late cretaceous (cenomanian–turonian) macroflora from the chatham islands, new zealand:...
TRANSCRIPT

�������� ����� ��
Late Cretaceous (Cenomanian–Turonian) macroflora from the ChathamIslands, New Zealand: Bryophytes, lycophytes and pteridophytes
Chris Mays, Anne-Marie P. Tosolini, David J. Cantrill, Jeffrey D. Stil-well
PII: S1342-937X(14)00108-7DOI: doi: 10.1016/j.gr.2014.03.017Reference: GR 1250
To appear in: Gondwana Research
Received date: 30 October 2013Revised date: 18 February 2014Accepted date: 14 March 2014
Please cite this article as: Mays, Chris, Tosolini, Anne-Marie P., Cantrill, David J., Stil-well, Jeffrey D., Late Cretaceous (Cenomanian–Turonian) macroflora from the ChathamIslands, New Zealand: Bryophytes, lycophytes and pteridophytes, Gondwana Research(2014), doi: 10.1016/j.gr.2014.03.017
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
1
Late Cretaceous (Cenomanian–Turonian) macroflora from the Chatham
Islands, New Zealand: bryophytes, lycophytes and pteridophytes
Chris Maysa,*, Anne-Marie P. Tosolini
b, David J. Cantrill
c,d and Jeffrey D. Stilwell
a
a School of Geosciences, Monash University, Victoria, 3800, Australia
b School of Earth Sciences, The University of Melbourne, Victoria, 3010, Australia
c National Herbarium of Victoria, Royal Botanic Gardens Melbourne, Private Bag 2000, South Yarra, Victoria,
3141, Australia
d School of Botany, The University of Melbourne, Victoria, 3010, Australia
* Corresponding author; School of Geosciences, Monash University, Wellington Road, Victoria, 3800, Australia;
+61 3 9905 5786; Fax +61 3 9905 4903; email: [email protected]
Mays, C., Tosolini, A.-M.P., Cantrill, D.J. and Stilwell, J., 2013. Late Cretaceous (Cenomanian–Turonian) macroflora from the
Chatham Islands, New Zealand: bryophytes, lycophytes and pteridophytes. Gondwana Research XX, XXX–XXX. ISSN
XXXXX-XXXXX.
Abstract. Late Cretaceous fossils from the Chatham Islands, New Zealand, represent an
important high palaeolatitude (~ 70–80°S) flora. Located between eastern and western
Gondwana, and prior to Late Cretaceous continental break-up, these plants grew during a period
of global greenhouse climates. Macrofloral remains of the Tupuangi Formation, Pitt Island,
accumulated in a deltaic floodplain setting with plant material occurring on well-developed,
hydromorphic soil horizons or entrained in sediments overlying the soils. The macroflora
includes a rich angiosperm-conifer-Ginkgo flora with subsidiary ferns, lycophytes and
bryophytes. The components of the assemblage described herein include those of probable
cryptogam affinity, and comprise one thalloid liverwort, one leafy moss (Muscites gracilis sp.
nov.), one lycopod shoot, and three taxa of pteridophytic affinity, including a fern of probable
osmundalean affinity (Cladophlebis auriculipilosus sp. nov.). The floral assemblage shares only
one element with coeval localities across the Southern Hemisphere (Sphenopteris sp. cf. S.
warragulensis), and is characterised by relatively low fern diversity. The unique assemblage is

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
2
attributed, at least in part, to ecological bias associated with local deltaic depositional settings.
However, the relatively low osmundalean fern component follows global floristic trends for the
mid-Cretaceous, which saw a decline in diversity and abundance of ferns during the early phases
of angiosperm diversification and rise to ecological dominance.
Keywords: Cretaceous, fern, bryophyte, lycopod, Chatham Islands.
1. Introduction
The early Late Cretaceous (Cenomanian–Turonian) has long been recognised as an interval of
global greenhouse conditions (Barron, 1983; Arthur et al., 1985; Miller et al., 2005). Marine
fossil evidence suggests that this warming was acutely expressed at polar latitudes, with a
temperature maximum during the Turonian (Huber et al., 1995; Huber, 1998; Bice et al., 2006).
This Cretaceous warming trend has been supported by fossil floral palaeotemperature proxies
from Antarctica (Hayes, 1999; Francis and Poole, 2002), and a Cenomanian floral assemblage
from New Zealand (mean annual temperature: ~ 10°C; Parrish et al., 1998). Atmospheric carbon
dioxide levels were higher than modern levels by a factor of between four and seven (Berner and
Kothavala, 2001; Bice et al., 2006; Wang et al., in press); it has been proposed that such high
CO2 concentrations in ancient atmospheres would lead to a state of ‘carbon fertilisation’,
enhancing polar forest productivity (e.g. Beerling and Osborne, 2002). These global conditions
coincide with major changes to terrestrial ecosystems.
The mid-Cretaceous is characterised as an interval of angiosperm diversification and rise to
ecological dominance (e.g. Wolfe et al., 1975; Crane et al., 1995; Lupia et al., 1999).
Angiosperm diversification was diachronous, occurring at equatorial palaeolatitudes prior to the
mid- and high latitudes. This pattern of diversification is well-established in the Northern

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
3
Hemisphere (Lidgard and Crane, 1988, 1990), with preliminary evidence supporting a
comparable trend for the Southern Hemisphere (Drinnan and Crane, 1990; Cantrill and Poole,
2002; Nagalingum et al., 2002). Over the same interval, relative diversity trends of free-sporing
plants (ferns, lycophytes and non-vascular plants) consistently show an overall decrease
(Northern Hemisphere, Lupia et al., 1999; Antarctica, Cantrill and Poole, 2002; Australia,
Nagalingum et al., 2002), raising the possibility of ecologic replacement (Lupia et al., 1999).
However, the decline of fern diversity during the Cretaceous was not a uniform trend across all
fern groups. Molecular data suggest that some fern groups, most notably the polypods, which are
the most diverse extant clade, show an overall increase in diversity (Pryer et al., 2004; Schneider
et al., 2004). Of the free-sporing plant groups that exhibit a decline in diversity and abundance
during the mid- to Late Cretaceous, osmundaceous ferns appear to have been particularly
affected (e.g. Nagalingum et al., 2002).
Most gondwanan Cretaceous free-sporing plant fossils have been recovered from Lower
Cretaceous strata as a consequence, at least in part, of this mid- to Late Cretaceous floral
overturn. Early Cretaceous cryptogam leaf floras of Gondwana have been primarily reported
from the Antarctic Peninsula (Jefferson, 1981; Cantrill, 1995, 1996, 1997, 1998; Nagalingum,
2003; Cantrill and Nagalingum, 2005; Nagalingum and Cantrill, 2006; Cantrill and Poole, 2012),
and Australia (Seward, 1904; Douglas, 1973; Drinnan and Chambers, 1986; McLoughlin, 1996;
McLoughlin et al., 1995, 2002). These studies consistently show a highly diverse fern
component, with rare lycopods and horsetails, and locally dominant, non-vascular plants. Indeed,
the upper Lower Cretaceous strata of Alexander Island, Antarctic Peninsula, have yielded a
macrofloral assemblage where ferns comprise the most diverse group (Falcon-Lang et al., 2001;
Nagalingum, 2003). This high diversity of free-sporing plants is reflected in the Early
Cretaceous palynological records of both southeast Australia (e.g. Dettmann, 1986, 1994;
Nagalingum et al., 2002; Wagstaff et al., 2013) and the Antarctic Peninsula (Dettmann and

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
4
Thomson, 1987; Cantrill and Poole, 2002), with both regions typically exhibiting a greater
diversity of free-sporing plants than of seed-plants.
There are only a few reports of mid-Cretaceous macrofloral assemblages from South Island,
New Zealand, by Ettingshausen (1887), McQueen (1956) and Daniel (1989). Daniel (1989)
illustrated several cryptogams (pls. 10–11) from the mid-Cretaceous (Albian/Cenomanian) strata
of the Clarence Valley, South Island, and included a comparison of various informally described
fossil pinnules of the extinct osmundaceous Phyllopteroides; several of these were regarded as
species by Parrish et al. (1998), but without formal taxonomic descriptions. Pole and Philippe
(2010) reported dispersed gymnosperm and angiosperm leaf cuticle taxa from the Cenomanian–
Turonian Tupuangi Formation of the Chatham Islands. Recent fieldwork on the Chatham Islands
has yielded an extensive macroflora, which serves as the basis for the present study — the first
of a series of contributions to document this floral assemblage. This is the first systematic study
of probable cryptogamic leaf fossils from eastern Zealandia. This early Late Cretaceous flora
will provide an important data-point for future studies of floristic change across the southern
high latitudes.
2. Geological background
The Chatham Islands are located ~ 850 km east of Christchurch, New Zealand (~ latitude:
44°S; longitude: 176°W), and are the only emergent part of an elongated plateau of continental
crust known as the Chatham Rise (Fig. 1A). The Chatham Rise extends eastward from South
Island, New Zealand, and is a segment of the coherent and mostly submerged subcontinent
Zealandia (including New Zealand, New Caledonia, the Lord Howe Rise and the Campbell
Plateau). Prior to Late Cretaceous continental breakup, Zealandia was conjoined with the
Australian and Antarctic sectors of eastern Gondwana (Fig. 2). The lower Upper Cretaceous

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
5
fossiliferous terrestrial sequence of the Chatham Islands (the Tupuangi Formation) was
deposited on the margins of the Bounty Trough, an extensional basin between the Chatham Rise
and the Campbell Plateau (Fig. 1A). This basin formed prior to, or concurrent with, Zealandia-
Gondwana rifting (Bradshaw et al., 1996; Eagles et al., 2004). During the early Late Cretaceous,
the Chatham Rise was primarily emergent (Wood et al., 1989; Campbell et al., 1993) and located
within the south polar circle (palaeolatitude: ~ 70–80°S; Markwick et al., 2000; Mukasa and
Dalziel, 2000; Stilwell et al., 2006).
<INSERT FIG. 1>
The Tupuangi Formation is the oldest sedimentary stratigraphic unit exposed on the Chatham
Islands (Campbell et al., 1993). On the basis of spore-pollen biostratigraphy, the formation was
originally correlated to the Motuan to Mangaotanean New Zealand chronostratigraphic stages
(upper Albian to Turonian; R.A. Couper in Hay et al., 1970), but Mildenhall (1994) proposed a
correlation to the Teratan New Zealand stage (Coniacian) for the uppermost strata. The
biostratigraphic range was later refined to Ngaterian to Mangaotanean (Cenomanian to
Turonian; Mays and Stilwell, 2013), as no autochthonous Motuan or Teratan spore-pollen
assemblages could be verified. The Tupuangi Formation is overlain by the Kahuitara Tuff. A
probable coeval unit of the Kahuitara Tuff, the Southern Volcanics, has been radiometrically
dated with a maximum age of 85.45 ± 0.6 Ma (Piripauan; Santonian; Panter et al., 2006). This
suggests an unconformity between the Tupuangi Formation and the Kahuitara Tuff (Mays and
Stilwell, 2013).
<INSERT FIG. 2>
The Tupuangi Formation has an estimated exposed thickness of ~ 400 m (Campbell et al.,
1993; Mays and Stilwell, 2013), and has only been reported from stratigraphic sections on Pitt
Island (Māori: Rangiauria; Moriori: Rangiaotea). Pitt Island is the second largest island of the
Chatham Islands archipelago, and is located 22 km southeast of Chatham Island (Fig. 1B). The

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
6
Tupuangi Formation consists primarily of interbedded, poorly-consolidated, quartzofeldspathic
fine sandstone and carbonaceous siltstone facies (Campbell et al., 1993). Lignitic layers (≤ 1.2 m
thick) and large in situ tree stumps with associated root systems are present at discrete horizons
throughout the interbedded silt-/sandstone succession. The Tupuangi Formation has been
interpreted as primarily fluvial to deltaic (Campbell et al., 1993), with palynological evidence of
restricted marine influence from the uppermost (Turonian) strata (Mays and Stilwell, 2013).
3. Materials and methods
<INSERT FIG. 3>
Fossil material from Pitt Island was collected by C. Mays, D.J. Cantrill, J.D. Stilwell and
volunteer field assistants from Monash University, Clayton, Australia, during two field seasons:
January–February, 2009, and February, 2012. Fossils were obtained from outcrops at Waihere
Bay, western Pitt Island (Fig. 3). All specimens were collected from the Tupuangi Formation.
Individual hand samples are provided with unique sample registration numbers, denoted by the
prefix “PL”, and are registered with GNS Science, Lower Hutt, New Zealand. An individual
fossil specimen may comprise multiple samples, e.g., part and counterpart pairs. Localities are
given Fossil Record File numbers, and provided with prefix “CH/f”. To provide stratigraphic
context, stratigraphic logs were made of all reported fossil localities, except for CH/f0774, which
was a non-coastal outcrop and could not be reliably correlated to the coastal Waihere Bay
succession (Fig. 4). Locality and sample details, including figure numbers and part/counterpart
relationships, are provided herein as supplementary data (Table 1).
<INSERT FIG. 4>
Macrophotographs were taken using a Canon Powershot SX120 IS; photomicroscopy was
undertaken using a Leica M205-C, with a Leica DFC290 camera. Line drawings were performed
with a Wacom Bamboo pen tablet. Bold lines of line drawings represent the preserved margins

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
7
of the fossil specimens, but due to a range of taphonomic factors, these may not necessarily be
the margin of the specimen prior to burial; dashed lines represent inferred margins or veins
(unless otherwise stated).
Terms employed in the systematic descriptions conform to the definitions of Ellis et al.
(2009) where relevant; all other taxonomic descriptive terms follow Harris and Woolf-Harris
(2001). Where referred to, extant subclasses, orders, families and genera follow the classification
of Christenhusz et al. (2011). Stratigraphic and sedimentological terms are after Tucker (2011);
palaeosol terms are after Retallack (2001), except where otherwise stated.
4. Results
4.1 Stratigraphic context of fossil localities
The fossil plants are found consistently within massive to weakly laminated to bedded, poorly
consolidated, grey to light brown (chroma: 1–2, as per Munsell, 1975), carbonaceous clay- to
siltstone facies. Laminae are typically defined by beds of leaf remains (generally comminuted);
siltstone fossiliferous lamina surfaces are slightly undulate. Locally, fossiliferous layers are bed-
scale size (1–15 cm) and composed of compressed, carbonised plant remains, but these
fossiliferous beds are not suitable for taxonomy due to the density of fossil material and
distortion of individual specimens. The fossiliferous fine-grained sedimentary packages
represent the upper sections of fining-upward successions 30 to 250 cm thick; these are overlain,
and commonly truncated, by beds of massive to cross-bedded fine- to medium-grained sandstone
(Fig. 4A, B). Fossil roots/rootlets extending below the fossil beds were identified at three
localities (CH/f0775, 0776 and 0778). The sediments at three localities appeared to be partly to
highly weathered as indicated by the light brown colouration (chroma: 2; CH/f0776–0778). The

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
8
fossils from these localities were typically preserved as oxidised impressions. Oxidation appears
to be most intense on outcrop surfaces and adjacent to faults/joints; as such, this feature was
most likely due to late-stage diagenetic factors such as surficial weathering and ground-water
penetration via rock fractures. Localities CH/f0776 and 0778 are rich in sulphide content,
typically pyrite and marcasite.
4.2 Systematic palaeontology
Marchantiophyta Crandall-Stotler and Stotler, 2000
Class incertae sedis
Hepaticites Walton, 1925
Type species
Hepaticites langii Walton, 1925 (lectotype chosen by Schuster, 1966)
Hepaticites sp. (Fig. 5A–C)
Material examined
One specimen as follows: PL1014 (Fig. 5A–C).
Description
Plant thalloid, spreading, prostrate, preserved portion 24 mm in diameter. Individual thallus
branches are 1.2–1.8 mm wide, ≤ 15.5 mm long, dichotomising up to five times, at irregular
intervals (1–3.8 mm); final branch dichotomy diverges at 26–44°, other dichotomies diverge at
39–86°. Thallus apices rounded, margins entire except notched at apices (Fig. 5C). Costa 0.1–0.5

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
9
mm wide where discernible, thinning towards terminal branches; thalli are darkest at costa and
gradationally lighten towards margins. Sporophytic remains not present.
Comparison and remarks
The present taxon is assigned to the fossil-genus Hepaticites, which includes all fossil taxa
having an apparent relationship to the “Hepaticae”, following the system employed by
Oostendorp (1987). Recent phylogenetic reviews of extant liverworts have disbanded Hepaticae,
and recognised three distinct classes under Marchantiophyta: Jungermanniopsida,
Haplomitriopsida and Marchantiopsida (Crandall-Stotler and Stotler, 2000; Crandall-Stotler et
al., 2009). The thalloid organisation of the present taxon is most similar to extant members of
Marchantiopsida or Jungermanniopsida, but the features necessary (e.g. sporophytic remains,
cellular structure, rhizoids) to satisfactorily attribute it to either of these extant classes are not
preserved. The gradual lightening of colour from the midribs suggests that the original tissue
was thinner away from the costal regions and in the thallus apices; this feature is similar to
Hepaticites undulatus Drinnan and Chambers, 1986 but the thalli of the present taxon are much
smaller. Hepaticites minutus Cantrill, 1997 and H. arcuatus (Lindley and Hutton, 1837) Harris,
1942 both feature similar branching patterns, but the thalli of the former are consistently smaller,
whilst those of the latter are larger, and both lack the apical notches that characterise the
presently described specimen. The anthocerophyte Dendroceros victoriensis Drinnan and
Chambers, 1986 is similar to the present specimen by having notched thallus apices and a
branching, prostrate, spreading habit, but the present specimen is smaller and lacks the
preservation quality to express the diagnostic arcuate ribs of D. victoriensis.
<INSERT FIG. 5>
Bryophyta Schimp.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
10
Class incertae sedis
Muscites Brongniart, 1828
Type species
Muscites tournali Brongniart, 1828
Muscites gracilis Mays and Cantrill sp. nov. (Fig. 5D–G)
Holotype
Holotype specimen comprises two hand samples, part (PL1027; Fig. 5D–F) and counterpart
(PL1028). Holotype is from locality CH/f0777, Waihere Bay, Pitt Island, Chatham Islands, New
Zealand, Tupuangi Formation, Cenomanian.
Other material
Two specimens as follows: PL1033, PL1034. Both from locality CH/f0778.
Etymology
Latin gracilis, meaning slender or narrow, describing the axis and the diagnostic leaf shape.
Diagnosis
Axes furcate into three–five branches; leaves decurrent, linear to lanceolate with involute
margins and prominent costae.
Description

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
11
Branched axis habit, preserved axis length ≤ 24 mm, 0.3–0.8 mm wide; axis furcates into
three–five separate branches at 25–50° at discrete nodes spaced 7–15 mm apart. Axes appear to
bear leaves along their entire length, but insertion relationship is not clear; leaf lengths increase
towards the distal ends of the axes. Leaves are sessile, decurrent and reflexed (deviating from
stem asymptotically, but reflexing typically ≤ 65°, but as much as 180° in rare cases; see Fig. 5E,
F), linear to lanceolate (1.8–3 mm length, 0.2–0.5 mm basal width) and apices are acute; leaf
margins are entire and are involute. Costae are prominent, extending to apices and may be
excurrent. Lamina colour lightens from costa to margins (see Fig. 5G). Sporophytes are not
present.
Comparison and remarks
The present taxon has a branched pattern of strand-like stems and sessile leaves with distinct
midribs, as per the original diagnosis of the fossil-genus Muscites (Brongniart, 1828). These
specimens are similar to various extant taxa of Polytrichopsida (e.g. Polytrichastrum G.L. Sm.).
The gross morphology of the present species resembles the branching fossil pleurocarp
Palaeodichelyma sinitzae Ignatov and Shcherbakov, 2007 which has a probable affinity to
Bryopsida. However, the cellular details are not discernible due to the resolution constraints
imposed by the enclosing coarse sediments. This factor, combined with an absence of preserved
sporophytic details, precludes assignment to class level.
The leaves of this taxon have been flattened against the enclosing sediments in dorsiventral,
lateral and intermediate orientations. The leaves have been described as involute because of the
basiscopic position of the costa on the laterally compressed leaves diverging from the medial
portions of the stem (see Fig. 5G). However, when the leaves are compressed dorsiventrally, as
at the distal end of the stem (see Fig. 5E, F), it is evident that the costae are situated in the
middle of the lamina.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
12
The linear to lanceolate leaves and abaxial midribs of Muscites gracilis sp. nov. are similar to
M. antarcticus Cantrill, 2000 recovered from Aptian strata of Snow Island, Antarctica; PL1034
from the Tupuangi flora (Fig. 5G) appears particularly similar, this specimen having slightly
broader leaves than the holotype. However, the present taxon consistently has more slender axes,
and longer, thinner leaves. The taxon denoted “gametophyte type 2” described by Drinnan and
Chambers (1986) from Lower Cretaceous strata of southeastern Australia features linear leaves
and a branching habit, but the leaves appear to be excurrent rather than consistently decurrent,
and the axes are too robust to be conspecific with the presently described taxon. This is the first
Cretaceous moss macrofossil to be described from New Zealand.
Lycopodiophyta Cronquist et al., 1966
Lycopodiopsida Bartling, 1830 (as "Lycopodineae")
Order incertae sedis
Lycopodites Lindley and Hutton, 1833 non Brongniart, 1822
Type species
Lycopodites falcatus Lindley and Hutton, 1833 emend. Harris, 1961
Lycopodites sp. (Fig. 5H)
Material examined
One specimen (PL1002).
Description

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
13
Dichotomous secondary axis branches from primary axis at 55°, primary axis width 0.19–
0.24 mm, secondary axis width 0.16–0.2 mm. Homophyllous; leaves sessile, erect, laterally
complanate, alternately arranged; leaves crowded but not overlapping, spaced 0.8–1.2 mm (as
measured from basal margins). Leaf elliptical, apex straight to acuminate (possibly mucronate),
proximal margin of lamina approximately at right angles to stem. Primary axis leaf length 1.4–
1.6 mm, width 0.8–1.0 mm; secondary axis leaf length 1.1–1.2 mm, width 0.5–0.6 mm. Lamina
thickenings evident on proximal (width: 80–150 µm) and distal (width: 60–90 µm) margins.
Fertile organs not present and cellular detail not preserved.
Comparison and remarks
A probable affinity to Lycopodiales has been proposed for various species of Lycopodites,
e.g. L. falcatus Lindley and Hutton, 1833 emend. Harris, 1961 (Harris, 1961); L. victoriae
Seward, 1904 (Douglas, 1973). The same conclusion cannot be made for the present taxon, due
to the absence of preserved homosporous or heterosporous strobili, which would aid in
distinguishing between an affinity with Lycopodiales or Selaginellales, respectively. There
appears to be a distinct size difference between the leaves of the two preserved stems of this
specimen, suggesting a growth habit with subsidiary stems branching from a primary stem. Such
a feature is expressed in various extant species of Selaginella (e.g. S. hordeiformis Baker; S.
novae‑hollandiae (Sw.) Spring). Conversely, the homophyllous habit is suggestive of a
lycopodiaceous affinity (see discussion by Harris, 1961). Furthermore, complanate leaves are
common amongst extant lycpodiaceous species (e.g. Diphasiastrum complanatum (L.) Holub, D.
digitatum (Dillenius ex A. Braun) Holub), whilst this character is rare amongst Selaginellales.
Lycopodites gracilis Oldham and Morris, 1863 and the type species L. falcatus Lindley and
Hutton, 1833 emend. Harris, 1961 are both heterophyllous, but the present taxon is
homophyllous with no evidence of decussate phyllotaxis. Lycopodites victoriae Seward, 1904 is

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
14
homophyllous, but the leaves are typically larger, falcate and have a prominent midrib. The
leaves of L. arberi Edwards, 1934 are erect, similar to the presently described specimen, but are
larger, tend to overlap and have mucronate apices; the present taxon may have this latter feature,
but the apices are not adequately preserved to be described as such. Walkom (1928) described
two lycopodiaceous taxa from Early Cretaceous (Burger, 1980) strata of northeast Australia:
?Lycopodites sp. A and B. The former has a prominent midrib and sub-opposite leaves, whereas
the presently described specimen has alternately arranged leaves with no visible midrib; the
latter has long, lanceolate leaves, rather than the short, elliptical leaves of the Tupuangi taxon.
Lycopodites sp. reported by Rigby (1977) from Middle Jurassic strata of Queensland, Australia
(redrawn and ascribed to L. gracilis (Morris) Seward and Sahni, 1920 by Mcloughlin et al., in
press), has falcate, rather than elliptical, leaves. McQueen (1956) recorded a specimen of
‘Lycopdium cf. volubile Forster f. 1786’ from the Seymour River coal measures (Warder
Formation), middle Clarence Valley, New Zealand, which have since been estimated as Albian–
Cenomanian in age (Douglas and Williams, 1982; Browne and Reay, 1993). The specimen
illustrated is much larger than the present specimen, and exhibits linear, falcate leaves. Upon re-
examination of the specimen recorded by McQueen, Daniel (1989) suggested that it has a
probable podocarpaceous affinity; indeed, the specimen appears to have close resemblance to the
conifer Pagiophyllum, a common element recorded from Albian strata of Alexander Island,
Antarctic Peninsula (as Pagiophyllum sp.; Cantrill and Falcon-Lang, 2001).
An interesting feature of the presently described specimen is the presence of thickened tissue
on both the proximal and distal lamina margins, giving the appearance of a bicostate leaf
architecture; however, no recorded extant member of Lycopodium has such venation (e.g. Chu,
1974). Further, Lycopodiaceae is defined by a single vein (Mirbel in Lamarck and Mirbel, 1802,
in Øllgaard, 1990), whereas a bicostate habit would imply an affinity outside this group. To

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
15
satisfactorily conclude that the present taxon has two marginate costae will require cellular
details that cannot be ascertained from the present specimen.
<INSERT FIG. 6>
Polypodiophyta Cronquist et al., 1966
Polypodiopsida Cronquist et al., 1966
?Osmundales Link, 1833 (as “Osmundaceae”)
Cladophlebis Brongniart, 1849
Type species
Cladophlebis albertsii (Dunker, 1846) Brongniart, 1849
Cladophlebis auriculipilosus Mays and Tosolini, sp. nov. (Fig. 6)
Holotype
Comprises two hand samples, part (PL1036; Fig. 6A, B) and counterpart (PL1035; Fig. 6C–
E). Holotype is from locality CH/f0778, Waihere Bay, Pitt Island, Chatham Islands, New
Zealand, Tupuangi Formation, Cenomanian.
Other material
One specimen from locality CH/f0778 comprising two hand samples: PL1031 (part), PL1032
(counterpart).
Etymology

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
16
Latin auriculi-, from auricula, meaning ear in reference to the diagnostic pinnule auricles;
Latin -pilosus, meaning covered in hair referring to the distinctive hair-like trichomes of the
rachises and rachillae.
Diagnosis
Rachises and rachillae have trichomes with longitudinally elongated bases. Alternating,
sessile pinnae. Pinnules adnate, ovate. Minor auricle on acroscopic pinnule margin, basiscopic
margin decurrent to rachilla, apices convex to rounded. Pecopteroid venation, primary vein
decurrent, distally reflexed.
Description
Frond at least bipinnate (complete frond not preserved), preserved frond 58 mm long (total
length unknown). Rachis ≤ 1.8 mm wide with no lamina wing. Rachis and rachillae feature
trichomes with longitudinally elongated decurrent bases (≤ 1.75 mm long, ≤ 0.2 mm wide; see
Fig. 6C). Preserved pinnae ≤ 45 mm long (total length not known) by ≤ 13 mm wide, distichous
and alternate arrangement, rachillae ≤ 1 mm wide, sessile and inserted laterally or slightly
dorsally on rachis at 65–70°. Pinna planes slightly tilted (5–10°) relative to the plane of the
rachis. Pinnules ≤ 7.5 mm long by ≤ 4.9 mm wide, distichous and sub-opposite arrangement,
inserted laterally on rachilla with adnate attachment; pinnules oblong, apex shape convex to
rounded. Pinnules are typically curved perpendicular to plane of the lamina (?revolute). Pinnules
along proximal rachilla typically discrete, medial pinnules conjoined at bases, complete fusion of
pinnule margins at apices. Isolated pinnules are crowded to partially superimposed, density of
isolated pinnules is approximately 2.3–3.6 per 1 cm along rachillae, increasing apically along
pinna. Pinnule margins entire to sinuous, acroscopic margins are straight to convex but widening
with small auricle and constricted basally, basiscopic margin is straight to convex and basally

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
17
decurrent to the rachilla; basiscopic margins of discrete pinnules typically exhibit overlap with
part of the acroscopic auricle of the subtending pinnule. Venation pattern is pecopteroid; primary
vein, ≤ 0.2 mm wide, diverges from rachilla decurrently, reflexing distally into pinnule, straight
to sympodial in the medial portion of pinnule, angle between medial primary vein and rachilla
65–90°. Primary vein persists for at least two thirds of pinnule length, but evanesces into two
apical secondary veins. From primary vein, ≤ 10 distichously arranged secondary veins diverge
at ≤ 55°, dichotomising once, or less commonly twice, before reaching pinnule margin. Vein
density is approximately 11–15 per 10 mm along the pinnule margin. Initial secondary vein
typically anadromic. Acroscopic secondary vein diverges from primary vein at an angle
approximately parallel to rachilla, dichotomising veinlets terminate at both auricle apex and
sinus. No fertile material was preserved.
Comparison and remarks
The diagnosis of Cladophlebis Brongniart, 1849 describes sterile fronds with pecopteroid
venation, whereas equivalent fertile fronds are referred to several other genera, most notably
Osmundopsis Harris, 1931 emend. Harris, 1961 and Todites Seward, 1900 emend. Harris, 1961
(see review by Balme, 1995). This association of Osmunda-like fertile fronds with their sterile
counterparts provided strong support for a phylogenetic relationship for various members of
Cladophlebis with Osmundales (Phipps et al., 1998). The rachises/rachillae of Osmundaceae
feature grooves and ridges, as per the original familial description by Berchtold and Presl (1820,
fides Kramer, 1990). As such, the presence of diagnostic trichomes on the rachises/rachillae
precludes a definitive association of the presently described species to Osmundaceae.
Furthermore, there have been reported specimens of Cladophlebis that are not likely to be allied
to Osmundales. Cladophlebis cyathifolia Villar de Seoane, 1996 has been found with in situ
Cyathidites-type spores, suggesting an affinity to Cyatheales (Villar de Seoane, 1996, 1999); this

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
18
taxon was later referred to Cyathea cyathifolia (Villar de Seoane, 1996) Villar de Seoane, 1999.
The spores associated with Cladophlebis tripinnata Archangelsky, 1963 emend. Villar de
Seoane, 1996 exhibit both monolete and trilete forms, which Villar de Seoane (1996) interpreted
as indicating an affinity with Dennstaedtiaceae (Polypodiales). However, this is not exclusively a
feature of Dennstaedtiaceae; many species of disparate extant fern groups have been known to
produce both monolete and trilete spores (Selling, 1944; Brown, 1960). A range of fossil species
also exhibit this feature, including members of Marattiales (e.g. Marattia anglica (Thomas)
Harris, 1961; see review by Murdock, 2008) and Dipteridaceae (Polypodiales; e.g. Polyphacelus
stormensis, Yao et al., 1991). This suggests that a mix of spore aperture types from an individual
fertile frond is not a reliable diagnostic feature for any specific fern group; however, no cases of
fertile fossil fronds of unequivocal osmundalean affinity have yet been found with a combination
of monolete and trilete spores. These examples emphasise the probability that Cladophlebis is
not limited to Osmundales. Because Cladophlebis is most commonly associated with
Osmundales, this study tentatively assigns an osmundalean affinity to C. auriculipilosus;
however, without the preservation of fertile material, or the observation of anomocytic stomata
on the lamina surface, uncertainty remains over the assignment of this species to order (or
family), following the recommendation of Seward (1904).
The sterile fronds of Todites williamsonii (Brongniart, 1828) Seward, 1900 emend. Harris,
1961 have sessile, cladophleboid, typically isolated pinnules, similar to the presently described
specimens, but have acute apices that curve acroscopically. The pinnules of Cladophlebis
octonerva Holmes, 2003 from the Triassic of New South Wales, Australia, lack both auricles on
the acroscopic margin and trichomes along rachis/rachilla (Holmes, 2003). Douglas (1973)
reported a similar taxon (Cladophlebis sp. “a”) from the Early Cretaceous (?Berriasian–
Hauterivian) strata of southeastern Australia; however, the pinnules were much more elongate,
were not decurrent on the basiscopic margin, and lacked auricles. There have been several

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
19
reported specimens of Cladophlebis from Cretaceous strata of New Zealand, yet none of these
taxa has equivalent pinnule shape (see Table 2). The species described herein is similar in many
respects to the specimen denoted Cladophlebis sp. by Drinnan and Chambers (1986) from the
Lower Cretaceous (Aptian–Albian; Dettmann, 1986) of Victoria, Australia. Shared features
include: pinnule sizes with entire margins; recurved and flexuose midveins; and variable
presence of an auricle on the acroscopic margins of the laminae. However, the alternate pinnule
arrangement, hastate to falcate pinnule shape, and the consistently catadromic initial secondary
veins of Cladophlebis sp., are not consistent with C. auriculipilosus. Furthermore, upon re-
examination of the original specimen of Cladophlebis sp., the specimen has a distinctly glabrous
rachilla, thus lacking the diagnostic trichomes on the rachillae of C. auriculipilosus.
<INSERT TABLE 2>
Sterile pinnules of extant members of Osmundales typically have better expression of the
venation on the abaxial, rather than adaxial, lamina surfaces (e.g. Osmunda claytoniana L.); if
we assume the same feature for the present specimens, then the prominent grooves on the
impression specimens of C. auriculipilosus (Fig. 6D) represent a mould of the abaxial surfaces
of the pinnae. Therefore, the consistent curvature of the pinnule surfaces would suggest revolute,
rather than involute, lamina margins. Furthermore, members of Osmundales with adnate and
discrete or basally fused pinnules, tend to exhibit sub-planar to revolute pinnule margins (e.g.
Osmundastrum cinnamomeum (L.) C. Presl). There are precedents of revolute pinnule margins
being preserved in various fossil ferns, and these have been attributed to original leaf
physiognomy rather than taphonomy (e.g. Rothwell, 1978; Delevoryas et al., 1992). The
inference of revolute lamina margins provides a method of orienting the frond relative to the
whole plant whilst it was in its living position, despite a lack of preserved cellular details. Where
the pinnule margins are superimposed, the adaxial surfaces of the pinnule auricles most often
underlie the decurrent acropetal pinnule margins (auricle underlap to overlap ratio = 72:29; data

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
20
pooled from both recovered impression samples). A reconstruction is presented (Fig. 7), which
highlights this tendency of the basiscopic margins of the pinnules to overlap the auricles of the
subtending pinnules.
<INSERT FIG. 7>
The pinna planes of the holotype are tilted 5–10° relative to the rachis axis; this angle was
likely greater prior to compaction under sedimentary overburden. All of the preserved pinnae
attached to a rachis have a consistent orientation, with the adaxial surfaces rotated towards the
frond apex. This observation is not a particularly useful diagnostic feature by itself, but this
consistent orientation may represent evidence of a heliotropic adaptation, as expressed by many
extant erect herbaceous ferns; e.g. Dryopteris erythrosora (D.C. Eaton) Kuntze has pinnae that
approach 90° from the plane of the rachis (Iino, 2001). Alternatively, such pinna orientation
could be the result of sedimentary processes, whereby the plant remains have become oriented
with respect to the flow direction during deposition (Bateman, 1999), as is common with oblate
pebble clasts in aqueous depositional environments with a unidirectional flow (SenGupta, 1966).
The fossiliferous beds from which C. auriculipilosus was recovered lacked any discernible
sedimentary structures with which to estimate palaeocurrent directions. Thus, it is not possible to
determine whether the preserved orientation of the pinnae whilst in outcrop was consistent with
the depositional current direction. If further specimens yield a consistent pinna orientation
relative to the rachis, regardless of their orientations relative to the strata, this would point
towards an inherent anatomical feature, consistent with an erect growth habit, rather than being
caused by taphonomic effects.
<INSERT FIG. 8>
Order incertae sedis
Sphenopteris (Brongniart, 1822) Sternberg, 1825

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
21
Type species
Sphenopteris elegans (Brongniart, 1822) Sternberg, 1825
Sphenopteris sp. cf. S. warragulensis McCoy in Stirling, 1892 (Fig. 8)
Synonymy
1995 Sphenopteris sp. cf. S. warragulensis McCoy in Stirling, 1892; McLoughlin et al., p.
283, fig. 4A.
Material examined
Fifteen samples comprising eleven individual specimens with four counterparts (PL1008–13,
PL1015–21, PL1023 and PL1024).
Description
Frond at least pinnate, base and apex not preserved. Preserved rachis length ≤ 62 mm, width ≤
2 mm, prominent longitudinal grooves, no lamina wing. Pinna attachment petiolulate,
distichously and alternately arranged at 10–18 mm intervals along rachis, insertion on rachis at ≤
35°. Pinnae are lanceolate with acute bases and alternate to sub-opposite, compound, pinnatifid
lobes. Lobes are ovate to lanceolate, have acute, lobulate to serrate distal margins (preserved
dimensions: ≤ 17 mm long by ≤ 5.5 mm wide); lobes are interconnected by narrow, interlobate
lamina (≤ 1.2 mm total width) parallel to primary vein (≤ 0.3 mm wide). Venation is
sphenopteroid; primary vein straight to sinuous, preserved length ≤ 39 mm. Single secondary
veins enter each lobe, diverging from the primary vein at ≤ 30°, spaced at ≤ 14 mm intervals,
and persisting to the distal margin. Secondary veins straight to sinuous, ≤ 0.2 mm wide. Tertiary

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
22
veins dichotomise from secondary veins at ≤ 30°, dichotomies spaced at ≤ 2.3 mm intervals.
Tertiary veins dichotomise ≤ six times into lateral veinlets, each terminating at the acute tips of
the distal serrate/lobulate margins; lateral veinlets do not dichotomise. The initial tertiary veins
of each lobe are catadromic. No fertile material was preserved.
Comparison and remarks
The presently described specimens have lamina lobes of widely variable length, width, degree
of vein dichotomy and margin dissection; however, the values of these variables appear to fall
upon a continuous spectrum, suggesting natural intraspecific variation. Thus, there are no
adequate grounds to assign the present specimens to more than one taxon. This conservative
approach is supported by the large, well-preserved specimens of Sphenopteris warragulensis
McCoy, 1892 described by Drinnan and Chambers (1986) from Aptian–Albian strata of
southeastern Australia, which have remarkable intraspecimen variability. Specifically, the
proximal pinnae and associated pinnules were much more likely to be discrete than those closer
to the distal ends of the fronds, the latter becoming “pinnatifid rather than pinnate” (Drinnan and
Chambers, 1986, p. 28). The synonymous Sphenopteris sp. cf. S. warragulensis of McLoughlin
et al. (1995) has a pinna habit described as pinnulate, but under the presently employed
taxonomic scheme (Ellis et al., 2009), these pinnae would be described as lobate rather than
pinnulate, due to the distinctive interconnecting laminar tissue between alternating secondary
veins. The degree of pinnatification for the present taxon is variable across specimens, with most
showing distinct interlobate laminae (e.g. Fig. 8C, D), whilst some specimens have lobes that
approach a pinnulate form (e.g. Fig. 8E, F). Larger preserved specimens will likely reveal that
the variants of the present taxon merely represent different sterile portions of fronds of the same
species.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
23
The Aptian–Albian southeast Australian specimens of Sphenopteris warragulensis described
by Drinnan and Chambers (1986) featured tertiary veins (‘secondary veins’ therein), which
dichotomise only once or twice, whereas those from the Chatham Islands exhibit up to six
dichotomies; all other features of the presently described specimens fall within the prescribed
variations for S. warragulensis. The similarity of this taxon to fertile fronds of the
leptosporangiate S. warragulensis from southeastern Australia facilitates the assignment of this
taxon to Polypodiopsida, but no order can be definitively ascribed at present. This taxon differs
from S. travisii Stirling, 1900 ex Drinnan and Chambers, 1986 by having longer and more
closely-spaced lobes (pinnules). Unlike the present taxon, the tertiary-order veins in
Sphenopteris göepperti Dunker, 1846 do not dichotomise. The pinnae of S. nordenskjoeldii
Halle, 1913 emend. Rees and Cleal, 2004 exhibit a “pseudo-dichotomous dissection” (Halle,
1913, p. 26), a description that emphasises the lobate (rather than pinnate) arrangement of the
pinnae as seen in the present specimens. However, the veins of S. nordenskjoeldii consistently
terminate in lobules (‘secondary lobes’ of Rees and Cleal, 2004), whereas the present taxon
commonly has serrate lobe margins. The Tupuangi specimens exhibit slightly larger lobes than
Sphenopteris sp. cf. S. warragulensis from Cenomanian strata of Queensland, as described by
McLoughlin et al. (1995), but all other features are comparable.
<INSERT FIG. 9>
Adiantites Göeppert, 1836 emend. Kidston, 1923
Type species
Adiantites oblongifolius Göeppert, 1836 (lectotype chosen by Andrews, 1970)
Adiantites sp. (Fig. 9)

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
24
Synonymy
cf. 2007 Adiantopteris tripinnata Cladera et al., pp. 53, 54, fig. 4B, C.
Material examined
Twenty-nine specimens (no counterparts) as follows: PL1000, PL1001, PL1005–7, PL1022,
PL1025, PL1026, PL1030, PL1039–58.
Description
Rachis ≤ 95 mm long, ≤ 2.2 mm wide, sympodially branching, with shallow longitudinal
grooves and no lamina wing. Pinnae have a distichous and alternative to sub-opposite
arrangement on rachis (at intervals of ≤ 12 mm), and are dimorphic. Petiolules attach at lamina
margin, and typically diverge from rachis decurrently but can be reflexed up to 100° relative to
rachis (Fig. 9E, F). Pinnae have cuneate bases and are alternately pinnatilobate, each lobe has
serrate distal margins. A single primary vein (≤ 0.4 mm wide) enters each pinna, is persistent for
less than a quarter of total pinna length, dichotomising evenly four–ten times at 10–20° each.
Each veinlet terminates at the tip of a lobe distal margin serration, ≤ 27 per lobe, one–two veins
per millimetre along distal margin. Elongated interlobate laminae extend to sinuses, and are
bordered by long, sub-parallel veinlets (Fig. 9B). Most pinnae are arranged alternately,
petiolulate to sub-petiolulate, elliptical to oblong, ≤ 31 mm long (28 mm excluding petiolule)
and ≤ 17.5 mm wide), with four–five distinct lobes; the two (or three) lobes situated on the
acroscopic side of the leaf are consistently longer than the two (or three) basiscopic lobes
(basiscopic to acroscopic lobe length average ratio approximately 1:1.7, as measured from the
initial vein dichotomy to lobe apex; Fig. 9E, F). Second type of pinna (?terminal) is larger (≤ 40
mm long by ≤ 28 mm wide) than lateral pinnae, basal shape is concave to acuminate, five–seven

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
25
alternating distal lobes; lobes are mostly fused along their adjacent margins, basiscopic lobes
approach a pinnatifid habit. Lobes of ?terminal pinnae consistently increase in length distally
(with maximum pinna length equal to the axis of alternation; Fig. 9D), cf. lateral pinnae which
feature acroscopic lobes that are consistently longer than the basiscopic lobes. No fertile material
was preserved.
Comparison and remarks
At present, there are two common generic designations employed to categorise sterile leaf
taxa with foliage similar to the extant Adiantum L.: 1) Adiantites Göeppert, 1836 emend.
Kidston, 1923; and 2) Adiantopteris Vassilevskaja, 1963. Adiantites was initially erected to
comprise all stipitate, fan-shaped fossil leaves with the distinctive anadromous venation pattern
typical of Adiantum. As suggested by his choice of name, Göeppert (1836) suspected a fern
affinity for the genus. He based the diagnosis on various leaf taxa from upper Palaeozoic strata;
however, many of these taxa have since been shown to have probable gymnosperm affinities.
Support for this association include the preservation of Adiantites-type foliage in association
with the seed taxon Wardia White, 1904 (see discussion by White, 1936), and complex axis
vascularity (typical of Medullosales; Jennings, 1985). In terms of stratigraphic distribution, there
appear to be two distinct populations of extinct Adiantum-like taxa in the fossil record: a
Palaeozoic assemblage and a Mesozoic to late Cenozoic assemblage. Adiantopteris was erected
to reflect this, and was intended to apply only to the latter group (Vassilevskaja et al., 1963). In
contrast to the Palaeozoic specimens, the fertile counterparts of the Mesozoic–Cenozoic
Adiantum-like population have consistently featured marginal sori, reflecting a pteridophytic
affinity (e.g. Adiantites lindsayoides Sewell, 1904 emend. Drinnan and Chambers, 1986).
However, the diagnosis of Adiantopteris is not significantly differentiated morphologically from
Adiantites. Furthermore, as noted by Moseychik and Ryabinkina (2012), stratigraphic

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
26
discrepancy does not provide adequate grounds for the reclassification of Adiantites taxa because
it does not account for the possible co-occurrence of gymnosperm and fern groups with
comparable sterile foliage. As such, the presently described material is assigned to Adiantites
rather than Adiantopteris, the former genus having priority.
A frond featuring the in situ attachment of the two described pinna types is not available
amongst any of the preserved specimens. Until the probable terminal pinnae are found in
association with the rachises, the interpretation of a single leaf species with a dimorphic habit
must be regarded as tentative. Based on the pinnate (typically bipinnate) leaf habit of similar
taxa (e.g. Adiantites dispersus Douglas, 1973, Adiantopteris tripinnata Cladera et al., 2007), it is
probable that the observed pattern of increasing sinus depth towards the base of the larger leaves
would continue basiscopically until leaves are pinnatisect, and ultimately petiolate further
proximally along the rachis; this would result in dimorphic leaves with an imparipinnate
arrangement on each axis. The present material is too degraded to gain a satisfactory description
of the pinna arrangement, and precludes assignment to any definite species. Despite this, the
preserved features allow for the exclusion of the present material from some species of
Adiantites. Adiantites lindsayoides from Lower Cretaceous strata of southeastern Australia
(Seward, 1904; Drinnan and Chambers, 1986) has two veins emerging from the base of each
pinna (cf. one for the presently described taxon). The pinnae of A. dispersus are typically 10 mm
long (up to 20 mm), are ‘wedge-shaped’ to weakly lobate and have veins that dichotomise up to
twice (Douglas, 1973); in contrast, the lateral pinnae of Adiantites sp. are much larger, have a
consistent, well-developed pinnatilobate shape, and veins that dichotomise several times. The
present specimens resemble Adiantites sewardi Yabe, 1905 [= Adiantopteris sewardii (Yabe,
1905) Vassilevskaja, 1963] from Lower Cretaceous strata of Korea (Yabe, 1905), eastern Russia
(Krassilov, 1967) and Japan (Kimura et al., 1979), but feature more prominent interlobate
sinuses. Adiantopteris tripinnata, as described by Cladera et al. (2007), matches the presently

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
27
described taxon in all features except the prescribed size range; the specimens herein are all
consistently larger; however, this is not deemed a feature diagnostic enough to exclude the
synonymy outright, as the size differences could be due to a range of uncontrolled environmental
variables. Assignment of the present material to A. tripinnata is not possible until an individual
leaf with both axial and terminal pinnae is found. Specimens of Adiantites have been noted (as
cf. Adiantites and Adiantites sp.) from Cretaceous strata of the Antarctic Peninsula (Cantrill,
1997, and Leppe et al., 2007, respectively), but no formal descriptions were provided therein. To
date, there have been no previously reported specimens of Adiantites (or Adiantopteris) foliage
from New Zealand strata of any age.
Due to the palaeogeographic and geochronological similarity of the present specimens to
Cretaceous Gondwanan foliage of definite leptosporangiate fern affinity (e.g. A. lindsayoides),
and the absence of recorded Mesozoic Adiantites-type pteridosperm material, an assignment to
Polypodiopsida affinity is tentatively proposed for the present taxon until fertile material can
provide a firmer taxonomic placement. Furthermore, this discussion highlights the ambiguity
surrounding the genera presently employed for the assignment of fossil Adiantum-like foliage,
and a taxonomic revision of Adiantites and Adiantopteris is sorely needed.
5. Discussion
5.1 Implications for mid-Cretaceous floral ecology of Gondwana
The Cenomanian Tupuangi Formation flora is one of a handful of high-latitude floras from
the early Late Cretaceous of the Southern Hemisphere and, as such, gives important insights into
ecological changes from this part of the world. Cenomanian tropical sea-surface mean annual
temperatures were ~ 31–37°C, with a Cretaceous maximum during the Turonian (Bice et al.,

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
28
2006); the warming trend was accompanied by a low thermal gradient, whereby the polar
regions experienced the greatest degree of warming over this interval (Huber et al., 2002). This
pattern of warming was mirrored by the rise to dominance of angiosperms in the vegetation and
concomitant ecological changes. Early Cretaceous macrofloras at high southern latitudes are rich
in liverworts (Douglas, 1973; Drinnan and Chambers, 1986; Cantrill, 1997, 2000; McLoughlin
et al., 2002), bryophytes (Drinnan and Chambers, 1986; Cantrill, 2000), and a particularly high
abundance and diversity of ferns (Ettingshausen, 1887; Seward, 1904; Halle, 1913; Arber, 1917;
Edwards, 1934; Medwell, 1954; McQueen, 1955; Douglas, 1973; Drinnan and Chambers, 1986;
Cantrill and Webb, 1987; Cantrill, 1995, 1996, 1998, 2000; Nagalingum, 2003; Cantrill and
Nagalingum, 2005; Nagalingum and Cantrill, 2006; Cantrill and Poole, 2012). These groups
appear to have been impacted by the rise to dominance of angiosperms across the southern high
latitudes (Cantrill and Poole, 2002; Nagalingum et al., 2002) but the severity, timing and the
degree of specific group interactions are not fully understood due to the scarcity of macrofloral
assemblages. For example, Antarctic studies have compared late Albian floras with Coniacian
floras (Cantrill and Poole, 2002; Poole and Cantrill, 2006), but the Tupuangi Formation flora
represents a critical intervening interval and demonstrates that, although these groups were still
present, their diversities and abundances were low. Approximately coeval assemblages have
been recorded from the Albian–Cenomanian (the “Unnamed Formation” at Bajo de los Corrales;
Passalia et al., 2001; Kachaike Formation; Passalia, 2007a, 2007b) and Cenomanian–Coniacian
(Mata Amarilla Formation; Iglesias et al., 2007) of Patagonia, South America (see review by
Archangelsky et al., 2009). Other important coeval floras include those of the Winton
Formation, Queensland, Australia (late Albian–Cenomanian; see review by McLoughlin et al.,
2010), and the Warder Formation, South Island, New Zealand (Ngaterian; ~ late Albian to mid-
Cenomanian; McQueen, 1956; Daniel, 1989; Parrish et al., 1998). With the exception of the
Cenomanian–Coniacian Mata Amarilla flora, all of these assemblages exhibit a diverse and

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
29
abundant fern component, in conjunction with abundant conifers and subsidiary dicotyledonous
angiosperms. In contrast to these assemblages, the Tupuangi Formation macrofloral assemblage
appears to exhibit a depauperate fern component (this study) and a high diversity of
gymnosperms (Pole and Philippe, 2010); however, this conclusion is preliminary until
quantitative intergroup macrofloral data can be obtained. Compared to the Winton assemblage,
the approximately coeval palaeofloras of Patagonia show similarities in angiosperm morphology
(Mata Amarilla flora; Iglesias et al., 2007), and overall floristic components (Kachaike flora;
Passalia, 2007a). It has been noted that these locations represent similar latitudes during the mid-
Cretaceous (Iglesias et al., 2007), and both of these regions were at substantially lower latitudes
than the Chatham Islands during the Cenomanian (Fig. 2). The climatic differences associated
with this latitudinal discrepancy would likely contribute to the unique floral ecology of the
Tupuangi Formation.
The floras of the Warder Formation, middle Clarence Valley, New Zealand, share a similar
palaeolatitude and stratigraphic range with the Tupuangi flora. Despite this, there appears to be
markedly lower fern abundance and diversity in the Tupuangi flora, the most notable absence is
the osmundaceous Phyllopteroides Medwell, 1954 emend. Cantrill and Webb, 1987. From the
Warder flora, Daniel (1989) identified thirty specimens of this genus, and classified them into
four distinct morphological groups. Furthermore, there have been three taxa of Cladophlebis
reported from the Ngaterian Clarence Valley strata (upper Albian to mid-Cenomanian;
McQueen, 1956; Parrish et al., 1998), whereas there were no recorded Cladophlebis from the
Ngaterian strata of the Tupuangi Formation, and only one species (C. auriculipilosus sp. nov.)
from Arowhanan strata (upper Cenomanian). This discrepancy in macrofloral assemblages is
likely due to local palaeoenvironmental influences, as explored in section 5.2 (below).
Because of the uncertainty over their affinities, the Tupuangi fern taxa have limited value for
interpreting the palaeobiogeography of, and ecological interrelationships between, fern groups.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
30
The fern taxon with arguably the greatest taxonomic certainty is Cladophlebis auriculipilosus,
because of various associations of members of this genus with Osmundaceae. Under the broad
diagnosis presented by Brongniart (1849), Cladophlebis has been a used for many extinct
species of sterile fronds with pecopteroid venation (see Dijkstra and Van Amerom, 1981, for a
review). Various species of Cladophlebis [particularly C. australis (Morris, 1845) Walkom,
1917 and C. denticulata (Brongniart, 1828) Fontaine, 1889 emend. Harris, 1961] were
particularly conservative and successful (Phipps et al., 1998). These have long been recognised
throughout Mesozoic strata across Eastern Gondwana, which includes Australia (Seward, 1904;
Herbst, 1978), Antarctica (Halle, 1913; Gee, 1989) and New Zealand (Arber, 1917; McQueen,
1956; Parrish et al., 1998). An affinity with Osmundaceae has been consistently proposed for
Cladophlebis, based on associations with osmundaceous stems (e.g. Kidston and Gwynne-
Vaughan, 1907; Medwell, 1954) and in situ spores (e.g. Litwin, 1985). In Australia, the relative
abundance of osmundaceous spores shows a progressive decrease during the Early Cretaceous
until the Cenomanian/Turonian, before their abundance stabilises for the most part throughout
the Late Cretaceous (Dettmann et al., 1992; Nagalingum et al., 2002). This relative decrease is
concurrent with an accelerating angiosperm diversification trend at southern high latitudes
(Drinnan and Crane, 1990; Cantrill and Poole, 2002). These data support the notion that the
Osmundales were significantly and adversely affected by Early to early Late Cretaceous
angiosperm diversification in the Southern Hemisphere (Nagalingum et al., 2002). Preliminary
macrofloral data appear to reflect this trend: Cladophlebis is typically the most abundant
component of high southern palaeolatitude macrofloral assemblages during the Early Cretaceous
(e.g. Herbst, 1971; Douglas, 1973; Gee, 1989), followed by a relative paucity of these fossils
during the mid- to Late Cretaceous. The presently recorded occurrence of only two specimens of
a single species of Cladophlebis and total absence of other osmundaceous ferns (e.g.
Phyllopteroides) from the Cenomanian strata of the Chatham Islands, amidst numerous

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
31
specimens of Sphenopteris and Adiantites, lends support for the aforementioned floristic pattern.
However, this support should be considered provisional until a more robust floral
abundance/diversity data-set throughout the stratigraphic succession can be obtained.
Liverwort-rich floras have been recovered from various Cretaceous strata of Gondwana,
including Antarctica (Cantrill, 1997), Australia (Douglas, 1973), India (Banerji, 1989) and South
Africa (Anderson and Anderson, 1985). These commonly form mats of spreading thalloid forms
on regularly disturbed overbank beds of high-energy rivers (e.g. Falcon-Lang et al., 2001). In
contrast, the presently described non-vascular plants were found as solitary elements amongst
leaf litter beds, overlying intermittent deltaic flood deposits (Fig. 4A, B; see section 5.2). From
Alexander Island, Antarctica, specimens of Adiantites (Adiantites sp.) were recorded in
association with the liverwort Marchantites arcuatus Cantrill, 1997 in leaf litter beds
comparable to the Tupuangi flood deposits. Within this context, Cantrill (1997) interpreted
Adiantites as a herbaceous understorey component, and this interpretation is inferred for the
Adiantites taxon of the Tupuangi flora, due to the consistently abundant conifer component (C.
Mays, pers. obs., 2012), which likely formed the overstorey.
5.2 Palaeoenvironmental and taphonomic considerations
The Tupuangi Formation was deposited as part of a vast fluviodeltaic depositional system
along the southern margin of the Chatham Rise (Wood et al., 1989; Campbell et al., 1993)
during the Cenomanian–Turonian (Mildenhall, 1994; Mays and Stilwell, 2013). A wide range of
macrofloral remains, including conifers, ginkgoes and angiosperms (C. Mays and D.J. Cantrill,
pers. obs., 2012), in addition to the free-sporing plants recorded herein, were deposited in leaf
litter horizons. The fining -upward stacking patterns of fine-grained sediments at these localities
are typical of low-flow rate river/delta floodplains (Bridge, 2003). Preserved root systems

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
32
indicate the formation of soils; the soil horizons are characterised by laminated, diverse leaf
beds, or thicker compacted carbonised plant beds. These suggest vegetation re-establishment on
the exposed floodplain deposits, with substantial peats forming in some cases. The massive to
cross-bedded sandstone facies immediately overlying these fossiliferous intervals typically lack
the concave-up geometry typical of meandering river channels, and are interpreted as being
deposited during intermittent fluviodeltaic sheet flow events (Bridge, 2003). Apart from sporadic
root traces, the palaeosols appear to have no discernible pedogenic structures, and sedimentary
depositional structures (e.g. planar bedding, ripples) are typically preserved within 10 cm below
the fossiliferous palaeosol surface. Due to their low chroma and high organic carbon and
sulphide content, these fossil soils are considered gleysols or histosols, which are typical of
regions with a consistently high water table (Mack et al., 1993; Retallack, 2001). The
palaeoenvironmental regime that this suggests for the examined fossil sites is: 1) a fluvial setting
with a sustained high precipitation rate; or 2) a coastal delta with a high groundwater base level.
The intermittent influxes of microplankton in the microfloral record (Mays and Stilwell, 2013),
and a lack of channel structures associated with the deposits, support the latter environmental
interpretation.
The Warder Formation, middle Clarence Valley, and the Tupuangi Formation, the Chatham
Islands, have similar ages and palaeolatitudes (see section 5.1). Furthermore, the Chatham Rise
has been interpreted as primarily emergent throughout the Cenomanian (Wood et al., 1989;
Laird and Bradshaw, 2004), thus no major geographic barriers are inferred between the
Tupuangi and Warder localities during the interval of preservation. Presumably, the
aforementioned factors would promote floral homogeneity between these localities; however,
the macrofloral assemblages appear to be starkly different. The discrepancy between the floras
of the Warder and Tupuangi formations may be explained by biases associated with the different
environments of deposition. The alluvial deposits of the Warder Formation have been interpreted

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
33
as being deposited in a meandering river system with localised lacustrine and swampy settings
(Browne and Reay, 1993; Parrish et al., 1998), whereas the Tupuangi flora localities examined
herein likely represent paralic, deltaic settings (Campbell et al., 1993; Mays and Stilwell, 2013).
Leaf litter beds, like those typical of the Tupuangi and Warder floras, reveal only the floral
ecology proximal to the site of deposition (Burnham, 1989, 1994). As such, differences in
palaeoenvironment may introduce ecological biases by promoting preservation of riparian taxa
within the Warder flora, and flora adapted to marginal marine settings within the Tupuangi
Formation. Local ecological or taphonomic bias is also suggested by the palynological record.
Studies of spore/pollen dispersal in modern deltaic and fluvial systems (e.g. Müller, 1959;
Darrell, 1973; Chmura, 1994) have revealed that levee and overbank deposits can accumulate
large numbers of distally-sourced palynomorphs, and can thus be misleading when interpreting
the local floral ecology; however, deltaic spore/pollen diversity data from such sediments
provides a reliable approximation of the regional floral ecology (e.g. Moss et al., 2005). The
diverse spore assemblage of the Tupuangi Formation (Mildenhall, 1994; Mays and Stilwell,
2013; Mays, in review) belies the relative scarcity of free-sporing plants in the macrofloral
assemblage. However, most of the spore elements of the Tupuangi Formation are common to
coeval strata of mainland New Zealand, including those from middle Clarence Valley (Raine,
1984). This suggests there was a significant degree of ecological homogeneity across Zealandia
during the Cenomanian. As such, non-vascular plants, lycopods and ferns were much more
diverse in the Chatham Islands region than indicated by the locally-derived macrofloral record
represented by the presently reported fossil beds. The differences in fern diversity and
abundance between the Warder and Tupuangi assemblages is likely explained by biases caused
by local depositional settings, rather than climatic, evolutionary or geographic discrepancies.
6. Conclusions

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
34
The Chatham Islands region during the mid-Cretaceous was geographically situated between
the Australian and the Antarctic/South American sectors of Gondwana on the Panthalassan
margin, and is arguably the highest southern latitude (~ 75–80ºS) floral assemblage of the
Cenomanian–Turonian, an interval of intense global greenhouse conditions. Within this context,
the Tupuangi flora represents a crucial biogeographic data-point between the well-described
coeval floras of Australia and South America/Antarctica. Two non-vascular plants, one lycopod
and three taxa of probable fern affinity are described, including two newly described species
(Cladophlebis auriculipilosus sp. nov. and Muscites gracilis sp. nov.), representing the first
macrofloral fossil record of these groups from the Chatham Islands, New Zealand.
The preserved sedimentary record of the fossiliferous beds reflects a deltaic floodplain
environment with a consistently high water table. Plant fossils were typically preserved as leaf
litter mats or thin coal beds immediately overlying gleyed palaeosols, which formed from
immature, waterlogged soils.
The Tupuangi flora fern diversity and abundance is markedly lower compared to various
coeval lower palaeolatitude localities of eastern and western Gondwana. Of particular note is the
relative scarcity of osmundalean ferns compared to Early Cretaceous localities of the high
southern latitudes. This provides provisional support for the decline of this group during the
mid-Cretaceous in the wake of the diversification of angiosperms across high southern latitudes.
Despite an apparent lack of geographic barriers during the Cenomanian, the Tupuangi flora has
yielded a starkly different assemblage, and is relatively depauperate in fern taxa, compared to the
coeval flora of the Warder Formation, mainland New Zealand. This is inferred to be due, at least
in part, to local ecological and/or taphonomic biases associated with differences in depositional
environments between the two localities.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
35
Acknowledgements
The authors would like to acknowledge the help of volunteer laboratory and field assistants:
Elyse Butterfield, Andrew Giles, Cameron McKenzie, David Pickering and Jesse Vitacca.
Special thanks go to chief preparator, Chava Rodriguez, who directed the efforts to ‘bring life
back’ to these fossils. We would also like to thank Assoc. Prof. Andrew Drinnan of the School
of Botany, University of Melbourne, for his assistance with laboratory facilities. CM would like
to thank the residents of Pitt Island, especially Mr John Preece, Mr Bill Gregory-Hunt and Mrs
Dianne Gregory-Hunt for allowing access to the outcrops on their land, as well as Mr Ken
Lanauze and Mrs Judy Lanauze for their assistance on Pitt Island. Terry and Donna Tuanui
kindly provided support on Chatham Island before and after fieldwork on Pitt Island. Fieldwork
and research supported by a Monash University Faculty of Science Bridging Research Grant and
an Australian Research Council Linkage Grant awarded to JDS. Further financial support was
provided by an Australian Postgraduate Award scholarship awarded to CM.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
36
References:
Anderson, J.M., Anderson, H.M., 1985. Palaeoflora of southern Africa: prodromus of South
African megafloras Devonian to Lower Cretaceous. AA Balkema, Rotterdam, The
Netherlands, 424 pp.
Andrews, H.N., 1970. Index of Generic Names of Fossil Plants, 1820–1965. United States
Government Printing Office, Washington D.C., U.S.A., 354 pp.
Arber, E.A.N., 1917. The earlier Mesozoic floras of New Zealand. New Zealand Geological
Survey Paleontological Bulletin 6, 1–80.
Archangelsky, S., Barreda, V., Passalia, M.G., Gandolfo, M., Prámparo, M., Romero, E., Cúneo,
R., Zamuner, A., Iglesias, A., Llorens, M., Puebla, G.G., Quattrocchio, M., Volkheimer, W.,
2009. Early angiosperm diversification: evidence from southern South America. Cretaceous
Research 30, 1073–1082.
Arthur, M.A., Dean, W.E., Schlanger, S.O., 1985. Variations in the global carbon cycle during
the Cretaceous related to climate, volcanism, and changes in atmospheric CO2. In: Sundquist,
E.T., Broecker, W.S. (Eds.), Chapman Conference on Natural Variations in Carbon Dioxide
and the Carbon Cycle. American Geophysical Union, Tarpon Springs, Florida, pp. 504–529.
Balme, B.E., 1995. Fossil in situ spores and pollen grains: An annotated catalogue. Review of
Palaeobotany and Palynology 87, 81–323.
Banerji, J., 1989. Some Mesozoic plant remains from Bhuj Formation with remarks on the
depositional environment of beds. Palaeobotanist 37, 159–168.
Barron, E.J., 1983. A warm, equable Cretaceous: the nature of the problem. Earth-Science
Reviews 19, 305–338.
Bartling, F.G., 1830. Ordines Naturales Plantarum. Dieterich, Göttingen, Germany, 498 pp.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
37
Bateman, R.M., 1999. Particle orientation and palaeoenvironments. In: Jones, T.P., Rowe, N.P.
(Eds.), Fossil Plants and Spores: Modern Techniques. The Geological Society, London, U.K.,
pp. 199–205.
Beerling, D.J., Osborne, C.P., 2002. Physiological ecology of Mesozoic polar forests in a high
CO2 environment. Annals of Botany 89, 329–339.
Berchtold, F.v., Presl, J.S., 1820. O přirozenosti rostlin, aneb, Rostlinář. J. Kraus, Prague, 322
pp.
Berner, R.A., Kothavala, Z., 2001. GEOCARB III: A revised model of atmospheric CO2 over
Phanerozoic time. American Journal of Science 301, 182–204.
Bice, K.L., Birgel, D., Meyers, P.A., Dahl, K.A., Hinrichs, K.-U., Norris, R.D., 2006. A multiple
proxy and model study of Cretaceous upper ocean temperatures and atmospheric CO2
concentrations. Paleoceanography 21, PA2002.
Bradshaw, J.D., Weaver, S.D., Muir, R.J., 1996. Mid-Cretaceous oroclinal bending of New
Zealand terranes. New Zealand Journal of Geology and Geophysics 39, 461–468.
Bridge, J.S., 2003. Rivers and Floodplains: Forms, Processes, and Sedimentary Record.
Blackwell Science, Oxford, U.K., 491 pp.
Brongniart, A.T., 1822. Sur la classification et la distribution des végétaux fossiles en général.
Memoires du Museum d'Histoire Naturelle, Paris 8, 203–348.
Brongniart, A.T., 1828. Histoire des v g taux fossiles: ou, Recherches botani ues et g ologi ues
sur les v g taux renferm s dans les diverses couches du globe. G. Dufour et E. d'Ocagne,
Paris, France, 488 pp.
Brongniart, A.T., 1849. Tableaux des genres de végétaux fossiles considerés sous le point de vue
de leur classification botanique et de leur distribution géologique. Dictionnaire Universal
Histoire Naturelle 13, 1–127.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
38
Brown, C.A., 1960. What is the role of spores in fern taxonomy? American Fern Journal 5, 6–
14.
Browne, G.H., Reay, M.B., 1993. The Warder Formation: Cyclic fluvial sedimentation during
the Ngaterian (late Albian–Cenomanian) of Marlborough, New Zealand. New Zealand
Journal of Geology and Geophysics 36, 27–35.
Burger, D., 1980. Early Cretaceous (Neocomian) microplankton from the Carpentaria Basin,
northern Queensland. Alcheringa 4, 263–279.
Burnham, R.J., 1989. Relationships between standing vegetation and leaf litter in a paratropical
forest: Implications for paleobotany. Review of Palaeobotany and Palynology 58, 5–32.
Burnham, R.J., 1994. Patterns in tropical leaf litter and implications for angiosperm paleobotany.
Review of Palaeobotany and Palynology 81, 99–113.
Campbell, H.J., Andrews, P.B., Beu, A.G., Maxwell, P.A., Edwards, A.R., Laird, M.G.,
Hornibrook, N.D.B., Mildenhall, D.C., Watters, W.A., Buckeridge, J.S., Lee, D.E., Strong,
C.P., Wilson, G.J., Hayward, B.W., 1993. Cretaceous–Cenozoic Geology and Biostratigraphy
of the Chatham Islands, New Zealand. Institute of Geological and Nuclear Sciences Limited,
Lower Hutt, New Zealand, 269 pp.
Cantrill, D.J., 1995. The occurrence of the fern Hausmannia Dunker (Dipteridaceae) in the
Cretaceous of Alexander Island, Antarctica. Alcheringa 19, 243–254.
Cantrill, D.J., 1996. Fern thickets from the Cretaceous of Alexander Island, Antarctica
containing Alamatus bifarius Douglas and Aculea acicularis sp. nov. Cretaceous Research 17,
169–182.
Cantrill, D.J., 1997. Hepatophytes from the Early Cretaceous of Alexander Island, Antarctica:
systematics and paleoecology. International Journal of Plant Sciences 158, 476–488.
Cantrill, D.J., 1998. Early Cretaceous fern foliage from President Head, Snow Island, Antarctica.
Alcheringa 22, 241–258.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
39
Cantrill, D.J., 2000. A Cretaceous (Aptian) flora from President Head, Snow Island, Antarctica.
Palaeontographica Abteilung B 253, 153–191.
Cantrill, D.J., Falcon-Lang, H.J., 2001. Cretaceous (Late Albian) coniferales of Alexander
Island, Antarctica. 2. Leaves, reproductive structures and roots. Review of Palaeobotany and
Palynology 115, 119–145.
Cantrill, D.J., Nagalingum, N.S., 2005. Ferns from the Cretaceous of Alexander Island,
Antarctica: Implications for Cretaceous phytogeography of the Southern Hemisphere. Review
of Palaeobotany and Palynology 137, 83–103.
Cantrill, D.J., Poole, I., 2002. Cretaceous patterns of floristic change in the Antarctic Peninsula.
In: Crame, J.A., Owen, A.W. (Eds.), Palaeobiogeography and Biodiversity Change: The
Ordovician and Mesozoic-Cenozoic Radiations. Geological Society, London, U.K., pp. 141–
152.
Cantrill, D.J., Poole, I., 2012. The Vegetation of Antarctica through Geological Time.
Cambridge University Press, Cambridge, U.K., 480 pp.
Cantrill, D.J., Webb, J.A., 1987. A reappraisal of Phyllopteroides Medwell (Osmundaceae) and
its stratigraphic significance in the Lower Cretaceous of eastern Australia. Alcheringa 11, 59–
85.
Chmura, G.L., 1994. Palynomorph distribution in marsh environments in the modern Mississippi
Delta plain. Geological Society of America Bulletin 106, 705–714.
Christenhusz, M.J.M., Zhang, X.-C., Schneider, H., 2011. A linear sequence of extant families
and genera of lycophytes and ferns. Phytotaxa 19, 7–54.
Chu, M.C.-Y., 1974. A comparative study of the foliar anatomy of Lycopodium species.
American Journal of Botany 61, 681–692.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
40
Cladera, G., Del Fueyo, G.M., Villar de Seoane, L., Archangelsky, S., 2007. Early Cretaceous
riparian vegetation in Patagonia, Argentina. Revista del Museo Argentino de Ciencias
Naturales, Nueva Serie 9, 49–58.
Crampton, J.S., Hollis, C.J., Raine, J.I., Roncaglia, L., Schiøler, P., Strong, C.P., G.J., W., 2004.
Chapter 10, Cretaceous. In: Cooper, R.A. (Ed.), The New Zealand Geological Timescale.
Institute of Geological and Nuclear Sciences Institute of Geological and Nuclear Sciences,
Lower Hutt, New Zealand, pp. 103–122.
Crandall-Stotler, B., Stotler, R.E., 2000. Morphology and classification of the Marchantiophyta.
In: Shaw, A.J., Goffinet, B. (Eds.), Bryophyte Biology. Cambridge University Press,
Cambridge, U.K., pp. 21–70.
Crandall-Stotler, B., Stotler, R.E., Long, D.G., 2009. Phylogeny and classification of the
Marchantiophyta. Edinburgh Journal of Botany 66, 155–198.
Crane, P.R., Friis, E.M., Pedersen, K.R., 1995. The origin and early diversification of
angiosperms. Nature 374, 27–33.
Cronquist, A., Takhtajan, A., Zimmermann, W., 1966. On the higher taxa of Embryobionta.
Taxon 15, 129–134.
Daniel, I.L., 1989. Taxonomic investigations of elements from the Early Cretaceous megaflora
from the Middle Clarence Valley, New Zealand. Ph.D. thesis, University of Canterbury,
Christchurch, New Zealand, 256 pp.
Darrell, J.H., 1973. Statistical evaluation of palynomorph distribution in the sedimentary
environments of the modern Mississippi River Delta. Ph.D. thesis, Louisiana State University,
Baton Rouge, Louisiana, U.S.A., 477 pp.
Delevoryas, T., Taylor, T.N., Taylor, E.L., 1992. A marattialean fern from the Triassic of
Antarctica. Review of Palaeobotany and Palynology 74, 101–107.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
41
Dettmann, M.E., 1986. Early Cretaceous palynoflora of subsurface strata correlative with the
Koonwarra Fossil Bed, Victoria. In: Jell, P.A., Roberts, J. (Eds.), Plants and Invertebrates
from the Lower Cretaceous Koonwarra Fossil Bed, South Gippsland, Victoria. Association of
Australasian Palaeontologists, Sydney, Australia, pp. 79–110.
Dettmann, M.E., 1994. Cretaceous vegetation: the microfossil record. In: Hill, R.S. (Ed.),
History of the Australian Vegetation: Cretaceous to Recent. Cambridge University Press,
London, pp. 143–170.
Dettmann, M.E., Molnar, R.E., Douglas, J.G., Burger, D., Fielding, C., Clifford, H.T., Francis,
J., Jell, P., Rich, T., Wade, M., Vickers-Rich, P., Pledge, N., Kemp, A., Rozefelds, A., 1992.
Australian Cretaceous terrestrial faunas and floras: biostratigraphic and biogeographic
implications. Cretaceous Research 13, 207–262.
Dettmann, M.E., Thomson, M.R.A., 1987. Cretaceous palynomorphs from the James Ross
Island area, Antarctica – a pilot study. British Antarctic Survey Bulletin 77, 13–59.
Dijkstra, S.J., Van Amerom, H.W.J., 1981. Filicales, Pteridospermae, Cycadales, 2. Supplement
A-C. Kugler Publications, Amsterdam, The Netherlands, 144 pp.
Douglas, J.G., 1973. The Mesozoic floras of Victoria. Part 3. Geological Survey of Victoria
Memoirs 29, 1–310.
Douglas, J.G., Williams, G.E., 1982. Southern polar forests: The Early Cretaceous floras of
Victoria and their palaeoclimatic significance. Palaeogeography Palaeoclimatology
Palaeoecology 39, 171–185.
Drinnan, A.N., Chambers, T.C., 1986. Flora of the Lower Cretaceous Koonwarra fossil bed
(Korumburra Group), South Gippsland, Victoria. Memoirs of the Association of Australasian
Palaeontologists 3, 1–77.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
42
Drinnan, A.N., Crane, P.R., 1990. Cretaceous paleobotany and its bearing on the biogeography
of austral angiosperms. In: Taylor, T.N., Taylor, E.L. (Eds.), Antarctic Palaeobiology and its
Role in the Reconstruction of Gondwana. Springer-Verlag, New York, U.S.A., pp. 192–219.
Dunker, W., 1846. Monographie der Norddeutschen Wealdenbildung. Oehme und Müller,
Braunschweig, Germany, 83 pp.
Eagles, G., Gohl, K., Larter, R.D., 2004. High-resolution animated tectonic reconstruction of the
South Pacific and west Antarctic margin. Geochemistry, Geophysics, Geosystems 5, Q07002.
Edwards, W.N., 1934. Jurassic plants from New Zealand. Annals and Magazine of Natural
History 7, 81–109.
Ellis, B., Daly, D.C., Hickey, L.J., Johnson, K.R., Mitchell, J.D., Wilf, P., Wing, S.L., 2009.
Manual of Leaf Architecture. Comstock Publishing Associates, Ithaca, New York, U.S.A.,
190 pp.
Ettingshausen, C.v., 1887. Beiträge zur Kenntniss der fossilen flora Neuseelands. Denkschriften
der Kaiserlichen Akademie der Wissenschaften, Wien 53, 143–194.
Falcon-Lang, H.J., Cantrill, D.J., Nichols, G.J., 2001. Biodiversity and terrestrial ecology of a
mid-Cretaceous, high-latitude floodplain, Alexander Island, Antarctica. Journal of the
Geological Society 158, 709–724.
Francis, J.E., Poole, I., 2002. Cretaceous and early Tertiary climates of Antarctica: evidence
from fossil wood. Palaeogeography Palaeoclimatology Palaeoecology 182, 47–64.
Gee, C.T., 1989. Revision of the Late Jurassic/Early Cretaceous flora from Hope Bay,
Antarctica. Palaeontographica B 213, 149–214.
Göeppert, H.R., 1836. Die fossilen Farrenkriuter (systema filicum fossilium). Nova Acta
Leopoldina 17, 1–486.
Halle, T.G., 1913. The Mesozoic flora of Graham Land. Wissenschaftliche Ergebnisse der
Schwedischen Südpolar-Expedition 1901–1903 3 (14), 124 pp.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
43
Harris, J.G., Woolf-Harris, M., 2001. Plant Identification Terminology: An Illustrated Guide,
2nd ed. Spring Lake Publishing, Payson, Utah, U.S.A., 206 pp.
Harris, T.M., 1961. The Yorkshire Jurassic Flora, vol. 1: Thallophyta-Pteridophyta. British
Museum (Natural History), London, U.K., 212 pp.
Hay, R.F., Mutch, A.R., Watters, W.A., 1970. Geology of the Chatham Islands. New Zealand
Geological Survey Bulletin 83, 1–86.
Hayes, P.A., 1999. Cretaceous Angiosperm Leaf Floras from Antarctica. Ph.D. thesis, University
of Leeds, Leeds, U.K., 310 pp.
Herbst, R., 1971. Paleophytología Kurtziana III. 7. Revisión de las especies argentinas del
género Cladophlebis. Ameghiniana 8, 265–281.
Herbst, R., 1978. Revision de las especies Australásicas de Cladophlebis (Bgt). Facena 2, 3–28.
Holmes, W.B.K., 2003. The Middle Triassic megafossil flora of the Basin Creek Formation,
Nymboida Coal Measures, New South Wales, Australia. Part 3. Fern-like foliage.
Proceedings of the Linnean Society of New South Wales 124, 53–108.
Huber, B.T., 1998. Tropical paradise at the Cretaceous poles? Science 282, 2199–2220.
Huber, B.T., Hodell, D.A., Hamilton, C.P., 1995. Middle–Late Cretaceous climate of the
southern high latitudes: stable isotopic evidence for minimal equator-to-pole thermal
gradients. Geological Society of America Bulletin 107, 1164–1191.
Huber, B.T., Norris, R.D., MacLeod, K.G., 2002. Deep-sea paleotemperature record of extreme
warmth during the Cretaceous. Geology 30, 123–126.
Iglesias, A., Zamuner, A.B., Poiré, D.G., Larriestra, F., 2007. Diversity, taphonomy and
palaeoecology of an angiosperm flora from the Cretaceous (Cenomanian–Coniacian) in
southern Patagonia, Argentina. Palaeontology 50, 445–466.
Iino, M., 2001. Phototropism in higher plants. In: Häder, D.-P., Lebert, M. (Eds.),
Photomovement. Elsevier Science B.V., Amsterdam, The Netherlands, pp. 659–811.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
44
Jefferson, T.H., 1981. Palaeobotanical Contributions to the Geology of Alexander Island,
Antarctica. Ph.D. thesis, Cambridge University, Cambridge, U.K., 265 pp.
Jennings, J.R., 1985. Fossil Plants from the Mauch Chunk Formation of Pennsylvania:
Morphology of Adiantites antiquus. Journal of Palaeontology 59, 1146–1157.
Kidston, R., 1923. Fossil plants of the Carboniferous rocks of Great Britain. Memoirs of the
Geological Survey of Great Britain, Palaeontology 2, 377–522.
Kidston, R., Gwynne-Vaughan, D.T., 1907. On the fossil Osmundaceae, Part 1. Transactions of
the Royal Society of Edinburgh 45, 759–780.
Kimura, T., Shigeru, S., Takao, T., 1979. Early Cretaceous plants from the Tokurazawa
Formation, Gumma Prefecture, in the inner zone of northeast Japan. Transactions and
proceedings of the Palaeontological Society of Japan, New Series 114, 87–96.
Kramer, K.U., 1990. Osmundaceae. In: Kubitzki, K. (Ed.), The Families and Genera of Vascular
Plants, 1. Pteridophytes and Gymnosperms. Springer-Verlag, Berlin, Germany, pp. 197–200.
Krassilov, V.A., 1967. Early Cretaceous Flora from South Primorye and its Significance for
Stratigraphy. Nauka, Мoscow, U.S.S.R., 363 pp.
Laird, M.G., Bradshaw, J.D., 2004. The break-up of a long-term relationship: the Cretaceous
separation of New Zealand from Gondwana. Gondwana Research 7, 273–286.
Leppe, M., Michea, W., Muñoz, C., Palma-Heldt, S., Fernandoy, F., 2007. Paleobotany of
Livingston Island: The first report of a Cretaceous fossil flora from Hannah Point. In: Cooper,
A.K., Barrett, P., Stagg, H., Storey, B., Stump, E., Wise, W., Anderson, J., Barron, J., Bart,
P., Blankenship, D., Davey, F., Diggles, M., Finn, C., Fitzgerald, P., Florindo, F., Francis, J.,
Futterer, D., Gamble, J., Goodge, J., Hammer, W., Helton, P., Ivins, E., Kyle, P.R.,
LeMasurier, W.E., Mayewski, P., Naish, T.R., Passchier, S., Pekar, S., Raymond, C., Ricci,
C.A., Studinger, M., Sugden, D., Thorn, V., Wilson, T. (Eds.), Antarctica: A Keystone in a
Changing World. The National Academies Press, Santa Barbara, California, U.S.A., pp. 1–4.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
45
Lidgard, S., Crane, P.R., 1988. Quantitative analyses of the early angiosperm radiation. Nature
331, 344–346.
Lidgard, S., Crane, P.R., 1990. Angiosperm diversification and Cretaceous floristic trends: a
comparison of palynofloras and leaf macrofloras. Paleobiology 16, 77–93.
Lindley, J., Hutton, W., 1833. The fossil flora of Great Britain: Or figures and descriptions of the
vegetable remains found in a fossil state in this country. James Ridgway, London, U.K., 218
pp.
Link, H.F., 1833. Hortus Regius Botanicus Berolinensis. G. Reimer, Berlin, Germany, 376 pp.
Litwin, R.J., 1985. Fertile organs and in situ spores of ferns from the Late Triassic Chinle
Formation of Arizona and New Mexico, with discussion of the associated dispersed spores.
Review of Palaeobotany and Palynology 44, 101–146.
Lupia, R., Lidgard, S., Crane, P.R., 1999. Comparing palynological abundance and diversity:
implications for biotic replacement during the Cretaceous angiosperm radiation. Paleobiology
25, 305–340.
Mack, G.H., James, W.C., Monger, H.C., 1993. Classification of paleosols. Geological Society
of America Bulletin 105, 129–136.
Markwick, P.J., Rowley, D.B., Ziegler, A.M., Hulver, M., Valdes, P.J., Sellwood, B.W., 2000.
Late Cretaceous and Cenozoic global palaeogeographies: mapping the transition from a "hot-
house" world to an "ice-house" world. GFF 12, 103.
Mays, C., Stilwell, J.D., 2013. Pollen and spore biostratigraphy of the mid-Cretaceous Tupuangi
Formation, Chatham Islands, New Zealand. Review of Palaeobotany and Palynology 192,
79–102.
McLoughlin, S., 1996. Early Cretaceous macrofloras of Western Australia. Records of the
Western Australian Museum 18, 19–65.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
46
McLoughlin, S., Drinnan, A.N., Rozefelds, A.C., 1995. A Cenomanian flora from the Winton
Formation, Eromanga Basin, Queensland, Australia. Memoirs of the Queensland Museum 38,
27–3313.
Mcloughlin, S., Jansson, I.-M., Vajda, V., in press. Megaspore and microfossil assemblages
reveal diverse herbaceous lycophytes in the Australian Early Jurassic flora. Grana.
McLoughlin, S., Pott, C., Elliott, D., 2010. The Winton Formation flora (Albian–Cenomanian,
Eromanga Basin): implications for vascular plant diversification and decline in the Australian
Cretaceous. Alcheringa 34, 303–323.
McLoughlin, S., Tosolini, A.-M.P., Nagalingum, N., Drinnan, A.N., 2002. The Early Cretaceous
(Neocomian) flora and fauna of the lower Strzelecki Group, Gippsland Basin, Victoria,
Australia. Memoirs of the Association of Australasian Palaeontologists 26, 1–144.
McQueen, D.R., 1955. Revision of supposed Jurassic angiosperms from New Zealand. Nature
175, 177–178.
McQueen, D.R., 1956. Leaves of Middle and Upper Cretaceous pteridophytes and cycads from
New Zealand. Transactions of the Royal Society of New Zealand - Geology 83, 673–685.
Medwell, L.M., 1954. A review and revision of the flora of the Victorian Lower Jurassic.
Proceedings of the Royal Society of Victoria 65, 63–111.
Mildenhall, D.C., 1994. Palynological reconnaissance of Early Cretaceous to Holocene
sediments, Chatham Islands, New Zealand, 1st ed. Institute of Geological and Nuclear
Sciences Limited, Lower Hutt, New Zealand, 204 pp.
Miller, K.G., Kominz, M.A., Browning, J.V., Wright, J.D., Mountain, G.S., Katz, M.E.,
Sugarman, P.J., Cramer, B.S., Christie-Blick, N., Pekar, S.F., 2005. The Phanerozoic record
of global sea-level change. Science 310, 1293–1298.
Moseychik, Y., Ryabinkina, N.N., 2012. To the knowledge of the Visean flora of Pechora, Ural
Mountains (in Russian). Lethaea Rossica 6, 2–22.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
47
Moss, P.T., Kershaw, A.P., Grindrod, J., 2005. Pollen transport and deposition in riverine and
marine environments within the humid tropics of northeastern Australia. Review of
Palaeobotany and Palynology 134, 55–69.
Mukasa, S.B., Dalziel, I.W.D., 2000. Marie Byrd Land, West Antarctica: Evolution of
Gondwana's Pacific margin constrained by zircon U-Pb geochronology and feldspar
common-Pb isotopic compositions. Geological Society of America Bulletin 112, 611–627.
Müller, J., 1959. Palynology of recent Orinoco Delta and shelf sediments Micropaleontology 5,
1–32.
Munsell, 1975. Munsell Soil Color Charts. Munsell Color, Baltimore, Maryland, U.S.A., 24 pp.
Murdock, A.G., 2008. Phylogeny of marattioid ferns (Marattiaceae): inferring a root in the
absence of a closely related outgroup. American Journal of Botany 95, 626–641.
Nagalingum, N.S., 2003. Cretaceous ferns from Australia and Antarctica. Ph.D. thesis,
University of Melbourne, Melbourne, Australia, 261 pp.
Nagalingum, N.S., Cantrill, D.J., 2006. Early Cretaceous Gleicheniaceae and Matoniaceae
(Gleicheniales) from Alexander Island, Antarctica. Review of Palaeobotany and Palynology
138, 73–93.
Nagalingum, N.S., Drinnan, A.N., Lupia, R., McLoughlin, S., 2002. Fern spore diversity and
abundance in Australia during the Cretaceous. Review of Palaeobotany and Palynology 119,
69–92.
Øllgaard, B., 1990. Lycopodiaceae. In: Kubitzki, K. (Ed.), The Families and Genera of Vascular
Plants, 1. Pteridophytes and Gymnosperms. Springer-Verlag, Berlin, Germany, pp. 31–39.
Oostendorp, C., 1987. The Bryophytes of the Palaeozoic and Mesozoic. J. Cramer, Berlin,
Germany, 112 pp.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
48
Panter, K.S., Blusztajn, J., Hart, S.R., Kyle, P.R., Esser, R., McIntosh, W.C., 2006. The origin of
HIMU in the SW Pacific: evidence from intraplate volcanism in southern New Zealand and
subantarctic islands. Journal of Petrology 47, 1673–1704.
Parrish, J.T., Daniel, I.L., Kennedy, E.M., Spicer, R.A., 1998. Paleoclimatic significance of mid-
Cretaceous floras from the middle Clarence Valley, New Zealand. Palaios 13, 149–154.
Passalia, M.G., 2007a. A mid-Cretaceous flora from the Kachaike Formation, Patagonia,
Argentina. Cretaceous Research 28, 830–840.
Passalia, M.G., 2007b. Nuevos registros para la flora cretácica descripta por Halle (1913) en
lago San Martín, Santa Cruz, Argentina. Ameghinana 44, 565–595.
Passalia, M.G., Romero, E.J., Panza, J.L., 2001. Improntas foliares del Cretácico de Santa Cruz,
Argentina. Ameghiniana 38, 73–84.
Phipps, C., Taylor, T.N., Taylor, E.L., Cúneo, R., Boucher, L.D., Yao, X., 1998. Osmunda
(Osmundaceae) from the Triassic of Antarctica: An example of evolutionary stasis. American
Journal of Botany 85, 888–895.
Pole, M.S., Philippe, M., 2010. Cretaceous plant fossils of Pitt Island, the Chatham group, New
Zealand. Alcheringa 34, 231–263.
Poole, I., Cantrill, D.J., 2006. Cretaceous and Cenozoic vegetation of Antarctica integrating the
fossil wood record. Geological Society, London, Special Publications 258, 63–81.
Pryer, K.M., Schuettpelz, E., Wolf, P.G., Schneider, H., Smith, A.R., Cranfill, R., 2004.
Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate
divergences. American Journal of Botany 91, 1582–1598.
Raine, J.I., 1984. Outline of a palynological zonation of Cretaceous to Paleogene terrestrial
sediments in West Coast region, South Island, New Zealand. New Zealand Geological Survey
Report 109, 1–82.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
49
Rees, P.M., Cleal, C.J., 2004. Lower Jurassic Floras from Hope Bay and Botany Bay,
Antarctica. Blackwell Publishing, London, U.K., 96 pp.
Retallack, G.J., 2001. Soils of the Past: An Introduction to Paleopedology, 2nd ed. Blackwell
Science, Oxford, U.K., 420 pp.
Rigby, J.F., 1977. New collections of Triassic plants from the Esk Formation, southeast
Queensland. Queensland Government Mining Journal 78, 320–325
Rothwell, G.W., 1978. Doneggia complura gen. et sp. nov., a filicalean fern from the Upper
Pennsylvanian of Ohio. Canadian Journal of Botany 56, 3096–3104.
Schellart, W.P., Lister, G.S., Toy, V.G., 2006. A Late Cretaceous and Cenozoic reconstruction
of the Southwest Pacific region: Tectonics controlled by subduction and slab rollback
processes. Earth-Science Reviews 76, 191–233.
Schneider, H., Schuettpelz, E., Pryer, K.M., Cranfill, R., Magallón, S., Lupia, R., 2004. Ferns
diversified in the shadow of angiosperms. Nature 428, 553–557.
Schuster, R.M., 1966. The Hepaticae and Anthocerotae of North America East of the Hundredth
Meridian, volume 1. Columbia University Press, New York, U.S.A., 802 pp.
Selling, O.H., 1944. Studies in the recent and fossil species of Schizaea, with particular reference
to their spore characters. Meddelanden fran Göteborgs Botanisk Trädgård 16, 1–112.
SenGupta, S., 1966. Studies on orientation and imbrication of pebbles with respect to cross-
stratification. Journal of Sedimentary Research 36, 362–369.
Seward, A.C., 1904. On a collection of Jurassic plants from Victoria. Records of the Geological
Survey of Victoria 1, 155–211.
Sternberg, G.K., 1825. Versuch einer geognostisch-botanischen Darstellung der Flora der
Vorwelt, vol. 1, pt. 4. Frederich Fleischer, Leipzig, Germany, 48 pp.
Stilwell, J.D., Consoli, C.P., Sutherland, R., Salisbury, S., Rich, T.H., Vickers-Rich, P.A.,
Currie, P.J., Wilson, G.J., 2006. Dinosaur sanctuary on the Chatham Islands, Southwest

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
50
Pacific: First record of theropods from the K–T boundary Takatika Grit. Palaeogeography
Palaeoclimatology Palaeoecology 230, 243–250.
Stirling, J., 1892. Notes on the fossil florula and faunae of the Gippsland carbonaceous area.
Victorian Department of Mines, Special Report 1, 10–13.
Tucker, M.E., 2011. Sedimentary Rocks in the Field: A Practical Guide, 4th ed. Wiley-
Blackwell, West Sussex, U.K., 276 pp.
Vassilevskaja, N.D., Vakhrameev, V.A., Vladimirovich, V.P., Dorofeyev, P.I., Kara-Murza,
E.N., Lepekhina, V., Novick, E., Radchenko, G.P., Sedov, M.A., Srebrodolskaya, N.N.,
Turutanova-Ketova, A.I., 1963. Subclass Leptofilitses. Leptosporangiate ferns. In:
Vakhrameev, V.A., Radchenko, G.P., Tahtadzhan, A.L. (Eds.), Algae, Bryophytes,
Psilophytes, Lycopsids, Sphenophytes and Ferns. Publishing House of the U.S.S.R. Academy
of Sciences, Moscow, U.S.S.R., pp. 569–601.
Veevers, J.J., 2006. Updated Gondwana (Permian–Cretaceous) earth history of Australia.
Gondwana Research 9, 231–260.
Villar de Seoane, L., 1996. Estudio morfológico de helechos fértiles hallados en la Formación
Baqueró (Cretácico Inferior), Provincia de Santa Cruz, Argentina. Revista Espanola de
Paleontologia 11, 83–90.
Villar de Seoane, L., 1999. Estudio comparado de Cyathea cyathifolia comb. nov. del Cretácico
Inferior de Patagonia, Argentina. Revista Espanola de Paleontologia 14, 157–163.
Wagstaff, B.E., Gallagher, S.J., Norvick, M.S., Cantrill, D.J., Wallace, M.W., 2013. High
latitude Albian climate variability: Palynological evidence for long-term drying in a
greenhouse world. Palaeogeography Palaeoclimatology Palaeoecology 386, 501–511.
Walkom, A.B., 1928. Fossil plants from Plutoville, Cape York Peninsula. Proceedings of the
Linnean Society of New South Wales 53, 145–150.
Walton, J., 1925. Carboniferous Bryophyta: I. Hepaticae Annals of Botany 39, 563–572.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
51
Wang, Y., Huang, C., Sun, B., Quand, C., Wu, J., Lin, Z., in press. Paleo-CO2 variation trends
and the Cretaceous greenhouse climate. Earth-Science Reviews.
White, C.M., 1936. Fossil Flora of the Wedington Sandstone Member of the Fayettevlle Shale.
United States Geological Survey Professional Papers. Government Printing Office,
Washington D.C., U.S.A., pp. 13–41.
Wolfe, J.A., Doyle, J.A., Page, V.M., 1975. The bases of angiosperm phylogeny: Paleobotany.
Annals of the Missouri Botanical Garden 62, 801–824.
Wood, R.A., Andrews, P.B., Herzer, R.H., Hornibrook, N.D.B., Hoskins, R.H., Beu, A.G.,
Maxwell, P.A., Keyes, I.W., Raine, J.I., Mildenhall, D.C., Wilson, G.J., Smale, D., Soong, R.,
Watters, W.A., 1989. Cretaceous and Cenozoic Geology of the Chatham Rise, South Island,
New Zealand. New Zealand Geological Survey, Lower Hutt, New Zealand, 75 pp.
Yabe, H., 1905. Mesozoic plants from Korea. Journal of the College of Sciences, Imperial
University of Tokyo 20, 1–89.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
52
Figure captions:
Figure 1: Geography of eastern Zealandia and the Chatham Islands. A) Map of modern eastern
Zealandia. Bold black outline = modern coastline; fine grey outline = 2000 m isobath. B) Map of
Chatham Islands. A detailed map of the studied fossil localities is provided in Figure 3. Adapted
from Campbell et al. (1993) with permission.
Figure 2: Palaeogeographic reconstruction of the Cenomanian (~ 95 Ma), south polar
perspective. Polar circle latitude is based on modern value (66° 33′ 44″S). Ant. = Antarctica;
Aus. = Australia; M. = Madagascar; S.A. = South America; Zeal. = Zealandia. Compiled from
Mukasa and Dalziel (2000), Schellart et al. (2006) and Veevers (2006). Emergent land areas are
modified from Markwick et al. (2000).
Figure 3: Geological map of Waihere Bay. This figure is the complementary inset map for
Figure 1. This map includes the fossil localities reported in this study.
Figure 4: Composite stratigraphic column of the Tupuangi Formation succession at Waihere
Bay, Pitt Island, with expanded stratigraphic logs of the coastal fossil localities reported in this
study. The stratigraphic column has been divided into lower (Fig. 4A) and upper (Fig. 4B) sub-
sections. Lithofacies are after the unit names of Hay et al. (1970), as discussed by Campbell et
al. (1993). New Zealand chronostratigraphic stages are defined by Crampton et al. (2004).
Stratigraphic columns and chronostratigraphic correlations are adapted from Mays and Stilwell
(2013). Ngat. = Ngaterian; Man. = Mangaotanean; ?Piri. = ?Piripauan; NZCS = New Zealand
Chronostratigraphic Stage; SST = sandstone.
Figure 5: Images of marchantiophyte, bryophyte and lycophyte specimens. A–C) Hepaticites
sp., PL1014. A) Microphotograph of specimen taken under ethanol, scale = 5 mm. B) Line
drawing of preserved thalli, dashed lines represent inferred margin, bold lines show where costae
are observed, shaded areas represent thalli that do not appear to be attached to the outlined
specimen, scale = 5 mm. C) Microphotograph taken under ethanol exhibiting notched apices

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
53
(arrows), scale = 0.5 mm. D–G) Muscites gracilis Mays and Cantrill sp. nov. D–F, holotype,
PL1027, scales = 2 mm. D) Composite micrograph of medial portion of stem exhibiting complex
branching habit. E) Distal portion of stem. F) Line drawing of distal portion of stem (as
illustrated in Fig. 5E), featuring examples of both dorsiventrally (D) and laterally (L) flattened
leaves, white areas represent leaves including costae where observed, dashed lines represent
inferred margins/costae of missing or obscured parts, shaded area represents the stem. G) Distal
portion showing costae along basiscopic margin of laterally flattened leaves, PL1034, scale = 1
mm. H) Lycopodites sp., preserved medial portion, PL1002, scale = 1 mm.
Figure 6: Cladophlebis auriculipilosus Mays and Tosolini sp. nov. holotype. A, B)
Compression, PL1036, scales = 10 mm. A) Possible basal portion of frond exhibiting alternating
organisation of pinnae. B) Venation detail under ethanol. C–E) Counterpart impression of
specimen illustrated in Fig. 6A, B, PL1035, scales = 2.5 mm. C) Microphotograph featuring the
elongated trichome bases on rachis and rachilla. D) Medial portion of pinna impression with
pinnule details, photograph taken with illumination at a low angle of incidence (from top of
image) to emphasise venation and curved lamina surface, arrows indicate probable revolute
sinuses on the apical sides of the auricles. Note that this impression represents the abaxial
surface of the pinna. E) Line drawing of pinna illustrated in Fig. 6D featuring pinnule basal
relationships including a tendency for the auricles to underlap the decurrent basiscopic margins
of the acropetal pinnules, dashed lines represent complementary details from compression,
shaded area represents the rachilla.
Figure 7: Artist’s reconstruction of the adaxial frond surface of Cladophlebis auriculipilosus
Mays and Tosolini sp. nov. Scale = 20 mm. Artist: Elyse Butterworth.
Figure 8: Sphenopteris sp. cf. S. warragulensis. A, B) Impression of degraded partial frond,
illustrating pinna arrangement, PL1015, scales = 1 mm. A) Photograph taken with low angle
light to accentuate venation details. B) Line drawing, shaded lobe is not attached to unshaded

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
54
frond. C, D) Partial pinna, PL1024, scales = 5 mm. C) Composite microphotograph, specimen
has been unevenly oxidised with only partial compression preserved. D) Line drawing showing
venation pattern and variably lobulate to serrate distal margins, shaded lobe may not have been
attached to non-shaded pinna. E, F) Impression of partial lobe, PL1019, scales = 2.5 mm. E)
Composite microphotograph. F. Line drawing.
Figure 9: Adiantites sp. A, B) Isolated (?terminal) pinna impression, PL1007, scales = 5 mm. A)
Microphotograph of specimen under ethanol. B) Line drawing, shaded areas indicate interlobate
lamina areas extending to sinuses, partitioning seven separate lobes. C, D) Isolated (?terminal)
pinna with preserved serrate margin details, PL1025, scales = 5 mm. C) Photograph. D) Line
drawing, AA = axis of alternation. E, F) Impression fossil of two leaves attached to rachis,
PL1005, scales = 5 mm. E) Photograph. F) Line drawing. G) Medial portion of rachis showing
arrangement of pinnae, PL1000, scale = 10 mm.
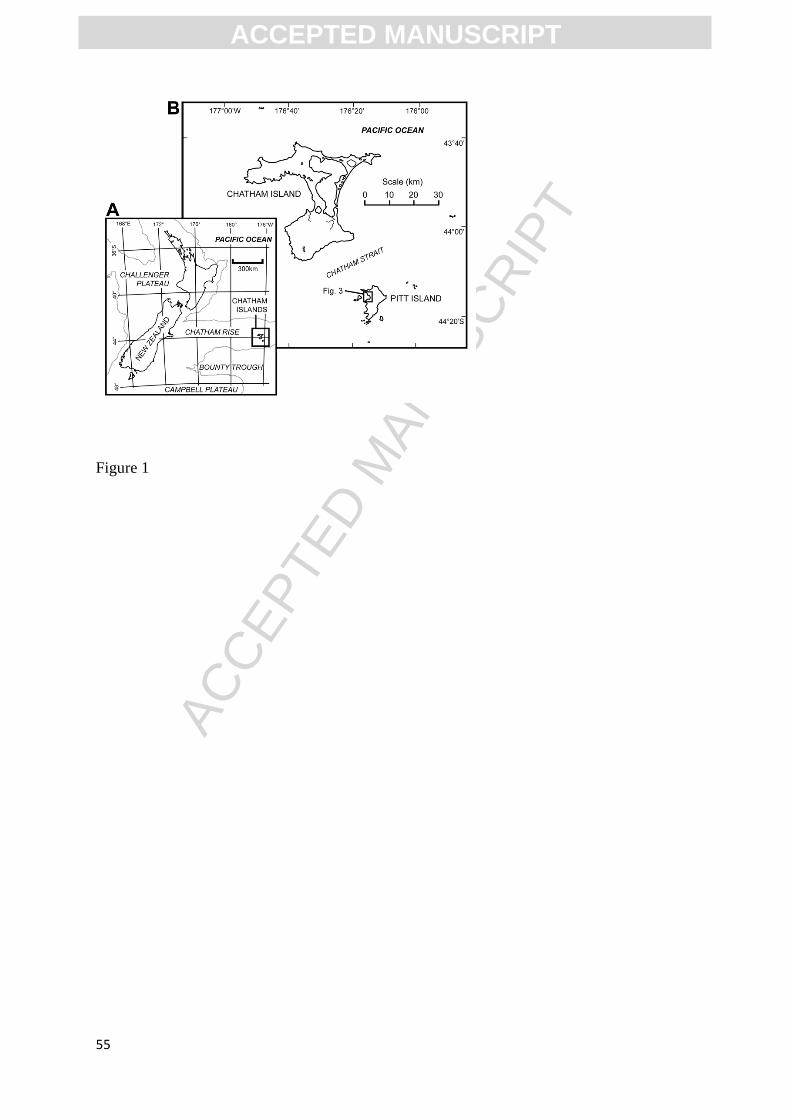
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
55
Figure 1
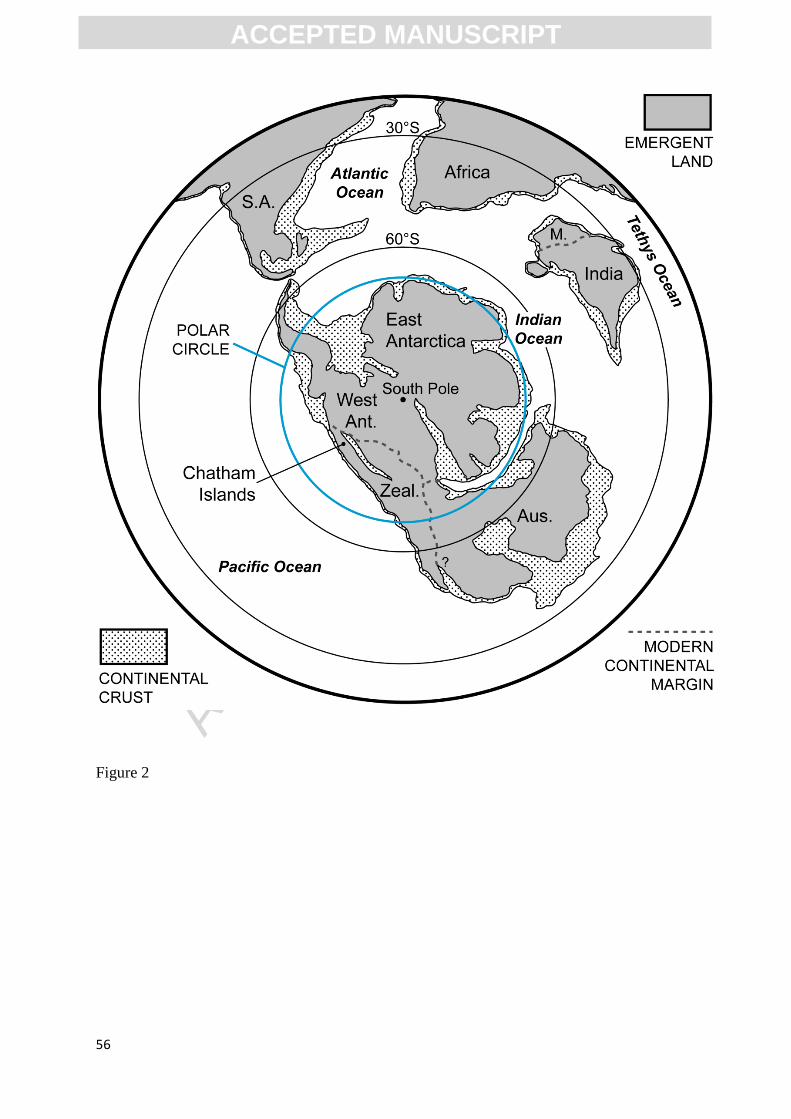
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
56
Figure 2
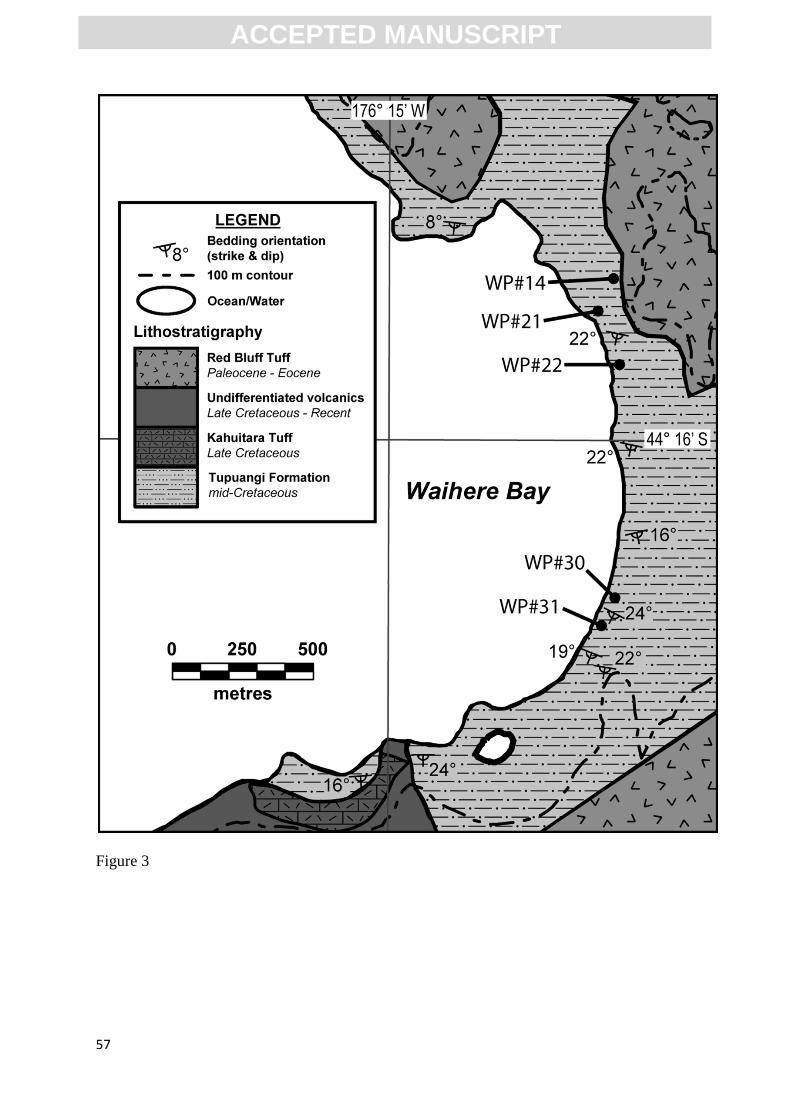
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
57
Figure 3
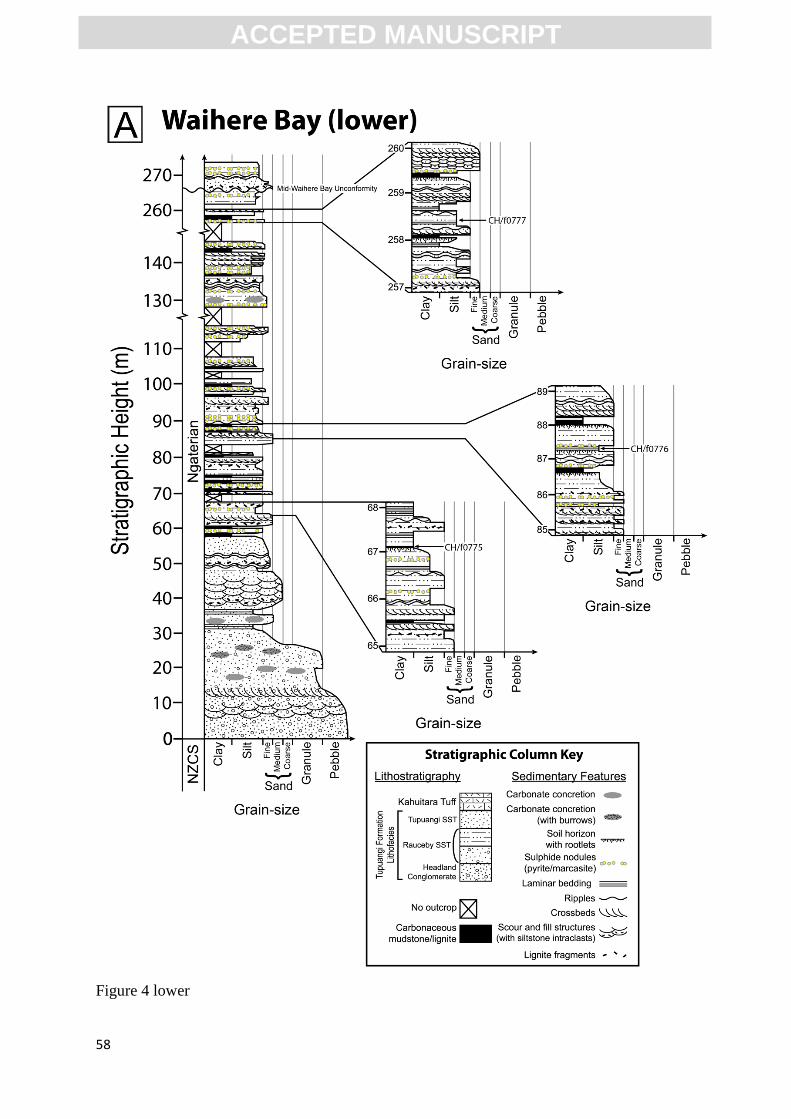
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
58
Figure 4 lower

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
59
Figure 4 upper

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
60
Figure 5

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
61
Figure 6

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
62
Figure 7

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
63
Figure 8

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
64
Figure 9

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
65
Table 2. List of Cladophlebis species recorded from probable Cretaceous strata of New Zealand. Locations for each species are listed in chronostratigraphic order; * = descriptive terms that have been substituted (as per Harris and Woolf-Harris, 2001, and Ellis et al., 2009) to allow for a standardised comparison; † = details not provided in original description, but inferred from illustrated specimen; 1 = Ettingshausen, 1890; 2
= Arber, 1917; 3 = McQueen, 1955; 4 = McQueen, 1956; 5 = Raine, 1981; 6 = Browne and Reay, 1993; 7 = Parrish et al., 1998; 8 = Raine in Kennedy, 2003; 9 = Mays and Stilwell, 2013.
Species Location Age Pinnule
arrangement
Pinnule
length
(mm)
Pinnule
width
(mm)
Pinnule
shape
Pinnule
apex
Pinnule
margin
Pinnule
attachment Comments
Cladophlebis sp. cf.
C. albertsi (Dunker
1846) Seward 1894
Waikato Heads
(2); Clarence
Valley (7)
"Neocomian" (4);
Cenomanian (6) sub-opposite ≤ 7 ≤ 4 falcate acuminate entire †
sessile,
adnate *
Cladophlebis
australis (Morris
1845) Walkom 1917
Waikato Heads
(2); Clarence
Valley (7);
Pakawau (2)
"Neocomian" (4);
Cenomanian (6);
Campanian/
Maastrichtian (3, 8)
not specified 7-33 3.5-8 lanceolate
to falcate
acute to
obtuse entire
sessile,
adnate *
fertile material
preserved
Cladophlebis
obscura
(Ettingshausen 1887)
McQueen 1956
Pakawau (1, 4) Campanian/
Maastrichtian (8)
alternate to
sub-opposite 25-30 5-6
lanceolate
* obtuse entire
sessile,
adnate *
Cladophlebis prisca
(Ettingshausen 1887)
McQueen 1956
Clarence Valley
(7); Pakawau
(1, 4)
Cenomanian (6);
Campanian/
Maastrichtian (3, 8)
alternate to
sub-opposite 25 6-7
lanceolate
*
acute to
obtuse entire
sessile,
adnate *;
acute insertion
Cladophlebis
wellmanii McQueen
1956
Paparoa Coal
Measures (4)
(?)Campanian/
Maastrichtian (5) N/A N/A N/A N/A N/A N/A N/A not pinnulate
Cladophlebis sp. Waikato Heads
(2) "Neocomian" (4)
?alternate to
sub-opposite † ≤ 9.5 † ≤ 6 †
lanceolate
†
?rounded
to obtuse †
?entire
†
sessile,
adnate †
single poorly
preserved
specimen

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
66
Cladophlebis
auriculipilosus Mays
and Tosolini sp. nov.
Pitt Island Cenomanian (9) sub-opposite ≤ 7.5 ≤ 4 ovate rounded entire sessile, partly
adnate

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
67
Graphical abstract

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
68
Late Cretaceous (Cenomanian–Turonian) macroflora from the
Chatham Islands, New Zealand: bryophytes, lycophytes and
pteridophytes
Chris Mays, Anne-Marie P. Tosolini, David J. Cantrill and Jeffrey D. Stilwell
Research highlights
The first record of Cretaceous non-seed macroflora from the Chatham Islands
Two new species described: Cladophlebis auriculipilosus and Muscites gracilis
Fossil fern content provisionally supports regional floral evolutionary trends