myeloperoxidase attracts neutrophils by physical forces
TRANSCRIPT

PHAGOCYTES, GRANULOCYTES, AND MYELOPOIESIS
Myeloperoxidase attracts neutrophils by physical forces*Anna Klinke,1 *Claudia Nussbaum,2 Lukas Kubala,3 Kai Friedrichs,1 Tanja K. Rudolph,1 Volker Rudolph,1
Hans-Joachim Paust,4 Christine Schroder,5 Daniel Benten,6 Denise Lau,1 Katalin Szocs,1 Paul G. Furtmuller,7
Peter Heeringa,8 Karsten Sydow,1 Hans-Jurgen Duchstein,9 Heimo Ehmke,10 Udo Schumacher,5 Thomas Meinertz,1
Markus Sperandio,2 and Stephan Baldus1
1Department of Cardiology, University Heart Center, Hamburg, Germany; 2Walter Brendel Centre of Experimental Medicine, Ludwig-Maximilians-University,Munich, Germany; 3Institute of Biophysics, Academy of Sciences of the Czech Republic, Brno, Czech Republic; Departments of 4Nephrology andRheumatology, 5Anatomy II, and 6Gastroenterology, University Hospital Eppendorf, Hamburg, Germany; 7Department of Chemistry, University of NaturalResources and Applied Life Sciences, Vienna, Austria; 8Department of Pathology and Medical Biology, University Medical Center Groningen, Groningen,The Netherlands; 9Department of Chemistry, University of Hamburg, Hamburg, Germany; and 10Department of Physiology, University Hospital Eppendorf,Hamburg, Germany
Recruitment of polymorphonuclear neu-trophils (PMNs) remains a paramount pre-requisite in innate immune defense and acritical cofounder in inflammatory vascu-lar disease. Neutrophil recruitment com-prises a cascade of concerted eventsallowing for capture, adhesion and extrava-sation of the leukocyte. Whereas PMNrolling, binding, and diapedesis are wellcharacterized, receptor-mediated pro-cesses, mechanisms attenuating the elec-trostatic repulsion between the nega-
tively charged glycocalyx of leukocyteand endothelium remain poorly under-stood. We provide evidence for myeloper-oxidase (MPO), an abundant PMN-derivedheme protein, facilitating PMN recruitmentby its positive surface charge. In vitro, MPOevoked highly directed PMN motility, whichwas solely dependent on electrostatic inter-actions with the leukocyte’s surface. In vivo,PMN recruitment was shown to be MPO-dependent in a model of hepatic ischemiaand reperfusion, upon intraportal delivery of
MPO and in the cremaster muscle exposedto local inflammation or to intraarterial MPOapplication. Given MPO’s affinity to both theendothelialandthe leukocyte’ssurface,MPOevolves as a mediator of PMN recruitmentbecause of its positive surface charge. Thiselectrostatic MPO effect not only displays aso far unrecognized, catalysis-independentfunction of the enzyme, but also highlights aprincipal mechanism of PMN attractiondriven by physical forces. (Blood. 2011;117(4):1350-1358)
IntroductionRecruitment of polymorphonuclear neutrophils (PMNs) is consid-ered a hallmark in host defense.1 With vessel- and tissue-infiltratingPMNs also being mechanistically linked to a broad range ofvascular inflammatory diseases including coronary artery disease,heart failure and ischemia/reperfusion injury, the pathophysiologicsignificance of PMN motility reaches far beyond innate immu-nity.2-4 So far, PMN migration is primarily viewed to be energy-dependent and cytoskeleton-dependent with G-protein coupledreceptors and integrins initiating and orchestrating signaling path-ways obligatory for neutrophil adhesion, spreading, diapedesis andchemotactic agitation.5-7 On activation, PMN releases myeloperox-idase (MPO), an abundant heme protein in PMN with potentbactericidal and vascular-inflammatory properties. The enzymeaccumulates along the endothelium and in the subendothelialspace,8 where it binds to anionic glycocalyx residues such asheparan sulfate glycosaminoglycans. Given the affinity of MPO toboth PMN and endothelial cells, we evaluated whether MPOaffects PMN locomotion and recruitment.
MethodsIsolation of PMN
Peripheral blood was drawn from healthy human volunteers and heparin-ized, and isolation of PMN was performed as previously described.9 Inbrief, after sedimentation in dextran solution (45 mg/mL), the supernatant
was placed over Histopaque 1077 (Sigma-Aldrich) for density gradientcentrifugation. Remaining red blood cells were eliminated by hypotoniclysis and the pellet was resuspended in Hanks balanced salt solution(HBSS; Invitrogen) containing 0.25% bovine serum albumin (BSA) (cellbuffer) and stored on ice until use.
Microslide motility experiments
Microslides (Ibidi �-slide I, ibitreat; IBIDI) were coated with fibrinogen(250 �g/mL). PMN or red blood cells (RBCs; 1 � 106/mL in cell buffer orplasma, where indicated) were incubated with 4-amino-benzoic acidhydrazide (ABAH; 50�M, Calbiochem) and H2O2 (50�M, 30 minutes,Merck), (-)-Blebbistatin (100�M, 30 minutes, Sigma-Aldrich), Cytochala-sin D (1�M, 30 minutes, Sigma-Aldrich), LY 294002 (200�M, 2 hours,Calbiochem), or with sodium azide and 2-deoyxglucose (50mM, 1 hour,Sigma-Aldrich). 100 �g of poly-L-arginine (PLA, Sigma-Aldrich), prota-mine (Protamin Valeant, MEDA Pharma), or histone H2A (Millipore) wasadded directly before application to microslides, where indicated. Forexperiments in pH 9.2, PMN were resuspended and lyophilized MPO wasreconstituted in cell buffer of pH 9.2. 100 �L of PMN suspension wasapplied into the microslide channel; cells were allowed to attach for5 minutes. Subsequently, 30 �L of MPO (120nM unless otherwise indi-cated, Planta Natural Products), recombinant, mutant MPO Q91T orM243T (120nM), IL-8 (25nM, PeproTech), or human serum albumin(120nM, Sigma-Aldrich) in cell buffer or plasma, optionally supplementedwith inhibitors, PLA, protamine, or histone, respectively, was added fromone side into the channel. Where indicated, Sepharose beads with anionic
Submitted May 10, 2010; accepted October 13, 2010. Prepublished online asBlood First Edition paper, October 27, 2010; DOI 10.1182/blood-2010-05-284513.
*A.K. and C.M. contributed equally to this study.
An Inside Blood analysis of this article appears at the front of this issue.
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 USC section 1734.
© 2011 by The American Society of Hematology
1350 BLOOD, 27 JANUARY 2011 � VOLUME 117, NUMBER 4
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom

(HiTrap SP HP, GE Healthcare/Amersham) or cationic (HiTrap Q HP, GEHealthcare) coating were used instead of PMN. The area of the steepestgradient was calculated as before (www.ibidi.com) and captured for20 minutes with a light microscope (CK 2, Olympus)–mounted charge-coupled device (CCD) camera (Retiga 1300, QImaging).
Image acquisition and processing of in vitro experiments
During leukocyte or Sepharose-bead motility experiments, pictures werecaptured at 0 minutes and 20 minutes, and time-lapse imaging was used tocreate a stack of images (iVision version 4.0, BioVision). The percentage ofcells oriented in an angle of less than 180° toward the segment of upwardgradient was evaluated (PMN in up-gradient segment). Twenty PMNs perstack were tracked and transformed to 2-dimensional plots, with thex-direction indicating the upward gradient orientation (manual tracking andchemotaxis-tool, created for ImageJ by IBIDI). The mean accumulateddistance and the x-forward-migration index (x-displacement � accumulateddistance�1) were calculated.
Quantification of f-actin with flow cytometry
PMN (1 � 106/mL cell buffer) were incubated with MPO (120nM) or IL-8(25nM). After indicated times, cells were fixed, permeabilized, andincubated with Alexa Fluor 488–Phalloidin (20 minutes, 5 U/mL in phos-phate buffer solution [PBS] with 1% BSA, Molecular Probes). Flowcytometric analysis was performed using a FACSCalibur flow cytometerand CellQuest Pro 5.1 software (BD Biosciences).
Mouse liver experimental protocols
Male C57BL/6J (wild-type; WT) and MPO_tm1lus (Mpo�/�) mice aged12-15 weeks were treated either to occlude the blood supply to the medianand left hepatic lobes with an atraumatic vascular clamp for 90 minutes, andreperfusion then initiated; or, their inferior mesenteric veins were injectedwith recombinant human serum albumin (Prospec); recombinant activeMPO (R&D Systems); inactive MPO M243T, Q91T MPO, or recombinantMPO preincubated with 50�M ABAH and H2O2 (4 �g in 200 �L ofphysiologic saline), respectively. Two hours after intraportal injection or20 hours after initiation of reperfusion, mice were again anaesthetized. Theliver was flushed by intraportal injection of 1 mL of PBS and excised. Allanimal experiments were approved by the local ethics committees (Ham-burg, Germany, G100/06) and in accordance with the US National ResearchCouncil’s “Guide for the Care and Use of Laboratory Animals.”
Quantitative assessment of hepatic PMN infiltration byimmunohistochemistry
Frozen liver specimens were fixed with acetone. Sections were incubatedwith anti-mouse Ly6G primary antibody (ratio of 1:40; Hycult Biodiagnos-tics), and endogenous peroxidase activity was blocked. The secondaryantibody was horseradish peroxidase (hrp)–labeled rabbit IgG to rat IgG(1:100, Dako), and the tertiary antibody was hrp-labeled goat anti–rabbitIgG (1:500, Vector Laboratories) in 3% normal mouse serum. PMNs werestained with 3-amino-9-ethylcarbazol (AEC) solution and tissue wascounterstained with hematoxyline. The number of neutrophils attached tosinusoids or extravasated into hepatic parenchyma was counted in30 high-power fields per animal (original magnification �600) under a lightmicroscope (Olympus).
Mouse cremaster muscle experimental protocols
Recombinant murine tumor necrosis factor (TNF)-� (500 ng per mouse,R&D) in sterile saline was injected into the scrotum of male C57BL/6J(WT) mice and MPO_tm1lus (Mpo�/�) mice aged 12-15 weeks. Two hoursafter injection, the carotid artery was cannulated and the cremaster musclesurgically prepared for intravital microscopy as previously described.10
During intravital microscopy, the cremaster muscle preparation was continu-ously superfused with a 37°C warm buffer solution. In separate experi-ments, the cremaster muscle was surgically prepared without prior treat-ment (trauma induced inflammation). Twenty minutes after surgical
preparation of the cremaster muscle, the animals received a bolus injectionof active MPO (20 �g per mouse) or recombinant, inactive MPO Q91T(20 �g per mouse) in a volume of 0.2 mL of sterile saline, or an injection ofsaline alone, into the carotid artery. During all experiments systemic bloodsamples were obtained (10 �L in 90 �L of Tuerk solution; Merck) forquantification of systemic white blood cell count. All animal experimentswere approved by the local ethics committees (Regierung von Oberbayern,Az. 55.2-1-54-2531-80-07) and in accordance with the “Guide for the Careand Use of Laboratory Animals.”
Intravital microscopy and data analysis
Intravital microscopy was performed on an upright microscope (OlympusBX51) with a saline immersion objective (original magnification 40�, 0.8numerical aperture) as described.10 Experiments were recorded on anS-VHS recorder using a CCD camera (model CF8/1, Kappa) for lateroffline analysis. Vessel diameter, segment length of postcapillary cremastermuscle venules, and PMN diameter were measured with a digital imageprocessing system.11 Centerline red blood cell velocity was determined by adual photodiode and a digital online cross-correlation program (CircusoftInstrumentation) and used for offline calculation of mean blood flowvelocity and wall shear rates as previously reported.12 Supplemental Tables1 and 2 (available on the Blood Web site; see the Supplemental Materialslink at the top of the online article) give an overview of microvascularparameters observed during intravital microscopy experiments. PMNadherence was quantified by counting the number of leukocytes per mm2 ofvessel surface that remained stationary for more than 30 seconds.
Whole-mount histology
To assess leukocyte extravasation on intrascrotal injection of TNF-�, wholemounts of cremaster muscles were prepared as previously described.13
Leukocyte extravasation was quantified by counting the number ofextravascular cells per mm2 of cremaster muscle tissue using a Zeiss uprightmicroscope with an oil immersion objective (original magnification 100�,1.3 numerical aperture).
MPO and Alcian blue staining
For hepatic MPO staining, human MPO (4 �g, Planta Natural Products) orhuman serum albumin (4 �g, Sigma-Aldrich) was injected to the mesen-teric vein of C57Bl/6J mice as described. Hepatic sections were incubatedwith the primary antibody to human MPO (rabbit, Calbiochem). Forcremasteric MPO staining, human MPO (20 �g) was injected into thecarotid artery of WT mice, and after 10 minutes an antibody to human MPO(40 �g, rabbit, Calbiochem) was injected. After another 10 minutes, thevena cava inferior was incised and 3 mL of PBS were flushed via the carotidcatheter. Where indicated, Alcian blue 8 GX (1 mL, 0.1% solution,Sigma-Aldrich) was injected into the artery and flushed with 1 mL of PBSand 1 mL of 4% formaldehyde. The cremaster muscle was excised asdescribed, placed on a microslide, and fixed with 4% formaldehyde. Allsamples were treated with secondary antibody Alexa 488 to rabbit IgG(Molecular Probes) and tissue was counterstained with DAPI. Images wereacquired with a Leica microscope (DMLB) and IVision software.
Statistical analysis
Data were tested for normal distribution using the Kolmogorov-Smirnovtest. When normality was shown, Student unpaired t test was used forpairwise comparison; otherwise, the Mann-Whitney rank sum test wasused. A before-after comparison was performed with the Student pairedt test. For multiple comparison, one-way ANOVA followed by a Bonferronipost hoc test, or Kruskal-Wallis ANOVA on ranks test followed by Dunnpost hoc test, was used as appropriate. A P value � .05 was consideredstatistically significant. All statistical analyses were carried out withSPSS Version 13 (SPSS). Data are presented as mean � SEM.
MYELOPEROXIDASE ATTRACTS NEUTROPHILS 1351BLOOD, 27 JANUARY 2011 � VOLUME 117, NUMBER 4
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom

Results
To test PMN motility in response to MPO, isolated human PMNadded to microslides were challenged with MPO administered toone pole of the chamber. The resulting gradient of MPO provokedPMN motility in a concentration-dependent manner (Figure 1A).The presence of human plasma did not affect MPO-directedmotility (Suppl. Figure 1A). The extent of directed PMN locomo-tion toward MPO was significantly elevated compared with humanserum albumin (HSA) and the chemokine interleukin-8 (IL-8,Figure 1B), with eosinophil peroxidase (EPO) attracting PMN to asimilar extent as MPO (supplemental Figure 1B). Interestingly,inactivation of MPO by the inhibitor 4-amino-benzoic acid hydra-zide (ABHA) did not attenuate MPO-dependent PMN motility.Furthermore, 2 MPO variants, which lack either total enzymaticactivity (Q91T) or the chlorinating, brominating, NO-oxidizing andNO-nitrating activity of the enzyme (M243T14), provoked PMNattraction to a similar extent as active MPO, thus indicating amechanism that operates irrespective of the enzyme’s catalyticactivity (Figure 1C, D).
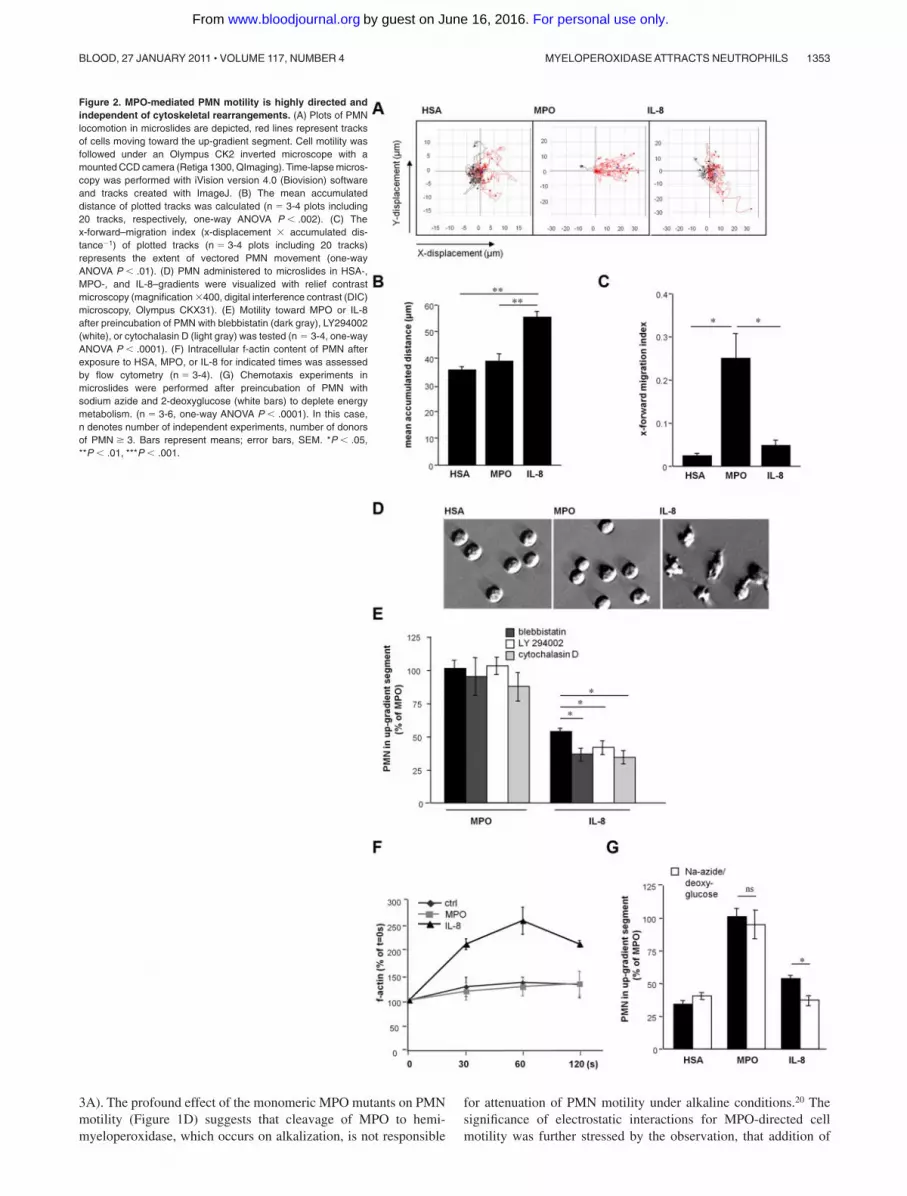
Remarkably, although MPO-exposed PMN traveled a shorterdistance compared with IL-8, MPO-dependent motility was strik-ingly more vectored (as expressed by the x-forward-migrationindex15; Figure 2A-C, supplemental Figure 2). These differencesbetween MPO-evoked compared with IL-8–evoked motility wereaccompanied by a discrepancy in cell morphology: PMN exposed
to MPO lacked the morphologic characteristics observed onmigration in response to IL-8, that is, polarization and formation oflamellipodia16 (Figure 2D). Given these directional and morpho-logic peculiarities in PMN motility toward MPO, characterizationof central intracellular signaling cascades involved in mediatingleukocyte migration were explored. Whereas inhibition of non-muscle myosin II by blebbistatin, phosphatidylinositol 3-kinaseactivity by LY 294002, and actin filament polymerization usingcytochalasin D impaired IL-8-induced migration,17-19 MPO-induced PMN motility remained unaffected. The rise in filamen-tous actin content (f-actin), which is a prerequisite of active PMNmigration and observed in IL-8 treated cells, was blunted inMPO-exposed cells. This indicates that PMN motility on MPOexposure is independent of cytoskeletal rearrangements (Figure 2E,F). Instead, MPO provoked passive PMN locomotion irrespectiveof energy consumption, as evidenced by preserved cell motility inthe presence of respiratory chain and glycolysis-inhibiting sodiumazide and 2-deoxyglucose, respectively (Figure 2G). Moreover,PMN movement toward MPO was retained after fixation of thecells using methanol (Figure 3A).
Given that MPO-dependent PMN motility was revealed to beindependent of energy-consuming cytoskeletal modifications, elec-trostatic interactions between the cationic MPO and the negativelycharged PMN-surface were explored. In support of electrostaticinteractions as a prerequisite for MPO-evoked PMN motility,alkalization of the assay buffer to the isoelectric point of MPO(pI 9.2) entirely prevented motility of methanol-fixed PMN (Figure
Figure 1. MPO-dependent PMN motility in vitro. PMN motilitywas evaluated at the steepest gradient of a chemotactic factor orcontrol in Ibidi microslides. (A) MPO provoked directed locomotionof PMN in a concentration-dependent manner (n � 4, one-wayANOVA; P � .0001). (B) Directed PMN motility after application ofhuman serum albumin (HSA, n � 21), MPO (n � 34), and interleu-kin-8 (IL-8, n � 9) was investigated (one-way ANOVA; P � .0001).(C) Directed motility toward HSA or MPO was evaluated onpreincubation with the MPO-inhibitor 4-amino-benzoic acid hydra-zide (ABAH; white bars, n � 3-4). (D) MPO-variants Q91T (n � 4)and M243T (n � 9) devoid of catalytic activity also yieldedincreased directed PMN motility (one-way ANOVA P � .003).Number of independent experiments denoted by n; number ofdonors of PMN � 3. Bars represent means; error bars, standarderror of the mean (SEM). *P � .05, **P � .01, ***P � .001.
1352 KLINKE et al BLOOD, 27 JANUARY 2011 � VOLUME 117, NUMBER 4
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom
3A). The profound effect of the monomeric MPO mutants on PMNmotility (Figure 1D) suggests that cleavage of MPO to hemi-myeloperoxidase, which occurs on alkalization, is not responsible
for attenuation of PMN motility under alkaline conditions.20 Thesignificance of electrostatic interactions for MPO-directed cellmotility was further stressed by the observation, that addition of
Figure 2. MPO-mediated PMN motility is highly directed andindependent of cytoskeletal rearrangements. (A) Plots of PMNlocomotion in microslides are depicted, red lines represent tracksof cells moving toward the up-gradient segment. Cell motility wasfollowed under an Olympus CK2 inverted microscope with amounted CCD camera (Retiga 1300, QImaging). Time-lapse micros-copy was performed with iVision version 4.0 (Biovision) softwareand tracks created with ImageJ. (B) The mean accumulateddistance of plotted tracks was calculated (n � 3-4 plots including20 tracks, respectively, one-way ANOVA P � .002). (C) Thex-forward–migration index (x-displacement � accumulated dis-tance�1) of plotted tracks (n � 3-4 plots including 20 tracks)represents the extent of vectored PMN movement (one-wayANOVA P � .01). (D) PMN administered to microslides in HSA-,MPO-, and IL-8–gradients were visualized with relief contrastmicroscopy (magnification �400, digital interference contrast (DIC)microscopy, Olympus CKX31). (E) Motility toward MPO or IL-8after preincubation of PMN with blebbistatin (dark gray), LY294002(white), or cytochalasin D (light gray) was tested (n � 3-4, one-wayANOVA P � .0001). (F) Intracellular f-actin content of PMN afterexposure to HSA, MPO, or IL-8 for indicated times was assessedby flow cytometry (n � 3-4). (G) Chemotaxis experiments inmicroslides were performed after preincubation of PMN withsodium azide and 2-deoxyglucose (white bars) to deplete energymetabolism. (n � 3-6, one-way ANOVA P � .0001). In this case,n denotes number of independent experiments, number of donorsof PMN � 3. Bars represent means; error bars, SEM. *P � .05,**P � .01, ***P � .001.
MYELOPEROXIDASE ATTRACTS NEUTROPHILS 1353BLOOD, 27 JANUARY 2011 � VOLUME 117, NUMBER 4
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom
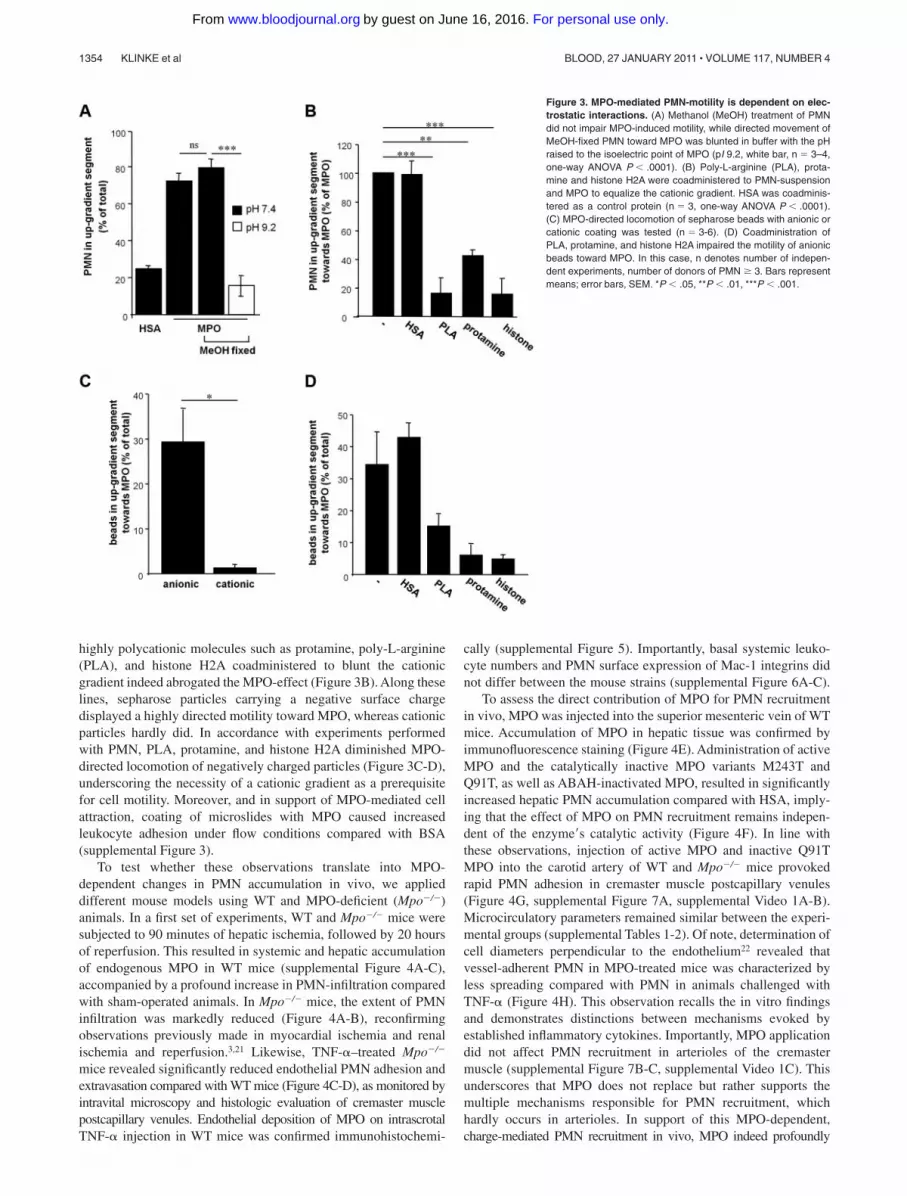
highly polycationic molecules such as protamine, poly-L-arginine(PLA), and histone H2A coadministered to blunt the cationicgradient indeed abrogated the MPO-effect (Figure 3B). Along theselines, sepharose particles carrying a negative surface chargedisplayed a highly directed motility toward MPO, whereas cationicparticles hardly did. In accordance with experiments performedwith PMN, PLA, protamine, and histone H2A diminished MPO-directed locomotion of negatively charged particles (Figure 3C-D),underscoring the necessity of a cationic gradient as a prerequisitefor cell motility. Moreover, and in support of MPO-mediated cellattraction, coating of microslides with MPO caused increasedleukocyte adhesion under flow conditions compared with BSA(supplemental Figure 3).
To test whether these observations translate into MPO-dependent changes in PMN accumulation in vivo, we applieddifferent mouse models using WT and MPO-deficient (Mpo�/�)animals. In a first set of experiments, WT and Mpo�/� mice weresubjected to 90 minutes of hepatic ischemia, followed by 20 hoursof reperfusion. This resulted in systemic and hepatic accumulationof endogenous MPO in WT mice (supplemental Figure 4A-C),accompanied by a profound increase in PMN-infiltration comparedwith sham-operated animals. In Mpo�/� mice, the extent of PMNinfiltration was markedly reduced (Figure 4A-B), reconfirmingobservations previously made in myocardial ischemia and renalischemia and reperfusion.3,21 Likewise, TNF-�–treated Mpo�/�
mice revealed significantly reduced endothelial PMN adhesion andextravasation compared with WT mice (Figure 4C-D), as monitored byintravital microscopy and histologic evaluation of cremaster musclepostcapillary venules. Endothelial deposition of MPO on intrascrotalTNF-� injection in WT mice was confirmed immunohistochemi-
cally (supplemental Figure 5). Importantly, basal systemic leuko-cyte numbers and PMN surface expression of Mac-1 integrins didnot differ between the mouse strains (supplemental Figure 6A-C).
To assess the direct contribution of MPO for PMN recruitmentin vivo, MPO was injected into the superior mesenteric vein of WTmice. Accumulation of MPO in hepatic tissue was confirmed byimmunofluorescence staining (Figure 4E). Administration of activeMPO and the catalytically inactive MPO variants M243T andQ91T, as well as ABAH-inactivated MPO, resulted in significantlyincreased hepatic PMN accumulation compared with HSA, imply-ing that the effect of MPO on PMN recruitment remains indepen-dent of the enzyme�s catalytic activity (Figure 4F). In line withthese observations, injection of active MPO and inactive Q91TMPO into the carotid artery of WT and Mpo�/� mice provokedrapid PMN adhesion in cremaster muscle postcapillary venules(Figure 4G, supplemental Figure 7A, supplemental Video 1A-B).Microcirculatory parameters remained similar between the experi-mental groups (supplemental Tables 1-2). Of note, determination ofcell diameters perpendicular to the endothelium22 revealed thatvessel-adherent PMN in MPO-treated mice was characterized byless spreading compared with PMN in animals challenged withTNF-� (Figure 4H). This observation recalls the in vitro findingsand demonstrates distinctions between mechanisms evoked byestablished inflammatory cytokines. Importantly, MPO applicationdid not affect PMN recruitment in arterioles of the cremastermuscle (supplemental Figure 7B-C, supplemental Video 1C). Thisunderscores that MPO does not replace but rather supports themultiple mechanisms responsible for PMN recruitment, whichhardly occurs in arterioles. In support of this MPO-dependent,charge-mediated PMN recruitment in vivo, MPO indeed profoundly
Figure 3. MPO-mediated PMN-motility is dependent on elec-trostatic interactions. (A) Methanol (MeOH) treatment of PMNdid not impair MPO-induced motility, while directed movement ofMeOH-fixed PMN toward MPO was blunted in buffer with the pHraised to the isoelectric point of MPO (pI 9.2, white bar, n � 3–4,one-way ANOVA P � .0001). (B) Poly-L-arginine (PLA), prota-mine and histone H2A were coadministered to PMN-suspensionand MPO to equalize the cationic gradient. HSA was coadminis-tered as a control protein (n � 3, one-way ANOVA P � .0001).(C) MPO-directed locomotion of sepharose beads with anionic orcationic coating was tested (n � 3-6). (D) Coadministration ofPLA, protamine, and histone H2A impaired the motility of anionicbeads toward MPO. In this case, n denotes number of indepen-dent experiments, number of donors of PMN � 3. Bars representmeans; error bars, SEM. *P � .05, **P � .01, ***P � .001.
1354 KLINKE et al BLOOD, 27 JANUARY 2011 � VOLUME 117, NUMBER 4
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom

attenuated the anionic endothelial surface charge in cremastermuscle venules, illustrated by diminished deposition of the cationic
dye Alcian blue, which binds to the glycocalyx in a charge-dependent fashion23-25 (Figure 4I).
Figure 4. MPO provokes neutrophil recruitment in vivo.(A) Representative liver sections of WT and Mpo�/� mice after90 minutes of hepatic ischemia with subsequent 20 hours ofreperfusion are shown (PMN � red-brown; �200, captured withZeiss AxioCam HRc mounted on a Zeiss Axioskop with AxioVi-sion 3.1. Tonal value correction, brightness, and contrast wereadjusted with Adobe Photoshop CS3 extended 10.0). (B) PMNwere quantified in liver sections of WT or Mpo�/� mice on hepaticischemia and reperfusion (hpf � high-power field; magnification�600; sham n � 5-8, treated n � 9-10, one-way ANOVAP � .0001). (C) Number of adherent leukocytes in postcapillaryvenules of the cremaster muscle of WT (19 vessels in 4 mice)and Mpo�/� mice (18 vessels in 4 mice) stimulated with TNF-�was assessed by intravital microscopy. (D) Number of perivascu-lar PMN in TNF-� stimulated cremaster muscle of WT (n � 4)and Mpo�/� mice (n � 4) was evaluated in Giemsa-stainedwhole mounts. (E) MPO-immunoreactivity (�MPO, green, AlexaFluor 488) on intraportal injection of HSA or human MPO isshown in hepatic sections of WT mice (blue � Dapi, originalmagnification �100, captured with Retiga 1300 CCD cameramounted on Leica DMLB fluorescence microscope by iVisionversion 4.0, processed as described in panel A). (F) HepaticPMN-infiltration was quantified on intraportal injection of HSA,active MPO (MPO), inactive MPO M243T and Q91T, or ABAH-inactivated MPO (n � 3-8, hpf � high-power field, magnification�600, one-way ANOVA P � .01). (G) Number of adherentleukocytes in cremaster muscle postcapillary venules was as-sessed before (basal, white bars) and after (post injection, blackbars) injection of saline solution (16 vessels in 5 mice), active(15 vessels in 3 mice) or inactive MPO Q91T (12 vessels in3 mice) into the carotid artery of WT mice. (H) Mean diameter ofPMN adherent to the endothelium of cremaster muscle venulesin mice on injection of saline, MPO, or Q91T MPO to the carotidartery or intrascrotal TNF-� application (500 ng) and a represen-tative image of vessel-adherent PMN in MPO and TNF-�–treatedmice are shown (magnification �400; intravital microscopy wasperformed with an Olympus BX51 microscope with a CF8/1CCD-camera [Kappa]), recorded on S-VHS recorder and digi-talized with MetaMorph software [Molecular Devices]). (I) Intralu-minal staining of MPO (green) and negatively charged glycocalyxresidues (Alcian blue) in WT mice on MPO injection of the carotidartery (MPO i.a.) or control (ctrl). Scale bars, 30 �m; fluores-cence images captured and processed as described in panelE and bright-field images as described in panel A. Bars representmeans; error bars, SEM. *P � .05, **P � .01, ***P � .001.
MYELOPEROXIDASE ATTRACTS NEUTROPHILS 1355BLOOD, 27 JANUARY 2011 � VOLUME 117, NUMBER 4
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom

Discussion
Recruitment of PMN during inflammation implies a complexmultistep process, which is initiated by capture, tethering, androlling of leukocytes followed by leukocyte adhesion, spreading,and crawling on the inflamed vessel wall, and finally diapedesisinto tissue. We herein propose a central role of MPO during theearly steps of PMN interaction with the endothelium, this rolebeing mediated by electrostatic forces between the cationic MPOsurface and the anionic PMN surface.
Whereas the initial steps of the migration cascade are primarilyconsidered to be selectin-dependent, nonselective physical forceshave been suggested as alternative and potentially precedingmechanisms allowing PMN interaction with the vessel wall.Previous reports speculated on cationic proteins being responsiblefor the initiation of leukocyte emigration by alteration of thesurface charge.26 The attenuation of electrostatic repulsive forceshas been hypothesized to permit short-range forces like Van derWaals and hydrophobic forces between leukocytes and the endothe-lium to act, and thereby facilitate leukocyte adhesion.27 In fact,reports in the 1970s and early 1980s suggested that PMN-derivedproteins enhance PMN margination and aggregation by decreasingnegative surface charge, but these reports failed to identify theunderlying effector molecules.28-30 In accordance with these obser-vations, sulfated glycosaminoglycans became established epitopesmediating the repulsion of blood cells and attenuating adhesion ofcirculating leukocytes, in part because of their negative charge, andby shielding endothelial adhesion molecules. Depletion of heparansulfate chains has been shown to increase leukocyte adhesion inpostcapillary venules,31 whereas the negative charge of the leuko-cyte surface protein leukosialin (CD 43) appears to exert antiadhe-sive properties.32
The current data now expand on these findings and introduceMPO as a powerful inducer of PMN recruitment. In fact, as theenzyme is capable of attracting PMN in vitro independently ofcytoskeletal-dependent and energy-dependent rearrangements ofthe leukocyte, electrostatic forces may indeed account for essentialsteps of PMN migration in vivo. Given its high affinity toglycosaminoglycans and matrix proteins such as collagen, fibronec-tin,8,33 and fibrinogen (data not shown), MPO, released at andcovering sites of inflammation, attenuates the antiadhesive proper-ties of the vessel wall. This cationic “coat” may ease the interplayof PMN with the endothelium and allow PMN to interact with
adhesion molecules harbored within the glycocalyx.34 Further-more, MPO deposition by adherent and crawling leukocytes on theendothelial surface layer may even help to guide upcoming PMN tothe exit point into tissue (Figure 5). This concept of MPOsupporting established mechanisms of PMN recruitment ratherthan unselectively promoting firm PMN adhesion is underscoredby the fact that the enzyme did not provoke PMN recruitment inarterioles, where leukocyte interaction with the endothelium re-mains restricted because of high shear forces and a limitedexpression of adhesion molecules.
Alternative mechanisms apart from electrostatic interactionsmay be responsible for the current observations. For example, onecould speculate that MPO increases vascular permeability asshown for other cationic molecules.35,36 Vascular permeability ismainly regulated by the adhesion molecule vascular endothelial(VE)–cadherin, with its blockade resulting in increased permeabil-ity and enhanced PMN emigration.37 However, research does notconfirm that MPO affects vascular permeability.8 Moreover, MPObinding to the neutrophil integrin Mac-138,39 may facilitate PMNadhesion and thus support electrostatically driven cell recruitment,as previously described for elastase and proteinase 3.40,41
It is possible to deduce from the current study that other cationicgranular proteins should exhibit similar effects as MPO. However,electrostatically driven PMN recruitment does not only depend onthe pI of the affecting protein: For example, lactoferrin, �-chymo-trypsinogen A, and histone, despite a similar pI, promote endothe-lial leukocyte adhesion in varying degrees.42 In fact, the differentbinding affinities of the proteins to the endothelium appear to be ofcritical importance as well.43 MPO is characterized by its highaffinity to heparan sulfate glycosaminoglycans,8,44 the major com-ponents of the endothelial surface layer. In conjunction with itsabundant granular expression, this results in profound depositionon the endothelial surface and might account for the enzyme’sprofound effects on PMN recruitment.
Of note, the proposed mechanism might also affect the affinityof other blood corpuscles toward the vessel wall as shown, forexample, for red blood cells in vitro (supplemental Figure 8).However, MPO-mediated affinity of leukocytes to the vessel wallmight be of particular significance, given that leukocytes subse-quently adhere to endothelial cells and extravasate in an MPO-independent fashion.
In conclusion, these findings describe a novel mechanismunderlying neutrophil motility and highlight a so-far unrecognizedrole for MPO in the recruitment of PMN, which is independent of
Figure 5. Scheme of the suggested mechanism of MPO-mediated PMN recruitment. Electrostatic repulsion between the negatively charged glycocalyx of PMN and thevessel wall prevents the interactions of PMN with the endothelium. Given the difference in height, the glycocalyx ( 500 nm) shields selectins ( 40 nm), which communicatethe definite contact of PMN to the vessel wall (for instance, binding of constitutively expressed P-selectin glycoprotein ligand-1 [PSGL-1] on the PMN membrane withP-selectins on the endothelial surface). Thus binding of MPO to glycosaminoglycans reduces the negative surface charge and allows for electrostatic attraction of PMN to theendothelium, which then mediates receptor-ligand interactions, resulting in PMN tethering and rolling, adhesion, and diapedesis.
1356 KLINKE et al BLOOD, 27 JANUARY 2011 � VOLUME 117, NUMBER 4
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom

the enzyme’s catalytic activity. Instead, PMN attraction is commu-nicated by MPO’s positive charge, an observation that calls forrevisiting the role of MPO in innate immunity and the impact ofphysical forces on the recruitment of neutrophils.
Acknowledgments
The authors thank Klaus Ley, Barbara Walzog, Jurgen Schymein-sky, and Henry Bourne for insightful discussions, and HartwigWieboldt and Susanne Bierschenk for expert technical assistance.
This work was supported by the Deutsche Forschungsgemein-schaft (S.B. and T.K.R.), the Deutsche Herzstiftung (V.R.), theWerner Otto Stiftung (S.B.) and the Academy of Science CzechRepublic (grant M200040908, L.K.). P.H. is supported by theNetherlands Organization for Scientific Research (VIDI 91766341).
Authorship
Contribution: A.K. designed the project, performed experiments,and wrote the manuscript; C.N. designed and performed the
intravital microscopy experiments and wrote the manuscript; L.K.,K.F., D.B., D.L., and K. Szocs provided suggestions on the projectand performed experiments; H.-J.P. performed flow cytometricexperiments; C.S. and U.S. were responsible for flow chamberexperiments; P.H. designed animal experiments; P.G.F. synthesizedMPO variants and made suggestions on the in vitro experiments;T.K.R., V.R., K. Sydow, H.-J.D., H.E., and T.M. provided sugges-tions on the project and revised the manuscript; and M.S. and S.B.supervised the project and wrote the manuscript.
Conflict-of-interest disclosure: The authors declare no compet-ing financial interests.
Correspondence: Anna Klinke, PhD, Cardiovascular ResearchCenter, University Hospital Hamburg Eppendorf and UniversityHeart Center Hamburg, Department of Cardiology, Martinistr52, Hamburg, Germany, 20246; e-mail: [email protected]; or Stephan Baldus, MD, Cardiovascular ResearchCenter, University Hospital Hamburg Eppendorf and UniversityHeart Center Hamburg, Department of Cardiology, Martinistr52, Hamburg, Germany, 20246; e-mail: [email protected].
References
1. Nathan C. Neutrophils and immunity: challengesand opportunities. Nat Rev Immunol. 2006;6(3):173-182.
2. Naruko T, Ueda M, Haze K, van der Wal AC, et al.Neutrophil infiltration of culprit lesions in acutecoronary syndromes. Circulation. 2002;106(23):2894-2900.
3. Askari AT, Brennan ML, Zhou X, et al. Myeloper-oxidase and plasminogen activator inhibitor 1play a central role in ventricular remodeling aftermyocardial infarction. J Exp Med. 2003;197(5):615-624.
4. Litt MR, Jeremy RW, Weisman HF, Winkelstein JA,Becker LC. Neutrophil depletion limited to reper-fusion reduces myocardial infarct size after 90minutes of ischemia. Evidence for neutrophil-me-diated reperfusion injury. Circulation. 1989;80(6):1816-1827.
5. Ley K, Laudanna C, Cybulsky MI, Nourshargh S.Getting to the site of inflammation: the leukocyteadhesion cascade updated. Nat Rev Immunol.2007;7(9):678-689.
6. Ridley AJ, Schwartz MA, Burridge K, et al. Cellmigration: integrating signals from front to back.Science. 2003;302(5651):1704-1709.
7. Phillipson M, Heit B, Colarusso P, Liu L,Ballantyne CM, Kubes P. Intraluminal crawling ofneutrophils to emigration sites: a molecularly dis-tinct process from adhesion in the recruitmentcascade. J Exp Med. 2006;203(12):2569-2575.
8. Baldus S, Eiserich JP, Mani A, et al. Endothelialtranscytosis of myeloperoxidase confers specific-ity to vascular ECM proteins as targets of tyrosinenitration. J Clin Invest. 2001;108(12):1759-1770.
9. Eiserich JP, Baldus S, Brennan ML, et al. Myelo-peroxidase, a leukocyte-derived vascular NO oxi-dase. Science. 2002;296(5577):2391-2394.
10. Sperandio M, Frommhold D, Babushkina I, et al.Alpha 2,3-sialyltransferase-IV is essential forL-selectin ligand function in inflammation. EurJ Immunol. 2006;36(12):3207-3215.
11. Pries AR. A versatile video image analysis systemfor microcirculatory research. Int J Microcirc ClinExp. 1988;7(4):327-345.
12. Long DS, Smith ML, Pries AR, Ley K, Damiano ER.Microviscometry reveals reduced blood viscosityand altered shear rate and shear stress profiles inmicrovessels after hemodilution. Proc Natl AcadSci U S A. 2004;101(27):10060-10065.
13. Frommhold D, Ludwig A, Bixel MG, et al. Sialyl-transferase ST3Gal-IV controls CXCR2-mediatedfirm leukocyte arrest during inflammation. J ExpMed. 2008;205(6):1435-1446.
14. Zederbauer M, Furtmuller PG, Ganster B,Moguilevsky N, Obinger C. The vinyl-sulfoniumbond in human myeloperoxidase: impact on com-pound I formation and reduction by halides andthiocyanate. Biochem Biophys Res Commun.2007;356(2):450-456.
15. Kessenbrock K, Frohlich L, Sixt M, et al. Protein-ase 3 and neutrophil elastase enhance inflamma-tion in mice by inactivating antiinflammatory pro-granulin. J Clin Invest. 2008;118(7):2438-2447.
16. Mitchison TJ, Cramer LP. Actin-based cell motilityand cell locomotion. Cell. 1996;84(3):371-379.
17. Xu J, Wang F, Van Keymeulen A, et al. Divergentsignals and cytoskeletal assemblies regulate self-organizing polarity in neutrophils. Cell.2003;114(2):201-214.
18. Zaslaver A, Feniger-Barish R, Ben-Baruch A.Actin filaments are involved in the regulation oftrafficking of two closely related chemokine re-ceptors, CXCR1 and CXCR2. J Immunol. 2001;166(2):1272-1284.
19. Willeke T, Scharffetter-Kochanek K, Gaehtgens P,Walzog B. A role for beta2 integrin CD11/CD18-mediated tyrosine signaling in extravasation ofhuman polymorphonuclear neutrophils. Biorheol-ogy. 2001;38(2-3):89-100.
20. Andrews PC, Krinsky NI. The reductive cleavageof myeloperoxidase in half, producing enzymi-cally active hemi-myeloperoxidase. J Biol Chem.1981;256(9):4211-4218.
21. Matthijsen RA, Huugen D, Hoebers NT, et al.Myeloperoxidase is critically involved in theinduction of organ damage after renal ischemiareperfusion. Am J Pathol. 2007;171(6):1743-1752.
22. Frommhold D, Mannigel I, Schymeinsky J, et al.Spleen tyrosine kinase Syk is critical for sus-tained leukocyte adhesion during inflammation invivo. BMC Immunol. 2007;8:31.
23. Ghitescu L, Fixman A. Surface charge distributionon the endothelial cell of liver sinusoids. J CellBiol. 1984;99(2):639-647.
24. Haldenby KA, Chappell DC, Winlove CP,Parker KH, Firth JA. Focal and regional variationsin the composition of the glycocalyx of large ves-sel endothelium. J Vasc Res. 1994;31(1):2-9
25. Klinger M, Kramarz S, Wendycz D, Kopec W. Dif-ferent erythrocyte and platelet surface electriccharge in various types of glomerulonephritis.Nephrol Dial Transplant. 1997;12(4):707-712.
26. Janoff A, Zweifach BW. Adhesion and Emigrationof Leukocytes Produced by Cationic Proteins ofLysosomes. Science. 1964;144:1456-1458.
27. Fehr J, Jacob HS. In vitro granulocyte adherenceand in vivo margination: two associatedcomplement-dependent functions. Studies basedon the acute neutropenia of filtration leukophore-sis. J Exp Med. 1977;146(3):641-652.
28. Gallin JI, Durocher JR, Kaplan AP. Interaction ofleukocyte chemotactic factors with the cell sur-face. I. Chemotactic factor-induced changes inhuman granulocyte surface charge. J Clin Invest.1975;55(5):967-974.
29. Gallin JI. Degranulating stimuli decrease theneagative surface charge and increase the adhe-siveness of human neutrophils. J Clin Invest.1980;65(2):298-306.
30. Schaack TM, Takeuchi A, Spilberg I, Persellin RH.Alteration of polymorphonuclear leukocyte sur-face charge by endogenous and exogenous che-motactic factors. Inflammation. 1980;4(1):37-44.
31. Constantinescu AA, Vink H, Spaan JA. Endothe-lial cell glycocalyx modulates immobilization ofleukocytes at the endothelial surface. ArteriosclerThromb Vasc Biol. 2003;23(9):1541-1547.
32. Woodman RC, Johnston B, Hickey MJ, Teoh D,Reinhardt P, et al. The functional paradox ofCD43 in leukocyte recruitment: a study usingCD43-deficient mice. J Exp Med. 1998;188(11):2181-2186.
33. Baldus S, Rudolph V, Roiss M, Ito WD, Rudolph TK,et al. Heparins increase endothelial nitric oxidebioavailability by liberating vessel-immobilizedmyeloperoxidase. Circulation. 2006;113(15):1871-1878.
34. Reitsma S, Slaaf DW, Vink H, van Zandvoort MA,oude Egbrink MG. The endothelial glycocalyx:composition, functions, and visualization.Pflugers Arch. 2007;454(3):345-359.
35. Peterson MW, Stone P, Shasby DM. Cationicneutrophil proteins increase transendothelial al-bumin movement. J Appl Physiol. 1987;62(4):1521-1530.
36. Dull RO, Dinavahi R, Schwartz L, Humphries DE,Berry D, et al. Lung endothelial heparan sulfates
MYELOPEROXIDASE ATTRACTS NEUTROPHILS 1357BLOOD, 27 JANUARY 2011 � VOLUME 117, NUMBER 4
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom

mediate cationic peptide-induced barrier dysfunc-tion: a new role for the glycocalyx. Am J PhysiolLung Cell Mol Physiol. 2003;285(5):L986-L995.
37. Engelhardt B, Wolburg H. Mini-review: Transen-dothelial migration of leukocytes: through thefront door or around the side of the house? EurJ Immunol. 2004;34(11):2955-2963.
38. Johansson MW, Patarroyo M, Oberg F, Siegbahn A,Nilsson K. Myeloperoxidase mediates cell adhe-sion via the alpha M beta 2 integrin Mac-1,CD11b/CD18. J Cell Sci. 1997;110 (Pt 9):1133-1139.
39. Lau D, Mollnau H, Eiserich JP, et al. Myeloperoxi-
dase mediates neutrophil activation by associa-tion with CD11b/CD18 integrins. Proc Natl AcadSci U S A. 2005;102(2):431-436.
40. Cai TQ, Wright SD. Human leukocyte elastase isan endogenous ligand for the integrin CR3CD11b/CD18, Mac-1, alpha M beta 2 and modu-lates polymorphonuclear leukocyte adhesion.J Exp Med. 1996;184(4):1213-1223.
41. David A, Kacher Y, Specks U, Aviram I. Interac-tion of proteinase 3 with CD11b/CD18 beta2 inte-grin on the cell membrane of human neutrophils.J Leukoc Biol. 2003;74(4):551-557.
42. Asako H, Kurose I, Wolf RE, Granger DN.
Mechanisms of lactoferrin-induced leukocyte-endothelial cell adhesion in postcapillary venules.Microcirculation. 1994;1(1):27-34.
43. Morgan DM, Larvin VL, Pearson JD. Biochemicalcharacterisation of polycation-induced cytotoxic-ity to human vascular endothelial cells. J Cell Sci.1989;94(Pt 3):553-559.
44. Daphna EM, Michaela S, Eynat P, Irit A, Rimon S.Association of myeloperoxidase with heparin: oxi-dative inactivation of proteins on the surface ofendothelial cells by the bound enzyme. Mol CellBiochem. 1998;183(1-2):55-61.
1358 KLINKE et al BLOOD, 27 JANUARY 2011 � VOLUME 117, NUMBER 4
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom

online October 27, 2010 originally publisheddoi:10.1182/blood-2010-05-284513
2011 117: 1350-1358
Schumacher, Thomas Meinertz, Markus Sperandio and Stephan BaldusFurtmüller, Peter Heeringa, Karsten Sydow, Hans-Jürgen Duchstein, Heimo Ehmke, UdoHans-Joachim Paust, Christine Schröder, Daniel Benten, Denise Lau, Katalin Szocs, Paul G. Anna Klinke, Claudia Nussbaum, Lukas Kubala, Kai Friedrichs, Tanja K. Rudolph, Volker Rudolph, Myeloperoxidase attracts neutrophils by physical forces
http://www.bloodjournal.org/content/117/4/1350.full.htmlUpdated information and services can be found at:
(554 articles)Phagocytes, Granulocytes, and Myelopoiesis Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society
For personal use only.on June 16, 2016. by guest www.bloodjournal.orgFrom