chondrichthyans from a cenomanian (late cretaceous) bonebed, saskatchewan, canada: canadian...
TRANSCRIPT

CHONDRICHTHYANS FROM A CENOMANIAN (LATE
CRETACEOUS) BONEBED, SASKATCHEWAN,
CANADA
by CHARLIE J. UNDERWOOD* and STEPHEN L. CUMBAA�*School of Earth Sciences, Birkbeck College, Malet Street, London WC1E 7HX, UK; e-mail [email protected]
�Earth Sciences, Canadian Museum of Nature, PO Box 3443 Stn ‘D’, Ottawa, ON K1P 6P4, Canada; e-mail [email protected]
Typescript received 20 August 2009; accepted in revised form 11 November 2009
Abstract: Acid preparation of samples of a bonebed from
the Cenomanian of central Canada yielded several thousand
well-preserved chondrichthyan teeth, in addition to numer-
ous other vertebrate remains. Teeth and other remains of
one species of chimaeroid, one species of hybodont shark,
three species of Ptychodus, 10 species of neoselachian sharks
and two species of batoid were recorded. The family Archae-
olamnidae fam. nov., genera Meristodonoides gen. nov. and
Telodontaspis gen. nov. and species Ptychodus rhombodus sp.
nov., Telodontaspis agassizensis gen et sp. nov., Eostriatolamia
paucicorrugata sp. nov., Roulletia canadensis sp. nov., Creto-
rectolobus robustus sp. nov. and Orectoloboides angulatus sp.
nov. are described. Status of the genus Palaeoanacorax and
the species Cretoxyrhina denticulata, Squalicorax curvatus and
‘Rhinobatos’ incertus are discussed, and reconstructed denti-
tions of Archaeolamna and Roulletia presented. The fauna is
of low diversity and dominated by active hunters, with many
species apparently endemic to the northern Western Interior
Seaway.
Key words: Canada, Cretaceous, hybodonts, lamniforms,
Ptychodus, ray, shark.
T he Western Interior Seaway was a broad and relatively
shallow body of water that, through the ‘mid’ and Late
Cretaceous, stretched across the North American conti-
nent from the Boreal Ocean in the North to the Tethys
Ocean in the south. Rocks deposited within the seaway
are well known for their fossil vertebrates, with chondri-
chthyans, teleosts, reptiles and birds all being well repre-
sented (e.g. Russell 1988, 1993). Despite these rich faunas,
study has been very uneven, and the sharks and rays have
received relatively little attention. In addition, it has been
recognized that there were strong biogeographical con-
trols on vertebrate distribution (Nicholls and Russell
1990), resulting in very different assemblages occurring in
rocks for the southern and northern parts of the seaway.
Chondrichthyan faunas from the Cenomanian are rela-
tively well known from Texas (e.g. Welton and Farish
1993; Cappetta and Case 1999), but far less so from the
mid and northern parts of the seaway. Abundant shark
teeth, mainly lamniforms, have been recorded from a
number of sites across the central and northern parts of
the USA (e.g. Cicimurri 2001a, b; Shimada et al. 2006),
but poor illustrations and conservative taxonomy have
sometimes rendered these difficult to interpret. Recorded
faunas from further north have typically been from shal-
low water to marginal facies (Case 2001, Cook et al.
2008). Cenomanian vertebrate faunas from more offshore
sites typically occur in bone-rich sandstones (e.g.
Cicimurri 2001a) or limestones (e.g. Cicimurri 2001b;
Shimada et al. 2006), presumably representing hiatal
horizons. Similar sandstones, limestones and mudstones
are also present in western Manitoba (McNeil and Cald-
well 1981, pers. obs.), whereas true bonebeds, usually with
a carbonate cement, occur in otherwise generally noncal-
careous-laminated mudstones in eastern Saskatchewan
(e.g. Cumbaa and Tokaryk 1999; Schroder-Adams et al.
1999, 2001). These bonebeds (Cumbaa and Tokaryk 1999;
Cumbaa et al. 2006) contain a very rich fauna dominated
by pelagic predatory vertebrates, in particular lamniform
sharks, enchodontid teleosts and hesperornithiform birds.
GEOLOGICAL SETTING
The Upper Cretaceous succession of the Western Interior
Seaway across North America has yielded Cenomanian
sharks from a number of sites throughout the region (as
summarized by Cook et al. 2008). The Cretaceous rocks
in central Canada rest unconformably on Palaeozoic and
Precambrian basement rocks and are dominated by mud-
stones, with coarser clastic material and carbonates both
[Palaeontology, Vol. 53, Part 4, 2010, pp. 903–944]
ª The Palaeontological Association doi: 10.1111/j.1475-4983.2010.00969.x 903

being very limited. On the eastern edge, this succession
forms the Manitoba Escarpment, a broken line of hills
and uplands running from south-western Manitoba and
terminating in the Pasquia Hills of eastern Saskatchewan.
The foot of the escarpment is largely obscured beneath
sediments deposited along the margins of glacial Lake
Agassiz. Exposures are limited, but a number of man-
made excavations and natural river cliffs are present.
Sandstones and siltstones of Albian age are overlain by a
series of mudstone formations ranging in age from Ceno-
manian to Late Campanian. These mudstones are typi-
cally laminated, and benthic organisms are very limited in
diversity, with only oysters and inoceramids being abun-
dant. Although vertebrate fossils are present throughout
the Upper Cretaceous succession, bone concentrations
and bonebeds are only known from the Belle Fourche
Member of the Ashville Formation. This unit is Cenoma-
nian in age, with the Cenomanian ⁄ Turonian boundary
being present within the lower part of the overlying Keld
member of the Favel Formation (see Cumbaa et al. 2006).
Although concentrations of vertebrate material have been
recorded at a number of localities in eastern Saskatche-
wan and western Manitoba, the bonebed from the expo-
sures on the Bainbridge River in the Pasquia Hills (e.g.
Cumbaa et al. 2006) has so far yielded the most abundant
vertebrate material (see Text-fig. 1). Material from other
sites is currently under study.
The main exposure on the Bainbridge River is a river
cliff known as Royal Saskatchewan Museum locality num-
ber 63E09-0003. Owing to the protected nature of the site,
full coordinates of the site are kept with the specimens at
the Royal Saskatchewan Museum. The majority of the cliff
comprises rocks of the Favel Formation (see Schroder-
Adams et al. 1999, fig. 7), with the Belle Fourche Member
largely obscured below scree. Small exposures do, however,
allow the stratigraphy to be recorded, and about 5 m of
carbonate-free laminated mudstones are seen. Within the
middle part of this are numerous discontinuous, thin beds
and starved ripples of very fine sandstone, many rich in fish
scales. Although the bonebed occurs within this interval, it
is rarely seen at outcrop. Poor exposures and fallen mate-
rial show that it comprises a level of very discontinuous
lenses within the Ostrea beloiti beds, a stratigraphic unit
associated with a widespread marker bentonite, believed to
be a northern continuation of the ‘X’ bentonite in the wes-
tern interior of the United States (Cumbaa and Bryant
TEXT -F IG . 1 . Map of the Manitoba
Escarpment, modified from Cumbaa
et al. (2006). The sections of Late
Cretaceous escarpment are: 1, Pasquia
Hills; 2, Porcupine Hills; 3, Duck
Mountain; 4, Riding Mountain; and 5,
Pembina Mountain. The sample site in
the Pasquia Hills region is marked with
a star.
904 P A L A E O N T O L O G Y , V O L U M E 5 3

2001; McNeil and Caldwell 1981). The only lenses found in
situ were just over 2 m below this marker bentonite (Cum-
baa and Bryant 2001). Bonebed lenses are up to 0.2 m thick
and often have a strongly scoured base and irregularly rip-
pled top. Large mudclasts of bentonite are common within
the lenses and sorting of clasts is strongly evident. The
bonebed is clast supported, with little or no matrix between
the clasts of bones, teeth, coprolites and phosphate
nodules, which are held together by calcite cement. Most
bioclasts in the bonebed are extremely well preserved. All
of the neoselachian teeth have their root preserved (with
the exception of those that appear to have been partly
digested, and there is very little abrasion). The general
lack of roots on hybodont teeth may be the result of
resorption. Many fragile bird long bones are complete, and
a large proportion of broken bones have longitudinal
breaks, suggesting early breakage prior to loss of organic
material. A small proportion of larger bones are highly
abraded, but this is almost unknown amongst smaller
specimens.
The Bainbridge River bonebed is sedimentologically
similar to one described from the Carrot River, 100 km
to the south-west (see Cumbaa et al. 2006), which has a
similar fauna but which is much more restricted (Phillips
2008). Both of these bonebeds lie within the ‘middle’ to
Late Cenomanian benthic foraminiferal Verneuilinoides
perplexus Zone (McNeil and Caldwell 1981), but radio-
metric age determinations have yet to be made on the
Bainbridge locality. The Carrot River bonebed is capped
by a bentonite dated by 40Ar ⁄ 39Ar at 95.17 Ma (Cumbaa
et al. 2006). Faunal and sedimentological characteristics
of the bioclastic units present elsewhere along the escarp-
ment have not yet been investigated, although they are
thought to be roughly synchronous.
MATERIAL AND METHODS
Samples of the Bainbridge River bonebed were collected
as loose blocks in the stream bed and banks. As clear size
sorting could be seen, the coarser-grained blocks were
preferentially collected, as these could be seen to contain
more large shark teeth and tetrapod bones. Some blocks
were weathered, and vertebrate fossils could be collected
from degraded heaps of bone material in the field, but
the majority were very well lithified and were collected
for later acid digestion.
Bonebed samples were broken down in buffered 5–10%
acetic acid, with the undissolved residue removed every
few days and washed and dried. Samples were sieved and
sorted down to 1-mm mesh size. For this study, about
20 kg of bonebed was dissolved and studied, with a fur-
ther large collection of specimens used for comparative
purposes. A single block of about 5 kg total weight
yielded 4432 identifiable shark teeth in the >1-mm mesh
size fraction.
Despite the large number of shark teeth within the
coarse and medium (>1 mm) sieve fractions, there were
very few identifiable remains of chondrichthyans in the
fine sieve fraction. Not only do the <1-mm sieve fractions
contain large quantities of fragmented teleost material,
making sorting for chondrichthyan material very time
consuming, but shark and ray teeth are genuinely very
rare in these fractions compared to the coarser material.
For these reasons, it was not practical to sort more than a
representative sample of the fine material, resulting in an
underrepresentation of the small-toothed taxa.
Larger specimens were ammonium chloride coated
before photography and greater focal depth was obtained
by digitally blending images of different focal distances.
Very small teeth were imaged by SEM, and therefore, have
a somewhat different appearance to optically photo-
graphed material and are presented on separate plates.
Dentitions were reconstructed by comparison with extant
taxa, but variations in the relative shapes of parasymphyse-
al and intermediate teeth and second and third upper ante-
rior teeth between extant taxa make the relative positions
of these teeth in the fossil reconstructions provisional.
Specimens were collected under Saskatchewan Heritage
Palaeontological Resource Investigation Permit # 06-03P,
and are deposited in the collections of the Royal Sas-
katchewan Museum, Regina, Saskatchewan, prefix RSM.
SYSTEMATIC PALAEONTOLOGY
Remarks. Descriptive tooth terminology largely follows
that of Cappetta (1987).
Class CHONDRICHTHYES Huxley, 1880
Subclass SUBTERBRANCHIALIA Zangerl, 1979
Order CHIMAERIFORMES (Berg, 1940) sensu Patterson, 1965
Family EDAPHODONTIDAE Owen, 1846
Genus EDAPHODON Buckland, 1838
Edaphodon sp.
Plate 1, figures 1–2
Material. One fragmentary palatine plate (P2989.43). Several
other indeterminate chimaeroid plate fragments may also be
referable to this taxon.
Description. The identifiable fragment is from the distal part of
the left palatine plate. In occlusal view, the anterior inner tritor
occupies over a third of the width of the plate and its entire
length, with neither end being preserved. The outer part of the
occlusal surface is concave and smooth, with the distal end of
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 905

the outer tritor being just visible at the posterior end of the
specimen. The symphyseal edge is smooth and short. The outer
part of the basal face of the plate is smoothly convex, becoming
flat or concave in the inner part. There is a well-developed
descending lamina which extends for half of the width of the
plate and is made up of tissue comprising packed longitudinal
tubes.
Remarks. There is no evidence to suggest that the frag-
ments of chimaeroid present here belong to more than
one species, although additional taxa may be present.
The overall morphology of the plates and the form of
the tritors suggests an edaphodontid affinity, and com-
parisons with better preserved coeval material by Evgeny
Popov suggested that they belonged to Edaphodon. All
of the remains seen belong to small tooth plates, proba-
bly not exceeding 50 mm long, contrasting to far larger
sizes reached by tooth plates of many of the species of
Edaphodon.
Subclass ELASMOBRANCHII Bonaparte, 1838
Cohort EUSELACHII Hay, 1902
Order HYBODONTIFORMES Maisey, 1989
Superfamily HYBODONTOIDEA Owen, 1846 sensu Zangerl,
1981
Family HYBODONTIDAE Owen, 1846
Subfamily HYBODONTINAE Owen, 1846
Genus MERISTODONOIDES gen. nov.
Derivation of name. From the general resemblance to the teeth
assigned to Meristodon by Agassiz (based on indeterminate mate-
rial), which has commonly been used for the genus named here.
Type species. Hybodus rajkovichi Case, 2001 from the Cenoma-
nian, Late Cretaceous of MN, USA.
Included species. It is considered here that several species can be
included into Meristodonoides gen. nov. These are as follows:
Hybodus butleri Thurmond, 1971 from the Aptian or Albian of
Texas; Hybodus montanensis Case, 1978 from the Campanian of
Montana and Wyoming; and Hybodus novojerseyensis Case and
Cappetta, 2004 from the Early Maastrichtian of New Jersey. A
number of other taxa described on fragmentary material are
probably also referrable to this genus.
Diagnosis. Hybodont shark known largely from teeth, but
partial fin spines and cephalic spines from sediments
where this genus occurs may be congeneric. There is a
low degree of heterodonty, which is mainly represented
by changes in the relative size of main cusp and lateral
cusplets and strength of ornamentation. Separated from
other genera by the following combination of characters:
(1) tooth close to symmetrical with single, well-developed
cusp being erect or slightly inclined lingually with only
slight distal inclination in lateral teeth; (2) main cusp
round to slightly labiolingually flattened in cross-section
with continuous but very weak cutting edge; (3) lateral
cusplets absent, incipient or very small relative to main
cusp and well separated from it; where they are present
there is a degree of heterodonty, presumably dignathic;
(4) ornament of longitudinal striae never coalescing to
form labial boss except very weakly in posterior teeth of
some species; (5) root very low and somewhat expanded
lingually, excavated on basal surface (6) root very strongly
vascularized, with row of many small foramina below
base of crown on labial face, but vascularization on the
basal face of root extremely variable. Fin spines probably
tuberculate, lacking strong longitudinal ridges.
Remarks. Teeth of this genus are separated from Hybodus
s.s. Agassiz, 1837 (H. reticulatus Agassiz, 1837 and similar
forms) by the presence of a single well-developed cusp,
very low root and, for some species, lack of a labial boss.
Teeth of Egertonodus Maisey, 1987 have multiple, well-
developed cusps but are in other respects similar. Teeth
of Planohybodus Rees and Underwood, 2008 have a simi-
lar overall shape but differ in being far more compressed,
having a far better developed cutting edge and having a
root with a basal face that is more strongly excavated
labially. Teeth of all other Jurassic and Cretaceous
hybodont genera have morphologies very different from
Meristodonoides gen. nov.
The subgenus Meristodon Agassiz, 1837 was erected for
a collection of isolated tooth crowns from the British
Early Cretaceous. The figures suggest that these are inde-
terminate, but are likely to be a mixture of teeth of Eger-
tonodus and Planohybodus. Despite this, the genus
Meristodon has been used to refer to teeth from the Late
Cretaceous (e.g. Glickman 1980; Underwood and Ward
EXPLANATION OF PLATE 1
Figs 1–2. Edaphodon sp.; ·5.5; P2989.43, partial palatine plate, 1, occlusal view. 2, basal view.
Figs 3–18. Meristodonoides rajkovichi (Case, 2001); all ·6.5. 3–5, P2989.44, posterolateral tooth. 3, labial view. 4, lingual view. 5, basal
view. 6–8, P2989.45, lower anterior tooth. 6, labial view. 7, basal view. 8, lingual view. 9–11, P2989.46, upper posterolateral tooth.
9, labial view. 10, lingual view. 11, basal view. 12–13, P2989.47, upper anterior tooth. 12, labial view. 13, lingual view. 14–15,
P2989.48, upper lateral tooth. 14, labial view. 15, lingual view. 16, P2989.49, lower anterior tooth, labial view. 17–18, P2989.50,
lower lateral tooth. 17, labial view. 18, lingual view.
906 P A L A E O N T O L O G Y , V O L U M E 5 3

PLATE 1
UNDERWOOD and CUMBAA, Edaphodon and Meristodonoides
1 2
3
6
5
4
7
8
9
10
11
12
16
18
15
17
14
11
13

2008), as the heterogeneous nature of the type material
had not been recognized.
The known range of Meristodonoides gen. nov. is from
Aptian or Albian (Thurmond 1971) to Early (e.g. Case and
Cappetta 2004) or possibly Late (Case 1979, as Hybodus
sp.) Maastrichtian. Most records of Meristodonoides gen.
nov. have been from within the Western Interior Seaway,
but it is also known from Kazakhstan (D. Ward, pers.
comm. 2008) and northern Europe (Biddle 1993, as Hybo-
dus sp.; Underwood and Ward 2008, as Meristodon sp.).
Meristodonoides rajkovichi (Case, 2001)
Plate 1, figures 3–18
?1993 Hybodus sp. Williamson et al., p. 449, fig. 3.1.
1999 Hybodus sp. 1. Cappetta and Case, p. 53, figs 1–2.
1999 Hybodus sp. Cumbaa and Tokaryk, p. 61, fig. 5
(pars, 8 teeth in left 2 columns).
2001 Hybodus rajkovichi Case, pl. 1, fig. 5, pl. 2, figs 1–2.
?2001b Hybodus sp. Cicimurri, p. 185, fig. 5a.
2006 Hybodus sp. Cumbaa et al., p. 146, fig. 4.1.
?2008 Hybodus sp. Cook et al., p. 1189, fig. 4A.
Material. About 2000 teeth studied including P2989.43–
P2989.50. Only 20 of the teeth studied have a root preserved.
Description. Teeth show weak monognathic and dignathic het-
erodonty. The crowns of teeth of this species are typically about
as wide as high, reaching about 8 mm. The tooth is relatively
symmetrical, with the main cusp in the centre of the crown. The
main cusp is 1.5–2 times as high as broad at the base and is the
only well-developed cusp on the majority of teeth. There are one
or two pairs of low to incipient lateral cusplets; where more
strongly developed the teeth are presumed to be (by analogy
with other hybodonts) from upper anterior positions. The main
cusp curves gently lingually and in some teeth has a very gentle
inclination towards the posterior. The labial face of the cusp is
gently convex, separated from a more strongly convex lingual
face by a small but continuous cutting edge. Labial ornament is
restricted to the lower quarter to half of the main cusp, where it
is represented by 6–12 longitudinal ridges, which become stron-
ger towards the base. In the lateral parts of the crown, these
ridges are similar or may be stronger and frequently reach the
apex of the cusplets or occlusal cutting edge. Ornament on the
lingual face is similar but finer, with the ridges sometimes reach-
ing nearly to the apex of the main cusp. The ornament reaches
the crown-root junction on the labial side and close to it on the
lingual face. The root is preserved in less than one per cent of
the teeth. The root is deeper (labiolingually) higher and wider
than the crown. The labial face of the root is very low, and flares
somewhat towards the base, where there is a large recess in the
basal face of the root on the labial side. A row of many very
small foramina are present just below the labial crown-root
junction. The root lingual face comprises a narrow and lingually
sloping shelf and a vertical face. In addition to a row of many
very small foramina just below the labial crown-root junction,
larger and more irregularly spaced foramina are present across
the root face. The basal root face is flat with a hollowed-out
recess. Very small foramina are present across the root basal
face, the number and size being very variable between individual
teeth. Larger and often rather elongate foramina are restricted to
the labial side of the root within the recess. Fragments of fin
spines tentatively referred to this taxon have an ornament of
regularly spaced, oval, tubercles.
Remarks. Many of the teeth of this species compare well
with type material but a greater degree of heterodonty
may be seen than previously recognized. Many of the
teeth described here have a finer and longer ornament of
fine ridges than the figured type assemblage (Case 2001),
but intermediate morphologies are present showing this
to be intraspecific variation. The presence of a proportion
of teeth with clearly developed lateral cusplets, but in all
other respects identical to the type material, suggests that
a greater degree of dignathic heterodonty is present
within this genus than previously recognized. This species
is very similar to Meristodonoides butleri (Thurmond
1971), but teeth have a higher and more slender cusp and
appear to have a weaker labial ornament and finer lingual
ornament (although it is hard to be sure from the draw-
ings of the type material).
The vast majority of well-preserved, isolated, post-
Triassic hybodont teeth lack a root, whereas teeth in
associated dentitions from the same deposits typically
have well-preserved roots (CJU, pers. obs.). This suggests
that the loss of the root is not commonly a result of
EXPLANATION OF PLATE 2
Figs 1–17. Ptychodus rhombodus sp. nov. Figs 1–10 · 4.75; Figs 11–17 · 9. 1–2, P2989.51, holotype, anteriolateral tooth. 1, occlusal
view. 2, lateral view. 3–4, P2989.52 lower symphyseal tooth. 3, occlusal view. 4, lateral view. 5–6, P2989.53? lower anterior tooth.
5, occlusal view. 6, lateral view. 7–8, P2989.54 lateral tooth. 7, occlusal view. 8, lateral view. 9–10, P2989.55 posterolateral tooth.
9, occlusal view. 10, lateral view. 11, P2989.56 posterolateral tooth, occlusal view. 12–13, P2989.57 posterolateral tooth. 12,
occlusal view. 13, lingual view. 14, P2989.58 posterolateral tooth, occlusal view. 15, P2989.59 upper symphyseal tooth, occlusal
view. 16–17, P2989.60 posterolateral tooth. 16, occlusal view. 17, lingual view.
Figs 18–21 Ptychodus sp.; all ·4.75. 18–19, P2989.62, lower symphyseal tooth. 18, occlusal view. 19, lateral view. 20–21, P2989.63
posterolateral tooth. 20, occlusal view. 21, lingual view.
908 P A L A E O N T O L O G Y , V O L U M E 5 3

PLATE 2
UNDERWOOD and CUMBAA, Ptychodus rhombodus
1
35
2
4
6
1211
9
7
810
16
1415 17
13
20
2119
18

taphonomic processes but was because of resorption of
the root-crown contact during tooth dehiscence.
Order INCERTAE SEDIS
Family PTYCHODONTIDAE Jaekel, 1898
Genus PTYCHODUS Agassiz, 1838
Type species. Ptychodus mammillaris Agassiz, 1839 from the Late
Cretaceous of Europe.
Ptychodus rhombodus sp. nov.
Plate 2, figures 1–17; Plate 8, figure 1
1999 Ptychodus sp. Cumbaa and Tokaryk, p. 61, fig. 5
(pars, 4 teeth in third column).
2006 Ptychodus anonymus Williston 1900; Cumbaa et al.,
p. 146, fig. 4.3.
Derivation of name. From strongly rhombic shape of most teeth.
Holotype. P2989.51 (Pl. 2, figs 1–2).
Additional material. One hundred and fifty-six teeth studied
including P2989.52–P2989.61.
Diagnosis. Teeth small and low. Strong heterodonty, sym-
physeal teeth quite square in occlusal view, all other teeth
asymmetrical and rhombic to some degree. Dentition with
many more files of small, rhombic, compressed lateral teeth
than larger and more square anterior teeth. Crown low and
outer part of crown flat, with small but conspicuous raised
‘cusp’ towards centre of anterior teeth, inclined to posterior
in lateral teeth. Ornament continues across all of occlusal
face with no clearly defined marginal area. Ornament of
four to 15 subparallel ridges (typically 6–10 in lateral teeth,
8–14 in anterior teeth), rarely bifurcating other than close
to edge, breaking up to tubercles at edges and often towards
labial margin of larger teeth. Root low and strongly lin-
gually inclined. Foramina below root-crown junction on all
sides, especially conspicuous on labial root face.
Description. Teeth are small for the genus, with the largest being
13 mm wide, and the majority less than 10 mm. They demon-
strate a high degree of heterodonty although very few symphy-
seal teeth are present. All teeth are wider than deep, although
this is only slightly so in symphyseal teeth; many of the smaller
lateral teeth are approximately twice as wide as deep. In occlusal
view, all but symphyseal teeth are strongly rhombohedral in
shape, with the lingual edge of the tooth displaced posteriorly.
The crown on all teeth is low, with the occlusal face being gently
convex but with a low but distinct swollen ‘cusp’ on all but the
smallest posterior teeth. In lateral teeth, this is on the posterior
side of the tooth. The sides of the crown are smoothly concave
on lateral and labial faces but there is a well-developed concave
notch on the lingual face. The ornament on the occlusal face is
rather variable but intermediate forms are present. The domi-
nant ornament are subparallel ridges running from the anterior
to posterior side of the crown. In anterior and anterolateral
teeth, there are between 10 and 15 ridges, with fewer present in
posterolateral teeth, with as few as four ridges in the most pos-
terior teeth. There are occasional examples of ridges bifurcating,
largely restricted to close to the lateral and labial margins of the
teeth. Towards the lingual edge of many teeth, the ridges
become more curved with the concave side towards the labial
face. The ridges reach close to the anterior and posterior edges
of the crown, breaking up into small tubercles at the crown
edge, especially in larger teeth. There is commonly an area of
broken ridges or tubercles towards the labial crown edge. There
is no marginal area to the crown, and ornament continues
unbroken to the crown edge. The crown overhangs the root on
all sides. The root is of similar height and narrower than the
crown. The root is strongly inclined lingually in all but symphy-
seal teeth, and in some lateral teeth the lingual margin of the
root extends out beyond the lingual margin of the crown. The
lateral faces of the root are all flat, and all have small foramina
just below the root-crown junction, although these are especially
well developed on the labial face. The basal face of the root is
flat and has small and irregularly spaced foramina, sometimes
with the addition of one or two larger foramina.
Remarks. These teeth are highly variable in overall shape
but intermediate morphologies demonstrate that they
clearly belong to a single taxon. The large proportion of
highly compressed, small and asymmetrical lateral teeth
has not been previously noted with any Ptychodus species;
articulated dentitions of Ptychodus decurrens Agassiz, 1838
(see Woodward 1912) do have several files of compressed
lateral teeth, but also possess several files of nearly sym-
metrical anterior teeth, whilst dentitions of other Ptycho-
dus species have proportionally far fewer reduced lateral
teeth. This species shares a lot of characteristics with
Ptychodus decurrens Agassiz, 1838 and P. anonymus Will-
iston, 1900, but can clearly be separated from these.
Teeth of P. decurrens share an almost identical occlusal
ornament to P. rhombodus sp. nov., but differ in having a
crown that has a far more evenly domed profile, lacking the
flat surface with central small cusp seen here, and a larger
maximum size. It is also probable that the dental formula
was different. Teeth of P. anonymus are also similar, but
differ in having a more clearly defined granular margin sur-
rounding the ridged occlusal ornament, and a higher and
more clearly defined cusp. Ptychodus janewayii Cope, 1874
was erected on some small and strongly rhombic teeth from
the Cenomanian of Kansas, which are considered here to
probably represent more than one species. Although it is
possible that at least one of the type series is conspecific
with the material described here (Williston 1900, fig. 9), the
910 P A L A E O N T O L O G Y , V O L U M E 5 3

two specimens mentioned in the diagnosis (therefore
including, by inference, the holotype) have only four and
five folds, respectively, far less than recorded on any similar
sized tooth of P. rhombodus sp. nov. As the low lateral and
posterior teeth of this taxon are not clearly differentiated
from those of P. anonymus, it is possible that P. rhombodus
sp. nov. has not been recognized in assemblages where its
remains are rare. It is likely that P. rhombodus sp. nov. and
P. anonymus are closely related, and it is possible that the
probable Middle Cenomanian P. rhombodus sp. nov. is
ancestral to the Late Cenomanian to Coniacian (Shimada
et al. 2006) P. anonymus.
Ptychodus sp.
Plate 2, figures 18–21
Material. One lower symphyseal tooth and one posterolateral
tooth (P2989.62 and P2989.63).
Description. The larger tooth is 12 mm wide and has a total
height of 10 mm, the smaller tooth is 5 mm wide. The crown of
the symphyseal tooth is roughly rectangular in occlusal view,
being somewhat expanded on the labial edge and having a
prominent indentation on the lingual edge. The majority of the
crown is taken up by a roughly conical cusp, which is somewhat
wider than high. This is surrounded by a narrow but distinct flat
marginal area on the lateral edges but is absent on the labial
edge. The posterolateral tooth is rather rhombic in profile, with
the posterior edge being ‘stretched out’. A prominent domed
cusp occupies the central region of the crown. The crown of
both teeth is ornamented by about 10 subparallel ridges, which
pass over the cusp, some bifurcating on the cusp flanks. In both
teeth, ornament fades out to a smooth outer edge of the crown,
although there has been some wear or abrasion of the lateral
parts of the larger tooth. The labial side of the crown of the lar-
ger tooth is ornamented by small but distinct granulation, which
is also present on the small area of vertical cusp face on the lin-
gual side. The crown of both teeth is flared basally and over-
hangs the root on all sides. The root is shallow, especially so in
the posterolateral tooth, and slightly displaced lingually. Foram-
ina of various sizes are present just below the crown-root junc-
tion on all sides but are especially prominent on the lingual face.
The basal face of the root is slightly concave and has several,
irregularly spaced, foramina of different sizes.
Discussion. Teeth are similar to those of both P. rhombo-
dus sp. nov. and P. anonymus, but appear to differ from
both. The symphyseal tooth is larger than any compara-
tive tooth of P. rhombodus sp. nov., and both teeth have
a far more conspicuous cusp and a more clearly defined
marginal area. It is, however, possible that these represent
atypical teeth of a large individual of P. rhombodus sp.
nov. The general shape and ornamentation are very
similar to that of P. anonymus but differ in the lack of a
well-defined granular marginal area.
Ptychodus ex. gr. decurrens Agassiz, 1838
Plate 3, figures 1–6
1839 Ptychodus decurrens Agassiz, p. 154, pl. 25b, figs 1–2,
4, 6–8.
1993 Ptychodus decurrens Agassiz, 1838; Welton and
Farish p. 59, figs 1–7.
1999 Ptychodus decurrens Agassiz, 1838; Cappetta and
Case p. 55, pl. 2, fig. 9.
?1999 Ptychodus decurrens Agassiz, 1838; Cappetta and
Case p. 55, pl. 2, figs 7–8.
2001a Ptychodus decurrens Agassiz, 1838; Cicimurri, p. 41,
fig. 6C–D.
2006 Ptychodus decurrens Agassiz, 1838; Shimada et al.
p. 7, fig. 6.3–6.4.
2006 Ptychodus decurrens Agassiz, 1838; Cumbaa et al.,
p. 146, fig. 4.5–4.6.
Material. Three teeth (P2989.64–P2989.66).
Description. The largest of the teeth referred to this species is
from an anterior position and has a maximum dimension of
12 mm, whilst the other, lateral and symphyseal, teeth are some-
what smaller. Both lateral teeth are somewhat asymmetrical and
rhombic in occlusal view, the anterior one having similar width
to depth, the other being slightly wider than deep. These lateral
teeth have a crown that is gently and evenly domed, without a
marginal area, with ornament continuing to the crown margins.
The symphyseal tooth is rather ‘arrowhead-shaped’ in occlusal
view, with a wide and concave lingual margin. The crown is
weakly domed, and the ornament does not reach the crown
margins. The crown ornament of the largest tooth comprises
about 10 well-developed ridges, which grade from smoothly
curved near the crown lingual edge to being folded back on
themselves near the labial edge. There is some bifurcation of the
ridges, and some breaking up of the ridges close to the crown
margins, but generally the ornament continues to the crown
edge. The ornament of the other teeth comprises numerous sub-
parallel, fine, ridges cover the lingual two-thirds of the crown,
with the lingual part being ornamented with irregularly broken
ridges and fine granulations. The crown of the largest tooth is
somewhat thicker than that of the others, but all have a similar
degree of overhang over the root. The edges of the crown are
convex on the labial and lateral edges, but concave on the lin-
gual side, especially so in the symphyseal tooth. The root is rela-
tively low and displaced to the lingual edge. Foramina are
present both close to the crown-root junction and on the root
basal face, with numerous small foramina and scattered larger
ones being present in both places.
Discussion. These teeth agree well with previous descrip-
tions of the species. The teeth considered to belong to
this species have rather different ornament, but all share a
similar tooth shape and fall within the morphological
range of teeth of P. decurrens. Although several subspecies
of P. decurrens have been named (e.g. Herman 1977), it
has been shown that ornament is very variable amongst
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 911

teeth from the same stratigraphical interval, and even
within the same articulated dentitions (e.g. Woodward
1912). The two teeth with the finer ornament are P. de-
currens var. oweni of Woodward 1912. Despite this varia-
tion, it is possible that more than one species or form
exists, as teeth from the Turonian ⁄ Coniacian boundary
(Cappetta and Case 1999) appear to have fewer and
stronger ridges on the crown than teeth figured from the
Albian to Turonian.
Subcohort NEOSELACHII Compagno, 1977
Order LAMNIFORMES Berg, 1958
Family CRETOXYRHINIDAE Glickman, 1958
Genus CRETOXYRHINA Glickman, 1958
Type species. Oxyrhina mantelli Agassiz, 1843 from the Late
Cretaceous of England.
Remarks. Teeth of Cretoxyrhina mantelli have been
recorded as extremely widespread in rocks of Albian to
Campanian age (for full synonymy and records of occur-
rence see Herman 1977; Siverson 1992, 1996; Siverson
and Lindgren 2005). Throughout the temporal range of
the species, distinct changes in tooth morphology occur,
with specimens from older rocks generally being smaller,
more gracile and having lateral cusplets in many tooth
positions, contrasting with the larger, more robust teeth
typically lacking lateral cusplets present in specimens
from younger rocks (e.g. Siverson 1996; Siverson and
Lindgren 2005). Cretoxyrhina denticulata (Glickman,
1957) was erected for Cenomanian material within which
lateral cusplets are present in posterolateral teeth. The
diagnosis of C. denticulata was vague, and it has been
unclear whether these are a temporal morph of C. man-
telli or represent a recognizably different species. Siverson
(1996) appears to suggest that the two are synonymous
but does not include the type material of C. denticulata
in the synonymy of C. mantelli. Reexamination of some
of the syntype specimens of C. mantelli (CJU, pers. obs.)
suggests that C. denticulata represents a species that can
be readily separated from C. mantelli.
Cretoxyrhina denticulata (Glickman, 1957)
Plate 3, figures 7–22
1912 Oxyrhina mantelli Agassiz; Woodward, plate XLIII,
figs 11–13.
1957 Isurus denticulatus Glickman, p. 105, figs 1–4,
6–17.
?1993 Cretoxyrhina mantelli (Agassiz); Welton and Farish
p. 101, figs 1–12, p. 102, figs 1–2.
?1996 Cretoxyrhina mantelli (Agassiz); Siverson, p. 821,
figs 1–18.
2000 Pseudisurus denticulatus (Glickman); Zhelezko,
plate II, figs 1–5.
?2001a Cretoxyrhina mantelli (Agassiz); Cicimurri, p. 42,
fig. 7.i–k.
?2001b Cretoxyrhina mantelli (Agassiz); Cicimurri, p. 185,
fig. 5j.
2006 Cretoxyrhina mantelli (Agassiz); Cumbaa et al.,
p. 146, fig. 4.13.
2006 Cretoxyrhina mantelli (Agassiz); Shimada et al.,
p. 15, fig. 10.9–10.10.
2008 ?Cretoxyrhina cf. C. mantelli (Agassiz); Cook et al.,
p. 1192, fig. 5A.
Material. Nineteen teeth studied including P2989.67–P2989.74.
Description. The teeth of this taxon appear to largely represent
juveniles, with only three teeth of larger, presumed adult, indi-
viduals. The largest tooth, probably from the fourth lower file,
is 30 mm in greatest dimension. The teeth show a high degree
of monognathic and dignathic heterodonty (see Shimada 1997;
Siverson 1996; Siverson and Lindgren 2005 for details), as well
as ontogenetic heterodonty; the largest tooth here lacks lateral
cusplets which are present in smaller teeth from similar jaw
positions. Anterior teeth have a cusp that is higher than the
tooth width and is about twice as high as wide. The cusp is
rather labiolingually compressed and straight but inclined to
the posterior. The cusp lingual face is almost flat, whilst the
labial face it smoothly convex. The edges of the cusp flare out-
wards at the base, and in all but a large adult tooth, have a
pair of incipient to very low lateral cusplets. The crown is un-
ornamented other than some faint wrinkles near the base of
the crown on the lingual face of some teeth. The base of the
crown is straight to faintly arched, and does not overhang the
root. The root comprises well-developed root lobes forming an
internal angle of about 90–100 degrees. The anterior root lobe
EXPLANATION OF PLATE 3
Figs 1–6. Ptychodus ex. gr. decurrens Agassiz, 1838; all ·4.5. 1–2, P2989.64, upper symphyseal tooth. 1, occlusal view. 2, lateral view.
3–4, P2989.65, anterior tooth. 3, occlusal view. 4, lateral view. 5–6, P2989.66, lateral tooth. 5, occlusal view. 6, lateral view.
Figs 7–22. Cretoxyrhina denticulata (Glickman, 1957); Figs 9–14 · 2.25; Figs 15–22 · 4.5. 7–8, P2989.67, anterior tooth of subadult. 7,
lingual view. 8, labial view. 9–10, P2989.68, ? fourth lower tooth of adult. 9, labial view. 10, lingual view. 11–12, P2989.69,
posterolateral tooth of adult. 11, labial view. 12, lingual view. 13–14, P2989.70, lower lateral tooth of subadult. 13, labial view. 14,
lingual view. 15–16, P2989.71, parasymphyseal tooth of adult. 15, lingual view. 16, labial view. 17–18, P2989.72, ? upper lateral
tooth of juvenile. 17, labial view. 18, lingual view. 19–20, P2989.73, posterolateral tooth. 19, labial view. 20, lingual view. 21–22,
P2989.74, ? third upper tooth of neonate. 21, labial view. 22, lingual view.
912 P A L A E O N T O L O G Y , V O L U M E 5 3
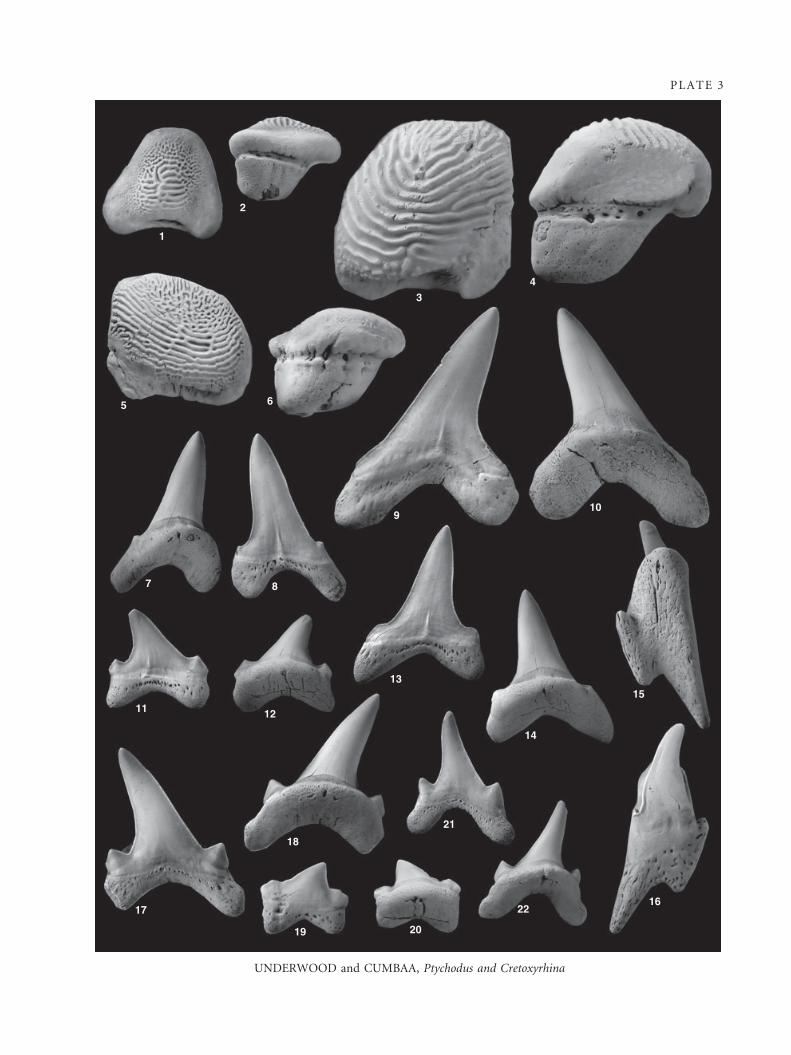
PLATE 3
UNDERWOOD and CUMBAA, Ptychodus and Cretoxyrhina
2
1
34
109
65
7 8
13
14
15
1622
2019
17
11 12
18
21

is longer than the posterior in all teeth seen. The root lobes
are relatively parallel sided and have a smoothly rounded to
slightly pointed termination. The root is labiolingually com-
pressed with a very wide and flat linguo-basal face. There is a
narrow shelf at the base of the crown on the lingual side but
no well-developed protuberance. There is a small foramen at
the top of the root linguo-basal face, with other foramina being
very small and largely confined to a row below the labial
crown-root junction. Lateral teeth have a similar overall design
to those of more anterior positions, but are wider and lower.
The cusp is strongly inclined to the posterior and may be
slightly curved in some teeth. This is flanked by a pair of small
but conspicuous lateral cusplets, which are roughly triangular
in shape. The root lobes are widely separated and the basal
angle is very obtuse.
Discussion. The larger specimens recorded here are simi-
lar to those figured by Welton and Farish (1993), which
are all large teeth presumably of adults. Teeth of
C. mantelli are recorded from younger rocks (e.g. Shi-
mada 1997; Siverson 1996; Siverson and Lindgren 2005)
and differ in having a greater maximum size, and have
wider cusps and less well-developed cusplets throughout
ontogeny, which are largely absent within adults. This
material suggests that teeth from almost all jaw positions
of juveniles of C. denticulata had lateral cusplets, which
are retained in lateral and posterior teeth of adults.
Some specimens from the Late Cenomanian only possess
lateral cusplets on posterolateral teeth (e.g. Welton and
Farish 1993; Siverson 1996) and are only tentatively
referred to this species. Although often uncommon,
C. denticulata has been recorded in many Cenomanian
sites where open marine facies have yielded extensive
chondrichthyan faunas. It is, therefore, likely to have
been a globally distributed generalist tolerant of a wide
range of local conditions.
Family OTODONTIDAE Glickman, 1964
Genus CRETALAMNA Glickman, 1958
Type species. Otodus appendiculatus Agassiz, 1843 from the
Turonian of England.
Remarks. The genus Cretalamna was suggested by Siver-
son (1999) to belong within the Otodontidae Glickman,
1964, and the dental formula (Shimada 2007) would seem
to support this interpretation. There has been discussion
about the spelling of Cretalamna (Cappetta 2006), but it
is here considered that the predominance of usage in
recent years and relative timing of change in ICZN rules
and reassessment of the spelling suggest that the spelling
used here is correct.
Cretalamna sp.
Plate 4, figures 1–2
?1993 Cretolamna woodwardi Herman; Welton and Farish,
p. 105, figs 1–3.
2006 Cretolamna appendiculata (Agassiz); Cumbaa et al.,
p. 146, fig. 4.12.
2006 Cretalamna appendiculata (Agassiz); Shimada et al.,
p. 15, fig. 10.5–10.8.
Material. One tooth (P2989.75).
Description. The single tooth is an upper lateral tooth with a
slightly imperfectly developed root. The tooth is slightly wider
than high with a slightly distally inclined main cusp being over
half the total tooth width. The main cusp is higher than wide
and has a gently convex anterior edge and concave posterior
edge. This is flanked by a pair of triangular lateral cusplets, each
about twice as wide as high. All of the cusps are labiolingually
compressed, with a very slightly convex lingual face and moder-
ately convex labial face. There is a strong and continuous cutting
edge across all cusps. There is no ornament on any of the
crown, the base of which is only slightly arched, so that the
cusps are all in a row. Enameloid does not extend down over
the root on either side. The root is strongly labiolingually com-
pressed, and root lobes are not clearly differentiated, with just a
gentle indentation in the central part of the basal edge of the
root. Both labial and linguo-basal faces of the root are flat.
Discussion. The single tooth recovered in this study, in
addition to rare teeth recorded from the Carrot River
(Cumbaa et al. 2006) appear to belong to a single species
that is conspecific with material figured by Welton and
Farish (1993) as Cretolamna woodwardi. These teeth differ
EXPLANATION OF PLATE 4
Figs 1–2. Cretalamna sp.; ·2.25; P2989.75, upper lateral tooth, 1, labial view. 2, lingual view.
Figs 3–23. Eostriatolamia paucicorrugata sp. nov; all ·6.5. 3–5, P2989.76, holotype, upper lateral tooth. 3, anterior view. 4, labial view.
5, lingual view. 6–8, P2989.77, ? first lower lateral tooth. 6, labial view. 7, lingual view. 8, anterior view. 9, P2989.78, upper lateral
tooth, labial view. 10, P2989.79, lower lateral tooth, labial view. 11, P2989.80, ? third upper anterior tooth, labial view. 12–13,
P2989.81, upper posterolateral tooth. 12, labial view. 13, lingual view. 14–16, P2989.82, ? first upper anterior tooth. 14, labial
view. 15, lingual view. 16, posterior view. 17, P2989.83, ? third lower anterior tooth, labial view. 18–19, P2989.84, ? lower
parasymphyseal tooth. 18, labial view. 19, lingual view. 20, P2989.85, ? second lower anterior tooth, labial view. 21–23, P2989.86,
? second upper anterior tooth. 21, labial view. 22, anterior view. 23, lingual view.
914 P A L A E O N T O L O G Y , V O L U M E 5 3
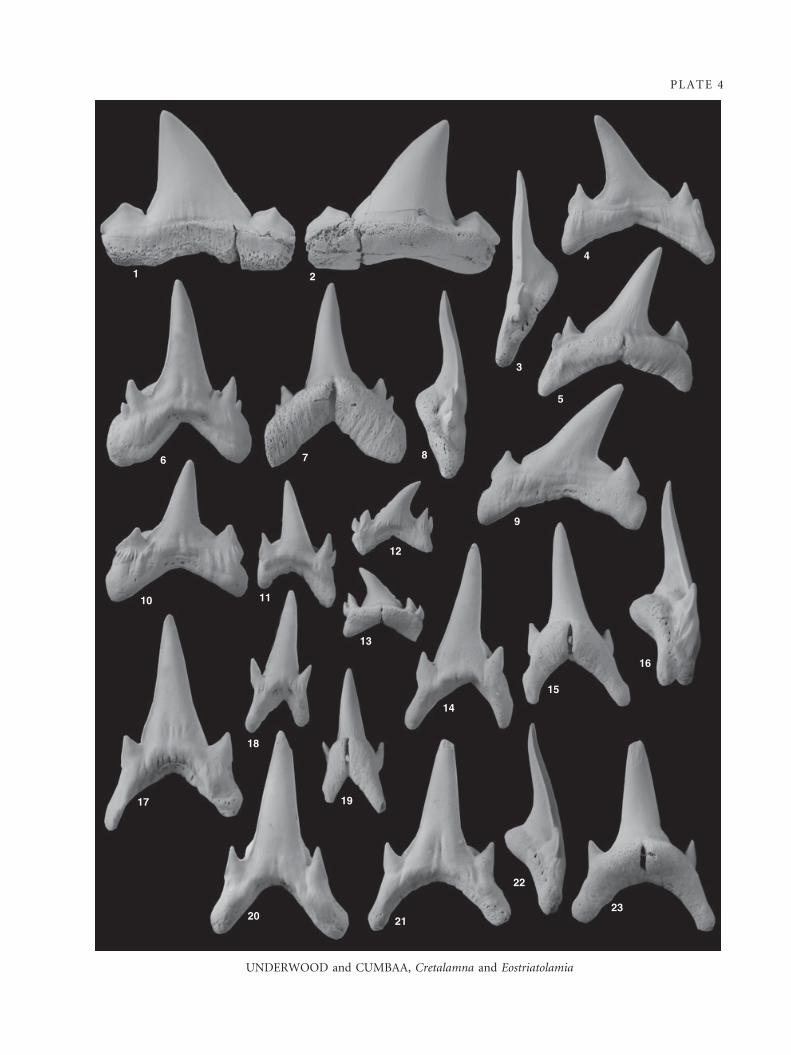
PLATE 4
UNDERWOOD and CUMBAA, Cretalamna and Eostriatolamia
1 2
4
3
5
6 7 8
9
12
11
13
14
15
16
10
17 19
18
20 2123
22

markedly from the type material of Dwardius woodwardi,
with smaller lateral cusplets in anterior teeth and greater
labiolingual compression and a straighter root in lateral
teeth. The overall tooth design is far closer to that of Cre-
talamna appendiculata but differs in possessing a more
robust root with a far more rounded profile. The tooth
described here is also considerably larger then in coeval
C. appendiculata. It is here considered that the species
C. appendiculata, as typically used, is in reality a number
of distinct and readily definable species, of which this is
but one.
Family ODONTASPIDIDAE Muller and Henle, 1839 s.l.
Remarks. Although the extant species Carcharias taurus
Rafinesque, 1810, Odontaspis ferox (Risso, 1810) and
Odontaspis noronhai (Maul, 1955) are all included into the
Odontaspididae by Compagno (2001), there is evidence
that the ‘family’ is polyphyletic and should be split into
two or even three separate families. The taxa within the
Odontaspididae have very different dentitions, and these
different dental formulae are readily recognized in the
fossil record. In addition, molecular phylogenetic work
(Human et al. 2006) suggests that O. ferox belongs in a
clade with Pseudocarcharias Cadenat, 1963 and Alopias
Rafinesque, 1810, whereas C. taurus is in a clade with the
Lamnidae Muller and Henle, 1838. Although defining new
families containing extant taxa is beyond the scope of this
paper and would involve considerable anatomical study of
modern specimens, it is considered here that the Odonta-
spididae should not be considered as a natural grouping.
Family ODONTASPIDIDAE Muller and Henle, 1839 s.l.,
?Odontaspis ferox group
Genus EOSTRIATOLAMIA Glickman, 1980
Type species. Lamna venusta Leriche, 1929 from the Campanian
of France.
Remarks. The genus Eostriatolamia was erected for Lamna
venusta and a number of other Carcharias-like taxa from
the Cretaceous, including Odontaspis striatula Dalinkevi-
cius, 1935. The genus was considered to be a junior syno-
nym of Carcharias by Cappetta (1987), who stated that
the teeth of L. venusta and O. striatula were similar. It is
here considered that L. venusta and O. striatula are con-
generic, but do not belong to Carcharias.
Teeth of E. striatula very closely resemble those of Ce-
nocarcharias tenuiplicatus Cappetta and Case, 1975, the
similarities being recognized by Welton and Farish
(1993). Cappetta and Case (1999) suggested that C. tenui-
plicatus differs from E. striatula in being smaller, thicker,
having a less inclined cusp and having stronger lingual
ornament. Despite this, figured specimens of both species
show great heterodonty and variation in ornament, with
no specimens representing the same probable tooth posi-
tion being figured from both species. It is here considered
that Cenocarcharias should be considered a junior syno-
nym of Eostriatolamia, with E. tenuiplicatus and E. striatu-
la being very closely related or possibly conspecific.
Eostriatolamia paucicorrugata sp. nov.
Plate 4, figures 3–23
?2001b Cenocarcharias tenuiplicatus (Cappetta and Case);
Cicimurri, p. 185, fig. 5r.
2006 Cenocarcharias tenuiplicatus (Cappetta and Case);
Cumbaa et al., p. 148, fig. 5.2.
Derivation of name. From weak corrugations on the labial face
of the crown.
Holotype. P2989.76 (Pl. 4, figs 3–5).
Additional material. One hundred and fifty teeth studied includ-
ing P2989.77–P2989.86.
Diagnosis. Heterodonty well developed and probably of
Odontaspis ferox type. Teeth small and gracile, typically less
than 10 mm high. Root and base of crown quite symmet-
rical in all teeth, with main cusp distally inclined or curved
in presumed upper lateral files. Main cusp higher than
wide in all teeth, widening towards the base, especially in
lateral teeth. Lingual curvature of cusp present in anterior
teeth. One or two pairs of short but robust lateral cusplets
present in all teeth, clearly separated from main cusp but
joined by broad labial base of crown. Cusplets sharply
pointed and higher than wide in anterior teeth but similar
width and height in lateral teeth. Lateral edge of outer cus-
plets overhangs outer edges of root, weakly in anterior
teeth, strongly in lateral teeth. Base of crown extends at
least half way down labial face of root lobes and does not
strongly overhang the root in lateral teeth. A continuous
and well-developed cutting edge extends over all cusps.
Crown ornament restricted to faint and irregular corruga-
tions at base of labial face and, in some teeth, very faint
longitudinal striations on lingual face of cusps. The root is
strongly bilobed with a very prominent nutritive groove in
all teeth. Root lobes form basal angle of 70–140 degrees.
Linguobasal face of root slightly arched and flattened.
Description. Teeth of this species are relatively small, with the
largest anterior teeth being 11 mm high. There is considerable
heterodonty that appears to follow the same pattern as in Odon-
taspis ferox.
916 P A L A E O N T O L O G Y , V O L U M E 5 3

All anterior teeth and most lower lateral teeth are nearly sym-
metrical, whilst the main cusp of upper lateral and some lower
posterolateral teeth are inclined to the posterior, even though
the base is still centrally situated on the tooth. Anterior teeth
have a straight and slender main cusp flanked by a pair of small
but clearly differentiated lateral cusplets; a second pair of very
small lateral cusplets is present on some teeth. The main cusp is
triangular in profile, with smoothly tapering sides, and is consid-
erably more than twice as high as wide. The main cusps of lower
anterior teeth are very slightly curved lingually but this curvature
is not seen in upper teeth. Lateral cusplets of anterior teeth are
longer than wide and are slightly divergent. The cusps are united
by a well-developed lower part of the crown, which extends for
at least half of the length of the root lobes on the labial side.
This basal part of the crown is rather swollen and overhangs the
root labially. At the crown-root junction on the lingual face, a
well-developed, recessed, neck is present around the base of the
main cusp. There is typically an indentation on the labial face of
the crown between the basal edge and the base of the lateral cus-
plets. The labial face of all cusps is gently convex, although this
may be flat or slightly concave at the base of the main cusp.
This face is separated from the strongly convex lingual face by a
small but continuous cutting edge. Ornament is weak on both
sides of anterior teeth. Ornamentation of the labial face is pres-
ent in most, but not all, teeth as irregular and short wrinkles at
the base of the cusps, usually concentrated in the area between
the main and lateral cusps. On the lingual face ornament is also
weak or absent, comprising numerous extremely fine longitudi-
nal ridges on the lower third of the main cusp. Roots of all ante-
rior teeth are strongly ‘V’ shaped, with the angles between the
root lobes varying from 70 to 110 degrees, largely depending on
jaw position. The root lobes gently taper towards somewhat
pointed terminations, and the main difference between anterior
and posterior root lobes is the degree of angularity of the lobe
termination. The labial face of the root is narrow and at least
partly overhung by the crown. The lingual side of the root has a
well-developed lingual protuberance in all teeth but this is espe-
cially well developed in lower anterior teeth where it is very pro-
nounced. A deep and very well-developed nutritive groove is
always present. Lingual faces of the root lobes are somewhat
concave and there is a marked, but not sharp, angle between the
labial and lingual faces. Lower parasymphyseal teeth are not
much smaller than other anterior teeth and similar in overall
form to other lower anterior teeth but are considerably more
compressed anterior distally. A single very small, fragmentary,
tooth is probably from an upper parasymphyseal position.
Lateral teeth are wider and lower than anterior teeth, in many
jaw positions being approximately as wide as high. In all teeth,
the base of the main cusp comprises about a third of the tooth
width, being flanked by a single, or more commonly double,
pair of lateral cusplets. In profile, the main cusp of all teeth is
somewhat concave towards the base, and the base merges with
the inner pair of cusplets. The main cusp is erect in lower teeth
but posteriorly inclined in upper teeth and does not show lin-
gual curvature in any tooth position. Lateral cusplets are higher
than wide when two pairs are present but similar width and
height on teeth possessing only one pair. The outer edge of the
cusplets overhangs the root lobes, giving a somewhat ‘spearhead’
profile. All cusps are united to a well-developed labial extension
of the crown, which extends at least half way down the labial
face of the root and forms a distinct overhang at its base in the
centre, but not on the root lobes. The lateral parts of the crown
on the labial side of the tooth are swollen, and there is typically
a concavity at the base of the lateral cusplets, especially in upper
teeth. The labial face of all cusps is weakly convex, and there is a
strong and continuous cutting edge separating crown lingual
and labial faces. Strong and irregular vertical corrugations are
usually present on the labial face of the crown but do not extend
onto the cusps. On some teeth from both lower and upper posi-
tions, these are restricted to a single ridge below the lateral cus-
plets or absent altogether. The lingual face of the crown is
convex and sometimes ornamented with very fine longitudinal
striations on the lower part of the main cusp. A well-developed
but narrow neck is present along the lingual crown-root junc-
tion. Both anterior and posterior root lobes are similar and have
an angle between the root lobes of between 115 and 140 degrees.
The root of upper teeth is more compressed labiolingually than
of lower teeth, and root lobes are somewhat wider but otherwise
they are similar. Root lobes are slightly longer than the lateral
extent of the crown, with the labial face of the root being partly
covered by the crown. The remainder of the root labial face is
shallow and somewhat flared towards the base. On the lingual
side of the tooth, there is a narrow but prominent shelf below
the base of the cusps. This shelf is separated from the linguo-
basal face of the root by a clear and relatively sharp edge. The
linguo-basal face of the root is quite flat and somewhat concave.
Below the base of the main cusp, the root extends lingually,
weakly in upper teeth but more strongly in lower teeth, and a
very prominent nutritive groove is present. A tooth of presumed
third upper anterior ⁄ first intermediate position has a morphol-
ogy intermediate between that of anterior and lateral teeth but is
more inclined to the posterior than either and, if it is from an
adult, somewhat smaller.
Remarks. Teeth of Eostriatolamia paucicorrugata sp. nov.
are similar to specimens referred to Eostriatolamia striatu-
la and E. tenuiplicatus, and probably represent a closely
related species. Although some specimens of E. striatula
lack ornament on the labial or both crown faces (speci-
mens of which Dalinkevicius (1935) only tentatively
referred to the species), an ornament of short and strong
ridges is usually present at the base of the labial face, and
fine striations are typically present on the lingual face of
the main cusp of teeth referred to E. striatula and E. ten-
uiplicatus. In contrast, ornament is very restricted on
teeth of E. paucicorrugata sp. nov. In addition, lateral cus-
plets are more robust in E. paucicorrugata sp. nov. than
in other, related, species, and there is a greater tendency
for a second pair of lateral cusplets to be present, whilst
the base of the labial crown face extends further along the
root lobes and does not overhang the root to the same
degree. Teeth of E. paucicorrugata sp. nov. also resemble
those of E. subulata (Agassiz, 1843), but teeth of E. subu-
lata differ in possessing larger lateral cusplets and a
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 917

straight labial crown base, which does not extend onto
the root lobes. E. paucicorrugata sp. nov. teeth differ from
those of ‘Odontaspis’ saskatchewanensis Case et al. 1990
(which is probably conspecific with Carcharias sp. A. of
Welton and Farish 1993 but that very small number of
specimens recorded for ‘O.’ saskatchewanensis prevents
detailed comparison) by its larger size, wider lateral cus-
plets and flared labial base of the crown.
Although general tooth morphology of Eostriatolamia
paucicorrugata sp. nov. is very similar to that of Carcha-
rias taurus, the lack of the numerous files of reduced pos-
terior teeth present in C. taurus confirm generic
separation (Glickman and Averianov 1998). Despite the
lack of sufficient material to reconstruct the dentition of
E. paucicorrugata sp. nov., it is tentatively suggested that
the dental formula would resemble that of Odontaspis fe-
rox. This is suggested by a moderate-sized tooth resem-
bling that of the third upper anterior ⁄ first intermediate
position of O. ferox, whilst two different sizes of parasym-
physeal teeth have been recognized in E. striatula by Da-
linkevicius (1935) and are probably also present in this
species.
Eostriatolamia paucicorrugata sp. nov. appears to be a
species endemic to the northern part of the Western
Interior Seaway, being known only from Canada and
possibly North Dakota and Wyoming (Cicimurri 2001b).
Further south it is absent but replaced by E. tenuiplica-
tus, which is present in Texas (Cappetta and Case 1975,
1999; Welton and Farish 1993) and Colorado (Shimada
et al. 2006). The limited ranges of these species contrasts
with the widespread distribution of E. striatula (e.g.
Dalinkevicius 1935; Landemaine 1991; Biddle 1993;
Siverson 1997).
Family ODONTASPIDIDAE Muller and Henle, 1839 s.l.,
Carcharias taurus group
Genus ROULLETIA Vullo et al., 2007
Type species. Roulletia bureaui Vullo et al., 2007 from the Ceno-
manian of France.
Roulletia canadensis sp. nov.
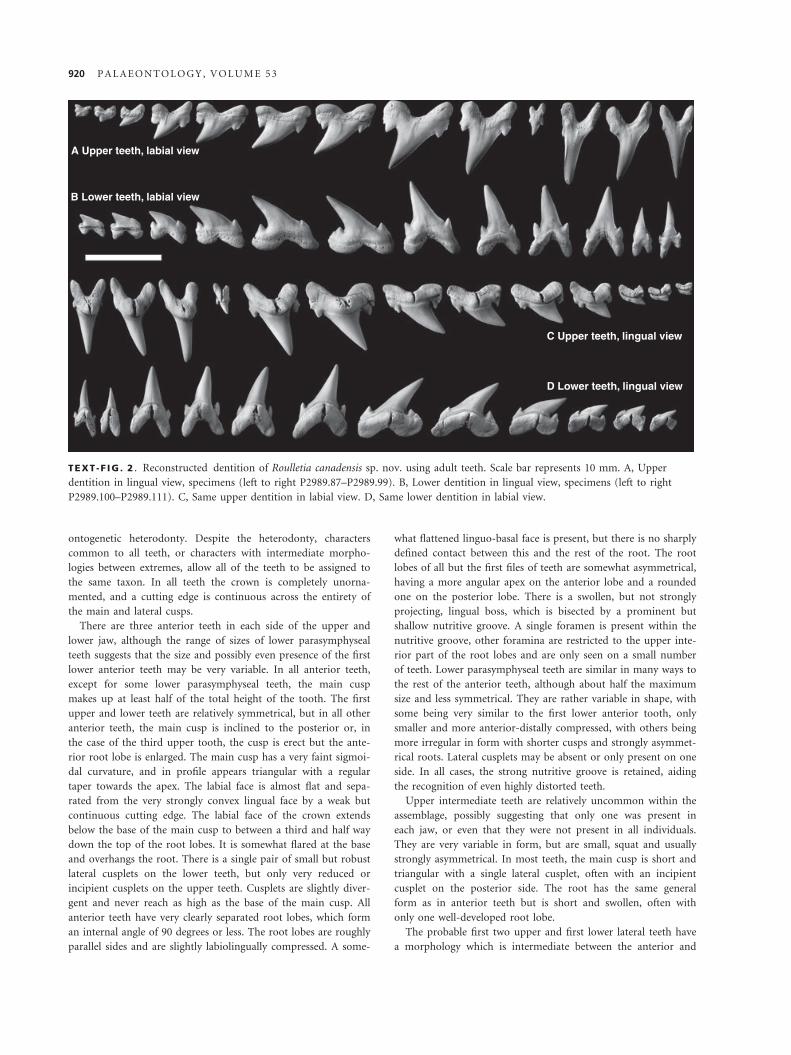
Plate 5, figures 1–24; Text-figure 2A–D
1999 Carcharias sp. Cumbaa and Tokaryk, p. 61, fig. 5
(pars; bottom tooth centre right column).
2006 Carcharias amonensis (Cappetta and Case); Cumbaa
et al., p. 148, fig. 5.1.
2006 Dallasiella willistoni Cappetta and Case; Cumbaa
et al., p. 148, fig. 5.3.
2006 Carcharias amonensis (Cappetta and Case); Shimada
et al., p. 12, fig. 9.1–9.2.
Derivation of name. From abundant specimens being apparently
restricted to Canada.
Holotype. P2989.107 (Pl. 5, figs 15–16), third lower lateral tooth.
Additional material. About 4500 teeth studied including
P2989.87–P2989.106; P2989.108–P2989.111.
Diagnosis. Dentition of Carcharias type with three differ-
entiated upper and three lower anterior teeth, upper inter-
mediate teeth and lower parasymphyseal teeth. All but
first upper and lower teeth somewhat asymmetrical. Orna-
ment lacking on all teeth. Crown rather flared on labial
face, overhanging top of root. Cutting edge continuous
across main and lateral cusps in all teeth. Anterior teeth
with steadily tapering, triangular main cusp and very small
to incipient lateral cusplets not reaching base of main
cusp. Root of all teeth with clearly defined lobes and well-
developed nutritive groove. Root of anterior and first lat-
eral teeth strongly ‘V’ shaped. Root lobes roughly parallel
sided with rounded terminations and only slightly flat-
tened basal face. Lateral teeth all have main cusp inclined
to posterior, in teeth posterior of second lateral position
main cusp strongly curved. One pair of lateral cusplets
sometimes with incipient outer pair present but short and
triangular. Root lobes in lateral teeth with wide and flat
basal face giving ‘swollen’ profile to posterior root lobe.
Description. The dentition of Roulletia canadensis sp. nov. shows
very pronounced monognathic and less pronounced dignathic
heterodonty. There is no evidence of sexual or significant
EXPLANATION OF PLATE 5
Figs 1–24. Roulletia canadensis sp. nov.; all ·6. 1–3, P2989.102, first lower anterior tooth. 1, labial view. 2, lingual view. 3, anterior
view. 4–6, P2989.89, third upper anterior tooth. 4, labial view. 5, lingual view. 6, anterior view. 7–9, P2989.88, second upper
anterior tooth. 7, labial view. 8, lingual view. 9, anterior view. 10, P2989.87, first upper anterior tooth, labial view. 11, P2989.103,
second lower anterior tooth, labial view. 12, P2989.95, fifth upper lateral tooth, labial view. 13–14, P2989.90, upper intermediate
tooth. 13, labial view. 14, lingual view. 15–17, P2989.104, holotype, third lower lateral tooth. 15, lingual view. 16, labial view. 17,
anterior view. 18, P2989.101, lower parasymphyseal tooth, labial view. 19–20, P2989.92, second upper lateral tooth. 19, labial
view. 20, lingual view. 21–22, P2989.110, lower posterior tooth. 21, labial view. 22, lingual view. 23–24, P2989.97, upper posterior
tooth. 23, labial view. 24, lingual view.
918 P A L A E O N T O L O G Y , V O L U M E 5 3
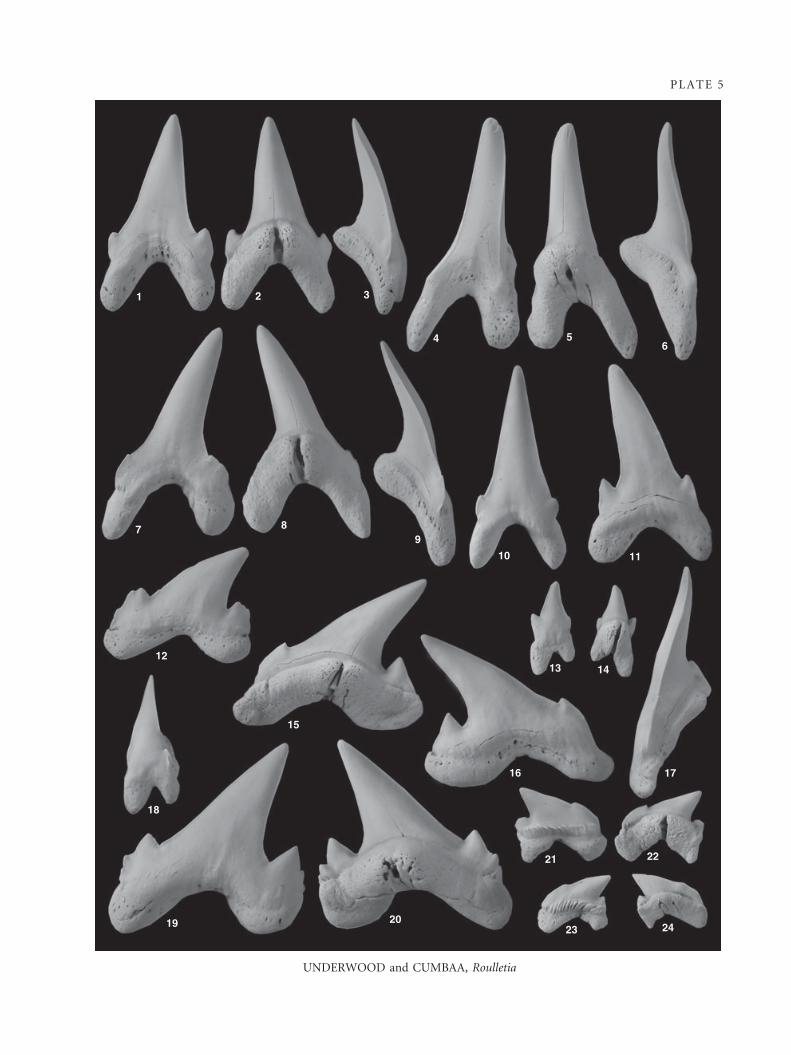
PLATE 5
UNDERWOOD and CUMBAA, Roulletia
21 3
4 56
7 89
10 11
13 1412
15
16 17
21 22
23 242019
18
ontogenetic heterodonty. Despite the heterodonty, characters
common to all teeth, or characters with intermediate morpho-
logies between extremes, allow all of the teeth to be assigned to
the same taxon. In all teeth the crown is completely unorna-
mented, and a cutting edge is continuous across the entirety of
the main and lateral cusps.
There are three anterior teeth in each side of the upper and
lower jaw, although the range of sizes of lower parasymphyseal
teeth suggests that the size and possibly even presence of the first
lower anterior teeth may be very variable. In all anterior teeth,
except for some lower parasymphyseal teeth, the main cusp
makes up at least half of the total height of the tooth. The first
upper and lower teeth are relatively symmetrical, but in all other
anterior teeth, the main cusp is inclined to the posterior or, in
the case of the third upper tooth, the cusp is erect but the ante-
rior root lobe is enlarged. The main cusp has a very faint sigmoi-
dal curvature, and in profile appears triangular with a regular
taper towards the apex. The labial face is almost flat and sepa-
rated from the very strongly convex lingual face by a weak but
continuous cutting edge. The labial face of the crown extends
below the base of the main cusp to between a third and half way
down the top of the root lobes. It is somewhat flared at the base
and overhangs the root. There is a single pair of small but robust
lateral cusplets on the lower teeth, but only very reduced or
incipient cusplets on the upper teeth. Cusplets are slightly diver-
gent and never reach as high as the base of the main cusp. All
anterior teeth have very clearly separated root lobes, which form
an internal angle of 90 degrees or less. The root lobes are roughly
parallel sides and are slightly labiolingually compressed. A some-
what flattened linguo-basal face is present, but there is no sharply
defined contact between this and the rest of the root. The root
lobes of all but the first files of teeth are somewhat asymmetrical,
having a more angular apex on the anterior lobe and a rounded
one on the posterior lobe. There is a swollen, but not strongly
projecting, lingual boss, which is bisected by a prominent but
shallow nutritive groove. A single foramen is present within the
nutritive groove, other foramina are restricted to the upper inte-
rior part of the root lobes and are only seen on a small number
of teeth. Lower parasymphyseal teeth are similar in many ways to
the rest of the anterior teeth, although about half the maximum
size and less symmetrical. They are rather variable in shape, with
some being very similar to the first lower anterior tooth, only
smaller and more anterior-distally compressed, with others being
more irregular in form with shorter cusps and strongly asymmet-
rical roots. Lateral cusplets may be absent or only present on one
side. In all cases, the strong nutritive groove is retained, aiding
the recognition of even highly distorted teeth.
Upper intermediate teeth are relatively uncommon within the
assemblage, possibly suggesting that only one was present in
each jaw, or even that they were not present in all individuals.
They are very variable in form, but are small, squat and usually
strongly asymmetrical. In most teeth, the main cusp is short and
triangular with a single lateral cusplet, often with an incipient
cusplet on the posterior side. The root has the same general
form as in anterior teeth but is short and swollen, often with
only one well-developed root lobe.
The probable first two upper and first lower lateral teeth have
a morphology which is intermediate between the anterior and
A Upper teeth, labial view
B Lower teeth, labial view
C Upper teeth, lingual view
D Lower teeth, lingual view
TEXT -F IG . 2 . Reconstructed dentition of Roulletia canadensis sp. nov. using adult teeth. Scale bar represents 10 mm. A, Upper
dentition in lingual view, specimens (left to right P2989.87–P2989.99). B, Lower dentition in lingual view, specimens (left to right
P2989.100–P2989.111). C, Same upper dentition in labial view. D, Same lower dentition in labial view.
920 P A L A E O N T O L O G Y , V O L U M E 5 3

remainder of the lateral teeth. The remainder of the lateral and
posterolateral teeth have a morphology that is quite distinct
from that of the anterior teeth. Lateral teeth are all strongly
asymmetrical with the main cusp strongly inclined or, on the
more posterior teeth, curved towards the posterior. Presumed
upper teeth are somewhat more compressed than lowers, with a
rather more ‘blade-like’ main cusp. In all lateral teeth the apex
of the main cusp reaches a point level with the posterior end of
the root. The main cusp is of a similar height to the root in all
but the posteriormost teeth, where it is somewhat shorter. The
base of the main cusp comprises somewhat over half of the total
width of the tooth, and in the majority of teeth is flanked by a
pair of short and triangular lateral cusplets. In some teeth from
both presumed upper and lower positions, incipient or very
small additional cusplets may be present, generally but not
always on the posterior side of the tooth. In posterolateral teeth,
the anterior lateral cusplet is commonly fused to the leading
edge of the main cusp to form a serration, or is absent alto-
gether. The labial face of all cusps is only slightly convex, and
there is typically a weak concavity at the base of the main cusp.
The lingual face of the cusps is more strongly convex than the
labial face, but all cusps are still distinctly labiolingually com-
pressed. The cutting edge separating labial and lingual crown
faces is well developed and continuous. The base of the crown
extends a short distance below the cusps on the labial face, with
very minor overhang of the root in the central part. The base of
the crown on the lingual face is gently arched over the root and
has a narrow but well-defined neck. The root of all lateral and
posterolateral teeth is strongly labiolingually compressed with a
wide and flat linguo-basal face. The root lobes are relatively
short, and the sharp angle between the linguo-basal edges of the
root lobes is from 90 to 110 degrees. The root lobes are of simi-
lar lengths, with a very well-rounded apex to the posterior lobe
and a somewhat more angular apex to the anterior lobe. A pro-
nounced shelf is present at the top of the root on the lingual
side of the tooth, and there is a sharp angle between this and
the root linguo-basal face. A well-developed nutritive groove
extends vertically across the entire root linguo-basal face.
Discussion. Teeth of Roulletia canadensis sp. nov. are
extremely common within the samples studied, and com-
prise the second most abundant chondrichthyan remains.
The reconstructed dentition gives a dental formula very
similar to that of Carcharias taurus, although differs from
it in lacking the small posterior teeth of C. taurus, and
probably in having less strongly compressed lower para-
symphyseal teeth. Teeth of R. canadensis sp. nov. are dis-
tinctly different from those of C. taurus, with greater
degree of heterodonty and more compressed and inclined
lateral teeth, further confirming the generic distinction.
Roulletia canadensis sp. nov. has teeth similar to those
of R. bureaui Vullo et al., 2007 from the Cenomanian of
France but differs in a number of respects. Teeth of
R. canadensis sp. nov. are more gracile and labiolingually
compressed. They have a less robust main cusp which is
more blade-like in lateral teeth, have wider lateral cusplets
and have a root which, in lateral teeth, is less strongly ‘V’
shaped and have wider and more rounded root lobes.
Despite these differences, the similarity in overall tooth
shape and heterodonty suggest that these two species are
congeneric. A second species that is very similar to
R. canadensis sp. nov. is ‘Carcharias’ amonensis (Cappetta
and Case, 1975) which is present elsewhere in North
America in the Cenomanian (e.g. Welton and Farish 1993;
Cappetta and Case 1999). Latero-posterior teeth of
‘C.’ amonensis and R. canadensis sp. nov. are very similar,
although teeth of R. canadensis sp. nov. have a more
curved main cusp and more rounded root lobes. Teeth
from other jaw positions are less similar, with R. canaden-
sis sp. nov. having a less erect main cusp and shorter lat-
eral cusplets in lateral teeth, and very much smaller lateral
cusplets and a complete cutting edge in anterior teeth.
Roulletia canadensis sp. nov. appears to have been
abundant within the northern part of the Western Inte-
rior Seaway, being common at a number of sites in cen-
tral Canada in addition to the one described here (SC,
pers. obs.). It is present, if previously unrecognized, in
the mid part of the Seaway (Shimada et al. 2006) where it
co-occurs with ‘Carcharias’ amonensis (Shimada et al.
2006, fig 9.3). R. canadensis sp. nov. has not been
recorded in Texas (Cappetta and Case 1975, 1999; Welton
and Farish 1993) where ‘C.’ amonensis is common. In
France, ‘Carcharias’ amonensis and R. bureaui are both
known but do not occur at the same stratigraphical levels.
Current records, therefore, suggest that R. canadensis sp.
nov. was a cool-water taxon, possibly endemic to the
northern WIS.
Family ARCHAEOLAMNIDAE fam. nov.
Derivation of name. From Archaeolamna Siverson, 1992, the
genus used to diagnose the family.
Diagnosis. Family known only from disassociated teeth.
Dentitions with well-developed ‘lamniform’ heterodonty
with distinct tooth morphologies in different jaw posi-
tions. Dentitions comprise three upper anterior and typi-
cally three lower anterior teeth. Small and variously
‘stunted’ teeth are present in upper parasymphyseal, lower
parasymphyseal and upper intermediate positions. No
distinct morphological break between lateral and poster-
ior teeth in upper and lower jaws, and largest teeth of
dentition in lateral positions. Teeth of typical lamniform
morphology with ‘V’ or ‘U’ shaped root and erect cusp
flanked by smaller cusplets. Crown ornament weak, irreg-
ular or absent. Lingual neck of crown narrow but well
defined. Teeth with serrations or cutting morphology are
unknown. Root large and dense, with the only well-devel-
oped foramen on apex of lingual bulge or within nutritive
groove. Nutritive groove absent, small or shallow. Teeth
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 921

with anaulacorhize vascularization restricted to large or
posterior teeth of taxa where a large foramen is typically
present.
Included taxa. Archaeolamna is the genus from which the family
is diagnosed. In addition, there are several other genera with
similar overall tooth morphology but within which the dental
formula is unknown. Archaeolamna is known from Albian to
Maastrichtian strata (Siverson 1996). Teeth of Cretodus Sokolov,
1965 are very similar to those of Archaeolamna, differing largely
in their much stronger ornament, probably lesser heterodonty
and greater maximum size. Cretodus is known from Cenomanian
through Campanian strata (Schwimmer et al. 2002). Teeth of
Dallasiella Cappetta and Case, 1999 are also very similar to those
of Archaeolamna, differing mainly in the greater compression of
the roots of lateral teeth, and this may also be a member of the
Archaeolamnidae fam. nov. Dallasiella is reported from Ceno-
manian to Coniacian strata (Cappetta and Case 1999; Cumbaa
et al. 2006; Cook et al. 2008). Inferred presence of the same
dental formula suggests that Telodontaspis gen. nov. (see below)
may also be included into the Archaeolamnidae fam. nov. Teeth
of Protolamna Cappetta, 1980a, included into the Eoptolamnidae
by Kriwet et al. (2008), also have a number of similarities to the
anterior teeth of Archaeolamna. Despite this, morphology of
teeth of other positions, and the presumed dental formula, of
known species of the Eoptolamnidae readily differentiate the two
families. The known range of the Archaeolamnidae fam. nov. is,
therefore, Albian to Maastrichtian and was apparently globally
distributed.
Remarks. The dental formula of Archaeolamna differs
from that of all other known Cretaceous and Cenozoic
lamniforms in possessing teeth in lower parasymphyseal,
upper parasymphyseal and intermediate tooth positions
(of Cappetta 1987), associated with three large upper
anterior teeth. Within Cretaceous lamniforms, upper
parasymphyseal teeth are present within Cretoxyrhina
(Siverson 1996; Shimada 1997), Cretalamna (Shimada
2007) and Cardabiodon (Siverson 1999; where the indi-
cated first upper anterior tooth may also represent a
parasymphyseal), indicating their presence within the
Cretoxyrhinidae Glickman, 1958, Otodontidae Glickman,
1964 and Cardabiodontidae Siverson, 1999, respectively,
whilst lower parasymphyseal teeth are either absent in all
three taxa, or possibly present only in Cretoxyrhina (Shi-
mada 1997). Intermediate teeth (sensu Cappetta 1987,
defined as stunted teeth growing on the bar between the
upper anterior and lateral hollows) are absent in all three
species, with the intermediate teeth of Shimada (1997,
2007) here considered as third upper anterior teeth. All
three of these tooth positions are absent in extant Lamni-
dae Muller and Henle, 1838.
Within the extant Odontaspididae Muller and Henle,
1839 s.l., upper parasymphyseal teeth are present in
Odontaspis ferox (Risso, 1810) but absent in O. noronhai
(Maul, 1955) and Carcharias taurus Rafinesque, 1810.
Lower parasymphyseal and intermediate teeth are present
in all three species, but in O. ferox the intermediate teeth
are especially numerous and preceded by only two files of
large upper anterior teeth, in contrast to the three of
almost all other lamniforms.
The apparently unique presence of all tooth families
within Archaeolamna suggests that the tooth formula is
the least derived, with progressive loss of files of small
teeth in more derived lamniform families, until in the
Lamnidae all of these ‘small’ teeth are lost and the third
upper anterior tooth is severely reduced.
Genus ARCHAEOLAMNA Siverson, 1992
Type species. Odontaspis kopingensis Davis, 1890 from the Camp-
anian of Sweden.
Archaeolamna ex. gr. kopingensis (Davis 1890)
Text-figures 3A–D, 4A–R
1990 Cretodus sp. Case et al., p. 1090, fig. 7a–f.
1996 Archaeolamna aff. kopingensis (Davis, 1890);
Siverson, p. 825, pl. 2, figs 1–6.
1997 Archaeolamna sp. Siverson, p. 462, fig. 4A–H.
1999 Carcharias sp. Cumbaa and Tokaryk, p. 61, fig. 5
(pars; upper 3 teeth centre right column).
2001 Cenocarcharias tenuiplicatus (Cappetta and Case);
Case, pl. 2, figs 4, 6.
?2001 ?Carcharias amonensis (Cappetta and Case); Case,
pl. 2, fig. 7.
2006 Archaeolamna sp. Cumbaa et al., p. 146, fig. 4.10.
2006 Cretodus semiplicatus (Munster in Agassiz); Cumbaa
et al., p. 146, fig. 4.11.
2006 Archaeolamna cf. A. kopingensis (Davis, 1890);
Shimada et al. p. 15, fig. 10.1.
2008 Dallasiella willistoni Cappetta and Case; Cook et al.,
p. 1192, fig. 5B.
2008 Archaeolamna cf. A. kopingensis (Davis, 1890); Cook
et al., p. 1192, fig. 5C.
Material. About 4000 teeth studied including P2989.112–
P2989.140.
Description. The assemblage of teeth of Archaeolamna ex. gr.
kopingensis shows very varied morphologies, revealing strong
monognathic and weak dignathic heterodonty (see above), as
well as some ontogenetic and probable sexual heterodonty.
Within the tooth assemblage, most of the teeth are reasonably
small, with a small number of larger teeth (as used in the denti-
tion reconstruction) and a very small minority of teeth that are
at least 50% larger than average. The very largest teeth differ
from others in that there are commonly strong but short and
irregular longitudinal folds at the base of the labial face of the
crown, especially prominent in teeth from posterolateral
922 P A L A E O N T O L O G Y , V O L U M E 5 3

positions. Larger teeth of all anterior and anterolateral tooth
positions appear to be present in both a robust and a gracile
morph. These have the same overall shape but the main cusp
and the root lobes of the gracile morph are considerably more
slender than in the robust morph. The main cusp of the gracile
teeth is also somewhat longer for the width of the tooth than in
the robust morph. By analogy with some modern lamniforms
and carcharhiniforms, it is probable that the gracile morph
represents teeth of adult male individuals, whilst the robust
morph represents females and immature individuals.
Anterior teeth have a main cusp that is rather higher than the
root and a single pair of slightly divergent lateral cusplets. The
main cusp is robust and tapers steadily to a point. In lower
anterior teeth and presumed first upper anterior teeth, the main
cusp is straight and vertical, becoming inclined in some upper
anterior teeth (file two) and curved in others (file three). The
lateral profile of the main cusp shows a very slight sigmoidal
curvature. In all anterior teeth the lateral cusplets on anterior
and posterior sides of the tooth are of similar size, and are
rather higher than wide. The crown unites the main and lateral
cusplets at the base on the labial side, but not on the lingual
face, where a narrow neck is present at the base of each of the
cusps. The labial face of the cusps is faintly convex, but com-
monly flat or slightly concave at the base of the main cusp. The
lingual face of all cusps is strongly convex, giving them a rather
semicircular cross-section. A well-defined cutting edge is contin-
uous across all cusps. The root is bulky and strongly ‘V’ shaped.
The root is somewhat asymmetrical in all anterior teeth, with
the asymmetry varying with tooth position. The root lobes form
an internal angle of 90 degrees or less, depending on jaw posi-
tion. Root lobes are oval in cross-section, largely compressed an-
tero-distally, but in the posterior lobe this may be twisted
towards the lingual side. The terminations of the root lobes are
variable in shape, ranging from smoothly rounded to sharply
pointed, but are never strongly flattened. The linguo-basal base
of the root is not strongly flattened. There is a rather inflated
root lingual boss, and in the centre of this is either a single well-
developed foramen or a cluster of up to four small foramina. In
a small proportion of smaller teeth, the main foramen is within
a short but deep groove.
Lower parasymphyseal teeth are very variable but are essen-
tially modified anterior teeth. There appears to be a morphologi-
cal continuum between first lower anterior teeth and highly
modified and stunted parasymphyseal teeth. It is, therefore,
likely that the dentition at the front of the lower jaw is less
clearly defined than in other lamniforms. The crown of lower
parasymphyseal teeth differs from that of other anterior teeth
largely in being smaller relative to the root and less symmetrical,
with lateral cusplets often being better formed on one side of
the tooth than the other, or even absent on one or both sides.
The root is antero-distally compressed with a very well-devel-
oped lingual boss. Root lobes are short and rarely of close to
equal length; in many of the smaller teeth, only a single root
lobe is fully developed.
A Upper teeth, labial view
B Lower teeth, labial view
C Upper teeth, lingual view
D Lower teeth, lingual view
TEXT -F IG . 3 . Reconstructed dentition of Archaeolamna ex. gr. kopingensis (Davis 1890) using large teeth of the robust (?female)
morph. Scale bar represents 10 mm. A, Upper dentition in lingual view, specimens (left to right P2989.112–P2989.125). B, Lower
dentition in lingual view, specimens (left to right P2989.126–P2989.138). C, Same upper dentition in labial view. D, Same lower
dentition in labial view.
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 923
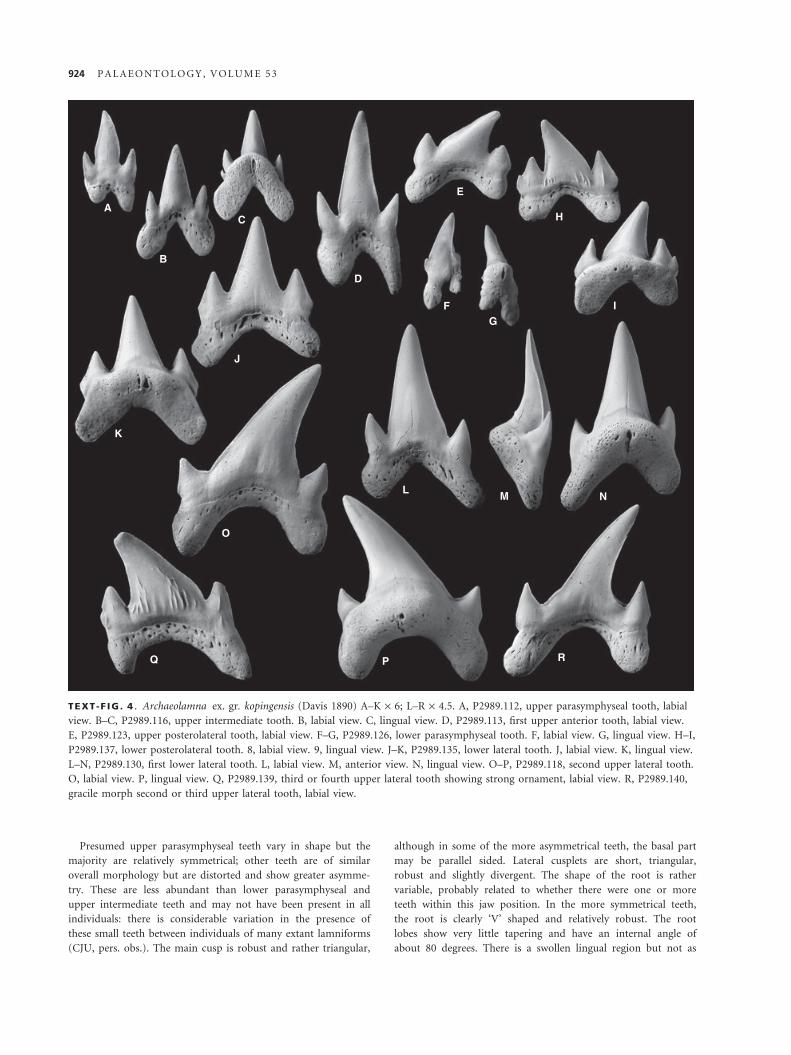
Presumed upper parasymphyseal teeth vary in shape but the
majority are relatively symmetrical; other teeth are of similar
overall morphology but are distorted and show greater asymme-
try. These are less abundant than lower parasymphyseal and
upper intermediate teeth and may not have been present in all
individuals: there is considerable variation in the presence of
these small teeth between individuals of many extant lamniforms
(CJU, pers. obs.). The main cusp is robust and rather triangular,
although in some of the more asymmetrical teeth, the basal part
may be parallel sided. Lateral cusplets are short, triangular,
robust and slightly divergent. The shape of the root is rather
variable, probably related to whether there were one or more
teeth within this jaw position. In the more symmetrical teeth,
the root is clearly ‘V’ shaped and relatively robust. The root
lobes show very little tapering and have an internal angle of
about 80 degrees. There is a swollen lingual region but not as
A
B
C
D
E
FG
H
I
NML
J
K
O
Q P R
TEXT -F IG . 4 . Archaeolamna ex. gr. kopingensis (Davis 1890) A–K · 6; L–R · 4.5. A, P2989.112, upper parasymphyseal tooth, labial
view. B–C, P2989.116, upper intermediate tooth. B, labial view. C, lingual view. D, P2989.113, first upper anterior tooth, labial view.
E, P2989.123, upper posterolateral tooth, labial view. F–G, P2989.126, lower parasymphyseal tooth. F, labial view. G, lingual view. H–I,
P2989.137, lower posterolateral tooth. 8, labial view. 9, lingual view. J–K, P2989.135, lower lateral tooth. J, labial view. K, lingual view.
L–N, P2989.130, first lower lateral tooth. L, labial view. M, anterior view. N, lingual view. O–P, P2989.118, second upper lateral tooth.
O, labial view. P, lingual view. Q, P2989.139, third or fourth upper lateral tooth showing strong ornament, labial view. R, P2989.140,
gracile morph second or third upper lateral tooth, labial view.
924 P A L A E O N T O L O G Y , V O L U M E 5 3

enlarged as in most anterior teeth, and the root lobes are oval in
cross-section without a well-defined linguo-basal face of the
root. In the remainder of the teeth from this proposed position,
the root is more compressed antero-posteriorly, creating a larger
lingual bulge. The root lobes are short and not often well
defined, comprising considerably less than half of the height of
the root.
Upper intermediate teeth are common within the assemblage,
and it is, therefore, likely that several files of these were present
in the jaw. They are symmetrical and have an erect and triangu-
lar main cusp flanked by a pair of slightly divergent cusplets. In
respects other than size, the crown of these teeth resembles that
of lower anterior teeth. The root is strongly ‘V’ shaped and has
an internal angle of 90 degrees or less. The root comprises about
half of the height of the tooth. The root is variably compressed
labiolingually. In some teeth, the root lobes are oval in cross-sec-
tion with a slightly flattened linguo-basal root base, whereas in
other teeth the linguo-basal face of the root is very flattened,
even including the region directly below the main cusp. This
flattening extends to the root lobes, which are about twice as
wide as they are thick. The lingual foramen is present but very
small. In addition, there are a very small number of probable
upper intermediate teeth that are small and squat. A triangular
main cusp is compressed labiolingually and flanked by a pair of
lateral cusplets that are similar in shape to those of lower lateral
teeth. The lingual face of the crown is wide at the base and is
either smooth or ornamented with irregular wrinkles similar to
those seen in other teeth. The lingual face of the crown is
smooth other than a very well-developed neck.
Upper lateral and posterior teeth all have a similar morphol-
ogy, varying largely in their size and the height of the cusps.
The first upper lateral tooth has a morphology somewhat inter-
mediate between that of the anterior teeth and the remaining
lateral teeth. The main cusp is robust and has a smooth curve
along both anterior and posterior edge. In all but in the small-
est posterior teeth, the main cusp is of similar height to the
root. The axis of the main cusp forms an angle of about 60
degrees with the base of the tooth. There is a single pair of
robust lateral cusplets of similar size to each other which are
slightly higher than wide. These are slightly divergent, but in
more posterolateral teeth, the posterior lateral cusplet is often
more erected than the main cusp. In the most posterior teeth,
the anterior cusplet may be reduced or incipient. The labial face
of the crown is smooth and slightly convex, although on some
very large teeth and some posterolateral teeth, small vertical
wrinkles may be present just above the crown-root junction.
The three cusps are united by a narrow basal strip of enameloid
on the labial side of the tooth, but not on the lingual side.
There, a very prominent neck is present at the base of each of
the cusps. The lingual face of the cusps is smooth and strongly
convex. There is a well-developed cutting edge across all of the
cusps. The root of upper lateral teeth is bulky and quite sym-
metrical, and the rather stubby root lobes form an internal
angle of 90–100 degrees. The outer edge of the anterior root
lobe continues from the edge of the main cusp, with a rounded
termination. The labial face of the root slopes gently away from
the base of the crown, and there is no labial crown overhang.
The linguo-basal face of the root is slightly flattened, with no
clear edge to the face. The lingual boss is large and rounded,
with one or sometimes two or three foramina at the apex. In
some smaller teeth, a very short but deep groove is present.
Some small foramina are seen on the upper parts of the root
labial face, but are rarely conspicuous.
Lower lateral teeth are very close to symmetrical, with the
exception of the probable first lower tooth, which is very similar
in shape to the third anterior tooth, and the smaller posterolat-
eral teeth. The main cusp is straight and quite short, with
slightly concave outer edges. There is a single pair of relatively
large lateral cusplets, that are higher than wide. The base of the
lateral cusplets is in line with the base of the main cusp, as
opposed to being some way down the outer edge of the root
lobes as in teeth from other positions in the jaw. The outer edge
of the cusplets overhangs the root slightly, and in some postero-
lateral teeth this projection develops into an additional incipient
cusplet. The labial face of the crown is very slightly convex to
almost flat, and has no ornament in the vast majority of teeth.
The basal edge of the enameloid on the labial side is gently
arched, as opposed to the strong curvature on other teeth. The
lingual face of the cusps is smooth and strongly convex, with a
well-developed neck at the base of each of the cusps. In poster-
ior teeth, the main cusp is strongly inclined as in the upper
teeth, and upper and lower teeth can only be differentiated by
the shape of the root. The root of lower lateral teeth is far more
‘square’ than on teeth from other positions; viewed lingually, the
top of the root is only slightly arched upwards, and the edges of
the root lobes are almost vertical below the outer edges of the
lateral cusplets, giving a very sharp angle between these edges.
The inner edges of the root lobes form a smooth semicircle in
most teeth, becoming more angular in posterior teeth. The root
lobes themselves have rounded terminations and are somewhat
flattened labiolingually. The labial face of the root smoothly
curves away from the crown, with a slight recess below the base
of the main cusp. The linguo-basal face of the root is large,
making up almost all of the root that can be seen in lingual
view, and relatively flat. There is a distinct but not sharply
angled edge to this face. The lingual boss projects slightly, giving
a narrow horizontal shelf below the cusps. There are one or
more foramina at the apex of the lingual boss, and in some
smaller teeth these are opened out to form a short but distinct
groove. Small foramina are present towards upper part of
the labial face of the root, but are small and irregularly
distributed.
Discussion. Archaeolamna is known from the Albian to
Maastrichtian and appears to have had a virtually global
distribution (see Siverson 1996). Of these occurrences, the
majority of records relate to Archaeolamna kopingensis
(Davis, 1890), although stratigraphical variations suggest
that this may represent a group of closely related taxa
rather than a single species. In addition to A. kopingensis
s.l., A. haigi Siverson, 1996 from the Cenomanian of Aus-
tralia and A. kopingensis judithensis Siverson, 1992 from
the Campanian of the USA (the latter probably deserving
of specific rank) are known.
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 925

Despite the very high degrees of heterodonty present, it
is possible to make general comparisons between the
Archaeolamna kopingensis s.l. specimens described here
and from other populations. Albian specimens from
Europe (Biddle 1993) are typically rather gracile and pos-
sess a poorly formed nutritive groove in most or all teeth.
Albian specimens from Australia (Siverson 1997) are
similar, but the nutritive groove is restricted to smaller
teeth. Archaeolamna kopingensis s.l. specimens from the
Cenomanian of Australia (Siverson 1996) and probably
France (Vullo et al. 2007), as well as younger occurrences,
lack a nutritive groove and possess weak folds on the
labial face of the larger teeth. The specimens described
here are therefore most similar to those from the Austra-
lian Albian, differing only in the possession of weak
ornament on the labial face of a small number of the very
largest and most posterior teeth. Although there are slight
differences between different populations of A. kopingen-
sis, it has not been possibly to produce a reliable set of
criteria for separating species within what almost certainly
represents a complex of morphologically similar taxa.
Within the ‘mid’ Cretaceous, A. kopingensis s.l. is wide-
spread but not global in occurrence. It is common at this
site and elsewhere in the Cenomanian of central Canada
(SC, pers. obs.) as well as in Colorado (Shimada et al.
2006). It is also present in western Europe (e.g. Biddle
1993; Vullo et al. 2007; CJU, pers. obs.), Russia (CJU,
pers. obs. of specimens mislabelled as other species) and
Australia (Siverson 1996, 1997). Despite this, it has not
been recorded in Texas (e.g. Welton and Farish 1993;
Cappetta and Case 1999) or elsewhere in areas associated
with Tethys. It, therefore, appears to have been a temper-
ate water inhabitant, present in both Northern and
Southern Hemispheres.
Family ?ARCHAEOLAMNIDAE fam. nov.
Genus TELODONTASPIS gen. nov.
Type species. Telodontaspis agassizensis sp. nov.
Derivation of name. From the spear-like tooth morphology and
an Odontaspis-like overall tooth shape.
Diagnosis. Strong monognathic and dignathic heterodont
dentition with anterior teeth up to 12 mm high. Several
files of upper and lower parasymphyseal teeth and upper
intermediate teeth present. Cusp unornamented, slender
and straight, at least 3 times as high as wide, inclined to
posterior in lateral teeth and faintly labiolingually sigmoi-
dal in anterior teeth. Single pair of small, triangular, lat-
eral cusplets present in upper lateral teeth, absent in all
other teeth. Base of crown with prominent neck on lin-
gual side, and extends along upper third to half of root
lobe on labial side. Cutting edge continuous in lateral
teeth but not reaching base of crown on anterior teeth.
Crown does not overhang root. Root with two clearly
defined lobes, of roughly equal size in lower teeth, but
with anterior lobe longer than posterior in upper teeth.
Lingual face of root with poorly developed lingual protu-
berance, below which root lobes are either labiolingually
compressed with a flat linguo-basal root face in upper
lateral teeth, or somewhat expanded lingually with no
clearly defined linguo-basal root face. There is a well-
developed round to slightly elongate foramen at the apex
of the lingual protuberance, with very small foramina
spread across the root surface.
Remarks. The relatively small number of teeth assigned to
the single known species of this genus, and the high
degree of heterodonty, make reconstruction of the denti-
tion less certain than with some other taxa present in this
assemblage. This is made more difficult by the presence
of different proportions of teeth in the fossil assemblage
than would be expected in the jaw of a living individual,
probably because of a combination of differential shed-
ding rates, a range of ontogenetic stages being present
and likelihood of random bias in a small sample size. The
teeth collected comprise a relatively large number of teeth
assigned to parasymphyseal files, and relatively few teeth
considered to be lower lateral positions. It is possible that
more material would require a different reconstruction of
the dentition to be developed. Despite this, it is evident
that the overall dental formula is similar to that of
Archaeolamna, with three upper anterior tooth positions,
upper and lower parasymphyseal teeth, and upper inter-
mediate teeth all being present. In addition, there appears
to be a morphological gradation from lower parasymphy-
seal to lower anterior teeth, as seen in Archaeolamna.
Despite this, the number of other lower and upper para-
symphyseal teeth suggest that multiple files of these were
present in life, a character present in lower jaws of Odon-
taspis noronhai but not present in the upper jaw of other
known lamniforms. The overall tooth shape is not unlike
that of Cretoxyrhina mantelli (Agassiz, 1843), but Tel-
odontaspis differs in possessing lower parasymphyseal and
upper intermediate teeth and a more strongly developed
heterodonty within the upper jaw. Teeth of T. agassizensis
sp. nov. are similar in overall morphology to teeth of neo-
nate individuals of Cretoxyrhina mantelli, and have previ-
ously been mistaken for that species. Despite this, teeth of
T. agassizensis sp. nov. differ in the range of tooth mor-
phologies present within the dentition, most of which
clearly do not fit in with the ontogenetic trend of C. man-
telli. Teeth are similar to those of Priscusurus adrupto-
dontus Kriwet, 2006, although this genus and species was
926 P A L A E O N T O L O G Y , V O L U M E 5 3

based on a very small sample of material, and so remains
poorly defined. Teeth of T. agassizensis sp. nov. differ from
those of P. adruptodontus in having a more slender main
cusp and, in upper lateral teeth, small lateral cusplets.
Telodontaspis agassizensis sp. nov.
Plate 6, figures 1–27
2006 Cretoxyrhina mantelli (Agassiz)(?); Shimada et al.,
p. 15, fig. 10.11–10.12.
Derivation of name. From the locality of the type material being
close to the shoreline of glacial Palaeolake Agassiz.
Holotype. P2989.144 (Pl. 6, figs 9–11).
Additional material. Forty-four teeth studied including P2989.
141–P2989.143; P2989.145–P2989.152.
Diagnosis. As for genus.
Description. The teeth of this species are very varied in shape
but occur in a number of discrete morphologies relating to posi-
tions within the jaw. The relatively small number of teeth pre-
vents the construction of a full dentition, and as a result, tooth
positions are partly inferred by comparison with other taxa.
There are two different morphologies of parasymphyseal teeth
(Pl. 6, figs 12–13, 18–20), with the commoner, often more asym-
metrical, teeth being considered here to come from the upper
dentition. These presumed upper parasymphyseal teeth have a
straight cusp that is up to half the total height of the tooth. The
cusp is straight and slender and, in labial view, tapers steadily
towards the apex. It is unornamented in all teeth. There is a very
slight lingual curvature in some teeth. The labial face of the cusp
is gently convex, separated from the strongly convex lingual face
by a narrow cutting edge, which is continuous in some teeth,
but only present on the upper half of either the anterior or pos-
terior edge of the cusp on others. There is a well-developed lin-
gual neck at the base of the cusp. The basal edge of the crown
on the labial side of the tooth is extended along the root lobes,
extending further along the anterior than posterior lobe. The
crown basal edge is flat and does not noticeably overhang the
root. On the majority of teeth, the cutting edge does not extend
along this basal extension of the crown. A small proportion of
teeth have an incipient cusplet on the mesial side of the crown.
There are two distinct root lobes, the posterior one being about
half the length of the anterior in the more asymmetrical teeth.
The anterior root lobe has a rounded apex, whilst that of the
posterior is usually more pointed. The labial face of both root
lobes is slightly concave and merges with the base of the crown,
There is a narrow shelf below the base of the crown on the lin-
gual side, terminating in a pointed protuberance with a well-
developed foramen at the apex. Below this, the root is strongly
labiolingually compressed and the linguo-basal root face is flat
to slightly concave. Small foramina are present over the entire
root surface.
Teeth referred to lower parasymphyseal positions are rather
different. The cusp is slender and is curved lingually and
slightly twisted to the posterior. The cutting edge is continuous
on the anterior edge, but does not reach the base on the pos-
terior side of most teeth. There is a pair of incipient cusplets
at the base of the cusp. The base of the crown is straight on
both faces, with a well-developed lingual neck. The labial face
of the crown does not overhang the root. The root is relatively
bulky, with two root lobes that form an angle of up to 90
degrees. Both root lobes are tapered and have variable rounded
terminations. The root lobes are not compressed in either ori-
entation, and the linguo-basal root face is convex. The root is
swollen below the lingual base of the crown and has a some-
what slit-like foramina at the apex. Small foramina are present
over the remainder of the root surface. Some teeth have a mor-
phology intermediate between this and presumed lower anterior
teeth, differing from the latter by being smaller and having
longer root lobes.
All of the anterior teeth have an elongate and slender cusp
that is higher than the root, and lateral cusplets are absent,
although an incipient distal cusplet is present on the presumed
third upper tooth. The cusp is weakly sigmoidal in all teeth, but
in labial view, is either straight or, in the probable first upper
teeth, very gently curved. It is inclined to the posterior in lower
and first upper anterior teeth, but inclined to the posterior in
the other upper anteriors. In the presumed first and second
lower teeth, and to a lesser extent other anterior teeth, the labial
face of the cusp narrows towards the base, with the cutting edges
moving onto the crown labial surface. At the base of the crown
is a well-developed neck in the lingual side, whilst on the labial
side the crown enameloid extends for about a third of the length
of the upper edge of the root lobes, the crown merging with the
root labial face without any overhang. There is a continuous
cutting edge which does not reach the crown base in any teeth.
The root is robust in all anterior teeth, but more gracile and
labiolingually compressed in the second and third upper teeth.
The root of lower anterior teeth is relatively expanded lingually,
resulting in a well-developed lingual protuberance. This has a
well-developed foramen at the apex. In some lower anterior
teeth, the root is incompletely developed, but the root lobes are
seen to be very short and stubby, considerably less than half the
total height of the root and both are of similar size and shape
with a rounded root lobe apex. In the third lower teeth, the root
lobes are somewhat longer and asymmetrical, with a more
pointed anterior root lobe apex and more rounded posterior
apex. There is a somewhat flattened area on the linguo-basal face
of the distal root lobe, but otherwise this face is not differenti-
ated. The upper anterior teeth have well-defined root lobes that
are asymmetrical, with the anterior lobe being longer than the
posterior. The root is expanded to form a lingual protuberance,
but this is not as distinct as in the lower teeth. The profile of
the root of the three presumed upper anterior teeth is rather dif-
ferent; in the first upper, the root lobes are short, and the ante-
rior lobe somewhat recurved to the posterior, whilst the roots of
the second and third teeth are strongly asymmetrical with a
longer anterior lobe. The root of the presumed third upper
tooth is somewhat more labiolingually compressed than that of
other anterior teeth, and the linguo-basal root face is more
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 927

clearly differentiated and flatter, thus resembling the root of the
upper lateral teeth.
Teeth presumed to be from upper lateral positions show some
morphological variation, with the first upper anterior being
clearly differentiated from the rest. Upper lateral teeth are
strongly asymmetrical, with the cusp strongly inclined to the
posterior. The cusp comprises up to half of the total height of
the tooth, being slender, straight and tapering smoothly to the
tip. There is no lingual curvature other than in the probable an-
teriormost teeth, where there is a gentle sigmoidal curve. The
labial face of the cusp is almost flat, and a continuous cutting
edge is present. At the base of the main cusp is a pair of very
small to (in the anteriormost teeth) incipient lateral cusplets,
which are triangular in profile and of similar size in both ante-
rior and posterior positions. The lingual face of the crown is
convex, nearly semicircular in section in most teeth but more
lingually expanded in the anteriormost teeth. There is a clear lin-
gual neck. The labial face of the crown extends along about half
of the upper edge of the root lobes, being flat and merging with
the root labial face. The root lobes are asymmetrical, becoming
less so in more posterior teeth. The anterior root lobe it slender
and parallel sided, with a rounded or slightly pointed apex. The
posterior root lobe is typically about half the length of the ante-
rior, and has a more angular termination. In the anteriormost
teeth, the most distal part of the posterior root lobe is twisted
lingually. The root is at its widest at the point of a poorly devel-
oped lingual protuberance, and then tapers gradually towards
the tips of the root lobes, resulting in the root lobes being very
labiolingually compressed and having a wide and flat linguo-
basal root face. The root of the anteriormost teeth is of the same
general shape, but with very little labiolingual compression,
resulting in a far more robust root. There is a prominent foram-
ina on the lingual protuberance, and small foramina are present
across the surface of the root of all teeth.
Upper intermediate teeth are similar in overall shape to those
from the mid part of the upper lateral files. They differ in being
smaller, reaching about half the maximum size, having a more
robust main cusp and having a pair of well-developed lateral
cusplets. Anterior and posterior cusplets are of similar size and
triangular in profile.
Presumed lower lateral teeth are in many respects similar to
anterior teeth. The teeth are relatively symmetrical, except in
posterior files where they are rather inclined to the posterior.
The cusp is straight and slender, with a very slight sigmoidal
curvature and comprises about half of the total height of the
tooth. The cusp labial face is weakly convex, and separated from
the strongly convex lingual face by a narrow but continuous cut-
ting edge. There are no lateral cusplets, and the base of the labial
face of the crown extends over the upper part of the root lobes,
where there is no basal overhang. There is a clear neck at the
lingual base of the crown. The outer edges of the root lobes
extend almost in continuation of the angle of the outer edges of
the cusp, giving the teeth a rather compressed ‘Y’ shaped profile.
The root lobes are clearly separated and rather gracile and paral-
lel sided. There is a well-developed lingual protuberance, and
below this the lingual side of the root lobes are somewhat
expanded lingually, with no clearly defined linguo-basal root
face. There is a single, well-developed, foramen at the apex of
the lingual protuberance, and numerous small foramina else-
where on the root.
Remarks. Teeth of Telodontaspis agassizensis sp. nov. are
present in the Cenomanian of Colorado (Shimada et al.
2006), and it is considered likely that this species has been
under recorded because of confusion with teeth of juve-
nile Cretoxyrhina.
Family ANACORACIDAE Casier, 1947
Genus SQUALICORAX Whitley, 1939
Type species. Corax pristodontus Agassiz, 1843 from the Maas-
trichtian of the Netherlands.
Squalicorax curvatus (Williston, 1900)
Plate 7, figures 1–23; Plate 8, figure 2
1900 Corax curvatus Williston pl. 30, figs 7–8.
1990 Squalicorax falcatus (Agassiz); Case et al., p. 1091,
fig. 10a–p, p. 1092, fig. 11a–b.
?1993 Squalicorax curvatus (Williston, 1900); Welton and
Farish, p. 116, figs 3–4.
1993 Squalicorax falcatus (Agassiz); Williamson et al.,
p. 457, fig. 7.4–7.5.
1999 Squalicorax sp. (group falcatus); Cappetta and Case,
p. 63, pl. 6, figs 4–9.
EXPLANATION OF PLATE 6
Figs 1–27. Telodontaspis agassizensis sp. nov.; all ·4.5. 1–2, P2989.141, ? lower parasymphyseal tooth. 1, labial view. 2, lingual view.
3–5, P2989.142, ? first upper anterior tooth. 3, labial view. 4, anterior view. 5, lingual view. 6–8, P2989.143, ? first upper lateral
tooth. 6, labial view. 7, anterior view. 8, lingual view. 9–11, P2989.144, holotype, ? third upper anterior tooth. 9, anterior view.
10, labial view. 11, lingual view. 12–13, P2989.145, ? upper parasymphyseal tooth. 12, labial view. 13, lingual view. 14–15,
P2989.146, ? upper intermediate tooth. 14, labial view. 15, lingual view. 16–17, P2989.147, ? upper lateral tooth. 16, lingual view.
17, labial view. 18–20, P2989.148, ? lower parasymphyseal tooth. 18, labial view. 19, anterior view. 20, lingual view. 21–22,
P2989.149, ? lower posterior tooth. 21, labial view. 22, lingual view. 23–24, P2989.150, ? second lower anterior tooth. 23, labial
view. 24, lingual view. 25–26, P2989.151, ? third upper anterior tooth of juvenile. 25, labial view. 26, lingual view. 27, P2989.152,
? incomplete first lower anterior tooth, labial view.
928 P A L A E O N T O L O G Y , V O L U M E 5 3
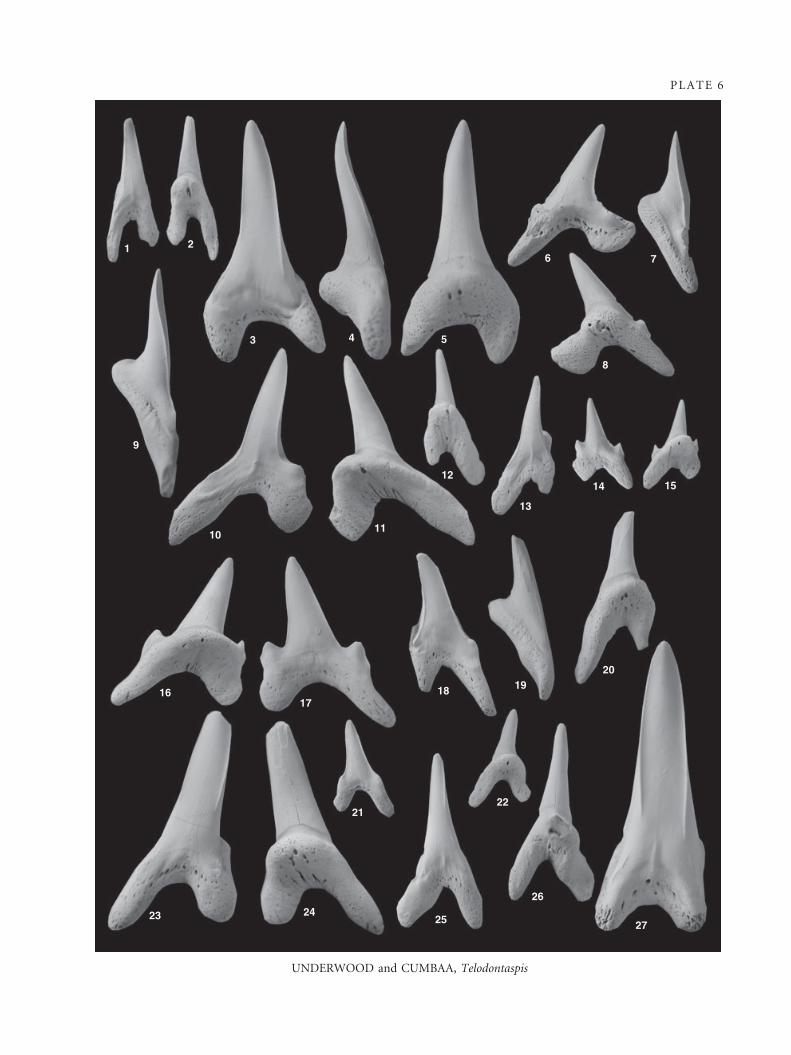
PLATE 6
UNDERWOOD and CUMBAA, Telodontaspis
21
3 4 5
8
76
9
1011
12
13
14 15
1617
18 1920
2122
23 2425
26
27

1999 Squalicorax curvatus (Williston); Cappetta and
Case, p. 65, pl. 7, figs 6–8.
1999 Squalicorax sp. Cumbaa and Tokaryk, p. 61, fig. 5
(pars; 5 teeth in right 2 columns).
?2001b Squalicorax sp. cf. S. curvatus (Williston);
Cicimurri, p. 185, fig. 5g.
2006 Squalicorax falcatus (Agassiz); Cumbaa et al.,
p. 146, fig. 4.9.
2006 Squalicorax curvatus (Williston); Shimada et al.,
p. 9, fig. 8.2–8.4.
2006 Squalicorax falcatus (Agassiz); Shimada et al., p. 9,
fig. 8.5–8.9.
2006 Squalicorax sp. Shimada et al., p. 9, fig. 8.10–8.14.
2008 Squalicorax sp. Cook et al., p. 1189, fig. 5B.
Material. About 800 teeth studied including P2989.153–
P2989.166.
Description. Teeth of this species are up to 14 mm wide and
show considerable gradient monognathic heterodonty as well as,
in anterior files, dignathic heterodonty. By comparison with
modern lamniform and carcharhiniform sharks, it is presumed
that teeth with more slender and erect cusps which are associ-
ated with a more swollen labial overhang with a somewhat ‘W’
shaped basal edge of the crown are from lower anterior posi-
tions.
The crown of the teeth varies from 75 to 200 per cent as
wide as high depending on jaw position, with only extreme
anterior teeth being higher than wide. The crown comprises a
main cusp and a small but clearly defined distal heel, the latter
being between one fifth and one-quarter of the total width of
the tooth. The main cusp is never symmetrical, other than in
some parasymphyseal teeth, and is inclined to the posterior to
a varying degree. The labial face is faintly convex, with a
strongly convex lingual face, neither with any ornamentation.
The anterior edge of the main cusp is very variable in shape.
In most teeth, the basal part of the anterior edge is straight
and at a lower angle than the bulk of the edge, with a clear
angular contact between the two parts of the edge. In pre-
sumed lower teeth, this basal section is shorter and less well
defined than in upper teeth. After this the anterior cusp edge
ranges from straight to clearly convex, but is faintly convex in
most teeth. The apex of the cusp typically forms and angle of
50–60 degrees. The posterior edge of the cusp is straight to
slightly convex and varies from near vertical (in lower anterior
teeth and some larger posterolateral teeth) to strongly over-
hanging (in most teeth). Only rare presumed upper anterolat-
eral teeth have the cusp apex reaching beyond the posterior
margin of the rest of the tooth. The distal heel is low and has
a straight upper edge and convex posterior edge. There is a
sharp angle between this and the main cusp of between 80 and
125 degrees, only being less than 90 degrees in some lower lat-
eral teeth. Continuous serrations are present over the cusp and
distal heel; these get smaller towards the tooth apex but are
otherwise very regular, with between 5 and 7 serrations per
millimetre. The serrations are roughly semicircular in profile
and symmetrical, with no sign of secondary serrations. The
base of the labial face of the crown extends down over
the upper third to half of the root. In presumed upper teeth,
the labial base of the crown clearly overhangs the root with a
straight to slightly concave basal edge, without flaring towards
the base. The base of the labial crown face of lower teeth
extends further in the anterior and posterior parts of the tooth
than in the centre, giving a rather ‘W’ shaped profile in some
teeth. There is a prominent overhang along all of the labial
crown base, but it is particularly evident where the crown
reaches its lowest points, where the crown edge is flared labi-
ally to form a narrow ledge. The lingual base of the crown has
a very obvious neck, which is of a relatively constant height
across the width of the tooth. The roots of all teeth are close
to symmetrical and are robust and thick. The root lobes are
poorly defined, with an angle of 100–140 degrees between the
anterior and posterior parts of the root. The root is of similar
width to the crown, but the relative width of root and crown
differ greatly with jaw position. The ends of the root lobes are
commonly rounded, but the anterior root lobe may be more
sharply pointed in lateral teeth, whilst a small linguo-distally
projecting boss is present at the posterior of the lingual face of
many lateral teeth. The labial root face is high and flat, and
may be somewhat concave in the central part. There is a clear
but rounded edge between this and the linguo-basal root face,
which is quite flat. The lingual face of the root also forms a
clear but rounded edge with the linguo-basal root face, and is
flat and of a constant height across the width of the tooth.
The central part of the lingual side of the root is rather swol-
len, and extends further lingually than the rest of the tooth.
There is a small but obvious foramen close to the junction of
the lingual and linguo-basal root faces in some teeth, but these
are in the minority. There is a row of large and obvious
foramina just below the base of the labial face of the crown in
EXPLANATION OF PLATE 7
Figs 1–23. Squalicorax curvatus (Williston, 1900); all ·4.5. 1–2, P2989.153, parasymphyseal tooth. 1, labial view. 2, lingual view. 3–4,
P2989.154, ? upper anterior tooth. 3, labial view. 4, lingual view. 5–6, P2989.155, ? upper anterior tooth. 5, labial view. 6, lingual
view. 7–8, P2989.156, ? upper anterolateral tooth. 7, labial view. 8, lingual view. 9–10, P2989.157, ? upper anterolateral
tooth showing occlusion damage. 9, labial view. 10, lingual view. 11, P2989.158, ? lower anterior tooth, labial view. 12, P2989.159,
? lower anterior tooth, labial view. 13–14, P2989.160, ? lower anterolateral tooth. 13, labial view. 14, lingual view. 15–16,
P2989.161, ? upper lateral tooth. 15, labial view. 16, lingual view. 17–18, P2989.162, posterolateral tooth. 17, labial view. 18,
lingual view. 19-20, P2989.163, posterolateral tooth. 19, labial view. 20, lingual view. 21–22, P2989.164, posterior tooth. 21, labial
view. 22, lingual view. 23, P2989.165, posterolateral tooth, labial view.
930 P A L A E O N T O L O G Y , V O L U M E 5 3

PLATE 7
UNDERWOOD and CUMBAA, Squalicorax
1
23 4
5 6
78
910
11
12 13
14
15 16
18
17
20
19
21 23 22

all teeth, and small and irregular foramina are present across
all faces of the root. The tip of the cusp and ends of the root
lobes are slightly bent labially so that the tooth has an overall
concave labial face in most teeth.
Discussion. The assemblage of teeth assigned to this spe-
cies includes a quite diverse range of morphologies, but
these all fit well within a single reconstructed dentition.
Similarities in the root, serration, proportions and size of
the teeth also suggest the presence of only a single species.
The type material of Squalicorax curvatus Williston, 1900
comprises two teeth, both of which compare well to teeth
described here, and probably represent anterolateral and
posterolateral positions. Subsequent descriptions of Squa-
licorax from the northern part of the Western Interior
Seaway have recognized two morphotypes, and have
either included all in S. falcatus Agassiz, 1843 (e.g. Case
et al. 1990) or have separated them into S. falcatus and
S. curvatus (e.g. Cumbaa et al. 2006; Shimada et al. 2006).
Although some teeth can readily be placed into one of
these morphotypes, others cannot, and Shimada et al.
(2006) comment that the teeth they identify as S. falcatus
appear to grade into S. curvatus and S. sp. (the last prob-
ably representing lower anterior teeth as described here).
It is now recognized that the diversity of anacoracids is
rather higher than was previously appreciated (e.g. Capp-
etta and Case 1999; Siverson et al. 2007), and some
names, especially Squalicorax falcatus, have been used for
a number of different species. The holotype of S. falcatus
(see Siverson et al. 2007) falls outside the morphological
range of the material described here, having a less clearly
defined distal heel, high cusp with smoothly convex ante-
rior edge and probably smaller serrations and more labio-
lingual compression. Although there has been confusion
between S. curvatus and S. baharijensis Stromer, 1927 (e.g.
Welton and Farish 1993; see Siverson et al. 2007), most
teeth of S. curvatus are considerably more erect and have
a shorter cusp, whilst presumed lower teeth are higher
and have a shorter and wider cusp with a more continu-
ously deep lingual neck.
Squalicorax curvatus was taken as the type species for
the genus Praeptychocorax Glickman, 1980, but the genus
is poorly defined and appears to be largely based on the
supposition that it is ancestral to Ptychocorax Glickman
and Istchenko in Glickman, 1980, and is here considered
invalid.
Genus PALAEOANACORAX Glickman and Shvazhaite, 1971
Type species. Palaeoanacorax volgensis Glickman in Glickman
and Shvazhaite, 1971 from the Cenomanian of Russia.
Remarks. The genus Palaeoanacorax was erected by Glick-
man (Glickman and Shavzhaite 1971) for a group of ana-
coracid species defined both on their morphology and
stratigraphical position. Although the definition of a genus
on stratigraphical grounds should be considered invalid,
the group of species, exemplified by Palaeoanacorax
volgensis Glickman, 1971, included into this genus are
morphologically distinct from Squalicorax Whitley, 1939.
Teeth of Palaeoanacorax differ from those of Squalicorax
s.s. in having a lower crown with well-defined cusp, thicker
and lower root which is overhung by the crown labially,
greater degree of heterodonty and serrations that are incip-
ient, irregular or missing altogether. Whilst Glickman
(1971) did not include Squalicorax falcatus and related
forms into Palaeoanacorax, they were included into the
genus by Sokolov (1978) in some parts of a paper that
elsewhere retains them in Squalicorax (of current usage).
It is considered here that Palaeoanacorax is a valid
genus and includes Squalicorax pawpawensis Siverson
et al., 2007, Squalicorax microserratodon Shimada, 2008,
Squalicorax primulus Muller and Diedrich, 1991, Squalico-
rax priscoserratus Siverson et al., 2007 and Palaeoanacorax
volgensis Glickman, 1971, as well as a number of
unnamed species from the Cenomanian (e.g. Siverson
1997), Turonian ⁄ Coniacian (Cappetta and Case 1999),
Santonian and Campanian (Underwood and Ward 2008).
Eoanacorax Glickman, 1971 is probably a separate valid
EXPLANATION OF PLATE 8
Fig. 1. Ptychodus rhombodus sp. nov.; ·25; P2989.61, posterior tooth, occlusal view.
Fig. 2. Squalicorax curvatus (Williston, 1900); ·25; P2989.166, parasymphyseal tooth, labial view.
Figs 3–5. Palaeoanacorax aff. pawpawensis (Siverson et al., 2007); all ·25. 3, P2989.175, parasymphyseal tooth, labial view. 4–5,
P2989.176, parasymphyseal tooth. 4, labial view. 5, lingual view.
Figs 6–18. Orectoloboides angulatus sp. nov.; all ·25. 6–8, P2989.177, posterior tooth. 6, labial view. 7, oblique view. 8, lingual view. 9–
12, P2989.178, holotype, anterior tooth. 9, labial view. 10, oblique view. 11, lingual view. 12, occlusal view. 13–15, P2989.179,
anterior tooth. 13, oblique view. 14, labial view. 15, oblique lingual view. 16–18, P2989.180, posterior tooth. 16, labial view. 17,
oblique view. 18, lingual view.
Figs 19–22. Cretorectolobus robustus sp. nov.; ·12.5; P2989.188, holotype, lateral tooth, 18, labial view. 19, occlusal view. 20, lingual
view. 21, oblique view.
932 P A L A E O N T O L O G Y , V O L U M E 5 3
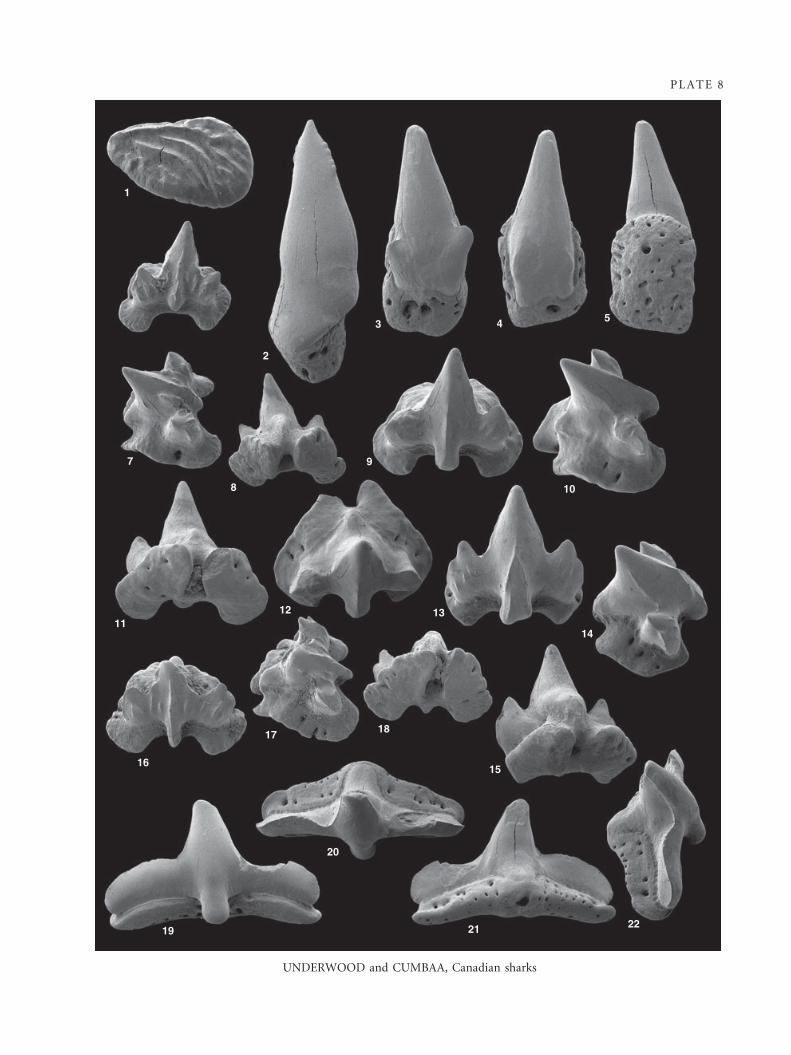
PLATE 8
UNDERWOOD and CUMBAA, Canadian sharks
1
2
3 4 5
7
8
9
10
1112 13
14
16
17 18
15
2221
20
19
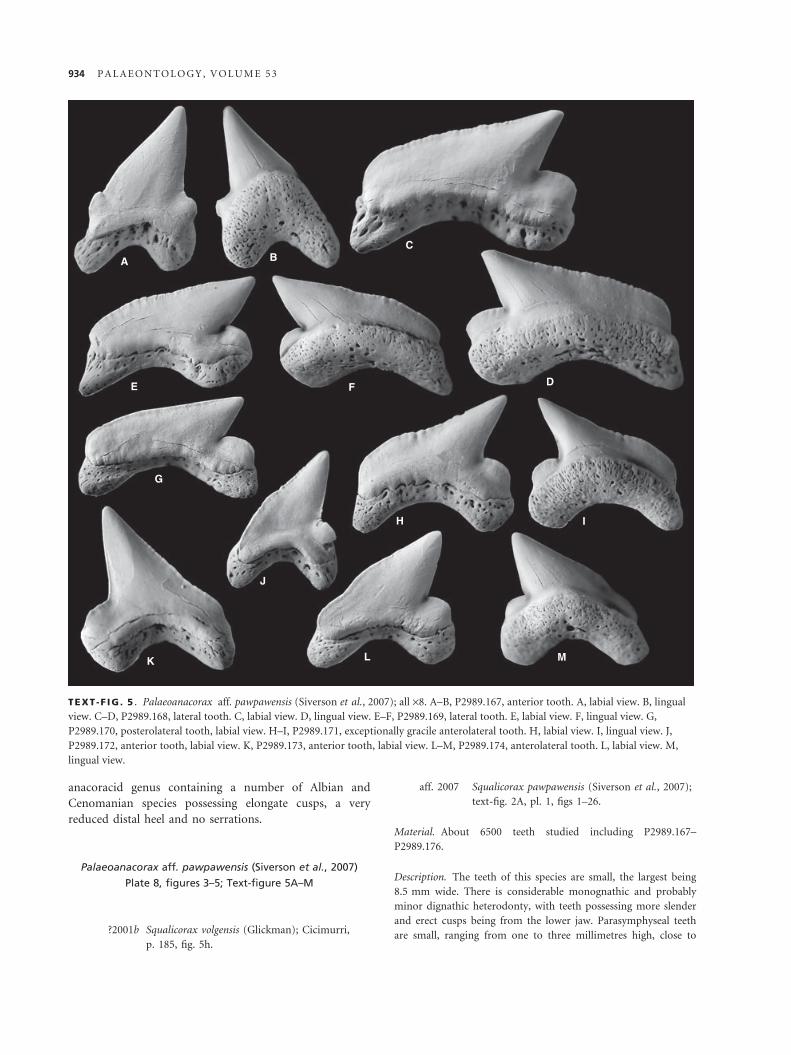
anacoracid genus containing a number of Albian and
Cenomanian species possessing elongate cusps, a very
reduced distal heel and no serrations.
Palaeoanacorax aff. pawpawensis (Siverson et al., 2007)
Plate 8, figures 3–5; Text-figure 5A–M
?2001b Squalicorax volgensis (Glickman); Cicimurri,
p. 185, fig. 5h.
aff. 2007 Squalicorax pawpawensis (Siverson et al., 2007);
text-fig. 2A, pl. 1, figs 1–26.
Material. About 6500 teeth studied including P2989.167–
P2989.176.
Description. The teeth of this species are small, the largest being
8.5 mm wide. There is considerable monognathic and probably
minor dignathic heterodonty, with teeth possessing more slender
and erect cusps being from the lower jaw. Parasymphyseal teeth
are small, ranging from one to three millimetres high, close to
A BC
DE F
H
G
I
J
K L M
TEXT -F IG . 5 . Palaeoanacorax aff. pawpawensis (Siverson et al., 2007); all ·8. A–B, P2989.167, anterior tooth. A, labial view. B, lingual
view. C–D, P2989.168, lateral tooth. C, labial view. D, lingual view. E–F, P2989.169, lateral tooth. E, labial view. F, lingual view. G,
P2989.170, posterolateral tooth, labial view. H–I, P2989.171, exceptionally gracile anterolateral tooth. H, labial view. I, lingual view. J,
P2989.172, anterior tooth, labial view. K, P2989.173, anterior tooth, labial view. L–M, P2989.174, anterolateral tooth. L, labial view. M,
lingual view.
934 P A L A E O N T O L O G Y , V O L U M E 5 3

symmetrical and more than twice as high as wide. The larger
parasymphyseal teeth are similar to other anterior teeth other
than being considerably more laterally compressed and, in some
cases, possessing a heel or incipient cusplet on both anterior and
posterior sides of the teeth. Smaller parasymphyseal teeth com-
prise a conical to slightly lingually curved cusp projecting from
an oval to near spherical root. In these teeth, the cutting edge
may be severely reduced or even absent.
Of the remainder of the teeth, the crown varies from 75 to
200 per cent as wide as high depending on jaw position, with
only extreme anterior teeth being higher than wide. The main
cusp is strongly inclined to the posterior in all but the most
anterior teeth and is followed by a small but clearly defined dis-
tal heel, the latter being between one sixth and one fifth of the
total width of the tooth. The main cusp is inclined to the pos-
terior, weakly in the most anterior teeth, but very strongly, with
the anterior cutting edge being almost horizontal, in posterolat-
eral teeth. The labial face is faintly convex to flat, with a strongly
convex lingual face, neither with any ornamentation. The ante-
rior edge of the main cusp is convex towards the base, becoming
straight or weakly concave higher up. The apex of the cusp typi-
cally forms and angle of 35–60 degrees, being more acutely
pointed in presumed lower teeth. The posterior edge of the cusp
is straight to very faintly convex. In all teeth there is some
degree of overhang of the main cusp over the distal heel. In
more erect anterior teeth, the apex of the main cusp extends as
far to the posterior as the rear edge of the tooth, but this is not
the case with most teeth. The distal heel is very low, with the
upper edge typically sloping downwards from its anterior end,
and evenly convex. A continuous cutting edge extends along the
main cusp and distal heel. True serrations are not seen on any
teeth, but faint crenulations or incipient serrations are present
on the anterior cutting edge of all teeth except some parasym-
physeal teeth and some posterolateral teeth. These crenulations
do not reach either the apex of the cusp or anterior edge of the
cutting edge. The base of the labial face of the crown extends
down over the upper half of the root, which it strongly over-
hangs. The labial base of the crown is gently concave in all teeth,
and is weakly flared towards the posterior end in some teeth.
The lingual base of the crown has a very obvious neck, which is
highest below the main cusp. The roots of all teeth are gently
curved, with a similar depth of the root (in lingual view) present
along much of its length, resulting in a lack of clearly defined
root lobes. The labial root face is low, being higher at the ante-
rior and posterior ends than near the centre. There is a clear but
rounded edge between this and the linguo-basal root face, which
is smoothly convex, and not clearly differentiated from the low
lingual root face. The central part of the lingual side of the root
is rather swollen, and extends further lingually than the rest of
the tooth. The roots of all teeth are heavily vascularized. There
is a row of large and obvious foramina just below the base of
the labial face of the crown in all teeth, and the overhanging
base of the crown may be somewhat indented next to the foram-
ina. Large and irregular foramina are present across the linguo-
basal root face, often joining to form irregular, branching
grooves in the root. Foramina are present on the lingual face of
the root, but these are smaller than elsewhere and present within
faint vertical grooves.
Discussion. These teeth are very abundant and the large
sample size allows the very high degree of heterodonty
shown by this species to be recognized. There are large
numbers of small parasymphyseal teeth recognized within
the collection, suggesting that several files were present on
at least one or possibly both, jaws. This has not been rec-
ognized in other anacoracid taxa (e.g. Shimada 2008).
The smaller parasymphyseal teeth may easily be mistaken
for teeth of co-occurring Cretomanta canadensis Case
et al., 1990, but may be separated from them by, amongst
other things, a less erect cusp and presence of a cutting
edge.
Although very similar to the topotype material of Pal-
aeoanacorax volgensis as figured by Siverson et al. (2007),
the labial basal margin of the crown is gently concave,
and not strongly scalloped. In addition, the majority of
the teeth described here have a more slender cusp and
wider distal heel than figured specimens of Palaeoanaco-
rax volgensis. The same differences separate Palaeoanaco-
rax aff. pawpawensis from the species present in the
Australian Cenomanian (described as Squalicorax volgensis
by Siverson (1996)) and specimens from the Turonian ⁄Coniacian boundary of Texas (described as Squalicorax
volgensis by Cappetta and Case (1999)). Palaeoanacorax
aff. pawpawensis is most similar to P. pawpawensis (Siver-
son et al. 2007) (including specimens figured as P. volgen-
sis by Cappetta and Case (1999) and Welton and Farish
(1993)), but appears to differ in typically possessing a
shorter and more robust cusp and larger but less well-
developed serrations on the majority of teeth, in addition
to having a smaller maximum size. Although it is
probable that Palaeoanacorax aff. pawpawensis and
P. pawpawensis are different species, there is considerable
morphological overlap between populations of both, and
it is at present not possible to clearly differentiate these
taxa. Palaeoanacorax aff. pawpawensis is also very similar
to P. primulus (Muller and Diedrich 1991), and there is a
possibility that they could be conspecific, but the poor
preservational and illustration quality of holotype and
paratype specimens (Siverson et al. 2007) make compari-
sons difficult. Palaeoanacorax aff. pawpawensis teeth differ
from those of the Albian P. priscoserratus (Siverson et al.
2007) and the Coniacian P. microserratodon (Shimada
2008) in the form of serration and root.
Order ORECTOLOBIFORMES Applegate, 1972
INCERTAE FAMILIAE
Genus ORECTOLOBOIDES Cappetta, 1977
Type species. Ginglymostoma parvula Dalinkevicius, 1935 from
the ?Albian of Lithuania.
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 935

Orectoloboides angulatus sp. nov.
Plate 8, figures 6–18
Derivation of name. After the angular overall appearance of the
teeth.
Holotype. P2989.178 (Pl. 8, figs 9–12).
Material. Three teeth studied (P2989.177; P2989.179–P2989.180).
Diagnosis. Teeth small, under 2 mm in greatest dimen-
sion, showing moderate heterodonty. Tooth crown of
similar width to height. All teeth close to symmetrical,
with a robust and conical main cusp (less than 1.5 times
as high as wide) flanked by a single pair of short and
robust lateral cusplets, with a second pair of incipient
cusplets sometimes being present. A single ridge forming
a sharp-edged keel is present on the labial face, and
extends from half way up the main cusp to the base of a
narrow but very prominent labial boss, sometimes bifur-
cating near the base. Other, shorter, ridges are present
elsewhere on the crown but are always widely spaced and
never join. Lingual crown face strongly convex with a
very well-developed lingual protuberance at the base.
Other than the labial boss, the crown does not strongly
overhang the root. The root is wider and deeper than the
crown, with a very well-developed nutritive groove seen
in all teeth. The distal parts of the root lobes are some-
what recurved labially and have flared bases.
Description. Teeth of this species are small and robust, with an
overall very ‘angular’ appearance. All teeth are a similar height
to width. The main cusp is conical and higher than wide with
the base making up about half of the width of the tooth crown.
This is flanked by a single pair of short and robust lateral cus-
plets, which may have a second pair of incipient cusplets on
their outer margin. The lateral cusplets are about as high as wide
and slightly divergent. A well-developed cutting edge runs from
the outer edge of the tooth crown and over the cusplets, but
does not reach the apex of the main cusp. The labial surface of
the tooth crown extends over the labial face of the tooth for at
least the same distance as the height of the lateral cusplets. A
very prominent labial protuberance is present below the main
cusp. This is roughly parallel sided with a rounded termination,
and is about half of the width of the base of the main cusp. The
labial face of all cusps is weakly convex. A very prominent,
sharp-edged ridge extends from the tip of the labial protuber-
ance to about half way up the main cusp. In one tooth, this
bifurcated into two ridges on the labial protuberance Pl. 8, figs
13–15). Additional, short longitudinal ridges are present on the
more lateral parts of the crown labial face. In the two larger
teeth, they are short and very weak with two or three ridges
being present below the lateral cusplets. On the two smaller
teeth, these ridges are more strongly developed, with up to five
on either side of the tooth. The lingual side of the cusps is
strongly convex, and below the main cusp this becomes
expanded downwards to form a lingual uvula. The uvula pro-
jects lingually to varying degrees, and extends downwards to the
lingual end of the root nutritive groove. Other than at the point
of the labial protuberance, the labial basal edge of the crown
only weakly overhangs the top of the root. There is no crown
overhang of the root lingually, and the basal lingual edge of the
crown has a narrow smooth neck at the junction with the root.
The root is bulky and slightly wider than the crown in all teeth.
The root is very strongly bilobate, with a wide and deep nutri-
tive groove extending for the whole depth of the root. The labial
surface of the root lobes is laterally and labially flared basally,
giving a rounded profile to the lateral edge of the root. The late-
ro-labial face of the root lobes also flares downwards, especially
on either side of the nutritive groove, where it forms a pointed
lingual projection. There is a sharply angled edge of the basal
face of the root all round. The basal face of the root lobes is
smooth and only slightly convex. Other than at least one very
large foramina within the nutritive groove, foramina are few but
quite prominent, with two or three circular to oval foramina on
each latero-labial root face and one to four on each basal root
face.
Remarks. Teeth of Orectoloboides angulatus sp. nov. are
very similar to those of O. parvulus Dalinkevicius, 1935,
but differ in several ways. The cusps of O. parvulus are
higher and more slender, with this being especially evi-
dent in lateral cusplets, where a second pair of clearly
separated cusplets is often present (CJU, pers. obs.). In
addition, teeth of O. parvulus appear to show a greater
degree of heterodonty with the distinctive lateral tooth
morphology (e.g. Cappetta 1977, fig. 2) not being recog-
nized here. Teeth of O. multistriatus Werner, 1989 have a
considerably finer and more densely spaced ornament.
Teeth of O. angulatus sp. nov. are superficially similar to
teeth of some sclerorhynchids, but differ in the presence
of lateral cusplets, a strong ornament and a more arched
and flared root.
Genus CRETORECTOLOBUS Case, 1978
Type species. Cretorectolobus olsoni Case, 1978, from the Campa-
nian of Montana, USA.
Cretorectolobus robustus sp. nov
Plate 8, figures 19–22
Derivation of name. From the relatively large and robust teeth
compared to others of the genus.
Holotype and only specimen. P2989.188 (Pl. 8, figs 19–22). This
was collected as part of the Canadian Museum of Nature study
and not from the same sub sample as the rest of the material
described here.
936 P A L A E O N T O L O G Y , V O L U M E 5 3

Diagnosis. Near symmetrical tooth with single erect cusp.
Tooth considerably wider than high, with blade-like lat-
eral crown extensions of similar width to root. Lateral
crown extensions of uniform height without cusplets but
noticeably arched to give convex occlusal surface. Occlu-
sal cutting edge of lateral extensions sigmoidal in occlusal
view. Labial protuberance prominent but narrow and par-
allel sided with rounded apex. Uvula tapering and quite
weakly developed. Root low and somewhat extended lin-
gually in region of uvula. Hemiaulacorhize root vasculari-
zation with well-developed small foramina on all faces of
root except labial face.
Description. The single tooth known is from a presumed lateral
position and is 4.4 mm wide. The tooth has a general ‘squati-
noid’ morphology, with an erect cusp at close to right angles
from the flattened and triangular root. The cusp is triangular
and has a height less than one third of the total tooth width
and slightly linguo-posteriorly curved. The cusp is triangular
with a convex labial surface and strong convex lingual surface.
At its base, the cusp merges with lateral blades on both mesial
and distal sides. These blades are of relatively constant height,
but are somewhat curved with a convex occlusal surface with
no incipient cusplets. The distal ends of the lateral blades are
smoothly rounded. There is a well-developed cutting edge
extending for the entire occlusal surface of the crown. When
viewed occlusally, there is a distinct sigmoidal curvature to the
cutting edge on the lateral blades. Below the labial face of the
cusp is a well-developed labial protuberance which extends to
at least the full depth of the root. This is narrower than the
base of the cusp, parallel sided and with a rounded tip. A wide
but shallow uvula is present on the lingual side of the tooth,
extending from the base of the cusp to the lingual edge of the
root. There is no ornament on the crown. Other than around
the labial protuberance, there is no crown overhang of the
root, but the crown-root contact is sharp and well defined.
The root is low and triangular in basal view. The root is about
three times as wide as the labiolingual depth, with a straight
labial face and expanded lingually to the tip of the uvula. The
labial root face is as wide as the crown and slightly flared at
the base. There is a sharp but not angled contact between this
and the basal face, which is broadly concave. The lingual root
face is slopes from the base of the crown to a distinct contact
with the basal face. There is a large foramen near the centre of
the root basal face and another just below the lingual tip of
the root. Small foramina are scattered across all faces of the
root except the labial face.
Remarks. A number of genera have been proposed for
teeth with a general ‘squatinoid’ morphology from the
Cretaceous, with different species being assigned to either
the Orectolobiformes or the Squatiniformes. The tooth
described here is assigned to the former because of the
low and rather robust cusp, high lateral blades and a root
that is only weakly expanded lingually. Cretorectolobus
robustus sp. nov. differs from C. olsoni Case, 1978 from
the Campanian of North America in having a more
robust cusp, no incipient lateral cusplets, a more slender
labial protuberance and more random spread of foramina
over the root. C. gracilis Underwood and Mitchell, 1999
from the British Albian and C. doylei Underwood et al.,
1999 from the British Hauterivian have a far smaller max-
imum size then C. robustus sp. nov. C. gracilis also differs
in having very reduced lateral blades and a higher root,
whilst C. doylei has incipient lateral cusplets and a rela-
tively high root. Teeth of species of the related genus Ced-
erstroemia Siverson, 1995 are more robust overall, with a
flattened and triangular cusp.
Superorder BATOMORPHII Cappetta, 1980b
Order RAJIFORMES Berg, 1940
Family RHINOBATIDAE Muller and Henle, 1838
Genus RHINOBATOS Linck, 1790 s.l.
Type species. Rhinobatos rhinobatos Linnaeus, 1758; extant.
Remarks. The genus Rhinobatos has been used for teeth
and skeletons of a large number of extinct species of rhin-
obatid (e.g. Cappetta 2006). Although many taxa previ-
ously assigned to Rhinobatos have subsequently been
moved to other genera, many fossil species of Rhinobatos
remain, many of which will probably need to be moved
to other genera in the future.
‘Rhinobatos’ cf. incertus Cappetta, 1973
Text-figure 6A–H
1973 Rhinobatos incertus Cappetta, p. 508, figs 26–33.
1973 Rhinobatos sp. Cappetta, p. 508, figs 18–25.
1990 ?Rhinobatos sp. Case et al., p. 1093, figs 12–13.
1993 Rhinobatos sp. Williamson et al., p. 458, fig. 8.
1993 Rhinobatos incertus Cappetta; Welton and Farrish,
p. 132, figs 1–2.
2001a ?Rhinobatos incertus Cappetta; Cicimurri, p. 185,
fig. 5X.
2004 ?Rhinobatos incertus Cappetta; Cicimurri, p. 9,
fig. 5J.
2006 Rhinobatos sp. Shimada et al., p. 19, fig. 11.3–11.4.
2007 Rhinobatos incertus Cappetta; Everhart, p. 230
(pars), fig 6B–D (non A).
Material. Two teeth (P2989.181 and P2989.182).
Description. Both teeth are less than 1 mm across and have low
crowns. The teeth are similar in width to height and have a
crown that is somewhat larger than the root. The labial face of
the crown is weakly convex and rather wider than high, and
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 937
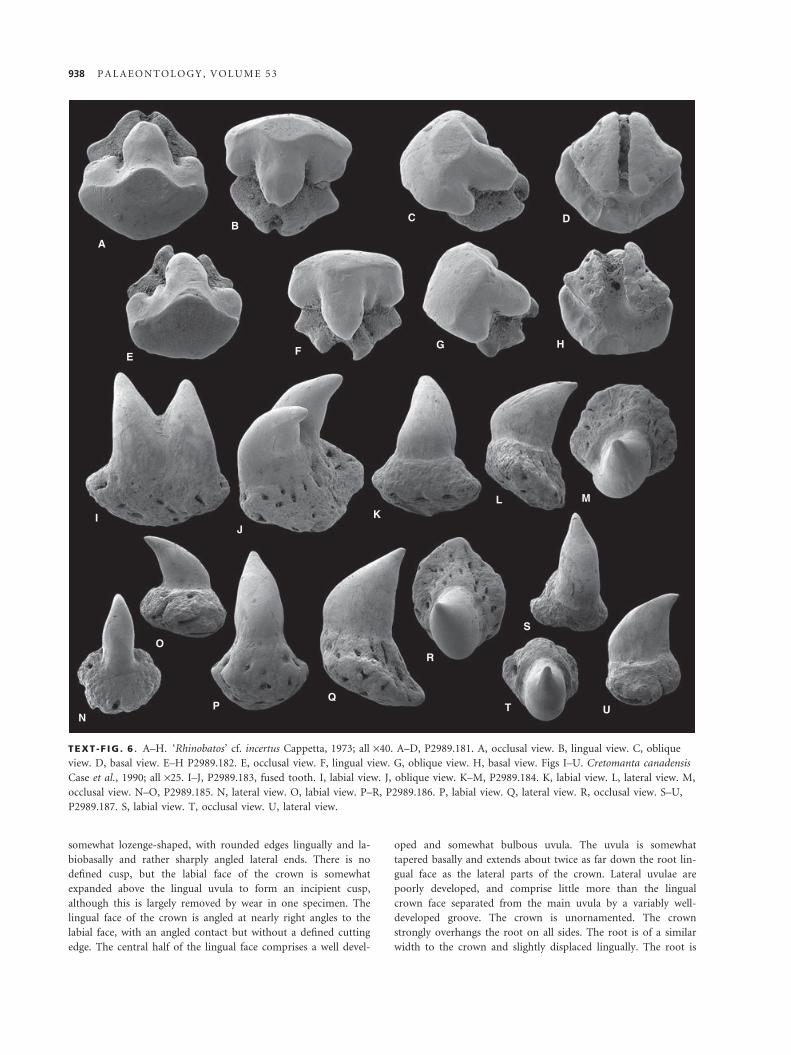
somewhat lozenge-shaped, with rounded edges lingually and la-
biobasally and rather sharply angled lateral ends. There is no
defined cusp, but the labial face of the crown is somewhat
expanded above the lingual uvula to form an incipient cusp,
although this is largely removed by wear in one specimen. The
lingual face of the crown is angled at nearly right angles to the
labial face, with an angled contact but without a defined cutting
edge. The central half of the lingual face comprises a well devel-
oped and somewhat bulbous uvula. The uvula is somewhat
tapered basally and extends about twice as far down the root lin-
gual face as the lateral parts of the crown. Lateral uvulae are
poorly developed, and comprise little more than the lingual
crown face separated from the main uvula by a variably well-
developed groove. The crown is unornamented. The crown
strongly overhangs the root on all sides. The root is of a similar
width to the crown and slightly displaced lingually. The root is
A
BC D
E FG H
JI K
L M
S
UT
R
QP
N
O
TEXT -F IG . 6 . A–H. ‘Rhinobatos’ cf. incertus Cappetta, 1973; all ·40. A–D, P2989.181. A, occlusal view. B, lingual view. C, oblique
view. D, basal view. E–H P2989.182. E, occlusal view. F, lingual view. G, oblique view. H, basal view. Figs I–U. Cretomanta canadensis
Case et al., 1990; all ·25. I–J, P2989.183, fused tooth. I, labial view. J, oblique view. K–M, P2989.184. K, labial view. L, lateral view. M,
occlusal view. N–O, P2989.185. N, lateral view. O, labial view. P–R, P2989.186. P, labial view. Q, lateral view. R, occlusal view. S–U,
P2989.187. S, labial view. T, occlusal view. U, lateral view.
938 P A L A E O N T O L O G Y , V O L U M E 5 3

strongly bilobate, with a wide and conspicuous nutritive groove.
The linguolateral root faces flare basally from beneath the crown,
and are relatively flat. The root basal faces are flat and slightly
angled laterally. In basal view, each root lobe basal face is trian-
gular, being of similar size in one tooth but rather asymmetrical
in the other, where the nutritive groove is wider. There is at
least one very large foramen within the nutritive groove, along
with one small foramen on each linguolateral root face and sev-
eral on each root lobe basal face.
Discussion. The teeth seen here appear to be the same
taxon as noncuspate teeth figured as Rhinobatos sp. by
Cappetta (1973). This species co-occurs with the cuspate
teeth that include the type material of R. incertus Cappet-
ta (1973), which it resembles in all but form of the occlu-
sal surface. In all instances where cuspate teeth referred
to, or resembling, R. incertus have been recorded, noncus-
pate teeth of the type recorded here are also present; in
addition, where an idea of proportions is recorded
(Cappetta 1973; Shimada et al. 2006), noncuspate teeth
far outnumber cuspate. It is therefore considered here (as
by Welton and Farish 1993 and Everhart 2007) that Rhin-
obatos sp. and R. incertus are conspecific, with the cuspate
morph representing teeth of breeding males.
Despite the poor quality of figures of some specimens
referred to this species (e.g. Case et al. 1990; Cicimurri
2001a, 2004), teeth appear to be widespread within the
Western Interior Seaway, and range from Upper Albian
to at least Coniacian (Everhart 2007). This species does
not, however, appear to be present in Europe or further
afield (e.g. Herman 1977; CJU, pers. obs.).
?BATOMORPHII incertae sedis
Genus CRETOMANTA Case et al., 1990
Type species. Cretomanta canadensis Case et al., 1990 from the basal
Turonian of Saskatchewan (published incorrectly as Coniacian).
Cretomanta canadensis Case et al., 1990
Text-figure 6I–U
1990 Cretomanta canadensis Case et al., p. 1093, fig. 14a–c.
1993 ?Rhincodontidae Welton and Farish, p. 86, figs 1–3.
2001a Cretomanta canadensis Case et al.; Cicimurri, p. 185,
fig. 5S–T.
2004 Cretomanta canadensis Case et al.; Cicimurri, p. 9,
fig. 5O.
2006 Cretomanta canadensis Case et al.; Shimada et al., p.
19, fig. 11.7–11.8.
Material. Forty-seven teeth studied including P2989.183–
P2989.187.
Description. Teeth of this species are up to 1.5 mm high and
are relatively homodont. The single cusp, or in the case of
one specimen, each of two equal sized cusps, comprises about
half of the total height of the tooth and is erect, being either
near vertical (relative to the root basal face) or slightly
inclined labially. The cusp is round in cross-section to slightly
laterally compressed, and lacks any cutting edge. The cusp is
one to 1.5 times as high as deep and strongly curved lin-
gually, with a curvature on the labial edge of 45–70 degrees.
There is a well-developed collar at the base of the cusp that
is somewhat constricted, with the thickest part of the cusp
being close to the centre, tapering to a slender and sharp
point.
The root is rounded and globular. In smaller teeth, it is
near spherical, but in most teeth the root is somewhat
expanded lingually. The labial face of the root is near vertical,
whereas other faces are flat to slightly concave, with the root
widening downwards and lingually to gently convex basal face.
Small foramina are spread across the root, but are concen-
trated on the mid part of the lateral root faces and on the
basal face.
In broken or transparent teeth, it is possible to see a slender,
tubular, pulp cavity running most of the length of the cusp.
Small tubules branch from this and are obvious within the
dentine.
Discussion. The affinity of Cretomanta canadensis is very
uncertain, largely because of the extremely convergent
and conservative morphology of teeth of microphagous
chondrichthyans. The overall morphology of the teeth is
extremely similar to that of the reduced parasymphyseal
teeth of Palaeoanacorax aff. pawpawensis. Teeth of
C. canadensis do, however, differ in having a more erect
and strongly curved cusp, a slightly fusiform labial cusp
profile, the lack of a cutting edge on all teeth, and a nar-
row pulp cavity. The form of vascularization suggests that
a lamniform affinity is not supported (contra Cappetta
2006), but may suggest a batomorph affinity. The pres-
ence of a ‘double’ tooth is similar to those present where
row splitting occurs in the dentition of some batoids (e.g.
Herman et al. 2000, textplate 5), and may further suggest
a batoid affinity. Although the tooth morphology is not
unlike that of some extant planktivorous batoids, it is
likely that this is convergently derived and as such family
level affinities are uncertain. Despite the morphology of
the teeth suggesting planktivory, faint longitudinal
scratches may be present, but it is not certain whether
these are a result of wear against rigid food items or
because of occlusion with teeth on the opposing jaw.
Teeth of similar morphology as the type material of Cre-
tomanta canadensis have been recognized (CJU, pers.
obs.) in rocks ranging in age from Cenomanian to Maas-
trichtian (Noubhani and Cappetta 1997), but it is cur-
rently uncertain whether this comprises several separate
species.
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 939

PALAEOECOLOGY
The assemblage recovered from the Bainbridge River
bonebed represents a relatively low diversity assemblage
dominated by pelagic predators. Although an abundance
of lamniforms appears to be a common feature of Creta-
ceous assemblages from the Western Interior Seaway (e.g.
Welton and Farish 1993; Williamson et al. 1993; Cappetta
and Case 1999), the almost complete lack of small necto-
benthic taxa in the Bainbridge River assemblage is excep-
tional. The elasmobranch fauna is associated with a very
rich and diverse assemblage of actinopterygians, with
remains of hesperornithiform birds also being common
(e.g. Cumbaa et al. 2006). Other nonavian reptiles are
relatively rare, with only plesiosaurs being regular finds.
The vast majority of the vertebrates appear to be active
predators, with teeth adapted for either clutching active
prey or, more rarely, cutting larger prey. Durophagous
taxa are relatively uncommon, although several species of
Ptychodus and pycnodonts are present. Teeth of Ptychodus
rarely appear to show wear, and so it is possible that the
main food source is not the same, or as hard shelled, as
that favoured by some Ptychodus species elsewhere. The
abundant remains of small teleosts and possibly teeth of
Cretomanta suggest that planktivores were common. The
extremely low number of teeth of benthic batoids and
orectolobids may be because of occasional transient
individuals. This assemblage therefore represents a very
high nutrient environment with a hostile sea floor.
Some of the species recorded here (Cretoxyrhina dentic-
ulata and Ptychodus ex. gr. decurrens) appear to have a
global distribution, but the majority are more restricted in
their range. Shallower water sites in the northern Western
Interior Seaway share some of the species with the fauna
described here (Case 2001; Cook et al. 2008) but contain
several lamniform and batoid taxa not present here, and
lack the Ptychodus. Comparisons of small tooth taxa are
not possible because of different collecting regimes. Many
of the taxa recorded here are also present (albeit previ-
ously unrecognized) at other sites within the offshore parts
of the northern Western Interior Seaway (e.g. Cappetta
1973; Cicimurri 2001b; Shimada et al. 2006), which also
contain Tethyan faunal elements not recorded at the Bain-
bridge River. The diverse chondrichthyan assemblages
known from the southern Western Interior Seaway in
Texas (e.g. Welton and Farish 1993; Cappetta and Case
1999) have little in common with those from the Bain-
bridge River, with only Squalicorax curvatus and ‘Rhino-
batos’ incertus present within Cenomanian rocks of both
regions but not seen outside the Seaway. This is consistent
with evidence from other vertebrate groups for at least
two distinct faunal provinces in the Western Interior Sea-
way (Nicholls and Russell 1990). In contrast, there are sev-
eral taxa present in Texas that are also present in Egypt
(Werner 1989), indicating a stronger Tethyan link for the
Texas assemblage than for a north-south connection
through the Western Interior Seaway.
The mid Cretaceous distribution of Archaeolamna ex.
gr. kopingensis is strongly indicative of it as a cool water
taxon, being also present in northern Europe (e.g. Biddle
1993; Vullo et al. 2007) and Australia (Siverson 1996;
1997) but absent in Texas. It is therefore likely that a
marine connection between the northern Western Interior
Seaway and Boreal Ocean existed at this time, allowing
direct connection to cool waters elsewhere.
If wide ranging taxa are excluded, there are no species
in common between the Bainbridge River and mid
Cretaceous faunas of northern Europe (e.g. Muller and
Diedrich 1991; Underwood and Mitchell 1999), and com-
mon European taxa appear to be completely lacking in
the Western Interior Seaway. This may suggest that there
was no direct connection in the Cenomanian between the
northern Western interior Seaway and the Proto Atlantic
across what is now Hudson Bay.
Acknowledgements. Support for this research came from the
Canadian Museum of Nature (CMN) and a Discovery Grant to
SLC for student support from the Natural Sciences and
Engineering Research Council of Canada and University of
London Central Research Fund GLRP5 to CJU. The field research
was conducted under Saskatchewan Heritage Palaeontological
Resource Investigation Permit # 06-03P. Harold Bryant and Tim
Tokaryk of the Royal Saskatchewan Museum ensured that the
specimens were properly catalogued. Richard Day (CMN) and
MSc. student Aaron Phillips (Carleton University) provided field
and laboratory assistance, and CMN volunteer Dale Patten helped
sort through the residue from acid preparation of the bonebed.
David Ward greatly assisted with the photography and his
knowledge was invaluable for reconstruction of the lamniform
dentitions. Kenshu Shimada is thanked for his helpful discussions
on WIS sharks. Michal Ginter, Jurgen Kriwet and David Ward
are all thanked for their contributions and comments as reviewers
of the manuscript.
Editor: Marcello Ruta
REFERENCES
A G A S S I Z , L. J. R. 1833–1844. Recherches sur les poissons fos-
siles. Text (5 vols; I., xlix+188 pp., II xii+310 + 366 pp., III
viii+390 pp., IV xvi+296 pp., V xii+122 + 160 pp.) and Atlas
(5 vols; I 10 pl., II., 149 pl., III 83 pl., IV, 61 pl., V, 91 pl.).
Neuchatel.
A P PL E G A T E , S. P. 1972. A revision of the higher taxa of
Orectolobids. Journal of the Marine Biological Association of
India, 14, 743–751.
B E R G , L. S. 1940. Classification of fishes, both recent and fossil.
Transactions of the Institute of Zoology, Academy of Sciences
USSR, 5, 85–517. [In Russian].
940 P A L A E O N T O L O G Y , V O L U M E 5 3

—— 1958. System der Rezenten und Fossilen Fischartigen und
Fische. Hochschulbucher fur Biologie, Berlin, 310 pp.
B I D DL E , J.-P. 1993. Les Elasmobranches de l’Albien inferieur
et moyen (Cretace inferieur) de la Marne et de la Haute-
Marne (France) in Elasmobranches et Stratigraphie. Belgian
Geological Survey Professional Paper, 264, 191–240.
B UC K L A N D, W. 1838. On the discovery of fossil fishes in the
Bagshot Sands at Goldworth Hill, 4 miles North of Guildford.
Proceedings of the Geological Society of London, 2, 687–688.
C A DE N A T, J. 1963. Notes d’Ichtyologie ouest-africaine.
XXXIX. – Notes sur les requines de la famille des Carchariidae
et des formes apparentees de L’Atlantique ouest-africaine (avec
la description d’une espece nouvelle: Pseudocarcharias pelagi-
cus, classee dans un sous-genre nouveau). Bulletin de l’Institut
Francais d’Afrique Noire A, 25, 526–543.
C A P PE T T A , H. 1973. Selachians from the Carlile Shale
(Turonian) of South Dakota. Journal of Paleontology, 47, 504–
514.
—— 1977. Selaciens nouveaux de l’Albien superieur de Wissant
(Pas-de-Calais). Geobios, 10, 967–973.
—— 1987 Chondrichthyes II, Mesozoic and Cenozoic Elasmo-
branchii. Handbook of Paleoichthyology. Vol. 3B, Stuttgart, 193
pp.
—— 1980a. Modification du statut generique de quelques
especes de selachiens cretaces et tertiaires. Palaeovertebrata, 10,
29–42.
—— 1980b. Les selaciens du Cretace superieur du Liban 2: Bato-
ides. Palaeontographica, Abteilung A, 168, 149–229.
—— 2006. Elasmobranchii Post-Triadici (Index generum et
specierum). 295–396. In R E I G R A F , W. (ed.). Fossilium Ca-
talogus I: Animalia, Pars 142. Backhuys Publishers, Leiden, 472
pp.
—— and C A S E , G. R. 1975. Selaciens nouveaux du Cretace du
Texas. Geobios, 8, 303–307.
—— —— 1999. Additions aux faunes de selaciens du Cretace du
Texas (Albien superieur–Campanien). Palaeo Ichthyologica, 9,
1–112.
C A S E , G. R. 1978. A new selachian fauna from the Judith River
formation (Campanian) of Montana. Palaeontographica, Abtei-
lung A, 160, 176–205.
—— 1979. Cretaceous selachians from the Peedee Formation
(Late Maestrichtian) of Duplin County, North Carolina.
Brimleyana, 2, 77–89.
—— 2001. A new selachian fauna from the Coleraine Formation
(Upper Cretaceous ⁄ Cenomanian) of Minnesota. Paleaeonto-
graphica Beitrage zur Naturgeschichte der Vorzeit, 261, 103–
112.
—— and CA PP E T TA , H. 2004. Additions to the elasmo-
branch fauna from the Late Cretaceous of New Jersey (Lower
Navesink Formation, Early Maastrichtian). Palaeovertebrata,
33, 1–16.
—— T OK A R Y K , T. T. and BA I R D, D. 1990. Selacians from
the Niobrara Formation of the Upper Cretaceous (Coniacian)
of Carrot River, Saskatchewan, Canada. Canadian Journal of
Earth Sciences, 27, 1084–1094.
C A S I E R, E. 1947. Constitution et evolution de la racine
dentaire des Euselachii. Bulletin du Musee Royal d’Histoire
Naturelle de Belgique, 23, 1–45.
C I C I M UR R I , D. J. 2001a. Cretaceous elasmobranchs of the
Greenhorn Formation (Middle Cenomanian – Middle Turo-
nian), Western South Dakota. National Park Service, Proceed-
ings of the 6th Fossil Resource Conference. Geological Resources
Division Technical Report, 27–43.
—— 2001b. Fossil Selachians from the Belle Fourche Shale (Cre-
taceous, Cenomanian), Black Hills Region of South Dakota
and Wyoming. The Mountain Geologist, 38, 181–192.
—— 2004. Late Cretaceous Chondrichthyans from the Carlile
Shale (Middle Turonian to Early Coniacian) of the Black Hills
Region, South Dakota and Wyoming. The Mountain Geologist,
41, 1–16.
C OM PA GN O , L. J. V. 1977. Phyletic relationships of Living
Sharks and Rays. American Zoologist, 17, 303–322.
—— 2001. FAO Species Catalogue for Fishery Purposes No. 1.
Sharks of the World: An Annotated and Illustrated Catalogue
of Shark Species Known to Date, vol. 2: Bullhead, Mackerel
and Carpet Sharks (Heterodontiformes, Lamniformes and
Orectolobiformes). FAO, Rome, 269 pp.
C OO K , T. D., W I L S O N , M. V. H. and M UR R A Y , A. M.
2008. A middle Cenomanian euselachian assemblage from the
Dunvegan Formation of northwestern Alberta. Canadian Jour-
nal of Earth Sciences, 45, 1185–1197.
—— M UR R A Y , A., W I L S O N , M., T A K E UC H I , G. and
S H I M A DA , K. in press. A partial skeleton of the late Creta-
ceous lamniform shark, Archaeolamna kopingensis, from the
Pierre Shale of western Kansas. Journal of Vertebrate Paleon-
tology.
C OP E , E. D. 1874. Review of the Vertebrata of the Cretaceous
period found west of the Mississippi River. Section I. On the
mutual relationships of the Cretaceous and Tertiary forma-
tions of the west. Section II. List of species of Vertebrata from
the Cretaceous formations of the west. Bulletin of the U.S.
Geological and Geographical Survey of the Territories, 1, 2–48.
C UM BA A , S. L. and BR Y A N T , H. N.. 2001. Stratigraphic
position of a Late Cretaceous (Cenomanian) bonebed, east-
central Saskatchewan. In Summary of Investigations 2001,
Volume 1, Saskatchewan Geological Survey, Saskatchewan
Energy Mines, Miscellaneous Reports, 2001-4.1, 121–124.
—— and TO K A R Y K , T. T. 1999. Recent discoveries of
Cretaceous marine vertebrates on the eastern margins of the
Western Interior Seaway. Saskatchewan Geological Survey,
Summary of Investigations, 1999, 1, 57–63.
—— S C H R OD E R - A DA M S , C., DA Y , R. G. and PH I L -
L I P S , A. J. 2006. Cenomanian bonebed faunas from the
northeastern margin, Western Interior Seaway, Canada. New
Mexico Museum of Natural History and Science Bulletin, 35,
139–155.
D A L I N K E V I C I U S , J. A. 1935. On the fossil fishes of the
Lithuanian Chalk. I. Selachii. Memoires de la Faculte des
Sciences de l’ universite de Vytautas le Grand, 9, 247–305.
D A V I S , J. W. 1890. On the fossil fish of the Cretaceous Forma-
tion of Scandinavia. Scientific Transactions of the Royal Dublin
Society, Series 2, 4, 363–434.
E V E R H A R T , M. J. 2007. New stratigraphic records (Albian to
Campanian) of Rhinobatos sp. (Chondrichthyes, Rajiformes)
from the Cretaceous of Kansas. Transactions of the Kansas
Academy of Science, 110, 225–235.
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 941

G L I C K M A N , L. S. 1957. On the systematic significance of
accessory denticles in the teeth of sharks of the families Lam-
nidae and Scapanorhynchidae. Trudy Geologischeskogo Muzeja
‘A. P. Karpinskogo’, Akademia Nauk SSSR, 1, 103–109. [In
Russian].
—— 1958. Rates of evolution in lamnoid sharks. Doklady Aka-
demii Nauk Soyuza Sovetskikh Sotsialisticheskikh Respublik, 123,
568–571. [In Russian].
—— 1964. Sharks of the Palaeogene and their stratigraphic signifi-
cance [In Russian]. Doklady Akademii Nauk Soyuza Sov-
etskikh Sotsialisticheskikh Respublik, Moscow, 228 pp.
—— 1980. Evolution of Cretaceous and Cenozoic lamnoid sharks
[In Russian]. Doklady Akademii Nauk Soyuza Sovetskikh Sots-
ialisticheskikh Respublik, Moscow, 247 pp.
—— and A V E R I A N O V , A. O. 1998. Evolution of the Creta-
ceous Lamnoid sharks of the genus Eostriatolamia. Paleonto-
logical Journal, 32, 376–384.
—— and S H A V Z H A I T E , R. A. 1971. Sharks of the family
anacoracidae from the Cenomanian and Turonian of Lithua-
nia, Pre–Volga Region and Middle Asia. Paleontologia I Stra-
tigrafia Pribaltiki i Belorussii, 3, 185–192. [In Russian].
H A Y , O. P. 1902. Bibliography and catalogue of the fossil ver-
tebrata of North America. Bulletin of the United States Geologi-
cal Survey, 179, 1–868.
H E R M A N , J. 1977. Les selaciens neocretaces et paleocenes de
Belgique et des contrees limitrophes elements d’une biostratig-
raphie intercontinentale. Memoires pour servir a l’explication
des cartes geologiques et minieres de la Belgique, 15, 1–450.
—— H OV E S TA DT - E U LE R, M., H OV E S TA DT , D. C. and
S T E H M A N N , M. 2000. Contributions to the comparative
morphology of teeth and other relevant ichthyodorulites in
living supraspecific taxa of chondrichthyan fishes. Part B:
Batomorphii. No. 4c: Order Rajiformes – Suborder Mylio-
batoidei – Superfamily Dasyatoidea – Family Dasyatididae –
Subfamily Dasyatinae – Genus: Urobatis – Subfamily Plesioba-
tiodea – Family: Plesiobatidae – Genus: Plesiobatis – Super-
family Myliobatidea – Family: Myliobatidae – Subfamily:
Myliobatinae – Genera: Aetobatus, Aetomylaeus, Myliobatis
and Pteromylaeus – Subfamily: Rhinopterinae – Genus: Rhin-
optera – Subfamily: Mobulinae – Genera: Manta and Mobula.
Addendum 1 to 4a: Erratum to genus Pteroplatytrygon. Bulle-
tin de l¢Institut Royal des Sciences Naturelles de Belgique, Biolo-
gie, 70, 5–67.
H UM A N , B. A., O W E N , E. P., CO M PA G N O , L. J. V. and
HA RL E Y , E. H. 2006. Testing morphologically based phylo-
genetic theories within the cartilaginous fishes with molecular
data, with special reference to the catshark family (Chondrich-
thyes; Scyliorhinidae) and the interrelationships within them.
Molecular Phylogenetics and Evolution, 39, 384–391.
H UX L E Y , T. H. 1880. On the application of the laws of evolu-
tion to the arrangement of the Vertebrata, and more particu-
larly of the Mammalia. Proceedings of the Zoological Society of
London, 1880, 649–662.
J A E KE L , O. 1898. Uber Hybodus Agassiz. Sitzungsberichte der
Gesellschaft Naturforschenden Freunde, 89, 135–146.
K R I W E T , J. 2006. Biology and dental morphology of Priscusu-
rus adruptodontus sp. nov. (Chondrichthyes, Lamniformes)
from the Albian (Early Cretaceous) of Peru. Journal of Verte-
brate Paleontology, 26, 538–543.
—— K L U G, S., CA N U DO , J. I. and C U E N CA - B E S C O S ,
G. 2008. A new Early Cretaceous lamniform shark (Chon-
drichthyes, Neoselachii). Zoological Journal of the Linnean Soci-
ety, 154, 278–290.
L A N D E M A I N E , O. 1991. Selaciens nouveaux du Cretace sup-
erieur du sud-ouest de la France. Quelques rapports a la sys-
tematique des Elasmobranches. Societe Amicale des Geologues
Amateurs, 1, 1–45.
L E R I C H E , M. 1929. Les poissons du Cretace marin de la Bel-
gique et du Limbourg hollandais (note preliminaire). Les re-
sultats stratigraphiques de leur etude. Bulletin de la Societe
Belge de Geologie, de Paleontologie et d’Hydrologie, 37, 199–
299.
L I N C K , H. F. 1790. Versuch einer Eintheilung der Fische nach
den Zahnen. Magazin fur das Neueste aus der Physik und Na-
turgeschichte, 6, 28–38.
L I N N A E US , C. 1758. Systema naturae. Tenth edition. Salvi,
Stockholm, 824 pp.
M A I S E Y , J. G. 1987. Cranial anatomy of the Lower Jurassic
Shark Hybodus reticulatus (Chondrichthyes: Elasmobranchii),
with comments on Hybodontid Systematics. American
Museum Novitates, 2878, 1–39.
—— 1989. Hamiltonichthys mapsi, g. & sp. nov. (Chondrich-
thyes; Elasmobranchii), from the Upper Pennsylvanian of
Kansas. American Museum Novitates, 2931, 1–42.
M A UL , G. E. 1955. Five species of rare sharks new for Madeira
including two new to science. Notulae Naturae (Philadelphia),
279, 1–13.
M C N E I L , D. H. and C A L DW E L L, W. G. E. 1981. Creta-
ceous rocks and their Foraminifera in the Manitoba Escarp-
ment. The Geological Association of Canada Special Paper, 21,
1–439.
M U L L E R , A. and DI E DR I C H , C. 1991. Selachier (Pisces,
Chondrichthyes) aus der Cenomanium von Ascheloh am
Teutoburger Wald (Nordrhein-Westfalen, NW-Deutchland).
Geologie und Palaontologie in Westfalen, 20, 5–105.
M U L L E R , J. and H E N L E , J. 1838–41. Systematische beshrei-
bung der plagiostomen. Veit, Berlin, 200 pp.
N I CH O LL S , E. L. and R US S E LL , A. P. 1990. Paleobiogeog-
raphy of the Cretaceous Western Interior Seaway of North
America: the vertebrate evidence. Palaeogeography, Palaeo-
climatology, Palaeoecology, 79, 149–169.
N O UB H A N I , A. and C A P PE T T A , H. 1997. Les Orectolobi-
formes, Carchariniformes, et Myliobatiformes (Elasmobran-
chii, Neoselachii) des Bassins a phosphate du Maroc
(Maastrichtian-Lutetien basal). Palaeo Ichthyologica, 8, 1–327.
O W E N , R. 1846. Lectures on comparative anatomy and Physiol-
ogy of Vertebrate Animals. Part 1. Fishes, London, 51 pp.
P A T TE RS ON , C. 1965. The phylogeny of the Chimaeroids.
Philosophical Transactions of the Royal Society of London, Series
B, No. 757, 249, 101–219.
P HI L L I P S , A. 2008. A Late Cretaceous (Cenomanian) marine
vertebrate-rich bioclastic horizon from the northeastern margin
of the Western Interior Seaway, Canada. MSc. thesis in Earth
Sciences, Carleton University, Ottawa, 237 pp.
942 P A L A E O N T O L O G Y , V O L U M E 5 3

R A F I N E S QU E , C. S. 1810. Caratteri di alcuni nuovi generi e
nouvi spedie di animali e piante della Sicilia, con varie osserv-
azione sopra i medesimi. Palermo, 105 pp.
R E E S , J. and U N DE R W O O D, C. J. 2008. Hybodont sharks
of the English Bathonian and Callovian (Middle Jurassic).
Palaeontology, 57, 117–147.
R I S S O , A. 1810. Ichthyologie de Nice, ou histoire naturelle des
poissons du departement des Alpes Maritimes, Paris, 388 pp.
R U S S E L L , D. A. 1988. A check list of North American marine
Cretaceous vertebrates including fresh water fishes. Occasional
Paper of the Tyrrell Museum of Paleontology, 4, 1–58.
—— 1993. Vertebrates in the Cretaceous Western Interior Sea.
In CA L D W E L L , W. G. E. and K A UF F M A N , E. G. (eds).
Evolution of the Western Interior Basin. Geological Association
of Canada Special Paper, 39, 665–680.
S C H R OD E R - A DA M S , C. J., L E C K I E , D. A., CR A I G , J.
and B L OC H , J. 1999. Upper Cretaceous Colorado Group in
the Pasquia Hills, Northeastern Saskatchewan: A multidisci-
plinary study in progress. In Summary of Investigations 1999,
Volume 1, Saskatchewan Geological Survey, Saskatchewan
Energy Mines, Miscellaneous Reports, 1999-4.1, 52–56.
—— CU M B A A , S. L., B L O CH , J., L E CK I E , D. A., C R A I G ,
J., S E I F E L - DE I N , S. A., S I M ON S , D.-J. H. A. E. and
KE N I G , F. 2001. Late Cretaceous (Cenomanian to Campa-
nian) paleoenvironmental history of the Eastern Canadian
margin of the Western Interior Seaway: bonebeds and anoxic
events. Palaeogeography, Palaeoclimatology, Palaeoecology, 170,
261–289.
S C H W I M M E R , D. R., H O OK S , G. E. III and J OH N S O N ,
B. 2002. Revised taxonomy, age, and geographic range of the
large lamniform shark Cretodus semiplicatus. Journal of Verte-
brate Paleontology, 22, 704–707.
S H I M A D A , K. 1997. Dentition of the Late Cretaceous lamni-
form shark, Cretoxyrhina mantelli, from the Niobrara Chalk of
Kansas. Journal of Vertebrate Paleontology, 17, 269–279.
—— 2007. Skeletal and dental anatomy of the lamniform shark
Cretalamna appendiculata, from the Upper Cretaceous Niobr-
ara Chalk of Kansas. Journal of Vertebrate Paleontology, 27,
584–602.
—— 2008. New anacoracid shark from the Upper Cretaceous
Niobrara Chalk of western Kansas, U.S.A. Journal of Vertebrate
Paleontology, 28, 1189–1194.
—— S CH U M A C H E R , B. A., P A R KI N , J. A. and
PA LE RM O, J. M. 2006. Fossil marine vertebrates from the
lowermost Greenhorn Limestone (Upper Cretaceous: Middle
Cenomanian) in southwestern Colorado. Journal of Paleontol-
ogy Memoir, 63, 1–45.
S I V E R S O N , M. 1992. Biology, dental morphology and taxon-
omy of Lamniform sharks from the Campanian of the Kris-
tianstad Basin, Sweden. Palaeontology, 35, 519–554.
—— 1995. Revision of Cretorectolobus (Neoselachii) and descrip-
tion of Cederstroemia n. gen., a Cretaceous carpet shark
(Orectolobiformes) with a cutting dentition. Journal of Paleon-
tology, 69, 974–979.
—— 1996. Lamniform sharks of the mid)Cretaceous Alinga For-
mation and Beedagong Claystone, Western Australia. Palaeon-
tology, 39, 813–849.
—— 1997. Sharks from the mid)Cretaceous Gearle Siltstone,
Southern Carnarvon Basin, Western Australia. Journal of
Vertebrate Paleontology, 17, 453–465.
—— 1999. Lamniform sharks of the Mid Cretaceous Alinga For-
mation and Beedagong Claystone, Western Australia. Palaeon-
tology, 39, 813–849.
—— and L I N D G R E N , J. 2005. Late Cretaceous sharks Creto-
xyrhina and Cardabiodon from Montana, USA. Acta Palaeon-
tologica Polonica, 50, 301–314.
—— —— and K E L LY , L. S. 2007. Anacoracid sharks from the
Albian (Lower Cretaceous) Pawpaw shale of Texas. Palaeontol-
ogy, 50, 939–950.
S O K OL O V , M. 1965. Teeth evolution of some genera of
Cretaceous sharks and reconstruction of their dentition.
Moskovkoe Obshchestvo Ispytatelie Prirody, Biulleten. Otodel
Geologicheskii, 40, 133–134.
—— 1978. Requins comme fossils-guides pour la zonation et la
subdivision descouches cretacees de Tourousk. Niedra.
Moscou, 61, 1–60. [In Russian].
S T R O M E R , E. 1927. Ergebnisse der Forschungsreisen Prof. E.
Stromers in den Wusten Agyptens. II. Wirbeltier-Reste der
Baharije-Stufe (unterstes Cenoman). 9: Die Plagiostomen, mit
einem Anhang uber kano- und mesozoische Ruckenflossensta-
cheln von Elasmobranchiern. Abhandlungen der Bayerischen
Akademie der Wissenschaften Mathematisch-naturwissenschaftli-
che Abteilung, 31, 1–64.
T H UR M ON D , J. T. 1971. Cartilaginous fishes of the Trinity
Group and related rocks (Lower Cretaceous) of North Central
Texas. Southeastern Geology, 13, 207–227.
U N DE R W O O D, C. J. and M I T C HE L L , S. H. 1999. Albian
and Cenomanian selachian assemblages from North East Eng-
land. Special Papers in Palaeontology, 60, 9–59.
—— and W A R D, D. 2008. Sharks of the Order Carcharhinifor-
mes from the British Coniacian, Santonian and Campanian
(Upper Cretaceous). Palaeontology, 51, 509–536.
—— M I T C H E L L, S. H. and V E L T K A M P , C. J. 1999. Shark
and ray teeth from the Hauterivian (Lower Cretaceous) of
north-east England. Palaeontology, 42, 287–302.
V U L L O, R., CA P PE TT A , H. and N E R A U D E A U , D.
2007. New sharks and rays from the Cenomanian and Turo-
nian of Charentes, France. Acta Palaeontologica Polonica, 52,
99–116.
W E L T O N , B. J. and FA R I S H , R. F., 1993. The Collector’s
Guide to Fossil Sharks and Rays from the Cretaceous of Texas.
Before Time, Texas, 203 pp.
W E R N E R, C. 1989. Die Elasmobranchier-Fauna des Gebel Dist
Member der Bahariya Formation (Obercenoman) der Oase
Bahariya, Agypten. Paleo Ichthyologica, 5, 1–112.
W H I TL E Y , G. P. 1939. Taxonomic notes on sharks and rays.
The Australian Zoologist, 9, 227–262.
W I L L I A M S ON , T. E., KI R KL A N D , J. I. and LU C A S , S. G.
1993. Selachians from the Greenhorn Cyclothem (‘Middle’
Cretaceous: Cenomanian–Turonian), Black Mesa, Arizona,
and the paleogeographic distribution of Late Cretaceous sela-
chians. Journal of Paleontology, 67, 447–474.
W I L L I S T O N , S. W. 1900. Cretaceous fishes, selachians and
ptychodonts. University Geological Survey, Kansas, 6, 237–256.
U N D E R W O O D A N D C U M B A A : C A N A D I A N C R E T A C E O U S C H O N D R I C H T H Y A N S 943

W O O DW A R D , A. S. 1912. The fishes of the English chalk.
Part 7. Monograph of the Palaeontolographical Society, 65, 225–
264.
Z A N G E R L , R. 1979. New Chondrichthyes from the Mazon
Creek Fauna (Pennsylvanian) of Illinois. 449–500. In N I T E -
CK I , M. H. (ed.). Mazon Creek Fossils. Academy Press, New
York, 581 pp.
—— 1981. Chondrichthyes I – Paleozoic Elasmobranchii. Hand-
book of Paleoichthyology, Vol. 3A, Stuttgart, 115 pp.
NOTE ADDED IN PROOF
Recent description of an articulated skeleton and dentition of Archaeolamna (Cook et al. in press) has some implications
for the dentition of this genus. In most respects the articulated dentition agrees with the dental reconstruction as given
here. Despite this, the articulated specimen demonstrates that the second and third upper anterior teeth as interpreted
here are reversed, a situation superficially more similar to that in extant lamnids than Carcharias. The articulated speci-
men also shows no upper intermediate tooth or other teeth resembling those interpreted here as being in that position.
It is therefore probable that, despite the relative frequency of upper intermediate teeth in the assemblage described here,
some individuals or populations of Archaeolamna lacked teeth in this position.
944 P A L A E O N T O L O G Y , V O L U M E 5 3