new pterosaur specimens from the kem kem beds (upper cretaceous, cenomanian) of morocco
TRANSCRIPT

RivistaItalianadiPaleontologiaeStratigrafia volume117 no.1 pp.149-160 April2011
NEW PTEROSAUR SPECIMENS FROM THE KEM KEM BEDS (UPPER CRETACEOUS, CENOMANIAN) OF MOROCCO
TAISSARODRIGUES1,ALEXANDERW.A.KELLNER1,BRYNJ.MADER2&DALEA.RUSSELL3
Received: October 11, 2010; accepted: February 1st, 2011
1 SetordePaleovertebrados,Dept.GeologiaePaleontologia,MuseuNacional/UFRJ.QuintadaBoaVistas/n,SãoCristóvão,20940-040,RiodeJaneiro,Brazil.E-mail:[email protected],[email protected]
2 QueensboroughCommunityCollege,222-0556thAvenueBayside,NY11364,USA.E-mail:[email protected] NorthCarolinaStateUniversity,Dept.ofMarine,EarthandAtmosphericSciences,JordanHall5145,Box8208,Raleigh,NC27695,
USA.E-mail:[email protected]
Key words:Pterosauria,Azhdarchidae,Morocco,Cretaceous,Cenomanian,KemKembeds.
Abstract. AlthoughpterosaursfromAfricaarestillrare,inre-centyearsseveralspecimenshavebeendescribedfromtheKemKembeds(UpperCretaceous,Cenomanian)ofMorocco.Herewedescribefour additional specimens from this informal lithostratigraphic unit:ajawfragment,twomid-cervicalvertebrae,andahumerus.Allthesespecimensshowthree-dimensionalpreservation,differingmuchfromtheflatconditionfoundinmostpterosaurmaterial.Thevertebraeareparticularly well preserved, and allow accurate observations on thepneumatizationoftheneuralarch.Basedoncomparablematerial,weshow that at least two edentulous pterosaur species were present inthis informal lithostratigraphicunit, thusaddingtothegrowingevi-denceofconsiderablepterosaurdiversityinnorthwesternAfricadur-ingthe“middle”Cretaceous.Sofar,theKemKembedshavethemostdiversepterosaurfaunainthiscontinent,withthepresenceofanhan-guerids,azhdarchids,pteranodontids,andtapejarids.
Riassunto. SebbeneglipterosauririnvenutiinAfricasianoan-corarari,negliultimiannidiversiesemplarisonostatidescrittidaico-siddettiKemKembedsdelMarocco(CretacicoSuperiore,Cenoma-niano).Nell’articolodescriviamoquattronuovirepertiprovenientidaquestaunità litostratigrafica informale:un frammentodimandibola,due vertebre medio-cervicali e un omero. Tutti questi esemplari of-frono una conservazione tridimensionale, differendo in questo dallecondizionidiappiattimentopropriedimoltirepertidipterosauri.Levertebre sono particolarmente ben conservate e consentono una ac-curata osservazione della pneumatizzazione dell’arco neurale. Sullabasedimaterialediconfrontonoimostriamochealmenoduespecieedentule di pterosauri sono presenti in questa unità litostratigraficainformale.Ciò incrementa l’evidenzadiunaconsiderevolediversitàtraglipterosaurichepopolaronol’Africanordoccidentaleduranteil“Cretacicomedio’’.IKemKembedscontengonolafaunaapterosauripiùdiversificatainquestocontinente,conlapresenzadianhangueridi,azhdarchidi,pteranodontidietapejaridi.
Introduction
African pterosaurs are still poorly known, lim-itedtorelativelyfewincompleteskeletonsandisolatedremains(Swinton1948;Monteilletetal.1982;Unwin& Heinrich 1999; Benton et al. 2000; Dalla Vecchiaetal.2001;Blackburn2002;DalSasso&Pasini2003;Ntamak-Nida et al. 2006; Blackbeard & Yates 2007;Kellneretal.2007;Costa&Kellner2009;Ibrahimetal.2010).Moroccorepresentsseveraloftherecentfindsofpterosaurfossils.
Sigogneau-Russell etal. (1998) first reportedonsome pterosaur material from a Lower CretaceouscalcareouslensinthelocalityofAnoual.Thesespeci-mens, nearly 300 isolated teeth, were described byKnoll(2000),whoidentifiedthemasbelongingtoor-nithocheiridsandgnathosaurines,thelatterquestionedbyRodrigues&Kellner (2010)whopointedout thatat least someof thegnathosaurin teethcouldactuallybelongtotheAnhangueridaeorcloselyrelatedtaxa.
Another Moroccan pterosaur occurrence is thesequence of cervical vertebrae from the Maastrich-tianoftheOuladAbdounPhosphaticBasinthatrep-resents the azhdarchid Phosphatodraco mauritanicusPereda-Suberbiola, Bardet, Jouve, Iarochène, Bouya&Amaghzaz,2003(seePereda-Suberbiolaetal.2003;Kellner2010).
Notwithstandingtheseoccurrences,mostptero-saur material comes from the Cenomanian Ifezouaneand Aoufous formations (informally known as theKemKembeds)(Serenoetal.1996;Cavinetal.2010).

150 Rodrigues T., Kellner A.W.A., Mader B.J. & Russell D.A.
Sofar,thesedepositshaveyieldedisolatedanhangueridteeth(Kellner&Mader1997;Wellnhofer&Buffetaut1999),isolatedazhdarchidvertebrae(Kellner&Mader1996; Rodrigues et al. 2006; Ibrahim et al. 2010) andfragmentaryjawsreferredtotheAnhangueridae,Azh-darchidae,Pteranodontidae,andTapejaridae(Mader&Kellner1999;Wellnhofer&Buffetaut1999;Kellneretal.2007;Ibrahimetal.2010),includingtwodescribedspecies: the anhanguerid Siroccopteryx moroccensisMader & Kellner, 1999 (see Mader & Kellner 1999;Rodrigues&Kellner2008;butseeIbrahimetal.2010for a different classification of this species) and theazhdarchid Alanqa saharica Ibrahim, Unwin, Martill,Baidder&Zouhri,2010(seeIbrahimetal.2010).Thepterosaurspecimens,althoughrelativelycommon,tendtobefragmentaryduetothehighenergydepositionalenvironment of both these units (Cavin et al. 2010).Herewedescribefourspecimensthathavebeenonlybriefly mentioned in the literature (Kellner & Mader1996;Rodrigueset al.2006), adding to thebetterun-derstandingoftheMoroccanpterosaurfauna.
Institutional abbreviations: BSP,BayerischeStaatssammlungfür Paläontologie und Geologie, Munich, Germany; CCMGE, Mu-seum of Central Scientific Research Institute for Geological Explo-ration,UniversityofSt.Petersburg,SaintPetersburg,Russia;CMN,Canadian Museum of Nature, Ottawa, Canada; FSAC, Faculté desSciences Ain Chock, Université Hassan II, Casablanca, Morocco;LINHM, Long Island Natural History Museum, New York City,USA; MDM, Mifune Dinosaur Museum, Mifune, Japan; MGUV,MuseodelDepartamentodeGeología/UniversidaddeValencia,Va-lencia,Spain;MN,MuseuNacional/UniversidadeFederaldoRiodeJaneiro,Riode Janeiro,Brazil;MTM,MagyarTermészettudományiMúzeum,Budapest,Hungary;NHMUK,NaturalHistoryMuseum,London,England;SMNK,StaatlichesMuseumfürNaturkundeKarl-sruhe, Karlsruhe, Germany; and TMM, Texas Memorial Museum,Austin,USA.
Anatomical abbreviations: cap, capitulumor radial condyleof thehumerus;co,cotyle;con,condyle;dpc,deltopectoralcrestofthe humerus; ect, ectepicondyle of the humerus; ent, entepicondyleofthehumerus;fo, foramen;fopn, foramenpneumaticum;hyp,hy-papophysis;mus,musclescar;nc,neuralcanal;ns,neuralspine;poex,postexapophysis; poz, postzygapophysis; prz, prezygapophysis; ?ri,possiblevestigial cervical rib;rid, ridge; sul, sulcus; tro, trochleaorulnar condyleof thehumerus; tub, tubercle; andulc,ulnar crestofthehumerus.
Geological Settings
Theso-called‘ContinentalIntercalaire’compris-esextensivedeltaicandbrackishdepositsintheLowerCretaceous of North Africa (Kilian 1931; Lapparent1960;Cavinetal.2010).InMorocco,itisknowninfor-mallyastheKemKembeds(Serenoetal.1996),whosefauna,paleoenvironmentandpaleogeographywerere-centlyreviewedbyCavinetal.(2010).TheageoftheKemKembedsisnoteasytobeestablishedduetothelackofmarine intercalations,butbytheconstraintof
the overlying Akrabou Formation, whose lower partcontains ammonoids characteristic of the lower partoftheupperCenomanian, andbycomparisontootherNorthAfricanfaunasofthe‘ContinentalIntercalaire’,ithasbeenconcludedthattheyweredepositedduringtheCenomanian(Serenoetal.1996;Cavinetal.2010).
Cavinetal.(2010)emphasizedthattheKemKembeds actually include two formations: the IfezouaneFormationatthebottomandtheAoufousFormationat the top, as pointed out before (e.g., Ettachfini &Andreu2004).TheIfezouaneFormationiscomposedmostly of sandstone, while the Aoufous Formationpresents marls with intercalations of sandstone andmicroconglomerates (Cavin et al. 2010). As most ofthefossilsfromtheKemKembedswerecollectedbycommercialdealers,itisnotpossibletodeterminetheirprovenience,both in termsof localityand lithostrati-graphical unit (e.g., McGowan & Dyke 2009). Thus,theKemKembedspaleovertebratefaunais, infact,acompoundassemblage,includingterrestrial,freshwaterandbrackishvertebrates(Cavinetal.2010).
Systematic Paleontology
PterosauriaKaup,1834
PterodactyloideaPlieninger,1901
DsungaripteroideasensuKellner2003
Dsungaripteroideaindet.
Material:?Lowerjaw(CMN50859)
The specimen CMN 50859 consists of the an-terior fragment (preserved length is 43 mm) from anedentulousjaw,possiblyamandible(Fig.1).Ittapersanteriorlyandshowsthemarginoppositetotheocclu-salsurfacerounded.Incross-section,thedorsalmarginisconcaveandthelateraledgesareelevated.Thelateralsidesareslightlyconvex,andacentralovallumencanalsobeobserved.There isnomedianridgeorgrooveontheocclusalsurface.Smallslit-likeforaminaarealsopresentontheocclusalandlateralsurfaces,suggestingthatthisportionofthejawwaswellvascularizedorin-nervated. The ventral margin lacks any evidence of asagittalcrestorkeel.
Proper identification of such a small fragmentcanonlybemadetentatively.TheDsungaripteridae,acladewhichcomprisespterosaursthatlackteethonthetip of the jaws (Young 1964, 1973), can be ruled outsince thecross-section isnotcircular inCMN50859.Among tapejarids sensu Kellner (2003), all species ofthe Tapejarinae possess dentary crests (Wellnhofer &Kellner1991;Lietal.2003;Wang&Zhou2003;Lü&Yuan2005;Lüetal.2006;Kellner&Campos2007),a

New pterosaur specimens from the Kem Kem beds (Upper Cretaceous, Cenomanian) of Morocco 151
featureabsentinthepreservedportionofCMN50859.ThenewmaterialalsodiffersfromthethalassodrominThalassodromeus sethi Kellner & Campos, 2002 bythelackofthedistinctivesharpdorsaledge(Kellner&Campos2002).
The tips of the jaws in the tapejarid genus Tu-puxuara,inthePteranodontidaesensuKellner(2003),Nyctosauridae,Chaoyangopteridae,andinmostoftheazhdarchids,aresimilartoCMN50859andwecannotproperlyassignthematerialtoanyofthese.ComparedwithotherMoroccantoothlessjaws,thepreservedpor-tionofCMN50859sharestheconcavedorsalsurfacewithBSP1993IX338(Wellnhofer&Buffetaut1999),here regarded as a lower jaw (contra Ibrahim et al.2010),andMN7054-V(Kellneretal.2007),tentativelyreferredtothePteranodontoidea(sensuKellner2003)(Wellnhofer & Buffetaut 1999; Kellner et al. 2007) orto the Azhdarchidae (BSP 1993 IX 338; Ibrahim etal.2010).CMN50859lacksthelowsagittalcrestandhasaquitedistinctcross-sectionfromtherecentlyde-scribedazhdarchidAlanqa saharica(seeIbrahimetal.2010),indicatingthatasecondedentulousspecieswaspresent intheKemKembedscompoundassemblage.Ascomparisonsare limitedandCMN50859alsocannotbeundoubtedlyidentifiedasalowerjaw,itisherereferred in the Dsungaripteroidea, a clade which in-cludesalltoothlesspterosaursbesidestheAnhangueri-dae,theIstiodactylidaeandtheDsungaripteridae(Kel-lner2003,Wangetal.2009;butseeUnwin2003foradifferenthypothesisofpterosaurphylogeny).
AzhdarchidaeNessov,1984
Azhdarchidaeindet.
Material: Mid-cervical vertebrae (CMN 50801 and LINHM014)
The twocervicalvertebraedescribedhere showsimilar basic morphology to one another. Both areelongatedandclearlybelongtothemiddleseries(be-tweencervicalvertebrae4and7).
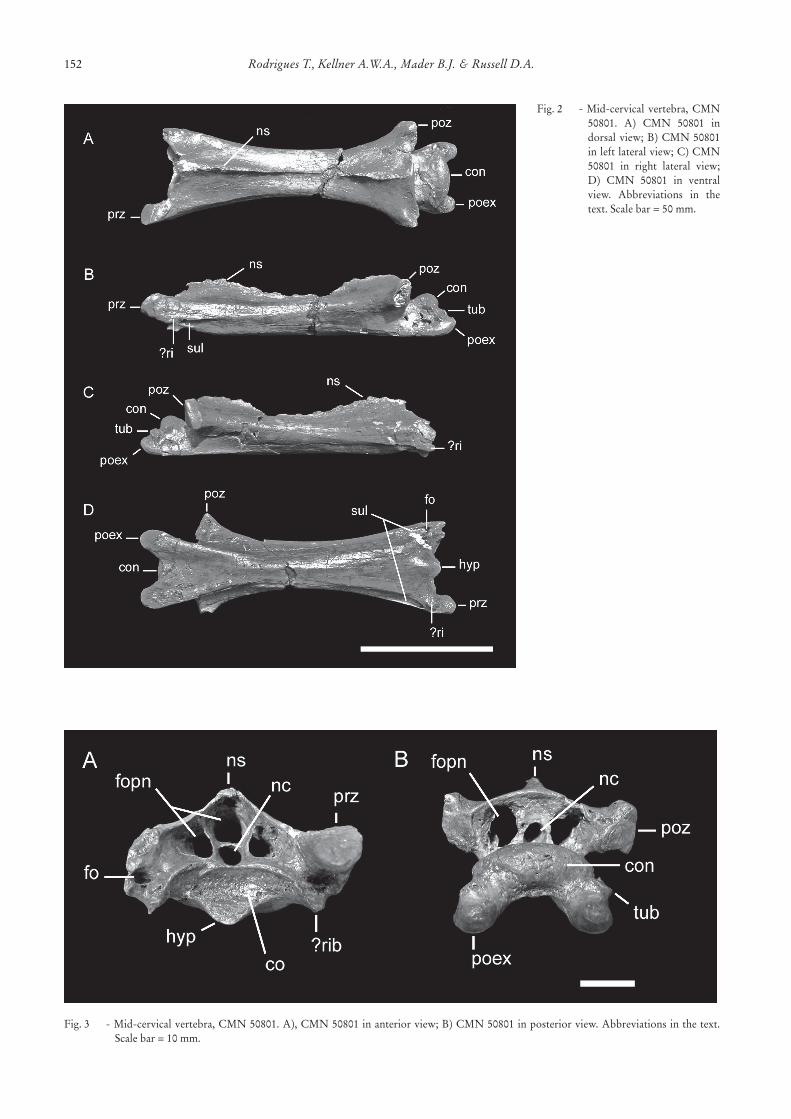
CMN 50801 (Figs 2, 3) is well preserved, withnomajor signsof compression.The centrum is elon-gate(119mmlong,18mmminimumwidth)andsome-what flattened dorsoventrally. No lateral pneumaticforamina are present. Two well-defined longitudinalsulciareobservedon theanteriorpart,at theventro-lateral surfaces. The anterior cotyle is saddle-shaped,bearingashortandblunthypapophysisontheventralsurface. The posterior condyle is expanded, sub-ovalinshape,somewhatflatteneddorsoventrally,andhasashallowdepressionunderneath.Theneuralspineislowand elongated anteroposteriorly, extending along thewhole lengthof theneural arch. In anterior andpos-
teriorviews,twoproportionallylarge,ovalpneumaticforaminaarepresentlateraltotheneuralcanal.Athirdpneumaticforamenisalsopresentontheanteriorside,locateddorsallytotheneuralcanal,butisabsentpos-teriorly.Thepresentspecimendoesnothavetherightprezygapophysis preserved. The articular facet of theleft prezygapophysis is broad, dorsally flattened, andorientedanteroventrally.Asmallossificationispresentventraltotheprezygapophysisandlateraltothecen-trum,whichappearstobethevestigeofthecervicalrib(eitherafusedcervicalriboranapophysisforthear-ticulationofanunfusedone),includingaforamen(fo-ramentransversarium).Ashallowsulcusstartsat thisforamen and ends at the middle part of the centrum.Thepostexapophysesarehorn-likeandslightlydiver-gentfromeachother.Eachbearssmalldorsolateraltu-bercles.Thearticularsurfacesofthepostzygapophysesarewide,flatanddirectedlateroposteriorly,somewhatinclineddownwards.
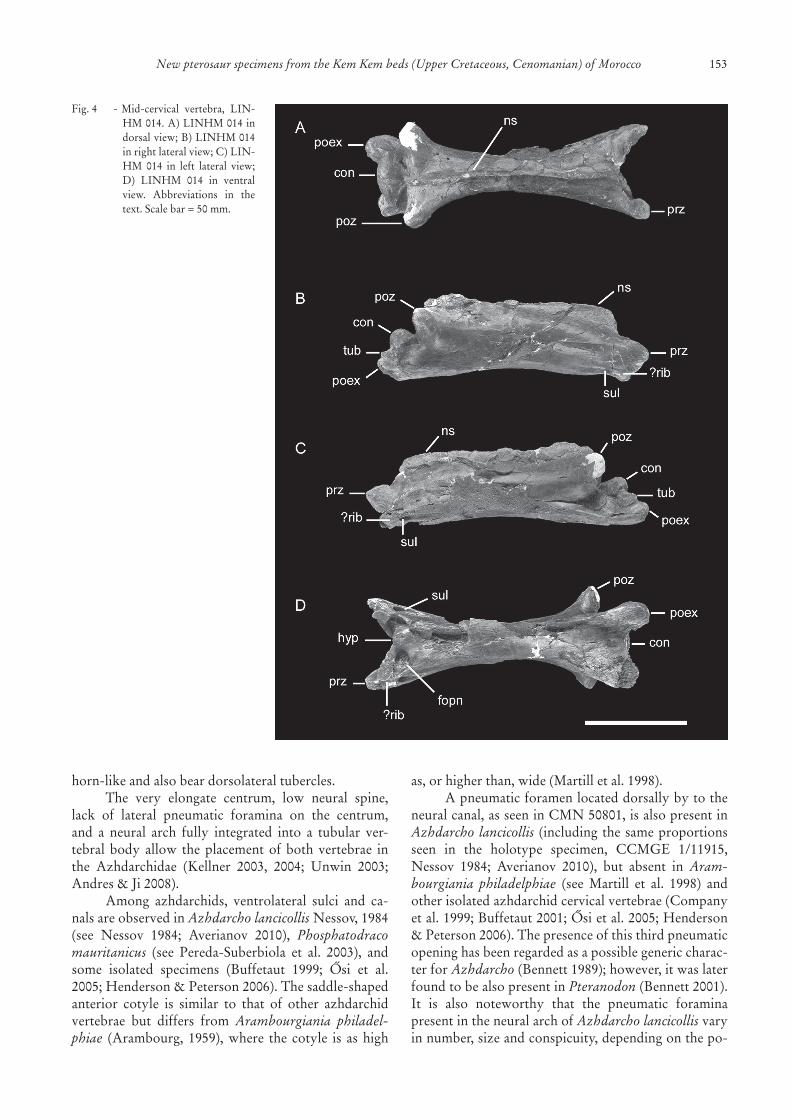
The vertebra LINHM 014 (Fig. 4) is also well-preserved,withaveryelongatecentrum(182mmlong,32 mm minimum width) that lacks lateral pneumaticforamina. Longitudinal, ventrolateral sulci are alsoclearly visible in this specimen, as is the ossificationregardedasthevestigeofthecervicalribs.Onthean-teriorside,ablunthypapophysisispresent.Ventrally,there are two large, round pneumatic openings, eachonelocatedlaterallytothehypapophysis.Beingpairedstructures, they can be considered original morpho-logicalfeaturesandnotartifactsderivedfromprepara-tion,pathologicalortaphonomicprocesses.Theneuralspine is lowand thecondyle laterally expanded.Twolarge pneumatic foramina are also observed lateral totheneuralcanal,inbothanteriorandposteriorviews.The articular region of the left prezygapophysis ismissing.Aprominentforamenispresentbeneatheachprezygapophysisandcontinuedasaventrolateralsul-cus,asseeninCMN50801.Thepostexapophysesare
Fig.1 -?Mandible,CMN50859.A)CMN50859 indorsalview;B)CMN50859inventralview;C)CMN50859inleftlat-eral view; D) CMN 50859 in proximal view. Scale bar =10mm.
152 Rodrigues T., Kellner A.W.A., Mader B.J. & Russell D.A.
Fig.2 -Mid-cervicalvertebra,CMN50801. A) CMN 50801 indorsalview;B)CMN50801inleftlateralview;C)CMN50801 in right lateral view;D) CMN 50801 in ventralview. Abbreviations in thetext.Scalebar=50mm.
Fig.3 -Mid-cervicalvertebra,CMN50801.A),CMN50801inanteriorview;B)CMN50801inposteriorview.Abbreviationsinthetext.Scalebar=10mm.
New pterosaur specimens from the Kem Kem beds (Upper Cretaceous, Cenomanian) of Morocco 153
horn-likeandalsobeardorsolateraltubercles.The very elongate centrum, low neural spine,
lack of lateral pneumatic foramina on the centrum,and a neural arch fully integrated into a tubular ver-tebral body allow the placement of both vertebrae inthe Azhdarchidae (Kellner 2003, 2004; Unwin 2003;Andres&Ji2008).
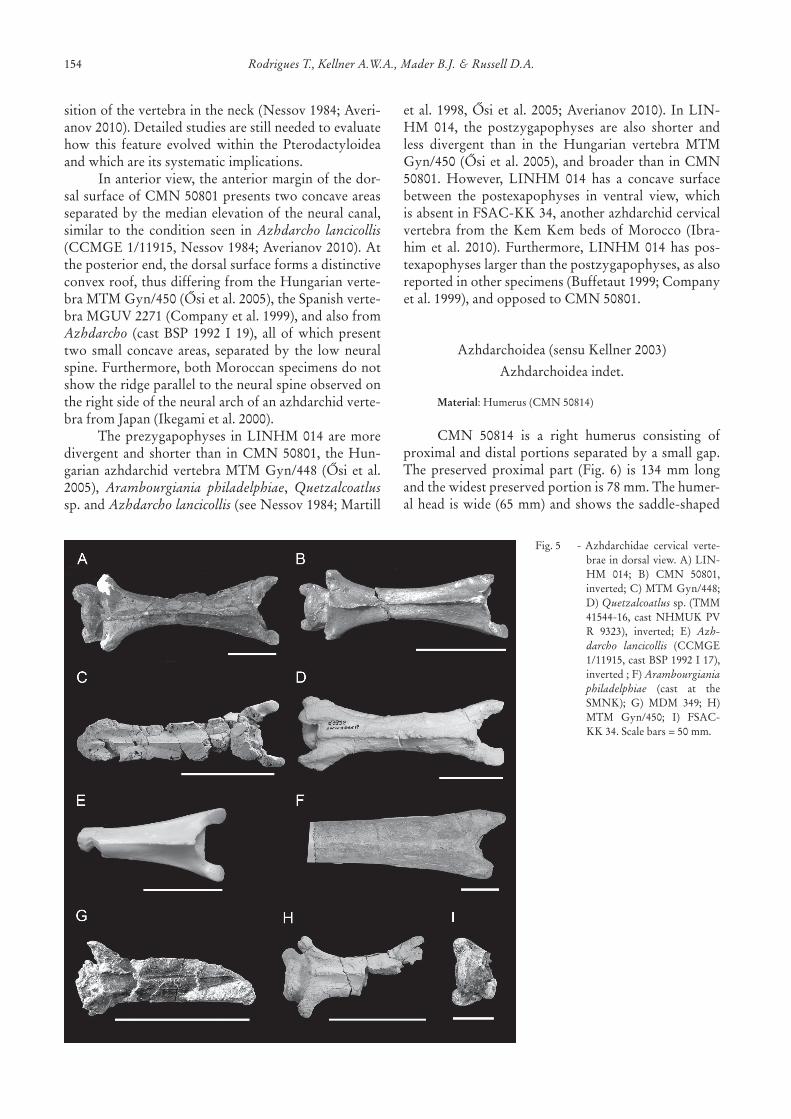
Among azhdarchids, ventrolateral sulci and ca-nalsareobservedinAzhdarcho lancicollisNessov,1984(see Nessov 1984; Averianov 2010), Phosphatodraco mauritanicus (see Pereda-Suberbiola et al. 2003), andsome isolated specimens (Buffetaut 1999; Osi et al.2005;Henderson&Peterson2006).Thesaddle-shapedanterior cotyle is similar to that of other azhdarchidvertebrae but differs from Arambourgiania philadel-phiae (Arambourg, 1959),where the cotyle is ashigh
as,orhigherthan,wide(Martilletal.1998).Apneumaticforamenlocateddorsallybytothe
neuralcanal,asseeninCMN50801,isalsopresentinAzhdarcho lancicollis (includingthesameproportionsseen in the holotype specimen, CCMGE 1/11915,Nessov 1984; Averianov 2010), but absent in Aram-bourgiania philadelphiae (see Martill et al. 1998) andotherisolatedazhdarchidcervicalvertebrae(Companyetal.1999;Buffetaut2001;Osietal.2005;Henderson&Peterson2006).Thepresenceofthisthirdpneumaticopeninghasbeenregardedasapossiblegenericcharac-terforAzhdarcho(Bennett1989);however,itwaslaterfoundtobealsopresentinPteranodon (Bennett2001).It is also noteworthy that the pneumatic foraminapresentintheneuralarchofAzhdarcho lancicollisvaryinnumber,sizeandconspicuity,dependingonthepo-
Fig.4 -Mid-cervical vertebra, LIN-HM014.A)LINHM014indorsalview;B)LINHM014inrightlateralview;C)LIN-HM 014 in left lateral view;D) LINHM 014 in ventralview. Abbreviations in thetext.Scalebar=50mm.
154 Rodrigues T., Kellner A.W.A., Mader B.J. & Russell D.A.
sitionofthevertebraintheneck(Nessov1984;Averi-anov2010).Detailedstudiesarestillneededtoevaluatehow this feature evolved within the Pterodactyloideaandwhichareitssystematicimplications.
Inanteriorview,theanteriormarginofthedor-salsurfaceofCMN50801presentstwoconcaveareasseparatedbythemedianelevationoftheneuralcanal,similar to the condition seen in Azhdarcho lancicollis(CCMGE1/11915,Nessov1984;Averianov2010).Attheposteriorend,thedorsalsurfaceformsadistinctiveconvexroof,thusdifferingfromtheHungarianverte-braMTMGyn/450(Osietal.2005),theSpanishverte-braMGUV2271(Companyetal.1999),andalsofromAzhdarcho (cast BSP 1992 I 19), all of which presenttwo small concave areas, separatedby the lowneuralspine.Furthermore,bothMoroccanspecimensdonotshowtheridgeparalleltotheneuralspineobservedontherightsideoftheneuralarchofanazhdarchidverte-brafromJapan(Ikegamietal.2000).
Theprezygapophyses inLINHM014aremoredivergent and shorter than inCMN50801, theHun-garianazhdarchidvertebraMTMGyn/448 (Osi etal.2005), Arambourgiania philadelphiae, Quetzalcoatlussp.andAzhdarcho lancicollis(seeNessov1984;Martill
etal.1998,Osietal.2005;Averianov2010). InLIN-HM 014, the postzygapophyses are also shorter andless divergent than in the Hungarian vertebra MTMGyn/450(Osietal.2005),andbroaderthaninCMN50801. However, LINHM 014 has a concave surfacebetween the postexapophyses in ventral view, whichisabsentinFSAC-KK34,anotherazhdarchidcervicalvertebra from the Kem Kem beds of Morocco (Ibra-him et al. 2010). Furthermore, LINHM 014 has pos-texapophyseslargerthanthepostzygapophyses,asalsoreportedinotherspecimens(Buffetaut1999;Companyetal.1999),andopposedtoCMN50801.
Azhdarchoidea(sensuKellner2003)
Azhdarchoideaindet.
Material:Humerus(CMN50814)
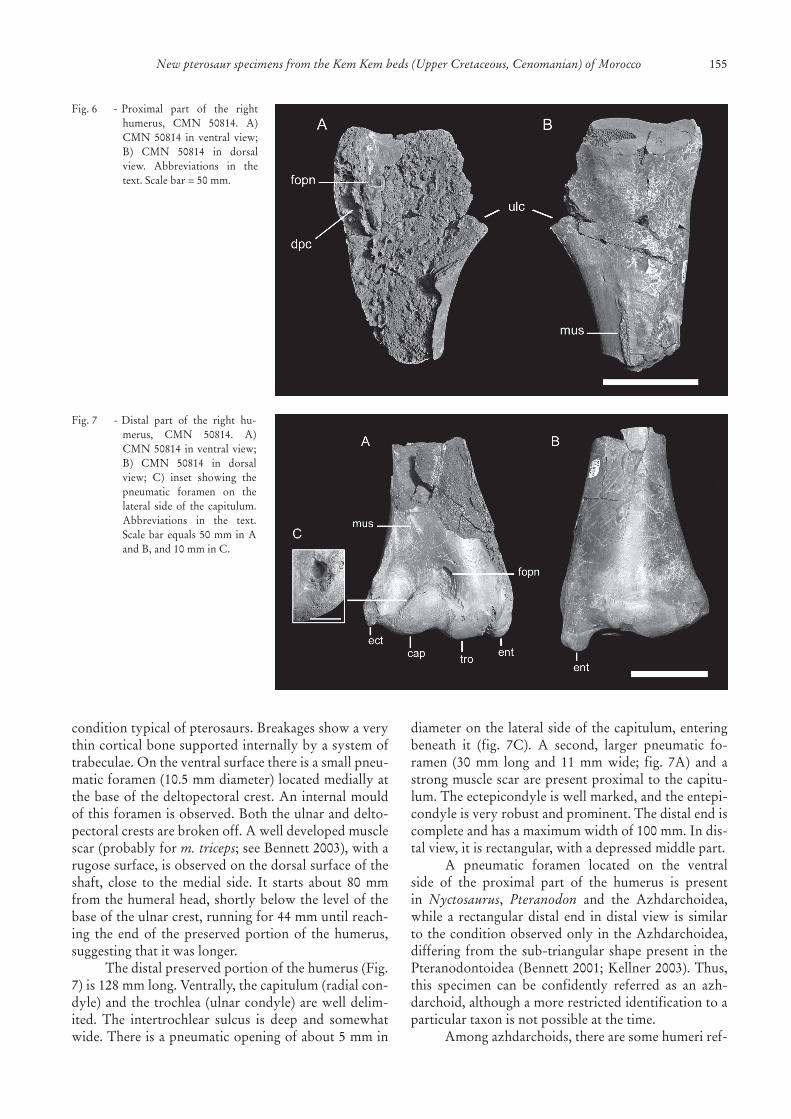
CMN 50814 is a right humerus consisting ofproximalanddistalportionsseparatedbyasmallgap.The preserved proximal part (Fig. 6) is 134 mm longandthewidestpreservedportionis78mm.Thehumer-alheadiswide(65mm)andshowsthesaddle-shaped
Fig.5 -Azhdarchidae cervical verte-braeindorsalview.A)LIN-HM 014; B) CMN 50801,inverted;C)MTMGyn/448;D)Quetzalcoatlussp.(TMM41544-16, castNHMUKPVR 9323), inverted; E) Azh-darcho lancicollis (CCMGE1/11915,castBSP1992I17),inverted;F)Arambourgiania philadelphiae (cast at theSMNK); G) MDM 349; H)MTM Gyn/450; I) FSAC-KK34.Scalebars=50mm.
New pterosaur specimens from the Kem Kem beds (Upper Cretaceous, Cenomanian) of Morocco 155
conditiontypicalofpterosaurs.Breakagesshowaverythincorticalbonesupportedinternallybyasystemoftrabeculae.Ontheventralsurfacethereisasmallpneu-maticforamen(10.5mmdiameter)locatedmediallyatthebaseofthedeltopectoralcrest.Aninternalmouldofthisforamenisobserved.Boththeulnaranddelto-pectoralcrestsarebrokenoff.Awelldevelopedmusclescar(probablyform. triceps;seeBennett2003),witharugosesurface,isobservedonthedorsalsurfaceoftheshaft, close to the medial side. It starts about 80 mmfromthehumeralhead,shortlybelowthelevelofthebaseoftheulnarcrest,runningfor44mmuntilreach-ing the end of the preserved portion of the humerus,suggestingthatitwaslonger.
Thedistalpreservedportionofthehumerus(Fig.7)is128mmlong.Ventrally,thecapitulum(radialcon-dyle)andthetrochlea(ulnarcondyle)arewelldelim-ited. The intertrochlear sulcus is deep and somewhatwide.Thereisapneumaticopeningofabout5mmin
diameteronthelateralsideofthecapitulum,enteringbeneath it (fig. 7C). A second, larger pneumatic fo-ramen (30 mm long and 11 mm wide; fig. 7A) and astrongmusclescararepresentproximaltothecapitu-lum.Theectepicondyleiswellmarked,andtheentepi-condyleisveryrobustandprominent.Thedistalendiscompleteandhasamaximumwidthof100mm.Indis-talview,itisrectangular,withadepressedmiddlepart.
A pneumatic foramen located on the ventralside of the proximal part of the humerus is presentin Nyctosaurus, Pteranodon and the Azhdarchoidea,while a rectangulardistal end indistalview is similartotheconditionobservedonlyintheAzhdarchoidea,differing fromthesub-triangular shapepresent in thePteranodontoidea(Bennett2001;Kellner2003).Thus,this specimen can be confidently referred as an azh-darchoid,althoughamorerestrictedidentificationtoaparticulartaxonisnotpossibleatthetime.
Amongazhdarchoids,therearesomehumeriref-
Fig.6 -Proximal part of the righthumerus, CMN 50814. A)CMN 50814 in ventral view;B) CMN 50814 in dorsalview. Abbreviations in thetext.Scalebar=50mm.
Fig.7 -Distal part of the right hu-merus, CMN 50814. A)CMN50814inventralview;B) CMN 50814 in dorsalview; C) inset showing thepneumatic foramen on thelateral side of the capitulum.Abbreviations in the text.Scalebarequals50mminAandB,and10mminC.
156 Rodrigues T., Kellner A.W.A., Mader B.J. & Russell D.A.
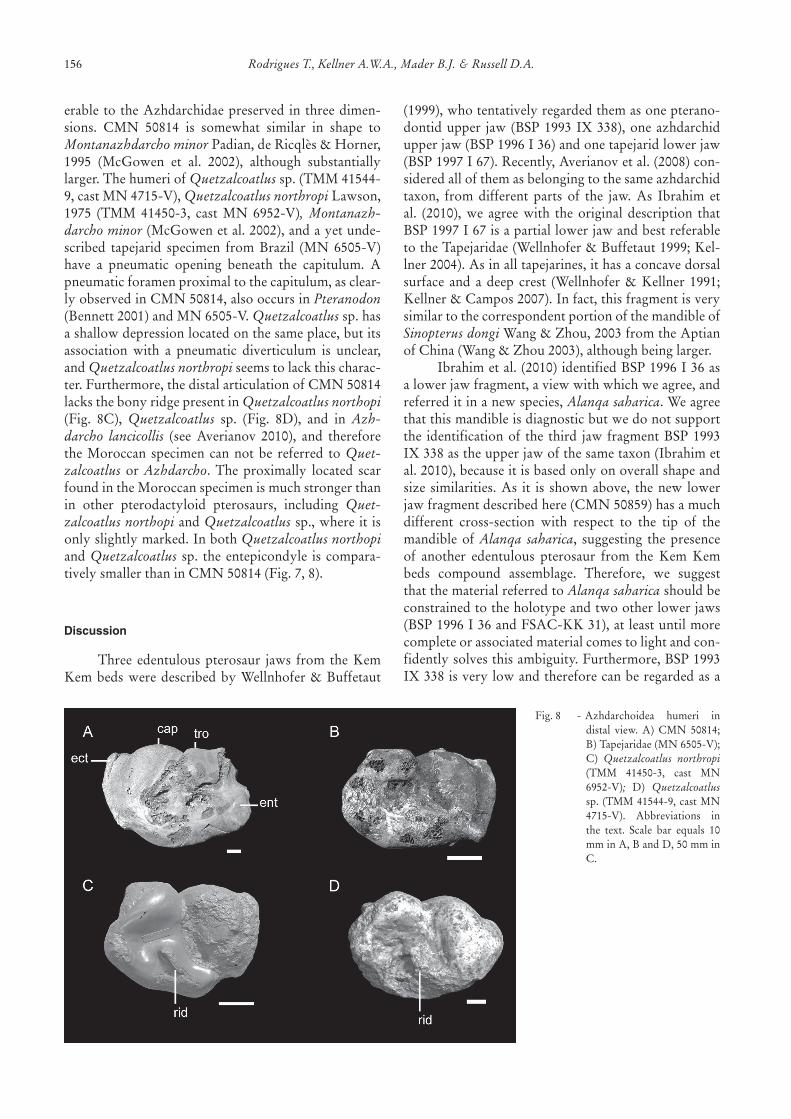
erable to theAzhdarchidaepreserved in threedimen-sions. CMN 50814 is somewhat similar in shape toMontanazhdarcho minor Padian,deRicqlès&Horner,1995 (McGowen et al. 2002), although substantiallylarger.ThehumeriofQuetzalcoatlus sp.(TMM41544-9,castMN4715-V),Quetzalcoatlus northropi Lawson,1975 (TMM 41450-3, cast MN 6952-V), Montanazh-darcho minor(McGowenetal.2002),andayetunde-scribed tapejarid specimen from Brazil (MN 6505-V)have a pneumatic opening beneath the capitulum. Apneumaticforamenproximaltothecapitulum,asclear-lyobservedinCMN50814,alsooccursinPteranodon (Bennett2001)andMN6505-V.Quetzalcoatlussp.hasashallowdepressionlocatedonthesameplace,butitsassociation with a pneumatic diverticulum is unclear,andQuetzalcoatlus northropiseemstolackthischarac-ter.Furthermore,thedistalarticulationofCMN50814lacksthebonyridgepresentinQuetzalcoatlus northopi (Fig. 8C), Quetzalcoatlus sp. (Fig. 8D), and in Azh-darcho lancicollis (see Averianov 2010), and thereforetheMoroccan specimencannotbe referred toQuet-zalcoatlus or Azhdarcho. The proximally located scarfoundintheMoroccanspecimenismuchstrongerthanin other pterodactyloid pterosaurs, including Quet-zalcoatlus northopi andQuetzalcoatlus sp.,whereitisonlyslightlymarked.InbothQuetzalcoatlus northopi andQuetzalcoatlus sp. theentepicondyle iscompara-tivelysmallerthaninCMN50814(Fig.7,8).
Discussion
Three edentulouspterosaur jaws from theKemKembedsweredescribedbyWellnhofer&Buffetaut
(1999),whotentativelyregardedthemasonepterano-dontidupper jaw(BSP1993IX338),oneazhdarchidupperjaw(BSP1996I36)andonetapejaridlowerjaw(BSP1997I67).Recently,Averianovetal.(2008)con-sideredallofthemasbelongingtothesameazhdarchidtaxon, from different parts of the jaw. As Ibrahim etal. (2010), we agree with the original description thatBSP1997I67isapartiallowerjawandbestreferabletotheTapejaridae(Wellnhofer&Buffetaut1999;Kel-lner2004).Asinalltapejarines,ithasaconcavedorsalsurface and a deep crest (Wellnhofer & Kellner 1991;Kellner&Campos2007).Infact,thisfragmentisverysimilartothecorrespondentportionofthemandibleofSinopterus dongiWang&Zhou,2003fromtheAptianofChina(Wang&Zhou2003),althoughbeinglarger.
Ibrahimetal.(2010)identifiedBSP1996I36asalowerjawfragment,aviewwithwhichweagree,andreferreditinanewspecies,Alanqa saharica.Weagreethatthismandibleisdiagnosticbutwedonotsupportthe identificationof the third jaw fragmentBSP1993IX338astheupperjawofthesametaxon(Ibrahimetal.2010),becauseitisbasedonlyonoverallshapeandsize similarities.As it is shownabove, thenew lowerjawfragmentdescribedhere(CMN50859)hasamuchdifferent cross-section with respect to the tip of themandible of Alanqa saharica, suggesting the presenceof another edentulous pterosaur from the Kem Kembeds compound assemblage. Therefore, we suggestthatthematerialreferredtoAlanqa saharicashouldbeconstrainedtotheholotypeandtwootherlowerjaws(BSP1996I36andFSAC-KK31),atleastuntilmorecompleteorassociatedmaterialcomestolightandcon-fidentlysolvesthisambiguity.Furthermore,BSP1993IX338isverylowandthereforecanberegardedasa
Fig.8 -Azhdarchoidea humeri indistal view. A) CMN 50814;B)Tapejaridae(MN6505-V);C) Quetzalcoatlus northropi(TMM 41450-3, cast MN6952-V); D) Quetzalcoatlussp. (TMM41544-9,castMN4715-V). Abbreviations inthe text. Scale bar equals 10mminA,BandD,50mminC.

New pterosaur specimens from the Kem Kem beds (Upper Cretaceous, Cenomanian) of Morocco 157
lowerjaw,sharingtheconcavedorsalsurfaceandlackofasagittalcrestwiththelowerjawofPteranodonandDawndraco(e.g.Bennett2001;Kellner2010).Forthetime being, we agree with the original attribution byWellnhofer & Buffetaut (1999) that BSP 1993 IX 338ismostprobablyaPteranodontidaeorcloselyrelatedtaxon(asalsopointedoutbyCavinetal.2010).
BothvertebraedescribedherearereferredtotheAzhdarchidae. Apart from their overall resemblance,LINHM014islargerandcomparativelylesselongatethanCMN50801.Besides,thepairedventralpneumat-icopeningspresentonlyinLINHM014areunknownin any other pterosaur specimen. As both vertebraewere found in isolation, it is not possible to know iftheypertainedtodifferentpositionsontheneck(suchthatthepresenceoftheseforaminawouldbetypicalofone of the cervical vertebrae but absent in others) orif thesefeaturessuggestthattwodifferentazhdarchidspeciescoexistedinthisregion.CMN50801sharesanoverall similarmorphologywith thespecimenCCM-GE1/11915,holotypeofAzhdarcho lancicollis,whichwasreferredasthe?fifthcervical(Bakhurina&Unwin1995). Comparisons to the published proportions ofPhosphatodraco mauritanicus(seePereda-Suberbiolaetal. 2003) indicate thatbothMoroccanvertebraecom-parewellwiththeputativesixthcervical;however,thedescribed fifth cervical actually consists of two sepa-rate,closelyarticulatedthirdandfourthcervicalverte-brae (Kellner2010).Therefore,bothMoroccanverte-braeheredescribedshowproportionssimilartowhatis actually the fifth cervical of Phosphatodraco mau-ritanicus. If these suppositions are correct, then bothspecimensarefifthcervicalsandthereforerepresenta-tivesoftwodifferentazhdarchidtaxa.
Both Moroccan vertebrae described here differfrom the fifth cervical of Quetzalcoatlus sp. (TMM41544-15, cast NHMUK PV R 9325), which is ex-tremely elongate, demonstrating that they concernrelatively small individuals when in comparison withderivedazhdarchids.
ThehumerusCMN50814allowsabetterestima-tiveof thewingspanof this individual. Itcanbeesti-matedthatat leastbetween35mmand50mmoftheshaft ismissing, renderingaminimumtotal lengthofthehumerusofabout300mm.Comparisonswiththeestimated wingspans of Quetzalcoatlus northropi andMontanazhdarcho minor (whose holotypes possesscomplete humeri) rendered an estimative of ca. 5.5-6mforthisindividual.Evenifdifferentgrowthratesinwingbonesofthesameindividualaretakenintocon-sideration (as shown by Sayão 2003), such estimativeindicates that CMN 50814 concerns a large but notgigantic pterosaur. As both distal epiphyses are wellfusedtothehumeralshaft,CMN50814canbeconsid-eredatleastasub-adultindividual(Bennett1993).
Conclusions
Despite the large number of pterosaurs knownto date, the attribution of isolated edentulous jaws isstill not well understood, as illustrated by the scarcenumber of characters in phylogenetic matrices thatcould be used in this regard (e.g., Kellner 2003; Un-win 2003; Andres & Ji 2008; Lü et al. 2009; Wang etal. 2009). Clades such as the Azhdarchoidea mightcomprisearelativelyhighmorphologicaldiversity,andmorecompletematerialisstillneededtobetterunder-standthis.Perhapssomewhatsurprisingly, the identi-fication of isolated postcranial material of edentuloustaxaislessofachallenge.Certainfeatures(e.g.,therec-tangulardistalendofthehumeri)providecladesgoodsupportandcanbeusedinreferralstomoreinclusivecladeswithconfidence.
Furthermore,thevertebraeheredescribedprovideahintonazhdarchidevolution,showingalowbutnotvestigialneuralspine,suggestingthattheheightoftheneuralspinereducedinatleastsomemorederivedandyounger azhdarchid forms, such as the MaastrichtianspeciesQuetzalcoatlus northropiandArambourgiania philadelphiae, which also possess an extreme elonga-tionofthecervicalvertebrae(Kellner&Langston1996;Martilletal.1998;Henderson&Peterson2006).
PterosaurspecimensfromtheIfezouaneandtheAoufous formations of Morocco, even though beingincompleteand found isolated, canbeextremelywellpreserved,asitisthecaseofthetwocervicalvertebraehere described. They comprise a rather wide rangeof dsungaripteroid clades, and represent so far themost diverse pterosaur fauna from Africa (Kellner etal.2007; Ibrahimetal.2010).Whether suchadiversefaunacoexistedtemporallyorisanartifactofthelackof detailed stratigraphic information still remains tobeaccessed.Inanycase,theexistenceofsuchhighdi-versityofcladesshowsthepotentialofnewfindsthattheseformationscouldyield.
Acknowledgments. The authors would like to thank OliverRauhut(BayerischeStaatssammlungfürPaläontologieundGeologie),LornaSteel(NaturalHistoryMuseum)andEberhardFrey(StaatlichesMuseumfürNaturkundeKarlsruhe)foraccesstothepterosaurcollec-tions,EricBuffetaut(CNRS,LaboratoiredeGéologiedel’EcoleNor-maleSupérieure)forprovidinginformationandbibliographyontheKemKembeds,JulioCompany(UniversidadPolitécnicadeValencia)forthearticles,photosandreplicasent,AttilaOsi(HungarianAcad-emyofSciences)forphotosofMTMGyn/448andMTMGyn/450,and Naoki Ikegami (Mifune Dinosaur Museum) for the photo ofMDM349.ThearticlewasgreatlyimprovedbythemanysuggestionsofthereviewersFabioDallaVecchia(InstitutCatalàdePaleontologia)andKevinPadian(UniversityofCalifornia).Thisstudywaspartiallyfundedby theConselhoNacionaldeDesenvolvimentoCientíficoeTecnológico (CNPq – grants 140407/2007-3 and 290019/2008-7 toT.R. and 307276/2009-0 to A.W.A.K.) and Fundação Carlos Cha-gasFilhodeAmparoàPesquisadoRiodeJaneiro(FAPERJ–grantE-26/102.779/2008 to A.W.A.K.), with support from the DeutscherAkademischerAustauschdienst(DAAD–toT.R.).

158 Rodrigues T., Kellner A.W.A., Mader B.J. & Russell D.A.
AndresB.&JiQ.(2008)-AnewpterosaurfromtheLiaon-ingprovinceofChina,thephylogenyofthePterodac-tyloidea, andconvergence in theircervicalvertebrae.Palaeontology,51:453-469.
ArambourgC.(1959)-Titanopteryx philadelphiaenov.gen.,nov. sp. ptérosaurien géant. Notes et Mém. Moyen Orient.,7:229-234.
AverianovA.O.(2010)-TheosteologyofAzhdarcho lanci-collisNessov,1984(Pterosauria,Azhdarchidae) fromtheLateCretaceousofUzbekistan.Proceedings ZIN,314:264-317.
Averianov A.O., Arkhangelsky M.S. & Pervushov E.M.(2008) - A new Late Cretaceous azhdarchid (Ptero-sauria,Azhdarchidae)fromtheVolgaRegion.Paleon-tol. J.,42:634-642.
BakhurinaN.N.&UnwinD.M.(1995)-Asurveyofptero-saursfromtheJurassicandCretaceousoftheFormerSovietUnionandMongolia.Historical Biol.,10:197-245.
Bennett S.C. (1989) - A pteranodontid pterosaur from theEarlyCretaceousofPeru,withcommentsonthere-lationshipsofCretaceouspterosaurs.J. Paleontol.,63:669-677.
BennettS.C.(1993)-TheontogenyofPteranodonandotherpterosaurs.Paleobiology,19:92-106.
Bennett S.C. (2001) - The osteology and functional mor-phology of the Late Cretaceous pterosaur Pterano-don.PartI.Generaldescriptionandosteology.Palae-ontogr. Abt. A,260:1-112.
Bennett S.C. (2003) - Morphological evolution of the pec-toralgirdleofpterosaurs:myologyandfunction.In:BuffetautE.&MazinJ.M.(Eds)-EvolutionandPal-aeobiology of Pterosaurs, Geol. Soc. London Spec. Publ.,217:191-215.
BentonM.J.,BouazizS.,BuffetautE.,MartillD.M.,OuajaM., Soussi M. & Trueman C.N. (2000) - Dinosaursand other fossil vertebrates from fluvial deposits intheLowerCretaceousofsouthernTunisia.Palaeoge-ogr. Palaeoclimatol. Palaeoecol.,157:227-246.
BlackbeardM.&YatesA.M.(2007)-ThetaphonomyofanEarlyJurassicdinosaurbonebedinthenorthernFreeState(SouthAfrica).J. Vert. Paleont.,27:49A.
Blackburn D. (2002) - Two Early Cretaceous pterosaursfromAfrica.J. Vert. Paleont.,22:37A.
BuffetautE.(1999)-PterosauriafromtheUpperCretaceousof Laño (Iberian Peninsula): a preliminary compara-tivestudy.Est. Mus. Cien. Nat. Alava,14:289-294.
BuffetautE.(2001)-AnazhdarchidpterosaurfromtheUp-perCretaceousofCruzy(Hérault,France).Cr. Acad. Sci. II A,333:357-361.
CavinL.,TongH.,BoudadL.,MeisterC.,PiuzA.,TabouelleJ.,AarabM.,AmiotR.,BuffetautE.,DykeG.,HuaS.&LeLoeuffJ. (2010)-Vertebrateassemblagesfromthe early Late Cretaceous of southeastern Morocco:Anoverview.J. Afr. Earth Sci.,57:391-412.
Company J., Ruiz-Omeñaca J.I. & Pereda-Suberbiola X.
(1999) - A long-necked pterosaur (Pterodactyloidea,Azhdarchidae)fromtheUpperCretaceousofValen-cia,Spain.Geol. Mijnbouw,78:319-333.
CostaF.R.&KellnerA.W.A.(2009)-Ontwopterosaurhu-meri fromtheTendagurubeds(UpperJurassic,Tan-zania).An. Acad. Bras. Cienc.,81:813-818.
DalSassoC.&PasiniG.(2003)-Firstrecordofpterosaurs(Diapsida, Archosauromorpha, Pterosauria) in theMiddleJurassicofMadagascar.Atti Soc. Ital. Sci. Nat. Mus. Civ. Stor. Nat. Milano,144:281-296.
Dalla Vecchia F.M., Arduini P. & Kellner A.W.A. (2001) -ThefirstpterosaurfromtheCenomanian(LateCreta-ceous)LagerstättenofLebanon.Cretaceous Res.,22:219-225.
EttachfiniE.M.&AndreuB.(2004)-LeCénomanienetleTuronien de la Plate-forme Préafricaine du Maroc.Cretaceous Res.,25:277-302.
Henderson M.D. & Peterson J.E. (2006) - An azhdarchidpterosaurcervicalvertebrafromtheHellCreekFor-mation (Maastrichtian) of southeastern Montana. J. Vert. Paleont.,26:192-195.
Ibrahim N., Unwin D.M., Martill D.M., Baidder L. &ZouhriS.(2010)-Anewpterosaur(Pterodactyloidea:Azhdarchidae) from the Upper Cretaceous of Mo-rocco. PLoS ONE 5(5): e10875. doi:10.1371/journal.pone.0010875.
IkegamiN.,KellnerA.W.A.&TomidaY.(2000)-Thepres-enceofanazhdarchidpterosaurintheCretaceousofJapan.Paleontol. Res.,4:165-170.
KaupJ.J.(1834)-VersucheinerEintheilungderSaugethierein6StämmeundderAmphibienin6Ordnungen.Isis,3:311-315.
Kellner A. W. A. (2003) - Pterosaur phylogeny and com-ments on the evolutionary history of the group. In:Buffetaut E. & Mazin J.M. (Eds) - Evolution andPalaeobiologyofPterosaurs,Geol. Soc. London Spec. Publ.,217:105-137.
KellnerA.W.A.(2004)-NewinformationontheTapejaridae(Pterosauria,Pterodactyloidea)anddiscussionof therelationshipsofthisclade.Ameghiniana,41:521-534.
KellnerA.W.A.(2010)-CommentsonthePteranodontidae(Pterosauria, Pterodactyloidea) with the descriptionoftwonewspecies.An. Acad. Bras. Cienc.,82:1063-1084.
KellnerA.W.A.&CamposD.A.(2002)-ThefunctionofthecranialcrestandjawsofauniquepterosaurfromtheEarlyCretaceousofBrazil.Science,297:389-392.
KellnerA.W.A.&CamposD.A.(2007)-ShortnoteontheingrouprelationshipsoftheTapejaridae(Pterosauria,Pterodactyloidea).Bol. Museu Nacional ser. Geologia,75:1-14.
KellnerA.W.A.&LangstonJr.W(1996)-Cranialremainsof Quetzalcoatlus (Pterosauria, Azhdarchidae) fromLate Cretaceous sediments of Big Bend NationalPark,Texas.J. Vert. Paleont.,16:222-231.
KellnerA.W.A.&MaderB.J.(1996)-FirstreportofPtero-
REFERENCES

New pterosaur specimens from the Kem Kem beds (Upper Cretaceous, Cenomanian) of Morocco 159
sauria (Pterodactyloidea,Azhdarchidae) fromCreta-ceousrocksofMorocco.J. Vert. Paleont.,16:45A.
KellnerA.W.A.&MaderB.J.(1997)-ArchosaurteethfromtheCretaceousofMorocco.J. Paleontol.,71:525-527.
KellnerA.W.A.,MelloA.M.S.&FordT.(2007)-Asurveyof pterosaurs from Africa with the description of anewspecimenfromMorocco.In:CarvalhoI.S.,Cas-sabR.C.T.,SchwankeC.,CarvalhoM.A.,FernandesA.C.S.,RodriguesM.A.C.,CarvalhoM.S.S.,AraiM.&OliveiraM.E.Q.(Eds)-Paleontologia:CenáriosdeVida.Vol.1:257-267,RiodeJaneiro.
Kilian C. (1931) - Des principaux complexes continentauxduSahara.C. R. Soc. Géol. France,9:109-111.
Knoll F. (2000) - Pterosaurs from the Lower Cretaceous(?Berriasian) of Anoual, Morocco. Ann. Paleontol.,86(3):157-164.
LapparentA.F.de(1960)-LesDinosauriensdu‘Continen-tal Intercalaire’ du Sahara central. Mém. Soc. Géol. France,n.ser.88A:1-57.
Lawson D.A. (1975) - Response to Could pterosaurs fly?Science,188:676-677.
LiJ.,LüJ.&ZhangB.(2003)-AnewLowerCretaceoussi-nopteridpterosaurfromthewesternLiaoning,China.Acta Palaeontol. Sin.,42:442-447.
LüJ.,JinX.,UnwinD.M.,ZhaoL.,AzumaY.&JiQ.(2006)-AnewspeciesofHuaxiapterus(Pterosauria:Ptero-dactyloidea) from the Lower Cretaceous of westernLiaoning,Chinawithcommentsonthesystematicsoftapejaridpterosaurs.Acta Geol. Sin.,80:315-326.
LüJ.,UnwinD.M.,JinX.,LiuY.&JiQ.(2009)-Evidenceformodularevolutioninalong-tailedpterosaurwithapterodactyloid skull.Proc. R. Soc. B, doi: 10.1098/rspb.2009.1603.
Lü J. & Yuan C. (2005) - New tapejarid pterosaur fromwesternLiaoning,China.Acta Geol. Sin.,79:453-458.
MaderB.J.&KellnerA.W.A. (1999) -AnewanhangueridpterosaurfromtheCretaceousofMorocco.Bol. Mu-seu Nacional ser. Geologia,45:1-11.
MartillD.M.,FreyE.,SadaqahR.M.&KhouryH.N.(1998)- Discovery of the holotype of the giant pterosaurTitanopteryx philadelphiaeArambourg,1959andthestatusofArambourgiania andQuetzalcoatlus.Neues Jahrb. Geol. Palaontol.-Abh., 207:57-76.
McGowanA.J.&DykeG.J.(2009)-AsurfeitoftheropodsintheMoroccanLateCretaceous?Comparingdiver-sityestimates fromfielddataandfossil shops.Geol-ogy,37:843-846.
McGowenM.R.,PadianK.,DeSosaM.A.&HarmonR.J.(2002) - Description of Montanazhdarcho minor, anazhdarchidpterosaurfromtheTwoMedicineForma-tion(Campanian)ofMontana.Paleobios,22:1-9.
MonteilletJ.,LappartientJ.R.&TaquetP.(1982)-Unpté-rosauriengéantdansleCrétacésupérieurdePaki(Sé-négal).C. R. Acad. Sci. II,295:409-414.
Nessov L.A. (1984) - [Upper Cretaceous pterosaurs andbirds fromCentralAsia]Paleontol. Zh., 1: 47-57 [inRussian].
Ntamak-Nida M.J., Ketchemen-Tandia B., Ewane R.V.,Lissock, J.P. & Courville P. (2006) - Nouvelles don-
nées sur les Mollusques et autres macro-organismescampaniensdeSikoum(Centre-EstdusousbasindeDouala-Cameroun): intérêts bio-chronologiques etpaléo-écologiques.Afr. Geosc. Rev.,13:385-394.
Osi A., Weishampel D.B. & Jianu C.M. (2005) - First evi-denceofazhdarchidpterosaursfromtheLateCreta-ceousofHungary.Acta Palaeontol. Pol.,50:777-787.
PadianK.,deRicqlèsA.J.&Horner,J.R.(1995)-Bonehis-tologydeterminesidentificationofanewfossiltaxonofpterosaur(Reptilia:Archosauria).C. R. Acad. Sci. Paris,320:77-84.
Pereda-Suberbiola X., Bardet N., Jouve S., Iarochène M.,BouyaB.&AmaghzazM.(2003)-Anewazhdarchidpterosaur from the Late Cretaceous phosphates ofMorocco.In:BuffetautE.&MazinJ.M.(Eds)-Ev-olution and Palaeobiology of Pterosaurs, Geol. Soc. London Spec. Publ.,217:79-90.
PlieningerF.(1901)-BeiträgezurKenntnisderFlugsaurier.Paläontogr.,48:65-90.
RodriguesT.&KellnerA.W.A.(2008)-Reviewofthepter-odactyloid pterosaur Coloborhynchus. Zitteliana B,28:219-228.
Rodrigues T. & Kellner A.W.A. (2010) - Note on the pte-rosaur material described by Woodward from theRecôncavo Basin, Lower Cretaceous, Brazil. Rev. Bras. Paleontol.,13:159-164.
Rodrigues T., Kellner A.W.A., Mader B.J. & Russell D.A.(2006) - Brief report on new pterosaur (Pterosauria,Pterodactyloidea) specimens from the Cretaceous ofMorocco.J. Vert. Paleont.,26:116A.
SayãoJ.M.(2003)-Histovariabilityinbonesoftwoptero-dactyloid pterosaurs from the Santana Formation,Araripe Basin, Brazil: preliminary results. In: Buffe-tautE.&Mazin J.M. (Eds) -EvolutionandPalaeo-biologyofPterosaurs,Geol. Soc. London Spec. Publ.,217:335-342,London.
Sereno P.C., Dutheil D.B., Iarochène M., Larsson H.C.E.,Lyon G.H., Magwene P.M., Sidor C.A., VarricchioD.J.&WilsonJ.A.(1996)-PredatorydinosaursfromtheSaharaandLateCretaceousfaunaldifferentiation.Science,272:986-991.
Sigogneau-Russell D., Evans S. E., Levine J. F. & RussellD.A.(1998)-TheEarlyCretaceousmicrovertebratelocality of Anoual, Morocco: a glimpse at the smallvertebrateassemblagesofAfrica.In:LucasS.G.,Kirk-landJ.I.&EstepJ.W.(Eds)-LowerandmiddleCre-taceousterrestrialecosystems,New Mexico Mus. Nat. Hist. Sci. Bull.,14:177-181.
Swinton W.E. (1948) - A Cretaceous pterosaur from theBelgianCongo.Bull. Soc. Belge Geol. Paléont. Hydr. Liége,77:234-238.
UnwinD.M.(2003)-Onthephylogenyandevolutionaryhistoryofpterosaurs. In:BuffetautE.&MazinJ.M.(Eds) - Evolution and Palaeobiology of Pterosaurs,Geol. Soc. London Spec. Publ.,217:139-190.
UnwinD.M.&HeinrichW-D.(1999)-Onapterosaurjawfrom the Upper Jurassic of Tendaguru (Tanzania).Mitt. Mus. Nat. Berl., Geowiss. Reihe,2:121-134.
WangX.,KellnerA.W.A.,JiangS.&MengX.(2009)-An

160 Rodrigues T., Kellner A.W.A., Mader B.J. & Russell D.A.
unusual long-tailed pterosaur with elongated neckfrom western Liaoning. An. Acad. Bras. Cienc., 81:793-812.
WangX.&ZhouZ.(2003)-Anewpterosaur(Pterodacty-loidea,Tapejaridae)fromtheEarlyCretaceousJiufo-tang Formation of western Liaoning, China and itsimplications for biostratigraphy. Chin. Sci. Bull., 48:16-23.
Wellnhofer P. & Buffetaut E. (1999) - Pterosaur remainsfrom the Cretaceous of Morocco. Palaontol. Z., 73:133-142.
WellnhoferP.&KellnerA.W.A.(1991)-TheskullofTape-jara wellnhoferi Kellner (Reptilia, Pterosauria) fromtheLowerCretaceousSantanaFormationoftheArar-ipe Basin, northeastern Brazil. Mitt. Bayer. Staatss. Paläon. hist. Geol.,31:89-106.
YoungC.C.(1964)-OnanewpterosaurianfromSinkiang,China.Vertebrata Palasiatica, 8:221-225.
YoungC.C.(1973)-[PterosaurianfaunafromWuerho,Sin-kiangII.In:ReportsofPaleontologicalExpeditionstoSinkiangII.] Inst. Vertebr. Paleontol. Paleoanthropol. Acad. Sin. Memoirs,11:18-35.[inChinese].