (FO) relative abundance (RA) impor-tance value (IA) ShannonndashWiener index (H) Simpson index (D) species richness (SR) andpercentage were used to estimate the AMF community structure [41ndash43]
Correlation analysis was performed using SPSS 260 to test for relationship betweenthe AMF community composition parameters and soil properties Venn was pictured forVenny 20 Graphical presentation was carried out using Origin 2022 Redundancy analysis(RDA) was conducted with Canoco 50 to reveal the influence of soil properties on theAMF communities
3 Results31 Effects of N Addition on Soil AMF Colonization Spore Density and External HyphalLength Density
AMF colonization (p = 0824) spore density (p = 0229) and external hyphal lengthdensity (p = 0435) were not significantly influenced by the nitrogen treatment (Table 1) TheAMF colonization ranged from 74 to 88 Spore density was highest in MN treatmentwhich averaged up to 365 spores per 1g air-dried soil and spore density was lowest value inthe control treatment External HLD was highest in the control treatment which averagedup to 510 m gminus1 and was the lowest in the LN treatment
Forests 2022 13 979 4 of 14
Table 1 Responses of arbuscular mycorrhizal fungi under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
Colonization () 8175 plusmn 225 8475 plusmn 160 8300 plusmn 310 8200 plusmn 270 030 082SD (spore gminus1 air-dried soil) 25569 plusmn 3984 29769 plusmn 3233 36463 plusmn 3785 31894 plusmn 4176 165 023
HLD (mg) 510 plusmn 095 353 plusmn 077 373 plusmn 046 453 plusmn 068 098 044SR 21 21 20 19 - -H 189 plusmn 011 196 plusmn 011 167 plusmn 021 194 plusmn 001 102 042D 079 plusmn 002 080 plusmn 003 069 plusmn 007 081 plusmn 001 192 018
SD spore density HLD external hyphal length density SR species richness H ShannonndashWiener index DSimpson index CK no fertilization LN 50 kg Nmiddothaminus1middotyrminus1 MN 100 kg Nmiddothaminus1middotyrminus1 HN 200 kg Nmiddothaminus1middotyrminus1Error bars represent plusmn SE of mean (n = 4)
32 Effects of N Addition on Soil AMF Community Composition
Although there was no significant effect of nitrogen treatment on the Shannon indexand Simpson index nitrogen addition decreased total spore richness (Table 1) The numberof unique AMF species decreased gradually with N addition (Figure 1)
Forests 2022 13 x FOR PEER REVIEW 4 of 16
Table 1 Responses of arbuscular mycorrhizal fungi under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
Coloniza-
tion () 8175 plusmn 225 8475 plusmn 160 8300 plusmn 310 8200 plusmn 270 030 082
SD (spore
gminus1 air-dried
soil)
25569 plusmn
3984
29769 plusmn
3233
36463 plusmn
3785
31894 plusmn
4176 165 023
HLD (mg) 510 plusmn 095 353 plusmn 077 373 plusmn 046 453 plusmn 068 098 044
SR 21 21 20 19 - -
H 189 plusmn 011 196 plusmn 011 167 plusmn 021 194 plusmn 001 102 042
D 079 plusmn 002 080 plusmn 003 069 plusmn 007 081 plusmn 001 192 018
SD spore density HLD external hyphal length density SR species richness H ShannonndashWiener
index D Simpson index CK no fertilization LN 50 kg Nmiddothaminus1middotyrminus1 MN 100kg Nmiddothaminus1middotyrminus1 HN
200kg Nmiddothaminus1middotyrminus1 Error bars represent plusmn SE of mean (n = 4)
32 Effects of N Addition on Soil AMF Community Composition
Although there was no significant effect of nitrogen treatment on the Shannon index
and Simpson index nitrogen addition decreased total spore richness (Table 1) The num-
ber of unique AMF species decreased gradually with N addition (Figure 1)
Figure 1 Venn diagram showing the unique and shared arbuscular mycorrhizal fungi species under
different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN
200kg N haminus1 yrminus1
In total 26 AMF species of spore were detected in four nitrogen treatments (Figure
2 Table 2 Figure A1) These AMF belonged to 12 genera (Acaulospora Archaeospora
Claroideoglomus Dentiscutata Diversispora Entrophospora Funneliformis Gigaspora
Glomus Rhizophagus Sclerocystis and Septoglomus) within six families (Acaulospo-
raceae Ambisporaceae Claroideoglomus Diversisporaceae Gigasporaceae and Glomer-
aceae)
Figure 1 Venn diagram showing the unique and shared arbuscular mycorrhizal fungi species underdifferent nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1HN 200 kg N haminus1 yrminus1
In total 26 AMF species of spore were detected in four nitrogen treatments (Figure 2Table 2 Figure A1) These AMF belonged to 12 genera (Acaulospora ArchaeosporaClaroideoglomus Dentiscutata Diversispora Entrophospora Funneliformis GigasporaGlomus Rhizophagus Sclerocystis and Septoglomus) within six families (AcaulosporaceaeAmbisporaceae Claroideoglomus Diversisporaceae Gigasporaceae and Glomeraceae)
Forests 2022 13 x FOR PEER REVIEW 5 of 16
(a) (b)
Figure 2 The frequency of occurrences (a) and relative abundance (b) among arbuscular mycor-
rhizal fungi under different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN
100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Table 2 Occurrence frequency relative abundance and important values of arbuscular mycorrhizal
fungi species in Chinese fir under nitrogen treatment
Species
Number AMF
Nitrogen
Treatment FO () RA () IV ()
CK LN MN HN
1 Acaulospora bierticulata + + + + 8125 1179 4652
2 Acaulospora denticulata minus + + minus 1250 159 705
3 Acaulospora excavat + + + + 9375 8163 8769
4 Acaulospora foveata + + + + 2500 499 1500
5 Archaeospora schenckii + minus minus minus 625 091 358
6 Claroideoglomus claroideum + + minus minus 1250 1500 1375
7 Claroideoglomus ethunicatum + + + + 4375 8500 6438
8 Dentiscutata heterogama minus minus + + 1250 10000 5625
9 Diversispora etunicatum minus + + + 2500 10000 6250
10 Entrophospora infrequens + minus minus minus 625 10000 5313
11 Funneliformis mosseae + + + + 3750 10000 6875
12 Funneliformis geosporum + + + + 3125 5441 4283
13 Gigaspora albida minus + + + 2500 4559 3530
14 Glomus aggregatum + minus + minus 625 10000 5313
15 Glomus ambisporum + minus minus minus 625 221 423
16 Glomus clarum + + + + 9375 012 4694
17 Glomus melanosporum + + + + 6875 639 3757
18 Glomus microaggregatum + + minus + 1875 314 1095
19 Glomus multicaule + + + + 6250 372 3311
20 Glomus multiforum + + + + 9375 2753 6064
21 Glomus reticulatum + + + + 10000 3461 6731
22 Rhizophagus aggregatus + + + + 10000 9739 9870
23 Rhizophagus intraradices minus + minus minus 625 261 443
24 Sclerocystis liquidambaris + + + + 3750 4098 3924
25 Sclerocystis sinuosum + + + + 8750 5902 7326
26 Septoglomus viscosum + + + + 10000 10000 10000
FO RA and IV are the frequency of occurrences relative abundance and importance value respec-
tively CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Figure 2 The frequency of occurrences (a) and relative abundance (b) among arbuscular mycor-rhizal fungi under different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Forests 2022 13 979 5 of 14
Table 2 Occurrence frequency relative abundance and important values of arbuscular mycorrhizalfungi species in Chinese fir under nitrogen treatment
Species Number AMFNitrogen Treatment
FO () RA () IV ()CK LN MN HN
1 Acaulospora bierticulata + + + + 8125 1179 46522 Acaulospora denticulata minus + + minus 1250 159 7053 Acaulospora excavat + + + + 9375 8163 87694 Acaulospora foveata + + + + 2500 499 15005 Archaeospora schenckii + minus minus minus 625 091 3586 Claroideoglomus claroideum + + minus minus 1250 1500 13757 Claroideoglomus ethunicatum + + + + 4375 8500 64388 Dentiscutata heterogama minus minus + + 1250 10000 56259 Diversispora etunicatum minus + + + 2500 10000 625010 Entrophospora infrequens + minus minus minus 625 10000 531311 Funneliformis mosseae + + + + 3750 10000 687512 Funneliformis geosporum + + + + 3125 5441 428313 Gigaspora albida minus + + + 2500 4559 353014 Glomus aggregatum + minus + minus 625 10000 531315 Glomus ambisporum + minus minus minus 625 221 42316 Glomus clarum + + + + 9375 012 469417 Glomus melanosporum + + + + 6875 639 375718 Glomus microaggregatum + + minus + 1875 314 109519 Glomus multicaule + + + + 6250 372 331120 Glomus multiforum + + + + 9375 2753 606421 Glomus reticulatum + + + + 10000 3461 673122 Rhizophagus aggregatus + + + + 10000 9739 987023 Rhizophagus intraradices minus + minus minus 625 261 44324 Sclerocystis liquidambaris + + + + 3750 4098 392425 Sclerocystis sinuosum + + + + 8750 5902 732626 Septoglomus viscosum + + + + 10000 10000 10000
FO RA and IV are the frequency of occurrences relative abundance and importance value respectively CK nofertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris andSeptoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum(100) gt Rhizophagus aggregatus (9870) gt Acaulospora excavat (8769) gt Sclerocystis sinuo-sum (7326) gt Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followedby Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomusetunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglomusviscosum had the highest IV followed by Acaulospora excavat (9042) Rhizophagus ag-gregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum (6956)(Figure 3)
Forests 2022 13 979 6 of 14
Forests 2022 13 x FOR PEER REVIEW 6 of 16
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-
mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris and
Septoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum
(100) Rhizophagus aggregatus (9870) Acaulospora excavat (8769) Sclerocystis sin-
uosum (7326) Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)
had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus
(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-
cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)
Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-
tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followed
by Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomus
etunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglo-
mus viscosum had the highest IV followed by Acaulospora excavat (9042) Rhizopha-
gus aggregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum
(6956) (Figure 3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitrogen treat-
ments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soil
pH However N addition increased soil N under LN treatment (+1505) Under LN treat-
ment soil AP and TP were the highest Soil NP increased slightly with N addition (Table
3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitro-gen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soilpH However N addition increased soil N under LN treatment (+1505) Under LNtreatment soil AP and TP were the highest Soil NP increased slightly with N addition(Table 3)
Table 3 Soil properties under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
pH 400 plusmn 008 402 plusmn 018 383 plusmn 008 393 plusmn 008 057 064TOC (gkg) 139 plusmn 009 124 plusmn 026 136 plusmn 014 150 plusmn 011 135 030
NH4+-N (mgkg) 2807 plusmn 104 2948 plusmn 192 3248 plusmn 291 3357 plusmn 305 116 037
AN (mgkg) 6143 plusmn 054 6090 plusmn 260 6452 plusmn 384 6465 plusmn 276 054 067AP (mgkg) 782 plusmn 096 898 plusmn 052 803 plusmn 073 805 plusmn 059 051 069
C (gkg) 8978 plusmn 406 9875 plusmn035 9310 plusmn 525 9168 plusmn 320 070 058N (gkg) 578 plusmn 025 665 plusmn 028 600 plusmn 028 565 plusmn 013 337 006
P (mgkg) 23464 plusmn 2257 23981 plusmn 2102 22852 plusmn 1505 22315 plusmn 2556 012 095CN 1554 plusmn 005 1567 plusmn 059 1655 plusmn 068 1622 plusmn 033 103 042NP 2531 plusmn 260 2562 plusmn 278 2673 plusmn 250 2681 plusmn 170 010 096
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus
C total carbon N total nitrogen P total phosphorus CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1 One-way ANOVA and LSD were used to analyze the differencesError bars indicate plusmn SE of mean (n = 4)
34 Relationship between Soil Properties and AMF
AMF colonization was negatively correlated with soil TOC (p lt 005) (Table 4) externalHLD was negatively connected with soil P (p lt 005) spore density was positively relatedto soil C (p lt 005) and species richness was in a positive correlation with soil TOC and C(p lt 005) The Shannon indices presented a positive correlation with soil pH (p lt 005)
Forests 2022 13 979 7 of 14
Table 4 Spearman correlation coefficients between soil properties and parameters of arbuscularmycorrhizal fungi
pH TOC NH4+-N AN AP C N P CN NP
Colonization 0012 minus0643 minus0437 minus0571 minus0245 minus014 minus0126 0142 0025 minus0112HLD minus0062 0267 0245 0288 minus0531 minus026 minus0197 minus0701 minus0154 0564SD minus0338 0214 0008 0251 0161 0679 0477 0283 0489 0002SR 0101 0604 0488 0551 0114 063 0478 0039 033 0117H 0623 0478 0341 0142 0081 0306 0328 minus0023 minus0098 0117D 0560 0408 0264 0105 012 0193 0209 0002 minus0066 007
Acaulospora minus0378 minus0575 minus0506 minus052 0004 minus0332 minus0418 0575 0332 minus0708 Archaeospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025
Claroideoglomus 0464 minus0303 0302 minus0029 minus0047 0077 0175 0071 minus0268 minus0045Dentiscutata 0014 0244 0434 0451 017 0453 0349 0165 0203 minus0047Diversispora 0057 0482 minus0205 minus0057 minus0051 minus0173 minus0304 0384 0399 minus0447
Entrophospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025Funneliformis 005 0764 0175 0266 minus0045 minus0163 minus0201 minus0093 0113 minus0033
Gigaspora minus0644 minus0385 minus0672 minus0466 0042 minus024 minus0419 0636 0581 minus0711 Glomus 0341 minus0133 0242 0033 minus0086 0199 0279 minus0363 minus0258 0393
Rhizophagus 0275 minus0152 minus0376 minus0438 minus0378 0023 0081 minus0056 minus0124 0208Septogolomus minus0138 0252 023 0404 021 0179 0193 minus0194 minus0104 0288
Ancestral minus0367 minus0508 minus0508 minus0524 minus0036 minus0425 minus0513 0575 0347 minus0757 Edaphophilic minus0083 0381 minus0341 minus0154 minus004 minus0218 minus0381 0505 0509 minus0582 Rhizophilic 0361 0431 053 052 004 0435 0541 minus0617 minus04 0801
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C
total carbon N total nitrogen P total phosphorus HLD external hyphal length density SD spore density SRspecies richness H ShannonndashWiener index D Simpson index p lt 005 p lt 001
Archaeospora was in a negative correlation with soil NP (p lt 005) Funneliformis waspositively correlated with soil TOC (p lt 001) and Gigaspora was negatively connectedwith soil pH NH4
+-N (p lt 005) and soil NP (p lt 001) but positively related to soil Pand CN (p lt 005) Ancestral AMF was in a negative correlation with soil NP (p lt 001)edaphophilic AMF was negatively correlated with soil NP (p lt 005) and rhizophilic AMFwas positively connected with soil NP (p lt 001) but negatively related to soil P (p lt 005)
RDA was conducted to evaluate the relationship between the soil chemical properties and theAMF community (Figure 4) RDA1 accounted for 6699 of the variations while RDA2 accountedfor 2189 Of the evaluated attributes the NP had significant conditional effects (p lt 001) whilethe soil TOC (p = 0076) and AN (p = 0092) caused referential effects to the AMF communities
Forests 2022 13 x FOR PEER REVIEW 8 of 16
density SD spore density SR species richness H ShannonndashWiener index D Simpson index p lt
005 p lt 001
Archaeospora was in a negative correlation with soil NP (p 005) Funneliformis was
positively correlated with soil TOC (p 001) and Gigaspora was negatively connected
with soil pH NH4+-N (p 005) and soil NP (p 001) but positively related to soil P and
CN (p 005) Ancestral AMF was in a negative correlation with soil NP (p 001)
edaphophilic AMF was negatively correlated with soil NP (p 005) and rhizophilic AMF
was positively connected with soil NP (p 001) but negatively related to soil P (p 005)
RDA was conducted to evaluate the relationship between the soil chemical properties
and the AMF community (Figure 4) RDA1 accounted for 6699 of the variations while
RDA2 accounted for 2189 Of the evaluated attributes the NP had significant condi-
tional effects (p 001) while the soil TOC (p = 0076) and AN (p = 0092) caused referential
effects to the AMF communities
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular
mycorrhizal fungi communities under different nitrogen treatments TOC total organic carbon
NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C total carbon
N total nitrogen P total phosphorus The red arrows represent soil properties the blue arrows
represent arbuscular mycorrhizal fungi communities
4 Discussion
N addition can make a difference on AMF diversity and community composition
because with the increase of soil nutrients plants would have a strategy to acquire nutri-
ents by fine roots or AMF [2644] Our results showed that NP is a factor that has a sig-
nificant effect on AMF communities (Figure 4) AMF community structure might have a
connection with the dynamics of soil conditions and C-to-P trade between AMF and
plants It has been agreed that AMF exchanges P derived from soil for plant carbon [1744ndash
47] In our study N addition had insignificant influence on AMF colonization external
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular mycorrhizalfungi communities under different nitrogen treatments TOC total organic carbon NH4
+-N ammoniumnitrogen AN available nitrogen AP available phosphorus C total carbon N total nitrogen P total phosphorusThe red arrows represent soil properties the blue arrows represent arbuscular mycorrhizal fungi communities
Forests 2022 13 979 8 of 14
4 Discussion
N addition can make a difference on AMF diversity and community compositionbecause with the increase of soil nutrients plants would have a strategy to acquire nutrientsby fine roots or AMF [2644] Our results showed that NP is a factor that has a significanteffect on AMF communities (Figure 4) AMF community structure might have a connectionwith the dynamics of soil conditions and C-to-P trade between AMF and plants It has beenagreed that AMF exchanges P derived from soil for plant carbon [1744ndash47] In our studyN addition had insignificant influence on AMF colonization external HLD spore densityor diversity This finding may be explained as the symbiotic relationship between AMFand root system being stable [48]
Generally there is little specificity between AMF and the host plant and severalAMF species could colonize one AM plant at the same time [4950] However differentpatterns of colonization with N addition suggest a preference for specific AMF species [51]In our study we observed that Glomeraceae was the most predominant family amongall treatments Glomus Septoglomus and Acaulospora were the predominant genera andSeptoglomus viscosum and Acaulospora foveata were the predominant species Our resultsslightly disagree with previous studies which presented that Glomus and Acaulospora asthe predominant species due to their smaller size of spore which helped them producemore spores in less time [485253] This discrepancy can be answered by the diversespecies and age of the host plants The high isolation frequencies of Septoglomus viscosumindicated that it has a strong ability to adapt to the environment (N deposition) Septoglomusviscosum can improve plant growth and production [54] which is mainly related to rootand aboveground biomass [55]
Archaeospora schenckii Entrophospora infrequens and Glomus ambisporum disappearedwith N treatments (Figure 3) which was in keeping with Eom [19] who also found thatEntrophospora infrequens decreased with N addition In P-rich (low NP) soils nitrogenapplication reduces underground carbon allocation and rare species decrease as N in-creases A previous study showed that the influence of N addition on AMF communityappeared different with added N promoting Glomeraceae in A Sieversiana while lesseningthem in L chinensis [16] Some Glomus species were sensitive to N application whichindicated that certain species of AMF play an indispensable role in host plant response toenvironmental change
As identified in most surveys conducted in other fields [56ndash58] we also discoveredFunneliformis mosseae presenting in our study which was reported to promote plant nutri-ents [59] GmosAAP1 an amino acid permease has been characterized from the extraradicalstructure of Funneliformis mosseae which indicates a potential for amino acid uptake [60]Our study showed that with N addition its importance decreased This decrease maybe explained by a decrease in the dependency on Funneliformis mosseae for N nutrientacquisition by the host plant
Above all our results showed that AMF participating in and manage the soil nutrientcycle could be sensitive to small changes in edatopes Therefore studying mycorrhizalsymbionts increases the knowledge of climatic variation [61ndash63]
AMF were grouped into three guilds namely edaphophilic rhizophilc and ancestralIn our study the percentage followed the order rhizophilic gt edaphophilic gt ancestral(Figure 5c) N addition altered AMF species but did not change functional groups Despitethe addition of nitrogen trees may reduce dependence on AMF Meanwhile a large groupof rhizophilic guilds function to reduce root pathogen infection and primarily benefit plantswith fine roots prone to pathogen infections
Forests 2022 13 979 9 of 14Forests 2022 13 x FOR PEER REVIEW 10 of 16
Figure 5 Cont
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-

Forests 2022 13 979 3 of 14
22 Soil Properties Analysis
All the soil samples were sieved through a 2 mm sieve Soil pH was measured in asoil-to-water ratio of 125 (vv) Soil NH4
+-N and NO3minus-N concentration were estimated
by colorimetric analysis [32] Soil available nitrogen (AN) is soil NH4+-N plus NO3
minus-N Soil available phosphorus (AP) and total phosphorus (TP) were extracted by sodiumbicarbonate first and their contents were followed by molybdenum blue colorimetry [33]Soil total organic carbon was measured by TOC-L CPHCPN [34] Soil total carbon (C) andtotal nitrogen (N) were measured by an elemental analyzer (Vario MACRO cube ElementarTrading Shanghai Shanghai China)
23 AMF Colonization and Extraradical Hphae Length Density
To highlight AMF structures roots samples were strained following the methodsdescribed by Philips and Hayman [35] and the percentage of mycorrhizal colonization wasestimated in the light of McGonigle et al [36] Extraradical HLD was determined on thebasis of the description by Jakoben et al [37]
24 AMF Spore Separation and Identification
The AMF spores used for identification were obtained from the 20 g air-dried rhizo-sphere soil samples following the wet sieving and decanting techniques as described byBrundrett [38] In order to determine the spore density the isolated intact healthy sporeswere extracted and placed on filter paper We separated them by color spore size orna-mentations hyphal attachments and wall layers and then used a dissecting microscopeat 45timesmagnification to observe them Spores were mounted on slides in polyvinylndashlacticacidndashglycerine (PVLG) and a mixture of PVLG with Melzerrsquos (11 vv) The spores wereidentified according to the International Culture Collection of Vesicular Arbuscular Mycor-rhizal fungi (INVAM) (httpinvamcafwvuedu last accessed in 21 May 2022) and recentadvances in Glomeromycota taxonomy [3940]
25 AMF Community Parameters and Statistical Analysis
The spore density (SD) frequency of occurrence (FO) relative abundance (RA) impor-tance value (IA) ShannonndashWiener index (H) Simpson index (D) species richness (SR) andpercentage were used to estimate the AMF community structure [41ndash43]
Correlation analysis was performed using SPSS 260 to test for relationship betweenthe AMF community composition parameters and soil properties Venn was pictured forVenny 20 Graphical presentation was carried out using Origin 2022 Redundancy analysis(RDA) was conducted with Canoco 50 to reveal the influence of soil properties on theAMF communities
3 Results31 Effects of N Addition on Soil AMF Colonization Spore Density and External HyphalLength Density
AMF colonization (p = 0824) spore density (p = 0229) and external hyphal lengthdensity (p = 0435) were not significantly influenced by the nitrogen treatment (Table 1) TheAMF colonization ranged from 74 to 88 Spore density was highest in MN treatmentwhich averaged up to 365 spores per 1g air-dried soil and spore density was lowest value inthe control treatment External HLD was highest in the control treatment which averagedup to 510 m gminus1 and was the lowest in the LN treatment
Forests 2022 13 979 4 of 14
Table 1 Responses of arbuscular mycorrhizal fungi under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
Colonization () 8175 plusmn 225 8475 plusmn 160 8300 plusmn 310 8200 plusmn 270 030 082SD (spore gminus1 air-dried soil) 25569 plusmn 3984 29769 plusmn 3233 36463 plusmn 3785 31894 plusmn 4176 165 023
HLD (mg) 510 plusmn 095 353 plusmn 077 373 plusmn 046 453 plusmn 068 098 044SR 21 21 20 19 - -H 189 plusmn 011 196 plusmn 011 167 plusmn 021 194 plusmn 001 102 042D 079 plusmn 002 080 plusmn 003 069 plusmn 007 081 plusmn 001 192 018
SD spore density HLD external hyphal length density SR species richness H ShannonndashWiener index DSimpson index CK no fertilization LN 50 kg Nmiddothaminus1middotyrminus1 MN 100 kg Nmiddothaminus1middotyrminus1 HN 200 kg Nmiddothaminus1middotyrminus1Error bars represent plusmn SE of mean (n = 4)
32 Effects of N Addition on Soil AMF Community Composition
Although there was no significant effect of nitrogen treatment on the Shannon indexand Simpson index nitrogen addition decreased total spore richness (Table 1) The numberof unique AMF species decreased gradually with N addition (Figure 1)
Forests 2022 13 x FOR PEER REVIEW 4 of 16
Table 1 Responses of arbuscular mycorrhizal fungi under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
Coloniza-
tion () 8175 plusmn 225 8475 plusmn 160 8300 plusmn 310 8200 plusmn 270 030 082
SD (spore
gminus1 air-dried
soil)
25569 plusmn
3984
29769 plusmn
3233
36463 plusmn
3785
31894 plusmn
4176 165 023
HLD (mg) 510 plusmn 095 353 plusmn 077 373 plusmn 046 453 plusmn 068 098 044
SR 21 21 20 19 - -
H 189 plusmn 011 196 plusmn 011 167 plusmn 021 194 plusmn 001 102 042
D 079 plusmn 002 080 plusmn 003 069 plusmn 007 081 plusmn 001 192 018
SD spore density HLD external hyphal length density SR species richness H ShannonndashWiener
index D Simpson index CK no fertilization LN 50 kg Nmiddothaminus1middotyrminus1 MN 100kg Nmiddothaminus1middotyrminus1 HN
200kg Nmiddothaminus1middotyrminus1 Error bars represent plusmn SE of mean (n = 4)
32 Effects of N Addition on Soil AMF Community Composition
Although there was no significant effect of nitrogen treatment on the Shannon index
and Simpson index nitrogen addition decreased total spore richness (Table 1) The num-
ber of unique AMF species decreased gradually with N addition (Figure 1)
Figure 1 Venn diagram showing the unique and shared arbuscular mycorrhizal fungi species under
different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN
200kg N haminus1 yrminus1
In total 26 AMF species of spore were detected in four nitrogen treatments (Figure
2 Table 2 Figure A1) These AMF belonged to 12 genera (Acaulospora Archaeospora
Claroideoglomus Dentiscutata Diversispora Entrophospora Funneliformis Gigaspora
Glomus Rhizophagus Sclerocystis and Septoglomus) within six families (Acaulospo-
raceae Ambisporaceae Claroideoglomus Diversisporaceae Gigasporaceae and Glomer-
aceae)
Figure 1 Venn diagram showing the unique and shared arbuscular mycorrhizal fungi species underdifferent nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1HN 200 kg N haminus1 yrminus1
In total 26 AMF species of spore were detected in four nitrogen treatments (Figure 2Table 2 Figure A1) These AMF belonged to 12 genera (Acaulospora ArchaeosporaClaroideoglomus Dentiscutata Diversispora Entrophospora Funneliformis GigasporaGlomus Rhizophagus Sclerocystis and Septoglomus) within six families (AcaulosporaceaeAmbisporaceae Claroideoglomus Diversisporaceae Gigasporaceae and Glomeraceae)
Forests 2022 13 x FOR PEER REVIEW 5 of 16
(a) (b)
Figure 2 The frequency of occurrences (a) and relative abundance (b) among arbuscular mycor-
rhizal fungi under different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN
100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Table 2 Occurrence frequency relative abundance and important values of arbuscular mycorrhizal
fungi species in Chinese fir under nitrogen treatment
Species
Number AMF
Nitrogen
Treatment FO () RA () IV ()
CK LN MN HN
1 Acaulospora bierticulata + + + + 8125 1179 4652
2 Acaulospora denticulata minus + + minus 1250 159 705
3 Acaulospora excavat + + + + 9375 8163 8769
4 Acaulospora foveata + + + + 2500 499 1500
5 Archaeospora schenckii + minus minus minus 625 091 358
6 Claroideoglomus claroideum + + minus minus 1250 1500 1375
7 Claroideoglomus ethunicatum + + + + 4375 8500 6438
8 Dentiscutata heterogama minus minus + + 1250 10000 5625
9 Diversispora etunicatum minus + + + 2500 10000 6250
10 Entrophospora infrequens + minus minus minus 625 10000 5313
11 Funneliformis mosseae + + + + 3750 10000 6875
12 Funneliformis geosporum + + + + 3125 5441 4283
13 Gigaspora albida minus + + + 2500 4559 3530
14 Glomus aggregatum + minus + minus 625 10000 5313
15 Glomus ambisporum + minus minus minus 625 221 423
16 Glomus clarum + + + + 9375 012 4694
17 Glomus melanosporum + + + + 6875 639 3757
18 Glomus microaggregatum + + minus + 1875 314 1095
19 Glomus multicaule + + + + 6250 372 3311
20 Glomus multiforum + + + + 9375 2753 6064
21 Glomus reticulatum + + + + 10000 3461 6731
22 Rhizophagus aggregatus + + + + 10000 9739 9870
23 Rhizophagus intraradices minus + minus minus 625 261 443
24 Sclerocystis liquidambaris + + + + 3750 4098 3924
25 Sclerocystis sinuosum + + + + 8750 5902 7326
26 Septoglomus viscosum + + + + 10000 10000 10000
FO RA and IV are the frequency of occurrences relative abundance and importance value respec-
tively CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Figure 2 The frequency of occurrences (a) and relative abundance (b) among arbuscular mycor-rhizal fungi under different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Forests 2022 13 979 5 of 14
Table 2 Occurrence frequency relative abundance and important values of arbuscular mycorrhizalfungi species in Chinese fir under nitrogen treatment
Species Number AMFNitrogen Treatment
FO () RA () IV ()CK LN MN HN
1 Acaulospora bierticulata + + + + 8125 1179 46522 Acaulospora denticulata minus + + minus 1250 159 7053 Acaulospora excavat + + + + 9375 8163 87694 Acaulospora foveata + + + + 2500 499 15005 Archaeospora schenckii + minus minus minus 625 091 3586 Claroideoglomus claroideum + + minus minus 1250 1500 13757 Claroideoglomus ethunicatum + + + + 4375 8500 64388 Dentiscutata heterogama minus minus + + 1250 10000 56259 Diversispora etunicatum minus + + + 2500 10000 625010 Entrophospora infrequens + minus minus minus 625 10000 531311 Funneliformis mosseae + + + + 3750 10000 687512 Funneliformis geosporum + + + + 3125 5441 428313 Gigaspora albida minus + + + 2500 4559 353014 Glomus aggregatum + minus + minus 625 10000 531315 Glomus ambisporum + minus minus minus 625 221 42316 Glomus clarum + + + + 9375 012 469417 Glomus melanosporum + + + + 6875 639 375718 Glomus microaggregatum + + minus + 1875 314 109519 Glomus multicaule + + + + 6250 372 331120 Glomus multiforum + + + + 9375 2753 606421 Glomus reticulatum + + + + 10000 3461 673122 Rhizophagus aggregatus + + + + 10000 9739 987023 Rhizophagus intraradices minus + minus minus 625 261 44324 Sclerocystis liquidambaris + + + + 3750 4098 392425 Sclerocystis sinuosum + + + + 8750 5902 732626 Septoglomus viscosum + + + + 10000 10000 10000
FO RA and IV are the frequency of occurrences relative abundance and importance value respectively CK nofertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris andSeptoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum(100) gt Rhizophagus aggregatus (9870) gt Acaulospora excavat (8769) gt Sclerocystis sinuo-sum (7326) gt Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followedby Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomusetunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglomusviscosum had the highest IV followed by Acaulospora excavat (9042) Rhizophagus ag-gregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum (6956)(Figure 3)
Forests 2022 13 979 6 of 14
Forests 2022 13 x FOR PEER REVIEW 6 of 16
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-
mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris and
Septoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum
(100) Rhizophagus aggregatus (9870) Acaulospora excavat (8769) Sclerocystis sin-
uosum (7326) Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)
had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus
(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-
cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)
Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-
tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followed
by Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomus
etunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglo-
mus viscosum had the highest IV followed by Acaulospora excavat (9042) Rhizopha-
gus aggregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum
(6956) (Figure 3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitrogen treat-
ments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soil
pH However N addition increased soil N under LN treatment (+1505) Under LN treat-
ment soil AP and TP were the highest Soil NP increased slightly with N addition (Table
3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitro-gen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soilpH However N addition increased soil N under LN treatment (+1505) Under LNtreatment soil AP and TP were the highest Soil NP increased slightly with N addition(Table 3)
Table 3 Soil properties under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
pH 400 plusmn 008 402 plusmn 018 383 plusmn 008 393 plusmn 008 057 064TOC (gkg) 139 plusmn 009 124 plusmn 026 136 plusmn 014 150 plusmn 011 135 030
NH4+-N (mgkg) 2807 plusmn 104 2948 plusmn 192 3248 plusmn 291 3357 plusmn 305 116 037
AN (mgkg) 6143 plusmn 054 6090 plusmn 260 6452 plusmn 384 6465 plusmn 276 054 067AP (mgkg) 782 plusmn 096 898 plusmn 052 803 plusmn 073 805 plusmn 059 051 069
C (gkg) 8978 plusmn 406 9875 plusmn035 9310 plusmn 525 9168 plusmn 320 070 058N (gkg) 578 plusmn 025 665 plusmn 028 600 plusmn 028 565 plusmn 013 337 006
P (mgkg) 23464 plusmn 2257 23981 plusmn 2102 22852 plusmn 1505 22315 plusmn 2556 012 095CN 1554 plusmn 005 1567 plusmn 059 1655 plusmn 068 1622 plusmn 033 103 042NP 2531 plusmn 260 2562 plusmn 278 2673 plusmn 250 2681 plusmn 170 010 096
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus
C total carbon N total nitrogen P total phosphorus CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1 One-way ANOVA and LSD were used to analyze the differencesError bars indicate plusmn SE of mean (n = 4)
34 Relationship between Soil Properties and AMF
AMF colonization was negatively correlated with soil TOC (p lt 005) (Table 4) externalHLD was negatively connected with soil P (p lt 005) spore density was positively relatedto soil C (p lt 005) and species richness was in a positive correlation with soil TOC and C(p lt 005) The Shannon indices presented a positive correlation with soil pH (p lt 005)
Forests 2022 13 979 7 of 14
Table 4 Spearman correlation coefficients between soil properties and parameters of arbuscularmycorrhizal fungi
pH TOC NH4+-N AN AP C N P CN NP
Colonization 0012 minus0643 minus0437 minus0571 minus0245 minus014 minus0126 0142 0025 minus0112HLD minus0062 0267 0245 0288 minus0531 minus026 minus0197 minus0701 minus0154 0564SD minus0338 0214 0008 0251 0161 0679 0477 0283 0489 0002SR 0101 0604 0488 0551 0114 063 0478 0039 033 0117H 0623 0478 0341 0142 0081 0306 0328 minus0023 minus0098 0117D 0560 0408 0264 0105 012 0193 0209 0002 minus0066 007
Acaulospora minus0378 minus0575 minus0506 minus052 0004 minus0332 minus0418 0575 0332 minus0708 Archaeospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025
Claroideoglomus 0464 minus0303 0302 minus0029 minus0047 0077 0175 0071 minus0268 minus0045Dentiscutata 0014 0244 0434 0451 017 0453 0349 0165 0203 minus0047Diversispora 0057 0482 minus0205 minus0057 minus0051 minus0173 minus0304 0384 0399 minus0447
Entrophospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025Funneliformis 005 0764 0175 0266 minus0045 minus0163 minus0201 minus0093 0113 minus0033
Gigaspora minus0644 minus0385 minus0672 minus0466 0042 minus024 minus0419 0636 0581 minus0711 Glomus 0341 minus0133 0242 0033 minus0086 0199 0279 minus0363 minus0258 0393
Rhizophagus 0275 minus0152 minus0376 minus0438 minus0378 0023 0081 minus0056 minus0124 0208Septogolomus minus0138 0252 023 0404 021 0179 0193 minus0194 minus0104 0288
Ancestral minus0367 minus0508 minus0508 minus0524 minus0036 minus0425 minus0513 0575 0347 minus0757 Edaphophilic minus0083 0381 minus0341 minus0154 minus004 minus0218 minus0381 0505 0509 minus0582 Rhizophilic 0361 0431 053 052 004 0435 0541 minus0617 minus04 0801
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C
total carbon N total nitrogen P total phosphorus HLD external hyphal length density SD spore density SRspecies richness H ShannonndashWiener index D Simpson index p lt 005 p lt 001
Archaeospora was in a negative correlation with soil NP (p lt 005) Funneliformis waspositively correlated with soil TOC (p lt 001) and Gigaspora was negatively connectedwith soil pH NH4
+-N (p lt 005) and soil NP (p lt 001) but positively related to soil Pand CN (p lt 005) Ancestral AMF was in a negative correlation with soil NP (p lt 001)edaphophilic AMF was negatively correlated with soil NP (p lt 005) and rhizophilic AMFwas positively connected with soil NP (p lt 001) but negatively related to soil P (p lt 005)
RDA was conducted to evaluate the relationship between the soil chemical properties and theAMF community (Figure 4) RDA1 accounted for 6699 of the variations while RDA2 accountedfor 2189 Of the evaluated attributes the NP had significant conditional effects (p lt 001) whilethe soil TOC (p = 0076) and AN (p = 0092) caused referential effects to the AMF communities
Forests 2022 13 x FOR PEER REVIEW 8 of 16
density SD spore density SR species richness H ShannonndashWiener index D Simpson index p lt
005 p lt 001
Archaeospora was in a negative correlation with soil NP (p 005) Funneliformis was
positively correlated with soil TOC (p 001) and Gigaspora was negatively connected
with soil pH NH4+-N (p 005) and soil NP (p 001) but positively related to soil P and
CN (p 005) Ancestral AMF was in a negative correlation with soil NP (p 001)
edaphophilic AMF was negatively correlated with soil NP (p 005) and rhizophilic AMF
was positively connected with soil NP (p 001) but negatively related to soil P (p 005)
RDA was conducted to evaluate the relationship between the soil chemical properties
and the AMF community (Figure 4) RDA1 accounted for 6699 of the variations while
RDA2 accounted for 2189 Of the evaluated attributes the NP had significant condi-
tional effects (p 001) while the soil TOC (p = 0076) and AN (p = 0092) caused referential
effects to the AMF communities
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular
mycorrhizal fungi communities under different nitrogen treatments TOC total organic carbon
NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C total carbon
N total nitrogen P total phosphorus The red arrows represent soil properties the blue arrows
represent arbuscular mycorrhizal fungi communities
4 Discussion
N addition can make a difference on AMF diversity and community composition
because with the increase of soil nutrients plants would have a strategy to acquire nutri-
ents by fine roots or AMF [2644] Our results showed that NP is a factor that has a sig-
nificant effect on AMF communities (Figure 4) AMF community structure might have a
connection with the dynamics of soil conditions and C-to-P trade between AMF and
plants It has been agreed that AMF exchanges P derived from soil for plant carbon [1744ndash
47] In our study N addition had insignificant influence on AMF colonization external
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular mycorrhizalfungi communities under different nitrogen treatments TOC total organic carbon NH4
+-N ammoniumnitrogen AN available nitrogen AP available phosphorus C total carbon N total nitrogen P total phosphorusThe red arrows represent soil properties the blue arrows represent arbuscular mycorrhizal fungi communities
Forests 2022 13 979 8 of 14
4 Discussion
N addition can make a difference on AMF diversity and community compositionbecause with the increase of soil nutrients plants would have a strategy to acquire nutrientsby fine roots or AMF [2644] Our results showed that NP is a factor that has a significanteffect on AMF communities (Figure 4) AMF community structure might have a connectionwith the dynamics of soil conditions and C-to-P trade between AMF and plants It has beenagreed that AMF exchanges P derived from soil for plant carbon [1744ndash47] In our studyN addition had insignificant influence on AMF colonization external HLD spore densityor diversity This finding may be explained as the symbiotic relationship between AMFand root system being stable [48]
Generally there is little specificity between AMF and the host plant and severalAMF species could colonize one AM plant at the same time [4950] However differentpatterns of colonization with N addition suggest a preference for specific AMF species [51]In our study we observed that Glomeraceae was the most predominant family amongall treatments Glomus Septoglomus and Acaulospora were the predominant genera andSeptoglomus viscosum and Acaulospora foveata were the predominant species Our resultsslightly disagree with previous studies which presented that Glomus and Acaulospora asthe predominant species due to their smaller size of spore which helped them producemore spores in less time [485253] This discrepancy can be answered by the diversespecies and age of the host plants The high isolation frequencies of Septoglomus viscosumindicated that it has a strong ability to adapt to the environment (N deposition) Septoglomusviscosum can improve plant growth and production [54] which is mainly related to rootand aboveground biomass [55]
Archaeospora schenckii Entrophospora infrequens and Glomus ambisporum disappearedwith N treatments (Figure 3) which was in keeping with Eom [19] who also found thatEntrophospora infrequens decreased with N addition In P-rich (low NP) soils nitrogenapplication reduces underground carbon allocation and rare species decrease as N in-creases A previous study showed that the influence of N addition on AMF communityappeared different with added N promoting Glomeraceae in A Sieversiana while lesseningthem in L chinensis [16] Some Glomus species were sensitive to N application whichindicated that certain species of AMF play an indispensable role in host plant response toenvironmental change
As identified in most surveys conducted in other fields [56ndash58] we also discoveredFunneliformis mosseae presenting in our study which was reported to promote plant nutri-ents [59] GmosAAP1 an amino acid permease has been characterized from the extraradicalstructure of Funneliformis mosseae which indicates a potential for amino acid uptake [60]Our study showed that with N addition its importance decreased This decrease maybe explained by a decrease in the dependency on Funneliformis mosseae for N nutrientacquisition by the host plant
Above all our results showed that AMF participating in and manage the soil nutrientcycle could be sensitive to small changes in edatopes Therefore studying mycorrhizalsymbionts increases the knowledge of climatic variation [61ndash63]
AMF were grouped into three guilds namely edaphophilic rhizophilc and ancestralIn our study the percentage followed the order rhizophilic gt edaphophilic gt ancestral(Figure 5c) N addition altered AMF species but did not change functional groups Despitethe addition of nitrogen trees may reduce dependence on AMF Meanwhile a large groupof rhizophilic guilds function to reduce root pathogen infection and primarily benefit plantswith fine roots prone to pathogen infections
Forests 2022 13 979 9 of 14Forests 2022 13 x FOR PEER REVIEW 10 of 16
Figure 5 Cont
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-
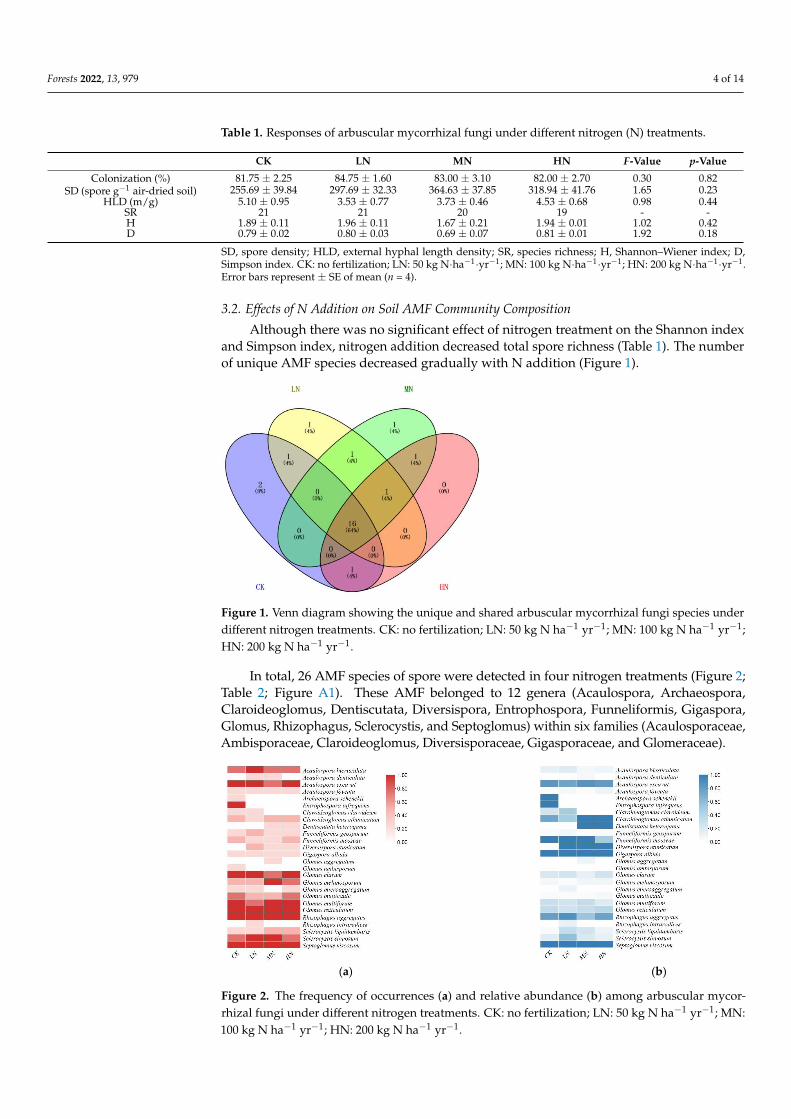
Forests 2022 13 979 4 of 14
Table 1 Responses of arbuscular mycorrhizal fungi under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
Colonization () 8175 plusmn 225 8475 plusmn 160 8300 plusmn 310 8200 plusmn 270 030 082SD (spore gminus1 air-dried soil) 25569 plusmn 3984 29769 plusmn 3233 36463 plusmn 3785 31894 plusmn 4176 165 023
HLD (mg) 510 plusmn 095 353 plusmn 077 373 plusmn 046 453 plusmn 068 098 044SR 21 21 20 19 - -H 189 plusmn 011 196 plusmn 011 167 plusmn 021 194 plusmn 001 102 042D 079 plusmn 002 080 plusmn 003 069 plusmn 007 081 plusmn 001 192 018
SD spore density HLD external hyphal length density SR species richness H ShannonndashWiener index DSimpson index CK no fertilization LN 50 kg Nmiddothaminus1middotyrminus1 MN 100 kg Nmiddothaminus1middotyrminus1 HN 200 kg Nmiddothaminus1middotyrminus1Error bars represent plusmn SE of mean (n = 4)
32 Effects of N Addition on Soil AMF Community Composition
Although there was no significant effect of nitrogen treatment on the Shannon indexand Simpson index nitrogen addition decreased total spore richness (Table 1) The numberof unique AMF species decreased gradually with N addition (Figure 1)
Forests 2022 13 x FOR PEER REVIEW 4 of 16
Table 1 Responses of arbuscular mycorrhizal fungi under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
Coloniza-
tion () 8175 plusmn 225 8475 plusmn 160 8300 plusmn 310 8200 plusmn 270 030 082
SD (spore
gminus1 air-dried
soil)
25569 plusmn
3984
29769 plusmn
3233
36463 plusmn
3785
31894 plusmn
4176 165 023
HLD (mg) 510 plusmn 095 353 plusmn 077 373 plusmn 046 453 plusmn 068 098 044
SR 21 21 20 19 - -
H 189 plusmn 011 196 plusmn 011 167 plusmn 021 194 plusmn 001 102 042
D 079 plusmn 002 080 plusmn 003 069 plusmn 007 081 plusmn 001 192 018
SD spore density HLD external hyphal length density SR species richness H ShannonndashWiener
index D Simpson index CK no fertilization LN 50 kg Nmiddothaminus1middotyrminus1 MN 100kg Nmiddothaminus1middotyrminus1 HN
200kg Nmiddothaminus1middotyrminus1 Error bars represent plusmn SE of mean (n = 4)
32 Effects of N Addition on Soil AMF Community Composition
Although there was no significant effect of nitrogen treatment on the Shannon index
and Simpson index nitrogen addition decreased total spore richness (Table 1) The num-
ber of unique AMF species decreased gradually with N addition (Figure 1)
Figure 1 Venn diagram showing the unique and shared arbuscular mycorrhizal fungi species under
different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN
200kg N haminus1 yrminus1
In total 26 AMF species of spore were detected in four nitrogen treatments (Figure
2 Table 2 Figure A1) These AMF belonged to 12 genera (Acaulospora Archaeospora
Claroideoglomus Dentiscutata Diversispora Entrophospora Funneliformis Gigaspora
Glomus Rhizophagus Sclerocystis and Septoglomus) within six families (Acaulospo-
raceae Ambisporaceae Claroideoglomus Diversisporaceae Gigasporaceae and Glomer-
aceae)
Figure 1 Venn diagram showing the unique and shared arbuscular mycorrhizal fungi species underdifferent nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1HN 200 kg N haminus1 yrminus1
In total 26 AMF species of spore were detected in four nitrogen treatments (Figure 2Table 2 Figure A1) These AMF belonged to 12 genera (Acaulospora ArchaeosporaClaroideoglomus Dentiscutata Diversispora Entrophospora Funneliformis GigasporaGlomus Rhizophagus Sclerocystis and Septoglomus) within six families (AcaulosporaceaeAmbisporaceae Claroideoglomus Diversisporaceae Gigasporaceae and Glomeraceae)
Forests 2022 13 x FOR PEER REVIEW 5 of 16
(a) (b)
Figure 2 The frequency of occurrences (a) and relative abundance (b) among arbuscular mycor-
rhizal fungi under different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN
100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Table 2 Occurrence frequency relative abundance and important values of arbuscular mycorrhizal
fungi species in Chinese fir under nitrogen treatment
Species
Number AMF
Nitrogen
Treatment FO () RA () IV ()
CK LN MN HN
1 Acaulospora bierticulata + + + + 8125 1179 4652
2 Acaulospora denticulata minus + + minus 1250 159 705
3 Acaulospora excavat + + + + 9375 8163 8769
4 Acaulospora foveata + + + + 2500 499 1500
5 Archaeospora schenckii + minus minus minus 625 091 358
6 Claroideoglomus claroideum + + minus minus 1250 1500 1375
7 Claroideoglomus ethunicatum + + + + 4375 8500 6438
8 Dentiscutata heterogama minus minus + + 1250 10000 5625
9 Diversispora etunicatum minus + + + 2500 10000 6250
10 Entrophospora infrequens + minus minus minus 625 10000 5313
11 Funneliformis mosseae + + + + 3750 10000 6875
12 Funneliformis geosporum + + + + 3125 5441 4283
13 Gigaspora albida minus + + + 2500 4559 3530
14 Glomus aggregatum + minus + minus 625 10000 5313
15 Glomus ambisporum + minus minus minus 625 221 423
16 Glomus clarum + + + + 9375 012 4694
17 Glomus melanosporum + + + + 6875 639 3757
18 Glomus microaggregatum + + minus + 1875 314 1095
19 Glomus multicaule + + + + 6250 372 3311
20 Glomus multiforum + + + + 9375 2753 6064
21 Glomus reticulatum + + + + 10000 3461 6731
22 Rhizophagus aggregatus + + + + 10000 9739 9870
23 Rhizophagus intraradices minus + minus minus 625 261 443
24 Sclerocystis liquidambaris + + + + 3750 4098 3924
25 Sclerocystis sinuosum + + + + 8750 5902 7326
26 Septoglomus viscosum + + + + 10000 10000 10000
FO RA and IV are the frequency of occurrences relative abundance and importance value respec-
tively CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Figure 2 The frequency of occurrences (a) and relative abundance (b) among arbuscular mycor-rhizal fungi under different nitrogen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Forests 2022 13 979 5 of 14
Table 2 Occurrence frequency relative abundance and important values of arbuscular mycorrhizalfungi species in Chinese fir under nitrogen treatment
Species Number AMFNitrogen Treatment
FO () RA () IV ()CK LN MN HN
1 Acaulospora bierticulata + + + + 8125 1179 46522 Acaulospora denticulata minus + + minus 1250 159 7053 Acaulospora excavat + + + + 9375 8163 87694 Acaulospora foveata + + + + 2500 499 15005 Archaeospora schenckii + minus minus minus 625 091 3586 Claroideoglomus claroideum + + minus minus 1250 1500 13757 Claroideoglomus ethunicatum + + + + 4375 8500 64388 Dentiscutata heterogama minus minus + + 1250 10000 56259 Diversispora etunicatum minus + + + 2500 10000 625010 Entrophospora infrequens + minus minus minus 625 10000 531311 Funneliformis mosseae + + + + 3750 10000 687512 Funneliformis geosporum + + + + 3125 5441 428313 Gigaspora albida minus + + + 2500 4559 353014 Glomus aggregatum + minus + minus 625 10000 531315 Glomus ambisporum + minus minus minus 625 221 42316 Glomus clarum + + + + 9375 012 469417 Glomus melanosporum + + + + 6875 639 375718 Glomus microaggregatum + + minus + 1875 314 109519 Glomus multicaule + + + + 6250 372 331120 Glomus multiforum + + + + 9375 2753 606421 Glomus reticulatum + + + + 10000 3461 673122 Rhizophagus aggregatus + + + + 10000 9739 987023 Rhizophagus intraradices minus + minus minus 625 261 44324 Sclerocystis liquidambaris + + + + 3750 4098 392425 Sclerocystis sinuosum + + + + 8750 5902 732626 Septoglomus viscosum + + + + 10000 10000 10000
FO RA and IV are the frequency of occurrences relative abundance and importance value respectively CK nofertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris andSeptoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum(100) gt Rhizophagus aggregatus (9870) gt Acaulospora excavat (8769) gt Sclerocystis sinuo-sum (7326) gt Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followedby Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomusetunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglomusviscosum had the highest IV followed by Acaulospora excavat (9042) Rhizophagus ag-gregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum (6956)(Figure 3)
Forests 2022 13 979 6 of 14
Forests 2022 13 x FOR PEER REVIEW 6 of 16
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-
mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris and
Septoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum
(100) Rhizophagus aggregatus (9870) Acaulospora excavat (8769) Sclerocystis sin-
uosum (7326) Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)
had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus
(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-
cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)
Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-
tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followed
by Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomus
etunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglo-
mus viscosum had the highest IV followed by Acaulospora excavat (9042) Rhizopha-
gus aggregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum
(6956) (Figure 3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitrogen treat-
ments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soil
pH However N addition increased soil N under LN treatment (+1505) Under LN treat-
ment soil AP and TP were the highest Soil NP increased slightly with N addition (Table
3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitro-gen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soilpH However N addition increased soil N under LN treatment (+1505) Under LNtreatment soil AP and TP were the highest Soil NP increased slightly with N addition(Table 3)
Table 3 Soil properties under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
pH 400 plusmn 008 402 plusmn 018 383 plusmn 008 393 plusmn 008 057 064TOC (gkg) 139 plusmn 009 124 plusmn 026 136 plusmn 014 150 plusmn 011 135 030
NH4+-N (mgkg) 2807 plusmn 104 2948 plusmn 192 3248 plusmn 291 3357 plusmn 305 116 037
AN (mgkg) 6143 plusmn 054 6090 plusmn 260 6452 plusmn 384 6465 plusmn 276 054 067AP (mgkg) 782 plusmn 096 898 plusmn 052 803 plusmn 073 805 plusmn 059 051 069
C (gkg) 8978 plusmn 406 9875 plusmn035 9310 plusmn 525 9168 plusmn 320 070 058N (gkg) 578 plusmn 025 665 plusmn 028 600 plusmn 028 565 plusmn 013 337 006
P (mgkg) 23464 plusmn 2257 23981 plusmn 2102 22852 plusmn 1505 22315 plusmn 2556 012 095CN 1554 plusmn 005 1567 plusmn 059 1655 plusmn 068 1622 plusmn 033 103 042NP 2531 plusmn 260 2562 plusmn 278 2673 plusmn 250 2681 plusmn 170 010 096
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus
C total carbon N total nitrogen P total phosphorus CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1 One-way ANOVA and LSD were used to analyze the differencesError bars indicate plusmn SE of mean (n = 4)
34 Relationship between Soil Properties and AMF
AMF colonization was negatively correlated with soil TOC (p lt 005) (Table 4) externalHLD was negatively connected with soil P (p lt 005) spore density was positively relatedto soil C (p lt 005) and species richness was in a positive correlation with soil TOC and C(p lt 005) The Shannon indices presented a positive correlation with soil pH (p lt 005)
Forests 2022 13 979 7 of 14
Table 4 Spearman correlation coefficients between soil properties and parameters of arbuscularmycorrhizal fungi
pH TOC NH4+-N AN AP C N P CN NP
Colonization 0012 minus0643 minus0437 minus0571 minus0245 minus014 minus0126 0142 0025 minus0112HLD minus0062 0267 0245 0288 minus0531 minus026 minus0197 minus0701 minus0154 0564SD minus0338 0214 0008 0251 0161 0679 0477 0283 0489 0002SR 0101 0604 0488 0551 0114 063 0478 0039 033 0117H 0623 0478 0341 0142 0081 0306 0328 minus0023 minus0098 0117D 0560 0408 0264 0105 012 0193 0209 0002 minus0066 007
Acaulospora minus0378 minus0575 minus0506 minus052 0004 minus0332 minus0418 0575 0332 minus0708 Archaeospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025
Claroideoglomus 0464 minus0303 0302 minus0029 minus0047 0077 0175 0071 minus0268 minus0045Dentiscutata 0014 0244 0434 0451 017 0453 0349 0165 0203 minus0047Diversispora 0057 0482 minus0205 minus0057 minus0051 minus0173 minus0304 0384 0399 minus0447
Entrophospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025Funneliformis 005 0764 0175 0266 minus0045 minus0163 minus0201 minus0093 0113 minus0033
Gigaspora minus0644 minus0385 minus0672 minus0466 0042 minus024 minus0419 0636 0581 minus0711 Glomus 0341 minus0133 0242 0033 minus0086 0199 0279 minus0363 minus0258 0393
Rhizophagus 0275 minus0152 minus0376 minus0438 minus0378 0023 0081 minus0056 minus0124 0208Septogolomus minus0138 0252 023 0404 021 0179 0193 minus0194 minus0104 0288
Ancestral minus0367 minus0508 minus0508 minus0524 minus0036 minus0425 minus0513 0575 0347 minus0757 Edaphophilic minus0083 0381 minus0341 minus0154 minus004 minus0218 minus0381 0505 0509 minus0582 Rhizophilic 0361 0431 053 052 004 0435 0541 minus0617 minus04 0801
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C
total carbon N total nitrogen P total phosphorus HLD external hyphal length density SD spore density SRspecies richness H ShannonndashWiener index D Simpson index p lt 005 p lt 001
Archaeospora was in a negative correlation with soil NP (p lt 005) Funneliformis waspositively correlated with soil TOC (p lt 001) and Gigaspora was negatively connectedwith soil pH NH4
+-N (p lt 005) and soil NP (p lt 001) but positively related to soil Pand CN (p lt 005) Ancestral AMF was in a negative correlation with soil NP (p lt 001)edaphophilic AMF was negatively correlated with soil NP (p lt 005) and rhizophilic AMFwas positively connected with soil NP (p lt 001) but negatively related to soil P (p lt 005)
RDA was conducted to evaluate the relationship between the soil chemical properties and theAMF community (Figure 4) RDA1 accounted for 6699 of the variations while RDA2 accountedfor 2189 Of the evaluated attributes the NP had significant conditional effects (p lt 001) whilethe soil TOC (p = 0076) and AN (p = 0092) caused referential effects to the AMF communities
Forests 2022 13 x FOR PEER REVIEW 8 of 16
density SD spore density SR species richness H ShannonndashWiener index D Simpson index p lt
005 p lt 001
Archaeospora was in a negative correlation with soil NP (p 005) Funneliformis was
positively correlated with soil TOC (p 001) and Gigaspora was negatively connected
with soil pH NH4+-N (p 005) and soil NP (p 001) but positively related to soil P and
CN (p 005) Ancestral AMF was in a negative correlation with soil NP (p 001)
edaphophilic AMF was negatively correlated with soil NP (p 005) and rhizophilic AMF
was positively connected with soil NP (p 001) but negatively related to soil P (p 005)
RDA was conducted to evaluate the relationship between the soil chemical properties
and the AMF community (Figure 4) RDA1 accounted for 6699 of the variations while
RDA2 accounted for 2189 Of the evaluated attributes the NP had significant condi-
tional effects (p 001) while the soil TOC (p = 0076) and AN (p = 0092) caused referential
effects to the AMF communities
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular
mycorrhizal fungi communities under different nitrogen treatments TOC total organic carbon
NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C total carbon
N total nitrogen P total phosphorus The red arrows represent soil properties the blue arrows
represent arbuscular mycorrhizal fungi communities
4 Discussion
N addition can make a difference on AMF diversity and community composition
because with the increase of soil nutrients plants would have a strategy to acquire nutri-
ents by fine roots or AMF [2644] Our results showed that NP is a factor that has a sig-
nificant effect on AMF communities (Figure 4) AMF community structure might have a
connection with the dynamics of soil conditions and C-to-P trade between AMF and
plants It has been agreed that AMF exchanges P derived from soil for plant carbon [1744ndash
47] In our study N addition had insignificant influence on AMF colonization external
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular mycorrhizalfungi communities under different nitrogen treatments TOC total organic carbon NH4
+-N ammoniumnitrogen AN available nitrogen AP available phosphorus C total carbon N total nitrogen P total phosphorusThe red arrows represent soil properties the blue arrows represent arbuscular mycorrhizal fungi communities
Forests 2022 13 979 8 of 14
4 Discussion
N addition can make a difference on AMF diversity and community compositionbecause with the increase of soil nutrients plants would have a strategy to acquire nutrientsby fine roots or AMF [2644] Our results showed that NP is a factor that has a significanteffect on AMF communities (Figure 4) AMF community structure might have a connectionwith the dynamics of soil conditions and C-to-P trade between AMF and plants It has beenagreed that AMF exchanges P derived from soil for plant carbon [1744ndash47] In our studyN addition had insignificant influence on AMF colonization external HLD spore densityor diversity This finding may be explained as the symbiotic relationship between AMFand root system being stable [48]
Generally there is little specificity between AMF and the host plant and severalAMF species could colonize one AM plant at the same time [4950] However differentpatterns of colonization with N addition suggest a preference for specific AMF species [51]In our study we observed that Glomeraceae was the most predominant family amongall treatments Glomus Septoglomus and Acaulospora were the predominant genera andSeptoglomus viscosum and Acaulospora foveata were the predominant species Our resultsslightly disagree with previous studies which presented that Glomus and Acaulospora asthe predominant species due to their smaller size of spore which helped them producemore spores in less time [485253] This discrepancy can be answered by the diversespecies and age of the host plants The high isolation frequencies of Septoglomus viscosumindicated that it has a strong ability to adapt to the environment (N deposition) Septoglomusviscosum can improve plant growth and production [54] which is mainly related to rootand aboveground biomass [55]
Archaeospora schenckii Entrophospora infrequens and Glomus ambisporum disappearedwith N treatments (Figure 3) which was in keeping with Eom [19] who also found thatEntrophospora infrequens decreased with N addition In P-rich (low NP) soils nitrogenapplication reduces underground carbon allocation and rare species decrease as N in-creases A previous study showed that the influence of N addition on AMF communityappeared different with added N promoting Glomeraceae in A Sieversiana while lesseningthem in L chinensis [16] Some Glomus species were sensitive to N application whichindicated that certain species of AMF play an indispensable role in host plant response toenvironmental change
As identified in most surveys conducted in other fields [56ndash58] we also discoveredFunneliformis mosseae presenting in our study which was reported to promote plant nutri-ents [59] GmosAAP1 an amino acid permease has been characterized from the extraradicalstructure of Funneliformis mosseae which indicates a potential for amino acid uptake [60]Our study showed that with N addition its importance decreased This decrease maybe explained by a decrease in the dependency on Funneliformis mosseae for N nutrientacquisition by the host plant
Above all our results showed that AMF participating in and manage the soil nutrientcycle could be sensitive to small changes in edatopes Therefore studying mycorrhizalsymbionts increases the knowledge of climatic variation [61ndash63]
AMF were grouped into three guilds namely edaphophilic rhizophilc and ancestralIn our study the percentage followed the order rhizophilic gt edaphophilic gt ancestral(Figure 5c) N addition altered AMF species but did not change functional groups Despitethe addition of nitrogen trees may reduce dependence on AMF Meanwhile a large groupof rhizophilic guilds function to reduce root pathogen infection and primarily benefit plantswith fine roots prone to pathogen infections
Forests 2022 13 979 9 of 14Forests 2022 13 x FOR PEER REVIEW 10 of 16
Figure 5 Cont
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-

Forests 2022 13 979 5 of 14
Table 2 Occurrence frequency relative abundance and important values of arbuscular mycorrhizalfungi species in Chinese fir under nitrogen treatment
Species Number AMFNitrogen Treatment
FO () RA () IV ()CK LN MN HN
1 Acaulospora bierticulata + + + + 8125 1179 46522 Acaulospora denticulata minus + + minus 1250 159 7053 Acaulospora excavat + + + + 9375 8163 87694 Acaulospora foveata + + + + 2500 499 15005 Archaeospora schenckii + minus minus minus 625 091 3586 Claroideoglomus claroideum + + minus minus 1250 1500 13757 Claroideoglomus ethunicatum + + + + 4375 8500 64388 Dentiscutata heterogama minus minus + + 1250 10000 56259 Diversispora etunicatum minus + + + 2500 10000 625010 Entrophospora infrequens + minus minus minus 625 10000 531311 Funneliformis mosseae + + + + 3750 10000 687512 Funneliformis geosporum + + + + 3125 5441 428313 Gigaspora albida minus + + + 2500 4559 353014 Glomus aggregatum + minus + minus 625 10000 531315 Glomus ambisporum + minus minus minus 625 221 42316 Glomus clarum + + + + 9375 012 469417 Glomus melanosporum + + + + 6875 639 375718 Glomus microaggregatum + + minus + 1875 314 109519 Glomus multicaule + + + + 6250 372 331120 Glomus multiforum + + + + 9375 2753 606421 Glomus reticulatum + + + + 10000 3461 673122 Rhizophagus aggregatus + + + + 10000 9739 987023 Rhizophagus intraradices minus + minus minus 625 261 44324 Sclerocystis liquidambaris + + + + 3750 4098 392425 Sclerocystis sinuosum + + + + 8750 5902 732626 Septoglomus viscosum + + + + 10000 10000 10000
FO RA and IV are the frequency of occurrences relative abundance and importance value respectively CK nofertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris andSeptoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum(100) gt Rhizophagus aggregatus (9870) gt Acaulospora excavat (8769) gt Sclerocystis sinuo-sum (7326) gt Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followedby Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomusetunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglomusviscosum had the highest IV followed by Acaulospora excavat (9042) Rhizophagus ag-gregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum (6956)(Figure 3)
Forests 2022 13 979 6 of 14
Forests 2022 13 x FOR PEER REVIEW 6 of 16
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-
mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris and
Septoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum
(100) Rhizophagus aggregatus (9870) Acaulospora excavat (8769) Sclerocystis sin-
uosum (7326) Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)
had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus
(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-
cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)
Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-
tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followed
by Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomus
etunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglo-
mus viscosum had the highest IV followed by Acaulospora excavat (9042) Rhizopha-
gus aggregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum
(6956) (Figure 3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitrogen treat-
ments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soil
pH However N addition increased soil N under LN treatment (+1505) Under LN treat-
ment soil AP and TP were the highest Soil NP increased slightly with N addition (Table
3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitro-gen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soilpH However N addition increased soil N under LN treatment (+1505) Under LNtreatment soil AP and TP were the highest Soil NP increased slightly with N addition(Table 3)
Table 3 Soil properties under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
pH 400 plusmn 008 402 plusmn 018 383 plusmn 008 393 plusmn 008 057 064TOC (gkg) 139 plusmn 009 124 plusmn 026 136 plusmn 014 150 plusmn 011 135 030
NH4+-N (mgkg) 2807 plusmn 104 2948 plusmn 192 3248 plusmn 291 3357 plusmn 305 116 037
AN (mgkg) 6143 plusmn 054 6090 plusmn 260 6452 plusmn 384 6465 plusmn 276 054 067AP (mgkg) 782 plusmn 096 898 plusmn 052 803 plusmn 073 805 plusmn 059 051 069
C (gkg) 8978 plusmn 406 9875 plusmn035 9310 plusmn 525 9168 plusmn 320 070 058N (gkg) 578 plusmn 025 665 plusmn 028 600 plusmn 028 565 plusmn 013 337 006
P (mgkg) 23464 plusmn 2257 23981 plusmn 2102 22852 plusmn 1505 22315 plusmn 2556 012 095CN 1554 plusmn 005 1567 plusmn 059 1655 plusmn 068 1622 plusmn 033 103 042NP 2531 plusmn 260 2562 plusmn 278 2673 plusmn 250 2681 plusmn 170 010 096
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus
C total carbon N total nitrogen P total phosphorus CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1 One-way ANOVA and LSD were used to analyze the differencesError bars indicate plusmn SE of mean (n = 4)
34 Relationship between Soil Properties and AMF
AMF colonization was negatively correlated with soil TOC (p lt 005) (Table 4) externalHLD was negatively connected with soil P (p lt 005) spore density was positively relatedto soil C (p lt 005) and species richness was in a positive correlation with soil TOC and C(p lt 005) The Shannon indices presented a positive correlation with soil pH (p lt 005)
Forests 2022 13 979 7 of 14
Table 4 Spearman correlation coefficients between soil properties and parameters of arbuscularmycorrhizal fungi
pH TOC NH4+-N AN AP C N P CN NP
Colonization 0012 minus0643 minus0437 minus0571 minus0245 minus014 minus0126 0142 0025 minus0112HLD minus0062 0267 0245 0288 minus0531 minus026 minus0197 minus0701 minus0154 0564SD minus0338 0214 0008 0251 0161 0679 0477 0283 0489 0002SR 0101 0604 0488 0551 0114 063 0478 0039 033 0117H 0623 0478 0341 0142 0081 0306 0328 minus0023 minus0098 0117D 0560 0408 0264 0105 012 0193 0209 0002 minus0066 007
Acaulospora minus0378 minus0575 minus0506 minus052 0004 minus0332 minus0418 0575 0332 minus0708 Archaeospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025
Claroideoglomus 0464 minus0303 0302 minus0029 minus0047 0077 0175 0071 minus0268 minus0045Dentiscutata 0014 0244 0434 0451 017 0453 0349 0165 0203 minus0047Diversispora 0057 0482 minus0205 minus0057 minus0051 minus0173 minus0304 0384 0399 minus0447
Entrophospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025Funneliformis 005 0764 0175 0266 minus0045 minus0163 minus0201 minus0093 0113 minus0033
Gigaspora minus0644 minus0385 minus0672 minus0466 0042 minus024 minus0419 0636 0581 minus0711 Glomus 0341 minus0133 0242 0033 minus0086 0199 0279 minus0363 minus0258 0393
Rhizophagus 0275 minus0152 minus0376 minus0438 minus0378 0023 0081 minus0056 minus0124 0208Septogolomus minus0138 0252 023 0404 021 0179 0193 minus0194 minus0104 0288
Ancestral minus0367 minus0508 minus0508 minus0524 minus0036 minus0425 minus0513 0575 0347 minus0757 Edaphophilic minus0083 0381 minus0341 minus0154 minus004 minus0218 minus0381 0505 0509 minus0582 Rhizophilic 0361 0431 053 052 004 0435 0541 minus0617 minus04 0801
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C
total carbon N total nitrogen P total phosphorus HLD external hyphal length density SD spore density SRspecies richness H ShannonndashWiener index D Simpson index p lt 005 p lt 001
Archaeospora was in a negative correlation with soil NP (p lt 005) Funneliformis waspositively correlated with soil TOC (p lt 001) and Gigaspora was negatively connectedwith soil pH NH4
+-N (p lt 005) and soil NP (p lt 001) but positively related to soil Pand CN (p lt 005) Ancestral AMF was in a negative correlation with soil NP (p lt 001)edaphophilic AMF was negatively correlated with soil NP (p lt 005) and rhizophilic AMFwas positively connected with soil NP (p lt 001) but negatively related to soil P (p lt 005)
RDA was conducted to evaluate the relationship between the soil chemical properties and theAMF community (Figure 4) RDA1 accounted for 6699 of the variations while RDA2 accountedfor 2189 Of the evaluated attributes the NP had significant conditional effects (p lt 001) whilethe soil TOC (p = 0076) and AN (p = 0092) caused referential effects to the AMF communities
Forests 2022 13 x FOR PEER REVIEW 8 of 16
density SD spore density SR species richness H ShannonndashWiener index D Simpson index p lt
005 p lt 001
Archaeospora was in a negative correlation with soil NP (p 005) Funneliformis was
positively correlated with soil TOC (p 001) and Gigaspora was negatively connected
with soil pH NH4+-N (p 005) and soil NP (p 001) but positively related to soil P and
CN (p 005) Ancestral AMF was in a negative correlation with soil NP (p 001)
edaphophilic AMF was negatively correlated with soil NP (p 005) and rhizophilic AMF
was positively connected with soil NP (p 001) but negatively related to soil P (p 005)
RDA was conducted to evaluate the relationship between the soil chemical properties
and the AMF community (Figure 4) RDA1 accounted for 6699 of the variations while
RDA2 accounted for 2189 Of the evaluated attributes the NP had significant condi-
tional effects (p 001) while the soil TOC (p = 0076) and AN (p = 0092) caused referential
effects to the AMF communities
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular
mycorrhizal fungi communities under different nitrogen treatments TOC total organic carbon
NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C total carbon
N total nitrogen P total phosphorus The red arrows represent soil properties the blue arrows
represent arbuscular mycorrhizal fungi communities
4 Discussion
N addition can make a difference on AMF diversity and community composition
because with the increase of soil nutrients plants would have a strategy to acquire nutri-
ents by fine roots or AMF [2644] Our results showed that NP is a factor that has a sig-
nificant effect on AMF communities (Figure 4) AMF community structure might have a
connection with the dynamics of soil conditions and C-to-P trade between AMF and
plants It has been agreed that AMF exchanges P derived from soil for plant carbon [1744ndash
47] In our study N addition had insignificant influence on AMF colonization external
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular mycorrhizalfungi communities under different nitrogen treatments TOC total organic carbon NH4
+-N ammoniumnitrogen AN available nitrogen AP available phosphorus C total carbon N total nitrogen P total phosphorusThe red arrows represent soil properties the blue arrows represent arbuscular mycorrhizal fungi communities
Forests 2022 13 979 8 of 14
4 Discussion
N addition can make a difference on AMF diversity and community compositionbecause with the increase of soil nutrients plants would have a strategy to acquire nutrientsby fine roots or AMF [2644] Our results showed that NP is a factor that has a significanteffect on AMF communities (Figure 4) AMF community structure might have a connectionwith the dynamics of soil conditions and C-to-P trade between AMF and plants It has beenagreed that AMF exchanges P derived from soil for plant carbon [1744ndash47] In our studyN addition had insignificant influence on AMF colonization external HLD spore densityor diversity This finding may be explained as the symbiotic relationship between AMFand root system being stable [48]
Generally there is little specificity between AMF and the host plant and severalAMF species could colonize one AM plant at the same time [4950] However differentpatterns of colonization with N addition suggest a preference for specific AMF species [51]In our study we observed that Glomeraceae was the most predominant family amongall treatments Glomus Septoglomus and Acaulospora were the predominant genera andSeptoglomus viscosum and Acaulospora foveata were the predominant species Our resultsslightly disagree with previous studies which presented that Glomus and Acaulospora asthe predominant species due to their smaller size of spore which helped them producemore spores in less time [485253] This discrepancy can be answered by the diversespecies and age of the host plants The high isolation frequencies of Septoglomus viscosumindicated that it has a strong ability to adapt to the environment (N deposition) Septoglomusviscosum can improve plant growth and production [54] which is mainly related to rootand aboveground biomass [55]
Archaeospora schenckii Entrophospora infrequens and Glomus ambisporum disappearedwith N treatments (Figure 3) which was in keeping with Eom [19] who also found thatEntrophospora infrequens decreased with N addition In P-rich (low NP) soils nitrogenapplication reduces underground carbon allocation and rare species decrease as N in-creases A previous study showed that the influence of N addition on AMF communityappeared different with added N promoting Glomeraceae in A Sieversiana while lesseningthem in L chinensis [16] Some Glomus species were sensitive to N application whichindicated that certain species of AMF play an indispensable role in host plant response toenvironmental change
As identified in most surveys conducted in other fields [56ndash58] we also discoveredFunneliformis mosseae presenting in our study which was reported to promote plant nutri-ents [59] GmosAAP1 an amino acid permease has been characterized from the extraradicalstructure of Funneliformis mosseae which indicates a potential for amino acid uptake [60]Our study showed that with N addition its importance decreased This decrease maybe explained by a decrease in the dependency on Funneliformis mosseae for N nutrientacquisition by the host plant
Above all our results showed that AMF participating in and manage the soil nutrientcycle could be sensitive to small changes in edatopes Therefore studying mycorrhizalsymbionts increases the knowledge of climatic variation [61ndash63]
AMF were grouped into three guilds namely edaphophilic rhizophilc and ancestralIn our study the percentage followed the order rhizophilic gt edaphophilic gt ancestral(Figure 5c) N addition altered AMF species but did not change functional groups Despitethe addition of nitrogen trees may reduce dependence on AMF Meanwhile a large groupof rhizophilic guilds function to reduce root pathogen infection and primarily benefit plantswith fine roots prone to pathogen infections
Forests 2022 13 979 9 of 14Forests 2022 13 x FOR PEER REVIEW 10 of 16
Figure 5 Cont
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-
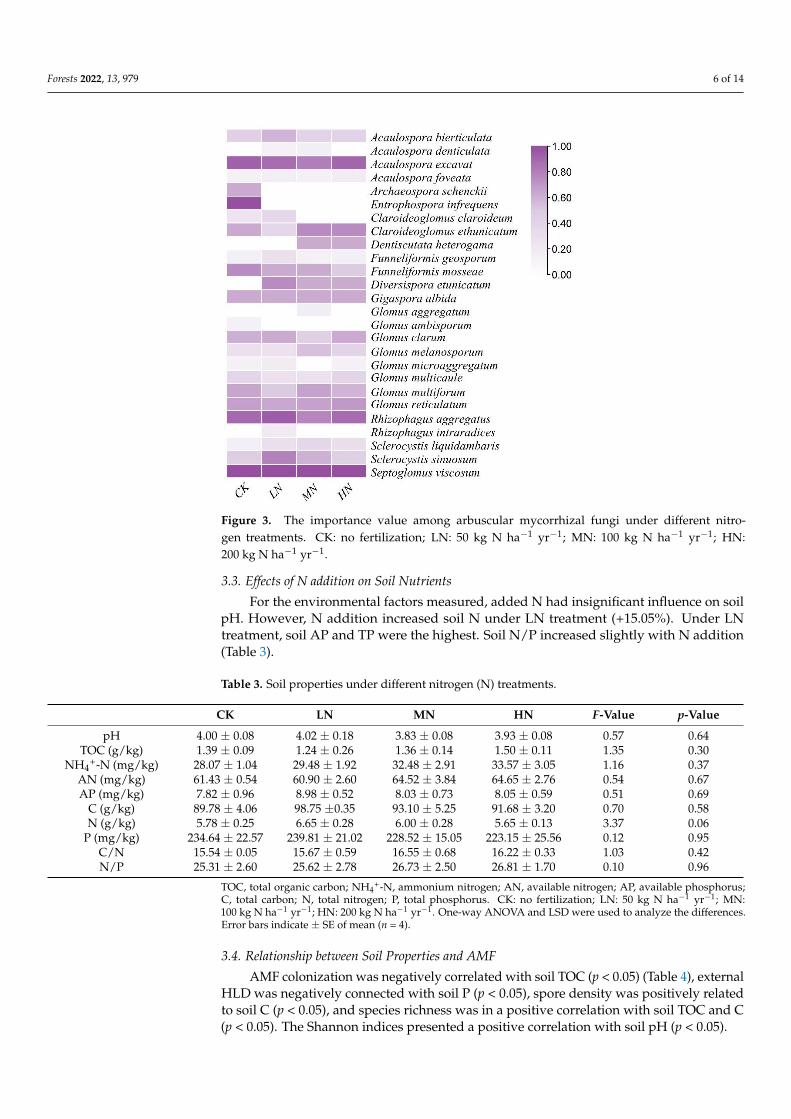
Forests 2022 13 979 6 of 14
Forests 2022 13 x FOR PEER REVIEW 6 of 16
Acaulospora bireticulata Acaulospora excacat Glomus clarum Glomus melanosporum Glo-
mus multicaule Glomus reticulatum Rhizophagus aggregatus Sclerocystis liquidambaris and
Septoglomus viscosum were the dominant species
Among all treatments we observed that IV top five were Septoglomus viscosum
(100) Rhizophagus aggregatus (9870) Acaulospora excavat (8769) Sclerocystis sin-
uosum (7326) Glomus reticulatum (6731) (Table 2)
For the CK treatment Septoglomus viscosum and Entrophospora infrequens (100)
had the highest IV followed by Acaulospora excavat (9213) Rhizophagus aggregatus
(8929) and Funneliformis mosseae (7500) For the LN treatment Septoglomus vis-
cosum (100) presented the highest IV followed by Rhizophagus aggregatus (9375)
Acaulospora excavat (8762) Sclerocystis sinuosum (7917) and Diversispora etunica-
tum (7500) For the MN treatment Septoglomus viscosum was the highest IV followed
by Acaulospora excavat (8056) Rhizophagus aggregatus (7738) Claroideoglomus
etunicatum (7500) and Glomus multiform (6635) For the HN treatment Septoglo-
mus viscosum had the highest IV followed by Acaulospora excavat (9042) Rhizopha-
gus aggregatus (8833) Claroideoglomus etunicatum (7500) and Glomus reticultaum
(6956) (Figure 3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitrogen treat-
ments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soil
pH However N addition increased soil N under LN treatment (+1505) Under LN treat-
ment soil AP and TP were the highest Soil NP increased slightly with N addition (Table
3)
Figure 3 The importance value among arbuscular mycorrhizal fungi under different nitro-gen treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100 kg N haminus1 yrminus1 HN200 kg N haminus1 yrminus1
33 Effects of N addition on Soil Nutrients
For the environmental factors measured added N had insignificant influence on soilpH However N addition increased soil N under LN treatment (+1505) Under LNtreatment soil AP and TP were the highest Soil NP increased slightly with N addition(Table 3)
Table 3 Soil properties under different nitrogen (N) treatments
CK LN MN HN F-Value p-Value
pH 400 plusmn 008 402 plusmn 018 383 plusmn 008 393 plusmn 008 057 064TOC (gkg) 139 plusmn 009 124 plusmn 026 136 plusmn 014 150 plusmn 011 135 030
NH4+-N (mgkg) 2807 plusmn 104 2948 plusmn 192 3248 plusmn 291 3357 plusmn 305 116 037
AN (mgkg) 6143 plusmn 054 6090 plusmn 260 6452 plusmn 384 6465 plusmn 276 054 067AP (mgkg) 782 plusmn 096 898 plusmn 052 803 plusmn 073 805 plusmn 059 051 069
C (gkg) 8978 plusmn 406 9875 plusmn035 9310 plusmn 525 9168 plusmn 320 070 058N (gkg) 578 plusmn 025 665 plusmn 028 600 plusmn 028 565 plusmn 013 337 006
P (mgkg) 23464 plusmn 2257 23981 plusmn 2102 22852 plusmn 1505 22315 plusmn 2556 012 095CN 1554 plusmn 005 1567 plusmn 059 1655 plusmn 068 1622 plusmn 033 103 042NP 2531 plusmn 260 2562 plusmn 278 2673 plusmn 250 2681 plusmn 170 010 096
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus
C total carbon N total nitrogen P total phosphorus CK no fertilization LN 50 kg N haminus1 yrminus1 MN100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1 One-way ANOVA and LSD were used to analyze the differencesError bars indicate plusmn SE of mean (n = 4)
34 Relationship between Soil Properties and AMF
AMF colonization was negatively correlated with soil TOC (p lt 005) (Table 4) externalHLD was negatively connected with soil P (p lt 005) spore density was positively relatedto soil C (p lt 005) and species richness was in a positive correlation with soil TOC and C(p lt 005) The Shannon indices presented a positive correlation with soil pH (p lt 005)
Forests 2022 13 979 7 of 14
Table 4 Spearman correlation coefficients between soil properties and parameters of arbuscularmycorrhizal fungi
pH TOC NH4+-N AN AP C N P CN NP
Colonization 0012 minus0643 minus0437 minus0571 minus0245 minus014 minus0126 0142 0025 minus0112HLD minus0062 0267 0245 0288 minus0531 minus026 minus0197 minus0701 minus0154 0564SD minus0338 0214 0008 0251 0161 0679 0477 0283 0489 0002SR 0101 0604 0488 0551 0114 063 0478 0039 033 0117H 0623 0478 0341 0142 0081 0306 0328 minus0023 minus0098 0117D 0560 0408 0264 0105 012 0193 0209 0002 minus0066 007
Acaulospora minus0378 minus0575 minus0506 minus052 0004 minus0332 minus0418 0575 0332 minus0708 Archaeospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025
Claroideoglomus 0464 minus0303 0302 minus0029 minus0047 0077 0175 0071 minus0268 minus0045Dentiscutata 0014 0244 0434 0451 017 0453 0349 0165 0203 minus0047Diversispora 0057 0482 minus0205 minus0057 minus0051 minus0173 minus0304 0384 0399 minus0447
Entrophospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025Funneliformis 005 0764 0175 0266 minus0045 minus0163 minus0201 minus0093 0113 minus0033
Gigaspora minus0644 minus0385 minus0672 minus0466 0042 minus024 minus0419 0636 0581 minus0711 Glomus 0341 minus0133 0242 0033 minus0086 0199 0279 minus0363 minus0258 0393
Rhizophagus 0275 minus0152 minus0376 minus0438 minus0378 0023 0081 minus0056 minus0124 0208Septogolomus minus0138 0252 023 0404 021 0179 0193 minus0194 minus0104 0288
Ancestral minus0367 minus0508 minus0508 minus0524 minus0036 minus0425 minus0513 0575 0347 minus0757 Edaphophilic minus0083 0381 minus0341 minus0154 minus004 minus0218 minus0381 0505 0509 minus0582 Rhizophilic 0361 0431 053 052 004 0435 0541 minus0617 minus04 0801
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C
total carbon N total nitrogen P total phosphorus HLD external hyphal length density SD spore density SRspecies richness H ShannonndashWiener index D Simpson index p lt 005 p lt 001
Archaeospora was in a negative correlation with soil NP (p lt 005) Funneliformis waspositively correlated with soil TOC (p lt 001) and Gigaspora was negatively connectedwith soil pH NH4
+-N (p lt 005) and soil NP (p lt 001) but positively related to soil Pand CN (p lt 005) Ancestral AMF was in a negative correlation with soil NP (p lt 001)edaphophilic AMF was negatively correlated with soil NP (p lt 005) and rhizophilic AMFwas positively connected with soil NP (p lt 001) but negatively related to soil P (p lt 005)
RDA was conducted to evaluate the relationship between the soil chemical properties and theAMF community (Figure 4) RDA1 accounted for 6699 of the variations while RDA2 accountedfor 2189 Of the evaluated attributes the NP had significant conditional effects (p lt 001) whilethe soil TOC (p = 0076) and AN (p = 0092) caused referential effects to the AMF communities
Forests 2022 13 x FOR PEER REVIEW 8 of 16
density SD spore density SR species richness H ShannonndashWiener index D Simpson index p lt
005 p lt 001
Archaeospora was in a negative correlation with soil NP (p 005) Funneliformis was
positively correlated with soil TOC (p 001) and Gigaspora was negatively connected
with soil pH NH4+-N (p 005) and soil NP (p 001) but positively related to soil P and
CN (p 005) Ancestral AMF was in a negative correlation with soil NP (p 001)
edaphophilic AMF was negatively correlated with soil NP (p 005) and rhizophilic AMF
was positively connected with soil NP (p 001) but negatively related to soil P (p 005)
RDA was conducted to evaluate the relationship between the soil chemical properties
and the AMF community (Figure 4) RDA1 accounted for 6699 of the variations while
RDA2 accounted for 2189 Of the evaluated attributes the NP had significant condi-
tional effects (p 001) while the soil TOC (p = 0076) and AN (p = 0092) caused referential
effects to the AMF communities
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular
mycorrhizal fungi communities under different nitrogen treatments TOC total organic carbon
NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C total carbon
N total nitrogen P total phosphorus The red arrows represent soil properties the blue arrows
represent arbuscular mycorrhizal fungi communities
4 Discussion
N addition can make a difference on AMF diversity and community composition
because with the increase of soil nutrients plants would have a strategy to acquire nutri-
ents by fine roots or AMF [2644] Our results showed that NP is a factor that has a sig-
nificant effect on AMF communities (Figure 4) AMF community structure might have a
connection with the dynamics of soil conditions and C-to-P trade between AMF and
plants It has been agreed that AMF exchanges P derived from soil for plant carbon [1744ndash
47] In our study N addition had insignificant influence on AMF colonization external
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular mycorrhizalfungi communities under different nitrogen treatments TOC total organic carbon NH4
+-N ammoniumnitrogen AN available nitrogen AP available phosphorus C total carbon N total nitrogen P total phosphorusThe red arrows represent soil properties the blue arrows represent arbuscular mycorrhizal fungi communities
Forests 2022 13 979 8 of 14
4 Discussion
N addition can make a difference on AMF diversity and community compositionbecause with the increase of soil nutrients plants would have a strategy to acquire nutrientsby fine roots or AMF [2644] Our results showed that NP is a factor that has a significanteffect on AMF communities (Figure 4) AMF community structure might have a connectionwith the dynamics of soil conditions and C-to-P trade between AMF and plants It has beenagreed that AMF exchanges P derived from soil for plant carbon [1744ndash47] In our studyN addition had insignificant influence on AMF colonization external HLD spore densityor diversity This finding may be explained as the symbiotic relationship between AMFand root system being stable [48]
Generally there is little specificity between AMF and the host plant and severalAMF species could colonize one AM plant at the same time [4950] However differentpatterns of colonization with N addition suggest a preference for specific AMF species [51]In our study we observed that Glomeraceae was the most predominant family amongall treatments Glomus Septoglomus and Acaulospora were the predominant genera andSeptoglomus viscosum and Acaulospora foveata were the predominant species Our resultsslightly disagree with previous studies which presented that Glomus and Acaulospora asthe predominant species due to their smaller size of spore which helped them producemore spores in less time [485253] This discrepancy can be answered by the diversespecies and age of the host plants The high isolation frequencies of Septoglomus viscosumindicated that it has a strong ability to adapt to the environment (N deposition) Septoglomusviscosum can improve plant growth and production [54] which is mainly related to rootand aboveground biomass [55]
Archaeospora schenckii Entrophospora infrequens and Glomus ambisporum disappearedwith N treatments (Figure 3) which was in keeping with Eom [19] who also found thatEntrophospora infrequens decreased with N addition In P-rich (low NP) soils nitrogenapplication reduces underground carbon allocation and rare species decrease as N in-creases A previous study showed that the influence of N addition on AMF communityappeared different with added N promoting Glomeraceae in A Sieversiana while lesseningthem in L chinensis [16] Some Glomus species were sensitive to N application whichindicated that certain species of AMF play an indispensable role in host plant response toenvironmental change
As identified in most surveys conducted in other fields [56ndash58] we also discoveredFunneliformis mosseae presenting in our study which was reported to promote plant nutri-ents [59] GmosAAP1 an amino acid permease has been characterized from the extraradicalstructure of Funneliformis mosseae which indicates a potential for amino acid uptake [60]Our study showed that with N addition its importance decreased This decrease maybe explained by a decrease in the dependency on Funneliformis mosseae for N nutrientacquisition by the host plant
Above all our results showed that AMF participating in and manage the soil nutrientcycle could be sensitive to small changes in edatopes Therefore studying mycorrhizalsymbionts increases the knowledge of climatic variation [61ndash63]
AMF were grouped into three guilds namely edaphophilic rhizophilc and ancestralIn our study the percentage followed the order rhizophilic gt edaphophilic gt ancestral(Figure 5c) N addition altered AMF species but did not change functional groups Despitethe addition of nitrogen trees may reduce dependence on AMF Meanwhile a large groupof rhizophilic guilds function to reduce root pathogen infection and primarily benefit plantswith fine roots prone to pathogen infections
Forests 2022 13 979 9 of 14Forests 2022 13 x FOR PEER REVIEW 10 of 16
Figure 5 Cont
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-
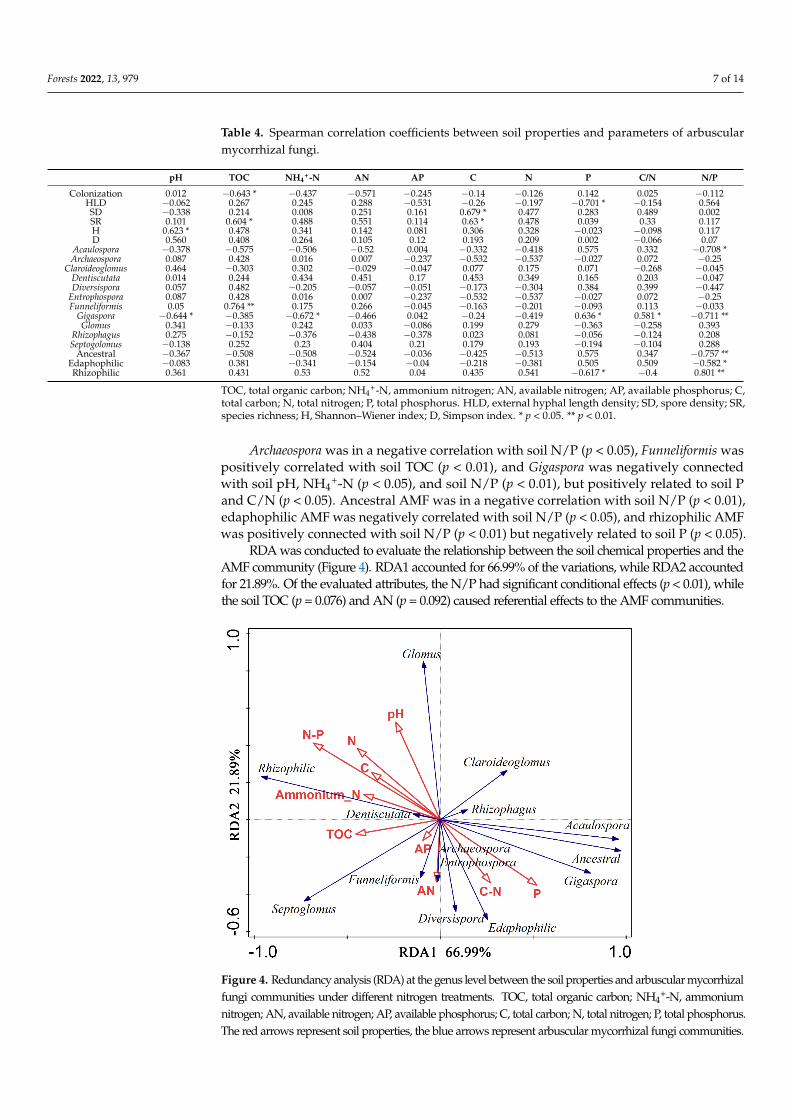
Forests 2022 13 979 7 of 14
Table 4 Spearman correlation coefficients between soil properties and parameters of arbuscularmycorrhizal fungi
pH TOC NH4+-N AN AP C N P CN NP
Colonization 0012 minus0643 minus0437 minus0571 minus0245 minus014 minus0126 0142 0025 minus0112HLD minus0062 0267 0245 0288 minus0531 minus026 minus0197 minus0701 minus0154 0564SD minus0338 0214 0008 0251 0161 0679 0477 0283 0489 0002SR 0101 0604 0488 0551 0114 063 0478 0039 033 0117H 0623 0478 0341 0142 0081 0306 0328 minus0023 minus0098 0117D 0560 0408 0264 0105 012 0193 0209 0002 minus0066 007
Acaulospora minus0378 minus0575 minus0506 minus052 0004 minus0332 minus0418 0575 0332 minus0708 Archaeospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025
Claroideoglomus 0464 minus0303 0302 minus0029 minus0047 0077 0175 0071 minus0268 minus0045Dentiscutata 0014 0244 0434 0451 017 0453 0349 0165 0203 minus0047Diversispora 0057 0482 minus0205 minus0057 minus0051 minus0173 minus0304 0384 0399 minus0447
Entrophospora 0087 0428 0016 0007 minus0237 minus0532 minus0537 minus0027 0072 minus025Funneliformis 005 0764 0175 0266 minus0045 minus0163 minus0201 minus0093 0113 minus0033
Gigaspora minus0644 minus0385 minus0672 minus0466 0042 minus024 minus0419 0636 0581 minus0711 Glomus 0341 minus0133 0242 0033 minus0086 0199 0279 minus0363 minus0258 0393
Rhizophagus 0275 minus0152 minus0376 minus0438 minus0378 0023 0081 minus0056 minus0124 0208Septogolomus minus0138 0252 023 0404 021 0179 0193 minus0194 minus0104 0288
Ancestral minus0367 minus0508 minus0508 minus0524 minus0036 minus0425 minus0513 0575 0347 minus0757 Edaphophilic minus0083 0381 minus0341 minus0154 minus004 minus0218 minus0381 0505 0509 minus0582 Rhizophilic 0361 0431 053 052 004 0435 0541 minus0617 minus04 0801
TOC total organic carbon NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C
total carbon N total nitrogen P total phosphorus HLD external hyphal length density SD spore density SRspecies richness H ShannonndashWiener index D Simpson index p lt 005 p lt 001
Archaeospora was in a negative correlation with soil NP (p lt 005) Funneliformis waspositively correlated with soil TOC (p lt 001) and Gigaspora was negatively connectedwith soil pH NH4
+-N (p lt 005) and soil NP (p lt 001) but positively related to soil Pand CN (p lt 005) Ancestral AMF was in a negative correlation with soil NP (p lt 001)edaphophilic AMF was negatively correlated with soil NP (p lt 005) and rhizophilic AMFwas positively connected with soil NP (p lt 001) but negatively related to soil P (p lt 005)
RDA was conducted to evaluate the relationship between the soil chemical properties and theAMF community (Figure 4) RDA1 accounted for 6699 of the variations while RDA2 accountedfor 2189 Of the evaluated attributes the NP had significant conditional effects (p lt 001) whilethe soil TOC (p = 0076) and AN (p = 0092) caused referential effects to the AMF communities
Forests 2022 13 x FOR PEER REVIEW 8 of 16
density SD spore density SR species richness H ShannonndashWiener index D Simpson index p lt
005 p lt 001
Archaeospora was in a negative correlation with soil NP (p 005) Funneliformis was
positively correlated with soil TOC (p 001) and Gigaspora was negatively connected
with soil pH NH4+-N (p 005) and soil NP (p 001) but positively related to soil P and
CN (p 005) Ancestral AMF was in a negative correlation with soil NP (p 001)
edaphophilic AMF was negatively correlated with soil NP (p 005) and rhizophilic AMF
was positively connected with soil NP (p 001) but negatively related to soil P (p 005)
RDA was conducted to evaluate the relationship between the soil chemical properties
and the AMF community (Figure 4) RDA1 accounted for 6699 of the variations while
RDA2 accounted for 2189 Of the evaluated attributes the NP had significant condi-
tional effects (p 001) while the soil TOC (p = 0076) and AN (p = 0092) caused referential
effects to the AMF communities
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular
mycorrhizal fungi communities under different nitrogen treatments TOC total organic carbon
NH4+-N ammonium nitrogen AN available nitrogen AP available phosphorus C total carbon
N total nitrogen P total phosphorus The red arrows represent soil properties the blue arrows
represent arbuscular mycorrhizal fungi communities
4 Discussion
N addition can make a difference on AMF diversity and community composition
because with the increase of soil nutrients plants would have a strategy to acquire nutri-
ents by fine roots or AMF [2644] Our results showed that NP is a factor that has a sig-
nificant effect on AMF communities (Figure 4) AMF community structure might have a
connection with the dynamics of soil conditions and C-to-P trade between AMF and
plants It has been agreed that AMF exchanges P derived from soil for plant carbon [1744ndash
47] In our study N addition had insignificant influence on AMF colonization external
Figure 4 Redundancy analysis (RDA) at the genus level between the soil properties and arbuscular mycorrhizalfungi communities under different nitrogen treatments TOC total organic carbon NH4
+-N ammoniumnitrogen AN available nitrogen AP available phosphorus C total carbon N total nitrogen P total phosphorusThe red arrows represent soil properties the blue arrows represent arbuscular mycorrhizal fungi communities
Forests 2022 13 979 8 of 14
4 Discussion
N addition can make a difference on AMF diversity and community compositionbecause with the increase of soil nutrients plants would have a strategy to acquire nutrientsby fine roots or AMF [2644] Our results showed that NP is a factor that has a significanteffect on AMF communities (Figure 4) AMF community structure might have a connectionwith the dynamics of soil conditions and C-to-P trade between AMF and plants It has beenagreed that AMF exchanges P derived from soil for plant carbon [1744ndash47] In our studyN addition had insignificant influence on AMF colonization external HLD spore densityor diversity This finding may be explained as the symbiotic relationship between AMFand root system being stable [48]
Generally there is little specificity between AMF and the host plant and severalAMF species could colonize one AM plant at the same time [4950] However differentpatterns of colonization with N addition suggest a preference for specific AMF species [51]In our study we observed that Glomeraceae was the most predominant family amongall treatments Glomus Septoglomus and Acaulospora were the predominant genera andSeptoglomus viscosum and Acaulospora foveata were the predominant species Our resultsslightly disagree with previous studies which presented that Glomus and Acaulospora asthe predominant species due to their smaller size of spore which helped them producemore spores in less time [485253] This discrepancy can be answered by the diversespecies and age of the host plants The high isolation frequencies of Septoglomus viscosumindicated that it has a strong ability to adapt to the environment (N deposition) Septoglomusviscosum can improve plant growth and production [54] which is mainly related to rootand aboveground biomass [55]
Archaeospora schenckii Entrophospora infrequens and Glomus ambisporum disappearedwith N treatments (Figure 3) which was in keeping with Eom [19] who also found thatEntrophospora infrequens decreased with N addition In P-rich (low NP) soils nitrogenapplication reduces underground carbon allocation and rare species decrease as N in-creases A previous study showed that the influence of N addition on AMF communityappeared different with added N promoting Glomeraceae in A Sieversiana while lesseningthem in L chinensis [16] Some Glomus species were sensitive to N application whichindicated that certain species of AMF play an indispensable role in host plant response toenvironmental change
As identified in most surveys conducted in other fields [56ndash58] we also discoveredFunneliformis mosseae presenting in our study which was reported to promote plant nutri-ents [59] GmosAAP1 an amino acid permease has been characterized from the extraradicalstructure of Funneliformis mosseae which indicates a potential for amino acid uptake [60]Our study showed that with N addition its importance decreased This decrease maybe explained by a decrease in the dependency on Funneliformis mosseae for N nutrientacquisition by the host plant
Above all our results showed that AMF participating in and manage the soil nutrientcycle could be sensitive to small changes in edatopes Therefore studying mycorrhizalsymbionts increases the knowledge of climatic variation [61ndash63]
AMF were grouped into three guilds namely edaphophilic rhizophilc and ancestralIn our study the percentage followed the order rhizophilic gt edaphophilic gt ancestral(Figure 5c) N addition altered AMF species but did not change functional groups Despitethe addition of nitrogen trees may reduce dependence on AMF Meanwhile a large groupof rhizophilic guilds function to reduce root pathogen infection and primarily benefit plantswith fine roots prone to pathogen infections
Forests 2022 13 979 9 of 14Forests 2022 13 x FOR PEER REVIEW 10 of 16
Figure 5 Cont
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-

Forests 2022 13 979 8 of 14
4 Discussion
N addition can make a difference on AMF diversity and community compositionbecause with the increase of soil nutrients plants would have a strategy to acquire nutrientsby fine roots or AMF [2644] Our results showed that NP is a factor that has a significanteffect on AMF communities (Figure 4) AMF community structure might have a connectionwith the dynamics of soil conditions and C-to-P trade between AMF and plants It has beenagreed that AMF exchanges P derived from soil for plant carbon [1744ndash47] In our studyN addition had insignificant influence on AMF colonization external HLD spore densityor diversity This finding may be explained as the symbiotic relationship between AMFand root system being stable [48]
Generally there is little specificity between AMF and the host plant and severalAMF species could colonize one AM plant at the same time [4950] However differentpatterns of colonization with N addition suggest a preference for specific AMF species [51]In our study we observed that Glomeraceae was the most predominant family amongall treatments Glomus Septoglomus and Acaulospora were the predominant genera andSeptoglomus viscosum and Acaulospora foveata were the predominant species Our resultsslightly disagree with previous studies which presented that Glomus and Acaulospora asthe predominant species due to their smaller size of spore which helped them producemore spores in less time [485253] This discrepancy can be answered by the diversespecies and age of the host plants The high isolation frequencies of Septoglomus viscosumindicated that it has a strong ability to adapt to the environment (N deposition) Septoglomusviscosum can improve plant growth and production [54] which is mainly related to rootand aboveground biomass [55]
Archaeospora schenckii Entrophospora infrequens and Glomus ambisporum disappearedwith N treatments (Figure 3) which was in keeping with Eom [19] who also found thatEntrophospora infrequens decreased with N addition In P-rich (low NP) soils nitrogenapplication reduces underground carbon allocation and rare species decrease as N in-creases A previous study showed that the influence of N addition on AMF communityappeared different with added N promoting Glomeraceae in A Sieversiana while lesseningthem in L chinensis [16] Some Glomus species were sensitive to N application whichindicated that certain species of AMF play an indispensable role in host plant response toenvironmental change
As identified in most surveys conducted in other fields [56ndash58] we also discoveredFunneliformis mosseae presenting in our study which was reported to promote plant nutri-ents [59] GmosAAP1 an amino acid permease has been characterized from the extraradicalstructure of Funneliformis mosseae which indicates a potential for amino acid uptake [60]Our study showed that with N addition its importance decreased This decrease maybe explained by a decrease in the dependency on Funneliformis mosseae for N nutrientacquisition by the host plant
Above all our results showed that AMF participating in and manage the soil nutrientcycle could be sensitive to small changes in edatopes Therefore studying mycorrhizalsymbionts increases the knowledge of climatic variation [61ndash63]
AMF were grouped into three guilds namely edaphophilic rhizophilc and ancestralIn our study the percentage followed the order rhizophilic gt edaphophilic gt ancestral(Figure 5c) N addition altered AMF species but did not change functional groups Despitethe addition of nitrogen trees may reduce dependence on AMF Meanwhile a large groupof rhizophilic guilds function to reduce root pathogen infection and primarily benefit plantswith fine roots prone to pathogen infections
Forests 2022 13 979 9 of 14Forests 2022 13 x FOR PEER REVIEW 10 of 16
Figure 5 Cont
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-

Forests 2022 13 979 9 of 14Forests 2022 13 x FOR PEER REVIEW 10 of 16
Figure 5 Cont
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-
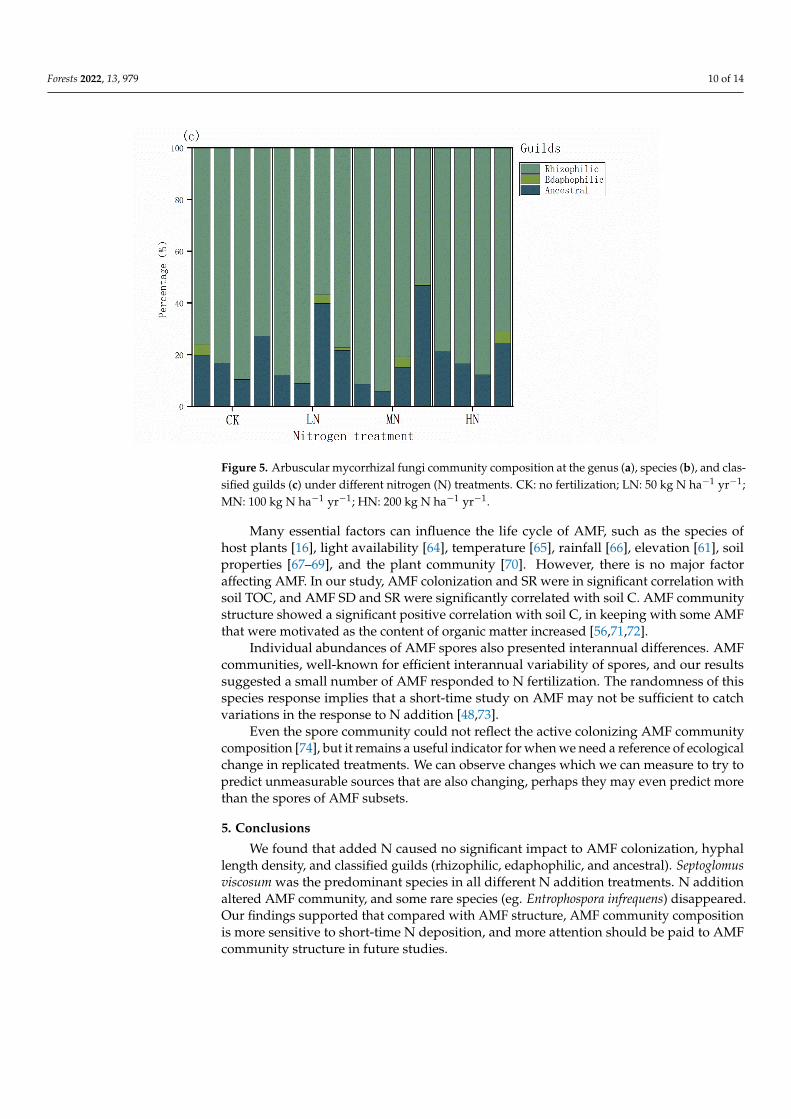
Forests 2022 13 979 10 of 14Forests 2022 13 x FOR PEER REVIEW 11 of 16
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and classified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1 MN 100kg N haminus1 yrminus1 HN 200kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species of host plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soil prop-erties [67ndash69] and the plant community [70] However there is no major factor affecting AMF In our study AMF colonization and SR were in significant correlation with soil TOC and AMF SD and SR were significantly correlated with soil C AMF community structure showed a significant positive correlation with soil C in keeping with some AMF that were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMF communities well-known for efficient interannual variability of spores and our results suggested a small number of AMF responded to N fertilization The randomness of this species response implies that a short-time study on AMF may not be sufficient to catch variations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF community composition [74] but it remains a useful indicator for when we need a reference of eco-logical change in replicated treatments We can observe changes which we can measure to try to predict unmeasurable sources that are also changing perhaps they may even predict more than the spores of AMF subsets
5 Conclusions We found that added N caused no significant impact to AMF colonization hyphal
length density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglo-mus viscosum was the predominant species in all different N addition treatments N addi-tion altered AMF community and some rare species (eg Entrophospora infrequens) disap-peared Our findings supported that compared with AMF structure AMF community composition is more sensitive to short-time N deposition and more attention should be paid to AMF community structure in future studies
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XG and FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZ and MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and
Figure 5 Arbuscular mycorrhizal fungi community composition at the genus (a) species (b) and clas-sified guilds (c) under different nitrogen (N) treatments CK no fertilization LN 50 kg N haminus1 yrminus1MN 100 kg N haminus1 yrminus1 HN 200 kg N haminus1 yrminus1
Many essential factors can influence the life cycle of AMF such as the species ofhost plants [16] light availability [64] temperature [65] rainfall [66] elevation [61] soilproperties [67ndash69] and the plant community [70] However there is no major factoraffecting AMF In our study AMF colonization and SR were in significant correlation withsoil TOC and AMF SD and SR were significantly correlated with soil C AMF communitystructure showed a significant positive correlation with soil C in keeping with some AMFthat were motivated as the content of organic matter increased [567172]
Individual abundances of AMF spores also presented interannual differences AMFcommunities well-known for efficient interannual variability of spores and our resultssuggested a small number of AMF responded to N fertilization The randomness of thisspecies response implies that a short-time study on AMF may not be sufficient to catchvariations in the response to N addition [4873]
Even the spore community could not reflect the active colonizing AMF communitycomposition [74] but it remains a useful indicator for when we need a reference of ecologicalchange in replicated treatments We can observe changes which we can measure to try topredict unmeasurable sources that are also changing perhaps they may even predict morethan the spores of AMF subsets
5 Conclusions
We found that added N caused no significant impact to AMF colonization hyphallength density and classified guilds (rhizophilic edaphophilic and ancestral) Septoglomusviscosum was the predominant species in all different N addition treatments N additionaltered AMF community and some rare species (eg Entrophospora infrequens) disappearedOur findings supported that compared with AMF structure AMF community compositionis more sensitive to short-time N deposition and more attention should be paid to AMFcommunity structure in future studies
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-
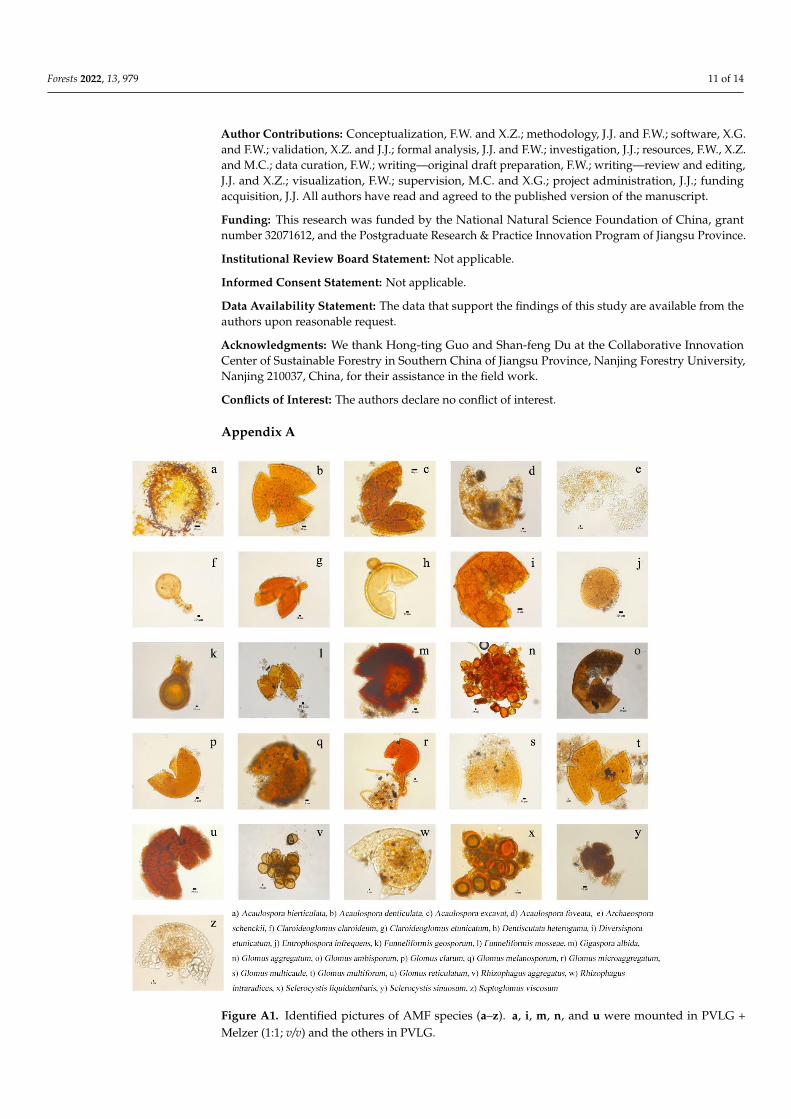
Forests 2022 13 979 11 of 14
Author Contributions Conceptualization FW and XZ methodology JJ and FW software XGand FW validation XZ and JJ formal analysis JJ and FW investigation JJ resources FW XZand MC data curation FW writingmdashoriginal draft preparation FW writingmdashreview and editingJJ and XZ visualization FW supervision MC and XG project administration JJ fundingacquisition JJ All authors have read and agreed to the published version of the manuscript
Funding This research was funded by the National Natural Science Foundation of China grantnumber 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Province
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from theauthors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative InnovationCenter of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry UniversityNanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Forests 2022 13 x FOR PEER REVIEW 12 of 16
editing JJ and XZ visualization FW supervision MC and XG project administration JJ
funding acquisition JJ All authors have read and agreed to the published version of the manu-
script
Funding This research was funded by the National Natural Science Foundation of China grant
number 32071612 and the Postgraduate Research amp Practice Innovation Program of Jiangsu Prov-
ince
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data that support the findings of this study are available from the
authors upon reasonable request
Acknowledgments We thank Hong-ting Guo and Shan-feng Du at the Collaborative Innovation
Center of Sustainable Forestry in Southern China of Jiangsu Province Nanjing Forestry University
Nanjing 210037 China for their assistance in the field work
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Figure A1 Identified pictures of AMF species (andashz) a i m n and u were mounted in PVLG +Melzer (11 vv) and the others in PVLG
Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-

Forests 2022 13 979 12 of 14
References1 Ackerman D Millet DB Chen X Global Estimates of Inorganic Nitrogen Deposition Across Four Decades Glob Biogeochem
Cycles 2018 33 100ndash107 [CrossRef]2 Liu D Guo X Xiao B What causes growth of global greenhouse gas emissions Evidence from 40 countries Sci Total Environ
2019 661 750ndash766 [CrossRef]3 Wen Z Wang R Li Q Liu J Ma X Xu W Tang A Collett JL Li H Liu X Spatiotemporal variations of nitrogen and
phosphorus deposition across China Sci Total Environ 2022 830 154740 [CrossRef]4 Thomas RQ Canham C Weathers KC Goodale C Increased tree carbon storage in response to nitrogen deposition in the
US Nat Geosci 2009 3 229ndash244 [CrossRef]5 Stevens CJ David TI Storkey J Atmospheric nitrogen deposition in terrestrial ecosystems Its impact on plant communities
and consequences across trophic levels Funct Ecol 2018 32 1757ndash1769 [CrossRef]6 Zhang T Chen HYH Ruan H Global negative effects of nitrogen deposition on soil microbes ISME J 2018 12 1817ndash1825
[CrossRef]7 Smith SE Smith FA Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth New Paradigms from Cellular to
Ecosystem Scales Annu Rev Plant Biol 2010 62 227ndash250 [CrossRef]8 Etesami H Jeong BR Glick BR Contribution of Arbuscular Mycorrhizal Fungi PhosphatendashSolubilizing Bacteria and Silicon
to P Uptake by Plant Front Plant Sci 2021 12 699618 [CrossRef]9 Liu H Wu Y Xu H Ai Z Zhang J Liu G Xue S N enrichment affects the arbuscular mycorrhizal fungi-mediated
relationship between a C4 grass and a legume Plant Physiol 2021 187 1519ndash1533 [CrossRef]10 Ma X Geng Q Zhang H Bian C Chen HYH Jiang D Xu X Global negative effects of nutrient enrichment on arbuscular
mycorrhizal fungi plant diversity and ecosystem multifunctionality New Phytol 2020 229 2957ndash2969 [CrossRef]11 Han Y Feng J Han M Zhu B Responses of arbuscular mycorrhizal fungi to nitrogen addition A meta-analysis Glob Chang
Biol 2020 26 7229ndash7241 [CrossRef]12 Weber SE Diez JM Andrews LV Goulden ML Aronson EL Allen MF Responses of arbuscular mycorrhizal fungi to
multiple coinciding global change drivers Fungal Ecol 2019 40 62ndash71 [CrossRef]13 Xiao D Che R Liu X Tan Y Yang R Zhang W He X Xu Z Wang K Arbuscular mycorrhizal fungi abundance was
sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems Biol Fertil Soils 2019 55457ndash469 [CrossRef]
14 Liu M Shen Y Li Q Xiao W Song X Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen andphosphorus additions affects leaf CNP stoichiometry in Chinese fir (Cunninghamia lanceolata) forests Plant Soil 2021 461 421ndash440[CrossRef]
15 Van Diepen LTA Lilleskov EA Pregitzer KS Miller RM Decline of arbuscular mycorrhizal fungi in northern hardwoodforests exposed to chronic nitrogen additions New Phytol 2007 176 175ndash183 [CrossRef]
16 Pan S Wang Y Qiu Y Chen D Zhang L Ye C Guo H Zhu W Chen A Xu G et al Nitrogen-induced acidification notN-nutrient dominates suppressive N effects on arbuscular mycorrhizal fungi Glob Chang Biol 2020 26 6568ndash6580 [CrossRef]
17 Lekberg Y Arnillas CA Borer ET Bullington LS Fierer N Kennedy PG Leff JW Luis AD Seabloom EW HenningJA Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils Nat Commun2021 12 3484 [CrossRef]
18 Wang X Sui X Liu Y Xiang L Zhang T Fu J Li A Yang P NP fertilization did not reduce AMF abundance or diversity butalter AMF composition in an alpine grassland infested by a root hemiparasitic plant Plant Divers 2018 40 117ndash126 [CrossRef]
19 Eom AH Hartnett DC Wilson GW Figge DA The Effect of Fire Mowing and Fertilizer Amendment on ArbuscularMycorrhizas in Tallgrass Prairie Am Midl Nat 1999 142 55ndash70 [CrossRef]
20 Zhang T Yang X Guo R Guo J Response of AM fungi spore population to elevated temperature and nitrogen addition andtheir influence on the plant community composition and productivity Sci Rep 2016 6 24749 [CrossRef]
21 Treseder KK Allen MF Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi A model and field testNew Phytol 2002 155 507ndash515 [CrossRef]
22 Camenzind T Homeier J Dietrich K Hempel S Hertel D Krohn A Leuschner C Oelmann Y Olsson PA Suaacuterez JPet al Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradientin tropical montane forests Soil Biol Biochem 2016 94 37ndash47 [CrossRef]
23 Camenzind T Hempel S Homeier J Horn S Velescu A Wilcke W Rillig MC Nitrogen and phosphorus additions impactarbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest Glob Chang Biol 2014 20 3646ndash3659[CrossRef]
24 Mueller RC Bohannan BJM Shifts in the phylogenetic structure of arbuscular mycorrhizal fungi in response to experimentalnitrogen and carbon dioxide additions Oecologia 2015 179 175ndash185 [CrossRef]
25 Corkidi L Rowland DL Johnson NC Allen EB Nitrogen fertilization alters the functioning of arbuscular mycorrhizas attwo semiarid grasslands Plant Soil 2002 240 299ndash310 [CrossRef]
26 Li L McCormack ML Chen F Wang H Ma Z Guo D Different responses of absorptive roots and arbuscular mycorrhizalfungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir For Ecol Manag 2019 433 64ndash72 [CrossRef]
Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-

Forests 2022 13 979 13 of 14
27 Sheldrake M Rosenstock NP Mangan S Revillini D Sayer EJ Olsson PA Verbruggen E Tanner EVJ Turner BLWright SJ Responses of arbuscular mycorrhizal fungi to long-term inorganic and organic nutrient addition in a lowland tropicalforest ISME J 2018 12 2433ndash2445 [CrossRef]
28 Xiang W Li L Ouyang S Xiao W Zeng L Chen L Lei P Deng X Zeng Y Fang J et al Effects of stand age on treebiomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations Forstwiss Centralblatt 2021140 317ndash332 [CrossRef]
29 Wu H Xiang W Ouyang S Xiao W Li S Chen L Lei P Deng X Zeng Y Zeng L et al Tree growth rate and soilnutrient status determine the shift in nutrient-use strategy of Chinese fir plantations along a chronosequence For Ecol Manag2020 460 117896 [CrossRef]
30 Lu N Xu X Wang P Zhang P Ji B Wang X Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequenceof Cunninghamia lanceolata Sci Rep 2019 9 18057 [CrossRef]
31 Cao J Lin T-C Yang Z Zheng Y Xie L Xiong D Yang Y Warming exerts a stronger effect than nitrogen addition onthe soil arbuscular mycorrhizal fungal community in a young subtropical Cunninghamia lanceolata plantation Geoderma 2020367 114273 [CrossRef]
32 Deng B-L Wang S-L Xu X-T Wang H Hu D-N Guo X-M Shi Q-H Siemann E Zhang L Effects of biochar anddicyandiamide combination on nitrous oxide emissions from Camellia oleifera field soil Environ Sci Pollut Res 2019 264070ndash4077 [CrossRef]
33 Zhou X Sun H Pumpanen J Sietiouml O-M Heinonsalo J Koumlster K Berninger F The impact of wildfire on microbial CNPstoichiometry and the fungal-to-bacterial ratio in permafrost soil Biogeochemistry 2010 142 1ndash17 [CrossRef]
34 Huang J Lin T-C Xiong D Yang Z Liu X Chen G Xie J Li Y Yang Y Organic carbon mineralization in soils of anatural forest and a forest plantation of southeastern China Geoderma 2019 344 119ndash126 [CrossRef]
35 Phillips JM Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapidassessment of infection Trans Br Mycol Soc 1970 55 158ndash161 [CrossRef]
36 Mcgonigle TP Miller MH Evans DG Fairchild GL Swan JA A new method which gives an objective measure ofcolonization of roots by vesicularmdashArbuscular mycorrhizal fungi New Phytol 1990 115 495ndash501 [CrossRef]
37 Jakobsen I Abbott L Robson AD External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifoliumsubterraneum L Spread of hyphae and phosphorus inflow into roots New Phytol 1992 120 371ndash380 [CrossRef]
38 Brundrett MC Mycorrhizas in Natural Ecosystems Adv Ecol Res 1991 21 171ndash31339 Kruumlger M Kruumlger C Walker C Stockinger H Schuumlszligler A Phylogenetic reference data for systematics and phylotaxonomy of
arbuscular mycorrhizal fungi from phylum to species level New Phytol 2011 193 970ndash984 [CrossRef]40 Oehl F Sieverding E Palenzuela J Ineichen K Silva G Advances in Glomeromycota taxonomy and classification IMA
Fungus 2011 2 191ndash199 [CrossRef]41 Wang J Wang GG Zhang B Yuan Z Fu Z Yuan Y Zhu L Ma S Zhang J Arbuscular Mycorrhizal Fungi Associated
with Tree Species in a Planted Forest of Eastern China Forests 2019 10 424 [CrossRef]42 Shannon CE Weaver W The Mathematical Theory of Communication The University of Illinois Press Urbana IL USA 194943 Simpson EH Measurement of Diversity Nature 1949 163 688 [CrossRef]44 Ma X Zhu B Nie Y Liu Y Kuzyakov Y Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen
deposition A meta-analysis Soil Biol Biochem 2021 163 108418 [CrossRef]45 Johnson NC Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytol
2010 185 631ndash647 [CrossRef]46 Avolio ML Koerner S La Pierre KJ Wilcox K Wilson GWT Smith MD Collins S Changes in plant community
composition not diversity during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrassprairie J Ecol 2015 102 1649ndash1660 [CrossRef]
47 Isbell F Reich PB Tilman D Hobbie SE Polasky S Binder S Nutrient enrichment biodiversity loss and consequentdeclines in ecosystem productivity Proc Natl Acad Sci USA 2013 110 11911ndash11916 [CrossRef]
48 Egerton-Warburton LM Johnson NC Allen EB Mycorrhizal community dynamics following nitrogen fertilization Across-site test in five grasslands Ecol Monogr 2007 77 527ndash544 [CrossRef]
49 Jansa J Mozafar A Kuhn G Anken T Ruh R Sanders IR Frossard EJEA Soil tillage affects the community structure ofmycorrhizal dungi in maize roots Ecol Appl 2003 13 1164ndash1176 [CrossRef]
50 Campos PDS Borie F Cornejo P Lopez-Raez JA Loacutepez-Garciacutea JA Seguel A Phosphorus Acquisition Efficiency Relatedto Root Traits Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping Front Plant Sci 2018 9 752 [CrossRef]
51 Loacutepez-Garciacutea Aacute Varela-Cervero S Vasar M Oumlpik M Barea JM Azcoacuten-Aguilar C Plant traits determine the phylogeneticstructure of arbuscular mycorrhizal fungal communities Mol Ecol 2017 26 6948ndash6959 [CrossRef]
52 Oehl F Sieverding E Ineichen K Ris E Boller T Wiemken A Community structure of arbuscular mycorrhizal fungiat different soil depths in extensively and intensively managed agroecosystems New Phytol 2005 165 273ndash283 [CrossRef][PubMed]
53 Jamiołkowska A Ksiezniak A Gałazka A Hetman B Kopacki M Skwaryło-Bednarz B Impact of abiotic factors ondevelopment of the community of arbuscular mycorrhizal fungi in the soil A Review Int Agrophys 2018 32 133ndash140 [CrossRef]
Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-

Forests 2022 13 979 14 of 14
54 Tedone L Ruta C De Cillis F De Mastro G Effects of Septoglomus viscosum inoculation on biomass yield and steviolglycoside concentration of some Stevia rebaudiana chemotypes Sci Hortic 2020 262 109026 [CrossRef]
55 Todeschini V AitLahmidi N Mazzucco E Marsano F Gosetti F Robotti E Bona E Massa N Bonneau L Marengo Eet al Impact of Beneficial Microorganisms on Strawberry Growth Fruit Production Nutritional Quality and Volatilome FrontPlant Sci 2018 9 1611 [CrossRef] [PubMed]
56 Alguacil MDM Torrecillas E Garciacutea-Orenes F Roldan A Changes in the composition and diversity of AMF communitiesmediated by management practices in a Mediterranean soil are related with increases in soil biological activity Soil Biol Biochem2014 76 34ndash44 [CrossRef]
57 Borriello R Lumini E Girlanda M Bonfante P Bianciotto V Effects of different management practices on arbuscularmycorrhizal fungal diversity in maize fields by a molecular approach Biol Fertil Soils 2012 48 911ndash922 [CrossRef]
58 Avio L Castaldini M Fabiani A Bedini S Sbrana C Turrini A Giovannetti M Impact of nitrogen fertilization and soiltillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem Soil Biol Biochem 2013 67 285ndash294[CrossRef]
59 Karasawa T Hodge A Fitter AH Growth respiration and nutrient acquisition by the arbuscular mycorrhizal fungus Glomusmosseae and its host plant Plantago lanceolata in cooled soil Plant Cell Environ 2012 35 819ndash828 [CrossRef]
60 Cappellazzo G Lanfranco L Fitz M Wipf D Bonfante P Characterization of an Amino Acid Permease from the Endomycor-rhizal Fungus Glomus mosseae Plant Physiol 2008 147 429ndash437 [CrossRef]
61 Zhang M Shi Z Yang M Lu S Cao L Wang X Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi atDifferent Elevations in Mt Taibai of Qinling Mountain Front Microbiol 2021 12 609386 [CrossRef]
62 Shi G Yao B Liu Y Jiang S Wang W Pan J Zhao X Feng H Zhou H The phylogenetic structure of AMF communitiesshifts in response to gradient warming with and without winter grazing on the QinghaindashTibet Plateau Appl Soil Ecol 2017 12131ndash40 [CrossRef]
63 Panneerselvam P Kumar U Senapati A Parameswaran C Anandan A Kumar A Jahan A Padhy SR Nayak AKInfluence of elevated CO2 on arbuscular mycorrhizal fungal community elucidated using Illumina MiSeq platform in sub-humidtropical paddy soil Appl Soil Ecol 2019 145 103344 [CrossRef]
64 Neuenkamp L Zobel M Koorem K Jairus T Davison J Oumlpik M Vasar M Moora M Light availability and light demandof plants shape the arbuscular mycorrhizal fungal communities in their roots Ecol Lett 2021 24 426ndash437 [CrossRef] [PubMed]
65 Bunn R Lekberg Y Zabinski C Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants Ecology2009 90 1378ndash1388 [CrossRef] [PubMed]
66 Wang J Zhang J Wang C Ren G Yang Y Wang D Precipitation exerts a strong influence on arbuscular mycorrhizal fungicommunity and network complexity in a semiarid steppe ecosystem Eur J Soil Biol 2021 102 103268 [CrossRef]
67 Chen Y-L Xu Z-W Xu T-L Veresoglou SD Yang G Chen B Nitrogen deposition and precipitation induced phylogeneticclustering of arbuscular mycorrhizal fungal communities Soil Biol Biochem 2017 115 233ndash242 [CrossRef]
68 Li X Qi Z Yu X Xu M Liu Z Du G Yang Y Soil pH drives the phylogenetic clustering of the arbuscular mycorrhizalfungal community across subtropical and tropical pepper fields of China Appl Soil Ecol 2021 165 103978 [CrossRef]
69 Antunes PM Lehmann A Hart MM Baumecker M Rillig MC Long-term effects of soil nutrient deficiency on arbuscularmycorrhizal communities Funct Ecol 2012 26 532ndash540 [CrossRef]
70 Zhang J Quan C Ma L Chu G Liu Z Tang X Plant community and soil properties drive arbuscular mycorrhizal fungaldiversity A case study in tropical forests Soil Ecol Lett 2021 3 52ndash62 [CrossRef]
71 Gryndler M Hršelovaacute H Sudovaacute R Gryndlerovaacute H Rezaacutecovaacute V Merhautovaacute V Hyphal growth and mycorrhiza formationby the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances Mycorrhiza 2005 15 483ndash488[CrossRef]
72 Oehl F Sieverding E Ineichen K Maumlder P Wiemken A Boller T Distinct sporulation dynamics of arbuscular mycorrhizalfungal communities from different agroecosystems in long-term microcosms Agric Ecosyst Environ 2009 134 257ndash268[CrossRef]
73 Mosbah M Philippe DL Mohamed M Molecular identification of arbuscular mycorrhizal fungal spores associated to therhizosphere of Retama raetam in Tunisia Soil Sci Plant Nutr 2018 64 335ndash341 [CrossRef]
74 Wolf J Johnson NC Rowland DL Reich PB Elevated CO2 and plant species richness impact arbuscular mycorrhizal fungalspore communities New Phytol 2003 157 579ndash588 [CrossRef] [PubMed]
- Introduction
- Materials and Methods
-
- Study Sites and Sampling
- Soil Properties Analysis
- AMF Colonization and Extraradical Hphae Length Density
- AMF Spore Separation and Identification
- AMF Community Parameters and Statistical Analysis
-
- Results
-
- Effects of N Addition on Soil AMF Colonization Spore Density and External Hyphal Length Density
- Effects of N Addition on Soil AMF Community Composition
- Effects of N addition on Soil Nutrients
- Relationship between Soil Properties and AMF
-
- Discussion
- Conclusions
- Appendix A
- References
-