montanide® isa 720 vaccines: quality control of emulsions, stability of formulated antigens, and...
TRANSCRIPT

Vaccine 23 (2005) 2530–2539
Montanide® ISA 720 vaccines: quality control of emulsions,stability of formulated antigens, and comparative
immunogenicity of vaccine formulations
Aaron P. Miles∗, Holly A. McClellan, Kelly M. Rausch, Daming Zhu, Michael D. Whitmore1,Sanjay Singh, Laura B. Martin, Yimin Wu, Birgitte K. Giersing2, Anthony W. Stowers3,
Carole A. Long, Allan SaulMalaria Vaccine Development Branch, National Institute of Allergy and Infectious Diseases, National Institutes of Health,
5640 Fishers Lane, Twinbrook I Room 1118, Rockville, MD 20852, USA
Received 22 June 2004; received in revised form 27 August 2004; accepted 30 August 2004Available online 30 October 2004
Abstract
al animals part, to thef storage ofI wered -injectiond hat meetst storageaP
K
1
huba
T
R
A
20)ad-De-
ater-n atn-imaland
ines
ct ofring
re-table
0d
Montanide® ISA 720 is an experimental adjuvant, formulated as water-in-oil emulsions, that induces high antibody titers in severpecies. It has been used in human vaccine trials with malaria and HIV vaccines. The heightened response is likely due, inormation of a depot at the injection site. However, post-formulation modifications were seen with seven proteins tested duringSA 720 formulations at 37◦C for 1 week and two proteins stored longer at 4◦C. Potency studies in mice, in which the stored vaccinesiluted into placebo emulsions for appropriate dosing, indicated that this instability could lead to loss of immunogenicity in the postepot, limiting the allowable storage time of preformed vaccines. We describe point-of-injection formulation for ISA 720 vaccines t
he requirement for in vitro stability. For preformed vaccines, addition of glycine or glycylglycine prevented antigen modification ont 37◦C, providing a potential way of stabilizing antigen/ISA 720 formulations for in vitro storage and the post-injection depot.ublished by Elsevier Ltd.
eywords:Montanide ISA 720; Adjuvant; Vaccine; Recombinant protein
. Introduction
Alum salts are the most widely used adjuvants in licenseduman vaccines. The need for more effective adjuvants forse in vaccines against diseases such as malaria is made cleary the generally inadequate immune responses to syntheticnd recombinant malarial antigens seen with the use of alum
∗ Corresponding author. Tel.: +1 301 435 2335; fax: +1 301 480 1962.E-mail address:[email protected] (A.P. Miles).
1 Present address: Amgen Inc., One Amgen Center Drive, M/S 30-1-A,housand Oaks, CA 91320, USA.2 Present address: Malaria Vaccine Initiative, 6290 Montrose Road,ockville, MD 20852, USA.3 Present address: CSL Limited, 45 Poplar Road, Parkville, Vic. 3052,ustralia.
[1–7]. One such adjuvant is Montanide ISA 720 (ISA 7(SEPPIC, Inc., Fairfield, NJ), an experimental squalenejuvant that contains a mannide monooleate emulsifier.signed for use in humans, this adjuvant forms stable win-oil droplets intended to give slow release of antigethe injection site[8]. It has been shown to elicit higher atibody titers than many other adjuvants in several anspecies[9], and has been used in limited human Phase 1Phase 2 trials with experimental malaria and HIV vacc[10–13,15].
Herbert suggested that the enhanced adjuvant effea water-in-oil emulsion could be explained by considethe depot of emulsion to be inert and capable of slowlyleasing antigen over a long period of time[14]. This effecimplies that the antigen in the emulsion must remain st
264-410X/$ – see front matter. Published by Elsevier Ltd.oi:10.1016/j.vaccine.2004.08.049

A.P. Miles et al. / Vaccine 23 (2005) 2530–2539 2531
for extended periods of time while in contact with all of thecomponents of the emulsion at body temperature (37◦C).
In two human Phase 1 vaccine trials with malaria antigensformulated in Montanide ISA 720, a loss of potency occurredfollowing formulation. In one case, the decreased potencywas found prior to vaccination and resulted in reformulationof one of the components[11]. In the other case, a vaccine trialwith thePlasmodium falciparumapical membrane antigen1 (AMA1), the loss of potency was not detected until aftervaccination commenced and the trial was compromised (Saulet al., in press). These problems could have been avoided byan efficient point-of-injection formulation method.
In this paper, we describe a convenient method that re-producibly achieves the 1�m droplet size recommended bythe manufacturer[8,16]. We also describe modifications toall six proteins tested after storage formulated with ISA 720at 37◦C, and describe methods for preventing these from oc-curring by the addition of stabilizers in the emulsion.
2. Materials and methods
2.1. Formulations
AMA1-C1 [17], a 1:1 mixture of two 62-kDa recombinanta ariapa con-c S,1 -t 20( 0 mLa steels ss to anO i In-t ndst e-b mul-sl 20p btaina
d inP sur-f edaI umeo forA isf siont iza-t
totalv texG L)
in 2 mL borosilicate glass vials (Catalog # 223683, Wheaton,Millville, NJ) for 30 min at maximum speed at room temper-ature. During vortexing, it was critical that the vials were heldupright.
Using this vortex method, emulsions were prepared con-taining bovine serum albumin (BSA, Pierce Biotechnol-ogy Inc., Rockford, IL), AMA1-FVO[17] (a componentof AMA1-C1), Pvs25H[19], the homologue of Pfs25 fromPlasmodium vivaxexpressed inSaccharomyces cerevisiae,and MSP142-FVO [20], a bacterially-expressed and refoldedprotein based on the 42-kDa carboxyl-terminal portion ofMSP1 fromP. falciparum, with 9 or 150�g/mL antigen inPBS/ISA 720 (30:70 aqueous to oil based on volume). Theseproteins were chosen so that comparisons could be made bothin terms of antigen recoveries from extractions from vaccineformulations and stability of the various proteins within theemulsions.
For antigen stabilization studies, Pfs25/ISA 720 mix-tures (Pfs25 at 9 and 150�g/mL) containing one of thefollowing additives at the indicated concentrations werevortexed as described above and stored at 37◦C for upto 3 weeks in the presence or absence of nitrogen: buty-lated hydroxytoluene (BHT), 1.36 mM; butylated hydrox-yanisole (BHA), 1.67 mM; (±)-�-tocopherol (Vitamin E),1.16 mM; cysteamine hydrochloride (CH), 17.6 mM;N-a y-c ).A ecyls ) atd
2
stri-b par-t dis-p A).I dis-p oil( ide8 sizea tot suit-a rcol5
2W
thea 0%( ina uga-t e-o sis byS
llelic forms of apical membrane antigen 1 from the malarasiteP. falciparum, AMA1-FVO and AMA1-3D7, wasseptically homogenized at 6000 rpm at a final vaccineentration of 40�g/mL in phosphate-buffered saline [PB55 mM NaCl, 1 mM KH2PO4, 3 mM Na2HPO·7H2O (Spec
rum Chemical Mfg. Corp., Gardena, CA)] and ISA 730:70 aqueous to oil based on volume) at a volume of 20t room temperature for 6 min using a 400 mL stainlessealed chamber assembly with a 20 mm× 145 mm stainlesteel saw tooth rotor stator generator probe attachedmni Mixer-ES homogenizer (Catalog # ES-115, Omn
ernational, Warrenton, VA). The manufacturer recommehat this formulation ratio be on a w/w basis. Our volumased ratios, however, consistently give rise to stable eions containing droplets with an average size of 1�m. Di-utions of this formulation were prepared in PBS/ISA 7lacebo emulsion, prepared in the same manner, to oppropriate antigen concentrations for immunizations.
Pfs25[18], a 20-kDa recombinant protein expresseichia pastorisbased on the 25-kDa zygote and ookinete
ace protein ofP. falciparum, was aseptically homogenizt 6000 rpm at a final concentration of 9�g/mL in PBS and
SA 720 (30:70 aqueous to oil based on volume) at a volf 200 mL at room temperature for 6 min as describedMA1. After storage at 37◦C for 8 weeks, dilutions of th
ormulation were prepared in PBS/ISA 720 placebo emulo obtain appropriate antigen concentrations for immunions.
For vortexed preparations, vaccine components in aolume of 1 mL were aseptically vortexed (Daigger Vorenie 2 vortexer, A. Daigger & Co., Inc., Vernon Hills, I
cetyl-l-cysteine (NALC), 12.3 mM; glycine, 267 mM; glylglycine, 267 mM (Sigma–Aldrich Co., St. Louis, MOntigens were extracted and analyzed by sodium dodulfate-polyacrylamide gel electrophoresis (SDS–PAGEays 0, 7, and 21 as described below.
.2. Droplet size analysis
Vaccine emulsions were analyzed for droplet size diution using a Mastersizer 2000 laser diffraction-basedicle size analyzer with an attached Hydro 2000S sampleersion unit (Malvern Instruments Inc., Southborough, M
SA 720 emulsions form water-in-oil droplets, which areersed in an oil phase consisting of Marcol 5 paraffinExxon Co., U.S.A., Houston, TX) and 2% (v/v) Montan0 surfactant (SEPPIC, Inc.) flowing through the particlenalyzer. Approximately 200�L per emulsion was added
he dispersion unit to obtain laser obscuration in a rangeble for each reading. The system was flushed with Maparaffin oil between all readings.
.3. Antigen extraction from emulsions, SDS–PAGE,estern blot, and N-terminal sequencing
Antigens were extracted from vaccine emulsions withddition of benzyl alcohol to a final concentration of 1v/v) followed by vortexing at room temperature for 20 mt maximum speed (Daigger Vortex Genie 2) and centrif
ion for 10 min at 16,100×g. The antigen-containing aquus phases were then removed and set aside for analyDS–PAGE.

2532 A.P. Miles et al. / Vaccine 23 (2005) 2530–2539
SDS–PAGE was performed on 4-20% gradient polyacry-lamide Tris–glycine–SDS gels using an XCell SureLock Ele-crophoresis Mini-Cell apparatus (Invitrogen Corp., Carlsbad,CA). Gels were run at 30 mA (constant) for approximately50 min and stained with Coomassie Blue R-350 (AmershamBiosciences Corp., Piscataway, NJ) or silver-stained as de-scribed[21]. For reducing conditions, samples were mixedwith dl-dithiothreitol (DTT, Sigma Chemical Co., St Louis,MO) at a final concentration of 50 mM prior to electrophore-sis.
Amino acid sequencing was performed by automated Ed-man degradation on samples separated by SDS–PAGE asabove and transferred to a polyvinylidene difluoride (PVDF)membrane (Invitrogen Corp.). Protein bands were visualizedwith Coomassie Blue R-350 and then isolated for sequencing.
A Western blot on nitrocellulose (Invitrogen Corp.) wasblocked in 3% (w/v) skim milk (Difco, Detroit, MI) in Tris-buffered saline (TBS), and the membrane was probed with amonoclonal antibody (mAb) that recognizes the 6×histidinetag on the C-terminus of Pfs25 (anti-pentaHis mAb, Qiagen,Inc., Valencia, CA). After washing in TBS/0.05% Tween 20(Sigma–Aldrich Co.), the membrane was incubated with goatanti-mouse immunoglobulin G (IgG) conjugated to alkalinephosphatase (Kirkegaard and Perry Labs, Gaithersburg, MD),washed, and developed with 5-bromo-4-chloro-3-indolyl-p andP con-s ed atd
2
ncew thea rovalp sixg ivedt t of0 rm 4f ndaryi n sixg ivedt t of0 lyd orf xingf eeks im-m -l ysp
d byE 4,Do t
protein in coating buffer (15 mM Na2CO3, 35 mM NaHCO3,pH 9.6) per well, washed with Tris-buffered saline-Tween20 [TBS-T, 25 mM Tris base, 137 mM NaCl, 3 mM KCl(Quality Biological, Inc., Gaithersburg, MD), 0.1% Tween 20(Sigma Chemical Co.)], and blocked with 5% (w/v) skim milk(Difco) in TBS at room temperature for two hours. Serumsamples diluted in 0.1% BSA, 0.05% Tween 20–TBS were in-cubated in the coated wells in triplicate for 2 h in a humidifiedchamber at room temperature. The plates were washed, incu-bated with a 1:1000 dilution of goat anti-mouse IgG conju-gated to alkaline phosphatase for 2 h at room temperature, andwashed again. One hundred microliters of substrate solution(p-nitrophenyl phosphate, Sigma Diagnostics, Inc., St. Louis,MO) was added per well, the plates were incubated for 20 minat room temperature, 25�L of stop solution (5 N sodium hy-droxide) was added to each well, and absorbance was readat 405 nm using a SPECTRAmax 340PC microplate spec-trophotometer (Molecular Devices, Sunnyvale, CA). Eachtest plate included a reference serum standard, which was se-rially diluted in duplicate in order to make a standard curve.Using the standard curve, the absorbances of individual testsera were converted to antibody units (SOFTmax PRO ver-sion 3, Molecular Devices). One antibody unit was defined asthe antibody concentration giving an optical density (O.D.)at 405 nm of 1.0 in this standard ELISA.
3
3
ndsfa nic-i relyc eachf undt ute,t ortantp .W urev iblyg niM and7 , or2 rpm,f ntsi mul-s %o ro-d
3
byt d in
hosphate/nitroblue tetrazolium (BCIP/NBT, Kirkegaarderry Labs). Primary and secondary antibodies were retituted according to manufacturers’ instructions and usilutions of 1:250 (primary) or 1:3000 (secondary).
.4. Potency studies
All animal potency studies were performed in accordaith National Institutes of Health guidelines and underuspices of an Animal Care and Use Committee approtocol. The AMA1 potency study was carried out inroups of ten female BALB/c mice. Each animal rece
wo 250�L intraperitoneal immunizations 28 days apar.01, 0.03, or 0.1�g of AMA1 in ISA 720 that was eitheade fresh (reference) or freshly diluted after storage at◦C
or 14 months. Sera were collected 14 days post-secommunization. The Pfs25 potency study was carried out iroups of eight female BALB/c mice. Each animal rece
wo 400�L intraperitoneal immunizations 28 days apar.24, 0.8, or 2.4�g of Pfs25 in ISA 720 that was either freshiluted after storage at 37◦C for 8 weeks (stored vaccine),
reshly prepared (reference vaccine) by aseptically vorteollowed by diluting as described above. After the 8-wtorage period, formulations to be used for the secondunizations were placed at 4◦C until use so that all formu
ations spent 8 weeks at 37◦C. Sera were collected 14 daost-secondary immunization.
Serum antibodies to AMA1 and Pfs25 were assayeLISA. Flat-bottom 96-well microtiter plates (Immulonynex Technology Inc., Chantilly, VA) were coated at 4◦Cvernight with 100�L of a 1�g/mL solution of recombinan
. Results
.1. Droplet size of vaccine formulations
The manufacturer of ISA 720 (SEPPIC, Inc.) recommeormulations with a droplet size of approximately 1�m di-meter as the optimal size for stability and immunoge
ty. Larger droplets may cause the emulsion to prematuollapse. We, therefore, sought to achieve this size forormulation, whether vortexed or homogenized. We fohat mixture volume, container size, revolutions per minime, and presence or absence of protein were each imparameters in reproducibly forming stable 1-�m emulsionshen vortexing, 2 mL glass vials containing 1 mL of mixt
ortexed upright at maximum speed for 30 min reproducave 0.5�m droplets. For homogenization with the Omixer ES, volumes of 9 and 20 mL in the 50 mL chamber5 mL in the 400 mL chamber homogenized at 8000 rpm00 mL in the 400 mL chamber homogenized at 6000
or three times 2 min cycles (with brief swirling of conten-between to ensure homogeneity) gave reproducible eification (Table 1). Volumes lower than approximately 18f the chamber volume, lower rpm, or fewer cycles all puced larger diameter droplets (data not shown).
.2. Antigen extraction
Following destabilization of the ISA 720 emulsionshe addition of benzyl alcohol, antigens were recovere

A.P. Miles et al. / Vaccine 23 (2005) 2530–2539 2533
Table 1Volume mean diameter andD(v, 0.9)a (in �m) of formulations obtained byvortexing or homogenizing representativeb samples in triplicate
Volume meandiameter
D(v, 0.9)
Vortexingc
Sample 1 0.511 1.019Sample 2 0.514 1.016Sample 3 0.553 1.092
Mean 0.526 1.042Standard deviation 0.023 0.043
Homogenizingb
50 mL chamber, 9 mL mixture 1 1.324 2.08650 mL chamber, 9 mL mixture 2 1.260 2.00750 mL chamber, 9 mL mixture 3 1.331 2.137
Mean 1.305 2.077Standard deviation 0.039 0.066
400 mL chamber, 200 mL mixture 1 1.084 1.672400 mL chamber, 200 mL mixture 2 1.079 1.673400 mL chamber, 200 mL mixture 3 1.074 1.663
Mean 1.079 1.669Standard deviation 0.005 0.006
a A percentile reading giving a droplet size below which 90% of the samplelies.
b For simplicity, results are not shown for a 50 mL chamber containinga 20 mL mixture or a 400 mL chamber containing a 75 mL mixture. Theseresults were consistent with those shown here.
c Formulation procedures were optimized with respect to mixture volume,container size, revolutions per minute, time, cycles (for homogenization) andpresence or absence of protein.
the aqueous layer. For some proteins (e.g., Pvs25, Pfs25, andBSA) there was close to a quantitative recovery, while for oth-ers (AMA1-FVO, AMA1-3D7, MSP142-FVO), recovery wasreproducibly lower (approximately 50–75%) and dependedupon the protein analyzed. For proteins giving low yields,we invariably saw precipitates in one or more of the resultinglayers. The precipitates were protein aggregates, and resolu-bilization with subsequent analysis by SDS–PAGE allowedvisualization (data not shown).
3.3. Stability of formulated antigens at 4◦C
The stability of recombinant AMA1 extracted from ISA720 following storage at 4◦C for 9 months was tested bySDS–PAGE of extracted antigen from two formulations: 40and 160�g/mL. Four to twenty percent gradient polyacry-lamide Tris–glycine–SDS gels were loaded with extracts andAMA1 standards. Non-reduced gels were run, followed byCoomassie Blue staining (Fig. 1). Significant modificationsof AMA1 took place upon storage at 4◦C for 9 months.Monomer band broadening is evident, along with multimerformation (especially dimers). The modifications of AMA1in ISA 720 stored at 4◦C for 9 months are similar to thoseseen with Pfs25 after storage for one to 3 weeks at 37◦C (seebelow).
Fig. 1. Non-reducing Coomassie Blue-stained SDS–PAGE analysis of ex-tracts of 40 and 160�g/mL formulations of AMA1-C1 in ISA 720 followingstorage at 4◦C for 9 months. Lanes 1–5 are 5, 4, 3, 2, or 1�g (respectively)reference AMA1-C1; lane 6, blank; lanes 7 and 8, duplicate 40�g/mL ex-tracts with approximately 0.9�g loaded per lane; lane 9, blank; lanes 10and 11, duplicate 160�g/mL extracts with approximately 3.4�g loaded perlane. Molecular weight standards are shown in kDa.
Pfs25 was extracted and analyzed by SDS–PAGE fromISA 720 formulations (2, 9, 35, and 150�g/mL) followingstorage at 4◦C for 3 weeks. Minor but detectible modificationwas present compared to bulk (unextracted) Pfs25 (data notshown).
3.4. Stability of formulated antigens at 37◦C
For testing stability at 37◦C, we chose vaccine formu-lations containing Pfs25, since these give nearly 100% re-coveries in the aqueous phase. Two 4–20% gradient poly-acrylamide Tris–glycine–SDS gels were loaded identicallywith similar amounts of Pfs25 extracted from 2, 9, 35, or150�g/mL ISA 720 formulations following storage at 37◦Cfor 3 weeks or unextracted Pfs25 (as reference). They werethen run non-reduced (Fig. 2A) and reduced (Fig. 2B) fol-lowed by silver staining. The modifications seen were pro-tein concentration dependent. Protein extracted from lowerconcentrations (i.e., 2 and 9�g/mL) showed broadening ofthe main band, and an increase in electrophoretic mobility,with minimal formation of multimers, while higher concen-trations (35 and 150�g/mL) underwent substantial formationof multimers with a corresponding reduction in the amount ofmonomer recovered. These multimers are not disulfide bonddependent, as they are insensitive to reduction by 50 mMD cedm thee oona d( am inal6 ctedf e
TT in the presence of SDS, conditions that fully reduonomers (Fig. 2B). These changes were not caused by
xtraction procedure itself, as samples from ISA 720 sfter emulsification (Fig. 2C) were identical to unextractereference) Pfs25 (seeFig. 2A and B). Western blotting withonoclonal antibody (mAb) directed against the C-termxhistidine tag and N-terminal sequencing of Pfs25 extra
rom the 9 and 150�g/mL formulations showed that th

2534 A.P. Miles et al. / Vaccine 23 (2005) 2530–2539
Fig. 2. SDS–PAGE of extracts of various concentrations of Pfs25 in ISA 720 following storage at 37◦C for 3 weeks. (A) Non-reducing and (B) reducing 4–20%gradient polyacrylamide Tris–glycine–SDS silver-stained gels. Reference lanes are 88 ng of Pfs25 (bulk antigen, not emulsified with ISA 720). Approximately88 ng total protein is loaded in all lanes. The concentrations shown are those of Pfs25 as originally formulated in ISA 720. Note that the multimers (visible inboth gels) are not sensitive to reduction by DTT at a concentration (50 mM) that fully reduces the 11 cystines present in Pfs25 (shown by an apparent massshift of +5 kDa). These gels show that the modifications take place in a concentration-dependent manner, with lower protein concentrations tending towardbroadening of the main band, and higher concentrations favoring multimer formation. (C) Pfs25 extracted from each formulated concentration shortly afteremulsification, showing appearance of Pfs25 following extraction at the start of the study. Lanes 1, 3, 5, and 7, non-reduced extracts of 150, 35, 9, and2�g/mLformulations, respectively. Lanes 2, 4, 6, and 8, reduced extracts of 150, 35, 9, and 2�g/mL formulations, respectively. Pfs25 loads: lanes 1 and 2, 7.5�g; lanes3 and 4, 1.75�g; lanes 5 and 6, 0.45�g; lanes 7 and 8, 0.1�g. Molecular weight standards are shown in kDa.
modified monomers and the multimers had intact N- and C-termini. Pfs25 extracted from formulations stored for 1 weekat 37◦C had significant changes, but less than those seen after3 weeks (data not shown). No Pfs25 could be extracted fromemulsions stored for 8 weeks at 37◦C.
As shown inFig. 3, BSA, AMA1-FVO, MSP142-FVO, orPvs25H extracted from 9�g/mL ISA 720 emulsions storedat 37◦C for 1 week showed a similar pattern of modification(band broadening and multimerization). Of particular note isthe absence of AMA1 and Pvs25H fragments in the reducedstored sample lanes (arrows inFig. 3).
3.5. Stability of formulated Pfs25 at 37◦C withantioxidants or anti-alkylating agents
Squalene is sensitive to oxidation, and SEPPIC recom-mends that ISA 720 be stored under nitrogen. Therefore,we investigated whether the modification of Pfs25 at 9 and150�g/mL stored at 37◦C for 3 weeks could be prevented
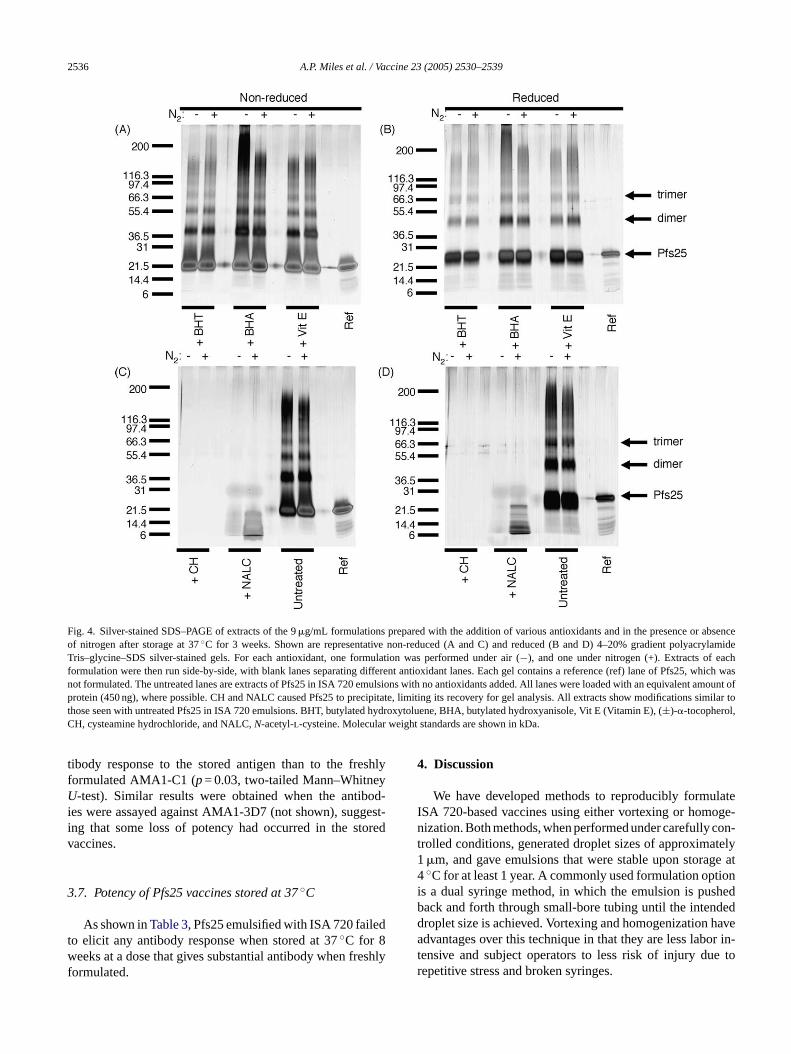
by antioxidants in the emulsion. We tested butylated hy-droxytoluene (BHT) at 1.36 mM, butylated hydroxyanisole(BHA) used at 1.67 mM, (±)-�-tocopherol at 1.16 mM, cys-teamine hydrochloride (CH) at 17.6 mM, andN-acetyl-l-cysteine (NALC) at 12.3 mM as examples of water- and oil-soluble antioxidants in concentrations that were not likely toresult in regulatory hurdles[22]. The samples were storedunder either nitrogen or air. Samples were extracted afterstorage for 0, 1, 7, and 21 days and were electrophoresedon 4–20% gradient polyacrylamide Tris–glycine–SDS gels,either non-reduced or reduced, followed by Coomassie Blueor silver staining. All samples treated with any antioxidantstored under nitrogen or air were indistinguishable from un-treated (reference vaccine) samples, indicating that at the con-centrations used, no antioxidant was able to protect Pfs25from modification.Fig. 4 shows representative SDS–PAGEgels with Pfs25 extracted from 9�g/mL formulations after 3weeks’ storage at 37◦C. Consistent results were obtained forother time points and for proteins formulated at 150�g/mL.

A.P. Miles et al. / Vaccine 23 (2005) 2530–2539 2535
Fig. 3. Composite image of silver-stained SDS–PAGE gels of extracts of9�g/mL individual formulations of four proteins in ISA 720. Each protein isshown in four consecutive lanes, with the first two lanes non-reduced (N) andthe last two lanes reduced (R). Lanes alternate between controls (C, samplesnot stored) and samples stored at 37◦C for 1 week (S). Equal volumes ofall extracts (12.5�l of 250�l total, corresponding to approximately 450 ng,depending on protein recovery) were loaded in each lane. For each protein,band broadening and multimerization not seen on day 0 are evident after 1week. The AMA1 and Pvs25H samples contain a proportion of moleculeswith cleavages in the polypeptide chains that result in lower molecular weightfragments on reducing gels (arrows). These are absent in the stored samples.Molecular weight standards are shown in kDa.
Storage under nitrogen had little or no impact on the degreeof modification of the protein. The inability of the antiox-idants to protect Pfs25 in ISA 720, as well as the lack ofprotection offered by storage under nitrogen, suggested thatthe observed modifications were not oxidative in nature, butrather a result of one or more reactive components of ISA720.
The pattern of modification of the proteins was consistentwith inter- and intramolecular cross-linking, potentially via
amino groups. To examine this possibility, glycine and gly-cylglycine were added to Pfs25 in ISA 720 formulations toact as scavengers. Glycine at an isotonic concentration (2%,267 mM) was chosen. ISA 720 formulations that contained 9or 150�g/mL Pfs25 and either glycine or glycylglycine wereaseptically prepared by vortexing. These were stored at 37◦Cunder air for 3 weeks prior to analysis. The samples were thenrun on 4–20% gradient polyacrylamide Tris–glycine–SDSgels, non-reduced or reduced, and silver-stained or stainedwith Coomassie Blue (Fig. 5). In both the 9 and 150�g/mLformulations, glycine or glycylglycine protected the protein,eliminating the broadening of the main Pfs25 band and pre-venting multimer formation.
3.6. Potency of AMA1 vaccines stored at 4◦C
A potency study of AMA1 in ISA 720 in mice was per-formed in an attempt to detect any immunological impactof the modifications detected on long-term storage at 4◦C.Six groups of ten female BALB/c mice each received two250�L intraperitoneal immunizations 28 days apart of 0.01,0.03, or 0.1�g of AMA1 in ISA 720 that was either madefresh (reference) or freshly diluted after storage at 4◦C for 14months. The dose range used had previously been found togive a sensitive dose–response curve in female BALB/c mice.S ationa thei gs
yf anti-A osts n-
Table 2Two weeks post-secondary immunization anti-AMA1-FVO ELISAa values of indiv )vaccines
Dose (0.01�g) Dose (0.03�g)
Dilution Source Dilution Source
Reference (T= 0) Stored (T= 14 months) Reference (T= 0) Store )
<67 <67 4080 236<67 <67 2720 179<67 <67 1524 412<67 <67 2470 112<67 <67 9450 <67<67 <67 247 1037<67 <67 233 101<67 <67 <67 117<67 <67 1540 <67<67 <67 2590 164
G rN 231 +
A at 40�g/mL givea
anti-AM of 1:35,000i ction of
Mb ± standard error GMb ± standard erro/A N/A 1,009 + 630/−388
MA1-C1 (equal amounts of FVO and 3D7 proteins) was formulatedppropriate dosing.a ELISA results are shown as units compared to a reference mouse
n the standard ELISA. Numbers below 67 represent the level of deteb GM, geometric mean.
era were collected 14 days post-secondary immuniznd antibody titers were then determined by ELISA with
mmunizing antigens (AMA1-FVO and AMA1-3D7) coatineparate plates.
As shown inTable 2, AMA1-C1/ISA 720, whether freshlormulated or stored for 14 months, gave increasingMA1-FVO antibody as a function of dose. At the mensitive dose (0.03�g), there was a significantly lower a
idual mice receiving diluted reference (fresh,T= 0) or stored (T= 14 months
Dose (0.1�g)
Dilution Source
d (T= 14 months) Reference (T= 0) Stored (T= 14 months
0 67 4,90033,600 27,65016,750 20,35015,650 6,32021,850 28,15020,600 33,85027,850 27,15044,150 29,55028,750 15,350
0 22,850 15,300
GMb ± standard error123/−80 10,443 + 8564/−4705 13,483 + 5082/−3691
in ISA 720, and dilutions were carried out into placebo emulsions to
A1 serum that has 35,000 units since it gives an O.D. = 1 at a dilutionthe assay and indicate no detectable antibody response.
2536 A.P. Miles et al. / Vaccine 23 (2005) 2530–2539
Fig. 4. Silver-stained SDS–PAGE of extracts of the 9�g/mL formulations prepared with the addition of various antioxidants and in the presence or absenceof nitrogen after storage at 37◦C for 3 weeks. Shown are representative non-reduced (A and C) and reduced (B and D) 4–20% gradient polyacrylamideTris–glycine–SDS silver-stained gels. For each antioxidant, one formulation was performed under air (−), and one under nitrogen (+). Extracts of eachformulation were then run side-by-side, with blank lanes separating different antioxidant lanes. Each gel contains a reference (ref) lane of Pfs25,which wasnot formulated. The untreated lanes are extracts of Pfs25 in ISA 720 emulsions with no antioxidants added. All lanes were loaded with an equivalent amount ofprotein (450 ng), where possible. CH and NALC caused Pfs25 to precipitate, limiting its recovery for gel analysis. All extracts show modifications similar tothose seen with untreated Pfs25 in ISA 720 emulsions. BHT, butylated hydroxytoluene, BHA, butylated hydroxyanisole, Vit E (Vitamin E), (±)-�-tocopherol,CH, cysteamine hydrochloride, and NALC,N-acetyl-l-cysteine. Molecular weight standards are shown in kDa.
tibody response to the stored antigen than to the freshlyformulated AMA1-C1 (p= 0.03, two-tailed Mann–WhitneyU-test). Similar results were obtained when the antibod-ies were assayed against AMA1-3D7 (not shown), suggest-ing that some loss of potency had occurred in the storedvaccines.
3.7. Potency of Pfs25 vaccines stored at 37◦C
As shown inTable 3, Pfs25 emulsified with ISA 720 failedto elicit any antibody response when stored at 37◦C for 8weeks at a dose that gives substantial antibody when freshlyformulated.
4. Discussion
We have developed methods to reproducibly formulateISA 720-based vaccines using either vortexing or homoge-nization. Both methods, when performed under carefully con-trolled conditions, generated droplet sizes of approximately1�m, and gave emulsions that were stable upon storage at4◦C for at least 1 year. A commonly used formulation optionis a dual syringe method, in which the emulsion is pushedback and forth through small-bore tubing until the intendeddroplet size is achieved. Vortexing and homogenization haveadvantages over this technique in that they are less labor in-tensive and subject operators to less risk of injury due torepetitive stress and broken syringes.

A.P. Miles et al. / Vaccine 23 (2005) 2530–2539 2537
Fig. 5. SDS–PAGE of extracts of 9 and 150�g/mL formulations of Pfs25 in ISA 720 containing glycine or glycylglycine following storage at 37◦C for 3weeks. Gels A and C are non-reduced, and B and D reduced. A and B are silver-stained, and C and D are stained with Coomassie Blue. Gels A and B containapproximately 450 ng of (left to right) unextracted Pfs25 (reference), Pfs25 + 267 mM glycine in ISA 720 extract, and Pfs25 + 267 mM glycylglycine in ISA720 extract. Blank lanes separate sample lanes. Gels C and D contain approximately 5�g of protein per lane, following the same loading as above. In boththe 9 and 150�g/mL formulations, the presence of either glycine or glycylglycine eliminates the broadening of the main Pfs25 band and prevents multimerformation. Slight main band smearing is evident and is probably a result of the extraction procedure (compared to Fig. 2C, extractions performed soonafterformulation). Molecular weight standards are shown in kDa.
In order to examine the stability of antigens, methodsmust be developed for routinely extracting them from theemulsions. The procedure described allowed near quantita-tive recovery of some, but not all proteins. The reasons forthe variable recovery were not clear, but recovery of smallproteins was often nearly 100%, while that of large proteinswas lower.
By using this extraction technique, we found that all pro-teins tested underwent rapid modification with time in storageat 37◦C. Similar changes were seen with two protein prepa-rations upon storage at 4◦C, although at a much slower ratethan with storage at 37◦C.
Extensive characterization of ISA 720-based vaccines hasnot been described previously. Without adequate vaccine test-
ing prior to use, clinical trials can potentially be compromiseddue to loss of potency. Indeed this is a risk with many experi-mental vaccines if they are not shown to be sufficiently stable.We suggest that at least a minimum set of assays be performedon clinical ISA 720-based vaccine batches, whether formu-lated and stored, or produced for point-of-injection (extempo-raneous) use. These assays include visual appearance, dropletsize, integrity by reduced and non-reduced SDS–PAGE, iden-tity by mAb recognition (preferably conformational), and po-tency. Ideally pre-clinical batches should also be analyzedwhere possible.
Pfs25 has 11 disulfide bonds. Probably because of thisextensive cross-linking, the non-reduced protein migratessubstantially faster on SDS–PAGE gels than the reduced

2538 A.P. Miles et al. / Vaccine 23 (2005) 2530–2539
Table 3Two weeks post-secondary immunization anti-Pfs25 ELISAa values of individual mice receiving diluted reference (fresh,T= 0) or stored (T= 8 weeks) vaccines
Dose (0.24�g) Dose (0.8�g) Dose (2.4�g)
Dilution Source Dilution Source Dilution Source
Reference (T= 0) Stored (T= 8 weeks) Reference (T= 0) Stored (T= 8 weeks) Referenceb (T= 0) Stored (T= 8 weeks)
<49 <49 <49 <49 9,020 <49<49 <49 <49 <49 8,980 <49<49 <49 <49 <49 12,700 <49<49 <49 <49 <49 7,820 <49166 <49 <49 <49 1,980 <49<49 <49 358 <49 1,770 <49<49 <49 <49 <49 24,440 <49<49 <49 20,605 <49 21,960 <49
Pfs25 was formulated at 9�g/mL in ISA 720, and dilutions were carried out into placebo emulsions to give appropriate dosing.a ELISA results are shown as units compared to a reference mouse anti-Pfs25 serum that has 25,000 units since it gives an O.D. = 1 at a dilution of 1:25,000
in the standard ELISA. Numbers below 50 represent the level of detection of the assay and indicate no detectable antibody response.b Geometric mean, 7897; Standard error, +3273/−2314.
form. After storage of emulsions at 37◦C, the monomermigrated faster on reducing SDS–PAGE gels. On non-reducing SDS–PAGE gels, the effect was not as pronounced.These changes in mobility and the presence of non-reducibleoligomers suggested that the proteins developed intra- and in-termolecular cross-links when emulsified with ISA 720. By8 weeks, the modification was so extensive that the proteincould no longer be recovered from the emulsions. A potencystudy in mice was performed using this modified materialdiluted into placebo emulsions to achieve appropriate dos-ing, and the results showed that the vaccines had lost allimmunogenicity. Since the antigen could not be extractedfrom the emulsion, this result might not seem surprising.Nonetheless, since the protein remained somewhere in theemulsion, the possibility existed that vaccine immunogenic-ity was intact. The lost immunogenicity shown in this studymade clear that the phenomenon observed by SDS–PAGEcarried a significant immunological impact. Several highlyreactive molecules, including malonyl dialdehyde, capableof cross linking proteins are generated from the oxidation ofsqualene[23]. Addition of either water-soluble or oil-solubleantioxidants and removal of oxygen failed to prevent modi-fication, suggesting that these molecules were present priorto dispensing the formulation. Furthermore, as the degree ofmodification, especially of the monomeric form of Pfs25, wasi elyt ation.T net viat
lbu-ma gt fromA od-i ac-t withc disul-fi d are
therefore, released under conditions of reducing SDS elec-trophoresis. These smaller fragments were missing under re-ducing SDS–PAGE of AMA1 and Pvs25H stored at 37◦C,indicating that they had been crosslinked to the remainder ofthe respective protein (Fig. 3).
Although not practical for use in the field, extemporaneousformulations are a short-term alternative to pre-formulationfollowed by extended cold storage. This means of formulat-ing can allow for more rapid testing of vaccine candidates inclinical trials, and many of the resources that would other-wise go toward establishing long-term stability can be spared.Nevertheless, characterization of the vaccines as we have de-scribed should still take place. Formulating in this way caneliminate modifications that would otherwise take place dur-ing storage, but stability could nonetheless remain an issuebecause vaccine persistence at the injection site for extendedperiods, and thus an opportunity for modifications, may stilllead to decreased immune responses, both in terms of peakantibody or T cell response and the length of the response.Addition of glycine or glycylglycine to recombinant protein-based vaccines that contain ISA 720 may provide a generalsolution.
Acknowledgments
arkG N-t andC are,a hiss itia-t
R
acy.
nversely proportional to the protein concentration, it is likhat the reactive species are present at low concentrhis is consistent with the ability of glycine or glycylglyci
o prevent protein modification, presumably by reactingheir amino groups with the modifying species.
Similar modifications were seen with bovine serum ain, with theP. falciparumproteins AMA1 and MSP142,nd with Pvs25H, theP. vivaxortholog of Pfs25, showin
hat this is not a protein specific phenomenon. The dataMA1 and Pvs25H provide support for the idea that the m
fications involve intramolecular crosslinking. As manufured, both of these proteins contain some moleculesleavages in the polypeptide chains. The fragments arede bonded to other points in the respective chains, an
The authors wish to express their gratitude to Marfield (Research Technologies Branch, NIAID) for
erminal sequencing, Lynn Lambert, Brian Keegan,heryl Kothe for expert animal immunizations and cnd Andrew Orcutt and Olga Muratova for ELISAs. Ttudy was supported in part by the Malaria Vaccine Inive, PATH.
eferences
[1] Ballou WR, Hoffman SL, Sherwood JA, et al. Safety and efficof a recombinant DNAPlasmodium falciparumsporozoite vaccineLancet 1987;1(8545):1277–81.

A.P. Miles et al. / Vaccine 23 (2005) 2530–2539 2539
[2] Amador R, Moreno A, Valero V, et al. The first field trials of thechemically synthesized malaria vaccine SPf66: safety, immunogenic-ity and protectivity. Vaccine 1992;10(3):179–84.
[3] Alonso PL, Smith T, Schellenberg JR, et al. Randomised trial ofefficacy of SPf66 vaccine againstPlasmodium falciparummalariain children in southern Tanzania. Lancet 1994;344(8931):1175–81.
[4] D’Alessandro U, Leach A, Drakeley CJ, et al. Efficacytrial of malaria vaccine SPf66 in Gambian infants. Lancet1995;346(8973):462–7.
[5] Keitel WA, Kester KE, Atmar RL, et al. Phase I trial of two re-combinant vaccines containing the 19 kd carboxy terminal fragmentof Plasmodium falciparummerozoite surface protein 1 (msp-1(19))and T helper epitopes of tetanus toxoid. Vaccine 1999;18(5/6):531–9.
[6] Kashala O, Amador R, Valero MV, et al. Safety, tolerability andimmunogenicity of new formulations of thePlasmodium falciparummalaria peptide vaccine SPf66 combined with the immunologicaladjuvant QS-21. Vaccine 2002;20(17/18):2263–77.
[7] Kaslow DC. Transmission-blocking vaccines. Chem Immunol2002;80:287–307.
[8] Aucouturier J, Dupuis L, Deville S, Ascarateil S, Ganne V. Mon-tanide ISA 720 and 51: a new generation of water in oil emulsions asadjuvants for human vaccines. Expert Rev Vaccines 2002;1(1):111–8.
[9] Pye D, Vandenberg KL, Dyer SL, et al. Selection of an adju-vant for vaccination with the malaria antigen, MSA-2. Vaccine1997;15(9):1017–23.
[10] Lawrence GW, Saul A, Giddy AJ, Kemp R, Pye D. Phase I trial inhumans of an oil-based adjuvant SEPPIC MONTANIDE ISA 720.
[ rialsISA
[ tagetsrial in
[13] Toledo H, Baly A, Castro O, et al. A phase I clinical trial ofa multi-epitope polypeptide TAB9 combined with Montanide ISA720 adjuvant in non-HIV-1 infected human volunteers. Vaccine2001;19(30):4328–36.
[14] Herbert WJ. The mode of action of mineral-oil emulsion adjuvantson antibody production in mice. Immunology 1968;14(3):301–18.
[15] Lawrence G, Cheng QQ, Reed C, et al. Effect of vaccinationwith 3 recombinant asexual-stage malaria antigens on initial growthrates ofPlasmodium falciparumin non-immune volunteers. Vaccine2000;18(18):1925–31.
[16] Aucouturier J, Dupuis L, Ganne V. Adjuvants designed for veterinaryand human vaccines. Vaccine 2001;19(17–19):2666–72.
[17] Kennedy MC, Wang J, Zhang Y, et al. In vitro studies with recom-binantPlasmodium falciparumapical membrane antigen 1 (AMA1):production and activity of an AMA1 vaccine and generation of amultiallelic response. Infect Immun 2002;70(12):6948–60.
[18] Zou L, Miles AP, Wang J, Stowers AW. Expression of malariatransmission-blocking vaccine antigen Pfs25 inPichia pastorisforuse in human clinical trials. Vaccine 2003;21(15):1650–7.
[19] Hisaeda H, Stowers AW, Tsuboi T, et al. Antibodies to malariavaccine candidates Pvs25 and Pvs28 completely block the abil-ity of Plasmodium vivaxto infect mosquitoes. Infect Immun2000;68(12):6618–23.
[20] Singh S, Kennedy MC, Long CA, Saul AJ, Miller LH, Stow-ers AW. Biochemical and immunological characterization ofbacterially expressed and refoldedPlasmodium falciparum42-kilodalton C-terminal merozoite surface protein 1. Infect Immun2003;71(12):6766–74.
[21] Sambrook J, Fritsch EF, Maniatis T. In: Molecular cloning: a labo-ratory manual, vols. 1–3. New York: Cold Spring Harbor Laboratory
[ ed.aceu-
[ e ofunsat-s. J
Vaccine 1997;15(2):176–8.11] Saul A, Lawrence G, Smillie A, et al. Human phase I vaccine t
of 3 recombinant asexual stage malaria antigens with Montanide720 adjuvant. Vaccine 1999;17:3145–59.
12] Genton B, Betuela I, Felger I, et al. A recombinant blood-smalaria vaccine reducesPlasmodium falciparumdensity and exerselective pressure on parasite populations in a phase 1–2b tPapua New Guinea. J Infect Dis 2002;185(6):820–7.
Press; 1989. p. 18.56.22] Kibbe A, editor. Handbook of pharmaceutical excipients. 3rd
London: American Pharmaceutical Association and the Pharmtical Press; 2000.
23] Rastrelli L, Passi S, Ippolito F, Vacca G, De Simone F. Ratdegradation of alpha-tocopherol, squalene, phenolics, and polyurated fatty acids in olive oil during different storage conditionAgric Food Chem 2002;50(20):5566–70.