new proteomic insights on the role of npr-a in regulating self-renewal of embryonic stem cells
TRANSCRIPT

New Proteomic Insights on the Role of NPR-A in RegulatingSelf-Renewal of Embryonic Stem Cells
SamehMagdeldin & Tadashi Yamamoto & Ikuo Tooyama &
Essam M. Abdelalim
# Springer Science+Business Media New York 2014
Abstract Embryonic stem cells (ESCs) have the ability toself-renew indefinitely and they can give unlimited source ofcells and tissues for cellular therapies. Recently, the natriureticpeptide receptor A (NPR-A) has been recognized as an im-portant regulator for the self-renewal of ESCs. To gain in-sights into possible novel mechanisms involved in NPR-Apathway that presumably regulates self-renewal and survivalof ESCs, we utilized a comprehensive label-free proteomicstechnology in our study. Targeting of NPR-A gene with smallinterfering RNA (siRNA) resulted in the inhibition of ESCsself-renewal. Coherently, quantitative label-free shotgun pro-teomic analysis identified differentially expressed proteinsinvolved in several biological processes, including cell cycleregulation, cell proliferation, cell fate specification, and apo-ptosis. Interestingly, in addition to Oct4 Nanog, and Sox2,
other proteins involved in ESCs self-renewal were down-regulated after NPR-A knockdown, such as heterogeneousnuclear ribonucleoprotein A2/B1 (ROA2), non-POUdomain-containing octamer-binding protein (Nono),nucleoplasmin (Npm1), histone H2A type 1-B/E (histoneH2A.2), SW1/SNF complex (Brg1), polycomb proteinSuz12 (Suz12), and cyclin-dependent kinase 4 (Cdk4). Fur-thermore, several protein candidates involved in early differ-entiation and cell death were up-regulated or down-regulatedas a result of NPR-A knockdown, including importin subunitalpha-4 (Impα4), importin-5 (Ipo5), H3 histones, core histonemacro-H2A.1 (H2A.y), apurine/apyrimidine endonuclease 1(Apex1), 78-kDa glucose-regulated protein (Grp78), and pro-grammed cell death 5 (Pdcd5). Overall, these findings depict acomprehensive view to our understanding of the pathwaysinvolved in the role of NPR-A in maintaining ESC functions.
Keywords ESCs . Proteome . Knockdown . Natriureticpeptide receptor A . Pluripotency . Differentiation . Label freequantification
Introduction
Embryonic stem cells (ESCs) are pluripotent cells, having thecapacity for unlimited self-renewal and differentiation into thethree germ layers: ectoderm, mesoderm, and endoderm. Theirability to differentiate into all cell types under appropriateconditions make them a promising tool for cell replacementtherapy [1]. ESCs maintain their unique characteristics byexpressing several intrinsic factors involved in maintainingself-renewal and pluripotency [2–5], which promote genesessential for self-renewal and pluripotency, and inhibits thosegenes which will be enhanced after ESC differentiation [3, 4].The core transcriptional factors Oct4, Sox2, and Nanog arerequired for maintaining self-renewal and pluripotency of
Electronic supplementary material The online version of this article(doi:10.1007/s12015-014-9517-0) contains supplementary material,which is available to authorized users.
S. Magdeldin : T. YamamotoDepartment of Structural Pathology Institute of Nephrology,Graduate School ofMedical and Dental Sciences, Niigata University,Tokyo, Japan
S. MagdeldinDepartment of Physiology, Faculty of Veterinary Medicine, SuezCanal University, Ismailia, Egypt
I. Tooyama : E. M. AbdelalimMolecular Neuroscience Research Center, Shiga University ofMedical Science, Setatsukinowa-cho, Otsu, Shiga 520-2192, Japan
E. M. Abdelalim (*)Qatar Biomedical Research Institute, Qatar Foundation, EducationCity, Doha 5825, Qatare-mail: [email protected]
E. M. AbdelalimDepartment of Cytology and Histology, Faculty of VeterinaryMedicine, Suez Canal University, Ismailia, Egypt
Stem Cell Rev and RepDOI 10.1007/s12015-014-9517-0

ESCs by regulating the expression of other genes as well asthemselves [6].
Recently, we have showed that natriuretic peptide receptorA (NPR-A) and its ligands atrial and brain natriuretic peptides(ANP and BNP), respectively [7], are expressed in pre-implantation embryos and in undifferentiated murine ESCs[8, 9]. Both ANP and BNP have been involved in severalfunctions during embryonic and adult stages of development[7, 10–12]. NPR-A has been recently found to be crucial forearly embryonic development [13]. Importantly, our previousstudy suggested that NPR-A might be implicated in maintain-ing several murine ESC functions related to self-renewal [8,14]. However, the molecular mechanisms involved in theNPR-A signalling pathways in ESCs are largely unknown.
Although several proteomic analysis have been performedto identify protein candidates that are possibly implicated indifferent biological functions in both murine ESCs [15, 16]and human ESCs [17, 18], there is a noted paucity of infor-mation regarding how NPR-A might affect ESCs proteome toachieve its peculiar role in self-renewal and pluripotency. Inaddition, it is unclear if this action is Per se or there might beother protein molecules regulated by NPR-A and implicatedin this prominent effect on ESCs. To this end, we sought toutilize the thorough coverage of the “shotgun” proteomicstechnology to access biologically relevant proteins modulatedfollowing NPR-A knockdown, as an endeavour to unravelNPR-A role in self-renewal.
Materials and Methods
Embryonic Stem Cell Culturing Condition
Murine ESCs (E14TG2a) (CRL-1821; American TypeCulture Collection, Manassas, VA) were maintained inDMEM/F-12 medium (Sigma) that was supplemented with1,000 U/mL Leukemia inhibitory factor (LIF) (Chemicon),11 % fetal bovine serum, 2 mM L-glutamine (Nacalai Tesque,Japan), 1 mM sodium pyruvate (Sigma), 1 % MEM nones-sential amino acids (GIBCO), 0.1 mM 2-mercaptoethanol(Sigma) and 1% penicillin-streptomycin. For small interferingRNA (siRNA) transfection, ESCs were cultured in the samemedium, except that fetal bovine serum was replaced with15 % Knockout Serum Replacement (KSR; GIBCO). MurineESCs were cultured under feeder-free conditions in the pres-ence of LIF.
RNA Interference (RNAi)
RNA interference in murine ESCs was carried out as previ-ously reported [8], and according to the manufacturer’s pro-tocol using Lipofectamine RNAiMAX (Invitrogen) in 6-wellplates. Two pairs of siRNAs (Invitrogen) were designed for
NPR-A (NM_008727) using the BLOCK-iT RNAi Designersoftware (http://rnaidesigner.lifetechnologies.com/rnaiexpress/). The appropriate siRNA negative controlDuplex (Cat. No. 12,935–300; Invitrogen) was selectedbased on the percentageG/C. Both NPR-A and control siRNAwere transfected at a final concentration of 40 μM for 24 h intriplicate for each treatment. Forty eight hours post-transfection, NPR-A knockdown procedure was confirmedby RT-PCR and western blotting. The sequences of theNPR-A siRNAs were as follows: NPR-A siRNA1—sense,5’-AGAUGUAGAUAACUCUGCCCUUGCG-3’; anti-sense, 5’-CGCAAGGGCAGAGUUAUCUAUACAUCU-3’; NPR-A siRNA2—sense, 5’-GCCUUCAGGGUUGUCCUGAACUUUA-3’; antisense, 5’-UAAAGUUCAGGACAACCCUGAAGGC-3’.
Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
Total RNAwas extracted using the RNeasy Mini kit (Qiagen)according to the manufacturer’s instructions. cDNA synthesiswas performed with 1 μg of total RNA using the SuperscriptIII first-strand cDNA synthesis kit (Invitrogen). The cDNA(0.5 μg) was used as a template in a mixture that containedAmpli Taq Gold polymerase (Applied Biosystems). Reversetranscription (RT)-polymerase chain reaction (PCR) was per-formed with an annealing temperature of 58 °C. The primersequences (sense and antisense) were as follows: NPR-A;sense: AGTACGCCAACAACCTGGAG, antisense: AAGAGCTGTAAAGCCCACGA. GAPDH, sense: GGCATTGTGGAAGGGCTCA, antisense: TCCACCACCCTGTTGCTGT.
Alkaline Phosphatase (AP) Activity Assay
To determine the ability of ESCs to maintain an undifferenti-ated phenotype, alkaline phosphatase (AP) activity assayswere performed using an AP kit (Chemicon) as previouslydescribed [9].
Protein Extraction and Cell Solubilization for Proteomics
Frozen aliquots (1.2×106) of murine ESCs were rinsed withcold PBS and centrifuged at 2,500 g for 5 min at 4 °C to pelletthe cells. After decanting the supernatant, 100 μl of urea lysisbuffer [8 M Urea, 500 mM Tris–HCl, pH 8.5, cOmplete ultramini protease inhibitors, Roche] weremixedwith cells, passedthrough needle (21Gx1.5) and briefly sonicated three times inice (output control three) for 5 s/each and incubated for 20minwith agitation in room temperature. Efficiency of homogeni-zation was confirmed using phase contrast microscopy (Sup-plemental Fig. 1). Protein lysate was centrifuged at 14.000 gfor 30 min to pellet the cell debris. Protein was assayed usingBCA method (Pierce, Rockford IL) at 562nm.
Stem Cell Rev and Rep

Filter-Aided Sample Preparation (FASP)
With minor modification of the original protocol, aliquots oflysate corresponding to 0.12 mg protein in 8 M urea, 0.5 MTris–HCl, pH 8.5 was processed in Vivacon 500 device (Sar-torius Stedim biotech) supplied with a hydrosart nitrate filtermembrane with molecular weight cutoff (MWCO) value 10KDa (FASP reactor) [19]. The device was centrifuged at14.000 g at 20 °C for 40 min. Subsequently, protein sampleon the FASP reactor was reduced for 20 min by addition of1.1 μl 1 M TCEP in 8 M urea, 0.1 M Tris–HCl, pH 8.5.Alkylation was done on the reactor by addition of 2.2 ul of0.5 M iodoacetamide in dark place. After brief centrifugation,the concentrate was diluted twice with 100 μl 8 M urea in0.1 M Tris–HCl, pH 8.0 and concentrated again. Resultedprotein concentrate on the FASP reactor was subjected toon-membrane endoprotease digestion by addition of Lys-C(enzyme to protein ratio; 1:25). The device was incubated inwet chamber overnight. Trypsin solution was added at a ratio(1:75) with 100 mM CaCl2 and incubated for additional 4 h.Recovery of peptides was achieved by centrifugation of thereactor in a new collection tube at 14.000 g for 40 min.Proteolytic activity were inhibited by acidifying the peptidemixture to pH 3 using trifluoroacetic acid.
Peptide Desalting with C18 StageTips (Stop-and-GoExtraction Tips)
For allowing high Lc-Ms/Ms reproducibility, peptide mixturewas cleaned up, and concentrated using reversed-phaseStageTips [20]. A homemade C18 disk with chromatographicbeads immobilized in a Teflon meshwork (Empore, 3 M) wasstacked in 200 μl low adsorption tip. To condition C18 carti-lage, 20 μl methanol was added and spin down at 1,500 g/1 min. The cartilage was rinsed with 20 μl 0.2 %trifluoroacetic acid (TFA), 80 % acetonitrile (ACN) mixtureand centrifuged at 1,500 g/1 min. Later on, 20μl of 0.2%TFAwas applied on the disk and spin down again at 1,500 g/1 min.Twenty μg of peptide mixture was placed on the C18 andcentrifuged at 150 g/3 min to trap peptides on the cartilage.For translation (transfer-elution), C18 cartilage was washedwith 20 μl of 0.2 % TFA and transferred to a new collectiontubes. Peptide mixture was collected by rinsing the cartilagewith 80 % acetonitrile (10 μl×3 times) where the collectedelute contains desalted peptides ready for mass spectrometricanalysis.
Nano-LC-MS/MS Analysis
Capillary columns were prepared in house for LC-MS/MSanalysis from particle slurries in methanol. An analyticalRPLC column was generated by pulling a 100 μm-ID/360 μm-OD capillary (Polymicro Technologies) to 5 μm-ID
tip. Reverse-phase particles (Aqua C18, 5 μm diameter, 90-Åpores, Phenomenex) were packed directly into the pulledcolumn at 800 p.s.i. until 12 cm long. The column was furtherequilibrated at 100 bar with buffer B followed by buffer A.Trapping column was prepared by creating a Kasil frit at oneend of an undeactivated 250 μm-ID/360 μm-OD capillary(Agilent Technologies) and then was successively packedwith 2.5 cm Aqua C18, 3 μm diameter, 125-Å pores,Phenomenex). The Kasil frit was prepared by briefly dippinga 15 cm capillary in well-mixed 300 μL Kasil 1,624 (PQCorporation) and 100 μL formamide, baked at 100 °C over-night, and cutting the frit to ~1.5 mm in length. The columnwas equilibrated using buffer A for10 min. Both trapping andanalytical columns were assembled using a zero dead-volumeunion (Upchurch Scientific). LC-MS/MS analysis was per-formed using Easy-nLC pump and Thermo Q Exactive usingan in-house-built electrospray stage. Electrospray was per-formed directly from the analytical column by applying theESI voltage at a tee (150μm-ID, Upchurch Scientific) directlydownstream of a 1:1,000 split flow used to reduce the flowrate to 250 nL/min through the columns. Replicates wereanalyzed for 2 h (120 min) gradient from 5 to 45 % buffer Bover 155 min, up to 100 % B over 50 min, held at 100 % B for5 min, then back to 100 % A for column re-equilibration.HPLC buffers (Honeywell) were 5 % acetonitrile with 0.1 %formic acid (A), and 80 % acetonitrile with 0.1 % formic acid(B). Precursor scanning in the Q Exactive was performed from400 to 1,600 m/z with the following settings, respectively: 1×106 target ions, 20 m maximum ion injection time and onemicroscan. Data-dependent acquisition of MS/MS spectrawas performed with the following settings: high-energycollision-induced dissociation (HCD) on the ten most intenseions per precursor scan, 30,000 automatic gain-control targetions, 100 m maximum injection time, 30 % normalized colli-sion energy and one microscan. Dynamic-exclusion settingsused were as follows: repeat count, 1; repeat duration, 40 s;exclusion list size, 500; and exclusion duration, 60 s.
Bioinformatic Data Analysis
Protein and peptide identification and comparison were donewith Integrated Proteomics Pipeline (IP2, http://integratedproteomics.com/). Tandem mass spectra wereextracted to MS1 and MS2 files from raw files usingRawExtract 1.9.9. MS/MS spectra were searched againstUniProtKB consisting of SwissProtKB/TrEMBL with re-versed mouse sequence databases using ProLuCID [19]. Anup to date database were downloaded at ftp://ftp.uniprot.org/pub/databases/uniprot/current_release/knowledgebase/complete/. The spectral search space included all fully andhalf tryptic peptide candidates within a 50-p.p.m. windowsurrounding the peptide candidate precursor mass. Carba-midomethylation (+57.02146) of cysteine was considered a
Stem Cell Rev and Rep

static modification and oxidation (+15.995) of both methio-nine and tryptophan and acetylation (+42.011) of lysine as avariable modification. Peptide candidates were filtered to 0.1 % FDR and proteins candidates to 1 % FDR usingDTASelect19 [21]. Chromatographic peak areas were extract-ed with Census [22]. Protein physicochemical properties werecalculated using IP2 pipeline. To generate a functionallygrouped gene ontology and pathway network of NPR-Aknockdown ESCs proteome, ClueGo (v 2.0.6) and CluePedia(v 1.0.7) plug-ins of Cytoscape (v 3.0.2) were setup to twosided hypergeometric statistical testing with significance level(P<0.05). False discovery rate (FDR p<0.05) correction wascalculated using Benjamini and Hochberg multiple testingcorrection. A customized and updated GO slim file (OBOv1.2; 32,150 term) and gene annotation file were downloadedfrom GO consortium and Kyoto encyclopedia of genes andgenomes (KEGG) pathway databases, respectively and usedin the current analysis [23].
Western Blotting
Total protein extracts were prepared from ESCs. The celllysates were mixed with a loading buffer containing 2 %SDS, electrophoresed by SDS-PAGE and transferred ontonitrocellulose membranes (Amersham Biosciences, Freiburg,Germany). Proteins were detected using antibodies againstNPR-A (1:1,000, sc-31,632; Santa Cruz Biotechnology),Oct4 (1:1,000, sc-5,279; Santa Cruz Biotechnology), Nanog(1:8,000, A300-397A; Bethyl Laboratories), cyclin D1 (1:1,000, sc-450; Santa Cruz Biotechnology) and β-actin(1:8,000, sc-47,778; Santa Cruz Biotechnology). The second-ary antibodies were peroxidase-conjugated anti-rabbit IgG,peroxidase-conjugated anti-goat IgG, or peroxidase-conjugated anti-mouse IgG (all 1:10,000; JacksonImmunoResearch Lab, Inc). The blots were developed usingSuperSignal West Pico Chemiluminescent substrate (Pierce),and visualized using an LAS-3,000 FujiFilm Lumino-ImageAnalyzer (FujiFilm, Tokyo, Japan).
Apoptosis Assay
Annexin V staining was performed as previously described[24] using flow cytometry and according to the manufac-turer’s guidelines (MBL, USA)
Biostatistical Analysis
In order to properly access differentially significant proteinsbetween control and NPR-A knockdown ESC proteomes,two-sided Student-T was carried out among replicates (n=6)between both groups. P<0.05 was considered statisticallysignificant threshold. Normalized spectral abundance factor,protein expression alteration (fold changes), log values and
confidence were calculated based on spectral peak intensitiesgenerated from the mass spectrometric analysis afterextracting confident protein spectra with P<0.01. Graph padprism 5 and Microsoft spreadsheets formulas were used instatistical evaluation.
Results
Efficiency of NPR-AKnockdown inMurine Embryonic StemCells
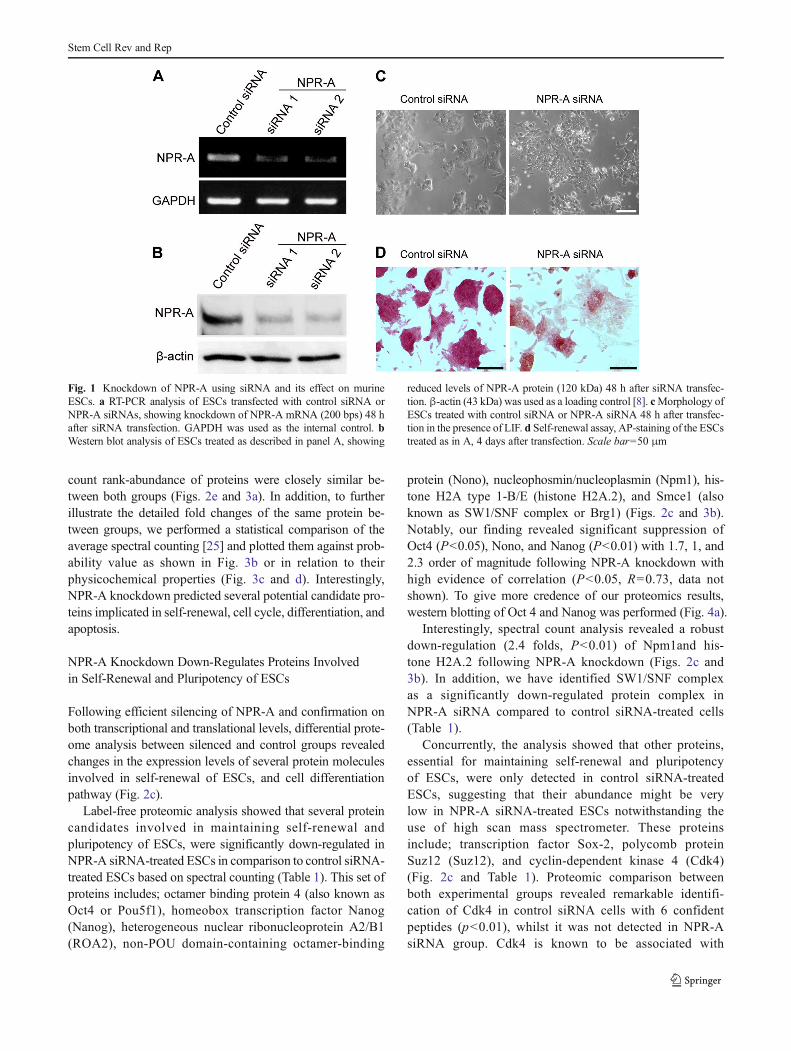
To further expand our knowledge on the role of NPR-A inESCs, we silenced NPR-A in undifferentiated murine ESCs inorder to depict a thorough and comprehensive understandingof the proteome changes provoked by genetic inhibition ofNPR-A. We used a siRNA-based technique to specificallyknockdown NPR-A in murine ESCs. The knockdown effi-ciency was examined 48 h after transfection of the siRNA.Following silencing, RT-PCR and western blotting revealedunambiguous reduction of NPR-A mRNA and protein levelsin the ESCs transfected with NPR-A-targeting siRNA (NPR-A siRNA1 and NPR-A siRNA2), compared with thosetransfected with a non-targeting siRNA (control siRNA) asshown in Fig. 1a and b. Furthermore, we observed that NPR-A knocking down led to changes in the morphology of ESCscolonies, where it became more flattened with differentiatedcharacteristics (Fig. 1c), as previously confirmed [8]. In addi-tion, alkaline phosphatase (AP) staining, a marker for undif-ferentiated ESCs, was less intense in cells underwent NPR-Aknockdown, indicating ESC differentiation (Fig. 1d).
Proteome Metrics of Embryonic Stem Cells Treatedwith siRNAs
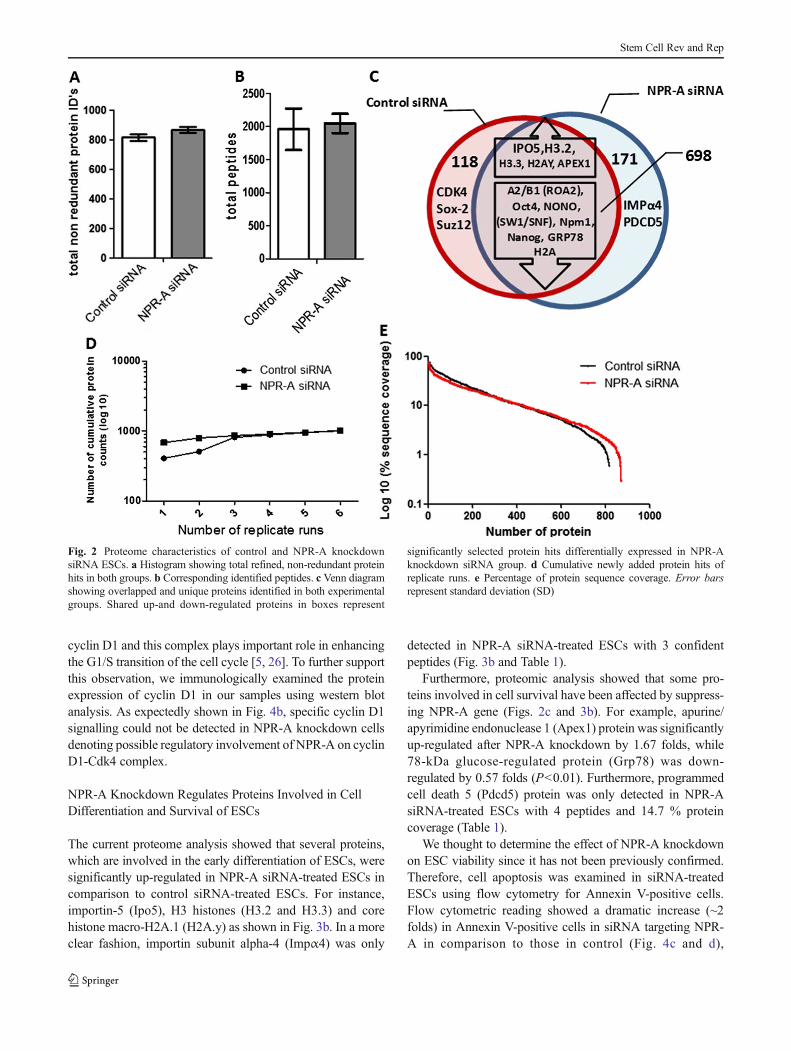
Comprehensive speculation on the proteome metrics betweencontrol siRNA- and NPR-A siRNA-treated ESCs was exten-sively examined in the current study. This experiment wasrepeated twice with high reproducibility (SupplementalFig. 1). As shown in Fig. 2, merged, non-redundant replicateanalysis of both control siRNA and NPR-A siRNA groupsshowed comparable protein identifications of 816 and 869,respectively. Detailed list of identified proteins in both exper-imental groups can be found in Supplemental Table 1. Similarnumber of peptides close to 2×103 was identified in eachgroup (Fig. 2b). Overlapping between identified proteinsshowed over 80 % shared proteins between experimentalgroups with unique, up-regulated and down-regulated proteincandidates (Fig. 2c) (as discussed below). The maximumidentification capabilities for both groups were attained andshowed by the plateau curve of cumulative newly addedprotein hits of replicates (Fig. 2d). In general perspective,the dynamic range of protein sequence coverage and spectral
Stem Cell Rev and Rep
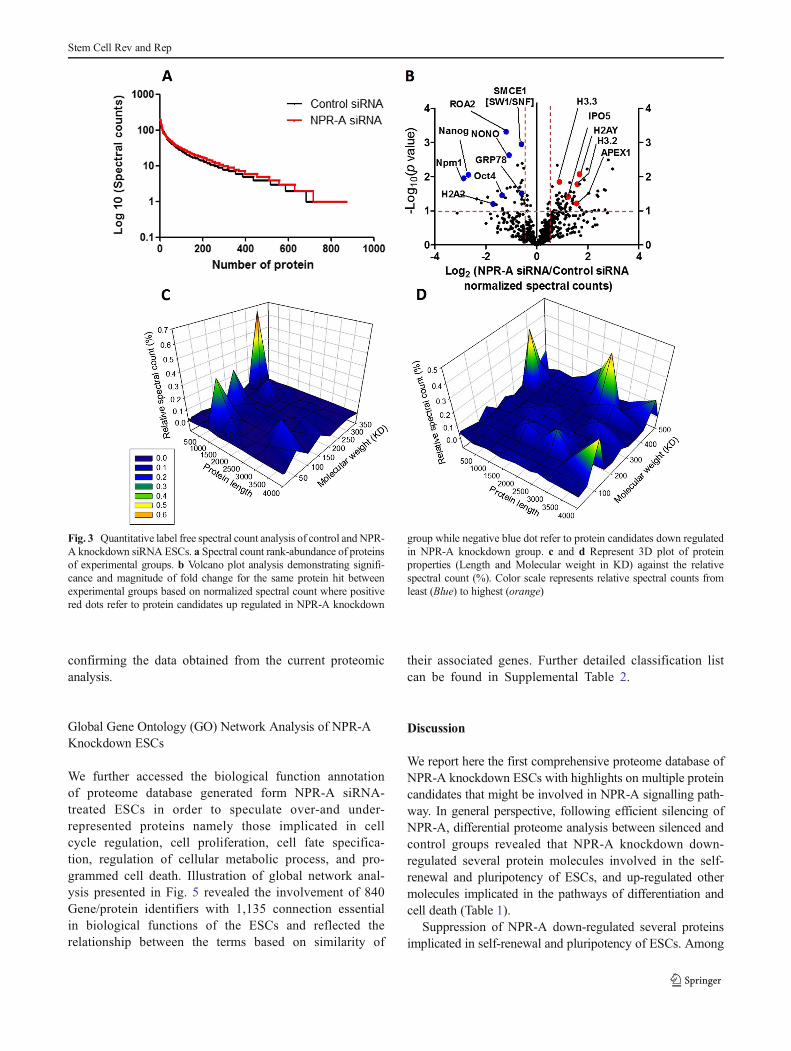
count rank-abundance of proteins were closely similar be-tween both groups (Figs. 2e and 3a). In addition, to furtherillustrate the detailed fold changes of the same protein be-tween groups, we performed a statistical comparison of theaverage spectral counting [25] and plotted them against prob-ability value as shown in Fig. 3b or in relation to theirphysicochemical properties (Fig. 3c and d). Interestingly,NPR-A knockdown predicted several potential candidate pro-teins implicated in self-renewal, cell cycle, differentiation, andapoptosis.
NPR-A Knockdown Down-Regulates Proteins Involvedin Self-Renewal and Pluripotency of ESCs
Following efficient silencing of NPR-A and confirmation onboth transcriptional and translational levels, differential prote-ome analysis between silenced and control groups revealedchanges in the expression levels of several protein moleculesinvolved in self-renewal of ESCs, and cell differentiationpathway (Fig. 2c).
Label-free proteomic analysis showed that several proteincandidates involved in maintaining self-renewal andpluripotency of ESCs, were significantly down-regulated inNPR-A siRNA-treated ESCs in comparison to control siRNA-treated ESCs based on spectral counting (Table 1). This set ofproteins includes; octamer binding protein 4 (also known asOct4 or Pou5f1), homeobox transcription factor Nanog(Nanog), heterogeneous nuclear ribonucleoprotein A2/B1(ROA2), non-POU domain-containing octamer-binding
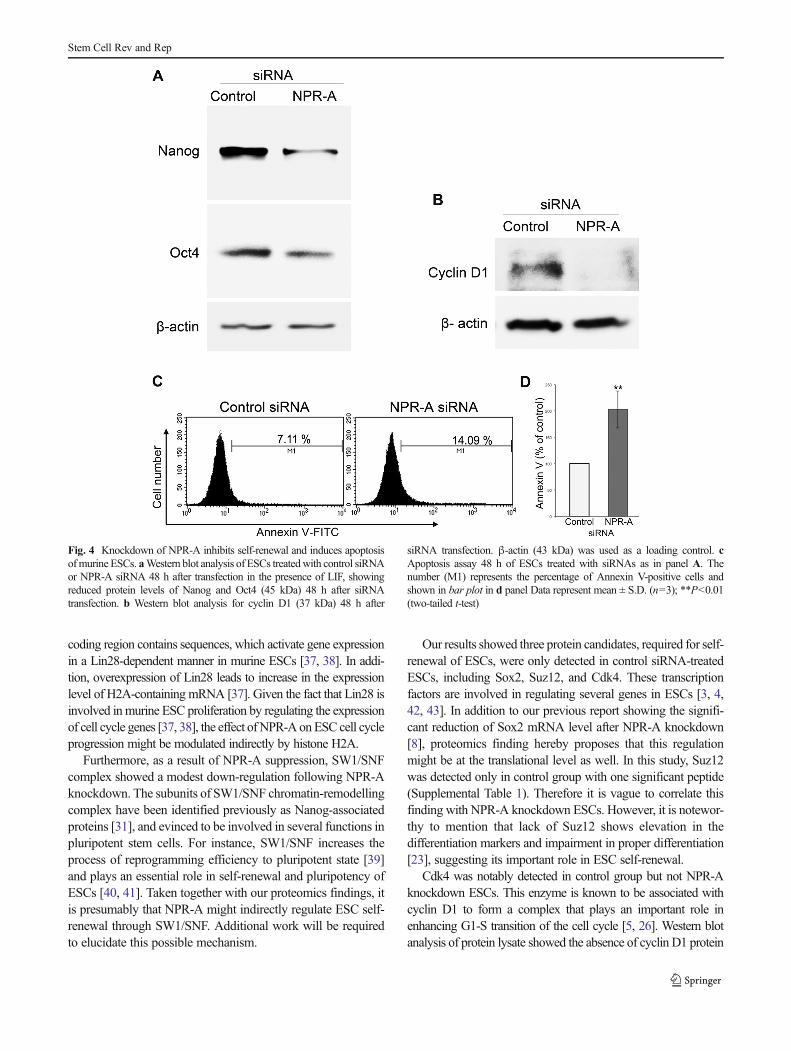
protein (Nono), nucleophosmin/nucleoplasmin (Npm1), his-tone H2A type 1-B/E (histone H2A.2), and Smce1 (alsoknown as SW1/SNF complex or Brg1) (Figs. 2c and 3b).Notably, our finding revealed significant suppression ofOct4 (P<0.05), Nono, and Nanog (P<0.01) with 1.7, 1, and2.3 order of magnitude following NPR-A knockdown withhigh evidence of correlation (P<0.05, R=0.73, data notshown). To give more credence of our proteomics results,western blotting of Oct 4 and Nanog was performed (Fig. 4a).
Interestingly, spectral count analysis revealed a robustdown-regulation (2.4 folds, P<0.01) of Npm1and his-tone H2A.2 following NPR-A knockdown (Figs. 2c and3b). In addition, we have identified SW1/SNF complexas a significantly down-regulated protein complex inNPR-A siRNA compared to control siRNA-treated cells(Table 1).
Concurrently, the analysis showed that other proteins,essential for maintaining self-renewal and pluripotencyof ESCs, were only detected in control siRNA-treatedESCs, suggesting that their abundance might be verylow in NPR-A siRNA-treated ESCs notwithstanding theuse of high scan mass spectrometer. These proteinsinclude; transcription factor Sox-2, polycomb proteinSuz12 (Suz12), and cyclin-dependent kinase 4 (Cdk4)(Fig. 2c and Table 1). Proteomic comparison betweenboth experimental groups revealed remarkable identifi-cation of Cdk4 in control siRNA cells with 6 confidentpeptides (p<0.01), whilst it was not detected in NPR-AsiRNA group. Cdk4 is known to be associated with
Fig. 1 Knockdown of NPR-A using siRNA and its effect on murineESCs. a RT-PCR analysis of ESCs transfected with control siRNA orNPR-A siRNAs, showing knockdown of NPR-A mRNA (200 bps) 48 hafter siRNA transfection. GAPDH was used as the internal control. bWestern blot analysis of ESCs treated as described in panel A, showing
reduced levels of NPR-A protein (120 kDa) 48 h after siRNA transfec-tion. β-actin (43 kDa) was used as a loading control [8]. cMorphology ofESCs treated with control siRNA or NPR-A siRNA 48 h after transfec-tion in the presence of LIF. d Self-renewal assay, AP-staining of the ESCstreated as in A, 4 days after transfection. Scale bar=50 μm
Stem Cell Rev and Rep
cyclin D1 and this complex plays important role in enhancingthe G1/S transition of the cell cycle [5, 26]. To further supportthis observation, we immunologically examined the proteinexpression of cyclin D1 in our samples using western blotanalysis. As expectedly shown in Fig. 4b, specific cyclin D1signalling could not be detected in NPR-A knockdown cellsdenoting possible regulatory involvement of NPR-A on cyclinD1-Cdk4 complex.
NPR-A Knockdown Regulates Proteins Involved in CellDifferentiation and Survival of ESCs
The current proteome analysis showed that several proteins,which are involved in the early differentiation of ESCs, weresignificantly up-regulated in NPR-A siRNA-treated ESCs incomparison to control siRNA-treated ESCs. For instance,importin-5 (Ipo5), H3 histones (H3.2 and H3.3) and corehistone macro-H2A.1 (H2A.y) as shown in Fig. 3b. In a moreclear fashion, importin subunit alpha-4 (Impα4) was only
detected in NPR-A siRNA-treated ESCs with 3 confidentpeptides (Fig. 3b and Table 1).
Furthermore, proteomic analysis showed that some pro-teins involved in cell survival have been affected by suppress-ing NPR-A gene (Figs. 2c and 3b). For example, apurine/apyrimidine endonuclease 1 (Apex1) protein was significantlyup-regulated after NPR-A knockdown by 1.67 folds, while78-kDa glucose-regulated protein (Grp78) was down-regulated by 0.57 folds (P<0.01). Furthermore, programmedcell death 5 (Pdcd5) protein was only detected in NPR-AsiRNA-treated ESCs with 4 peptides and 14.7 % proteincoverage (Table 1).
We thought to determine the effect of NPR-A knockdownon ESC viability since it has not been previously confirmed.Therefore, cell apoptosis was examined in siRNA-treatedESCs using flow cytometry for Annexin V-positive cells.Flow cytometric reading showed a dramatic increase (~2folds) in Annexin V-positive cells in siRNA targeting NPR-A in comparison to those in control (Fig. 4c and d),
Fig. 2 Proteome characteristics of control and NPR-A knockdownsiRNA ESCs. a Histogram showing total refined, non-redundant proteinhits in both groups. b Corresponding identified peptides. c Venn diagramshowing overlapped and unique proteins identified in both experimentalgroups. Shared up-and down-regulated proteins in boxes represent
significantly selected protein hits differentially expressed in NPR-Aknockdown siRNA group. d Cumulative newly added protein hits ofreplicate runs. e Percentage of protein sequence coverage. Error barsrepresent standard deviation (SD)
Stem Cell Rev and Rep
confirming the data obtained from the current proteomicanalysis.
Global Gene Ontology (GO) Network Analysis of NPR-AKnockdown ESCs
We further accessed the biological function annotationof proteome database generated form NPR-A siRNA-treated ESCs in order to speculate over-and under-represented proteins namely those implicated in cellcycle regulation, cell proliferation, cell fate specifica-tion, regulation of cellular metabolic process, and pro-grammed cell death. Illustration of global network anal-ysis presented in Fig. 5 revealed the involvement of 840Gene/protein identifiers with 1,135 connection essentialin biological functions of the ESCs and reflected therelationship between the terms based on similarity of
their associated genes. Further detailed classification listcan be found in Supplemental Table 2.
Discussion
We report here the first comprehensive proteome database ofNPR-A knockdown ESCs with highlights on multiple proteincandidates that might be involved in NPR-A signalling path-way. In general perspective, following efficient silencing ofNPR-A, differential proteome analysis between silenced andcontrol groups revealed that NPR-A knockdown down-regulated several protein molecules involved in the self-renewal and pluripotency of ESCs, and up-regulated othermolecules implicated in the pathways of differentiation andcell death (Table 1).
Suppression of NPR-A down-regulated several proteinsimplicated in self-renewal and pluripotency of ESCs. Among
Fig. 3 Quantitative label free spectral count analysis of control and NPR-A knockdown siRNA ESCs. a Spectral count rank-abundance of proteinsof experimental groups. b Volcano plot analysis demonstrating signifi-cance and magnitude of fold change for the same protein hit betweenexperimental groups based on normalized spectral count where positivered dots refer to protein candidates up regulated in NPR-A knockdown
group while negative blue dot refer to protein candidates down regulatedin NPR-A knockdown group. c and d Represent 3D plot of proteinproperties (Length and Molecular weight in KD) against the relativespectral count (%). Color scale represents relative spectral counts fromleast (Blue) to highest (orange)
Stem Cell Rev and Rep
those proteins, ROA2, which decreased ~1 fold followingNPR-A knockdownwith 42.5% sequence coverage (Table 2).This ribonucleoprotein showed high expression in undifferen-tiated ESCs [27–29]. Knockdown of ROA2 in human ESCsled to induction of differentiation, suppression of proliferationand cell cycle arrest in the G0/G1 phase [29]. Similar pheno-types have been reported in murine ESCs deficient in NPR-A[8], Taken together with our data, it is relevant that suppress-ing NPR-A level in ESCs is accompanied with reduction inthe expression of ROA2. Our finding showed significantsuppression of Oct4, Nanog, and Nono following NPR-Aknockdown. This could be explained biologically by the factthat Nono protein binds to the CpG island of Oct4 promoter,and positively regulates Oct4 gene expression in murineESCs. A previous report demonstrated that knockdown ofNono expression using siRNA led to Oct4 down-regulation[30]. Furthermore, it is well known that Oct4 and Nanog areessential for maintaining the pluripotency and their loss leadsto differentiation of ESCs [31, 32]. Our western blot analysis
confirmed that Oct4 and Nanog were significantly down-regulated, and supported the fidelity and accuracy of ourproteomic data.
Mass spectrometric quantification detected Npm1 proteinin a significant low level (P<0.01) compared to control coun-terpart although this protein is generally expressed at highlevels in murine [33] and human [34] ESCs. It is believed tobe crucial for ESC self-renewal since Npm1 knockdowninhibits ESC proliferation [35]. Several line of evidence in-criminated Npm1 in the interaction with all the three coretranscription factors (Oct4, Nanog, and Sox2) in ESCs, andits inhibition induces ESC differentiation into ectodermal andmesodermal lineages [36]. These findings suggest that NPR-Amight regulate ESC self-renewal through its effect on Npm1.Further investigation is required to confirm this possible cor-relation. Additionally, histone H2A.2 was significantly down-regulated in NPR-A knockdown group. Recent evidence dem-onstrated that production of histones, particularly H2A sub-type, is regulated by Lin28 inmurine ESCs [37]. Histone H2A
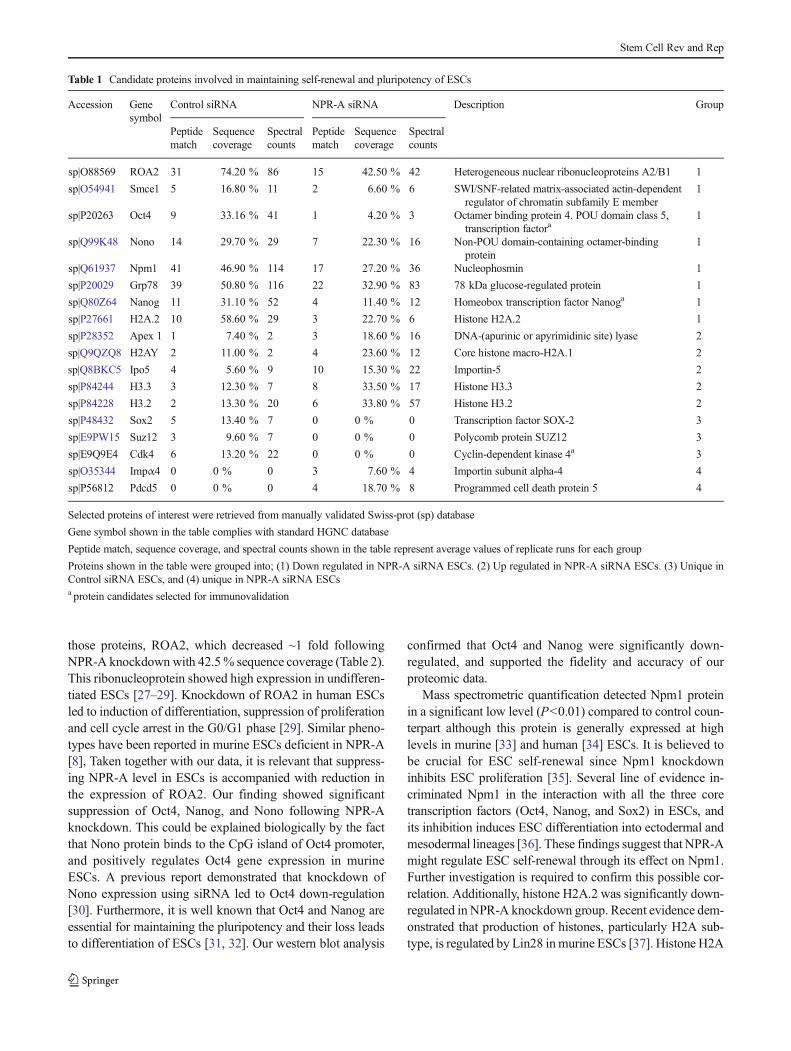
Table 1 Candidate proteins involved in maintaining self-renewal and pluripotency of ESCs
Accession Genesymbol
Control siRNA NPR-A siRNA Description Group
Peptidematch
Sequencecoverage
Spectralcounts
Peptidematch
Sequencecoverage
Spectralcounts
sp|O88569 ROA2 31 74.20 % 86 15 42.50 % 42 Heterogeneous nuclear ribonucleoproteins A2/B1 1
sp|O54941 Smce1 5 16.80 % 11 2 6.60 % 6 SWI/SNF-related matrix-associated actin-dependentregulator of chromatin subfamily E member
1
sp|P20263 Oct4 9 33.16 % 41 1 4.20 % 3 Octamer binding protein 4. POU domain class 5,transcription factora
1
sp|Q99K48 Nono 14 29.70 % 29 7 22.30 % 16 Non-POU domain-containing octamer-bindingprotein
1
sp|Q61937 Npm1 41 46.90 % 114 17 27.20 % 36 Nucleophosmin 1
sp|P20029 Grp78 39 50.80 % 116 22 32.90 % 83 78 kDa glucose-regulated protein 1
sp|Q80Z64 Nanog 11 31.10 % 52 4 11.40 % 12 Homeobox transcription factor Nanoga 1
sp|P27661 H2A.2 10 58.60 % 29 3 22.70 % 6 Histone H2A.2 1
sp|P28352 Apex 1 1 7.40 % 2 3 18.60 % 16 DNA-(apurinic or apyrimidinic site) lyase 2
sp|Q9QZQ8 H2AY 2 11.00 % 2 4 23.60 % 12 Core histone macro-H2A.1 2
sp|Q8BKC5 Ipo5 4 5.60 % 9 10 15.30 % 22 Importin-5 2
sp|P84244 H3.3 3 12.30 % 7 8 33.50 % 17 Histone H3.3 2
sp|P84228 H3.2 2 13.30 % 20 6 33.80 % 57 Histone H3.2 2
sp|P48432 Sox2 5 13.40 % 7 0 0 % 0 Transcription factor SOX-2 3
sp|E9PW15 Suz12 3 9.60 % 7 0 0 % 0 Polycomb protein SUZ12 3
sp|E9Q9E4 Cdk4 6 13.20 % 22 0 0 % 0 Cyclin-dependent kinase 4a 3
sp|O35344 Impα4 0 0 % 0 3 7.60 % 4 Importin subunit alpha-4 4
sp|P56812 Pdcd5 0 0 % 0 4 18.70 % 8 Programmed cell death protein 5 4
Selected proteins of interest were retrieved from manually validated Swiss-prot (sp) database
Gene symbol shown in the table complies with standard HGNC database
Peptide match, sequence coverage, and spectral counts shown in the table represent average values of replicate runs for each group
Proteins shown in the table were grouped into; (1) Down regulated in NPR-A siRNA ESCs. (2) Up regulated in NPR-A siRNA ESCs. (3) Unique inControl siRNA ESCs, and (4) unique in NPR-A siRNA ESCsa protein candidates selected for immunovalidation
Stem Cell Rev and Rep
coding region contains sequences, which activate gene expressionin a Lin28-dependent manner in murine ESCs [37, 38]. In addi-tion, overexpression of Lin28 leads to increase in the expressionlevel of H2A-containing mRNA [37]. Given the fact that Lin28 isinvolved inmurine ESC proliferation by regulating the expressionof cell cycle genes [37, 38], the effect ofNPR-A onESC cell cycleprogression might be modulated indirectly by histone H2A.
Furthermore, as a result of NPR-A suppression, SW1/SNFcomplex showed a modest down-regulation following NPR-Aknockdown. The subunits of SW1/SNF chromatin-remodellingcomplex have been identified previously as Nanog-associatedproteins [31], and evinced to be involved in several functions inpluripotent stem cells. For instance, SW1/SNF increases theprocess of reprogramming efficiency to pluripotent state [39]and plays an essential role in self-renewal and pluripotency ofESCs [40, 41]. Taken together with our proteomics findings, itis presumably that NPR-A might indirectly regulate ESC self-renewal through SW1/SNF. Additional work will be requiredto elucidate this possible mechanism.
Our results showed three protein candidates, required for self-renewal of ESCs, were only detected in control siRNA-treatedESCs, including Sox2, Suz12, and Cdk4. These transcriptionfactors are involved in regulating several genes in ESCs [3, 4,42, 43]. In addition to our previous report showing the signifi-cant reduction of Sox2 mRNA level after NPR-A knockdown[8], proteomics finding hereby proposes that this regulationmight be at the translational level as well. In this study, Suz12was detected only in control group with one significant peptide(Supplemental Table 1). Therefore it is vague to correlate thisfinding with NPR-A knockdown ESCs. However, it is notewor-thy to mention that lack of Suz12 shows elevation in thedifferentiation markers and impairment in proper differentiation[23], suggesting its important role in ESC self-renewal.
Cdk4 was notably detected in control group but not NPR-Aknockdown ESCs. This enzyme is known to be associated withcyclin D1 to form a complex that plays an important role inenhancing G1-S transition of the cell cycle [5, 26]. Western blotanalysis of protein lysate showed the absence of cyclin D1 protein
Fig. 4 Knockdown of NPR-A inhibits self-renewal and induces apoptosisofmurine ESCs. aWestern blot analysis of ESCs treatedwith control siRNAor NPR-A siRNA 48 h after transfection in the presence of LIF, showingreduced protein levels of Nanog and Oct4 (45 kDa) 48 h after siRNAtransfection. b Western blot analysis for cyclin D1 (37 kDa) 48 h after
siRNA transfection. β-actin (43 kDa) was used as a loading control. cApoptosis assay 48 h of ESCs treated with siRNAs as in panel A. Thenumber (M1) represents the percentage of Annexin V-positive cells andshown in bar plot in d panel Data represent mean ± S.D. (n=3); **P<0.01(two-tailed t-test)
Stem Cell Rev and Rep

in NPR-A siRNA-treated ESCs. This result comes along with ourprevious findings where NPR-A knockdown leads to cell cyclearrest at the G1 phase by down-regulating the transcript of cyclinD1 [8]. Taken together, we assume that the positive effect of NPR-A on ESC proliferation is dependent, at least partially, on acceler-ating the G1-S transition through regulating the Cdk4-cyclin D1complex protein. In addition, it supports the notion that NPR-A isrequired for maintaining self-renewal and pluripotency of ESCsby regulating the expression of several important proteins in-volved in maintaining ESC characteristics.
In the current experiment, NPR-A knockdown was possiblyimplicated in up-regulating the expression of some proteins in-volved inESCdifferentiation, including Impα4, Ipo5,H3 histonesand H2A.y. Previously, it was reported that undifferentiated
murine ESCs express Impα4 and Ipo5, which are significantlyup-regulated during early stages of differentiation [44]. Overex-pression of Impα4 in murine ESCs induces differentiation bydown-regulating Oct4 protein levels [44]. These findings suggestthat increased levels of Impα4 and Ipo5, which is induced byNPR-A knockdown, might be required during the early differen-tiation of ESCs. Also, we noticed that H3 histones and H2A.ywere significantly elevated in NPR-A siRNA-treated ESCs (Ta-ble 1). Recently, the role of histones in ESCs has been welldocumented. It was reported that expression levels of some his-tones are linked to ESC differentiation [45]. For instance, H3phosphorylation increases during early differentiation of ESCs[46]. A recent report demonstrated that macroH2A is expressedat high levels in the differentiated cells during embryogenesis, and
Fig. 5 Biological function network analysis of NPR-A knockdownESCs reflecting relationship between identified terms based on similarityof their associated genes. Node size correlates with the number of geneterms. Gene Term might be included in several groups. Degree of
connectivity between terms (edges) and definition of functional groups(node color) are calculated using kappa statistics. Network is laid outusing organic layout algorithm supported by cytoscape
Stem Cell Rev and Rep

its loss enhances the efficiency of the reprogramming topluripotency [47]. These findings suggest that suppressing NPR-A in murine ESCs might triggers the differentiation through up-regulating proteins involved in different lineage specifications.
Here we showed that NPR-A knockdown led to regulation ofseveral proteins related to cell survival, including Apex1, Grp78,and Pdcd5. Apex1 was found to be up-regulated as a result ofNPR-A knockdown. This enzyme is activated in response toDNA damage, which leads to increase the apoptotic response[48]. A recent study showed that Apex1 is expressed in thenucleus of human ESCs, and its knockdown after ionizingirradiation (IR) leads to significant reduction in the double strandbreaks (DSB) release and γ-H2AX expression in human ESCs[49]. We reported that Grp78 was down-regulated in NPR-Atreated ESCs. Grp78 is essential for the survival of pre-implantation embryos and ESCs [21, 22]. These findings suggestthat the observed high Apex1 and low Grp78 protein levelsinduced by NPR-A knockdown triggered the apoptotic signal-ling pathways.We also detected Pdcd5 inNPR-A siRNA-treatedESCs but not in control counterpart group. Pdcd5 dramaticallyincreases in cells undergoing apoptosis and plays an essentialrole in accelerating the apoptotic process of cells [50]. Takentogether with our apoptosis assay results, these findings indicatethat suppressing NPR-A induces cell death by activating pro-apoptotic proteins and reducing anti-apoptotic proteins.
Finally, the biological function interpreted by the GO an-notation network of NPR-A proteome further confirmed theinvolvement of several groups of genes, responsible for pro-duction of the above-mentioned proteins, in several functionssuch as cell cycle regulation, cell proliferation, cell fate spec-ification, regulation of cellular metabolic process, and pro-grammed cell death. Taken together with our previous find-ings, these evidences suggest the possible incrimination ofNPR-A in regulating several stem cell functions.
Conclusion
Our dataset finding adds new information to the growingknowledge of NPR-A role in ESCs. Here, we pinpointedseveral proteins that were possibly implicated in NPR-A path-way using innovative proteomics technology. Proteomicscreening of NPR-A knockdown ESCs was concurrently as-sociated with suppression of self-renewal proteins and induc-tion of other proteins involved in cell differentiation. In addi-tion, we showed that knockdown of NPR-A mediated theapoptotic process in ESCs by probable activation of pro-apoptotic proteins and suppression of anti-apoptotic proteins.These effects might be not strictly confined to NPR-A knock-down, thus further investigation, focusing on these proteins isrequired for additional confirmation. The current proteomicsstudy depicts a promising insight on the relevant role of NPR-A in controlling self-renewal and provides, at the meantime, a
useful reference for further studies. In the foreseeable future, athorough understanding on the biological implication of NPR-Awill be disclosed.
Competing interests The authors have declared no conflict of interest.
Funding This work was supported by JSPS (Japan Society for Promo-tion of Science) Grant-in-Aid for scientific research (B) to SM(23790933), and standard JSPS grant for foreign researcher (P 14105)to SM from Ministry of Education, Culture, Sports, Science and Tech-nology of Japan. The funders had no role in study design, data collectionand analysis, decision to publish, or preparation of the manuscript.
References
1. Chen, L., & Daley, G. Q. (2008). Molecular basis of pluripotency.Human Molecular Genetics, 17(R1), R23–R27.
2. Avilion, A. A., Nicolis, S. K., Pevny, L. H., Perez, L., Vivian, N., &Lovell-Badge, R. (2003). Multipotent cell lineages in early mousedevelopment depend on SOX2 function. Genes and Development,17(1), 126–140.
3. Boyer, L. A., Lee, T. I., Cole, M. F., Johnstone, S. E., Levine, S. S.,Zucker, J. P., et al. (2005). Core transcriptional regulatory circuitry inhuman embryonic stem cells. Cell, 122(6), 947–956.
4. Loh, Y. H., Wu, Q., Chew, J. L., Vega, V. B., Zhang, W., Chen, X., et al.(2006). The Oct4 and Nanog transcription network regulates pluripotencyin mouse embryonic stem cells. Nature Genetics, 38(4), 431–440.
5. Abdelalim, E. M. (2013). Molecular mechanisms controlling the cellcycle in embryonic stem cells. Stem Cell Reviews, 9(6), 764–773.
6. Niwa, H. (2007). How is pluripotency determined and maintained?Development, 134(4), 635–646.
7. Pandey, K. N. (2005). Biology of natriuretic peptides and theirreceptors. Peptides, 26(6), 901–932.
8. Abdelalim, E. M., & Tooyama, I. (2011). NPR-A regulates self-renewal and pluripotency of embryonic stem cells. Cell DeathDiseases, 2, e127.
9. Abdelalim, E.M., & Tooyama, I. (2009). BNP signaling is crucial forembryonic stem cell proliferation. PLoS ONE, 4(4), e5341.
10. Morishita, R., Gibbons, G. H., Pratt, R. E., Tomita, N., Kaneda, Y.,Ogihara, T., et al. (1994). Autocrine and paracrine effects of atrialnatriuretic peptide gene transfer on vascular smooth muscle and endo-thelial cellular growth. Journal of Clinical Investigation, 94(2), 824–829.
11. Levin, E. R., Gardner, D. G., & Samson, W. K. (1998). Natriureticpeptides. New England Journal of Medicine, 339(5), 321–328.
12. Silberbach, M., & Roberts, C. T., Jr. (2001). Natriuretic peptidesignalling: molecular and cellular pathways to growth regulation.Cellular Signalling, 13(4), 221–231.
13. Scott, N. J., Ellmers, L. J., Lainchbury, J. G.,Maeda, N., Smithies, O.,Richards, A. M., et al. (2009). Influence of natriuretic peptidereceptor-1 on survival and cardiac hypertrophy during development.Biochimica et Biophysica Acta, 1792(12), 1175–1184.
14. Abdelalim, E. M., & Tooyama, I. (2012). Regulation of self-renewaland pluripotency of embryonic stem cells: role of natriuretic peptidereceptor A. In M. A. Hayat (Ed.), Stem cells and cancer stem cells;therapeutic applications in disease and injury (Vol. 8, pp. 123–131).Netherlands: Springer.
15. Graumann, J., Hubner, N. C., Kim, J. B., Ko, K., Moser, M., Kumar,C., et al. (2008). Stable isotope labeling by amino acids in cell culture(SILAC) and proteome quantitation of mouse embryonic stem cellsto a depth of 5,111 proteins. Molecular and Cellular Proteomics,7(4), 672–683.
Stem Cell Rev and Rep

16. O’Brien, R. N., Shen, Z., Tachikawa, K., Lee, P. A., & Briggs, S. P.(2010). Quantitative proteome analysis of pluripotent cells by iTRAQmass tagging reveals post-transcriptional regulation of proteins re-quired for ES cell self-renewal. Molecular and Cellular Proteomics,9(10), 2238–2251.
17. Collier, T. S., Sarkar, P., Rao, B., & Muddiman, D. C. (2010).Quantitative top-down proteomics of SILAC labeled human embry-onic stem cells. Journal of the American Society for MassSpectrometry, 21(6), 879–889.
18. Novak, A., Amit, M., Ziv, T., Segev, H., Fishman, B., Admon, A.,et al. (2012). Proteomics profiling of human embryonic stem cells inthe early differentiation stage. Stem Cell Reviews, 8(1), 137–149.
19. Magdeldin, S., K. Yamamoto, Y. Yoshida, B. Xu, Y. Zhang, H.Fujinaka, et al. (2014). Deep proteome mapping of mouse kidneybased on OFFGel prefractionation reveals remarkable protein post-translational modifications. J Proteome Res,
20. Rappsilber, J., Mann, M., & Ishihama, Y. (2007). Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides forproteomics using StageTips. Nature Protocols, 2(8), 1896–1906.
21. Cociorva, D., L.T. D, and J.R. Yates. (2007). Validation of tandemmass spectrometry database search results using DTASelect. CurrProtoc Bioinformatics, Chapter 13, Unit 13 4.
22. Park, S. K., Venable, J. D., Xu, T., & Yates, J. R., 3rd. (2008). Aquantitative analysis software tool for mass spectrometry-based pro-teomics. Nature Methods, 5(4), 319–322.
23. Magdeldin, S., Yoshida, Y., Li, H., Maeda, Y., Yokoyama,M., Enany,S., et al. (2012). Murine colon proteome and characterization of theprotein pathways. BioData Mining, 5(1), 11.
24. Abdelalim, E. M., & Tooyama, I. (2012). NPR-C protects embryonicstem cells from apoptosis by regulating p53 levels. Stem Cells andDevelopment, 21(8), 1264–1271.
25. Carvalho, P. C., Hewel, J., Barbosa, V. C., & Yates, J. R., 3rd. (2008).Identifying differences in protein expression levels by spectralcounting and feature selection. Genetics and Molecular Research,7(2), 342–356.
26. Jirmanova, L., Afanassieff, M., Gobert-Gosse, S., Markossian, S., &Savatier, P. (2002). Differential contributions of ERK and PI3-kinaseto the regulation of cyclin D1 expression and to the control of the G1/S transition in mouse embryonic stem cells. Oncogene, 21(36),5515–5528.
27. Assou, S., Le Carrour, T., Tondeur, S., Strom, S., Gabelle, A., Marty,S., et al. (2007). A meta-analysis of human embryonic stem cellstranscriptome integrated into a web-based expression atlas. StemCells, 25(4), 961–973.
28. Van Hoof, D., Munoz, J., Braam, S. R., Pinkse, M. W., Linding, R.,Heck, A. J., et al. (2009). Phosphorylation dynamics during earlydifferentiation of human embryonic stem cells. Cell Stem Cell, 5(2),214–226.
29. Choi, H.S., H.M. Lee, Y.J. Jang, C.H. Kim, and C.J. Ryu. (2013).Heterogeneous Nuclear Ribonucleoprotein A2/B1 Regulates theSelfrenewal and Pluripotency of Human Embryonic Stem Cells viathe Control of the G1/S Transition. Stem Cells,
30. Park, Y., Lee, J. M., Hwang, M. Y., Son, G. H., & Geum, D. (2013).NonO binds to the CpG island of oct4 promoter and functions as atranscriptional activator of oct4 gene expression. Molecules andCells, 35(1), 61–69.
31. Niwa, H., Miyazaki, J., & Smith, A. G. (2000). Quantitative expres-sion of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nature Genetics, 24(4), 372–376.
32. Xie, X., Shi, Z., & Gu, W. (2012). Late-stage Freiberg’s diseasetreated with dorsal wedge osteotomy and joint distractionarthroplasty: technique tip. Foot and Ankle International, 33(11),1015–1017.
33. Elliott, S. T., Crider, D. G., Garnham, C. P., Boheler, K. R., & VanEyk, J. E. (2004). Two-dimensional gel electrophoresis database ofmurine R1 embryonic stem cells. Proteomics, 4(12), 3813–3832.
34. Richards, M., Tan, S. P., Tan, J. H., Chan, W. K., & Bongso, A.(2004). The transcriptome profile of human embryonic stem cells asdefined by SAGE. Stem Cells, 22(1), 51–64.
35. Wang, B. B., Lu, R., Wang, W. C., & Jin, Y. (2006). Inducible andreversible suppression of Npm1 gene expression using stably inte-grated small interfering RNA vector in mouse embryonic stem cells.Biochemical and Biophysical Research Communications, 347(4),1129–1137.
36. Johansson, H., & Simonsson, S. (2010). Core transcription factors,Oct4, Sox2 and Nanog, individually form complexes withnucleophosmin (Npm1) to control embryonic stem (ES) cell fatedetermination. Aging (Albany NY), 2(11), 815–822.
37. Xu, B., & Huang, Y. (2009). Histone H2a mRNA interacts withLin28 and contains a Lin28-dependent posttranscriptional regulatoryelement. Nucleic Acids Research, 37(13), 4256–4263.
38. Xu, B., Zhang, K., & Huang, Y. (2009). Lin28 modulates cell growthand associates with a subset of cell cycle regulator mRNAs in mouseembryonic stem cells. RNA, 15(3), 357–361.
39. Peric-Hupkes, D., Meuleman, W., Pagie, L., Bruggeman, S. W.,Solovei, I., Brugman, W., et al. (2010). Molecular maps of thereorganization of genome-nuclear lamina interactions during differ-entiation. Molecular Cell, 38(4), 603–613.
40. Chambers, I., Colby, D., Robertson,M., Nichols, J., Lee, S., Tweedie,S., et al. (2003). Functional expression cloning of Nanog, apluripotency sustaining factor in embryonic stem cells. Cell, 113(5),643–655.
41. Yan, Z.,Wang, Z., Sharova, L., Sharov, A. A., Ling, C., Piao, Y., et al.(2008). BAF250B-associated SWI/SNF chromatin-remodeling com-plex is required to maintain undifferentiated mouse embryonic stemcells. Stem Cells, 26(5), 1155–1165.
42. Chen, X., Xu, H., Yuan, P., Fang, F., Huss, M., Vega, V. B., et al.(2008). Integration of external signaling pathways with the core tran-scriptional network in embryonic stem cells. Cell, 133(6), 1106–1117.
43. Kim, J., Chu, J., Shen, X., Wang, J., & Orkin, S. H. (2008). Anextended transcriptional network for pluripotency of embryonic stemcells. Cell, 132(6), 1049–1061.
44. Young, J. C., Major, A. T., Miyamoto, Y., Loveland, K. L., & Jans, D.A. (2011). Distinct effects of importin alpha2 and alpha4 on Oct3/4localization and expression in mouse embryonic stem cells. FASEBJournal, 25(11), 3958–3965.
45. Golob, J. L., Paige, S. L., Muskheli, V., Pabon, L., & Murry, C. E.(2008). Chromatin remodeling during mouse and human embryonicstem cell differentiation.Developmental Dynamics, 237(5), 1389–1398.
46. Lee, E. R., McCool, K. W., Murdoch, F. E., & Fritsch, M. K. (2006).Dynamic changes in histone H3 phosphoacetylation during earlyembryonic stem cell differentiation are directly mediated bymitogen-and stress-activated protein kinase 1 via activation ofMAPK pathways. Journal of Biological Chemistry, 281(30),21162–21172.
47. Pasque, V., Radzisheuskaya, A., Gillich, A., Halley-Stott, R. P.,Panamarova, M., Zernicka-Goetz, M., et al. (2012). Histone variantmacroH2A marks embryonic differentiation in vivo and acts as anepigenetic barrier to induced pluripotency. Journal of Cell Science,125(Pt 24), 6094–6104.
48. Unnikrishnan, A., Raffoul, J. J., Patel, H. V., Prychitko, T. M.,Anyangwe, N., Meira, L. B., et al. (2009). Oxidative stress alters baseexcision repair pathway and increases apoptotic response in apurinic/apyrimidinic endonuclease 1/redox factor-1 haploinsufficient mice.Free Radical Biology and Medicine, 46(11), 1488–1499.
49. Kruta, M., Balek, L., Hejnova, R., Dobsakova, Z., Eiselleova, L.,Matulka, K., et al. (2013). Decrease in abundance of apurinic/apyrimidinic endonuclease causes failure of base excision repair inculture-adapted human embryonic stem cells. StemCells, 31(4), 693–702.
50. Chen, Y., Sun, R., Han,W., Zhang, Y., Song, Q., Di, C., et al. (2001).Nuclear translocation of PDCD5 (TFAR19): an early signal forapoptosis? FEBS Letters, 509(2), 191–196.
Stem Cell Rev and Rep