monograp late miocene artiodactyles
TRANSCRIPT

THE LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN TYPE LOCALITY OF THE DHOK PATHAN FORMATION, THE MIDDLE SIWALIKS, PAKISTAN
Muhammad Akbar Khan1* Muhammad Akhtar2 and Mehboob Iqbal3 1Zoology Department, Government College University, Faisalabad
2Zoology Department, Punjab University, Quid-e-Azam Campus, Lahore 3Zoology Department, Government Science College, Wahdat Road, Lahore
PAKISTAN JOURNAL OF ZOOLOGY SUPPLEMENTARY SERIES Number 10, December 2010
Pages 1-87
Published by
THE ZOOLOGICAL SOCIETY OF PAKISTAN

Pakistan Journal of Zoology
Supplement Series
Number 10, pp. 1-87, December 2010
THE LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN TYPE LOCALITY OF THE DHOK PATHAN FORMATION, THE MIDDLE SIWALIKS, PAKISTAN
MUHAMMAD AKBAR KHAN* MUHAMMAD AKHTAR AND MEHBOOB IQBAL
Published by
THE ZOOLOGICAL SOCIETY OF PAKISTAN

Pakistan J. Zool. Suppl. Ser., No.10, pp. 1-87, 2010.
The Late Miocene Artiodactyls in the Dhok Pathan Type Locality of the Dhok Pathan Formation, the Middle Siwaliks, Pakistan
Muhammad Akbar Khan1* Muhammad Akhtar2 and Mehboob Iqbal3 1Zoology Department, Government College University, Faisalabad 2Zoology Department, Punjab University, Quid-e-Azam Campus, Lahore 3Zoology Department, Government Science College, Wahdat Road, Lahore
CONTENTS
Abstract ..................................................................................................................................................................... 1 Introduction .............................................................................................................................................................. 1 Materials and methods .............................................................................................................................................. 7 Systematic palaeontology ........................................................................................................................................... 8 Palaeoenvironment and palaeobiogeography of the Dhok Pathan artiodactyls............................................................. 76 Conclusions ............................................................................................................................................................. 79 References ............................................................................................................................................................... 80 Abstract.– The late Miocene artiodactyls are presented on the basis of description of abundant material from the Dhok Pathan type locality of the Dhok Pathan Formation, the Middle Siwaliks and the remains increasingly indicate taxonomic diversity. Quantitatively, the taxa of bovids are the most predominant. But cervids, tragulids, giraffids and suids are approximately as common as each other at Dhok Pathan. Comparative morphometric features of the late Miocene artiodactyls from the locality are studied in here. More than 130 artiodactyl fossil specimens including skulls, horn cores, isolated teeth and fragments of maxillae or mandibles were studied. These fossils document twenty three artiodactyl species belonging to fifteen genera and five families. This assemblage includes two new taxa: Miotragocerus large sp. and ?Hydaspitherium sp. boselaphines, antilopines, reduncines, cervids, tragulids, giraffids and suids are abundant in the locality whereas tragelaphines and alcelaphines are absent. Biogeographically, the late Miocene artiodactyls indicate strong relationships with Eurasian and African late Miocene sites. In this study, palaeoenvironmental interpretations of the type locality provide important evidence regarding late Miocene palaeoenvironments. Most of these taxa indicate a predominance of woodland to savannah habitat during the deposition of the Dhok Pathan Formation. Key words: Mammals, Late Miocene, Systematics, Bovids, cervids, tragulids, giraffids.
INTRODUCTION
The late Miocene was a time of active speciation and origination of higher taxa in mammals (Table I) and it is a time period that is crucial to our understanding the evolution of the extant artiodactyls. The Dhok Pathan type locality has yielded several species of artiodactyls, perissodactyls, the oldest Siwalik cercopithecoid, carnivores, proboscideans and rodents (Table I), clearly distinct from that found in the upper part of the Nagri Formation and its equivalents. The artiodactyls comprise almost 50% of the total faunal elements (Fig. 1). Artiodactyls are found abundantly in the late Miocene sites of the Siwaliks, Africa and Eurasia (Vrba, 2000). The fossil record of Artiodactyla from
the Siwaliks (Table I) is generally very good in Dhok Pathan Mammalian Fauna
Artiodactyla46%
Perissodactyla 12%
Proboscidea17%
Rodentia3%
Carnivora20%
Cercopithecidae2%
Fig. 1. Percentage of species represented in the Dhok Pathan mammalian fauna.
comparison with that of other parts of the world and the late Miocene locality of the Dhok Pathan has yielded one of the richest fossil artiodactyl faunas from south Asia (Lydekker, 1878; Matthew, 1929; Colbert, 1935; Pilgrim, 1926a,b, 1937, 1939;

M.A. KHAN ET AL.
2
Thomas, 1984, Pickford, 1988; Akhtar, 1992; Gentry, 1994; Haile-Selassie et al., 2004; Bibi, 2007; Bibi et al., 2009). Many Siwalik artiodactyls Table I.- Mammalian faunas of Dhok Pathan. Many
species are under taxanomic revision. Artiodactyla Bovidae Tragoportax browni T. salmontanus T. perimense T. punjabicus Miotragocerus gluten Proleptobos birmanicus Selenoportax vexillarius S. lydekkeri Pachyportax latidens P. nagrii P. giganteus Eotragus sp. Gazella lydekkeri G. padriensis Taurotragus latidens Perimia falconeri Elaschistoceros khauristanensis Giraffidae Vishnutherium iravaticum Bramatherium
perimense Hydaspitherium megacephalum H. grande H. magnum Giraffa punjabiensis Cervidae Rucervus simplicidens Cervus triplidens C. sivalensis C. punjabiensis C. rewati Tragulidae Dorcabune anthracotherioides D. nagrii Dorcatherium majus D. minus Anthracotheriidae Chocromeryx silistrense Merycopotamus
dissimilis Suidae Tetraconodon magnus T. mirabilis Propotamochoerus uliginosus P. hysudricus Hippopotamodon sivalensis Hyosus punjabiensis H. tenuis Sivahyus hollandi Hippohyus lydekkeri H. grandis Sus comes S. adolescens Perissodactyla Equidae Hipparion antelopinum H. theobaldi H. perimense H. nagriensis Rhinocerotidae Aceratherium perimense A. Lydekkeri Chilotherium intermedium C. blanfordi Subchilotherium intermedium Alicornops sp. Brachypotherium perimense Chalicotheriidae Proboscidea Dinotherium indicum D. angustidens (?) Trilophodon hasnotensis Tetralophodon
falconeri T. punjabiensis Rhynchotherium
chinjiensis Synconolophus dhokpathanensis S. propathanensis
S. corrugatus S. ptychodus S. hasnoti Anancus perimensis Stegolophodon latidens S. cautleyi Stegodon bombifrons S. cliftii S. elephantoides Cercopithecidae Cercopithecus hasnoti Macacus sivalensis Rodentia Rhizomys sivalensis Rhizomys sp. Hystrix sivalensis Carnivora Arctamphicyon lydekkeri Indarctos punjabiensis Promellivora punjabiensis Enhydriodon falconeri Sivaonyx bathygnathus Vishnuictis salmontanus Ictitherium sivalense I. indicum Lycyaena macrostoma L. macrostoma-cinayaki Crocuta carnifex C. gigantean C. gigantean-latro C. mordax Mellivorodon palaeindicus, Acluropsis anneclens Paramachacrodus pilgrimi P. indicus Propontosmilus sivalensis, Felis sp.
are represented by cranial and postcranial material and show strong relationship with Eurasian and African artiodactyls. Fossil remains have been known in the area since the nineteenth century and large mammals have been the object of several monographies (Lydekker, 1878; Matthew, 1929; Colbert, 1935; Pilgrim, 1926a,b, 1937, 1939; Pickford, 1988). The late Miocene artiodactyls probably descended from Lophiomerycidae in the late middle Eocene of south Asia (Guo et al., 2000). The early-middle Eocene Kuldana Formation in northern Pakistan has yielded several forms of bunolo-phodont artiodactyls, all of which are endemic to the Indian subcontinent (Metais et al., 2004). The Oligocene fossils from the Bugti Hills of Pakistan may also document the first appearance and subsequent radiation of giraffes and bovids, two groups that dominate later Neogene and modern herbivore faunas. In Pakistan true bovids may appear as early as 19.5 Ma, and the family is certainly present at 18.5 Ma. Since they are not known earlier elsewhere, their appearance in southern Asia may document the beginning of this very important radiation (Barry et al., 2005). This period of time may represent a window of early representatives of giraffids, cervids and tragulids in

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
3
Pakistan. Eotragus, an early boselaphine described from the Kamlial Formation of the Siwaliks is considered the oldest bovid of the world (Solounias et al., 2005). It is apparent that the artiodactyl fauna from the Dhok Pathan is dominated by bovids (Table I). Nevertheless, there are other species of the artiodactyls in the fauna tragulids, cervids, giraffids and suids. Out of bovids, boselaphines are abundant in the late Miocene of the Dhok Pathan. The Siwalik was inhabited by various boselaphines during the late Miocene and many new forms of boselaphines appeared, while their diversification was reduced in the Early Pliocene (Khan et al., 2009a). Moya-Sola (1983) defines two generic or suprageneric groups of boselaphines comprising, the medium sized Siwalik boselaphines genera Miotragocerus Stromer, 1928 and Tragoportax Pilgrim, 1937 and the large sized Siwalik boselaphines genera Selenoportax and Pachyportax as well as allied forms such as Austroportax latifrons from the middle Miocene of Europe. Tragoportax were among the widest spread bovids in the late Miocene of the Siwaliks (Pilgrim, 1937, 1939; Thomas, 1984; Akhtar, 1992). The genus is also widespread in Oriental and southeastern Europe (Geraads et al., 2003; Kostopoulos, 2009). They appear for the first time in the middle Miocene but they are best known from the Turolian wherein, alongside Gazella, they are considered as the most widespread of all bovids. This group went extinct around the end of the Miocene (Gentry, 1999). Tragoportax sp. is present in the Turolian of the Graeco-Irano-Afganian province as well as in the Siwaliks (Solounias, 1981; Bernor, 1986; Kohler, 1987; Gentry and Heizmann, 1996; Kostopoulos, 2005, 2009). Tragoportax cyrenaicus at its type locality (France) is a morphologically advanced species of the very late Miocene (Geraads, 1989: 790). Allied species to T. cyrenaicus are found in the presumed early Pliocene locality of Langebaanweg, south Africa (Tragoportax acrae) and in the late Miocene of the Siwaliks (Tragoportax salmontanus; Gentry, 1999). Tragoportax salmontanus resembles closely the late Miocene Siwalik species Prostrepsiceros vinayaki. Pilgrim (1937, 1939) mentioned variety of the medium sized boselaphines in the Siwaliks and
some merely based on the minor variations (e.g. Sivaceros gradiens). Thomas (1984) referred to this group all the specimens that came from the Nagri and the Dhok Pathan formations, and allotted the name Tragocerina. More recently, Bibi et al. (2009) assigned the name ‘Tragoportacini’ for the Miotragocerus-Tragoportax complex. Akhtar (1992) recovered a huge material of Tragoportacini from the type locality Dhok Pathan and named it Tragocerus punjabiensis, Tragocerus browni, and Miotragocerus dhokpathanensis Akhtar. According to Akhtar (1992) that Miotragocerus dhokpathanensis is larger than Miotragocerus gluten. The species is illustrated by a single specimen opisthocranium and the characteristics of the opisthocranium allow us to compare it to Tragoportax. The skull shows all the main features of the Tragoportax (see systematics) and it is perhaps a synonym of Tragoportax. Furthermore, many remains of various taxa have been recovered from the type locality during the last four decades and erroneously linked to Tragoportax, Tragocerus and Miotragocerus. The revised description is proposed on the basis of the new finds (Appendix I) and the already recovered material from the type locality Dhok Pathan. This paper describes and interprets the entire collection of artiodactyls from the Dhok Pathan type locality (Appendix II), recovered during 1980 and 2009, and at present in the Palaeontology Laboratory of the Zoology Department of the Punjab University, Lahore, Pakistan and in the Zoology Department of GC University, in Faisalabad, Pakistan. The present paper deals with a taxonomic investigation of the late Miocene artiodactyl fauna of Dhok Pathan, the Middle Siwaliks, Pakistan. The late Miocene artiodactyls have a great significance in the World’s palaeontological history, as the period is marked by a great faunal diversity with a high degree of completeness. The aim of this work is to expose as brief as possible the richness and diversity of the Dhok Pathan artiodactyls. The main aim of this study has been to provide the first complete documentation of the artiodactyls found in the Dhok Pathan type locality by tackling aspects of taxonomy, stratigraphy, palaeontology and paleo-ecology of the Siwaliks of Pakistan. An ecologically important group, the artiodactyls, was selected for

M.A. KHAN ET AL.
4
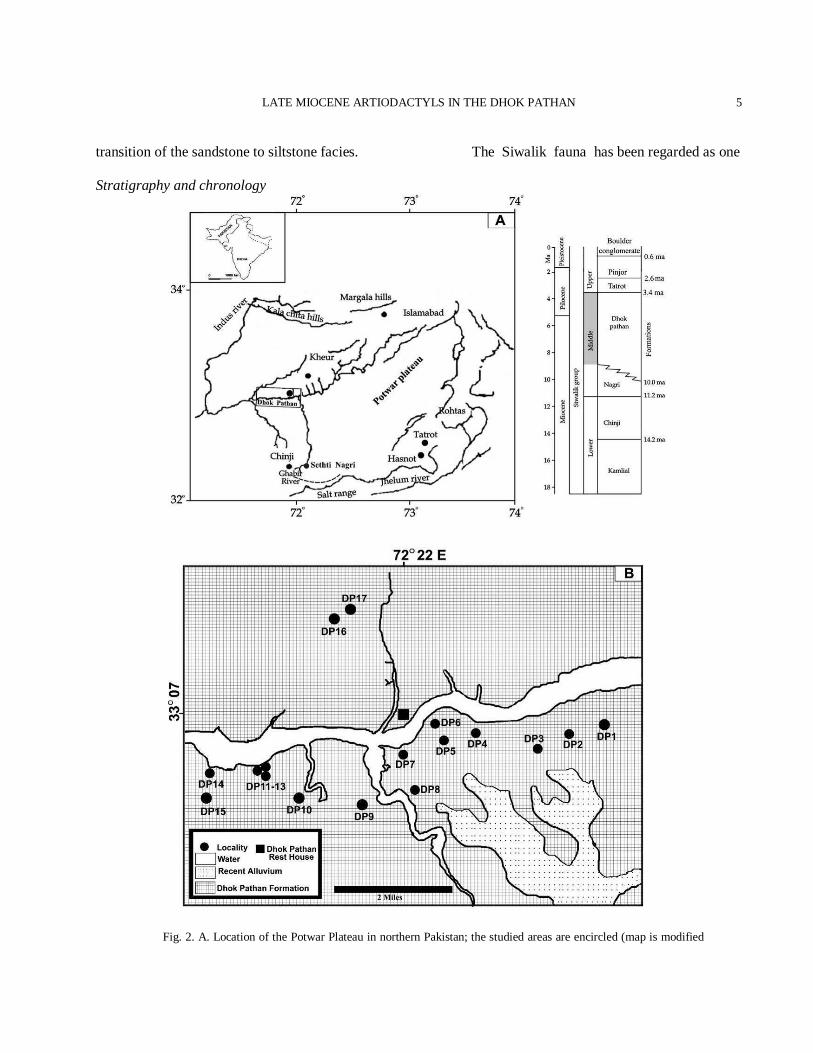
the study as the collected artiodactyl material presented notable diversity and thus could provide significant taxonomic, ecological, biostratigraphic and palaeoenvironmental information. Abbreviations Ca, Circa; Myr, Million years; Ma, Million years ago; MN, Mein Zones; GPTS, Geomagnetic Polarity Time Scale; GRTS, Geomagnetic Reversal Time Scale; DP1-DP17, Localities of Dhok Pathan from numbers1 to 17; AMNH, American Museum of Natural History; BMNH, British Museum of Natural History; PMNH, Pakistan Museum of Natural History; PUPC, Punjab University Palaeontological Collection, Lahore, Pakistan; PC-GCUF, Palaeontological Collection of GC University, Faisalabad, Punjab, Pakistan; P or p, Upper or lower premolar; M or m, upper or lower molar; GSI, Geological survey of India; GSP, Geological Survey of Pakistan; DP, Deciduous Premolar; W/L, Width/Length ratio; Fms/Fm, Formations/Formation; r, Right; l, Left; mm, Millimeters. Geography and geology The village Dhok Pathan (Lat. 33° 07' N: Long. 72° 14' E) is situated in the district Chakwal of northern Pakistan (Fig. 2A). The type locality is situated at Soan River about 75 km from Rawalpindi on Rawalpindi-Sargodha road and it is surrounded by extensive Neogene freshwater sedimentary rocks. The Dhok Pathan fossiliferous area is situated in the vicinity of the Dhok Pathan Rest House near the Soan River at an altitude of 1073 feet (Fig. 2B). The average thickness of the sequence around this area is about 497 m (Barry et al., 2002). Sites surrounding the village present an abundance of vertebrate fossils that represent almost all major Eutherian groups. The region of the Dhok Pathan exposes the most complete sequence of the Siwalik Group and yields a diversified assemblage of the Middle Siwalik faunas (Table I). The Dhok Pathan bed near the Dhok Kundrali is one of the most fossiliferous beds of the Siwaliks (Akhtar, 1992) and most of the material described in this paper is recovered from the Kundrali unit (DP9-13). This bed has yielded a large number of artiodactyl samples but these are not homogeneous. This is particularly important because much of the Dhok Pathan fauna of Pilgrim,
Brown, and Lewis apparently came from the vicinity of the bed (Barry et al., 2002). It is situated in the vicinity of the Dhok Pathan Rest House (Lat. 33° 07' N: Long. 72° 22' E) which is placed about at the bank of the Soan River near the village Dhok Pathan. Four new localities have been excavated and they are DP8, DP9, DP16 and DP17 (Fig. 2B). The geology of the Siwaliks has been well studied and documented by national and international researchers since the 18th century. The British and American workers did the pioneering work on the Siwalik geology and palaeontology (Lydekker, 1878; Colbert, 1935; Behrensmeyer and Tauxe, 1982; Quade and Cerling, 1995; Barry et al., 2002; Suresh et al., 2004). The stratigraphic sequence preserves a continuous record of the continental sedimentation (sediment thickness > 6 km at places) as well as an equally comparable continuous record of vertebrates, especially that of the mammals (Flynn et al., 1995; Flynn, 2003; Basu, 2004). The Middle Siwaliks (Nagri, Dhok Pathan formations) are dominantly arenaceous, consisting of multistoried coarse to medium-grained, blue-gray, massive sandstones (30 to > 60 m) with subordinate representation of clays, mudstones and siltstones. The Dhok Pathan type locality is characterized by the light colored sandstone with alternate clay and minor layers of conglomerates. The sediments have orange shale with less compacted gray sandstone bodies and red-brown mudstone with a few thin conglomerate interbeds. Sandstone and superposed red mudstone often form fining-upward couplets where the lower contact is erosional and lined with ripped-up clasts of the underlying mottled and red-brown clay stone. At a few places, thin crevasse-splay sheets, around 30 cm thick, clast-supported conglomerates occur. These conglomerate beds often contain unidentifiable bone and tooth fragments. Sandstone beds upsection gradually get thicker as well as multistoried. These substantially thicker, vertically stacked and laterally extensive individual gray sandstone units form a fining-upward sequence with thinner dull red to brown siltstones on top (Behrensmeyer and Tauxe, 1982). Varicolored, mottled, highly bioturbated paleosol horizons form distinct and laterally extensive units within the siltstone or at the
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
5
transition of the sandstone to siltstone facies. Stratigraphy and chronology
The Siwalik fauna has been regarded as one
Fig. 2. A. Location of the Potwar Plateau in northern Pakistan; the studied areas are encircled (map is modified
M.A. KHAN ET AL.
6
from Behrensmeyer and Barry, 2005 and the boundary dates are from Dennell et al., 2008 and Nanda, 2008). B. Simplified geology map of the Dhok Pathan area indicating the fossiliferous localities around the Dhok Pathan Rest House (DP – abbreviation for Dhok Pathan) from where the studied material is recovered (map is modified from Colbert, 1935).
C D
E F
G H
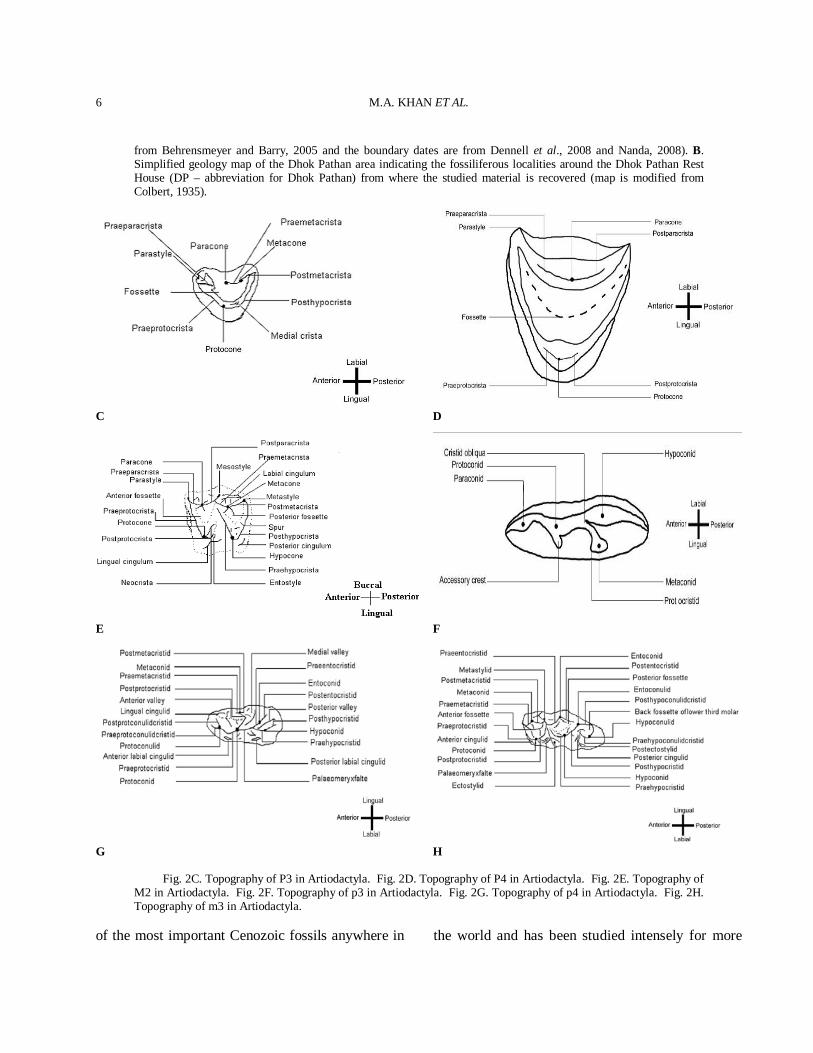
Fig. 2C. Topography of P3 in Artiodactyla. Fig. 2D. Topography of P4 in Artiodactyla. Fig. 2E. Topography of M2 in Artiodactyla. Fig. 2F. Topography of p3 in Artiodactyla. Fig. 2G. Topography of p4 in Artiodactyla. Fig. 2H. Topography of m3 in Artiodactyla.
of the most important Cenozoic fossils anywhere in the world and has been studied intensely for more

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
7
than a century. Several American and Pakistani scientists did post-independence stratigraphical work in the Siwaliks of Pakistan (Pilgrim, 1910, 1913; Colbert, 1935; Shah, 1977; 1980; Pilbeam et al., 1977; Stix, 1982; Behrensmeyer, 1987; Willis, 1993a, b; Behrensmeyer et al., 1995; Willis and Behrensmeyer, 1995). As a result, a fine-scale magnetostratigraphic work has emerged, supplemented by vertebrate faunal zone. Because the sediments are well suited for magnetostratigraphic studies, it has been possible to establish a regional chronostratigraphic framework that now spans the Early Miocene through the Pleistocene (Opdyke et al., 1979; Johnson G. D. et al., 1982; Tauxe and Opdyke, 1982; Johnson, N. M. et al., 1982, 1985; Kappelman, 1986; Friedman et al., 1992; Downing et al., 1993). Johnson N. et al. (1982, 1985) have shown that the Potwar Siwalik sediments range between 18.3 and 0.6 Ma. During this period only one major depositional hiatus has been identified (Opdyke et al., 1979). With reference to the Geomagnetic Reversal Time Scale of Berggern et al. (1985) the ages of the Miocene Potwar formations (Kamlial, Chinji, Nagri, and Dhok Pathan formations) and the Late Pliocene through Pleistocene formations are now fairly well understood. More recently, the geochronology of the Dhok Pathan Formation has been refined and a reliable chronostratigraphy has been established by geomagnetic polarity time scale. The magnetic polarity and stratigraphic dating has constrained the age of the Dhok Pathan Formation to between 10.1–ca. 3.5 Ma. The lower part of the Dhok Pathan Formation is dated 10.1-9.0 Ma and the upper part is considered to be dated at ca. 9.0-5.5 Ma (Cande and Kent, 1995; Barry et al., 2002). The Dhok Pathan Rest House section yielded an age of 8–6.5 Ma. Nevertheless, paleomagnetic and biostratigraphic correlations have yielded a precise geochronology of the latest Miocene/earliest Pliocene of the Dhok Pathan (Fig. 1A) (Pickford, 1988; Barry et al., 1982, 2002). Biostratigraphy The Dhok Pathan Formation faunas are mainly very similar to those of the underlying sediments; thus, the suids, the carnivores and the bovids continue virtually unchanged. Miotragocerus
and Tragoportax are represented by many species; Reduncini make their first appearance together with a species very similar to Prostrepsiceros vinayaki from which Thomas (1984) accepts the age of the late Miocene. The earliest reduncine (Kobus porrecticornis) records in Eurasia and in the Siwalik Formation of India and Pakistan also date to the late Miocene (Vrba, 1995). Miotragocerus and Tragoportax appeared in the Siwaliks during the latest middle Miocene and diversified during the late Miocene, before going extinct at the end of the Miocene. Selenoportax record indicates that it was present in the Siwaliks from the Late Middle Miocene to the Pliocene (Akhtar, 1992; Barry et al., 2002; Khan et al., 2009a). Nevertheless, Barry et al. (2002) recognized a maximum range of 10.3 to 7.9 Ma for this taxon. Qiu and Qiu (1995) list Selenoportax sp. from the Lufeng fauna (Chinese Miocene), age of 11.1–8.0 Ma (Steininger, 1999; Flynn and Qi, 1982). Pachyportax, on the other hand, is considered as a typical Late Miocene taxon. Pachyportax occurs in the Nagri and the Dhok Pathan zones of the Siwaliks, until Proamphibos replaces it soon after the start of the Tatrot. Hence, a Latest Miocene date around 7.0 Ma (Barry et al., 1991) would be considered as a possible date for a fauna containing P. latidens. Overall, the Dhok Pathan Formation faunas resemble those from the Turolian Land Mammal ‘Age’ as defined in Europe, North Africa and West Asia. There rather restricted time range seems to fall close to the boundary between the Vallesian and the Turolian Land Mammal ‘Ages’. So, the age of the Dhok Pathan, is considered as the late Miocene. Barry et al. (2002) suggested that the age of the Dhok Pathan was the late Miocene, however, the cervids, the bovids, the giraffids, and the suids suggest a late Miocene to early Pliocene age. The faunal association of the locality indicates an age to date between the late Miocene and early Pliocene (Fig. 2).
MATERIALS AND METHODS
The described material comprises skull, horn-cores, maxilla and mandible fragments, and isolated molars (Appendix II). The material includes the

M.A. KHAN ET AL.
8
newly collected specimens from the Dhok Pathan type locality during field trips that took place from 1980 to 2009 by the Palaeontology team of the Punjab University, Lahore, Pakistan. In addition some of the specimens used in this study already belonged to the collections of the Palaeontology Laboratory, University of the Punjab, Lahore, Pakistan and had been collected in the past decades. Almost all fossil specimens were found weathering out from or in situ within the light colored sandstone with alternate clay and orange shale. Fossils were generally very well preserved. The material came from 17 localities (Fig. 2B). The specimens excavated from these localities were generally in excellent condition, with little surface damage, often complete, and sometimes in articulation. Most specimens found on erosional surfaces were also well preserved, particularly those that had not been exposed for long, as on steep, actively eroding slopes. The spatial distribution of fossil material was non-random. Over 70% came from 6 localities of the Kundrali unit, the largest of that (localities DP11-13) had over 65 fossils on its surface. This pattern seems typical of the Siwaliks as earlier workers already noted that fossils were typically found in pockets in the Siwaliks (Gaur, 1987; Raza et al., 2002). Piercing instruments like chisels and geological hammers were employed for the excavation of partially embedded fossils. Careful measures were taken so as to prevent the fossils from disintegrating during excavation. Each specimen was wrapped with a cotton piece to avoid the shocks of transportation. Eventually the collected specimens were brought in the laboratory for taxonomic and morphological analysis. Clay and other hardly adjoined sedimentary particles were removed with the help of fine needles and brushes. Accidentally broken fragments of the specimens were rejoined by using gums and resins such as Magic Stone, Elfy, and Fixings etc. A hand lens was used for keen observation of very small and ambiguous morphological characters. The studied material is housed in the Palaeontology Laboratory of the Zoology Department of the Punjab University, in Lahore, Pakistan and in the Zoology Department of GC University, in Faisalabad, Pakistan. Comparisons of specimens were made with those at the Natural
History Museum, London (BMNH), the Natural History Museum, America (AMNH), the Geological Survey of Pakistan (GSP), the Geological Survey of India (GSI), the Palaeontological collection of Government College University, Faisalabad, Pakistan (PC-GCUF) and the specimens from the Palaeontology laboratory of the Zoology Department of the Punjab University, Lahore, Pakistan (PUPC). The specimens were catalogued by giving them a number which consists of the year of collection followed by a serial catalogue number (e.g. 83/261). Upper case letters stand for upper teeth and lower case letters for lower teeth. The selected photos of the studied taxa showing reflective morphometric features are given in the figures. The measurements of the horn-cores are taken at the base. The anteroposterior diameter (DAP) corresponds to the large diameter of the horn-core base and it may not be parallel to the sagittal plane. The mediolateral diameter (DT) is perpendicular to the DAP. Measurements of the studied teeth specimens were taken with a caliper as maximum lengths, widths and height at the occlusal level and are expressed in millimeters (mm). Tooth cusp terminology (Figs. 2C-H) follows the nomenclature of Gentry and Hooker (1988), and horn core terminology follows Pilgrim (1937, 1939) and Spassov and Geraads (2004). Taxonomic description Careful and extensive morphometric comparison led to the taxonomical identification of twenty three artiodactyl species. The identified artiodactyl species are listed in systematic order with information on holotype, diagnosis, differential diagnosis, geographic distribution, description, comparison, and discussion.
SYSTEMATIC PALAEONTOLOGY
Order Artiodactyla Owen, 1848 Family Bovidae Gray, 1821
Tribe Boselaphini Knottnerus-Meyer, 1907
Genus TRAGOPORTAX Pilgrim, 1937 Type species

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
9
Tragoportax salmontanus Pilgrim, 1937. Generic diagnosis (Pilgrim, 1937) Boselaphinae of moderate size; skull wide at the frontals and occipital; face bent down on the cranial axis; occiput long or moderately short; horn-cores rather short, with or without a slight twist, situated far apart on the frontals, but with their inner base expanded considerably so as to form a swelling on the frontals which sometimes bridges the entire interval between the horn-cores as an elevated belt falling abruptly to the rear; with prominent antero-internal and postero-external keels; keel axis very oblique to skull axis; horn-cores directed strongly backward and moderately or strongly outward, with or without an inward curve; cross section at base subtriangular, laterally compressed, compression increasing above the base, outer face between the keels weakly convex, third angle internal, about two thirds of the way from the anterior keel to the rear, rounded; supraorbital pits small, circular; temporal ridges very strong; frontal rugose; lachrymal fossa deep; basioccipital short, subtriangular, with or without a shallow median groove; supraoccipital exposed as a rectangular area on the upper surface of the skull; upper molars quadrate, moderately hypsodont, outer folds and ribs rather strong, median rib in posterior lobe weaker than in anterior lobe, entostyles present in all the molars. Generic diagnosis (Spassov and Geraads, 2004) Size generally large, approximately that of European Cervus elaphus. The postcornual fronto-parietal surface is a flat or slightly concave well defined depressed area, usually bordered laterally by well marked temporal ridges and caudally by a step leading to a slightly raised plateau. The basi-occipital has a longitudinal groove between the anterior and the posterior tuberosities, in the bottom of which often runs a weak sagittal crest. Adult male horn-cores are long and slender, usually curved backwards, with a triangular to subtriangular cross-section, well marked posterolateral keel and flattened lateral sides, but are less compressed than in Miotragocerus. Anterior rugosities growing downwards from the anterior keel at the basis of horn-cores are absent or weak, and usually do not extend onto the frontal. Demarcations (steps) on the
anterior keel are often found, but are few when present. Horn-cores have a heteronymous torsion (anti-clockwise on the right horn), so that the anterior keels first diverge in anterior view, but they re-approach towards the tips. The intercornual plateau is rather short antero-posteriorly, broad and almost rectangular between the horn-cores. The occipital is not much broader ventrally than dorsally, giving it a trapezoid (rather than triangular) outline. Teeth rather hypsodont; labial walls of upper teeth, and lingual ones of lower teeth with less accentuated ribs and styles than in Miotragocerus. Premolars relatively shorter than in Miotragocerus. P2 short relatively to P3, especially its anterior part, and parastyle curved backwards. P3 with lingually inflated hypocone. Metaconid of p3-p4 larger than in Miotragocerus, splayed lingually and T-shaped on p4, with an open anterior valley. Stratigraphic range Middle Siwaliks. Geographic distribution From the Vallesian/Turolian boundary (or lateVallesian?) to the end of the Turolian, perhaps earliest Pliocene in Africa. From southeastern Europe and the northern Paratethys region through Asia minor and the Middle East to Africa and the northern part of the Indian subcontinent (and possibly central Asia) (Pilgrim, 1937, 1939; Spassov and Geraads, 2004).
Tragoportax punjabicus (Pilgrim, 1910)
Type specimen Skull: GSI B486. Emended diagnosis (modified from Pilgrim 1937, 1939 and, Spassov and Geraads, 2004) A Tragoportax with moderately long, curved horn-cores with a large antero-posterior diameter, faintly twisted, slight torsion, cross-section triangular to subtriangular, well marked posterolateral and flattened sides. The postcornual fronto-parietal surface is a flat or slightly concave well defined depressed area, usually bordered laterally by well marked temporal ridges and caudally by a step leading to a slightly raised

M.A. KHAN ET AL.
10
plateau. The basi-occipital has a longitudinal groove between the anterior and the posterior tuberosities, in the bottom of which often runs a weak sagittal crest. Occipital is high and basioccipital is short. Premolar series is short, upper molars hypsodont with divergent styles and prominent median ribs. P2 has a distally sloping parastyle with a short parastyle-mesostyle part and bilobed lingually. P3 has a lingually inflated hypocone. The p4 has a strong paraconid, metaconid and entoconid. The entoconid is fused with the endostylid. The anterior valley of p4 is open. Studied material Skull: PUPC 86/149, opisthocranium. PUPC 86/150, opisthocranium. Horn-core: PUPC 86/264, broken at the apex, left side. Upper dentition: PUPC 83/743, isolated right P3. PUPC 87/327, isolated left P3. PUPC 00/21, isolated left P3. PUPC 00/89, right P4-M1. PUPC 83/671, isolated right M1. PUPC 87/677, isolated right M1. PUPC 83/673, isolated left M1. PUPC 83/30, isolated left M1. PUPC 83/676, isolated right M1. PUPC 83/674, isolated right M1. PUPC 97/13, isolated right M1. PUPC 00/80, right M1-2. PUPC 85/242, isolated right M2. PUPC 97/06, isolated left M2. PUPC 83/603, isolated right M2. PUPC 83/10, isolated left M2. PUPC 83/666, isolated right M2. PUPC 00/01, isolated left M2. PUPC 83/693, isolated right M2. PUPC 08/126, palatine parts of both maxillae with right P3-M3 and left P2-M3. PUPC 83/275, right maxillary fragment with P3-M3. PUPC 97/10, right maxillary fragment with M1-M3. PUPC 83/665, isolated left M3. PUPC 97/7, isolated right M3. PUPC 83/688, isolated right M3. Lower dentition: PUPC 83/712, isolated left p2. PUPC 97/12, left mandible fragment with p2-p4. PUPC 83/204, isolated left p3. PUPC 87/324, isolated left p3. PUPC 83/711, isolated left p4. PUPC 68/237, isolated left p4. PUPC 97/3, isolated right m1. PUPC 87/282, isolated left m1. PUPC 86/26, isolated left m1. PUPC 83/700, isolated right m1. PUPC 86/25, isolated left m2. PUPC 83/617, isolated left m2. PUPC 99/28, isolated right m2. PUPC 83/701, isolated left m2. PUPC 83/680, isolated left m3. PUPC 83/767, isolated left m3. PUPC 83/277, left mandible fragment with m3.
Description Skull PUPC 86/149 and PUPC 86/150 opisthocraniums are incomplete specimens having only the occipital region, right occipital condyle, paraoccipital process, parietals, and basisphenoid (Fig. 3). PUPC 86/150 opisthocranium (Fig. 3(2)) is more fragile than PUPC 86/149 (Fig. 3(1)). The facial part, horn-core, one occipital condyle and dentition are missing. The sutures separating the various bones are not visible. From the occipital foramen to the sphenoid, a continuous groove runs along the basioccipital, often with a weak sagittal crest in its middle. The groove is very prominent in PUPC 86/150 and the sagittal crest is getting weak towards the posterior tuberosities (Fig. 3(2)). The anterior tuberosities are fairly wide apart and not very large while the posterior tuberosities are large and expanded laterally. The parietal surface is a flat and slightly concave well defined depressed area, bordered laterally by well marked temporal ridges and caudally by a step leading to a slightly raised plateau. The temporal outer ridge runs back less obliquely and finally bends round by the side of the supraoccipital to join the lambdoid crest. The basioccipital is subtriangular in shape and the paraoccipital process points ventrally. The comparative measurements of the opisthocraniums are provided in Table II. Horn-core The horn-core is fragment and incomplete, lost upper part but the basal part is complete (Fig. 4). The horn-core is moderately long, compressed laterally. It is curved backward, weak torsion and faintly twisted. The lateral face of the horn-core is flat while the internal one is slightly convex. It has a prominent antero-internal keel and a rounded postero-external keel. The cross section of the horn-core at the base is triangular. The medio-lateral compression increases from the base to the top. Weak discontinuous longitudinal grooves run along the horn-core surface. The total length measured along the anterior face is 143 mm and along the posterior face is 140 mm. The transverse diameter at the base is 31 mm and antero-posterior diameter at the base is 53 mm. The mean index DT × 100/DAP is 58.4 at the base.

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
11
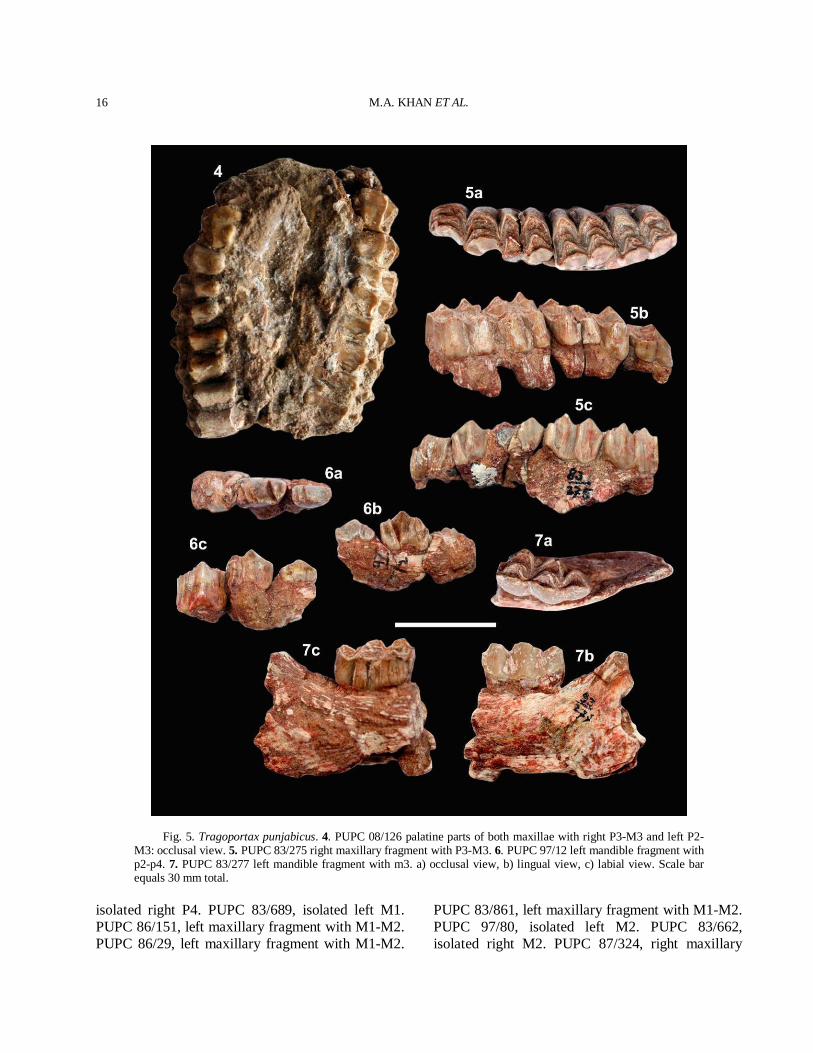
Upper dentition The premolar row is relatively shorter than the molar row. PUPC 08/126 is only one sample which shows complete cheek teeth series (Fig. 5(4)). The length of the premolar series is 32.5 mm and the length of the molar series is 52 mm in PUPC 08/126. P2 is similar to P3 but it is short to P3. P2
M.A. KHAN ET AL.
12
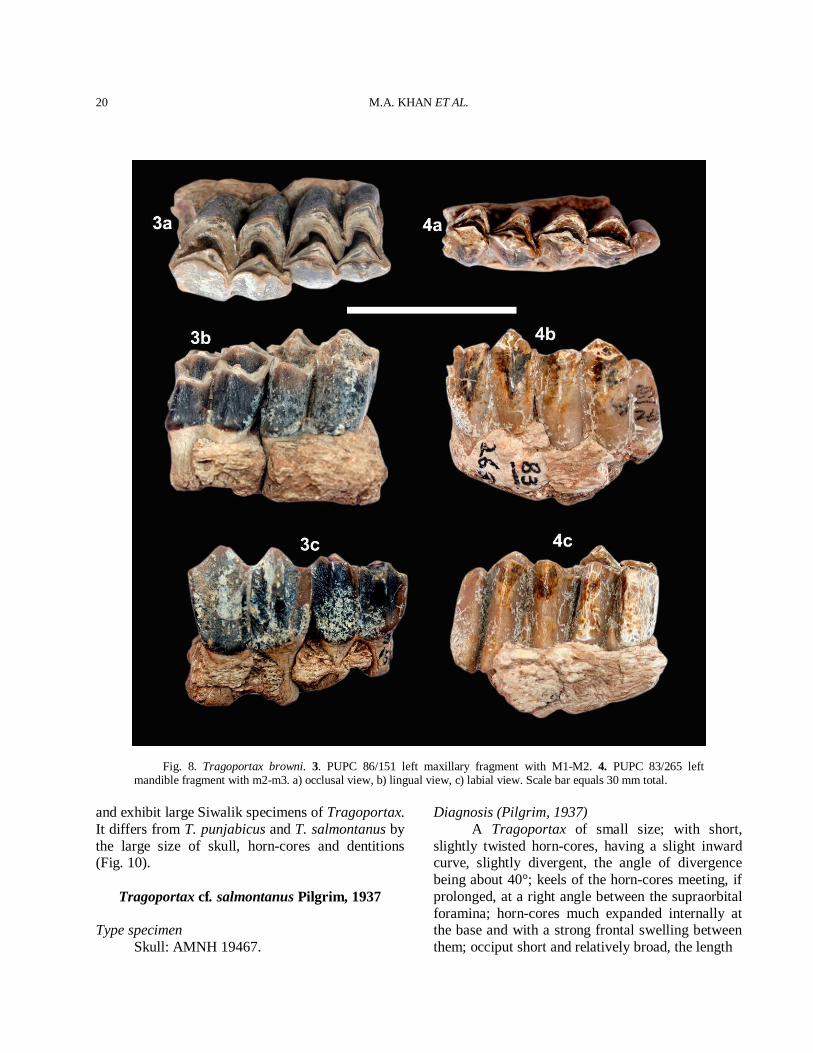
has a distally sloping parastyle with a short parastyle-mesostyle part and bilobed lingually. It has a weak hypocone. P3 is larger than P2. It has a narrow but very pronounced parastyle. P3 has a broad and strong posterior fold with a mesostyle anteriorly. P3 has a lingually inflated hypocone. P4 is a horse shoe shaped with strong anterior and posterior folds. A slight median rib is present. A very prominent and divergent parastyle is present in P4. A metastyle is moderately present. A short spur projects in the central fossette of the posterior side of the inner crescent in P4. The occlusal outline of the premolar is nearly rectangular, being much broader transversely than long. A sharp praeprotocrista unites a protocone with a parastyle. The molars are quadrate and hypsodont. The enamel is finely rugose. The molar row is longer than the premolar row. The small entostyle is present in all molars lingually. The entostyles are slightly shifted towards the hypocone. The parastyle and mesostyle are stronger than the metastyle. The metastyle is strong and posteriorly extended on M3. The anterior median rib is more prominent than the posterior one. A faint cingulum is present on the antero-internal and postero-internal surface of the molars. The main morphological features of the upper molars are the well projected mesostyle, the wider hypocone to the protocone comparatively and strong parastyle directed anteriorly (Fig. 5(4-5)). Lower dentition The lower premolars and molars are rugose (Fig. 5(6-7)). The paraconid of p2 is weaker than p3. The metaconid of p3 is backwardly directed. The entoconid is stronger than the metaconid. The paraconid of p3 is stronger than parastylid and placed antero-posterior axis of the premolar. The p4 is extended antero-posteriorly and similar to p3 (Fig. 5(6)). The p4 has a strong paraconid, metaconid and entoconid. The entoconid is fused with the endostylid. The anterior valley of p4 is open. A vestigial ectostylid is present in the molars. The hypoconulid of m3 is rather large and variable. The comparative measurements of teeth are provided in Table III. Comparison The general morphological characters of the
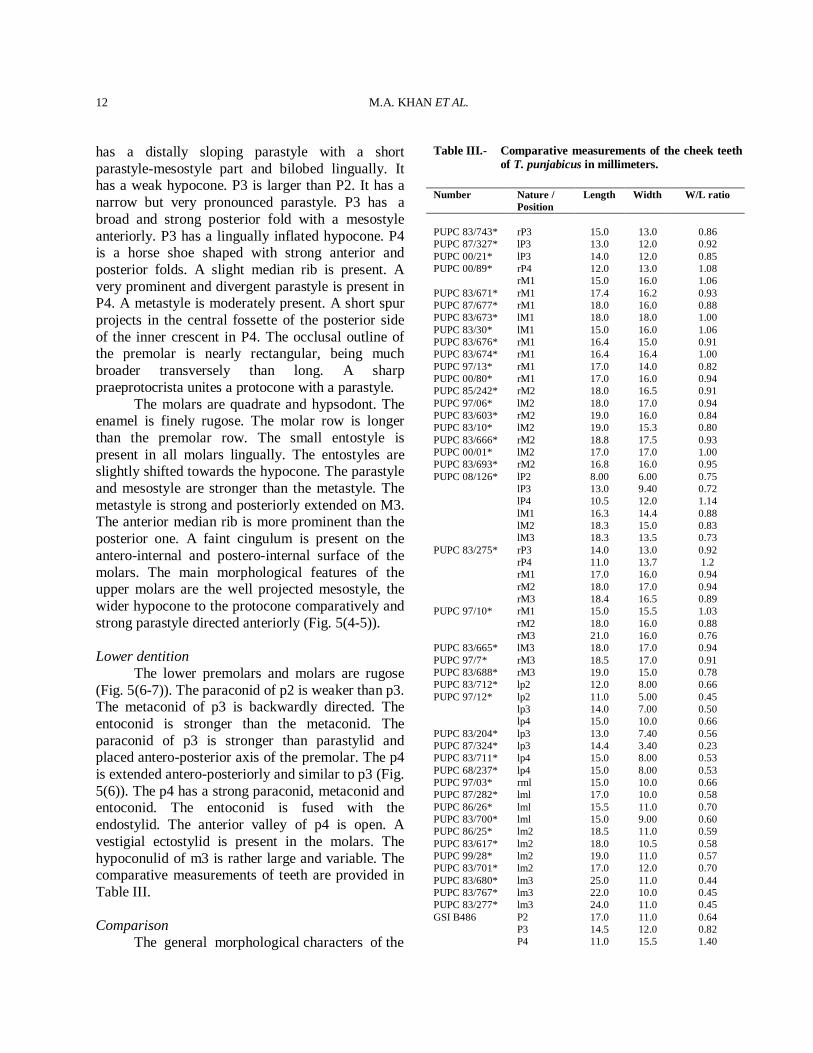
Table III.- Comparative measurements of the cheek teeth of T. punjabicus in millimeters.
Number Nature /
Position Length Width W/L ratio
PUPC 83/743* rP3 15.0 13.0 0.86 PUPC 87/327* lP3 13.0 12.0 0.92 PUPC 00/21* lP3 14.0 12.0 0.85 PUPC 00/89* rP4 12.0 13.0 1.08 rM1 15.0 16.0 1.06 PUPC 83/671* rM1 17.4 16.2 0.93 PUPC 87/677* rM1 18.0 16.0 0.88 PUPC 83/673* lM1 18.0 18.0 1.00 PUPC 83/30* lM1 15.0 16.0 1.06 PUPC 83/676* rM1 16.4 15.0 0.91 PUPC 83/674* rM1 16.4 16.4 1.00 PUPC 97/13* rM1 17.0 14.0 0.82 PUPC 00/80* rM1 17.0 16.0 0.94 PUPC 85/242* rM2 18.0 16.5 0.91 PUPC 97/06* lM2 18.0 17.0 0.94 PUPC 83/603* rM2 19.0 16.0 0.84 PUPC 83/10* lM2 19.0 15.3 0.80 PUPC 83/666* rM2 18.8 17.5 0.93 PUPC 00/01* lM2 17.0 17.0 1.00 PUPC 83/693* rM2 16.8 16.0 0.95 PUPC 08/126* lP2 8.00 6.00 0.75 lP3 13.0 9.40 0.72 lP4 10.5 12.0 1.14 lM1 16.3 14.4 0.88 lM2 18.3 15.0 0.83 lM3 18.3 13.5 0.73 PUPC 83/275* rP3 14.0 13.0 0.92 rP4 11.0 13.7 1.2 rM1 17.0 16.0 0.94 rM2 18.0 17.0 0.94 rM3 18.4 16.5 0.89 PUPC 97/10* rM1 15.0 15.5 1.03 rM2 18.0 16.0 0.88 rM3 21.0 16.0 0.76 PUPC 83/665* lM3 18.0 17.0 0.94 PUPC 97/7* rM3 18.5 17.0 0.91 PUPC 83/688* rM3 19.0 15.0 0.78 PUPC 83/712* lp2 12.0 8.00 0.66 PUPC 97/12* lp2 11.0 5.00 0.45 lp3 14.0 7.00 0.50 lp4 15.0 10.0 0.66 PUPC 83/204* lp3 13.0 7.40 0.56 PUPC 87/324* lp3 14.4 3.40 0.23 PUPC 83/711* lp4 15.0 8.00 0.53 PUPC 68/237* lp4 15.0 8.00 0.53 PUPC 97/03* rml 15.0 10.0 0.66 PUPC 87/282* lml 17.0 10.0 0.58 PUPC 86/26* lml 15.5 11.0 0.70 PUPC 83/700* lml 15.0 9.00 0.60 PUPC 86/25* lm2 18.5 11.0 0.59 PUPC 83/617* lm2 18.0 10.5 0.58 PUPC 99/28* lm2 19.0 11.0 0.57 PUPC 83/701* lm2 17.0 12.0 0.70 PUPC 83/680* lm3 25.0 11.0 0.44 PUPC 83/767* lm3 22.0 10.0 0.45 PUPC 83/277* lm3 24.0 11.0 0.45 GSI B486 P2 17.0 11.0 0.64 P3 14.5 12.0 0.82 P4 11.0 15.5 1.40

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
13
M1 18.0 18.0 1.00 M2 20.0 20.0 1.00 M3 20.0 18.5 0.92 GSI B574 M1 18.0 19.0 1.05 M2 20.5 22.0 1.07 M3 21.5 20.5 0.97 GSI B563 p2 12.5 7.00 0.56 m1 19.0 12.5 0.65 m2 21.0 13.0 0.61 m3 30.0 13.5 0.45 GSI B564 p2 13.0 7.00 0.53 p3 16.0 8.50 0.53 p4 17.5 10.0 0.57 m1 17.5 12.0 0.68 m2 20.5 14.0 0.68 m3 26.0 13.5 0.51 * The studied specimens. Referred data are taken from Pilgrim (1939) and Akhtar (1992).
available opisthocraniums, horn-core and dentitions show clear affinities with Tragoportax (Spassov and Geraads, 2004; Bibi and Gulec, 2008; Kostopoulos, 2005). The material (Figs. 3-5) is very close in morphology as well as in dimensions to the material from the already described material of the type locality (Pilgrim, 1937, 1939). The skull (Fig. 3) shows the Tragoportax postcornual and basioccipital features and basal horn-core cross
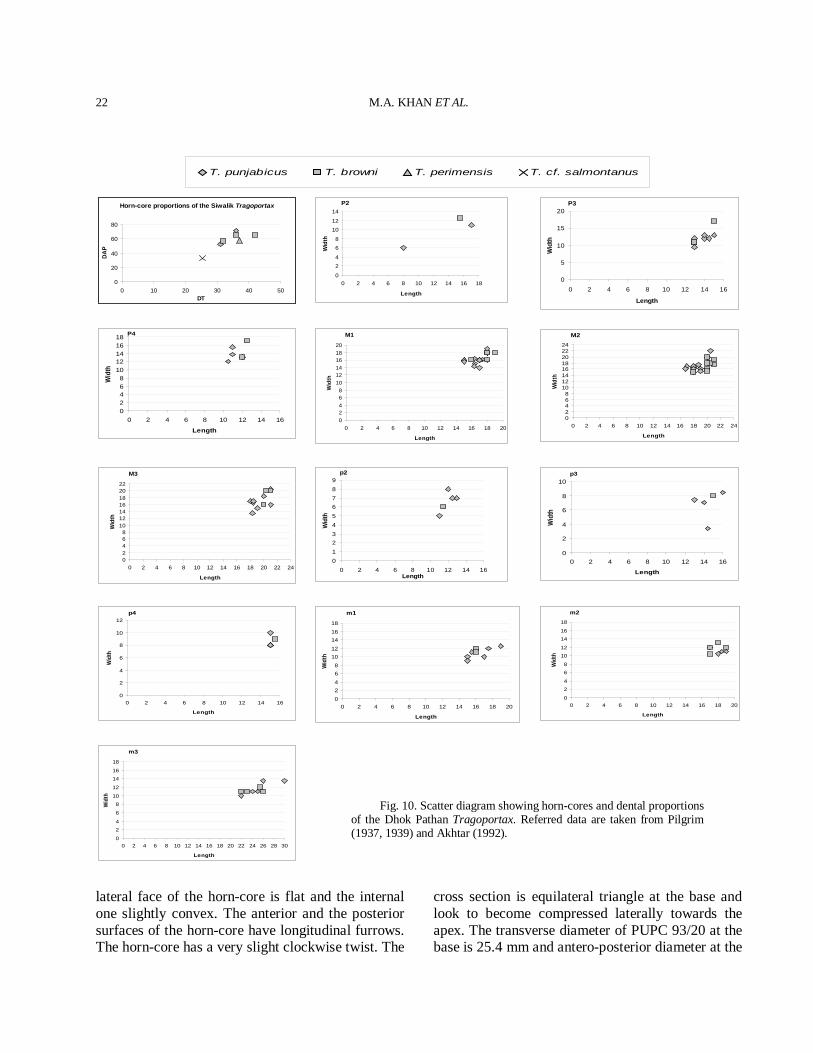
section close to Tragoportax (Spassov and Geraads, 2004; page 349). The skulls are almost identical to the type specimens of T. punjabicus and sharing in common a similar flat postcornual fronto-parietal surface with slightly concave well defined depressed area, well marked temporal ridges and caudally by a step leading to a slightly raised plateau and basi-occipital has a longitudinal groove between the anterior and the posterior tuberosities, in the bottom of which often runs a weak sagittal crest. The basal horn-core cross section is close to T. punjabicus. The dental proportions of the T. punjabicus fall within the range of the type specimens (Fig. 10), being clearly distinct from those of large Siwalik Tragoportax, T. browni and small Tragoportax, T. salmontanus (Tables III-IV). It is therefore no doubt for the inclusion of the studied material in T. punjabicus (Pilgrim, 1937, 1939). The species is also present in the Miocene of Eurasia (Kostopoulos, 2009; Bibi and Gulec, 2008). It differs from T. browni by the small size of skull, horn-cores and dentitions (Fig. 10).

Fig. 3. Tragoportax punjabicus. 1. PUPC 86/149 opisthocranium: a) dorsal view, b) ventral view, c) occipital view. 2. PUPC 86/150 opisthocranium: ventral view. Scale bar equals 30 mm total.

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
15
Fig. 4. Tragoportax punjabicus. 3. PUPC 86/264 left horn core: a) lateral view, b) medial view. Scale bar equals 30 mm total.
Tragoportax browni (Pilgrim, 1937)
Type specimen Skull: AMNH 19662. Emended diagnosis (modified from Pilgrim, 1937, 1939) A Tragoportax of large size. The size is larger than Eurasian T. curvicornis and the Siwalik T. punjabicus. Skull is broader at the orbits than in many other species. Horn-cores are long but with a
smaller antero-posterior diameter than other Tragoportax species, tilted backwards and strongly curved. Dentitions are hypsodont and larger than T. punjabicus. Studied material Opisthocranium: PUPC 86/204. Horn-cores: PC-GCUF 09/11, almost complete, side indeter-minate. PUPC 87/1, fragment, side indeterminate Upper dentition: PUPC 87/281, isolated left P3. PC-GCUF 09/42, isolated right P3. PC-GCUF 09/43,
M.A. KHAN ET AL.
16
Fig. 5. Tragoportax punjabicus. 4. PUPC 08/126 palatine parts of both maxillae with right P3-M3 and left P2-M3: occlusal view. 5. PUPC 83/275 right maxillary fragment with P3-M3. 6. PUPC 97/12 left mandible fragment with p2-p4. 7. PUPC 83/277 left mandible fragment with m3. a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
isolated right P4. PUPC 83/689, isolated left M1. PUPC 86/151, left maxillary fragment with M1-M2. PUPC 86/29, left maxillary fragment with M1-M2.
PUPC 83/861, left maxillary fragment with M1-M2. PUPC 97/80, isolated left M2. PUPC 83/662, isolated right M2. PUPC 87/324, right maxillary

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
17
fragment with M2-M3. PUPC 83/283, right maxillary fragment with M2-M3. Lower dentition: PUPC 99/27, right mandible fragment with m1-m2. PUPC 83/265, left mandible fragment with m2-m3. PUPC 83/269, right mandible fragment with m2-m3. PUPC 97/11, left mandible fragment with m3. Description Skull PUPC 86/204 opisthocranium is a fragment and incomplete specimen retaining only the occipital region, one occipital condyle, paraoccipital process, parietals, frontals, base of the right horn-core and basisphenoid (Fig. 6(1)). The opisthocranium is not compressed dorso-ventrally but the facial part, horn-core, one occipital condyle and dentition are missing. The sutures separating the various bones are pretty visible indicating relatively young animal. The suture between the parietal and the supraoccipital is more prominent than the parietal bones and the coronal suture. The intercornual area is rectangular and short antero-posteriorly. The fronto-parietal postcornual area is depressed and slightly concaves (Fig. 6(1a)). The occipital surface is rectangular, its dorsal part being broad. From the occipital foramen to the sphenoid, a continuous groove runs along the basioccipital, often with a weak sagittal crest in its middle. The opisthocranium is wide at the orbits and the occipital. The horn-cores are situated above the orbits and the cross section of the horn-core at the base is subtriangular. There is a shallow depression on the frontals behind each horn-core. The temporal ridges are strongly developed. They bifurcate a little behind the fronto-parietal suture. The inner ridge runs straight backwards and joins its fellow in the middle of the suture between the parietal and supraoccipital, enclosing a flat elevated area. The outer ridge runs back less obliquely and finally bends round by the side of the supraoccipital to join the lambdoid crest. The supraoccipital occupies a wide area of the skull dorsally i.e. 30 mm. The occipital is slightly concave in vertical direction above the foramen magnum. Nevertheless, the knob which forms the top of the occipital crest is prominent and wide and the occipital condyle projects to the rear of
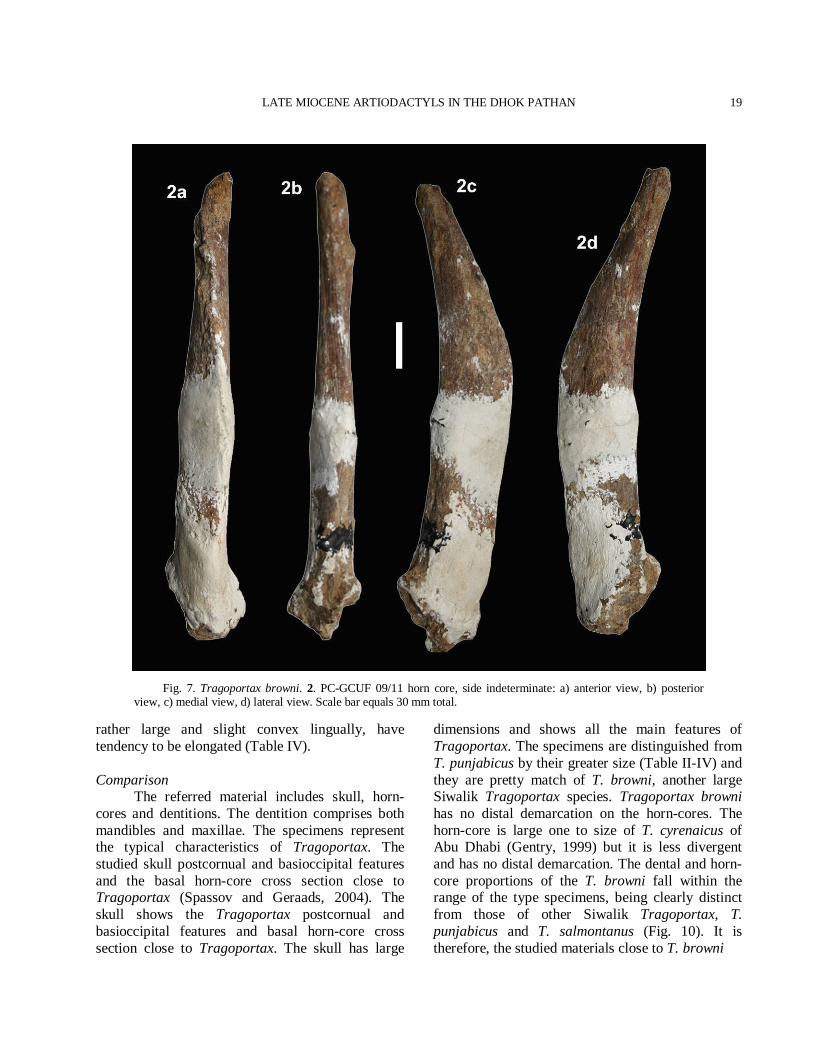
the occipital crest (Table III). The foramen magnum is large and the lamboidal crest is strongly developed. The basioccipital is subtriangular in shape. The anterior tuberosities are fairly wide apart and not very large while the posterior tuberosities are large and expanded laterally. A faint median crest is present between the tuberosities. The auditory bulla is long, laterally compressed and considerably inflated. Horn-core The horn-core is long, curved, inclined backward and almost complete. The horn-core divergence decreases towards the tip. The torsion is weak and the basal cross section is subtriangular. The horn-core is compressed laterally and straight in its lower portion (Fig. 7(2)). The antero-posterior diameter is large with a sharp anterior keel and a base with rounded corners. The antero-internal and postero-external keels are prominent. The surface is regularly ornamented by numerous longitudinal furrows that are sharper at the posterior than on the anterior. The longitudinal furrows indicate torsion. The concavity at antero-internal is prominent towards the apex and the tip of the horn-core is obtuse. The total length of PC-GCUF 09/11 measured along the anterior face is 355 mm. The transverse diameter at the base is 32 mm and antero-posterior diameter at the base is 57 mm. The mean index DT × 100/DAP is 56 at the base. Dentition The P3 has inflated hypocone. The paracone and the parastyle are strong. It has a narrow and prominent anterior fold, a broad and strong posterior fold. The median rib is placed anteriorly. The P4 is horse shoe-shaped with a rounded lingual wall. The molars are moderately high crowned. The upper molars are rugose with shining enamel. A rudimentary entostyle is present on the M1-M3 (Fig. 8(3)). A weak cingulum is present in the upper molars. The styles are very strong. The anterior median rib is more prominent than the posterior one. A spur is present in the posterior cavity. The lower molars have wrinkled enamel. The rugosity is very prominent in the lower molars. An ectostylid appears on the m1-m3 and goes up to the centre of the transverse valley (Fig. 8(4)). The hypoconulid is

M.A. KHAN ET AL.
18
Fig. 6. Tragoportax browni. 1. PUPC 86/204 opisthocranium: a) dorsal view, b) ventral view, c) occipital view. Scale bar equals 30 mm total.
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
19
Fig. 7. Tragoportax browni. 2. PC-GCUF 09/11 horn core, side indeterminate: a) anterior view, b) posterior view, c) medial view, d) lateral view. Scale bar equals 30 mm total.
rather large and slight convex lingually, have tendency to be elongated (Table IV). Comparison The referred material includes skull, horn-cores and dentitions. The dentition comprises both mandibles and maxillae. The specimens represent the typical characteristics of Tragoportax. The studied skull postcornual and basioccipital features and the basal horn-core cross section close to Tragoportax (Spassov and Geraads, 2004). The skull shows the Tragoportax postcornual and basioccipital features and basal horn-core cross section close to Tragoportax. The skull has large
dimensions and shows all the main features of Tragoportax. The specimens are distinguished from T. punjabicus by their greater size (Table II-IV) and they are pretty match of T. browni, another large Siwalik Tragoportax species. Tragoportax browni has no distal demarcation on the horn-cores. The horn-core is large one to size of T. cyrenaicus of Abu Dhabi (Gentry, 1999) but it is less divergent and has no distal demarcation. The dental and horn-core proportions of the T. browni fall within the range of the type specimens, being clearly distinct from those of other Siwalik Tragoportax, T. punjabicus and T. salmontanus (Fig. 10). It is therefore, the studied materials close to T. browni
M.A. KHAN ET AL.
20
Fig. 8. Tragoportax browni. 3. PUPC 86/151 left maxillary fragment with M1-M2. 4. PUPC 83/265 left mandible fragment with m2-m3. a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
and exhibit large Siwalik specimens of Tragoportax. It differs from T. punjabicus and T. salmontanus by the large size of skull, horn-cores and dentitions (Fig. 10).
Tragoportax cf. salmontanus Pilgrim, 1937
Type specimen Skull: AMNH 19467.
Diagnosis (Pilgrim, 1937) A Tragoportax of small size; with short, slightly twisted horn-cores, having a slight inward curve, slightly divergent, the angle of divergence being about 40°; keels of the horn-cores meeting, if prolonged, at a right angle between the supraorbital foramina; horn-cores much expanded internally at the base and with a strong frontal swelling between them; occiput short and relatively broad, the length
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
21
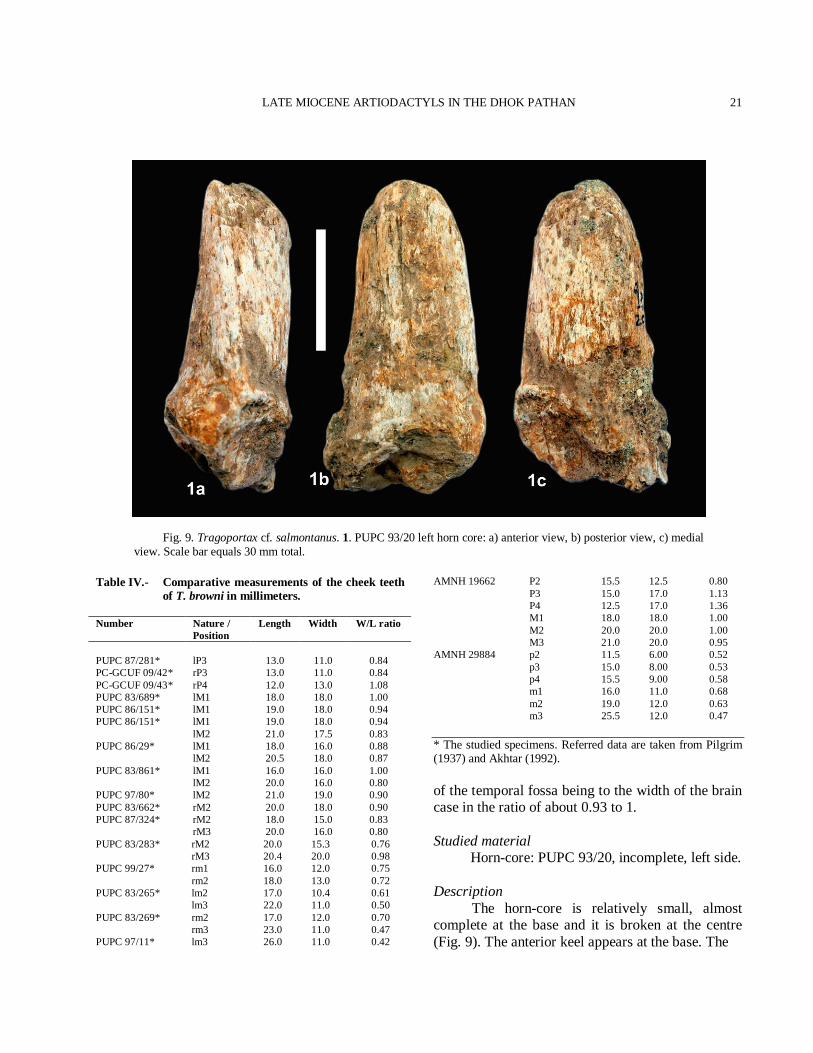
Fig. 9. Tragoportax cf. salmontanus. 1. PUPC 93/20 left horn core: a) anterior view, b) posterior view, c) medial view. Scale bar equals 30 mm total.
Table IV.- Comparative measurements of the cheek teeth
of T. browni in millimeters. Number Nature /
Position Length Width W/L ratio
PUPC 87/281* lP3 13.0 11.0 0.84 PC-GCUF 09/42* rP3 13.0 11.0 0.84 PC-GCUF 09/43* rP4 12.0 13.0 1.08 PUPC 83/689* lM1 18.0 18.0 1.00 PUPC 86/151* lM1 19.0 18.0 0.94 PUPC 86/151* lM1 19.0 18.0 0.94 lM2 21.0 17.5 0.83 PUPC 86/29* lM1 18.0 16.0 0.88 lM2 20.5 18.0 0.87 PUPC 83/861* lM1 16.0 16.0 1.00 lM2 20.0 16.0 0.80 PUPC 97/80* lM2 21.0 19.0 0.90 PUPC 83/662* rM2 20.0 18.0 0.90 PUPC 87/324* rM2 18.0 15.0 0.83 rM3 20.0 16.0 0.80 PUPC 83/283* rM2 20.0 15.3 0.76 rM3 20.4 20.0 0.98 PUPC 99/27* rm1 16.0 12.0 0.75 rm2 18.0 13.0 0.72 PUPC 83/265* lm2 17.0 10.4 0.61 lm3 22.0 11.0 0.50 PUPC 83/269* rm2 17.0 12.0 0.70 rm3 23.0 11.0 0.47 PUPC 97/11* lm3 26.0 11.0 0.42
AMNH 19662 P2 15.5 12.5 0.80 P3 15.0 17.0 1.13 P4 12.5 17.0 1.36 M1 18.0 18.0 1.00 M2 20.0 20.0 1.00 M3 21.0 20.0 0.95 AMNH 29884 p2 11.5 6.00 0.52 p3 15.0 8.00 0.53 p4 15.5 9.00 0.58 m1 16.0 11.0 0.68 m2 19.0 12.0 0.63 m3 25.5 12.0 0.47 * The studied specimens. Referred data are taken from Pilgrim (1937) and Akhtar (1992). of the temporal fossa being to the width of the brain case in the ratio of about 0.93 to 1. Studied material Horn-core: PUPC 93/20, incomplete, left side. Description The horn-core is relatively small, almost complete at the base and it is broken at the centre (Fig. 9). The anterior keel appears at the base. The
M.A. KHAN ET AL.
22
T. punjabicus T. browni T. perimensis T. cf. salmontanus
Horn-core proportions of the Siwalik Tragoportax
0
20
40
60
80
0 10 20 30 40 50DT
DAP
P2
0
24
68
1012
14
0 2 4 6 8 10 12 14 16 18
Length
Wid
th
P3
0
5
10
15
20
0 2 4 6 8 10 12 14 16
Length
Wid
th
P4
02468
1012141618
0 2 4 6 8 10 12 14 16
Length
Wid
th
M1
02468
101214161820
0 2 4 6 8 10 12 14 16 18 20
Length
Wid
th
M2
02468
1012141618202224
0 2 4 6 8 10 12 14 16 18 20 22 24
Length
Wid
th
M3
02468
10121416182022
0 2 4 6 8 10 12 14 16 18 20 22 24
Length
Wid
th
p2
0123456789
0 2 4 6 8 10 12 14 16Length
Wid
th
p3
0
2
4
6
8
10
0 2 4 6 8 10 12 14 16
Length
Wid
th
p4
0
2
4
6
8
10
12
0 2 4 6 8 10 12 14 16
Length
Wid
th
m1
02468
1012141618
0 2 4 6 8 10 12 14 16 18 20
Length
Wid
th
m2
0
24
68
1012
1416
18
0 2 4 6 8 10 12 14 16 18 20
Length
Wid
th
m3
0
24
6
810
12
1416
18
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Length
Wid
th
Fig. 10. Scatter diagram showing horn-cores and dental proportions of the Dhok Pathan Tragoportax. Referred data are taken from Pilgrim (1937, 1939) and Akhtar (1992).
lateral face of the horn-core is flat and the internal one slightly convex. The anterior and the posterior surfaces of the horn-core have longitudinal furrows. The horn-core has a very slight clockwise twist. The
cross section is equilateral triangle at the base and look to become compressed laterally towards the apex. The transverse diameter of PUPC 93/20 at the base is 25.4 mm and antero-posterior diameter at the

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
23
base is 33 mm giving its mean index DT × 100/DAP 77%. Comparison The cross section of the horn-core is typically of Tragoportax in having triangular shape and prominent anterior keel. The horn-core is shorter than in T. punjabicus and T. browni (Table II and Fig. 10). Tragoportax salmontanus has short horn-cores and the antero-posterior diameter is less (Pilgrim, 1937). The horn-core shape, size and the cross section is almost identical with T. salamontanus (Pilgrim, 1937, 1939; Kostopolous, 2009). However, the material is imperfect, based only on the horn-core and more perfect material will be required for the confirmation. Therefore, T. cf. salamontanus can be reasonably assigned for the horn-core, based on the morphometric characteristic. The horn-core of European Hadjidimovo Tragoportax are longer, less inclined and not so twisted than those of T. salmontanus from the Siwaliks (Pilgrim, 1939). Discussion Boselaphines become especially diversified in the Siwaliks. The genus Tragoportax created by Pilgrim (1937) for the Siwalik species T. salmontanus. The sample of Tragoportax exhibits fairly large range of size and morphology (Fig. 10). The two large Siwalik species T. punjabicus and T. browni shows overlapping in size (Fig. 10). However, the diagnostic features are enough to distinguish them for each other. Tragoportax browni might be synonym of T. punjabicus as proposed by Kostopoulos (2009) but it needs a more perfect material to confirm this hypothesis. Tragoportax is widespread in much of Europe and southern Asia during the Turolian (Bibi and Gulec, 2008) and recently, named the group Tragoportacini (Bibi et al., 2009). The different species of Tragoportax are best distinguished on the basis of horn-core characters. The horn-cores of the poorly known T. perimensis from the Middle Siwaliks (Pilgrim, 1939) are much shorter. Another poorly known species from the Siwaliks is T. islami. Tragoportax islami and T. perimensis are a possible synonym of T. salmontanus. The difference in horn-core curvature and in size is probably due to individual variability
(Kostopoulos, 2009). Three species of Tragoportax could be identical from the Siwaliks, based on the present investigation, one is small T. salmontanus and the second is intermediate T. punjabicus and the third is large T. browni. There is variability in diagnostic characters of Tragoportax and we can include already described skull Miotragocerus dhokpathanensis Akhtar, 1992 in T. browni. These species could be identical. The differences mentioned by Akhtar (1992) are metrical. Akhtar (1992) was not considered variability of Tragoportax. The skull is very close in morphology as well as in dimensions to the already described material of Tragoportax (Table II). Therefore, Miotragocerus dhokpathanensis is a possible synonym of T. browni. The skull is identical with the skull of T. browni and represents the same species.
Genus MIOTRAGOCERUS Stromer, 1928
Type species Miotragocerus monacensis Stromer, 1928.
Generic diagnosis (Solounias, 1981) Horn-cores triangular in cross section; above the orbits; not particularly compressed; converging anteriorly and forming a pronounced frontal buttress; not particularly twisted but with twist restricted to tips; gently diverging with tips neither turned in nor out; posterior grooves; anterior keel blunt, stopping about two-thirds from base forming at least one characteristic anterior demarcation (bump); horn-core thinner and rounder in cross section from point at which keel stops to tip; horn-core axis more vertical and bases broader anteroposteriorly, pedicals more poorly formed than in Tragoportax. Anterior keel blunter, often with several distinct growth bumps in males; less blunt, with a single bump in females. No postcornual pits; frontals strongly depressed behind horns; basicranium not particularly angled in relation to palate; preorbital fossa deep; supraorbital pits small, variable in number and position. Premolars longer in relation to the molars than in Tragoportax; P2 long; P3 with small hypocone in relation to protocone;

M.A. KHAN ET AL.
24
upper molar central cavities connect at mid-wear; entostyle small on upper molars. p4 cavity between paraconid and metaconid open; p4 paraconid tends to be larger than parastylid. Differs from Mesembriportax in having less sinused frontals; differs from Protragocerus in having longer, more compressed horn-cores with an anterior keel. Generic diagnosis (Spassov and Geraads, 2004) (The shape of horn-cores and those of associated structures are those of adult males). Size small (about that of fallow deer). The postcornual area of the skull is not depressed or raised as a low plateau. Basioccipital, definitely known in the subgenus Pikermicerus, without median longitudinal groove between the anterior and the posterior tuberosities, but with a faint sagittal keel. Strong temporal crests (at least in males) in early forms (Miotragocerus), weaker in more recent ones (Pikermicerus). Horn-cores moderately long to long in early forms and short in later ones, mediolaterally compressed, with flattened lateral and medial surfaces. Sharp anterior keel, but postero-lateral keel absent or poorly marked, and posterior face not well delimited. The section is therefore sub-elliptic. Anterior rugosities at base of horn-cores usually strong, extending onto the frontal along the keel, which often has several demarcations (steps) along its course. In front view, due to a slight torsion of the horn-cores bases, the keels are often slightly convergent upwards in the basal portion, then diverge towards the tips. The intercornual area is much longer than broad, especially narrow anteriorly. The occipital surface is high, much broader basally than at its top. Teeth brachydont, with strongly folded walls. Compared to Tragoportax, well documented forms have a long premolar row, with especially long P2 compared to P3, due to lengthening of its anterior part. Hypocone of P3 poorly expanded lingually. Metaconid of p3-p4 weak, anterior valley with incipient lingual wall. Stratigraphic range Lower and Middle Siwaliks. Geographic distribution Europe and south Asia (Spassov and Geraads, 2004).
Miotragocerus large sp.
Abbreviated Diagnosis (Pilgrim, 1939; Solounias, 1981; Spassov and Geraads, 2004) The premolars are long in relation to the molars. The premolars are as long as in Tragoportax which is a relatively larger animal. P2 has a well developed anterior metastyle unlike Tragoportax. The protocone of P3 is medially situated unlike Tragoportax. The upper molar central cavities connect at mid-wear and the entostyles are smaller than in Tragoportax. The lower dentition is more primitive than Tragoportax. The p4 cavity between the paraconid and the metaconid is open and therefore p4 is similar to p3. The lower molars have transversely situated protoconids and hypoconids and the hypoconulid of m3 is large.
Studied material Upper dentition: PUPC 83/275A, isolated right P2. PUPC 86/33, isolated right P3. PUPC 86/39, isolated left P3. PUPC 09/44, right maxillary ramus with P2-P4. PUPC 83/209, left maxillary ramus with P2-M1. PUPC 87/02 and PUPC 86/308, right maxillary ramus with DP4-M1. PUPC 87/246, left maxillary ramus with DP3-M2. PUPC 87/115, left maxillary ramus with DP4-M2. PUPC 87/116 and PUPC 87/117, right and left maxillary rami with M1-M2. Lower dentition: PUPC 88/740, left mandibular ramus with p2. PUPC 83/703, isolated left p2. PUPC 83/708 and PUPC 83/709, isolated right and left p3s. PUPC 83/742 and PUPC 87/294, isolated right and left p4s. PUPC 93/277, left mandibular ramus with p4-m1. PUPC 09/86, left mandibular ramus with p4-m3. PUPC 96/05, left mandibular ramus with p4-m1. PUPC 83/277, right mandibular ramus with m2 and broken m3.
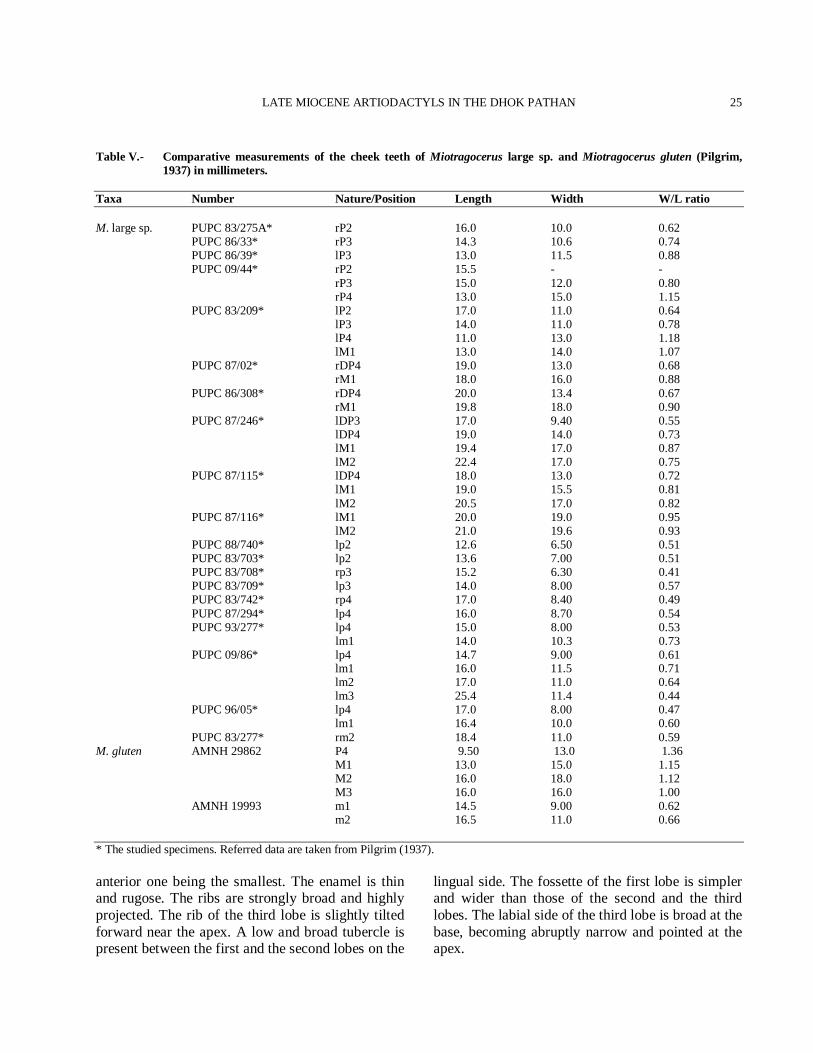
Description Upper dentition The deciduous molars are trilobed with thin enamel. The styles are bulky and divergent comparatively (Fig. 11(2)). The antero-posterior length of the deciduous molar is considerably greater than the transverse diameter (Table V). Height of the lobes increases antero-posteriorly,
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
25
Table V.- Comparative measurements of the cheek teeth of Miotragocerus large sp. and Miotragocerus gluten (Pilgrim, 1937) in millimeters.
Taxa Number Nature/Position Length Width W/L ratio M. large sp. PUPC 83/275A* rP2 16.0 10.0 0.62 PUPC 86/33* rP3 14.3 10.6 0.74 PUPC 86/39* lP3 13.0 11.5 0.88 PUPC 09/44* rP2 15.5 - - rP3 15.0 12.0 0.80 rP4 13.0 15.0 1.15 PUPC 83/209* lP2 17.0 11.0 0.64 lP3 14.0 11.0 0.78 lP4 11.0 13.0 1.18 lM1 13.0 14.0 1.07 PUPC 87/02* rDP4 19.0 13.0 0.68 rM1 18.0 16.0 0.88 PUPC 86/308* rDP4 20.0 13.4 0.67 rM1 19.8 18.0 0.90 PUPC 87/246* lDP3 17.0 9.40 0.55 lDP4 19.0 14.0 0.73 lM1 19.4 17.0 0.87 lM2 22.4 17.0 0.75 PUPC 87/115* lDP4 18.0 13.0 0.72 lM1 19.0 15.5 0.81 lM2 20.5 17.0 0.82 PUPC 87/116* lM1 20.0 19.0 0.95 lM2 21.0 19.6 0.93 PUPC 88/740* lp2 12.6 6.50 0.51 PUPC 83/703* lp2 13.6 7.00 0.51 PUPC 83/708* rp3 15.2 6.30 0.41 PUPC 83/709* lp3 14.0 8.00 0.57 PUPC 83/742* rp4 17.0 8.40 0.49 PUPC 87/294* lp4 16.0 8.70 0.54 PUPC 93/277* lp4 15.0 8.00 0.53 lm1 14.0 10.3 0.73 PUPC 09/86* lp4 14.7 9.00 0.61 lm1 16.0 11.5 0.71 lm2 17.0 11.0 0.64 lm3 25.4 11.4 0.44 PUPC 96/05* lp4 17.0 8.00 0.47 lm1 16.4 10.0 0.60 PUPC 83/277* rm2 18.4 11.0 0.59 M. gluten AMNH 29862 P4 9.50 13.0 1.36 M1 13.0 15.0 1.15 M2 16.0 18.0 1.12 M3 16.0 16.0 1.00 AMNH 19993 m1 14.5 9.00 0.62 m2 16.5 11.0 0.66 * The studied specimens. Referred data are taken from Pilgrim (1937). anterior one being the smallest. The enamel is thin and rugose. The ribs are strongly broad and highly projected. The rib of the third lobe is slightly tilted forward near the apex. A low and broad tubercle is present between the first and the second lobes on the
lingual side. The fossette of the first lobe is simpler and wider than those of the second and the third lobes. The labial side of the third lobe is broad at the base, becoming abruptly narrow and pointed at the apex.
M.A. KHAN ET AL.
26
Fig. 11. Miotragocerus large sp. 1. PUPC 83/209 left maxillary ramus with P2-M1. 2. PUPC 87/246 left maxillary ramus with DP3-M2. 3. PUPC 09/86 left mandibular ramus with p4-m3. a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
The P2 is longer than the P3 and the P4 (Fig. 11(1)). The P2 is bilobed with strongly developed paracone-parastyle-protocone complex. The paracone rib is strong and placed anteriorly. The P2
is more moralized than the P3. The P3 is quadrangular and less molarized than P2 (Fig. 11(1)). The hypocone of P3 is poorly expended lingually. A weak furrow is present lingually,

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
27
separating the hypocone from the protocone. The protruding paracone rib and the parastyle develop a narrow furrow anteriorly. The P4 has a rounded lingual wall and centrally placed median rib. The P4 has a weaker styles and a weaker paracone than P3. The molars are brachydont. The labial styles and ribs are well developed and strongly projected in upper molars (Fig. 11(2)). The anterior median rib is well projected than the posterior one. The posterior one is slightly flat. The mesostyle is robust pillar like structure. The styles are divergent. A small tubercle like entostyle is present in upper molars with strong folded walls. Lower dentition The lower dentition is brachydont with shining enamel (Fig. 11(3)). The lower premolars show tendency of molarization, lingually. The p2 is simple with a weak paraconid and strong medio-lingual cuspid. The weak praeprotoconulidcristid is present which is hardly distinguished from the fine postprotoconulidcristid. The postmetacristid is elongated and directs strongly backwards, tending to reach the postentocristid. The p3 has stronger paraconid than parastylid, situated vertically antero-posterior axis of the tooth but projected posteriorly towards the base. The preprotoconulidcristid distinguishes from the postprotoconulidcristid. The triangular metaconid is placed just behind the protoconid. It has flat lingual wall and enlarges from the tip to the base, tending to close the medial valley. The postentocristid and the posthypocristid are elongated and fused together lingually, forming a narrow, shallow and closed lingually posterior valley. The anterior valley is narrow while the medial one is wide. The paraconid is separated from the parastylid. The metaconid of p3 is free, elongated and backwardly directed. The entoconid is subtriangular and stronger than hypoconid. The hypoconid is separated from the protoconid through a labial groove. The p4 is similar to p3 but it is larger than p3. The paraconid of p4 is strong and the metaconid is extended anteroposteriorly. The entoconid of p4 is strong, fused rapidly with the endostylid. The hypoconid and the protoconid are angular labially. The labial groove between the protoconid and the
hypoconid is moderately deep. Both the prae- and post-metacristid are more developed and extend anteroposteriorly, tending to close the medial valley. The lower molars have a weak anterior transverse flange and a small ectostylid that increases from m1 to m3. The talonid of m3 is single-tubercled. The metaconid and entoconid are equally developed. The protoconid and the hypoconid are slightly constricted labially and the metastylid is stronger than the parastylid and the entostylid (Table V).
Comparison The studied material comprises the upper and the lower dentitions of medium size bovid. The morphology of these specimens is typical of Miocene boselaphines in general; the divergent styles of the teeth make their inclusion in boselaphines. There are many differences among the boselaphines from the Dhok Pathan. Selenoportax and Pachyportax are large size boselaphines found in the Dhok Pathan Formation (Khan et al., 2007; 2009a). The Helicoportax, Elachistocerus and Eotragus are comparatively small size boselaphines (Pilgrim, 1937, 1939; Akhtar, 1992; Khan et al., 2009a). The medium size boselaphines include Tragoportax and Miotragocerus (Khan et al., 2009a). The studied premolars and molars are well accentuated distinguished than those of Tragoportax (Spassov and Geraads, 2004). Furthermore, the P2 is longer than the P3-P4, the P3 poorly expanded lingually and the metaconid of the p3-p4 is weak. The morphology of the maxillar teeth and the mandibular teeth show that the samples reflects the diagnostic features of Miotragocerus and differentiate them to Tragoportax, other Siwalik medium size boselaphine of the common stratigraphic range. The morphology of the Dhok Pathan material allows it to be referred to Miotragocerus but the material metrically, is large in size than the Siwalik M. gluten (Table V; Fig. 13). Therefore, we suggest referring the Dhok Pathan material to M. large sp., while in the absence of horn-cores and cranial material species identification is impossible. However, the sample collected from the Dhok Pathan type locality is larger than the already known sample of Miotragocerus (Table V).

M.A. KHAN ET AL.
28
Miotragocerus cf. gluten (Pilgrim, 1937)
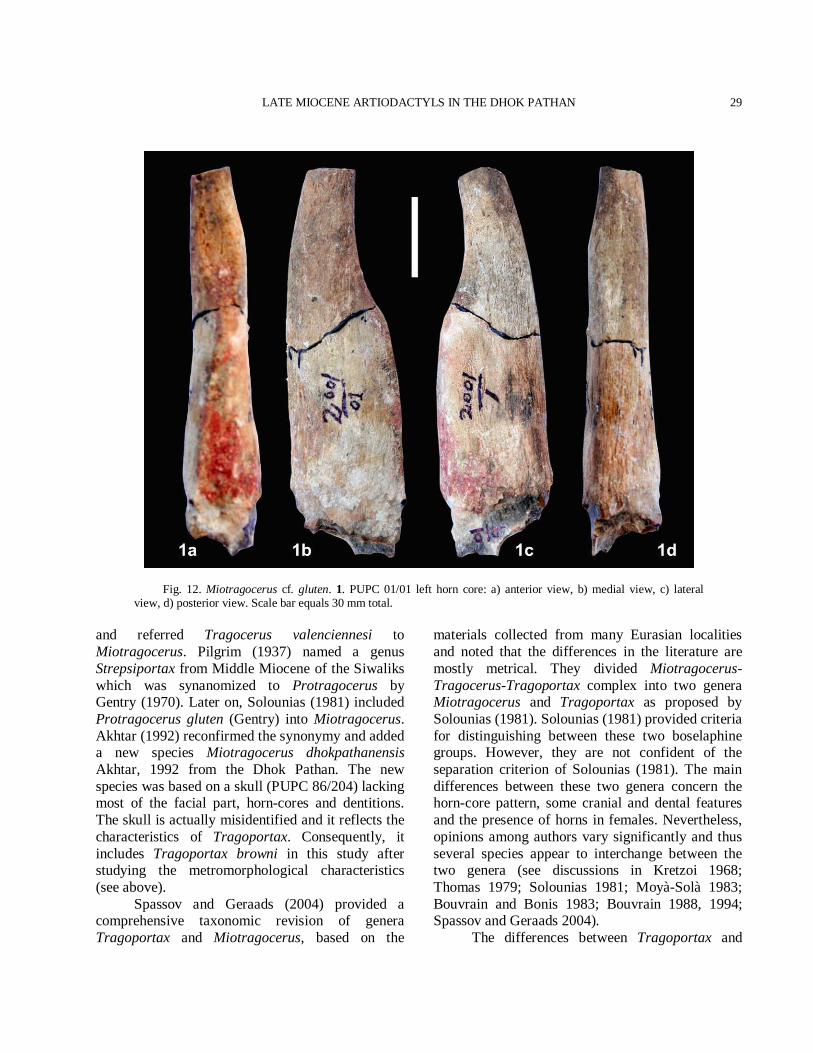
Type specimen Skull lacking most of the face and the dentition: AMNH 19746. Abbreviated Diagnosis (Pilgrim, 1937, 1939; Solounias, 1981; Spassov and Geraads, 2004) The horn-core is rather short or of moderate length, slender, slightly twisted, laterally compressed, situated immediately above the orbits and far apart on the frontals with moderately prominent antero-internal and postero-external keels with flattened lateral and slightly convex medial surfaces. The section is sub-elliptic. Anterior rugosities at base of horn-cores usually strong, extending onto the frontal along the keel, which often has several demarcations (steps) along its course. In front view, due to a slight torsion of the horn-cores bases, the keels are often slightly convergent upwards in the basal portion, then diverge towards the tips. Studied material Horn-core: PUPC 01/01, incomplete, left side. Description Horn-core The horn-core is broken at the apex. The horn-core is short, stout, slightly inclined backwards and compressed transversally (Fig. 12(1)). The antero-posterior diameter (DAP) is greater than the medio-lateral diameter (DT). The horn-core has a weak clockwise twist. The horn-core has prominent anterior keel and weak posterior keel. Between the keels the lateral surface is flat and the medial one is convex. The demarcations are well represented along the anterior keel. The anterior keel between the demarcations is well marked. The anterior keel torsion is weak and it is slightly convergent upwards at base then diverges towards the tips. The basal cross section is sub triangular. The horn-core’s surface is without longitudinal grooves but the rugosities on the anterior keel are common. The transverse diameter at the base is 21 mm and antero-posterior diameter at the base is 30 mm. The mean index DT × 100/DAP is 70% at the base. The
length of the studied horn-core is ca. 113 mm. The transverse diameter of the type AMNH 19746 at the base is 24.5 mm and its antero-posterior diameter at the base is 37 mm. The mean index DT × 100/DAP of the type is 66.2% at the base (Pilgrim, 1937). Comparison Demarcations (steps) on the anterior keel are found in Miotragocerus and Tragoportax (Spassov and Geraads, 2004). The horn-cores of Tragoportax are long and slender, usually curved backwards, with a triangular to subtriangular cross-section, well marked posterolateral keel and flattened lateral sides, but are less compressed than in Miotragocerus. Anterior rugosities growing downwards from the anterior keel at the basis of horn-cores are absent or weak, and usually do not extend onto the frontal. Horn-cores have a heteronymous torsion (anti-clockwise on the right horn), so that the anterior keels first diverge in anterior view, but they re-approach towards the tips. The studied horn-core is laterally compressed, small, clockwise torsion, weak posterolateral keel and clearly demarcated (Fig. 12(1)). All the characteristics are diagnostic enough to include the sample in genus Miotragocerus. The horn-core cross section is elliptical. In lateral view the posterior face of the horn-core is appears flat, whereas the anterior one at the base becomes concave at the upper half, following an abrupt reduction of the horn-core dimensions towards the apex. The general morphology of this horn-core is clearly similar that of Miotragocerus gluten described from the Middle Siwaliks (Pilgrim, 1937, 1939). Therefore, we provisionally assign the horn-core to Miotragocerus gluten. Discussion The type species of Miotragocerus M. monacensis Stromer, 1928, was described from a partial skull from the Astaracian/Vallesian of Oberföhring. The genus Miotragocerus is known from several Upper Miocene localities of Europe (Solounias, 1981; Spassov and Geraads, 2004) and the middle Pliocene of the Siwaliks in south Asia (Pilgrim, 1937; 1939; Akhtar, 1992). It is also known from Africa (Thomas, 1979). Solounias (1981) reviewed the genus, redescribed the species
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
29
Fig. 12. Miotragocerus cf. gluten. 1. PUPC 01/01 left horn core: a) anterior view, b) medial view, c) lateral view, d) posterior view. Scale bar equals 30 mm total.
and referred Tragocerus valenciennesi to Miotragocerus. Pilgrim (1937) named a genus Strepsiportax from Middle Miocene of the Siwaliks which was synanomized to Protragocerus by Gentry (1970). Later on, Solounias (1981) included Protragocerus gluten (Gentry) into Miotragocerus. Akhtar (1992) reconfirmed the synonymy and added a new species Miotragocerus dhokpathanensis Akhtar, 1992 from the Dhok Pathan. The new species was based on a skull (PUPC 86/204) lacking most of the facial part, horn-cores and dentitions. The skull is actually misidentified and it reflects the characteristics of Tragoportax. Consequently, it includes Tragoportax browni in this study after studying the metromorphological characteristics (see above). Spassov and Geraads (2004) provided a comprehensive taxonomic revision of genera Tragoportax and Miotragocerus, based on the
materials collected from many Eurasian localities and noted that the differences in the literature are mostly metrical. They divided Miotragocerus-Tragocerus-Tragoportax complex into two genera Miotragocerus and Tragoportax as proposed by Solounias (1981). Solounias (1981) provided criteria for distinguishing between these two boselaphine groups. However, they are not confident of the separation criterion of Solounias (1981). The main differences between these two genera concern the horn-core pattern, some cranial and dental features and the presence of horns in females. Nevertheless, opinions among authors vary significantly and thus several species appear to interchange between the two genera (see discussions in Kretzoi 1968; Thomas 1979; Solounias 1981; Moyà-Solà 1983; Bouvrain and Bonis 1983; Bouvrain 1988, 1994; Spassov and Geraads 2004). The differences between Tragoportax and
M.A. KHAN ET AL.
30
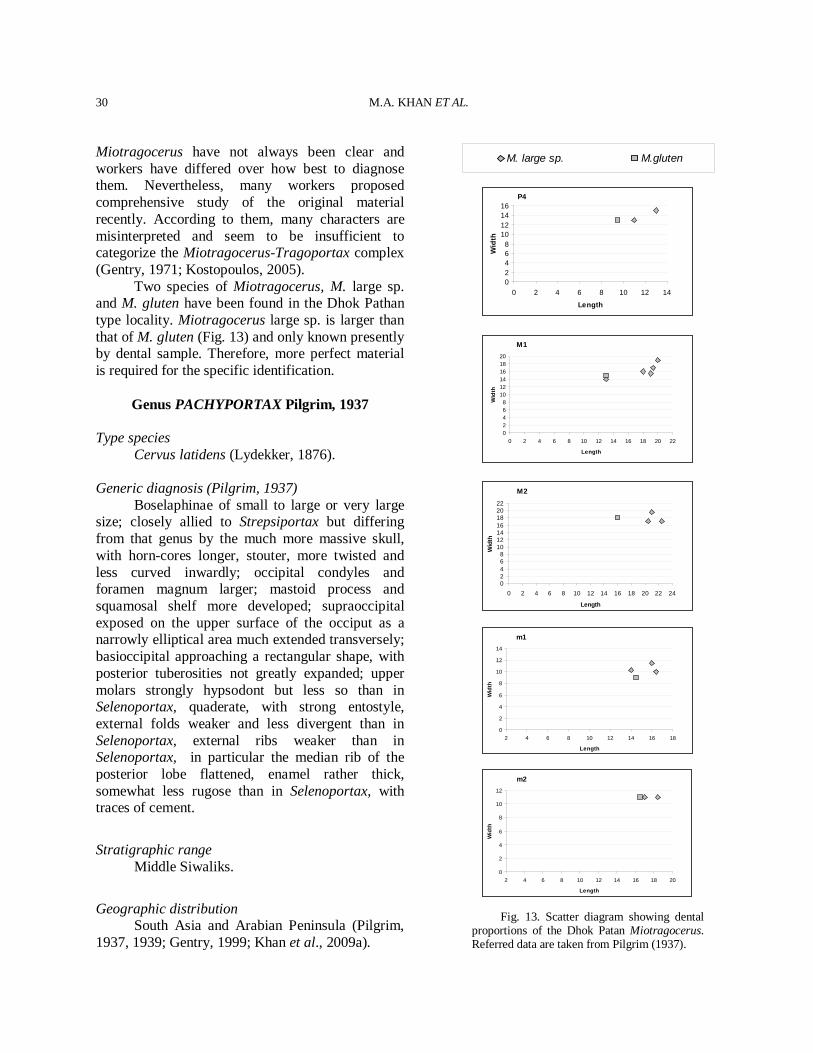
Miotragocerus have not always been clear and workers have differed over how best to diagnose them. Nevertheless, many workers proposed comprehensive study of the original material recently. According to them, many characters are misinterpreted and seem to be insufficient to categorize the Miotragocerus-Tragoportax complex (Gentry, 1971; Kostopoulos, 2005). Two species of Miotragocerus, M. large sp. and M. gluten have been found in the Dhok Pathan type locality. Miotragocerus large sp. is larger than that of M. gluten (Fig. 13) and only known presently by dental sample. Therefore, more perfect material is required for the specific identification.
Genus PACHYPORTAX Pilgrim, 1937
Type species Cervus latidens (Lydekker, 1876). Generic diagnosis (Pilgrim, 1937) Boselaphinae of small to large or very large size; closely allied to Strepsiportax but differing from that genus by the much more massive skull, with horn-cores longer, stouter, more twisted and less curved inwardly; occipital condyles and foramen magnum larger; mastoid process and squamosal shelf more developed; supraoccipital exposed on the upper surface of the occiput as a narrowly elliptical area much extended transversely; basioccipital approaching a rectangular shape, with posterior tuberosities not greatly expanded; upper molars strongly hypsodont but less so than in Selenoportax, quaderate, with strong entostyle, external folds weaker and less divergent than in Selenoportax, external ribs weaker than in Selenoportax, in particular the median rib of the posterior lobe flattened, enamel rather thick, somewhat less rugose than in Selenoportax, with traces of cement.
Stratigraphic range Middle Siwaliks.
Geographic distribution South Asia and Arabian Peninsula (Pilgrim, 1937, 1939; Gentry, 1999; Khan et al., 2009a).
P4
02468
10121416
0 2 4 6 8 10 12 14
Length
Wid
th
M1
02468
101214161820
0 2 4 6 8 10 12 14 16 18 20 22
Length
Wid
th
M2
02468
10121416182022
0 2 4 6 8 10 12 14 16 18 20 22 24
Length
Wid
th
m1
0
2
4
6
8
10
12
14
2 4 6 8 10 12 14 16 18
Length
Wid
th
m2
0
2
4
6
8
10
12
2 4 6 8 10 12 14 16 18 20
Length
Wid
th
Fig. 13. Scatter diagram showing dental proportions of the Dhok Patan Miotragocerus. Referred data are taken from Pilgrim (1937).
M. large sp. M.gluten

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
31
Pachyportax latidens (Lydekker, 1876) Type Specimen An isolated M3: GSI B219. Diagnosis (Pilgrim, 1937, 1939; Gentry, 1999; Khan et al., 2009a) Horn-cores are slender and situated directly above the orbits. They are stout with large size. They are less divergence and twisted clockwise (right horn-core). The cross section is subtriangular. The antero-internal and the postero-external keels are present. The third angle of the horn-cores is sufficient rounded to form a semicircle on the inner side of the keel. A very large sized Pachyportax, with quadrate upper molars and strong entostyle much extended transversely which through its close association with the wings of the inner lobes produces one or more fossettes in the centre of the tooth; the crown is not constricted at the apex, relatively strong styles and ribs, enamel moderately thick and rugose with traces of cement. Crown is narrow at the base and broad at the apex in Selenoportax whereas in Pachyportax the crown is not constricted at the apex. Entostyle is strong and much extending transversely in Pachyportax while in Selenoportax it is not much extending transversely. In Pachyportax posterior median rib is flattened whereas in Selenoportax it is strong like anterior median rib. Studied material Horn-core: PUPC 96/18, basal fragment, right side. Upper dentition: PUPC 80/646, isolated M2. PUPC 80/203, isolated right M2. PUPC 09/74, isolated M2. Lower dentition: PUPC 08/87, right mandibular ramus with m3 with broken hypoconulid and an anterior part of m2. Description The described material includes a horn-core (Fig. 14(1)), three upper molars and a mandibular fragment (Fig. 15(2-5)). The horn-core and the molars are partially damaged and somewhat deformed. Horn-core The anterior side of the horn-core pedicel is
preserved and it is very well rugose. The dorsal orbital rim and the surface of the orbit are also preserved (Fig. 14(1)). The horn-core’s inclination is very low in side view and it is less divergent. The cross section of the horn-core is subtriangular. The lateral side is slightly convex. The medial surface is somewhat rounded. The level of maximum transverse thickness lies posteriorly at the base. The surface of the horn-core is furnished with numerous longitudinal furrows; nevertheless a deep longitudinal ridge running along the centre of the anterior side is present. Both antero-internal and postero-external keels are present. The third angle does not form a keel but is sufficiently rounded to form a semicircle on the inner side of the keel axis. The length of the horn-core fragment is 92.5 mm. The anteroposterior (DAP) and transverse basal diameters (DT) would be 50 × 43 mm, giving it an index of compression at 86%. Upper dentition The cingulum is absolutely absent and all principal cusps the protocone, the paracone, the metacone and the hypocone are present. The enamel is fairly thick and rugose. The dentine is visible due to the early stage of wear. The entostyle is slightly damaged in PUPC 80/646 and extended transversely. The fossettes are broad and filled with sand stone (Fig. 15(2-4)). The posterior part of the anterior fossette is wider than the anterior part. The entostyle of PUPC 80/203 is badly damaged and its small portion can be seen at the base of the transverse valley of the molar. The styles are well developed; the mesostyle is strongly developed while the metastyle is moderately developed. The postprotocrista is clearly at the anterior end of the hypocone. The molars are quadrangular in shape (Fig. 15(2-4)). A prominent and distinct pressure mark lies at the anterior and the posterior sides of the molars verifying their second position in the molar series. Lower dentition It is a hypsodont and narrow crowned specimen (Fig. 15(5)). The metaconid and entoconid are slightly damaged at the apex while all ribs and stylids are well developed. The average thickness of the enamel all over the crown is 2.0 mm and it is
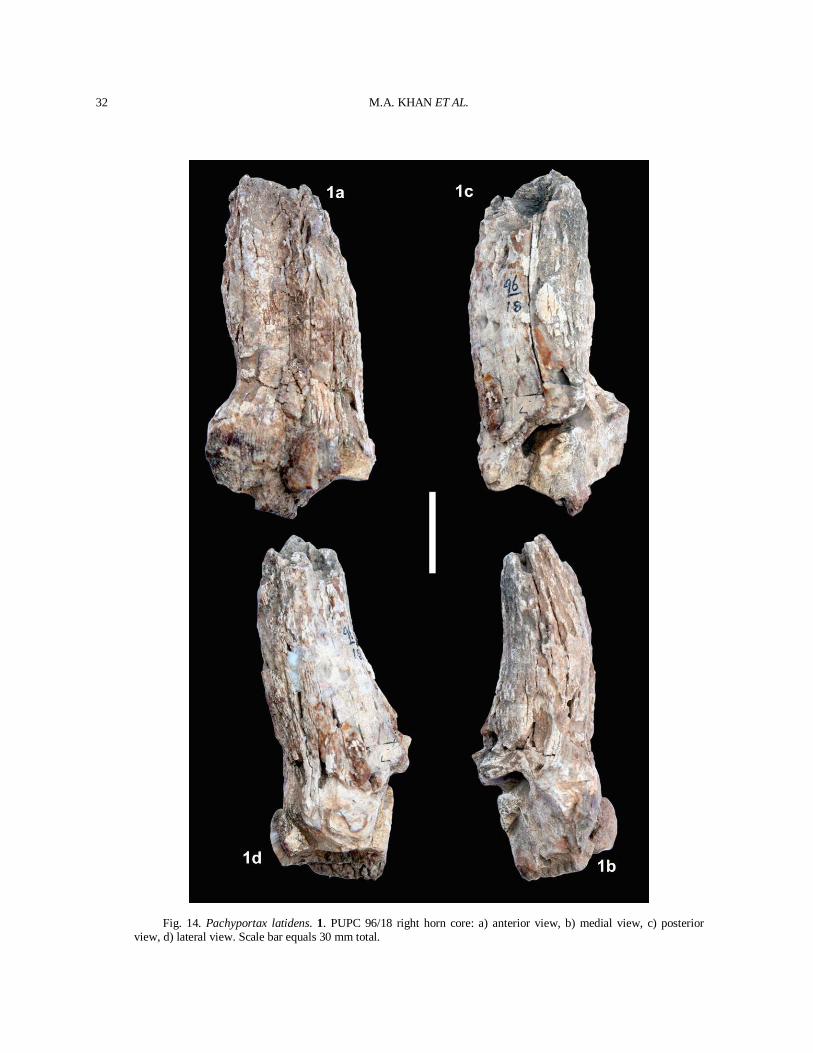
M.A. KHAN ET AL.
32
Fig. 14. Pachyportax latidens. 1. PUPC 96/18 right horn core: a) anterior view, b) medial view, c) posterior view, d) lateral view. Scale bar equals 30 mm total.
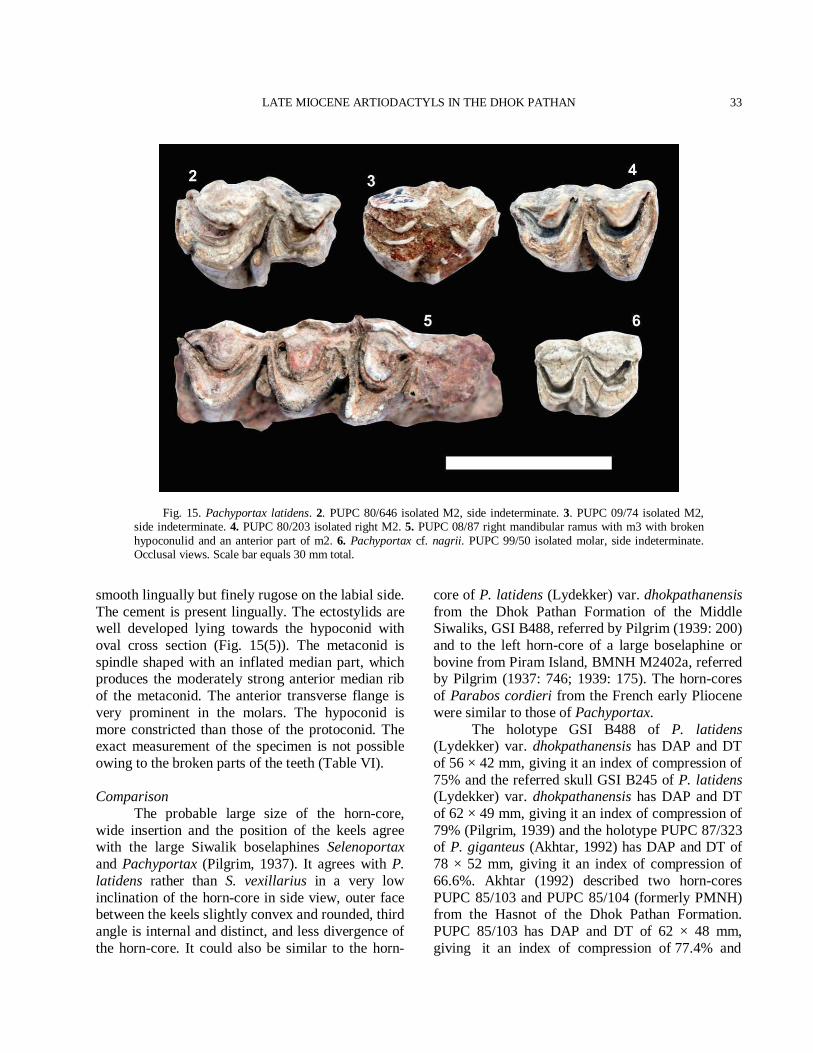
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
33
Fig. 15. Pachyportax latidens. 2. PUPC 80/646 isolated M2, side indeterminate. 3. PUPC 09/74 isolated M2, side indeterminate. 4. PUPC 80/203 isolated right M2. 5. PUPC 08/87 right mandibular ramus with m3 with broken hypoconulid and an anterior part of m2. 6. Pachyportax cf. nagrii. PUPC 99/50 isolated molar, side indeterminate. Occlusal views. Scale bar equals 30 mm total.
smooth lingually but finely rugose on the labial side. The cement is present lingually. The ectostylids are well developed lying towards the hypoconid with oval cross section (Fig. 15(5)). The metaconid is spindle shaped with an inflated median part, which produces the moderately strong anterior median rib of the metaconid. The anterior transverse flange is very prominent in the molars. The hypoconid is more constricted than those of the protoconid. The exact measurement of the specimen is not possible owing to the broken parts of the teeth (Table VI). Comparison The probable large size of the horn-core, wide insertion and the position of the keels agree with the large Siwalik boselaphines Selenoportax and Pachyportax (Pilgrim, 1937). It agrees with P. latidens rather than S. vexillarius in a very low inclination of the horn-core in side view, outer face between the keels slightly convex and rounded, third angle is internal and distinct, and less divergence of the horn-core. It could also be similar to the horn-
core of P. latidens (Lydekker) var. dhokpathanensis from the Dhok Pathan Formation of the Middle Siwaliks, GSI B488, referred by Pilgrim (1939: 200) and to the left horn-core of a large boselaphine or bovine from Piram Island, BMNH M2402a, referred by Pilgrim (1937: 746; 1939: 175). The horn-cores of Parabos cordieri from the French early Pliocene were similar to those of Pachyportax. The holotype GSI B488 of P. latidens (Lydekker) var. dhokpathanensis has DAP and DT of 56 × 42 mm, giving it an index of compression of 75% and the referred skull GSI B245 of P. latidens (Lydekker) var. dhokpathanensis has DAP and DT of 62 × 49 mm, giving it an index of compression of 79% (Pilgrim, 1939) and the holotype PUPC 87/323 of P. giganteus (Akhtar, 1992) has DAP and DT of 78 × 52 mm, giving it an index of compression of 66.6%. Akhtar (1992) described two horn-cores PUPC 85/103 and PUPC 85/104 (formerly PMNH) from the Hasnot of the Dhok Pathan Formation. PUPC 85/103 has DAP and DT of 62 × 48 mm, giving it an index of compression of 77.4% and
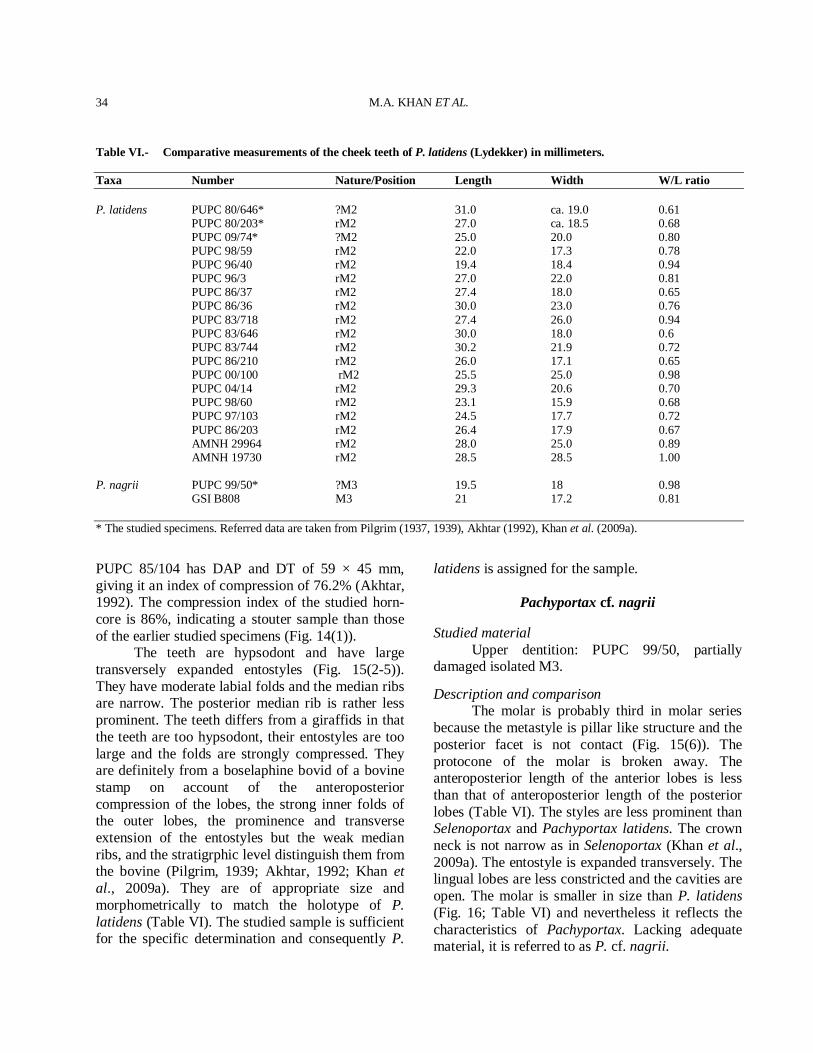
M.A. KHAN ET AL.
34
Table VI.- Comparative measurements of the cheek teeth of P. latidens (Lydekker) in millimeters. Taxa Number Nature/Position Length Width W/L ratio P. latidens PUPC 80/646* ?M2 31.0 ca. 19.0 0.61 PUPC 80/203* rM2 27.0 ca. 18.5 0.68 PUPC 09/74* ?M2 25.0 20.0 0.80 PUPC 98/59 rM2 22.0 17.3 0.78 PUPC 96/40 rM2 19.4 18.4 0.94 PUPC 96/3 rM2 27.0 22.0 0.81 PUPC 86/37 rM2 27.4 18.0 0.65 PUPC 86/36 rM2 30.0 23.0 0.76 PUPC 83/718 rM2 27.4 26.0 0.94 PUPC 83/646 rM2 30.0 18.0 0.6 PUPC 83/744 rM2 30.2 21.9 0.72 PUPC 86/210 rM2 26.0 17.1 0.65 PUPC 00/100 rM2 25.5 25.0 0.98 PUPC 04/14 rM2 29.3 20.6 0.70 PUPC 98/60 rM2 23.1 15.9 0.68 PUPC 97/103 rM2 24.5 17.7 0.72 PUPC 86/203 rM2 26.4 17.9 0.67 AMNH 29964 rM2 28.0 25.0 0.89 AMNH 19730 rM2 28.5 28.5 1.00 P. nagrii PUPC 99/50* ?M3 19.5 18 0.98 GSI B808 M3 21 17.2 0.81 * The studied specimens. Referred data are taken from Pilgrim (1937, 1939), Akhtar (1992), Khan et al. (2009a).
PUPC 85/104 has DAP and DT of 59 × 45 mm, giving it an index of compression of 76.2% (Akhtar, 1992). The compression index of the studied horn-core is 86%, indicating a stouter sample than those of the earlier studied specimens (Fig. 14(1)). The teeth are hypsodont and have large transversely expanded entostyles (Fig. 15(2-5)). They have moderate labial folds and the median ribs are narrow. The posterior median rib is rather less prominent. The teeth differs from a giraffids in that the teeth are too hypsodont, their entostyles are too large and the folds are strongly compressed. They are definitely from a boselaphine bovid of a bovine stamp on account of the anteroposterior compression of the lobes, the strong inner folds of the outer lobes, the prominence and transverse extension of the entostyles but the weak median ribs, and the stratigrphic level distinguish them from the bovine (Pilgrim, 1939; Akhtar, 1992; Khan et al., 2009a). They are of appropriate size and morphometrically to match the holotype of P. latidens (Table VI). The studied sample is sufficient for the specific determination and consequently P.
latidens is assigned for the sample.
Pachyportax cf. nagrii Studied material Upper dentition: PUPC 99/50, partially damaged isolated M3.
Description and comparison The molar is probably third in molar series because the metastyle is pillar like structure and the posterior facet is not contact (Fig. 15(6)). The protocone of the molar is broken away. The anteroposterior length of the anterior lobes is less than that of anteroposterior length of the posterior lobes (Table VI). The styles are less prominent than Selenoportax and Pachyportax latidens. The crown neck is not narrow as in Selenoportax (Khan et al., 2009a). The entostyle is expanded transversely. The lingual lobes are less constricted and the cavities are open. The molar is smaller in size than P. latidens (Fig. 16; Table VI) and nevertheless it reflects the characteristics of Pachyportax. Lacking adequate material, it is referred to as P. cf. nagrii.
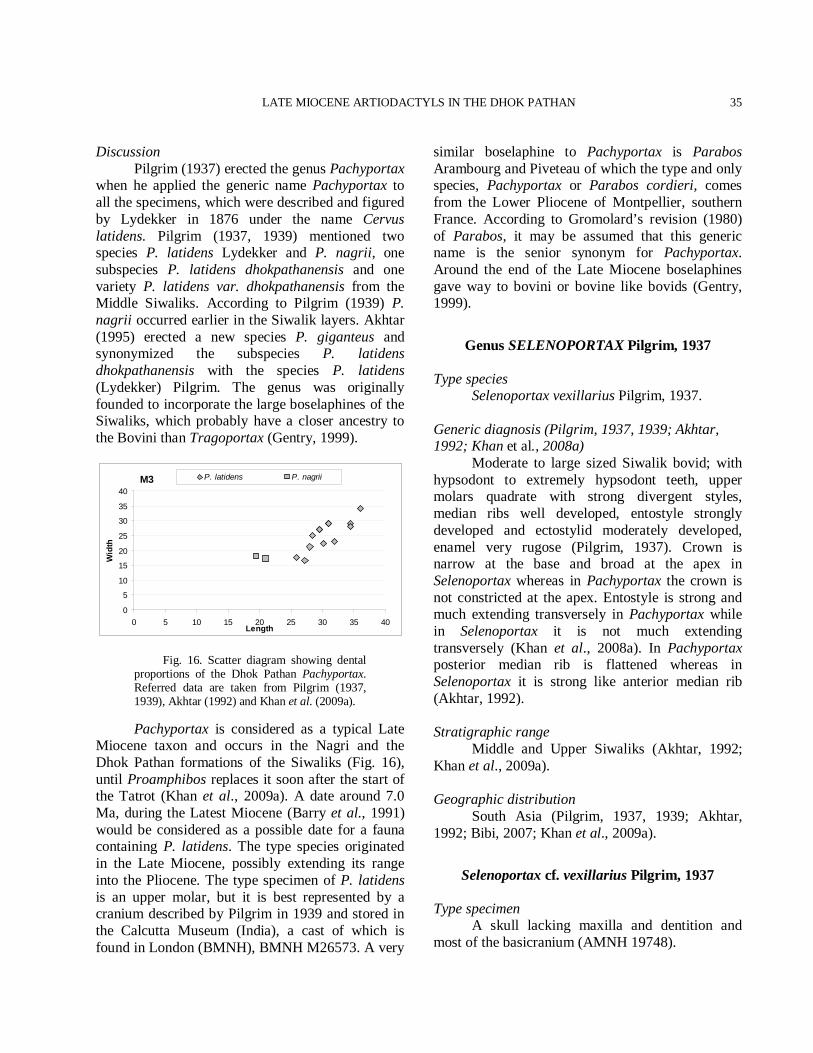
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
35
Discussion Pilgrim (1937) erected the genus Pachyportax when he applied the generic name Pachyportax to all the specimens, which were described and figured by Lydekker in 1876 under the name Cervus latidens. Pilgrim (1937, 1939) mentioned two species P. latidens Lydekker and P. nagrii, one subspecies P. latidens dhokpathanensis and one variety P. latidens var. dhokpathanensis from the Middle Siwaliks. According to Pilgrim (1939) P. nagrii occurred earlier in the Siwalik layers. Akhtar (1995) erected a new species P. giganteus and synonymized the subspecies P. latidens dhokpathanensis with the species P. latidens (Lydekker) Pilgrim. The genus was originally founded to incorporate the large boselaphines of the Siwaliks, which probably have a closer ancestry to the Bovini than Tragoportax (Gentry, 1999).
M3
0
5
10
15
20
25
30
35
40
0 5 10 15 20 25 30 35 40Length
Wid
th
P. latidens P. nagrii
Fig. 16. Scatter diagram showing dental proportions of the Dhok Pathan Pachyportax. Referred data are taken from Pilgrim (1937, 1939), Akhtar (1992) and Khan et al. (2009a).
Pachyportax is considered as a typical Late Miocene taxon and occurs in the Nagri and the Dhok Pathan formations of the Siwaliks (Fig. 16), until Proamphibos replaces it soon after the start of the Tatrot (Khan et al., 2009a). A date around 7.0 Ma, during the Latest Miocene (Barry et al., 1991) would be considered as a possible date for a fauna containing P. latidens. The type species originated in the Late Miocene, possibly extending its range into the Pliocene. The type specimen of P. latidens is an upper molar, but it is best represented by a cranium described by Pilgrim in 1939 and stored in the Calcutta Museum (India), a cast of which is found in London (BMNH), BMNH M26573. A very
similar boselaphine to Pachyportax is Parabos Arambourg and Piveteau of which the type and only species, Pachyportax or Parabos cordieri, comes from the Lower Pliocene of Montpellier, southern France. According to Gromolard’s revision (1980) of Parabos, it may be assumed that this generic name is the senior synonym for Pachyportax. Around the end of the Late Miocene boselaphines gave way to bovini or bovine like bovids (Gentry, 1999).
Genus SELENOPORTAX Pilgrim, 1937
Type species Selenoportax vexillarius Pilgrim, 1937. Generic diagnosis (Pilgrim, 1937, 1939; Akhtar, 1992; Khan et al., 2008a) Moderate to large sized Siwalik bovid; with hypsodont to extremely hypsodont teeth, upper molars quadrate with strong divergent styles, median ribs well developed, entostyle strongly developed and ectostylid moderately developed, enamel very rugose (Pilgrim, 1937). Crown is narrow at the base and broad at the apex in Selenoportax whereas in Pachyportax the crown is not constricted at the apex. Entostyle is strong and much extending transversely in Pachyportax while in Selenoportax it is not much extending transversely (Khan et al., 2008a). In Pachyportax posterior median rib is flattened whereas in Selenoportax it is strong like anterior median rib (Akhtar, 1992). Stratigraphic range Middle and Upper Siwaliks (Akhtar, 1992; Khan et al., 2009a). Geographic distribution South Asia (Pilgrim, 1937, 1939; Akhtar, 1992; Bibi, 2007; Khan et al., 2009a).
Selenoportax cf. vexillarius Pilgrim, 1937
Type specimen A skull lacking maxilla and dentition and most of the basicranium (AMNH 19748).

M.A. KHAN ET AL.
36
Abbreviated diagnosis (Khan et al., 2008a, 2009a) Cheek teeth large and strongly hypsodont, enamel very rugose. Upper molars quadrate with strong and divergent styles near the neck of crown, ribs quite large, entostyle/ectostylid strongly developed. Fossettes without indentations and simple in outline, anterior transverse flange poorly developed at front of lower molars. S. vexillarius is smaller than that of S. lydekkeri which was the large one species of the Siwalik Selenoportax. Studied material Lower dentition: PUPC 99/47, right mandibular ramus with m2 and roots of m1. PUPC 99/22, isolated left semierupted molar. PUPC 00/45, isolated left m2. PUPC 07/16, partially damaged molar. PUPC 97/02, isolated right m3. PUPC 97/01, isolated right m3. Description The sample of Selenoportax comprises only lower dentition (Fig. 17(1-3)). Three out of six are preserved with all morphological features whereas rest of the three is partially damaged. A mandibular ramus having length 73 mm bears m2. Lower dentition The specimens of the lower dentition present well developed second and third molars (Fig. 17(1-3); Table VII). The molars have fully developed principal conids. The conids are narrower than the cones and the anterior transverse flange is moderately developed in the lower molars. The molars present various stages of wear and have prominent ectostylids. In the third molars the hypoconulid is well developed, long and narrow, with a wide and inflated central area having two sloping cristids. The ribs and stylids are divergent lingually. Comparison The specimens are large enough to include them the large Siwalik boselaphines of the late Miocene Selenoportax and Pachyportax (Pilgrim, 1937; Khan et al., 2009a). The general contour of the studied specimens, the rugosity of the enamel, the strong ectostylids, the prominent median ribs, constricted labial lobes, the strong and divergent
stylids (Fig. 17(1-3)) exclude the specimens from the genus Pachyportax and favor their inclusion in the genus Selenoportax. Selenoportax is represented by two species in the Siwaliks S. vexillarius and S. lydekkeri. The dimensions and the morphology of the studied material (Fig. 18) reveal all the features of the species S. vexillarius as cited by Pilgrim in 1937 and 1939 (Table VII). The studied specimens present the same morphological features of the holotype (Pilgrim, 1937). The paraconid, the parastylid, the protoconid, the entoconid, and the development of the stylids are fairly similar. The second molars are narrow crowned, as indicated by its width and length ratio, and resemble with the specimens AMNH 19844, AMNH 19514 and AMNH 29917 as discussed by Pilgrim in 1937. The enamel layer of the third molar is finely plicated and it compares very favorably with AMNH 19514 (Pilgrim, 1937). The studied sample morphometrically (Fig. 18; Table VII) resembles with S. vexillarius, however, the material only comprises lower dentition and for the exact specific determination more material is required. Therefore, S. cf. vexillarius is assigned for the lower dentition. Discussion Pilgrim (1937) described two species of genus Selenoportax, S. vexillarius and S. lydekkeri based on a collection from various Siwalik localities in Pakistan and India. Akhtar (1992) ascribed both S. vexillarius and S. lydekkeri from the Middle and Upper Siwaliks in the genus Selenoportax and added two new species in it, S. dhokpathanensis and S. tatrotensis; the former was based on a damaged cranium (PUPC 86/248) and the later was based upon a maxillary portion bearing right P3-M3 and left P3-M3 (PUPC 87/19). More recently, Khan et al. (2009a) synanomyzed S. dhokpathanensis with S. lydekkeri and S. tatrotensis with S. vexillarius. Therefore, the genus Selenoportax consists only of two valid species and these are S. vexillarius and S. lydekkeri. S. vexillarius is smaller in size than S. lydekkeri (Fig. 18). In general the cheek teeth of S. vexillarius are smaller than those of S. lydekkeri (Fig. 18). Selenoportax records from the late middle Miocene and Pliocene of the Siwaliks (Akhtar, 1992; Barry et al., 2002; Bibi, 2007), nevertheless some uncertain occurrences are mentioned out of
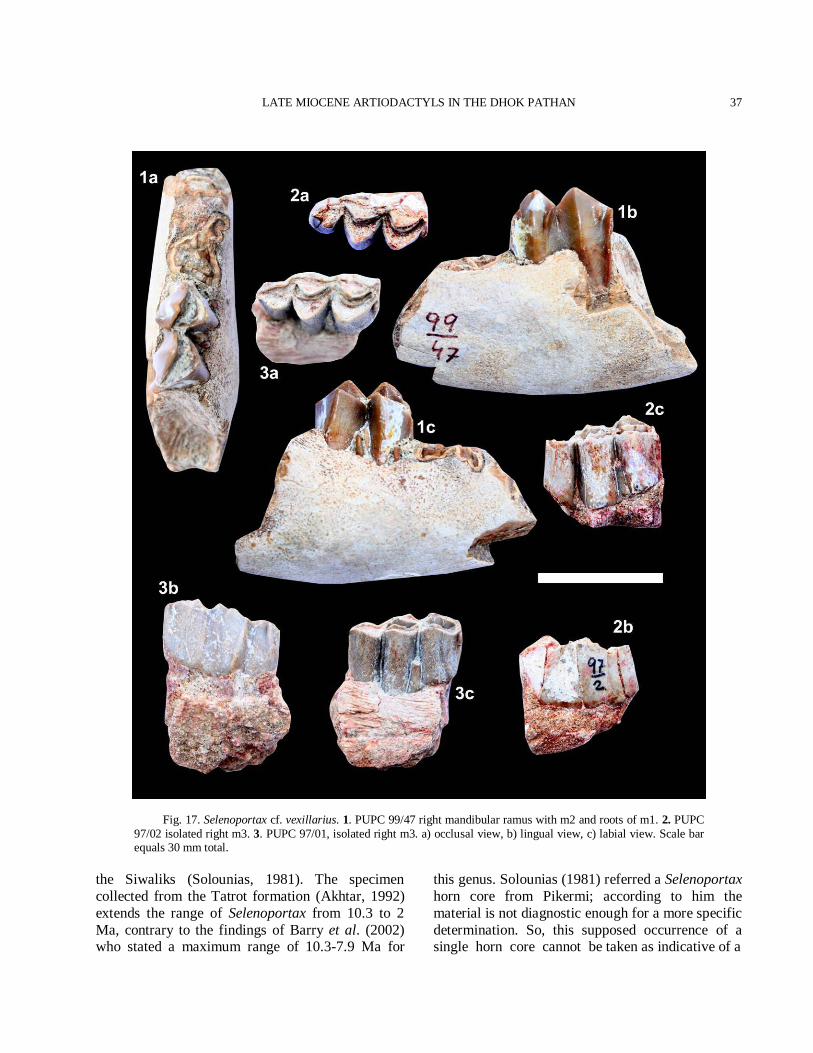
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
37
Fig. 17. Selenoportax cf. vexillarius. 1. PUPC 99/47 right mandibular ramus with m2 and roots of m1. 2. PUPC 97/02 isolated right m3. 3. PUPC 97/01, isolated right m3. a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
the Siwaliks (Solounias, 1981). The specimen collected from the Tatrot formation (Akhtar, 1992) extends the range of Selenoportax from 10.3 to 2 Ma, contrary to the findings of Barry et al. (2002) who stated a maximum range of 10.3-7.9 Ma for
this genus. Solounias (1981) referred a Selenoportax horn core from Pikermi; according to him the material is not diagnostic enough for a more specific determination. So, this supposed occurrence of a single horn core cannot be taken as indicative of a
M.A. KHAN ET AL.
38
Table VII.- Comparative measurements of the cheek teeth of S. cf. vexillarius in millimeters. Taxa Number Nature/Position Length Width W/L ratio S. cf. vexillarius PUPC 99/47* rm2 23.0 12.0 0.52 PUPC 99/22* lm? 26.5 - - PUPC 00/45* lm2 26.0 13.4 0.51 PUPC 07/16* m? 25.4 13.0 0.51 PUPC 97/02* rm3 27.0 14.0 0.51 PUPC 97/01* rm3 28.0 13.0 0.46 S. vexillarius PUPC 04/1 lm2 27.9 16.1 0.57 lm3 31.4 16.0 0.50 PUPC 98/78 lm2 25.0 16.0 0.64 lm3 36.0 15.0 0.41 PUPC 85/40 lm1 19.7 12.5 0.63 PUPC 04/12 lm2 20.0 12.5 0.62 PUPC 87/90 lm3 38.0 16.5 0.43 AMNH 10514 lm3 33.0 15.0 0.45 AMNH 29917 lm1 18.0 13 0.72 AMNH 19844 lm2 25.9 16.5 0.63 AMNH 19514 lm2 22.0 15.5 0.70 AMNH 29917 lm2 21.0 15.0 0.71 AMNH 19514 lm3 33.0 21.5 0.65 * The studied specimens. Referred data are taken from Pilgrim (1937, 1939), Akhtar (1992) and Khan et al. (2009a). Selenoportax presence in Europe. Qiu and Qiu (1995) list Selenoportax sp. from the Lufeng fauna (Chinese Miocene), age of 11.1-8.0 Ma (Steininger, 1999; Flynn and Qi, 1982). These specimens represent the sole record of Selenoportax from the Miocene of northern and central Asia (Tedford et al., 1991; Nakaya, 1994; Qiu and Qiu, 1995).
Subfamily Antilopinae Gray, 1821 Tribe Antilopini Gray, 1821
Genus PROSTREPSICEROS Major, 1891
Type species Prostrepsiceros houtumschindleri (Rodler and Weithofer, 1890). Generic diagnosis (Bibi and Gulec, 2008) Horn-cores with anticlockwise torsion, open spiraling, amounting to about 180° total torsion (half a rotation); moderately strongly divergent; inserted above orbit; mediolaterally compressed; cross-section ovate to kidney-shaped, with lateral surface flat basally to slightly concave distally; posterolateral keel present; anterior keel variably
present, always less sharp than posterolateral keel; surfaces with longitudinal grooving present; horn-cores extending onto pedicels anteriorly. Supraorbital foramina small, circular, opening anteriorly, and placed in semi-triangular pit. Post-cornual fossa present, wide and deep. Frontal sinuses small, not extending into pedicel. The p3 bearing separate paraconid parastylid; backwardly-slanted metaconid that fuses with entoconid in early wear stages; large open lingual valley between paraconid and metaconid; entoconid and entostylid well separated but fusing in mid-wear stages; entoconid much larger than entostylid; hypoconid weak; some cementum present in the lingual valleys. Upper teeth differ from those of Gazella in molars of larger size; with metacone more rounded; wider paracone rib; mesostyle wider and more projecting; and metastyle wider. Stratigraphic range Lower and Middle Siwaliks. Geographic distribution From Middle Miocene to Late Miocene. Prostrepsiceros is a bovid common in the late
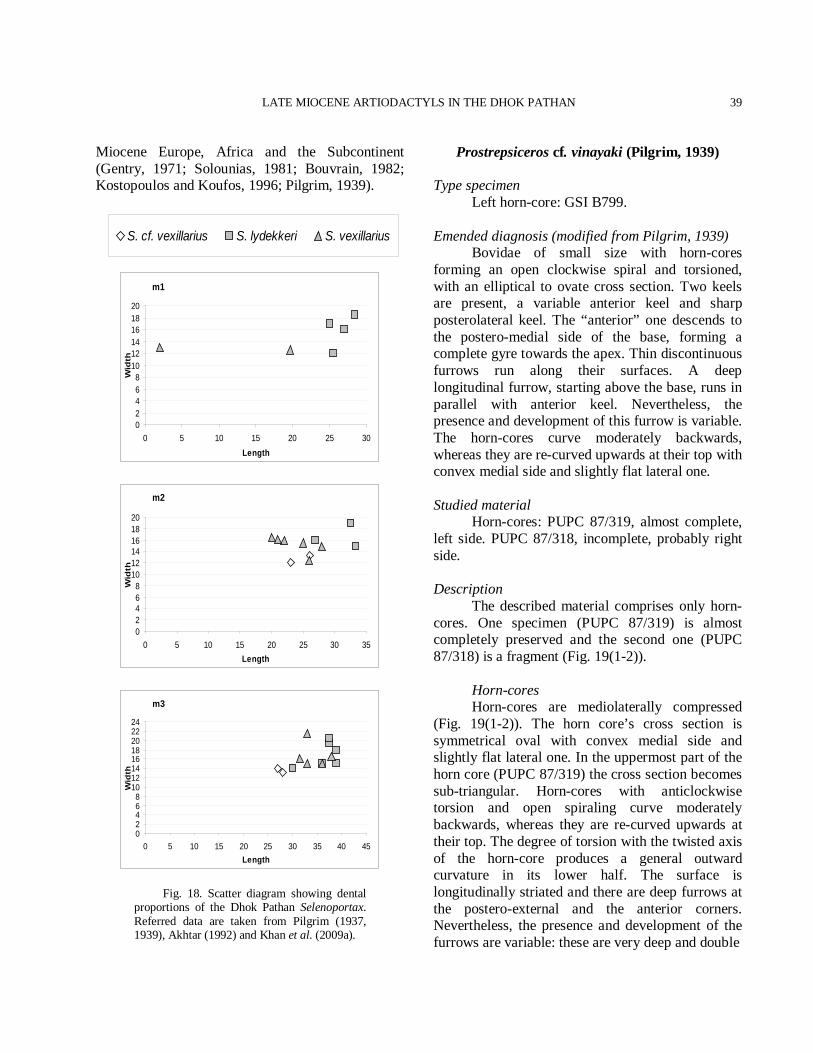
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
39
Miocene Europe, Africa and the Subcontinent (Gentry, 1971; Solounias, 1981; Bouvrain, 1982; Kostopoulos and Koufos, 1996; Pilgrim, 1939).
m1
02468
101214161820
0 5 10 15 20 25 30Length
Wid
th
m2
02468
101214161820
0 5 10 15 20 25 30 35Length
Wid
th
m3
02468
1012141618202224
0 5 10 15 20 25 30 35 40 45Length
Wid
th
Fig. 18. Scatter diagram showing dental proportions of the Dhok Pathan Selenoportax. Referred data are taken from Pilgrim (1937, 1939), Akhtar (1992) and Khan et al. (2009a).
Prostrepsiceros cf. vinayaki (Pilgrim, 1939)
Type specimen Left horn-core: GSI B799. Emended diagnosis (modified from Pilgrim, 1939) Bovidae of small size with horn-cores forming an open clockwise spiral and torsioned, with an elliptical to ovate cross section. Two keels are present, a variable anterior keel and sharp posterolateral keel. The “anterior” one descends to the postero-medial side of the base, forming a complete gyre towards the apex. Thin discontinuous furrows run along their surfaces. A deep longitudinal furrow, starting above the base, runs in parallel with anterior keel. Nevertheless, the presence and development of this furrow is variable. The horn-cores curve moderately backwards, whereas they are re-curved upwards at their top with convex medial side and slightly flat lateral one. Studied material Horn-cores: PUPC 87/319, almost complete, left side. PUPC 87/318, incomplete, probably right side. Description The described material comprises only horn-cores. One specimen (PUPC 87/319) is almost completely preserved and the second one (PUPC 87/318) is a fragment (Fig. 19(1-2)). Horn-cores Horn-cores are mediolaterally compressed (Fig. 19(1-2)). The horn core’s cross section is symmetrical oval with convex medial side and slightly flat lateral one. In the uppermost part of the horn core (PUPC 87/319) the cross section becomes sub-triangular. Horn-cores with anticlockwise torsion and open spiraling curve moderately backwards, whereas they are re-curved upwards at their top. The degree of torsion with the twisted axis of the horn-core produces a general outward curvature in its lower half. The surface is longitudinally striated and there are deep furrows at the postero-external and the anterior corners. Nevertheless, the presence and development of the furrows are variable: these are very deep and double
S. cf. vexillarius S. lydekkeri S. vexillarius

M.A. KHAN ET AL.
40
Fig. 19. Prostrepsiceros cf. vinayaki. 1. PUPC 87/319 left horn-core. 2. PUPC 87/318 ?right horn-core. a) anterior view, b) medial view, c) lateral view, d) posterior view. Scale bar equals 30 mm total.

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
41
in PUPC 87/318 but less deep in PUPC 87/319. Two keels are present. The “anterior” one descends to the postero-medial side of the base, forming a complete gyre towards the apex. It is smooth in the lower part of the horn-core but becomes sharper in its upper half. A deep longitudinal furrow, starting 2-3 cm above the base, runs in parallel with this keel or even replaces it. The second keel descends over the anterior margin of the postcornual groove and also forms a complete gyre towards the top. The preserved length of PUPC 87/319 is 158 mm, the DAP is 33 at the base and the DT is 26 mm at the base. The mean index DT × 100/DAP is 78% at the base.
Comparison The general morphological characters of the available horn-cores allot the medium sized antelope of the Middle Siwaliks to Prostrepsiceros (Pilgrim, 1939; Thomas, 1984). The deep anterior and posterior furrows, extremely torsioned, ovate cross section, two keels and mediolaterally compressed (DAP > DT) observed in the studied sample places them into Prostrepsiceros (Fig. 19(1-2)). The late Miocene Prostrepsiceros is poorly known from the Dhok Pathan type locality (Pilgrim, 1939; Thomas, 1984; Akhtar, 1992; Kostopoulos, 2005). In our knowledge the Dhok Pathan sample constitutes at present time one of the best known evidence of this genus in the Middle Siwaliks of Pakistan. Prostrepsiceros vinayaki (Pilgrim, 1939) is a known species of the Siwalik Prostrepsiceros (Pilgrim, 1939; Gentry, 1999; Kostopoulos, 2005). The observed characters in the studied sample are present in the Siwalik P. vinayaki and the material is very good matches for P. vinayaki. It agrees with the Siwalik P. vinayaki in having deep anterior and posterior furrows, extremely torsioned, ovate cross section, two keels and mediolaterally compression. It is very similar to the holotype horn-core (GSI B799) of P. vinayaki in having almost same index of compression. The DAP and DT at the base of GSI B799 being 25.5 × 19.5 mm, giving an index of compression of 76%. It is, however, slightly larger, in the studied sample, has index of compression 78%. Prostrepsiceros aff. vinayaki of Emirates of Abu Dhabi has anteroposterior and transverse basal
diameters of 29.6 × 20.9 mm, giving it an index of compression of 71%. Nisidorcas planicorni (Pilgrim, 1939) of Peram Island and Graeco-Turkish Turolian (Bouvrain, 1979; Kohler, 1987) is similar sized, and compressed to about the same extent, but has a posterior keel strengthening distally and no anterior keel (Gentry, 1999). According to the available data from Eurasian several localities (Pilgrim 1939; Gentry 1999; Kostopoulos, 2005; Bibi and Gulec, 2008) P. vinayaki differs from Eurasian Prostrepsiceros P. vallesiensis, P. houtumschindleri, P. rotundicornis, P. fraasi and P. zitteli in the longer and slenderer horn-cores, which are more torsioned than spiraled and extremely compressed mediolaterally (Bouvrain, 1982; Gentry, 1999).
Discussion Pilgrim (1939, p. 42) created a new species Gazellinae (?) gen. indet. (aff. Helicotragus) vinayaki, based on a very fragmentary left horn-core. Gentry (1970, p. 259) supposed that the holotype left horn-core GSI B799 reflects characters of young Sivoreas eremite. The holotype was collected from the locality Nila of Dhok Pathan Formation. Thomas (1984) studied two more horn-coresYGSP 4082 and YGSP 2631, collected from the same level of the Dhok Pathan Formation and ascribed species ‘vinayaki’. He observed that the holotype and the new sample clearly distinguished from Sivoreas eremite. Having observed the close resemblance of the sample with Prostrepsiceros, Thomas (1984) allotted the material to P. vinayaki (Pilgrim, 1939). The studied horn-cores collected from the type locality Dhok Pathan morphometrically resemble with the previous sample and reflect the morphometric characteristics of P. vinayaki. We therefore consider the horn-cores to P. vinayaki and confirm here its presence in the type locality of the Dhok Pathan Formation. Prostrepsiceros occurs in a wide geographic area of Saudi Arabia, North Africa, Greece, Tibetan Plateau and the Subcontinent (Pilgrim, 1939; Thomas, 1984; Bouvrain 1982; Bouvrain and Thomas 1992; Gentry 1999, 2003). Prostrepsiceros has two or more lineages and is the most diversified genus. Some species have keels and inclined

M.A. KHAN ET AL.
42
insertions of the horn-cores; others have no vestigial keels, open spiraling, more or upright insertions and little inclination of their cranial roofs. The radiation and distribution of the genus, as well as the phylogenetic relationships among the quite numerous affiliated taxa are not yet clear enough (Kostopoulos, 2009). The genus appeared in 10 Ma (Gentry, 2003) and exists hereafter with a variety of forms (Bouvrain and Thomas, 1992; Kostopoulos and Koufos, 1996; Bonis and Koufos, 1999), suggesting a fast radiation and distribution. Early representatives of Prostrepsiceros share in common a weak cranio-facial angle, a long basioccipital with a wide central groove, small supraorbital foramens not into pits and away from the frontal base, small postcornual grooves and slender horn-cores comparatively to the skull size, inserted on the posterior part of the orbital roof, strongly inclined backwards and widely separated on the frontals. Some of these apparently primitive features can also be seen in the poorly known Sinapodorcas from Middle Sinap, Turkey (Bouvrain et al., 1994; Gentry, 2003), a genus that seems to me very closely related to Prostrepsiceros. Bouvrain’s (1982) suggestion in dividing Prostrepsiceros in two distinct subgenera might indeed reflect an early division in two distinct lineages that evolved independendly from late Vallesian onwards. The first one includes P. vallesiensis, P. houtumschindleri, P. lybicus and P. vinayaki and the second Sinapodorcas, P. syridisi, P. rotundicornis, P. fraasi, P. zitteli and P. axiosi. According to Lehmann and Thomas (1987) a relationship between P. lybicus and the Jebel Hamrin form seems undeniable while Gentry (1999) looks for the possible ancestor stock of Sahabi form in P. fraasi from Samos. Kostopoulos (2009) suggests a possible relation between P. lybicus, P. vinayaki and P. vallesiensis. According to Gentry et al. (1999) Antilope itself probably descends from within Prosterpsiceros. The ancestry of Antilope cervicapra, the living blackbuck of India lies within Prostrepsiceros (Gentry, 1999).
Genus GAZELLA Blainville, 1816
Type species Gazella dorcas Linn., 1758.
Generic diagnosis (Pilgrim, 1939; Kostopoulos, 2009) Upper molar hypsodont, with prominent narrow styles, without entostyles, fossettes crescentic in shape, median ribs moderately strong in primitive forms, weak or absent in progressive forms, P2 longer than P3 and P4. Lower molars with goat folds, small ectostylids, fossettes with fairly simple outline, stylids and ribs moderately developed. Horn-cores moderately long, spaced, inserted over the orbits, not very divergent, sub circular or elliptical in cross section, unkeeled, inward or forward curvature near the tip, surface covered by longitudinal striae. Stratigraphic range Lower to Middle Siwaliks (Middle Miocene to Pliocene). Geographic distribution The occurrence of Gazella is recorded from the Lower Pliocene of Eurasia and several middle Miocene and Pleistocene localities of Africa (Gentry, 1999). It is abundantly found in the Lower Pliocene fauna of Asia and the southern parts of Europe (Kostopoulos, 2009). It is recorded from the Siwaliks of the Subcontinent (Pilgrim, 1937, 1939; Thomas, 1984; Akhtar, 1992; Khan, 2007).
Gazella lydekkeri Pilgrim, 1939
Type specimen A skull and conjoined mandible (AMNH 19663). Diagnosis (Pilgrim, 1937, 1939) Upper molars moderately hypsodont, styles narrow and strong with entostyles very small or absent, enamel moderately thick and rugose, fossettes narrow and deep, anterior median rib stronger than posterior one, premolar series rather long. Lower molars extremely hypsodont approaching quadrate shape, with small ectostylids, prominent goat folds, fossettes fairly simple in outline, stylids and ribs moderately developed. Horn cores moderately long, spaced, slightly curved backward, broadly elliptical in cross-section, fine ribs becoming rudimentary near the tips, one deep furrow posteriorly.

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
43
Fig. 20. Gazella lydekkeri. 1. PUPC 83/319 left horn-core. 2. PC-GCUF 09/45 left horn-core. a) anterior view, b) medial view, c) lateral view, d) posterior view. 3. PUPC 08/136 right isolated m2: occlusal view. 4. PUPC 07/13 isolated left m3. 5. PUPC 09/76 isolated left m3. a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
Studied material Horn-cores: PUPC 83/319, almost complete with a piece of frontal, broken at the apex, left side. PC-GCUF 09/45, almost complete with supraorbital pit, left side. Lower dentition: PUPC 08/136, right isolated m2. PUPC 07/13 and PUPC 09/76, isolated
left m3s. Description The material comprises two horn-cores, a second lower molar and two lower third molars (Fig. 20).

M.A. KHAN ET AL.
44
Horn-cores The horn cores curve gently upwards and backwards (Fig. 20(1-2)). The horn core looks to diverge very slightly and taper towards the tip. The supraorbital pit is preserved in PC-GCUF 09/45. It is pear shaped. The horn cores are relatively short, smart and present directly above the orbit. The bases of the horn cores are strongly elliptical, without keels, untwisted and slightly more convex on the inner side than on the outer side. The surfaces of the horn cores are furnished by the longitudinal furrows and there is one deep furrow present posteriorly. The horn cores arise close together and diverging gradually. The horn cores are supported by the smooth pedicels. The horn cores look alike morphologically and metrically. The preserved length of PUPC 83/319 is 72.6 mm, the DAP is 23.5 mm at the base and the DT is 18.5 mm at the base. The mean index DT × 100/DAP is 78.7% at the base. For PC-GCUF 09/45 the length is 60.5 mm, the DAP is 21.5 mm, the DT is 17.5 giving the index of compression is 81.3%. Lower dentition The m2 is well preserved and subhypsodont (Fig. 20(3)). The ribs are broader at the base and gradually narrow toward the tips. The anterior median rib is more evident than the posterior one. The mesostylid on the lingual side is very weak, visible only on the upper half of the tooth crown. On the lingual side, a short tubercle is present at the base between the metaconid and the entoconid. The labial transverse valley is occupied at the base by a short and robust ectostylid. A prominent goat fold is present along the anterior surface of the tooth. All the cusps are well preserved and conical in form. The lingual cusps are higher than the labial ones. The fossettes are moderately wide and simple. The metaconid is slightly higher than the entoconid. PUPC 07/13 and PUPC 09/76 are well preserved left isolated third lower molars (Fig. 20(4-5)). The enamel is rugose especially on the lingual side. All the conids are well preserved and the lingual ones are pointed. The anterior goat fold is broad and prominent, and the hypoconulid is narrow and crescentic (Fig. 20(3-5)). The ectostylid is robust, rounded and relatively longer. The mesostylid is narrow, gradually prominent towards
the apex. The comparative measurements are provided in Table VIII. Table VIII.- Comparative measurements (mm) of the cheek
teeth of G. lydekkeri. Number Nature /
position Length Width W/L ratio
PUPC 84/67 rm2 16.6 10.0 0.60 PUPC 87/160 lm2 14.0 9.00 0.64 PUPC 04/02* rm2 14.0 8.70 0.62 rm3 19.0 8.60 0.45 PC-GCUF 09/16
lm2 16.0 11.0 0.68
lm3 21.0 10.5 0.5 PUPC 04/08 lm3 20.0 9.00 0.45 PUPC 86/04 lm2 15.0 10.0 0.66 PUPC 87/ 162 lm3 22.0 10.5 0.47 PUPC 67/42 rm3 20.0 9.00 0.45 PUPC 08/136* rm2 15.5 7.50 0.48 PUPC 07/13* lm3 22.0 9.50 0.43 PUPC 0976* lm3 23.7 10.6 0.44 * The studied specimens. Referred data are taken from Pilgrim (1937, 1939), Akhtar (1992) and Khan (2008). Comparison The horn cores of Gazella are rising almost directly above the orbits and gently curving upwards and backwards. The horn cores characterizing an oval cross section with mediolateral compression are indicative of Gazella (Kostopoulos, 2009, Pilgrim, 1937, 1939; Akhtar, 1992). The studied horn cores reflect similar morphological characters with G. lydekkeri characteristically arises immediately above the orbit, moderated length, without keel, less mediolaterally compression, small but deep lachrymal fossa and has elliptical cross section (Fig. 20; Table VIII). The horn cores refer to G. lydekkeri while noting its morphometric characteristics. The small size and the less laterally compression of the horn-cores is somewhat distinguish it to male G. lydekkeri and indicates that the horn-cores belong to female animal (Gentry, 1970). The small and deep lachrymal fossa is present in G. lydekkeri as in living G. granti and very different from the large and deep fossa in living G. thomsoni (Bibi and Gulec, 2008). The horn cores differ from those of G. capricornis because of not having circular cross section. It is different from G. pilgrimi which has almost straight horn cores with more persist few longitudinal ribs (Kostopoulos,
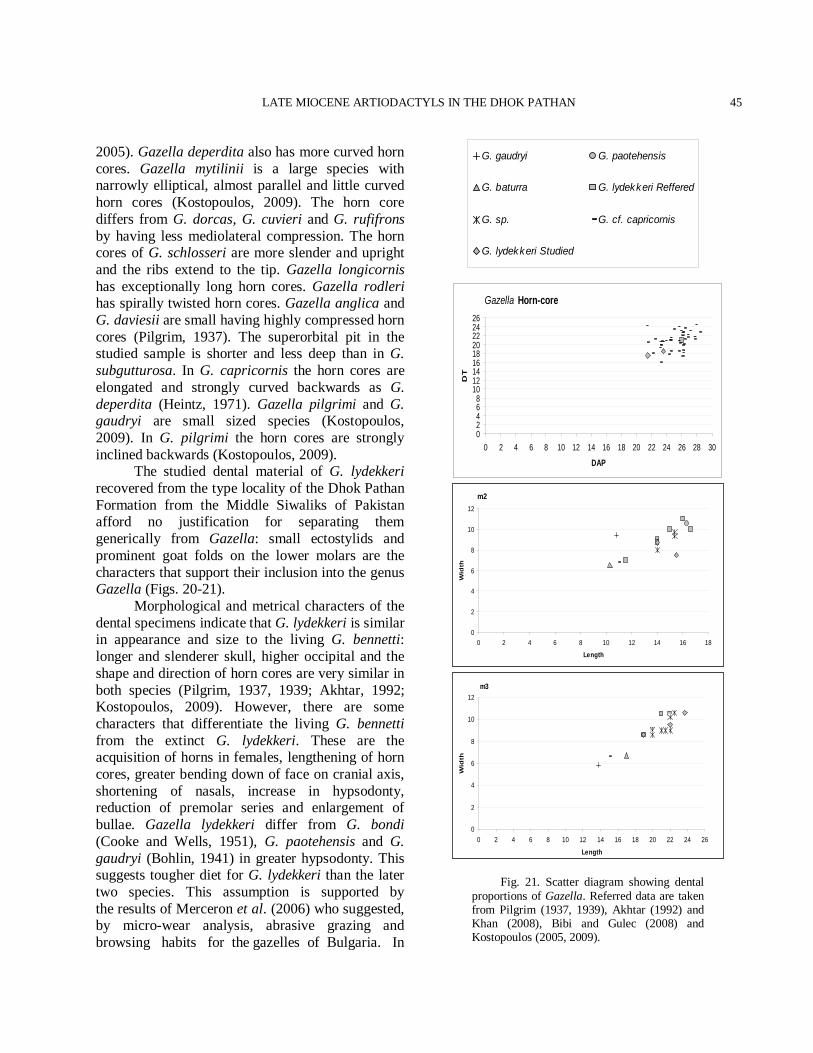
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
45
2005). Gazella deperdita also has more curved horn cores. Gazella mytilinii is a large species with narrowly elliptical, almost parallel and little curved horn cores (Kostopoulos, 2009). The horn core differs from G. dorcas, G. cuvieri and G. rufifrons by having less mediolateral compression. The horn cores of G. schlosseri are more slender and upright and the ribs extend to the tip. Gazella longicornis has exceptionally long horn cores. Gazella rodleri has spirally twisted horn cores. Gazella anglica and G. daviesii are small having highly compressed horn cores (Pilgrim, 1937). The superorbital pit in the studied sample is shorter and less deep than in G. subgutturosa. In G. capricornis the horn cores are elongated and strongly curved backwards as G. deperdita (Heintz, 1971). Gazella pilgrimi and G. gaudryi are small sized species (Kostopoulos, 2009). In G. pilgrimi the horn cores are strongly inclined backwards (Kostopoulos, 2009). The studied dental material of G. lydekkeri recovered from the type locality of the Dhok Pathan Formation from the Middle Siwaliks of Pakistan afford no justification for separating them generically from Gazella: small ectostylids and prominent goat folds on the lower molars are the characters that support their inclusion into the genus Gazella (Figs. 20-21). Morphological and metrical characters of the dental specimens indicate that G. lydekkeri is similar in appearance and size to the living G. bennetti: longer and slenderer skull, higher occipital and the shape and direction of horn cores are very similar in both species (Pilgrim, 1937, 1939; Akhtar, 1992; Kostopoulos, 2009). However, there are some characters that differentiate the living G. bennetti from the extinct G. lydekkeri. These are the acquisition of horns in females, lengthening of horn cores, greater bending down of face on cranial axis, shortening of nasals, increase in hypsodonty, reduction of premolar series and enlargement of bullae. Gazella lydekkeri differ from G. bondi (Cooke and Wells, 1951), G. paotehensis and G. gaudryi (Bohlin, 1941) in greater hypsodonty. This suggests tougher diet for G. lydekkeri than the later two species. This assumption is supported by the results of Merceron et al. (2006) who suggested, by micro-wear analysis, abrasive grazing and browsing habits for the gazelles of Bulgaria. In
Gazella Horn-core
02468
101214161820222426
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
DAPD
T
m2
0
2
4
6
8
10
12
0 2 4 6 8 10 12 14 16 18
Length
Wid
th
m3
0
2
4
6
8
10
12
0 2 4 6 8 10 12 14 16 18 20 22 24 26
Length
Wid
th
Fig. 21. Scatter diagram showing dental proportions of Gazella. Referred data are taken from Pilgrim (1937, 1939), Akhtar (1992) and Khan (2008), Bibi and Gulec (2008) and Kostopoulos (2005, 2009).
G. gaudryi G. paotehensis
G. baturra G. lydekkeri Reffered
G. sp. G. cf. capricornis
G. lydekkeri Studied

M.A. KHAN ET AL.
46
G. dorcadoides and G. altidens the teeth are more hypsodont than in G. lydekkeri. Gazella lydekkeri is similar to G. dorcadoides in hypsodonty but is different due to equally strong anterior and posterior median ribs of the later. Gazella lydekkeri is smaller with longer premolars, shorter, slenderer and remarkably less compressed horn cores than G. pilgrimi and G. capricornis from Turkey (Kostopoulos, 2005). The length of post canine teeth series in G. lydekkeri is greater than that of G. capricornis. In both these species P2 is longer than P3, P3 than P4 and p3 than p4. Premolars of G. lydekkeri are, however, narrower than those of G. capricornis. Entostyles are most frequently found in M1 and M2 of G. capricornis and less frequently in G. gaudryi, G. dorcadoides and G. lydekkeri. Fairly strong paracone rib, quite flat metacone labial surface and small basal tubercles are the characters of G. capricornis (Bibi and Gulec, 2008) that are very close to G. lydekkeri. The ribs on the inner side of lower molars are less pronounced in G. lydekkeri but extremely weak in G. dorcadoides and G. pilgrimi in which the inner surfaces of the lower molars are almost flat, much like the living forms of Gazella. The posterior cleft of the p4 of G. lydekkeri is restricted to the half of the tooth towards the base of the crown, whereas in G. capricornis it is well distinguishable down towards the base. The posterior valley in p4 of G. pilgrimi is more blocked than in G. lydekkeri. The p4 of G. dorcadoides seems to resemble closely to that of G. lydekkeri. Gazella lydekkeri is different from G. capricornis in skull and horn core characters. In case of G. lydekkeri, basioccipital is comparatively narrower, horn cores are less upright, less curved and have narrower cross section as opposed to G. capricornis. Gazella lydekkeri differs from G. soriae (Alcalá and Morales, 2006) by its smaller size and greater hypsodonty but similar in the reduction of the premolar row with respect to the molars. Gazella lydekkeri shares hypsodonty and short premolar sequence as a common character with G. gaozhuangensis (Chen, 1997) but distinct in p4 morphology and posterolabial crest of M3. Discussion The most species rich and wide spread antilopine is Gazella. Gazella and the Antilopini
became widespread in the upper Miocene, probably from their possible African or Siwalik origin (Gentry, 1994) and the fossils attributed to this taxon go back to the mid Miocene (Gentry, 1970). Gazella became widespread in Eurasia in the late Miocene, originating probably from Homoiodorcas (Gentry, 1999). The genus Gazella was erected by Blainville (1816), who recorded the first fossil horn-cores of Gazella stehlini in the European upper Vindobonian (Gentry, 1966). Most species of the Gazella are found as fragmentary material, such as isolated teeth and horn cores. All the European species are small. Pilgrim (1937, 1939) described two species of Gazella from the sediments of the Middle Siwaliks, G. lydekkeri, and ?G. superba, on the basis of the horn core size variation and the curvature. The holotype of G. lydekkeri is almost complete, comprising a skull and a conjoined mandible (AMNH 19663). The studied material defines sufficient similarity with G. lydekkeri. Pilgrim (1939) did not consider the variability owing to lack of sufficient material. Morphological horn core variability within a species is due to the great changes in horn core shape (Kostopoulos, 2005) and should be credited to interaspecific variability (Fig. 21). The sample of the type locality confirms the presence of G. lydekkeri in the Dhok Pathan Formation. G. lydekkeri shows horn core morphology with the Eurasian G. capricornis (Fig. 21). The horn core exhibits the characteristic features of G. capricornis (Pilgrim, 1937; Senyurek, 1954; Pilgrim and Hopwood, 1928). Recently, Kostopoulos (2005) observed the same horn core pattern as in G. lydekkeri. Earlier authors (Gentry, 1970, 1971; Bibi and Gulec, 2008) already proposed affinity of the Siwalik G. lydekkeri with Pontian G. capricornis.
Subfamily Hippotraginae Gray, 1821 Tribe Reduncini Gray, 1821
Genus KOBUS A. Smith, 1840
Type species Kobus ellipsiprymnus (Ogilby, 1833). Generic diagnosis (Pilgrim, 1939; Gentry, 1980) Reduncine of moderate to small size; with

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
47
large, relatively divergent, and backwardly tilted, slightly curved horn-cores, having a slight clockwise twist, with an oval, laterally compressed cross section, insertions at a low inclination; face somewhat bent down on the cranial axis; supraorbital pits rather large and deep with pointed posterior end. Stratigraphic range Middle Siwaliks. Geographic distribution Late Miocene to Pliocene of the Subcontinent and Eurasia (Pilgrim, 1939, Gentry, 1980; Thomas, 1980).
Kobus porrecticornis (Lyddeker, 1878)
Type specimen Frontlet with the horn-cores: GSI B229. Emended diagnosis (modified from 1939) A faint keel like structure is present proximally. The horn-cores are curved moderately backwards having a slight clockwise twist. Horn-cores are rather slender and decrease gradually in diameter towards the tip; their basal cross section is subovate. The surface of the horn-cores is furnished with longitudinal furrows. The presence of longitudinal furrows is variable. The lateral surface of the horn-cores is slightly flat and the medial one is convex. Studied material Horn-cores: PUPC 87/124 A+B, left and right horn-cores, proximally preserved fragments, perhaps same individual. PUPC 02/126 and PUPC 87/180, partial left horn-cores. Description The recovered material of Kobus porrecticornis comprises only horn-cores (Fig. 22). Horn-cores The surface of the horn-cores bears continuous longitudinal ribs. Most of these are remarkably strong. The cross section is ovate to subovate. The horn-cores are slightly twisted in a clockwise direction. The horn-cores are curved
backwardly and weakly mediolaterly compressed. The mediolateral compression persists towards the apexes. Their medial face is convex at the base while the lateral one is slightly flat. The horn-cores become tapers towards the tips (Fig. 22(1-4)). The longitudinal furrows are strongly grooved anteriorly in PUPC 87/124. The dimensions of the horn-cores at the base are: PUPC 02/126 – DAP 32.4 mm, DT 27.6 mm; PUPC 87/180 – DAP 36 mm, DT 29 mm. The mean index at the base DT × 100/DAP for PUPC 87/124A is 72%, PUPC 02/126 is 85% and PUPC 87/124B is 80.5%. Comparison The morphometric characters of the sample allot the medium size antelope. The horn-core’s slight clockwise twist excludes the sample to Gazella (Fig. 22(1-4)). Nevertheless, the torsion in Gazella may be observed in the horn tip of some species of Gazella but this does not extend to the horn-cores (Pilgrim, 1939). The material is distinguished from G. rodleri in having clockwise twist because G. rodleri have anticlockwise twist (Pilgrim, 1939). The described horn-cores are clearly differentiated from those Antilope, Helicotragus and Lithocranius in absence of strong torsion (Fig. 22(1-4)). The overall direction, compression, slight twist and sculpture of the specimens are pretty fit for the species. The basal proportions of the studied horn-cores indicate variation in compression but this may be due to allometric growth of the horns (e.g. see description). The described sample is being closer to reduncines Kobus in regard to the degree of torsion, backward tilt, divergence and sculpture of the horn-cores. The studied specimens indicate have a faint keel than that of the early reduncines K. subdolus Gentry, 1980 and K. sp. 2 at Langebaanweg, the Redunca aff. darti of Lehmann, and K. porrecticornis (Lydekker, 1878) of the Siwaliks and a character that could be explained by the earlier stage of the antelope. However, the studied specimens differ from the common African reduncine by being more transversely compressed, having less of a flattened lateral surface, a less pronounced posterolateral angle and perhaps by having less strong longitudinal grooving posteriorly. The horn-cores are like the Siwaliks
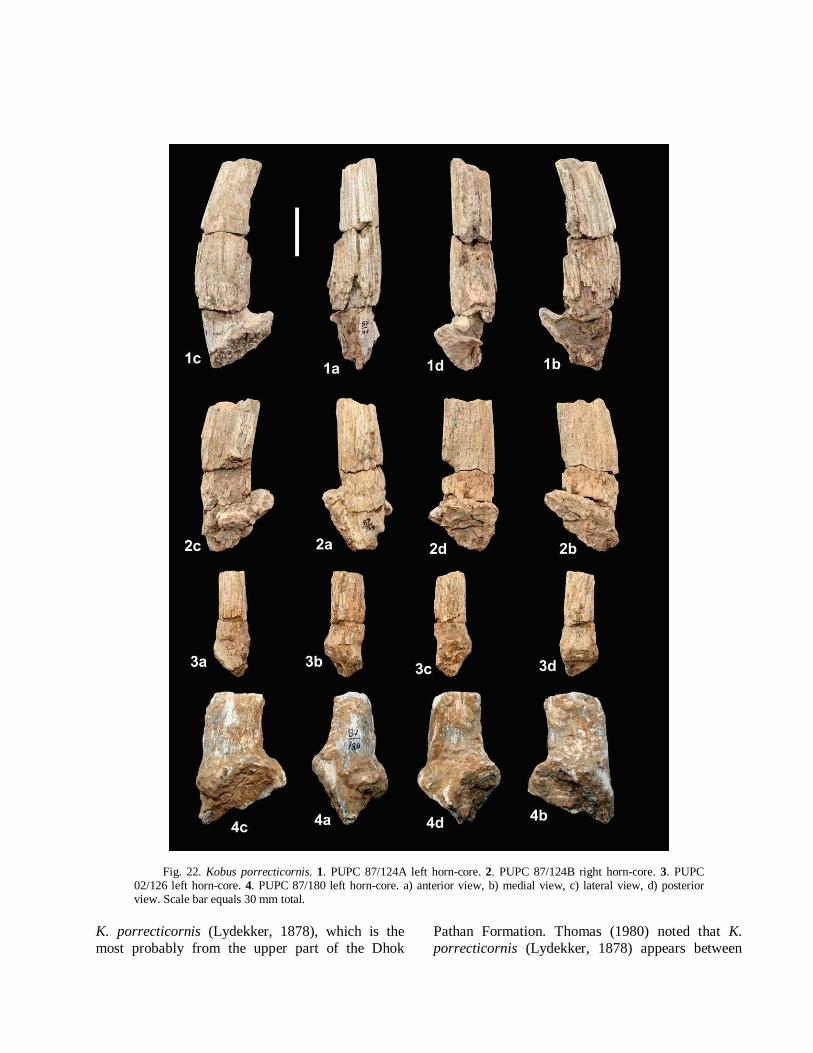
Fig. 22. Kobus porrecticornis. 1. PUPC 87/124A left horn-core. 2. PUPC 87/124B right horn-core. 3. PUPC 02/126 left horn-core. 4. PUPC 87/180 left horn-core. a) anterior view, b) medial view, c) lateral view, d) posterior view. Scale bar equals 30 mm total.
K. porrecticornis (Lydekker, 1878), which is the most probably from the upper part of the Dhok
Pathan Formation. Thomas (1980) noted that K. porrecticornis (Lydekker, 1878) appears between

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
49
sandstones 6 and 7 at the base of the Kundrali unit in the type zone of the Dhok Pathan. The findings of the studied horn-cores in the Dhok Pathan type locality confirm its stratigraphic occurrence in the upper part of the Middle Siwaliks. Discussion The common reduncine horn-cores vary quite a lot in size. They show little transverse compression, the lateral surface more flattened than the medial one, their widest mediolateral diameter lying rather anteriorly, a strong posterolateral angle, deep longitudinal grooving posteriorly, transverse ridges on their anterior surfaces, insertions above the backs of the orbits, insertions at a low angle in side view, much divergence, little backward curvature in their proximal parts, slight upward curvature distally, limited sinus development in the vicinity of the supraorbital pit but no sinuses in the pedicels, and deep postcornual fossae. The frontals are quite deep at the suture between the horn-core bases, and the dorsal part of the orbital rim projects quite strongly (Gentry, 1997). The braincase is moderately long, the cranial roof is steeply inclined but not curved downwards in its posterior part, the mastoids are moderately sized and face partly laterally, the ventral rim or edge just below the mastoid is weak and there is no dorsal rim (Gentry, 1985), the nuchal crest is quite strong and forms the effective boundary of the occipital surface, the occipital edge is not rounded in the form of a semicircle, a median vertical ridge was present on the occipital, the temporal lines approach fairly closely, and the auditory bulla was probably large. The main tooth characters of living reduncines are: moderate hypsodonty; teeth rather small in relation to skull and mandible size; upper and lower molars with entostyle/ectostylid; lingual lobes of upper molars and labial lobes of lowers constricted; upper molars with strong styles; localized ribs between the styles; central fossettes of upper molars with indentations (spurs) of their outlines and tending in later wear to have long, labially directed, transverse arms; lower molars with goat folds and front and rear outbowings of lingual walls; upper and lower P2s small; lower premolars with an appearance of anteroposterior compression; P4s with a strongly projecting hypoconid and often
a deep and narrow labial valley in front of it; p4s usually without paraconid-metaconid fusion to form a complete lingual wall (Gentry, 1997). In general earlier reduncine teeth are less distinctive than later ones in having lower crowned teeth, entostyle/ectostylid smaller or even absent, less constricted lingual lobes of upper molars and labial lobes of lower molars, ribs on upper molars less localized, central fossettes of upper molars less complicated, smaller goat folds on lower molars, lingual walls of lower molars less strongly outbowed, longer premolar rows and the entostylid more distinct from the entoconid on the back of p4. The Siwaliks K. porrecticornis indicates the earlier features of reduncines and is considered more primitive than those of African reduncines (Gentry, 1997). The Siwalik Kobus holotype GSI B229 named Dorcadoxa porrecticornis by Pilgrim in 1939 and placed in the Antilopini instead of Reduncini. Pilgrim (1939) had followed Lydekker and suggested a relationship of the holotype with Aepyceros considering it the first representative of African Aepyceros. He suggested that Dorcadoxa was the ancestral form of the living African Aepyceros and it may have been one of the early members of antelope, having migrated to Pakistan in the Miocene (Pilgrim, 1939). Gentry (1970) suggested a reduncine affinity. Thomas (1980) and Gentry (1980) treated the species as belonging to early member to Kobus. Horn-cores of K. subdolus, an earlier member of African Kobus may show an evolutionary advance over K. porrecticornis in that they are more fully reduncine in characters such as a less compressed cross section, a more flattened lateral surface, a more posterolateral keel, and a diminished backward curvature basally. The studied sample reflects the primitive features of reduncine and consequently, K. porrecticornis is the earliest member of Reduncini recovered from the sediments of Dhok Pathan type locality of the Dhok Pathan Formation.
cf. Kobikeryx sp. Pilgrim, 1939 Studied material Horn-cores: PUPC 99/49, distally preserved fragments, side indeterminate.

M.A. KHAN ET AL.
50
Description Horn-core It is not known from how far above the base it comes but it looks to break from the base (Fig. 23). The length of the fragment is 63 mm. The horn-core appears to be less compressed medio-laterally. The slight degree of compression probably increases above the base. There is no flattening of the surfaces of the horn-core and no keels. The horn-core is tilted back and the anterior border is slightly convex. The cross section is elongate oval without keels and the surface is ornamented with irregular longitudinal furrows. A prominent longitudinal furrow presents posteriorly (Fig. 23). The dimensions of the horn-core at the base are: PUPC 99/49 – DAP 25 mm, DT 20 mm giving the mean index DT × 100/DAP is 80%. The mean index at the base of already recovered specimens from the Dhok Pathan Formation (Pilgrim, 1939): for GSI B827 is 70%, for GSI B826 is 75.5% and for BMNH 15482 is 76.2%. Comparison and discussion The reduncines show little transverse compression, the lateral surface more flattened than the medial one, their widest mediolateral diameter lying rather anteriorly, a strong posterolateral angle, deep longitudinal grooving posteriorly, little backward curvature in their proximal parts (Gentry, 1997). The studied sample reflects the characteristics of reduncines and metrically accords well with the Siwalik Kobikeryx recovered from the Dhok Pathan Formation. However the material is not enough for the specific determination.
Subfamily Caprinae Gray, 1821
Tribe Caprini Gray, 1821
Genus CAPROTRAGOIDES Thenius, 1979 Type species Caprotragoides potwaricus (Pilgrim, 1939). Generic diagnosis (Pilgrim, 1939; Anzana and Morales, 1994) Small size, skull relatively wide at the orbits and narrow at the brain case; face bent down on the parietal surface at an angle of no more than 30°; horn-cores rather small, far apart, relatively rather
upright, inclined to the face at an angle of about 150° and to the parietal surface at an angle of about 60°, somewhat oblique to the skull axis, slightly curved to the rear, cross section elliptical, laterally compressed, without keels, slightly more convex on the medial than on the lateral side. Stratigraphic range Lower and Middle Siwaliks. Geographic distribution Middle Miocene to Pliocene of the Subcontinent and Eurasia (Pilgrim, 1939; Thomas, 1984; Kohler, 1987; Gentry, 1990).
Caprotragoides cf. potwaricus (Pilgrim, 1939)
Type specimen Fragmentary skull with the partial horn-cores: GSI B630. Emended abbreviated diagnosis (modified from 1939) Surfaces marked by very deep longitudinal grooving, particularly on the medial and posterior aspects; cross-section ovate, mediolaterally compressed with flattened lateral face; gently curved backwards; slight torsion (clockwise on right side), approach to a keel posteriorly and upright insertion. Studied material Horn-core: PC-GCUF 09/44, right side. Description Horn-core PC-GCUF 09/44 is the single most complete specimen, including a complete right horn core (Fig. 24). The horn-core is slightly curved backwards and tapers rapidly towards the tip. The cross section is ovate and slightly more convex on the medial than on the lateral side. The surface is covered with longitudinal grooves; some are comparatively deep medially and posteriorly (Fig. 24). Two deep longitudinal furrows, starting above the base, runs in parallel and become deep towards the tip of the horn-core approach to a keel posteriorly. The horn-core has slight twist along the grooves. The length

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
51
Fig. 23. cf. Kobikeryx sp. 1. PUPC 99/49 horn core, side indeterminate: a) anterior view, b) posterior view. Scale bar equals 30 mm total.
of the studied horn-core is 189 mm. The transverse diameter at the base is 28 mm and antero-posterior diameter at the base is 35 mm having mean index DT × 100/DAP is 80% at the base. The transverse diameter of the holotype (Pilgrim, 1939) at the base is 31.5 mm and the antero-posterior diameter at the base is 41 mm having mean index DT × 100/DAP is 77% at the base. Comparison This is one of the complete horn-core of Caprotragoides potwaricus ever recovered from the Siwaliks (Fig. 24). The specimen is much larger than the Siwalik gazelle. This specimen is immediately distinguished from Gazella by horn cores that are relatively much longer, and stouter. Horns inserted above the orbit; little inclined against the parietal surface; moderately divergent; gently curved backwards; with surfaces marked by very deep longitudinal grooving, particularly on the medial and posterior aspects; cross-section ovate (mediolateral compression) with slightly flattened
lateral face; with high pedicels, highest anteriorly. The medial surface is convex. The specimen is distinguished from African Gentrytragus and Tethytragus by its backwards curvature, and less mediolateral compression (Gentry, 1990; Bibi and Gulec, 2008). The specimen is a very good match of C. potwaricus based on horn core which is relatively long, curved backwards, moderately divergent, less stout and marked by deep longitudinal grooves. In Caprotragoides, the horn-cores are without keels, slightly compressed with a lateral surface a little flattened, slight backward curvature, the front of the pedicel higher than the back. Restricted sinuses are present in the frontols around the supraorbital pits and abutting the origin of horn-core pedicels. Tooth enamel is rugose, m1-2 have small or tiny ectostylid, those on m3 are smaller again. m3 never has a talonid fossette. Premolar rows are shorter than in Eotragus, otherwise the differences are almost non-existent. African Caprotragoides have larger and more hypsodont teeth than those of C. potwaricus (Gentry, 1970; Thomas, 1981).

M.A. KHAN ET AL.
52
Fig. 24. Caprotragoides cf. potwaricus. 1. PC-GCUF 09/44 right horn-core: a) anterior view, b) medial view, c) lateral view, d) posterior view. Scale bar equals 30 mm total.
Kubanotragus seems to occur as early in Asia as Eotragus in Europe (Chen, 1988) and has horn-cores of rounded cross section and slight or moderate torsion (clockwise on right side). However, both are without keels and have upright insertions (Gentry, 1990). Caprotragoides differed additionally from Kubanotragus by the hypsodonty of its cheek teeth. Caprotragoides species from Ngorora was larger and showed dental advances, such as a more complicated outline of central cavities on its molars and paraconid-metaconid fusion on p4 (Thomas, 1981). An earlier Caprotragoides from Nyakach had an approach to a keel on its horn-core and thereby suggested a relationship to Benicerus looks as if it had a more inclined cranial roof than did Caprotragoides. The African Pachytragus differs from Caprotragoides in
the inclined braincase roof, less upright horn-core insertions in side view, and less hypsodont teeth, and is difficult to envisage as a direct descendent of that Caprotragoides (Gentry, 1990). Discussion Caprotragoides is a non boselaphine bovid, founded for Hippotraginae gen. indet. (cf. Tragoreas) potwaricus Pilgrim, 1939, based on a frontlet from the Nagri Formation of the Siwaliks (Thomas, 1984). Anzana and Morales (1994) proposed the classification and new nomenclature for some mainly middle Miocene bovids. They ascribed C. potwaricus (Pilgrim, 1939) type species for the genus, founded from the middle–late Miocene of India and Pakistan. Thomas (1984) recovered C. potwaricus from the pre-Hipparion

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
53
sediments of Indian Siwaliks. The holotype frontlet of C. potwaricus and some records of C. gentri already extend into the upper Miocene. The present sample found in the Dhok Pathan type locality which belongs to upper Miocene (Barry et al., 2002) extends its presence in the Dhok Pathan Formation of the Middle Siwaliks. Prior to this, it is only known from the Nagri Formation of the Middle Siwaliks. Caprotragoides (Caprini) is the earliest bovid other than Eotragus, Miotragocerus and Homoiodorcas (Gentry, 1990; Thomas, 1984). These occur in pre-Hipparion faunas of Asia, Africa, and Europe. Caprotragoides has a similar relationship to Protoryx and Caprini (Kohler, 1987) as does Homoiodorcas to the Antilopini. Hence, if Boselaphini, Antilopini and Caprini are successive branches of bovids (Gentry, 1992), the same would be true of Eotragus, Homoiodorcas and Caprotragoides. Caprotragoides shares an immediate common ancestory with Homoiodorcas (Gentry, 1994).
Family Cervidae Goldfuss, 1820 Subfamily Cervinae Goldfuss, 1820
Tribe Cervini Weber, 1928
Genus CERVUS Linnaeus, 1758
Type species Cervus elaphus Linnaeus, 1758. Generic diagnosis (Colbert, 1935; Gentry et al., 1999) The cheek teeth evolved a greater crown height, fusion of the outer cristae, selenodont as well as intensified and higher. The styles are stronger than the stylids and the first molar is brachydont. The goat fold and neocrista are either in a vestigial form or absent. The upper premolars show molarization by becoming more rounded and retaining only a weak incision on the lingual wall. The ectostylid is present. Stratigraphic range Middle and Upper Siwaliks. Geographic distribution Asia, North America and Europe (Romer,
1974; Savage and Russel, 1983; Pitra et al., 2004).
Cervus cf. sivalensis Lydekker, 1884
Type specimen A fragment of right mandible with m2-3: GSI B215. Diagnosis (Colbert, 1935; Khan, 2007) A large cervid with relatively hypsodont molars. The skull and antlers resemble these portions in Cervus duvaucelli. A skull with frontal concavity at the orbits, and the forward swell at the pedcels. The lacrymal vacuity is smaller than in C. duvaucelli. The brow tines of the antler arise immediately above the burr, and form an obtuse angle with the beam. Studied material Lower dentition: PUPC 04/18, right mandibular ramus with m2-3. PUPC 09/73, right mandibular ramus with m2. Description Lower dentition The molars are in middle wear and the length of the mandibular ramus PUPC 04/18 is 87 mm (Fig. 25(1)). The major conids with the roots show the exact morphology of the molars. The conids are preserved up to their pointed ends, which are running laterally towards their respective cristids. The dentine is exposed over the crown surface. The crown shows the comparatively strong accessory columns. The ectostylid is strong and presents up to the middle of the transverse valley. The ectostylid, anteriorly, is connected to the postprotocristid and to the praehypocristid posteriorly. The median ribs are weak and the mesostylid seems to be absent. The metastylid and the entostylid are present on the lingual side and the metastylid is stronger than the entostylid, which is projected outwardly near the summit of the crown (Fig. 25(1)). The anterior and the posterior fossettes are prominent. The anterior fossette is formed by the union of the protoconid and the metaconid, while the posterior fossette is formed by the union of the hypoconid and the entoconid. The hypoconulid is broad and it is constricted posteriorly. The lateral lobes are not
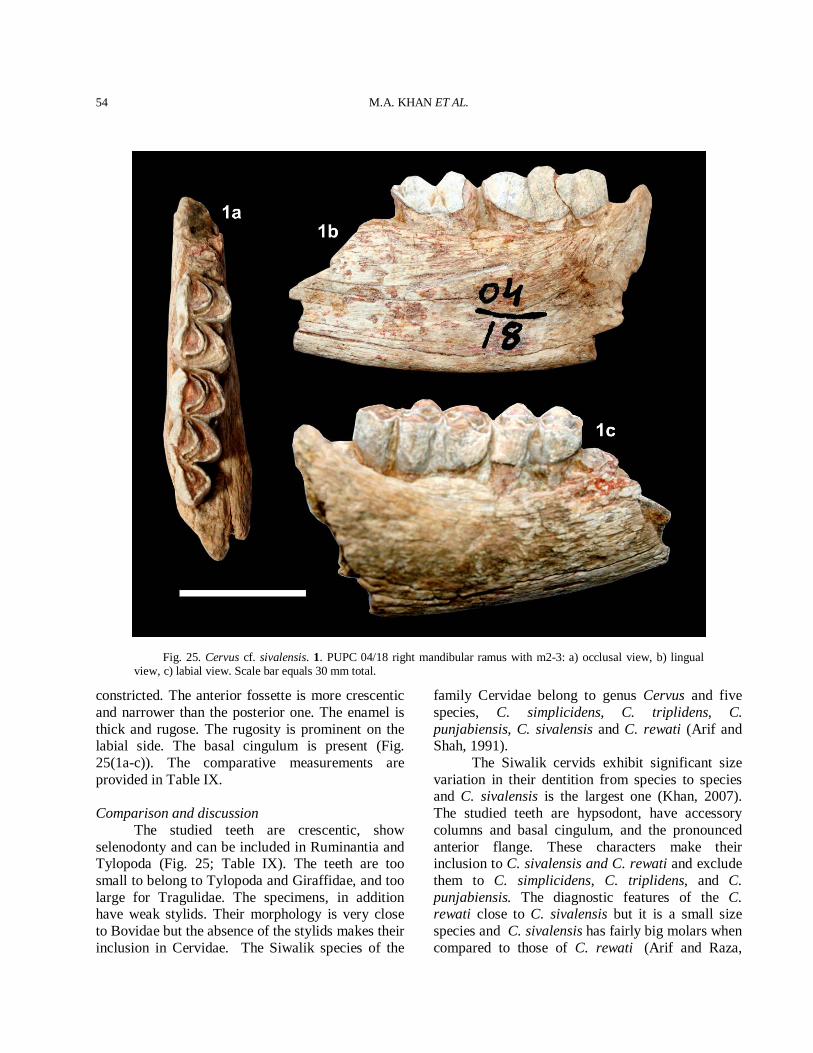
M.A. KHAN ET AL.
54
Fig. 25. Cervus cf. sivalensis. 1. PUPC 04/18 right mandibular ramus with m2-3: a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
constricted. The anterior fossette is more crescentic and narrower than the posterior one. The enamel is thick and rugose. The rugosity is prominent on the labial side. The basal cingulum is present (Fig. 25(1a-c)). The comparative measurements are provided in Table IX. Comparison and discussion The studied teeth are crescentic, show selenodonty and can be included in Ruminantia and Tylopoda (Fig. 25; Table IX). The teeth are too small to belong to Tylopoda and Giraffidae, and too large for Tragulidae. The specimens, in addition have weak stylids. Their morphology is very close to Bovidae but the absence of the stylids makes their inclusion in Cervidae. The Siwalik species of the
family Cervidae belong to genus Cervus and five species, C. simplicidens, C. triplidens, C. punjabiensis, C. sivalensis and C. rewati (Arif and Shah, 1991). The Siwalik cervids exhibit significant size variation in their dentition from species to species and C. sivalensis is the largest one (Khan, 2007). The studied teeth are hypsodont, have accessory columns and basal cingulum, and the pronounced anterior flange. These characters make their inclusion to C. sivalensis and C. rewati and exclude them to C. simplicidens, C. triplidens, and C. punjabiensis. The diagnostic features of the C. rewati close to C. sivalensis but it is a small size species and C. sivalensis has fairly big molars when compared to those of C. rewati (Arif and Raza,
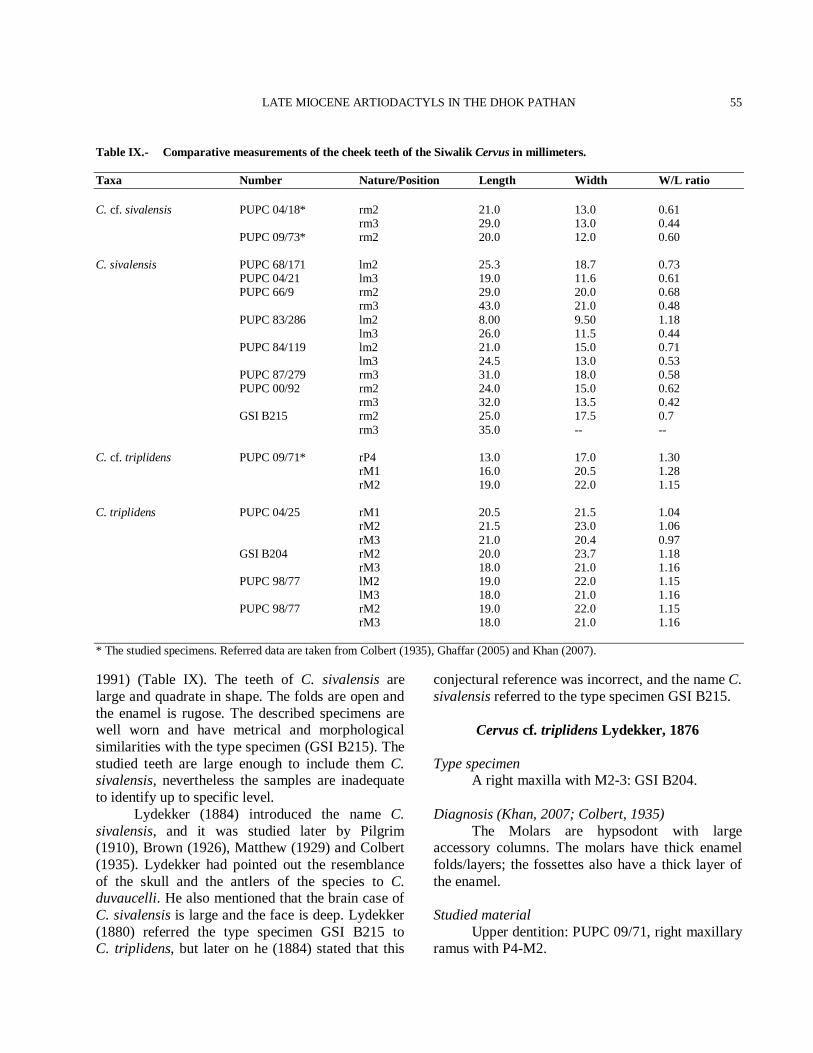
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
55
Table IX.- Comparative measurements of the cheek teeth of the Siwalik Cervus in millimeters. Taxa Number Nature/Position Length Width W/L ratio C. cf. sivalensis PUPC 04/18* rm2 21.0 13.0 0.61 rm3 29.0 13.0 0.44 PUPC 09/73* rm2 20.0 12.0 0.60 C. sivalensis PUPC 68/171 lm2 25.3 18.7 0.73 PUPC 04/21 lm3 19.0 11.6 0.61 PUPC 66/9 rm2 29.0 20.0 0.68 rm3 43.0 21.0 0.48 PUPC 83/286 lm2 8.00 9.50 1.18 lm3 26.0 11.5 0.44 PUPC 84/119 lm2 21.0 15.0 0.71 lm3 24.5 13.0 0.53 PUPC 87/279 rm3 31.0 18.0 0.58 PUPC 00/92 rm2 24.0 15.0 0.62 rm3 32.0 13.5 0.42 GSI B215 rm2 25.0 17.5 0.7 rm3 35.0 -- -- C. cf. triplidens PUPC 09/71* rP4 13.0 17.0 1.30 rM1 16.0 20.5 1.28 rM2 19.0 22.0 1.15 C. triplidens PUPC 04/25 rM1 20.5 21.5 1.04 rM2 21.5 23.0 1.06 rM3 21.0 20.4 0.97 GSI B204 rM2 20.0 23.7 1.18 rM3 18.0 21.0 1.16 PUPC 98/77 lM2 19.0 22.0 1.15 lM3 18.0 21.0 1.16 PUPC 98/77 rM2 19.0 22.0 1.15 rM3 18.0 21.0 1.16 * The studied specimens. Referred data are taken from Colbert (1935), Ghaffar (2005) and Khan (2007). 1991) (Table IX). The teeth of C. sivalensis are large and quadrate in shape. The folds are open and the enamel is rugose. The described specimens are well worn and have metrical and morphological similarities with the type specimen (GSI B215). The studied teeth are large enough to include them C. sivalensis, nevertheless the samples are inadequate to identify up to specific level. Lydekker (1884) introduced the name C. sivalensis, and it was studied later by Pilgrim (1910), Brown (1926), Matthew (1929) and Colbert (1935). Lydekker had pointed out the resemblance of the skull and the antlers of the species to C. duvaucelli. He also mentioned that the brain case of C. sivalensis is large and the face is deep. Lydekker (1880) referred the type specimen GSI B215 to C. triplidens, but later on he (1884) stated that this
conjectural reference was incorrect, and the name C. sivalensis referred to the type specimen GSI B215.
Cervus cf. triplidens Lydekker, 1876 Type specimen A right maxilla with M2-3: GSI B204. Diagnosis (Khan, 2007; Colbert, 1935) The Molars are hypsodont with large accessory columns. The molars have thick enamel folds/layers; the fossettes also have a thick layer of the enamel. Studied material Upper dentition: PUPC 09/71, right maxillary ramus with P4-M2.
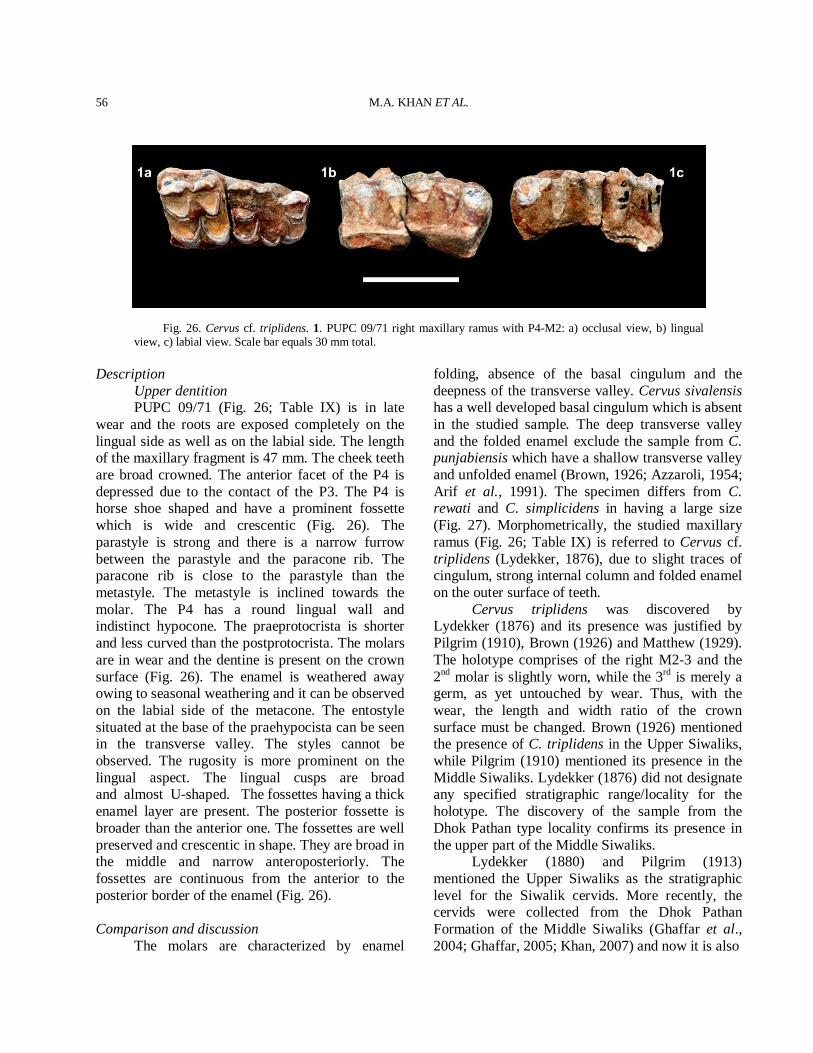
M.A. KHAN ET AL.
56
Fig. 26. Cervus cf. triplidens. 1. PUPC 09/71 right maxillary ramus with P4-M2: a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
Description Upper dentition PUPC 09/71 (Fig. 26; Table IX) is in late wear and the roots are exposed completely on the lingual side as well as on the labial side. The length of the maxillary fragment is 47 mm. The cheek teeth are broad crowned. The anterior facet of the P4 is depressed due to the contact of the P3. The P4 is horse shoe shaped and have a prominent fossette which is wide and crescentic (Fig. 26). The parastyle is strong and there is a narrow furrow between the parastyle and the paracone rib. The paracone rib is close to the parastyle than the metastyle. The metastyle is inclined towards the molar. The P4 has a round lingual wall and indistinct hypocone. The praeprotocrista is shorter and less curved than the postprotocrista. The molars are in wear and the dentine is present on the crown surface (Fig. 26). The enamel is weathered away owing to seasonal weathering and it can be observed on the labial side of the metacone. The entostyle situated at the base of the praehypocista can be seen in the transverse valley. The styles cannot be observed. The rugosity is more prominent on the lingual aspect. The lingual cusps are broad and almost U-shaped. The fossettes having a thick enamel layer are present. The posterior fossette is broader than the anterior one. The fossettes are well preserved and crescentic in shape. They are broad in the middle and narrow anteroposteriorly. The fossettes are continuous from the anterior to the posterior border of the enamel (Fig. 26). Comparison and discussion The molars are characterized by enamel
folding, absence of the basal cingulum and the deepness of the transverse valley. Cervus sivalensis has a well developed basal cingulum which is absent in the studied sample. The deep transverse valley and the folded enamel exclude the sample from C. punjabiensis which have a shallow transverse valley and unfolded enamel (Brown, 1926; Azzaroli, 1954; Arif et al., 1991). The specimen differs from C. rewati and C. simplicidens in having a large size (Fig. 27). Morphometrically, the studied maxillary ramus (Fig. 26; Table IX) is referred to Cervus cf. triplidens (Lydekker, 1876), due to slight traces of cingulum, strong internal column and folded enamel on the outer surface of teeth. Cervus triplidens was discovered by Lydekker (1876) and its presence was justified by Pilgrim (1910), Brown (1926) and Matthew (1929). The holotype comprises of the right M2-3 and the 2nd molar is slightly worn, while the 3rd is merely a germ, as yet untouched by wear. Thus, with the wear, the length and width ratio of the crown surface must be changed. Brown (1926) mentioned the presence of C. triplidens in the Upper Siwaliks, while Pilgrim (1910) mentioned its presence in the Middle Siwaliks. Lydekker (1876) did not designate any specified stratigraphic range/locality for the holotype. The discovery of the sample from the Dhok Pathan type locality confirms its presence in the upper part of the Middle Siwaliks. Lydekker (1880) and Pilgrim (1913) mentioned the Upper Siwaliks as the stratigraphic level for the Siwalik cervids. More recently, the cervids were collected from the Dhok Pathan Formation of the Middle Siwaliks (Ghaffar et al., 2004; Ghaffar, 2005; Khan, 2007) and now it is also
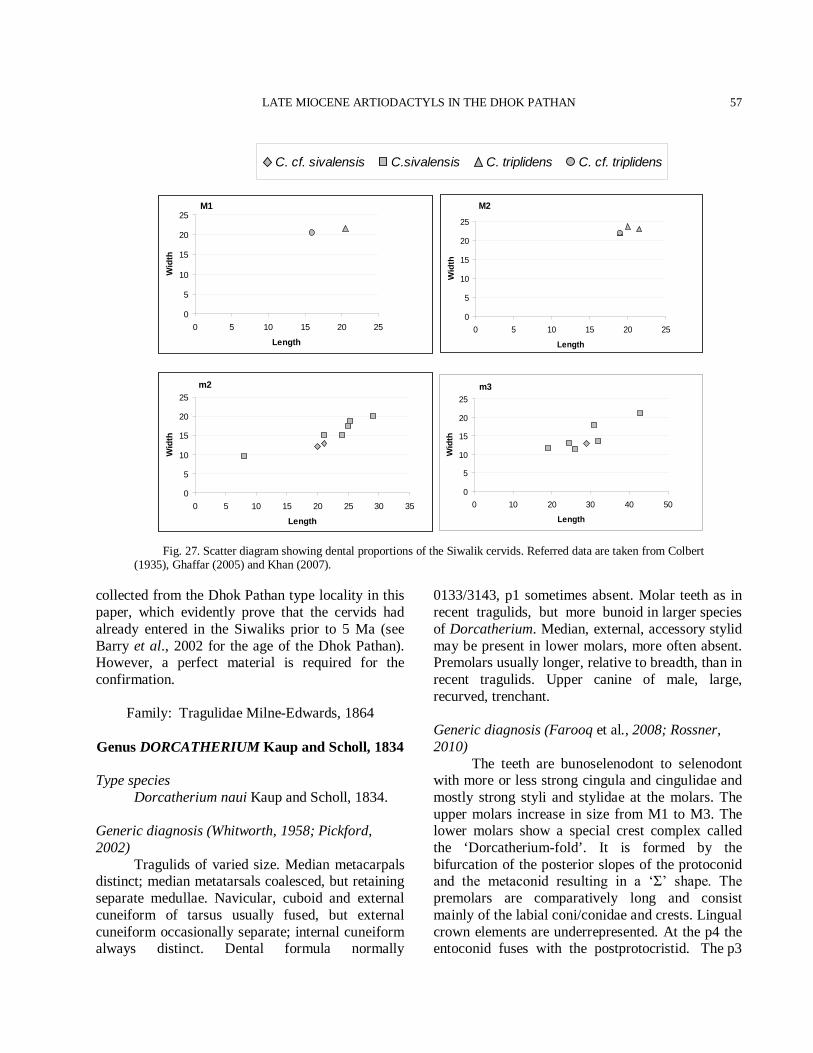
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
57
Fig. 27. Scatter diagram showing dental proportions of the Siwalik cervids. Referred data are taken from Colbert (1935), Ghaffar (2005) and Khan (2007).
collected from the Dhok Pathan type locality in this paper, which evidently prove that the cervids had already entered in the Siwaliks prior to 5 Ma (see Barry et al., 2002 for the age of the Dhok Pathan). However, a perfect material is required for the confirmation.
Family: Tragulidae Milne-Edwards, 1864 Genus DORCATHERIUM Kaup and Scholl, 1834
Type species Dorcatherium naui Kaup and Scholl, 1834. Generic diagnosis (Whitworth, 1958; Pickford, 2002) Tragulids of varied size. Median metacarpals distinct; median metatarsals coalesced, but retaining separate medullae. Navicular, cuboid and external cuneiform of tarsus usually fused, but external cuneiform occasionally separate; internal cuneiform always distinct. Dental formula normally
0133/3143, p1 sometimes absent. Molar teeth as in recent tragulids, but more bunoid in larger species of Dorcatherium. Median, external, accessory stylid may be present in lower molars, more often absent. Premolars usually longer, relative to breadth, than in recent tragulids. Upper canine of male, large, recurved, trenchant. Generic diagnosis (Farooq et al., 2008; Rossner, 2010) The teeth are bunoselenodont to selenodont with more or less strong cingula and cingulidae and mostly strong styli and stylidae at the molars. The upper molars increase in size from M1 to M3. The lower molars show a special crest complex called the ‘Dorcatherium-fold’. It is formed by the bifurcation of the posterior slopes of the protoconid and the metaconid resulting in a ‘Σ’ shape. The premolars are comparatively long and consist mainly of the labial coni/conidae and crests. Lingual crown elements are underrepresented. At the p4 the entoconid fuses with the postprotocristid. The p3
m2
0
5
10
15
20
25
0 5 10 15 20 25 30 35
Length
Wid
th
M1
0
5
10
15
20
25
0 5 10 15 20 25
Length
Wid
th
M2
0
5
10
15
20
25
0 5 10 15 20 25
Length
Wid
th
m3
0
5
10
15
20
25
0 10 20 30 40 50
Length
Wid
th
C. cf. sivalensis C.sivalensis C. triplidens C. cf. triplidens
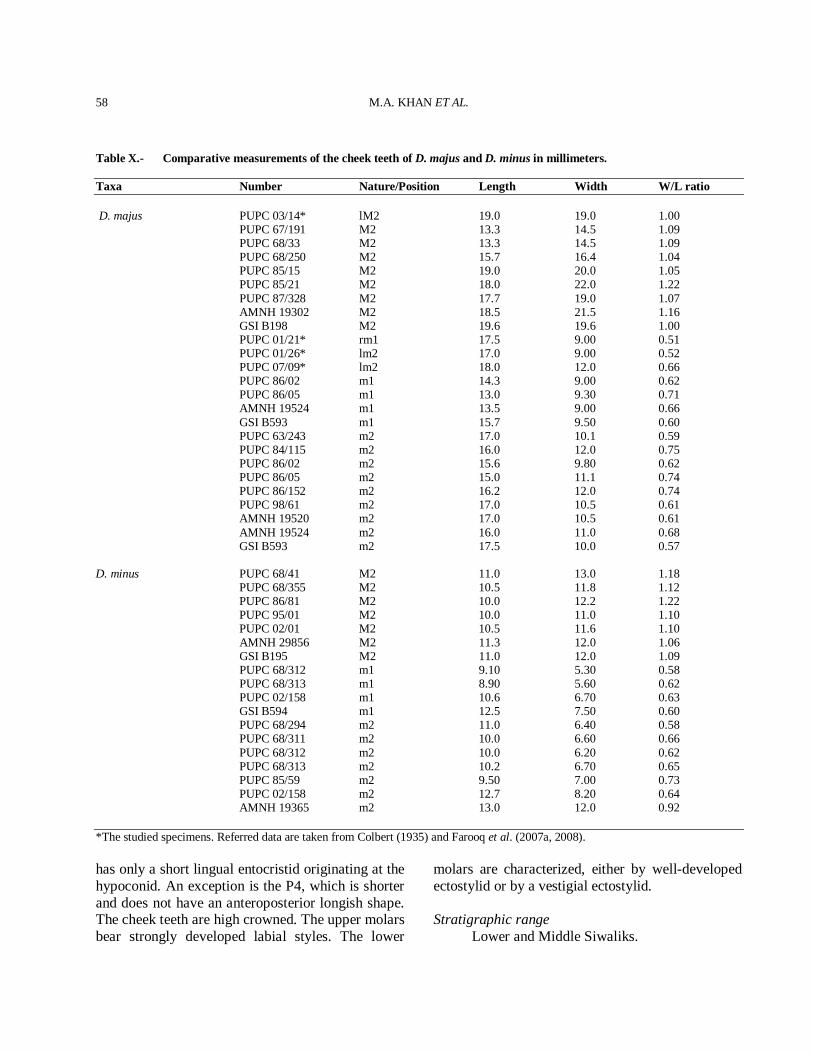
M.A. KHAN ET AL.
58
Table X.- Comparative measurements of the cheek teeth of D. majus and D. minus in millimeters. Taxa Number Nature/Position Length Width W/L ratio D. majus PUPC 03/14* lM2 19.0 19.0 1.00 PUPC 67/191 M2 13.3 14.5 1.09 PUPC 68/33 M2 13.3 14.5 1.09 PUPC 68/250 M2 15.7 16.4 1.04 PUPC 85/15 M2 19.0 20.0 1.05 PUPC 85/21 M2 18.0 22.0 1.22 PUPC 87/328 M2 17.7 19.0 1.07 AMNH 19302 M2 18.5 21.5 1.16 GSI B198 M2 19.6 19.6 1.00 PUPC 01/21* rm1 17.5 9.00 0.51 PUPC 01/26* lm2 17.0 9.00 0.52 PUPC 07/09* lm2 18.0 12.0 0.66 PUPC 86/02 m1 14.3 9.00 0.62 PUPC 86/05 m1 13.0 9.30 0.71 AMNH 19524 m1 13.5 9.00 0.66 GSI B593 m1 15.7 9.50 0.60 PUPC 63/243 m2 17.0 10.1 0.59 PUPC 84/115 m2 16.0 12.0 0.75 PUPC 86/02 m2 15.6 9.80 0.62 PUPC 86/05 m2 15.0 11.1 0.74 PUPC 86/152 m2 16.2 12.0 0.74 PUPC 98/61 m2 17.0 10.5 0.61 AMNH 19520 m2 17.0 10.5 0.61 AMNH 19524 m2 16.0 11.0 0.68 GSI B593 m2 17.5 10.0 0.57 D. minus PUPC 68/41 M2 11.0 13.0 1.18 PUPC 68/355 M2 10.5 11.8 1.12 PUPC 86/81 M2 10.0 12.2 1.22 PUPC 95/01 M2 10.0 11.0 1.10 PUPC 02/01 M2 10.5 11.6 1.10 AMNH 29856 M2 11.3 12.0 1.06 GSI B195 M2 11.0 12.0 1.09 PUPC 68/312 m1 9.10 5.30 0.58 PUPC 68/313 m1 8.90 5.60 0.62 PUPC 02/158 m1 10.6 6.70 0.63 GSI B594 m1 12.5 7.50 0.60 PUPC 68/294 m2 11.0 6.40 0.58 PUPC 68/311 m2 10.0 6.60 0.66 PUPC 68/312 m2 10.0 6.20 0.62 PUPC 68/313 m2 10.2 6.70 0.65 PUPC 85/59 m2 9.50 7.00 0.73 PUPC 02/158 m2 12.7 8.20 0.64 AMNH 19365 m2 13.0 12.0 0.92 *The studied specimens. Referred data are taken from Colbert (1935) and Farooq et al. (2007a, 2008). has only a short lingual entocristid originating at the hypoconid. An exception is the P4, which is shorter and does not have an anteroposterior longish shape. The cheek teeth are high crowned. The upper molars bear strongly developed labial styles. The lower
molars are characterized, either by well-developed ectostylid or by a vestigial ectostylid. Stratigraphic range Lower and Middle Siwaliks.

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
59
Geographic distribution Lower to Middle Siwaliks (Middle Miocene to early Pliocene) (Farooq et al., 2007a), MN4 (late Early Miocene, early Karpatian, middle Orleanian)
(Sach and Heizmann, 2001) to MN13 (latest Late Miocene, late Pontian or early Dacian, late Turolian) (Arambourg and Piveteau, 1929), eventually Pliocene (Bakalov and Nikolov, 1962), in Europe, Middle Miocene (Qiu and Gu, 1991) to Early Pliocene (Lydekker, 1876) in Asia, late Early Miocene (Pickford, 2001) to Early Pliocene (Pickford et al., 2004) in Africa.
Dorcatherium majus Lydekker, 1876
Type specimen Two upper molars: GSI B197. Diagnosis (Farooq et al., 2008) Dorcatherium majus is a species larger than Dorcatherium minus and is equal in size to Dorcabune anthracotherioides. It is characterized by strong parastyle and mesostyle, well-developed cingulum in upper molars and stoutly developed ectostylid. Studied material Upper dentition: PUPC 03/14, isolated left M2. Lower dentition: PUPC 01/21, isolated right m1. PUPC 01/26, left mandibular ramus with m2 and posterior portion of m1. PUPC 07/09, isolated left m2. Description Upper dentition The molar is almost quadrate in shape (Table X) and in an early wear (Fig. 28(1)). The basal shelf like cingulum surrounds the lingual part of the molar and it is moderately developed at the antero-posterior face of the molar. It is present at the entrance of the transverse valley between the protocone and the hypocone and it is entirely absent around the labial cones. The entostyle is absent (Fig. 28(1)). The enamel is wrinkled and shows a uniform thickness all over the crown of the molar. The anterior and the posterior cavities are wide and deep. The principal cusps are inclined towards the median longitudinal line of the molar and this
inclination is more conspicuous in the lingual cusps than those of the labial ones. The protocone is more worn than the other ones. It exhibits semi-crescentic shape, as the praeprotocrista is longer than the postprotocrista. The innermost lingual part of the protocone is not pointed. The paracone is relatively smaller in antero-posterior length than the metacone. The antero-buccal cusp has a strong anterior groove descending from its apex to the base of the crown, which separates the anterior style from the main cusp. The buccal notch is V-shaped. The parastyle is very well developed and connected to the base of the anterior median rib. The anterior median rib is more prominent than the posterior one. The mesostyle is very well developed and it is in the form of an isolated pillar. The metastyle is almost equal to that of parastyle in height and its basal part is broken. The hypocone is crescentic in shape. The posthypocrista is thin than the praehypocrista. The posthypocrista enters the metaconus leaving the metastyle outside the fossette and making the shape asymmetrical. Labial styles and columns are strong (Fig. 28(1)). Lower dentition The Dorcatherium-fold is present on the distal surface of the antero-lingul conids in the lower molars (28(2-4)). The antero-lingual cusp reaches forwards to join a small accessory cusplet in the midline, which gives a pointed contour to the front of the crown. On its distal surface there is a low crest that descends obliquely towards the midline where it joins the anterior crest of the disto-buccal cusp. The metaconid and the entoconid are higher than those of the protoconid and the hypoconid. The enamel is fairly thick and uniform in thickness. The cingulid is present on the posterior as well as on the anterior side. It can be observed around the hypoconid. The cingulid is weakly developed on the labial side especially at the entrance of the transverse valley along the praehypocristid. A small ectostylid is present at the entrance of the transverse valley towards the labial side, and it is closely associated with the postprotocristid. The central cavities are narrow. The protoconid is anteroposteriorly longer than the metaconid. It is crescentic in shape with the praeprotocristid and the postprotocristid. The post-
M.A. KHAN ET AL.
60
Fig. 28. Dorcatherium majus. 1. PUPC 03/14 isolated left M2. 2. PUPC 01/21 isolated right m1. 3. PUPC 07/09 isolated left m2. 4. PUPC 01/26 left mandibular ramus with m2 and posterior portion of m1. a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
protocristid is bifurcated at the point where it is attached to the postmetacristid. The dentinal islet of the protoconid is roughly triangular. The metaconid is pointed at the middle with the anterior and the posterior sloping cristids. The postmetacristid is also bifurcated in PUPC 01/21 (Fig. 28(2)) and PUPC 07/09 (Fig. 28(2)) and is slightly more worn than the praemetacristid. The praemetacristid has produced a
narrow and elongated dentinal islet. The entoconid is well developed but it is slightly damaged at the posterior side. The hypoconid is more crescentic than the protoconid. The comparative measurements are provided in Table X. Comparison and discussion Morphologically, the specimens show

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
61
typically tragulid features. The bunoselenodonty, isolated strong styles, basal cingulum, prominent anterior median ribs and the presence of Dorcatherium-fold full corroborate the tragulid status of the Dhok Pathan type locality fossils (Fig. 28). The upper molars of Dorcatherium majus are characterized by subhypsodont and selenodont teeth with isolated pillar like parastyle. The metastyle is always found weakly formed (Lydekker 1876; Pilgrim, 1915; Colbert, 1935; Farooq et al., 2007a; Rossner, 2010). The lower molars of D. majus are high comparatively and narrow crowned. They have ectostylids between the protoconids and the hypoconids. The metaconid is recognized as the highest one among all the conids. Stylids are weakly or improperly developed. Their anterior median ribs are well pronounced. All these characteristics exhibited by the present collection of lower dentition indicate very clearly that it belongs to the species D. majus (Colbert, 1935; Farooq et al., 2008). The material assigned to this species is morphometrically (Fig. 28; Table X) similar to the specimens from the Siwaliks described by Colbert (1935) and Farooq et al. (2007a, 2008). The first undoubted fossil tragulid is the early Miocene genus Dorcatherium from Africa and Eurasia. The extinct genus Dorcatherium is known with many species from Asia (Lydekker, 1876; Matthew, 1929; Farooq et al., 2007a, 2008), Europe (Rossner, 2010) and Africa (Pickford, 2001, 2002; Pickford et al., 2004). The numerous species referred to Dorcatherium mainly differ by their size (West 1980). Four Pakistani species of the genus Dorcatherium are D. minimus, D. minus, D. nagrii, and D. majus. D. minimus, the smallest species, known only by isolated upper third molar from the Chinji Formation of the Lower Siwaliks, Pakistan and D. nagrii from the Nagri Formation of the Siwaliks are rare found and probable invalid species. Dorcatherium majus and D. minus appears consistently in the Lower and the Middle Siwaliks and metrically, these are clearly distinguished to each other in size (Table X; Fig. 29) (Farooq, 2006, Farooq et al., 2007a, 2008).
Genus DORCABUNE Pilgrim, 1910
Type species Dorcabune anthracotherioides Pilgrim, 1910.
Generic diagnosis (Pilgrim, 1910, 1915; Colbert, 1935; Farooq et al., 2007b) Very large tragulids having bunodont teeth. Isolated parastyle and mesostyle, prominent cingulum and enamel rugosity are the diagnostic characteristics of the upper molars, whereas lower molars are characterized by their broadness, a wide talonid in the third molar, and a pyramidal protoconid with two posteriorly directed folds. The upper molars of Dorcabune anthracotherioides are characterized by their brachydonty and bunodonty. The inner cusps of upper molars are truly selenodont, whereas the outer ones are quite bunodont and absolutely conical in their general appearance. The median rib on the labial face of the paracone and metacone is so broad and prominent that it occupies almost all the space between the styles. This feature is very much pronounced in the paracone, the labial surface of which is in fact entirely rib. The parastyle and mesostyle are strong, massive and isolated, whereas the metastyle is very weakly developed. With wear, the mesostyle clearly displays its closer association with the metacone instead of fusing equally to both paracone and metacone. The protocone, instead of being a simple crescent, is more pyramidal in shape and displays three equally strong folds, one proceeding forwards and outwards, the second backwards and a third backwards with a tendency sometimes inwards and sometimes outwards. A strong cingulum runs antero-posteriorly, but is very much pronounced round the protocone. It often rises into a small tubercle at the entrance of the transverse valley between the protocone and hypocone. The enamel is heavy and has moderately fine rugosity (Pilgrim, 1915; Colbert, 1935). The lower molars are also characterized by well pronounced brachydonty and bunodonty. The anterior arm of protoconid terminates on a broad shelf almost parallel to the anterior margin of the tooth. Entoconid is conical, producing out anteriorly a short process in the direction of the mid line between the two anterior cusps. The hypoconid is crescentic; its anterior arm touches to the external process of the protoconid, while its posterior arm runs inward and completely encircles the posterior base of the entoconid.
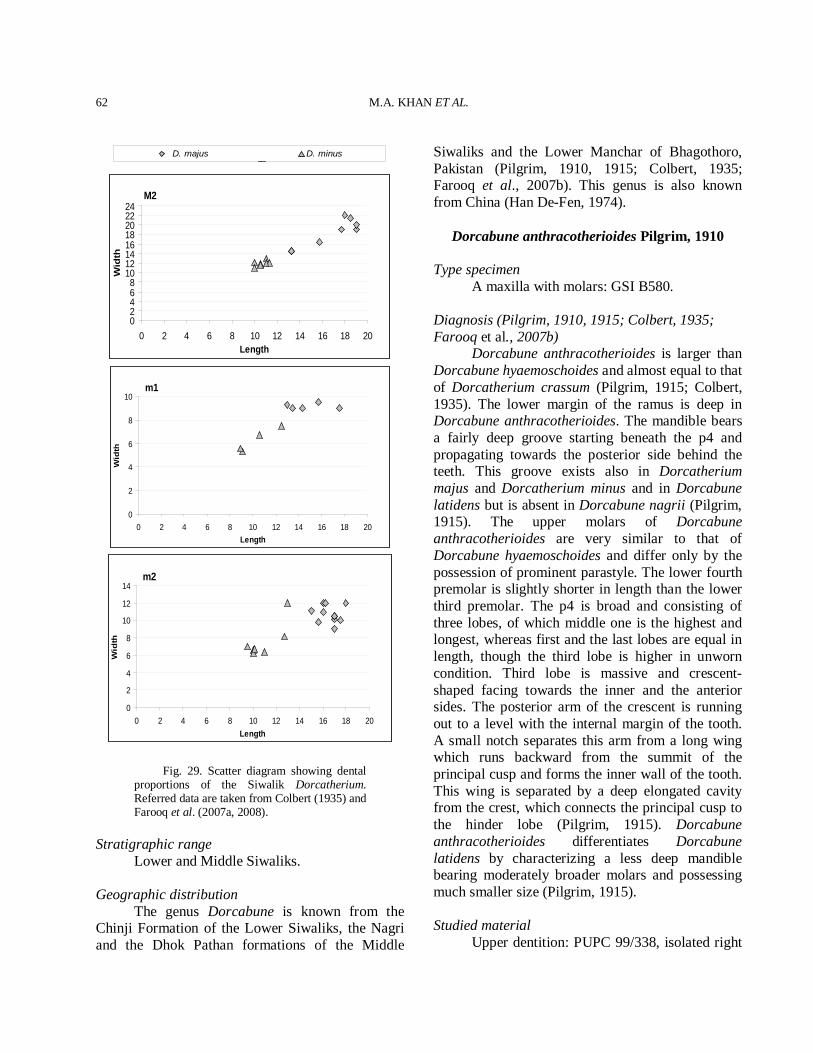
M.A. KHAN ET AL.
62
M2
02468
1012141618202224
0 2 4 6 8 10 12 14 16 18 20Length
Wid
th
m1
0
2
4
6
8
10
0 2 4 6 8 10 12 14 16 18 20Length
Wid
th
m2
0
2
4
6
8
10
12
14
0 2 4 6 8 10 12 14 16 18 20Length
Wid
th
Fig. 29. Scatter diagram showing dental proportions of the Siwalik Dorcatherium. Referred data are taken from Colbert (1935) and Farooq et al. (2007a, 2008).
Stratigraphic range Lower and Middle Siwaliks. Geographic distribution The genus Dorcabune is known from the Chinji Formation of the Lower Siwaliks, the Nagri and the Dhok Pathan formations of the Middle
Siwaliks and the Lower Manchar of Bhagothoro, Pakistan (Pilgrim, 1910, 1915; Colbert, 1935; Farooq et al., 2007b). This genus is also known from China (Han De-Fen, 1974).
Dorcabune anthracotherioides Pilgrim, 1910
Type specimen A maxilla with molars: GSI B580. Diagnosis (Pilgrim, 1910, 1915; Colbert, 1935; Farooq et al., 2007b) Dorcabune anthracotherioides is larger than Dorcabune hyaemoschoides and almost equal to that of Dorcatherium crassum (Pilgrim, 1915; Colbert, 1935). The lower margin of the ramus is deep in Dorcabune anthracotherioides. The mandible bears a fairly deep groove starting beneath the p4 and propagating towards the posterior side behind the teeth. This groove exists also in Dorcatherium majus and Dorcatherium minus and in Dorcabune latidens but is absent in Dorcabune nagrii (Pilgrim, 1915). The upper molars of Dorcabune anthracotherioides are very similar to that of Dorcabune hyaemoschoides and differ only by the possession of prominent parastyle. The lower fourth premolar is slightly shorter in length than the lower third premolar. The p4 is broad and consisting of three lobes, of which middle one is the highest and longest, whereas first and the last lobes are equal in length, though the third lobe is higher in unworn condition. Third lobe is massive and crescent-shaped facing towards the inner and the anterior sides. The posterior arm of the crescent is running out to a level with the internal margin of the tooth. A small notch separates this arm from a long wing which runs backward from the summit of the principal cusp and forms the inner wall of the tooth. This wing is separated by a deep elongated cavity from the crest, which connects the principal cusp to the hinder lobe (Pilgrim, 1915). Dorcabune anthracotherioides differentiates Dorcabune latidens by characterizing a less deep mandible bearing moderately broader molars and possessing much smaller size (Pilgrim, 1915). Studied material Upper dentition: PUPC 99/338, isolated right
D. majus D. minus

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
63
M2. Lower dentition: PUPC 04/01, right mandibular ramus with m3. Description Upper dentition The molar is in an early wear, squarish, bunodont and broad crowned (Fig. 30(1); Table XI). The enamel is extensively wrinkled and thick. The cingulum is moderately developed at the antero-posterior face of the tooth and heavily developed at the base of the protocone especially at the entrance of the transverse valley. The principal cones are inclined towards the median longitudinal line of the molar. The labial cones are relatively higher vertically than the lingual ones. The protocone is semi-crescentic in shape and displays three equally strong folds, one proceeding forwards and outwards, the second backwards and a third backwards with an inwards tendency. The innermost lingual part of the protocone is pointed. The postprotocrista enters the posterior part of the anterior fossette leaving the postparacrista outside the fossette. The praeprotocrista is larger than the postprotocrista and it is continuous with the parastyle of the paracone. The hypocone is relatively smaller in antero-posterior length than the protocone (Fig. 30(1)). The praehypocrista is very small, not fused with the postprotocrista and the praemetacrista, and consequently it is free distally and mesially by deep furrows. The posthypocrista slopes downwards larger than the praehypocrista and fuses with the posterior cingulum. The paracone is comparatively higher than the protocone. It is almost spindle-shaped, with thickness in the middle. The anterior median rib, heavy pillar like robust body siutuated labially of the paracone is separated to parastyle by a vertical groove. The metacone is smarter than the paracone. The praemetacrista is connected with the postparacrista and the postmetacrista is linked with the posthypocrista through a small ridge. It is transversely thick in the middle unlike the paracone as its median rib is much weaker than that of the paracone. The styles are moderately developed. The parastyle is well developed and it is connected to the base of the anterior median rib. The mesostyle is strongly developed and it is in the form of an isolated pillar. It seems to be more associated with the metacone than the paracone. It is thin towards its
apex and very thick at the base. The metastyle is weakly developed. The central cavities are wide and very deep. Both central cavities are continuous with one another. Of these cavities, the anterior one is closed at its anterior end and the posterior one is closed at its posterior end. The posterior central cavity is also continuous with the transverse valley. The transverse valley is deep and narrow (Fig. 30(1)). Lower dentition The third lower molar is in a good state of preservation and a bunodont (Fig. 30(2)). The metaconid and the entoconid tips are slightly damaged. It is in an early wear. It is hypsodont and narrow crowned. The enamel is heavy, thick and very rugose. The cingulid is well developed anteriorly and it is absent labially, lingually and posteriorly. A small singular tubercle is also present between the hypoconid and the talonid on the labial side. The praeprotocristid terminates in a broad shelf, almost parallel to the anterior margin of the tooth. The postprotocristid is bifurcated and one limb of the bifurcation is attached to the postmetacristid while the other one is attached to the praehypocristid producing M structure (Fig. 30(2)). The hypoconid is crescentic in shape; the praehypocristed touches the postprotocristid, whereas the posthypocristid runs inwards and completely encircles the posterior base of the entoconid. The metaconid is conical and bunodont. The entoconid is conical with a short anterior process proceeding between the two anterior conids. There is a wide vertical groove between the metaconid and the entoconid lingually. The talonid is broad bearing a crenulated tubercle. The lingual and the labial arms of the talonid are almost equal in height. Comparison and discussion The described molars exhibit buno-selenodonty pattern. The selenodonty are found in families of Bovidae, Cervidae, Giraffidae and Camilidae and the semi-selenodonty with bunodont pattern is found in family Tragulidae. The studied specimens reflect semi-selenodonty with bunodont pattern and belong to family Tragulidae. There are two Siwalik genera Dorcatherium and Dorcabune
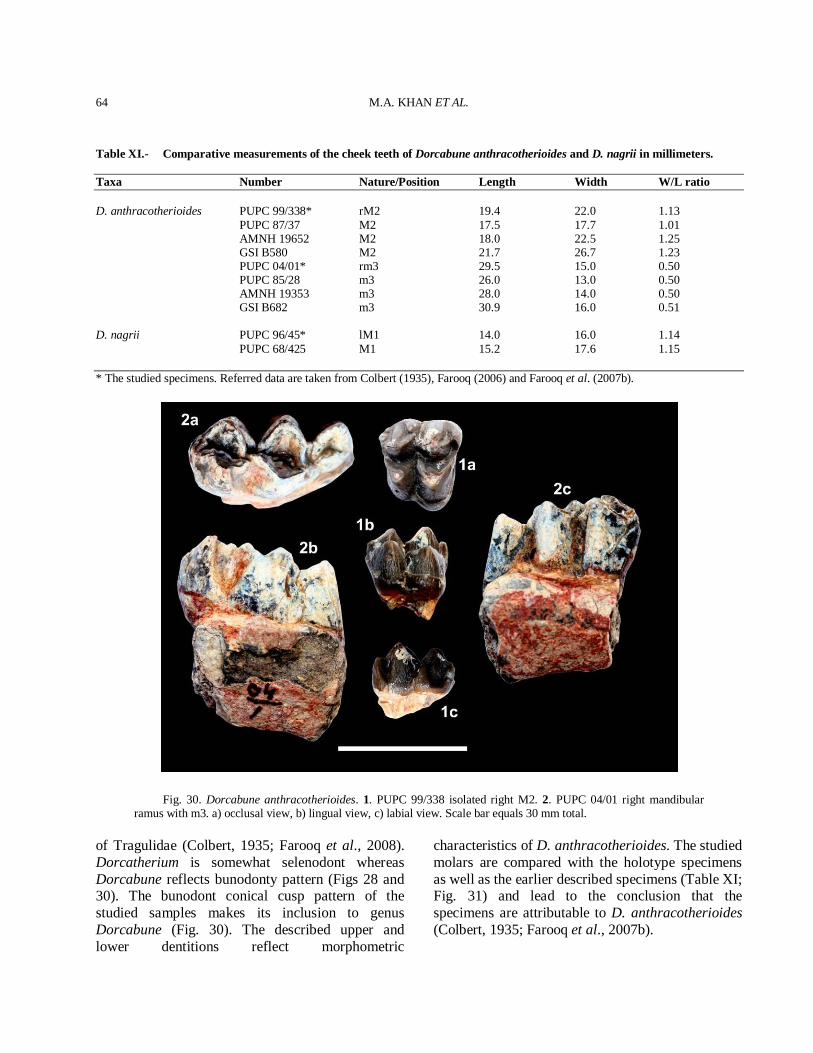
M.A. KHAN ET AL.
64
Table XI.- Comparative measurements of the cheek teeth of Dorcabune anthracotherioides and D. nagrii in millimeters. Taxa Number Nature/Position Length Width W/L ratio D. anthracotherioides PUPC 99/338* rM2 19.4 22.0 1.13 PUPC 87/37 M2 17.5 17.7 1.01 AMNH 19652 M2 18.0 22.5 1.25 GSI B580 M2 21.7 26.7 1.23 PUPC 04/01* rm3 29.5 15.0 0.50 PUPC 85/28 m3 26.0 13.0 0.50 AMNH 19353 m3 28.0 14.0 0.50 GSI B682 m3 30.9 16.0 0.51 D. nagrii PUPC 96/45* lM1 14.0 16.0 1.14 PUPC 68/425 M1 15.2 17.6 1.15 * The studied specimens. Referred data are taken from Colbert (1935), Farooq (2006) and Farooq et al. (2007b).
Fig. 30. Dorcabune anthracotherioides. 1. PUPC 99/338 isolated right M2. 2. PUPC 04/01 right mandibular ramus with m3. a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
of Tragulidae (Colbert, 1935; Farooq et al., 2008). Dorcatherium is somewhat selenodont whereas Dorcabune reflects bunodonty pattern (Figs 28 and 30). The bunodont conical cusp pattern of the studied samples makes its inclusion to genus Dorcabune (Fig. 30). The described upper and lower dentitions reflect morphometric
characteristics of D. anthracotherioides. The studied molars are compared with the holotype specimens as well as the earlier described specimens (Table XI; Fig. 31) and lead to the conclusion that the specimens are attributable to D. anthracotherioides (Colbert, 1935; Farooq et al., 2007b).
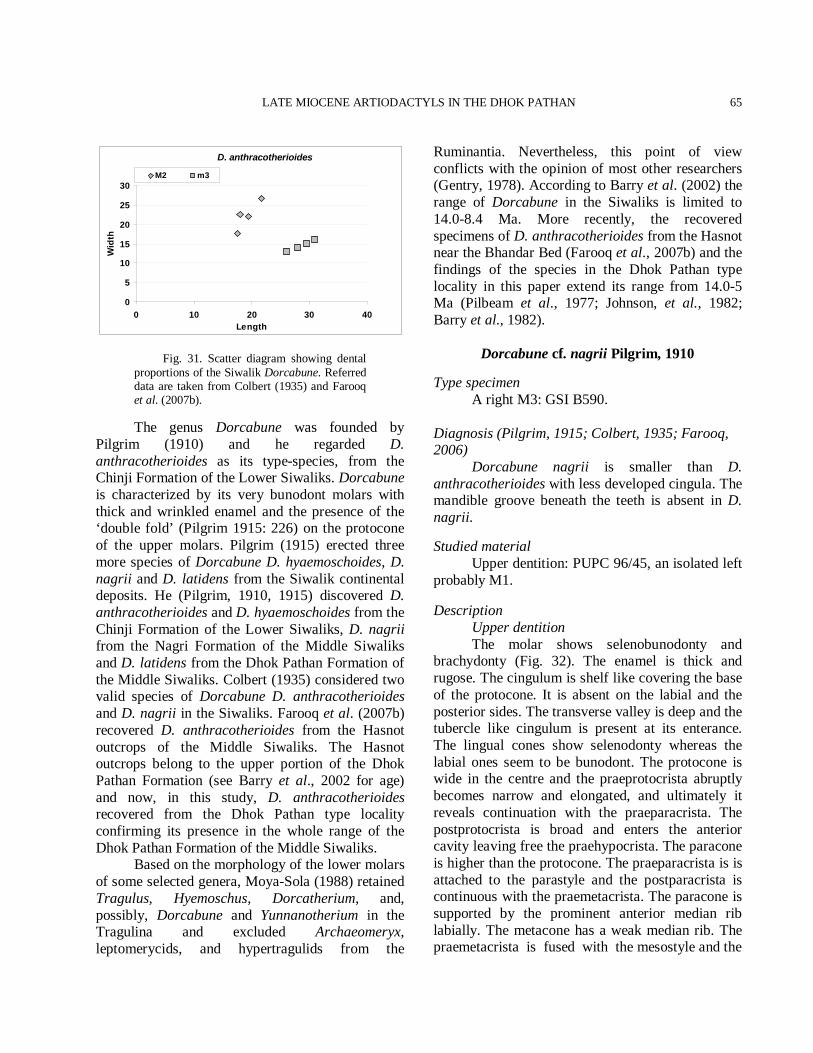
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
65
D. anthracotherioides
0
5
10
15
20
25
30
0 10 20 30 40Length
Wid
th
M2 m3
Fig. 31. Scatter diagram showing dental proportions of the Siwalik Dorcabune. Referred data are taken from Colbert (1935) and Farooq et al. (2007b).
The genus Dorcabune was founded by Pilgrim (1910) and he regarded D. anthracotherioides as its type-species, from the Chinji Formation of the Lower Siwaliks. Dorcabune is characterized by its very bunodont molars with thick and wrinkled enamel and the presence of the ‘double fold’ (Pilgrim 1915: 226) on the protocone of the upper molars. Pilgrim (1915) erected three more species of Dorcabune D. hyaemoschoides, D. nagrii and D. latidens from the Siwalik continental deposits. He (Pilgrim, 1910, 1915) discovered D. anthracotherioides and D. hyaemoschoides from the Chinji Formation of the Lower Siwaliks, D. nagrii from the Nagri Formation of the Middle Siwaliks and D. latidens from the Dhok Pathan Formation of the Middle Siwaliks. Colbert (1935) considered two valid species of Dorcabune D. anthracotherioides and D. nagrii in the Siwaliks. Farooq et al. (2007b) recovered D. anthracotherioides from the Hasnot outcrops of the Middle Siwaliks. The Hasnot outcrops belong to the upper portion of the Dhok Pathan Formation (see Barry et al., 2002 for age) and now, in this study, D. anthracotherioides recovered from the Dhok Pathan type locality confirming its presence in the whole range of the Dhok Pathan Formation of the Middle Siwaliks. Based on the morphology of the lower molars of some selected genera, Moya-Sola (1988) retained Tragulus, Hyemoschus, Dorcatherium, and, possibly, Dorcabune and Yunnanotherium in the Tragulina and excluded Archaeomeryx, leptomerycids, and hypertragulids from the
Ruminantia. Nevertheless, this point of view conflicts with the opinion of most other researchers (Gentry, 1978). According to Barry et al. (2002) the range of Dorcabune in the Siwaliks is limited to 14.0-8.4 Ma. More recently, the recovered specimens of D. anthracotherioides from the Hasnot near the Bhandar Bed (Farooq et al., 2007b) and the findings of the species in the Dhok Pathan type locality in this paper extend its range from 14.0-5 Ma (Pilbeam et al., 1977; Johnson, et al., 1982; Barry et al., 1982).
Dorcabune cf. nagrii Pilgrim, 1910
Type specimen A right M3: GSI B590. Diagnosis (Pilgrim, 1915; Colbert, 1935; Farooq, 2006) Dorcabune nagrii is smaller than D. anthracotherioides with less developed cingula. The mandible groove beneath the teeth is absent in D. nagrii. Studied material Upper dentition: PUPC 96/45, an isolated left probably M1. Description Upper dentition The molar shows selenobunodonty and brachydonty (Fig. 32). The enamel is thick and rugose. The cingulum is shelf like covering the base of the protocone. It is absent on the labial and the posterior sides. The transverse valley is deep and the tubercle like cingulum is present at its enterance. The lingual cones show selenodonty whereas the labial ones seem to be bunodont. The protocone is wide in the centre and the praeprotocrista abruptly becomes narrow and elongated, and ultimately it reveals continuation with the praeparacrista. The postprotocrista is broad and enters the anterior cavity leaving free the praehypocrista. The paracone is higher than the protocone. The praeparacrista is is attached to the parastyle and the postparacrista is continuous with the praemetacrista. The paracone is supported by the prominent anterior median rib labially. The metacone has a weak median rib. The praemetacrista is fused with the mesostyle and the
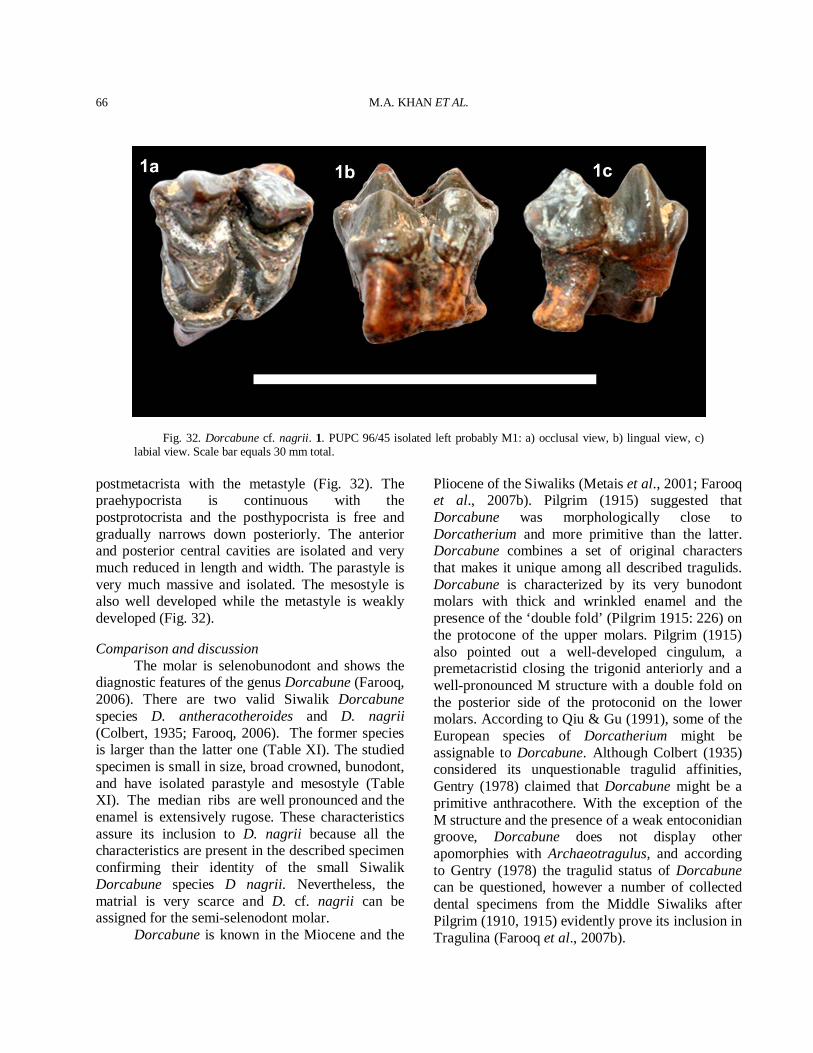
M.A. KHAN ET AL.
66
Fig. 32. Dorcabune cf. nagrii. 1. PUPC 96/45 isolated left probably M1: a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
postmetacrista with the metastyle (Fig. 32). The praehypocrista is continuous with the postprotocrista and the posthypocrista is free and gradually narrows down posteriorly. The anterior and posterior central cavities are isolated and very much reduced in length and width. The parastyle is very much massive and isolated. The mesostyle is also well developed while the metastyle is weakly developed (Fig. 32). Comparison and discussion The molar is selenobunodont and shows the diagnostic features of the genus Dorcabune (Farooq, 2006). There are two valid Siwalik Dorcabune species D. antheracotheroides and D. nagrii (Colbert, 1935; Farooq, 2006). The former species is larger than the latter one (Table XI). The studied specimen is small in size, broad crowned, bunodont, and have isolated parastyle and mesostyle (Table XI). The median ribs are well pronounced and the enamel is extensively rugose. These characteristics assure its inclusion to D. nagrii because all the characteristics are present in the described specimen confirming their identity of the small Siwalik Dorcabune species D nagrii. Nevertheless, the matrial is very scarce and D. cf. nagrii can be assigned for the semi-selenodont molar. Dorcabune is known in the Miocene and the
Pliocene of the Siwaliks (Metais et al., 2001; Farooq et al., 2007b). Pilgrim (1915) suggested that Dorcabune was morphologically close to Dorcatherium and more primitive than the latter. Dorcabune combines a set of original characters that makes it unique among all described tragulids. Dorcabune is characterized by its very bunodont molars with thick and wrinkled enamel and the presence of the ‘double fold’ (Pilgrim 1915: 226) on the protocone of the upper molars. Pilgrim (1915) also pointed out a well-developed cingulum, a premetacristid closing the trigonid anteriorly and a well-pronounced M structure with a double fold on the posterior side of the protoconid on the lower molars. According to Qiu & Gu (1991), some of the European species of Dorcatherium might be assignable to Dorcabune. Although Colbert (1935) considered its unquestionable tragulid affinities, Gentry (1978) claimed that Dorcabune might be a primitive anthracothere. With the exception of the M structure and the presence of a weak entoconidian groove, Dorcabune does not display other apomorphies with Archaeotragulus, and according to Gentry (1978) the tragulid status of Dorcabune can be questioned, however a number of collected dental specimens from the Middle Siwaliks after Pilgrim (1910, 1915) evidently prove its inclusion in Tragulina (Farooq et al., 2007b).

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
67
Family Giraffidae Gray, 1821 Subfamily Sivatheriinae Zittel, 1893
Genus HYDASPITHERIUM Lydekker, 1876
Type species Hydaspitherium megacephalum Lydekker, 1876. Abbreviated generic diagnosis (Matthew, 1929; Colbert, 1935; Bhatti, 2005) Teeth large, quadrate, with rugose enamel. Lower teeth broader and somewhat longer. Stratigraphic range Middle Siwaliks. Geographic distribution South Asia (Indian subcontinent). The occurrence of the genus is recorded from various localities of the Potwar Plateau (Middle Siwaliks) of Pakistan i.e., Dhok Pathan, Bhimber, Hasnot and it is also recovered from the Pliocene sediments of India (Lydekker, 1876; Pilgrim, 1910; Matthew, 1929; Colbert, 1935; Bhatti, 2005; Khan, 2007).
Hydaspitherium cf. megacephalum Lydekker, 1876
Type specimen A skull: GSI D150. Emended abbreviated diagnosis (modified from Colbert, 1935; Khan, 2007) The parastyle of Hydaspitherium megacephalum is prominent as compared to the mesostyle and the metastyle. The interior median rib is moderately developed, whereas the posterior median rib is weakly developed. The enamel is rugose. The stylids are present and the anterior fossette is transversely compressed. H. megacephalum generally is smaller than those of H. grande and H. magnum. Studied material Lower dentition: PUPC 09/78, left mandibular ramus with broken m1, complete m2 and partially erupted m3.
Description Lower dentition The material comprises a complete series of lower molars (Fig. 33; Table XII). PUPC 09/78 is a fragile and it is furnished with numerous cracks. PUPC 09/78 looks a subadult animal as the m1 in early wear, the m2 unworn and the m3 semierupted (Fig. 33). Furthermore, the mandible (PUPC 09/78) is shallow and its depth at the m2 base is 42 mm and the length of the mandibular ramus is 131 mm. The molars are in early wear, rugose and hypsodont. The conids are not interconnected with each other. The ectostylid is absent in the transverse valley. The cingulum is poorly developed. The fossettes are deep and narrow. The fossette between entoconid and hypoconid is closed at its rear end. The protoconid is V-shaped and the metaconid is spindle-shaped. The praehypocristid is longer than the posthypocristid and both the cristids are simple. The mesostylid is more developed than those of the metastylid and the entostylid. The anterior rib is more prominent than the posterior one. The hypoconulid is not fully erupted yet and it is lower in height than the other major conids (Fig. 33). Comparison The dental giraffid remains are difficult to assign at species level since size is in practice the only distinguishing criterion (Gentry, 1997). The described molars are typical giraffid molars showing basic giraffid features like the depth of the enamel folds, the obliquity of the barrels, the rugose sculpture of the enamel (Fig. 33). The teeth are large in size and can be excluded from the genus Giraffokeryx. In addition, flat entoconid wall and brachydonty present in Eurasian palaeotragines, the characters are absent in the studied sample. The studied molars are characterized by more narrowly pointed labial lobes, transverselly compressed anterior fossette, hypsodonty and out bowing of lingual ribs. These seemingly primitive characters give the teeth more resemblance to the upper Miocene Siwalik sevatheres. Among the large upper Miocene giraffids the genera Hydaspitheium, Giraffa, Sivatherium, Vishnutherium, and Bramatherium are possible candidates. Giraffa differs from the studied sample in brachydont teeth, lower molar set less obliquely to axis of jaw and not
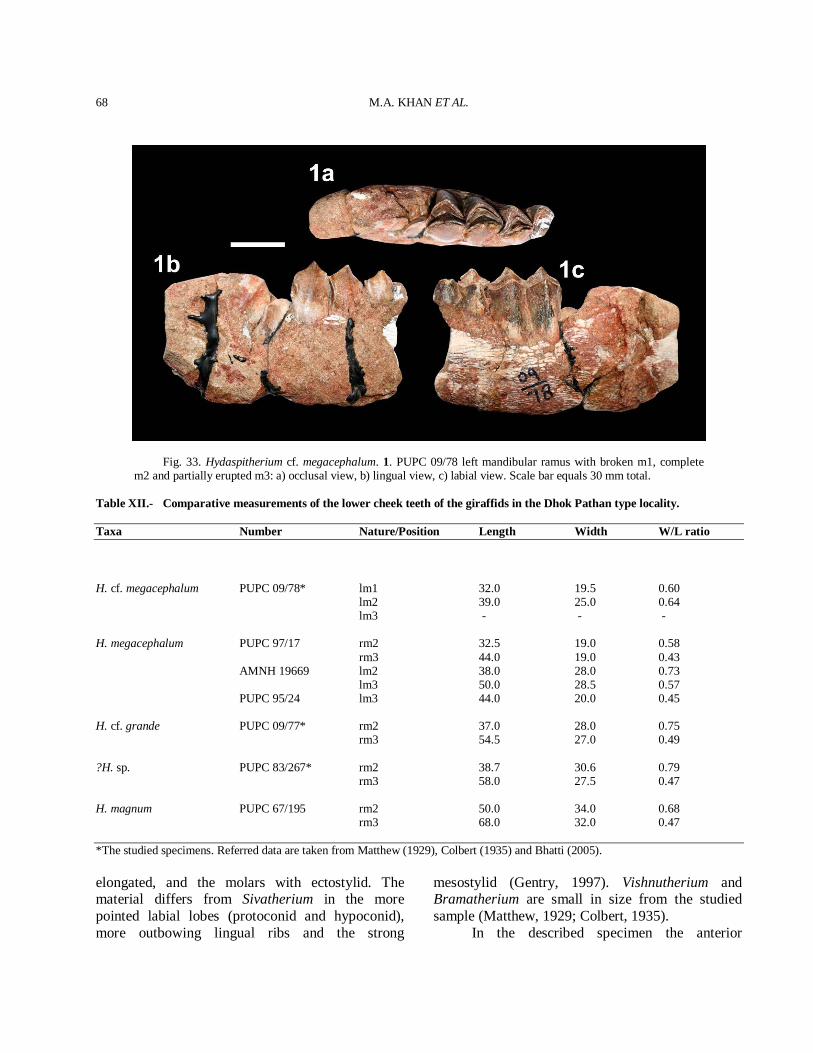
M.A. KHAN ET AL.
68
Fig. 33. Hydaspitherium cf. megacephalum. 1. PUPC 09/78 left mandibular ramus with broken m1, complete m2 and partially erupted m3: a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
Table XII.- Comparative measurements of the lower cheek teeth of the giraffids in the Dhok Pathan type locality. Taxa Number Nature/Position Length Width W/L ratio H. cf. megacephalum PUPC 09/78* lm1 32.0 19.5 0.60 lm2 39.0 25.0 0.64 lm3 - - - H. megacephalum PUPC 97/17 rm2 32.5 19.0 0.58 rm3 44.0 19.0 0.43 AMNH 19669 lm2 38.0 28.0 0.73 lm3 50.0 28.5 0.57 PUPC 95/24 lm3 44.0 20.0 0.45 H. cf. grande PUPC 09/77* rm2 37.0 28.0 0.75 rm3 54.5 27.0 0.49 ?H. sp. PUPC 83/267* rm2 38.7 30.6 0.79 rm3 58.0 27.5 0.47 H. magnum PUPC 67/195 rm2 50.0 34.0 0.68 rm3 68.0 32.0 0.47 *The studied specimens. Referred data are taken from Matthew (1929), Colbert (1935) and Bhatti (2005). elongated, and the molars with ectostylid. The material differs from Sivatherium in the more pointed labial lobes (protoconid and hypoconid), more outbowing lingual ribs and the strong
mesostylid (Gentry, 1997). Vishnutherium and Bramatherium are small in size from the studied sample (Matthew, 1929; Colbert, 1935). In the described specimen the anterior

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
69
fossette is compressed transversely, the stylids are present, the anterior median rib is prominent and the enamel sculpture is coarse (Fig. 33). These characteristics are found in the genus Hydaspitherium (Pilgrim, 1911; Matthew, 1929; Colbert, 1935; Bhatti, 2005). So the described specimens belong to the genus Hydaspitherium. The described lower molars are clearly smaller than those of H. grande and H. magnum (Fig. 36; Table XII). The studied material’s dimensions close to species H. megacephalum. The described specimen and the referred specimens of H. megacephalum appear same in size and crown structure (Pilgrim, 1911; Colbert, 1935; Bhatti, 2005). So the material belongs to the genus Hydaspitherium and more specifically to the species H. cf. megacephalum which is one of the smallest species of the genus.
Hydaspitherium cf. grande
Type specimen A left upper molar: GSI B155. Emended abbreviated diagnosis (modified from Colbert, 1935) A large species, considerably larger than H. megacephalum, but otherwise similar to it. The mandible is relatively smart and have mandibular groove beneath the teeth. The enamel is strongly rugose. The lower molars are broader and somewhat larger. Studied material Lower dentition: PUPC 09/77, right mandibular ramus with m2-3. Description and comparison The length of the mandibular ramus is 175 mm and its depth at m3 is 73 mm. A portion of the ascending ramus is also preserved which extends upwards to the rear end of the talonid (Fig. 34). The mandibular ramus is relatively slim and a groove is present beneath the teeth, which runs along the horizontal ramus. The groove is not observed in the samples assigned to H. megacephalum and H. magnum. The lower teeth resemble to H. megacephalum in the enamel sculpture,
transverselly compressed anterior fossette, structure of the crown, obliquely to axis of jaw, pointed labial lobes and outbowing of lingual ribs but metrically the sample is considerably larger than that of H. megacephalum (Fig. 36) and assign to H. cf. grande (Matthew, 1929; Colbert, 1935), as the material is insufficient for the more specifically identification (Fig. 34; Table XII).
?Hydaspitherium sp.
Studied material Lower dentition: PUPC 83/267, right mandibular ramus with m2-3.
Description and comparison This is one of the difficult specimens to identify. The length of the mandibular ramus is 158 mm, its depth at the m3 is 74 mm and the mandible maximum width is 40 mm. The sample is finely rugose and somewhat stout (Fig. 35). The mandible and teeth’s dimensions are about the same as in PUPC 09/77, here assigned to H. grande (Fig. 36; Table XII). The sample differs from H. grande in the rugosity of the enamel, the mandible stoutness and the width of the mandible which is 33 mm in PUPC 09/77 contrary to 40 mm of the studied specimem (Fig. 35). These characters give the sample more resemblance to Helladotherium and Camelus (Matthew, 1929; Colbert, 1935). The enamel is finely rugose in Helladotherium (Pilgrim, 1911) but Matthew (1929) stated that Helladotherium never entered in the Siwaliks. The Camelidae is not known in the Dhok Pathan Formation and in addition, Camelus lower molars are characterized by the fine rugose enamel, flat lingual walls and the retention of the anteroexternal fold. Unlike Camelus and Helladotherium, the lower molars are about the size of H. cf. grande, labial lobes are pointed, more outbowing lingual ribs and have weak stylids. The anterior fossette is compressed transversely and the molars are obliquely to axis of jaw. Therefore, the problematic sample (Fig. 35) could be doubtfully referred to Hydaspitherium sp. on the basis of its teeth characters and not to Helladotherium or Camelus.
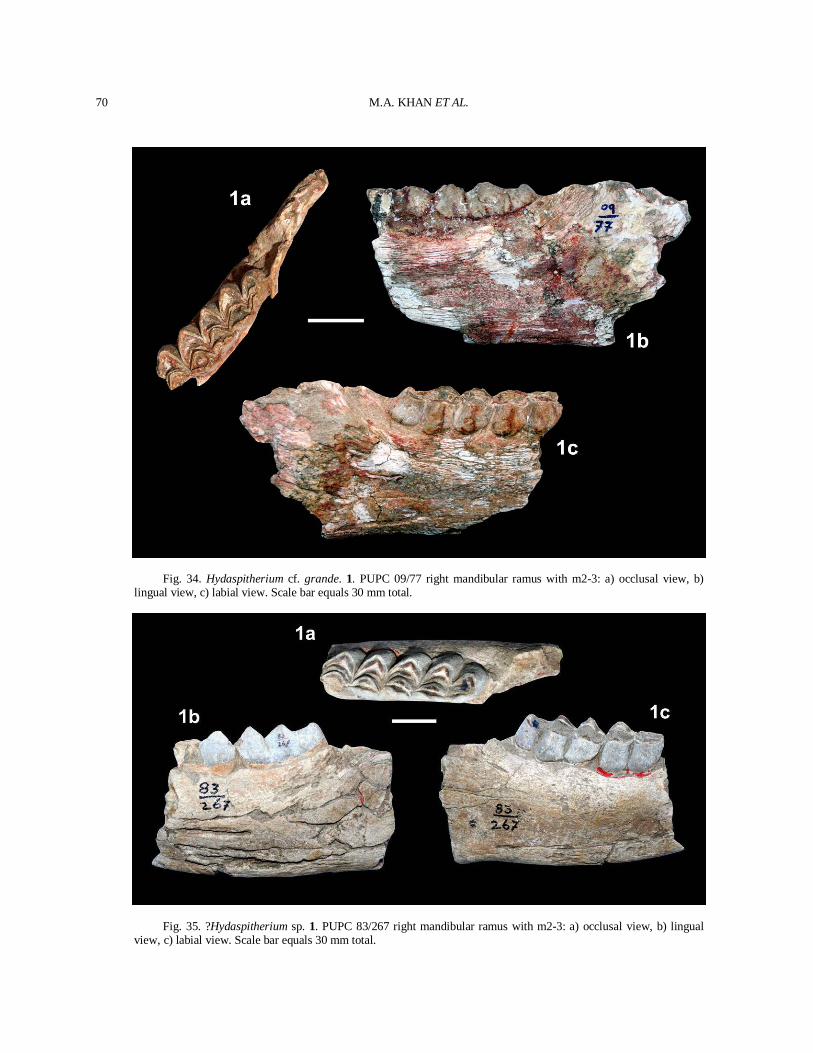
M.A. KHAN ET AL.
70
Fig. 34. Hydaspitherium cf. grande. 1. PUPC 09/77 right mandibular ramus with m2-3: a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
Fig. 35. ?Hydaspitherium sp. 1. PUPC 83/267 right mandibular ramus with m2-3: a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
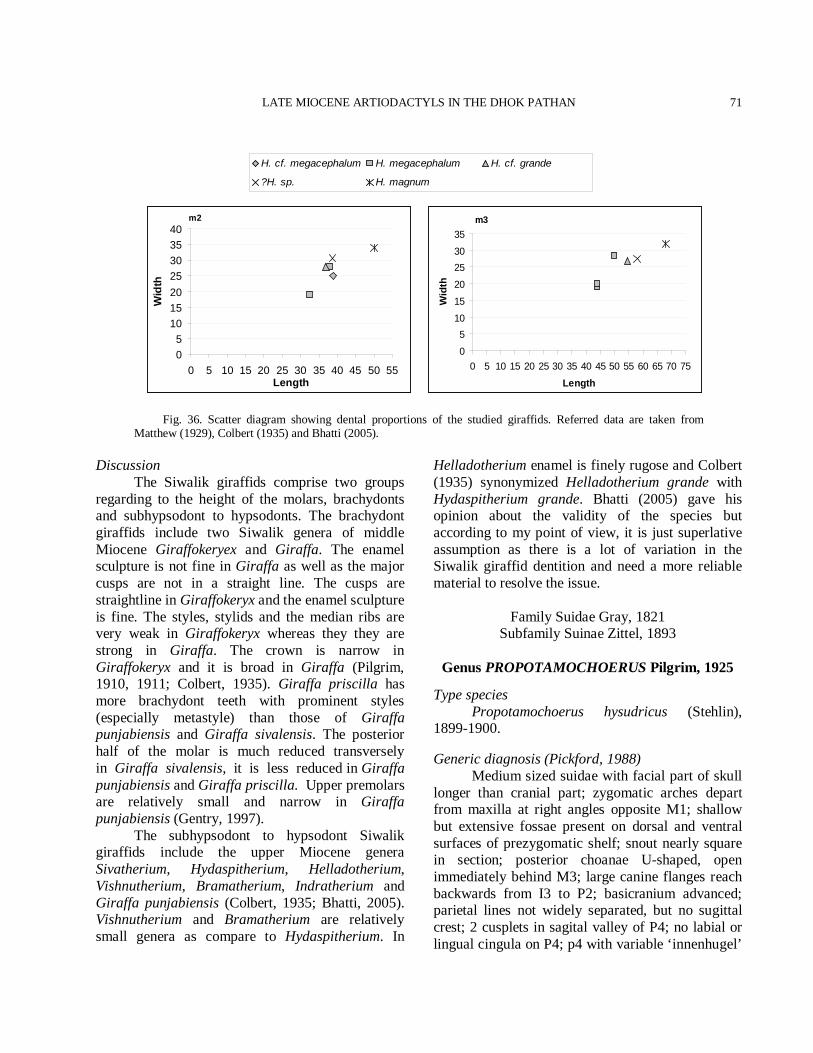
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
71
Fig. 36. Scatter diagram showing dental proportions of the studied giraffids. Referred data are taken from Matthew (1929), Colbert (1935) and Bhatti (2005).
Discussion The Siwalik giraffids comprise two groups regarding to the height of the molars, brachydonts and subhypsodont to hypsodonts. The brachydont giraffids include two Siwalik genera of middle Miocene Giraffokeryex and Giraffa. The enamel sculpture is not fine in Giraffa as well as the major cusps are not in a straight line. The cusps are straightline in Giraffokeryx and the enamel sculpture is fine. The styles, stylids and the median ribs are very weak in Giraffokeryx whereas they they are strong in Giraffa. The crown is narrow in Giraffokeryx and it is broad in Giraffa (Pilgrim, 1910, 1911; Colbert, 1935). Giraffa priscilla has more brachydont teeth with prominent styles (especially metastyle) than those of Giraffa punjabiensis and Giraffa sivalensis. The posterior half of the molar is much reduced transversely in Giraffa sivalensis, it is less reduced in Giraffa punjabiensis and Giraffa priscilla. Upper premolars are relatively small and narrow in Giraffa punjabiensis (Gentry, 1997). The subhypsodont to hypsodont Siwalik giraffids include the upper Miocene genera Sivatherium, Hydaspitherium, Helladotherium, Vishnutherium, Bramatherium, Indratherium and Giraffa punjabiensis (Colbert, 1935; Bhatti, 2005). Vishnutherium and Bramatherium are relatively small genera as compare to Hydaspitherium. In
Helladotherium enamel is finely rugose and Colbert (1935) synonymized Helladotherium grande with Hydaspitherium grande. Bhatti (2005) gave his opinion about the validity of the species but according to my point of view, it is just superlative assumption as there is a lot of variation in the Siwalik giraffid dentition and need a more reliable material to resolve the issue.
Family Suidae Gray, 1821 Subfamily Suinae Zittel, 1893
Genus PROPOTAMOCHOERUS Pilgrim, 1925
Type species Propotamochoerus hysudricus (Stehlin), 1899-1900. Generic diagnosis (Pickford, 1988) Medium sized suidae with facial part of skull longer than cranial part; zygomatic arches depart from maxilla at right angles opposite M1; shallow but extensive fossae present on dorsal and ventral surfaces of prezygomatic shelf; snout nearly square in section; posterior choanae U-shaped, open immediately behind M3; large canine flanges reach backwards from I3 to P2; basicranium advanced; parietal lines not widely separated, but no sugittal crest; 2 cusplets in sagital valley of P4; no labial or lingual cingula on P4; p4 with variable ‘innenhugel’
m3
0
510
15
20
2530
35
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75
Length
Wid
th
m2
05
10152025303540
0 5 10 15 20 25 30 35 40 45 50 55Length
Wid
thH. cf. megacephalum H. megacephalum H. cf. grande
?H. sp. H. magnum

M.A. KHAN ET AL.
72
and low to medium hypsodont anterior cusp and cingulum; dental formula 3143/3143; upper canine almost surrounded by cementum; molar enamel thickness moderate (2 ± 0.4 mm); Furchen shallow; no cingula on molars, except for occasional basal pillars in ends of median valleys; talon/talonid of M3/m3 relatively simple, no extra cusp pairs; mandible deep and robust; symphysis reaches back to rear of p2; canines not at corners of anterior ends of symphysis; anterior and posterior cusps of premolars variable, but never reach the same height as the main cusps. Stratigraphic range Middle Siwaliks. Geographic distribution Propotamochoerus occurs in the upper Miocene sediments of the Middle Siwaliks. The age range is from about 10 million to about 6 million years, a range that is comparable to the European Korynochoerus (Pickford, 1988). The localities of the Siwalik Hills that yield Propotamochoerus remains are Hasnot, Kotal Kund, Lehri, Kundvali, Gandakas, Utran, Dhok Mila, Jabi, Nagri, Khushalghar, Khaur, Kaulial, Mari, Ava, Bandar, Perim Island, Haritalyangar (Pilgrim, 1926b; Matthew, 1929; Colbert, 1935; Pickford, 1988).
Propotamochoerus hysudricus (Stehlin), 1899-
1900 Type specimen Right mandibular ramus with p3-m3 in worn condition: GSI B30. Diagnosis (Colbert, 1935; Pickford, 1988) No diastemata in upper or lower cheek tooth rows; orbits open over talon of M3; zygomatic arches leave face at midline of M1; lower premolars compressed with a long and complex third molar. Studied material Lower dentition: PUPC 07/32, left mandibular ramus with p4-m3; the p3 roots are also preserved in the alveoli. PUPC 99/02, left mandibular ramus with p4-m1. PUPC 94/65, right mandibular ramus with p4-m2. PUPC 97/15, right isolated m3.
Description and comparison The collected samples include the lower dentitions with mandibular rami (Fig. 37; Table XIII). The best preserved specimen is labeled as PUPC 07/32 (37(1)) and has a complete series of the lower molar, m1-3; the length of the molar series is 64 mm. It is broken posteriorly from where the ascending ramus hides the talonid of m3 in side view. The ascent begins well behind m3, so that this tooth is completely visible in lateral view. The length of the mandibular ramus (PUPC 07/32) is 112 mm and the depth of the mandibular ramus at m3 is 39 mm. The length of the mandibular ramus (PUPC 94/65) is 80 mm and its depth at m2 is 41 mm (Fig. 37(2)). The mandibular rami have robust bodies. The mandible of P. hysudricus reflects these above mentioned characteristics. The fourth premolar is a molarized tooth and the central cusp has migrated lingually to form the innenhugel and it appears subsymmetrical in lateral view. The metaconid is weak and occurs posteriorly to the protoconid. The posterior accessory cusplet is enlarged and is bordered by an inflated cingulum. The anterior accessory cusp and anterior cingulum are moderately high. The first molar is a four cusped tooth with anterior, median, and posterior accessory cusplets in the midline of the crown (Fig. 37; Table XIII). The posterior accessory cusplet is prominent and centrally placed. The molar is bunodont with the usual suid layout of four main cusps arranged in two lophs. The median basal pillar is present buccally in the median valley. The second molar is the large version of the m1. The m3 differs from the m2 by the presence of talonid and wide anterior lobes. The talonid of the m3 is moderately complex having a pair of cusplets side by side. There is moderate buccal flare in the molars, but lingual flare is minimal. The lingual cusps are slightly higher than the labial ones. The lower cheek teeth enamel is moderately thick and wrinkled. The giant suids of the Siwalik belong to the genera Tetraconodon, Sivachoerus and Hippopotamadon. The first two genera have characteristics inflated p3-4. The cheek teeth of the genus Hippopotamodon are extremely large. The studied material is too large to include Hippoptomadon. Propotamochoerus hysudricus is accompanied throughout the Middle Siwaliks by the

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
73
species Hippopotamodon sivalense. The specimens described here contain evidence about the species Propotamochoerus hysudricus and morpho-metrically close to the material of the species, described by Pickford in 1988 (Fig. 39; Table XIII). Propotamochoerus seems to have been derived from Hyotherium which occurs at the Chinji Formation, and it is possible that the changeover from one form to the other occurred at about 11 million years, just where the record is poor. The main advances made by Propotamochoerus over Hyotherium are the addition of sagittal cusplets in P4, increase in size, alteration in the shape of the anterior zygomata, and squaring off of the snout section (Pickford, 1988).
Genus HIPPOPOTAMODON Lydekker, 1877
Type species Hippopotamodon sivalense Lydekker, 1877. Generic diagnosis (Pickford, 1988) Giant Suinae in which the males have large flaring canines; molar enamel relatively thin; molars relatively simple with well developed furchenplan; labial cusps in lower molars lower crowned than lingual ones; P4 with posterior accessory cusp almost as large as two main labial cusps; posterior choanae U-shaped, open immediately behind M3; p4 with prominent innenhugel and 2-3-4 cusp, anterior cingulum and accessory-1 cusp moderately high; diastema between C-P1-2 short; broad flat dorsal surface to brain case. Stratigraphic range Middle Siwaliks. Geographic distribution Hippopotamodon is common in the upper Miocene of the Siwaliks. It is almost always associated with Hipparion and its range is very similar to that of Propotamochoerus hysudricus and Microstonyx (Pickford, 1988).
?Hippopotamodon sp.
Studied material Lower dentition: PUPC 00/14, left
mandibular ramus with broken p4 and partially complete m1. PUPC 08/26, badly crushed right mandibular ramus with damaged m1-3, probably juvenile animal.
Description and comparison The recovered samples are badly damaged unfortunately. However, the general contour of the molars allows to study the morphological features of the specimens. The features are based on the sample, labeled as PUPC 00/14 (Fig. 38; Table XIII). The premolar crown is damaged but its basal part is preserved. It is somewhat large tooth (Table XIII). The molars are simple with thin enamel. The median valley is wide, the furchen moderately deep and narrow. The basal pillar is present in the median valley. However, the mandibular rami are damaged but they preserve enough to throw light on their diagnostic features. The morphological features of the samples resemble with Hippopotamodon, described from the Dhok Pathan Formation of the Siwaliks by Pickford (1988). The metrical values of the sample are smaller than those of the already described material, as the material belongs to juvenile animal and furthermore, there is a trend towards increase in size with time in Hippopotamodon, specimens from the Nagri being smaller than those from the Dhok Pathan, and these smaller than those from the Hasnot area (Pickford, 1988). The material may be assigned to ?Hippopotamodon sp. based on the morphometric features (Figs. 38-39; Table XIII).
Discussion In the Siwaliks lower Miocene (20 Ma to 15 Ma) suids are very poorly known, being represented by two specimens, one each of Libycochoerus affinis and Libycochoerus fategadensis. Hyotherium, Listriodon and Conohyus are represented in the Siwalik middle Miocene (15 Ma to 10.5 Ma). Suid diversification found considerably in the Siwalik upper Miocene (10.5 Ma to 5.5 Ma). During the early part of the period (from base of the Nagri to top of the type Dhok Pathan) the species Propotamochoerus hysudricus and Hippopotamodon sivalense are the most abundant, with Tetraconodon magnus, Conohyus indicus and
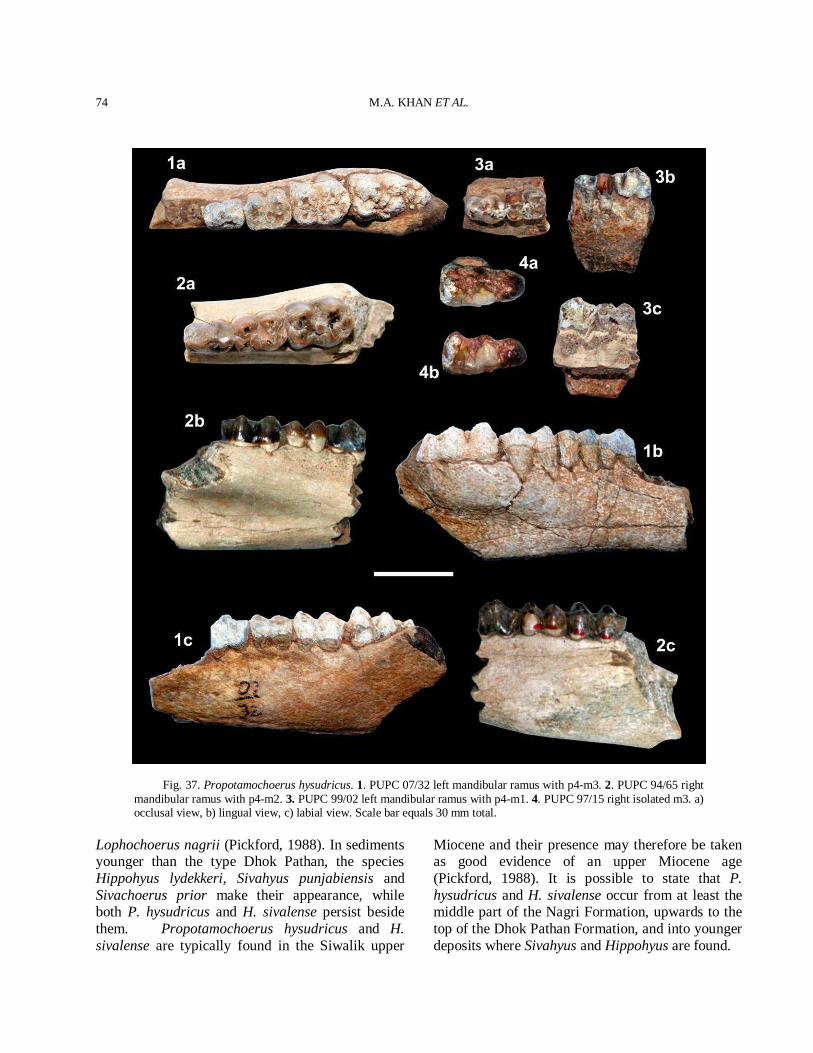
M.A. KHAN ET AL.
74
Fig. 37. Propotamochoerus hysudricus. 1. PUPC 07/32 left mandibular ramus with p4-m3. 2. PUPC 94/65 right mandibular ramus with p4-m2. 3. PUPC 99/02 left mandibular ramus with p4-m1. 4. PUPC 97/15 right isolated m3. a) occlusal view, b) lingual view, c) labial view. Scale bar equals 30 mm total.
Lophochoerus nagrii (Pickford, 1988). In sediments younger than the type Dhok Pathan, the species Hippohyus lydekkeri, Sivahyus punjabiensis and Sivachoerus prior make their appearance, while both P. hysudricus and H. sivalense persist beside them. Propotamochoerus hysudricus and H. sivalense are typically found in the Siwalik upper
Miocene and their presence may therefore be taken as good evidence of an upper Miocene age (Pickford, 1988). It is possible to state that P. hysudricus and H. sivalense occur from at least the middle part of the Nagri Formation, upwards to the top of the Dhok Pathan Formation, and into younger deposits where Sivahyus and Hippohyus are found.
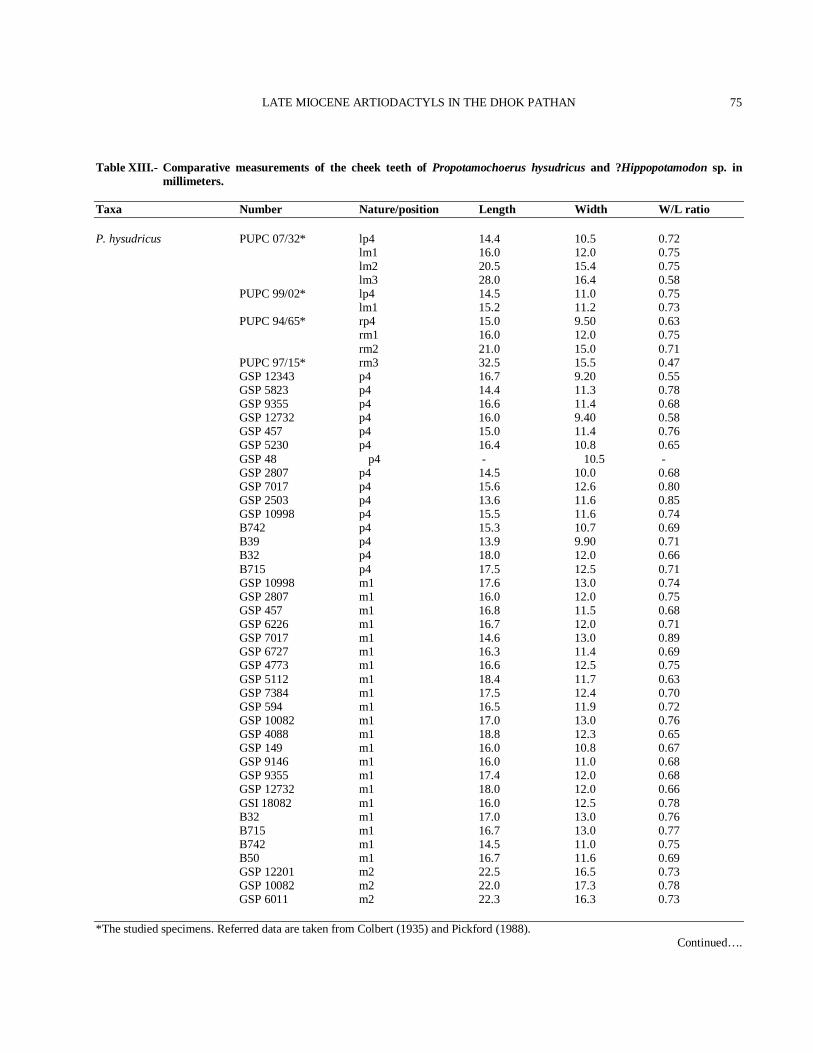
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
75
Table XIII.- Comparative measurements of the cheek teeth of Propotamochoerus hysudricus and ?Hippopotamodon sp. in
millimeters. Taxa Number Nature/position Length Width W/L ratio P. hysudricus PUPC 07/32* lp4 14.4 10.5 0.72 lm1 16.0 12.0 0.75 lm2 20.5 15.4 0.75 lm3 28.0 16.4 0.58 PUPC 99/02* lp4 14.5 11.0 0.75 lm1 15.2 11.2 0.73 PUPC 94/65* rp4 15.0 9.50 0.63 rm1 16.0 12.0 0.75 rm2 21.0 15.0 0.71 PUPC 97/15* rm3 32.5 15.5 0.47 GSP 12343 p4 16.7 9.20 0.55 GSP 5823 p4 14.4 11.3 0.78 GSP 9355 p4 16.6 11.4 0.68 GSP 12732 p4 16.0 9.40 0.58 GSP 457 p4 15.0 11.4 0.76 GSP 5230 p4 16.4 10.8 0.65 GSP 48 p4 - 10.5 - GSP 2807 p4 14.5 10.0 0.68 GSP 7017 p4 15.6 12.6 0.80 GSP 2503 p4 13.6 11.6 0.85 GSP 10998 p4 15.5 11.6 0.74 B742 p4 15.3 10.7 0.69 B39 p4 13.9 9.90 0.71 B32 p4 18.0 12.0 0.66 B715 p4 17.5 12.5 0.71 GSP 10998 m1 17.6 13.0 0.74 GSP 2807 m1 16.0 12.0 0.75 GSP 457 m1 16.8 11.5 0.68 GSP 6226 m1 16.7 12.0 0.71 GSP 7017 m1 14.6 13.0 0.89 GSP 6727 m1 16.3 11.4 0.69 GSP 4773 m1 16.6 12.5 0.75 GSP 5112 m1 18.4 11.7 0.63 GSP 7384 m1 17.5 12.4 0.70 GSP 594 m1 16.5 11.9 0.72 GSP 10082 m1 17.0 13.0 0.76 GSP 4088 m1 18.8 12.3 0.65 GSP 149 m1 16.0 10.8 0.67 GSP 9146 m1 16.0 11.0 0.68 GSP 9355 m1 17.4 12.0 0.68 GSP 12732 m1 18.0 12.0 0.66 GSI 18082 m1 16.0 12.5 0.78 B32 m1 17.0 13.0 0.76 B715 m1 16.7 13.0 0.77 B742 m1 14.5 11.0 0.75 B50 m1 16.7 11.6 0.69 GSP 12201 m2 22.5 16.5 0.73 GSP 10082 m2 22.0 17.3 0.78 GSP 6011 m2 22.3 16.3 0.73 *The studied specimens. Referred data are taken from Colbert (1935) and Pickford (1988).
Continued….
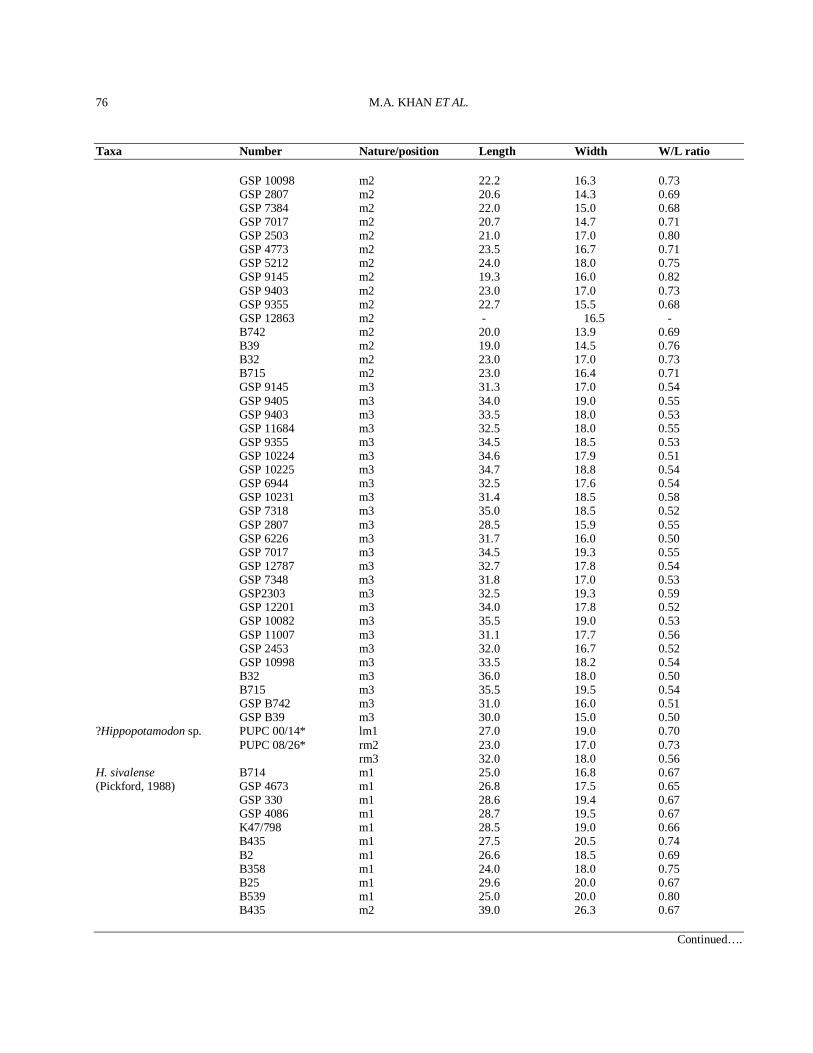
M.A. KHAN ET AL.
76
Taxa Number Nature/position Length Width W/L ratio GSP 10098 m2 22.2 16.3 0.73 GSP 2807 m2 20.6 14.3 0.69 GSP 7384 m2 22.0 15.0 0.68 GSP 7017 m2 20.7 14.7 0.71 GSP 2503 m2 21.0 17.0 0.80 GSP 4773 m2 23.5 16.7 0.71 GSP 5212 m2 24.0 18.0 0.75 GSP 9145 m2 19.3 16.0 0.82 GSP 9403 m2 23.0 17.0 0.73 GSP 9355 m2 22.7 15.5 0.68 GSP 12863 m2 - 16.5 - B742 m2 20.0 13.9 0.69 B39 m2 19.0 14.5 0.76 B32 m2 23.0 17.0 0.73 B715 m2 23.0 16.4 0.71 GSP 9145 m3 31.3 17.0 0.54 GSP 9405 m3 34.0 19.0 0.55 GSP 9403 m3 33.5 18.0 0.53 GSP 11684 m3 32.5 18.0 0.55 GSP 9355 m3 34.5 18.5 0.53 GSP 10224 m3 34.6 17.9 0.51 GSP 10225 m3 34.7 18.8 0.54 GSP 6944 m3 32.5 17.6 0.54 GSP 10231 m3 31.4 18.5 0.58 GSP 7318 m3 35.0 18.5 0.52 GSP 2807 m3 28.5 15.9 0.55 GSP 6226 m3 31.7 16.0 0.50 GSP 7017 m3 34.5 19.3 0.55 GSP 12787 m3 32.7 17.8 0.54 GSP 7348 m3 31.8 17.0 0.53 GSP2303 m3 32.5 19.3 0.59 GSP 12201 m3 34.0 17.8 0.52 GSP 10082 m3 35.5 19.0 0.53 GSP 11007 m3 31.1 17.7 0.56 GSP 2453 m3 32.0 16.7 0.52 GSP 10998 m3 33.5 18.2 0.54 B32 m3 36.0 18.0 0.50 B715 m3 35.5 19.5 0.54 GSP B742 m3 31.0 16.0 0.51 GSP B39 m3 30.0 15.0 0.50 ?Hippopotamodon sp. PUPC 00/14* lm1 27.0 19.0 0.70 PUPC 08/26* rm2 23.0 17.0 0.73 rm3 32.0 18.0 0.56 H. sivalense B714 m1 25.0 16.8 0.67 (Pickford, 1988) GSP 4673 m1 26.8 17.5 0.65 GSP 330 m1 28.6 19.4 0.67 GSP 4086 m1 28.7 19.5 0.67 K47/798 m1 28.5 19.0 0.66 B435 m1 27.5 20.5 0.74 B2 m1 26.6 18.5 0.69 B358 m1 24.0 18.0 0.75 B25 m1 29.6 20.0 0.67 B539 m1 25.0 20.0 0.80 B435 m2 39.0 26.3 0.67
Continued….
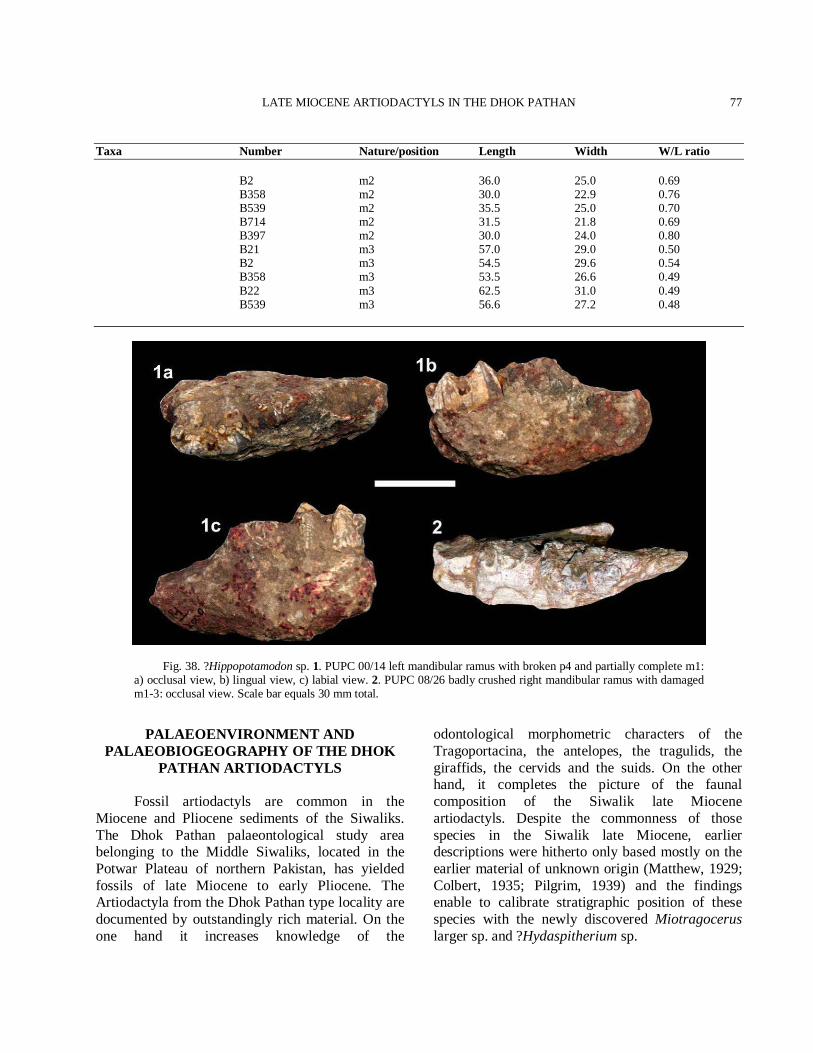
LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
77
Taxa Number Nature/position Length Width W/L ratio B2 m2 36.0 25.0 0.69 B358 m2 30.0 22.9 0.76 B539 m2 35.5 25.0 0.70 B714 m2 31.5 21.8 0.69 B397 m2 30.0 24.0 0.80 B21 m3 57.0 29.0 0.50 B2 m3 54.5 29.6 0.54 B358 m3 53.5 26.6 0.49 B22 m3 62.5 31.0 0.49 B539 m3 56.6 27.2 0.48
Fig. 38. ?Hippopotamodon sp. 1. PUPC 00/14 left mandibular ramus with broken p4 and partially complete m1: a) occlusal view, b) lingual view, c) labial view. 2. PUPC 08/26 badly crushed right mandibular ramus with damaged m1-3: occlusal view. Scale bar equals 30 mm total.
PALAEOENVIRONMENT AND PALAEOBIOGEOGRAPHY OF THE DHOK
PATHAN ARTIODACTYLS Fossil artiodactyls are common in the Miocene and Pliocene sediments of the Siwaliks. The Dhok Pathan palaeontological study area belonging to the Middle Siwaliks, located in the Potwar Plateau of northern Pakistan, has yielded fossils of late Miocene to early Pliocene. The Artiodactyla from the Dhok Pathan type locality are documented by outstandingly rich material. On the one hand it increases knowledge of the
odontological morphometric characters of the Tragoportacina, the antelopes, the tragulids, the giraffids, the cervids and the suids. On the other hand, it completes the picture of the faunal composition of the Siwalik late Miocene artiodactyls. Despite the commonness of those species in the Siwalik late Miocene, earlier descriptions were hitherto only based mostly on the earlier material of unknown origin (Matthew, 1929; Colbert, 1935; Pilgrim, 1939) and the findings enable to calibrate stratigraphic position of these species with the newly discovered Miotragocerus larger sp. and ?Hydaspitherium sp.
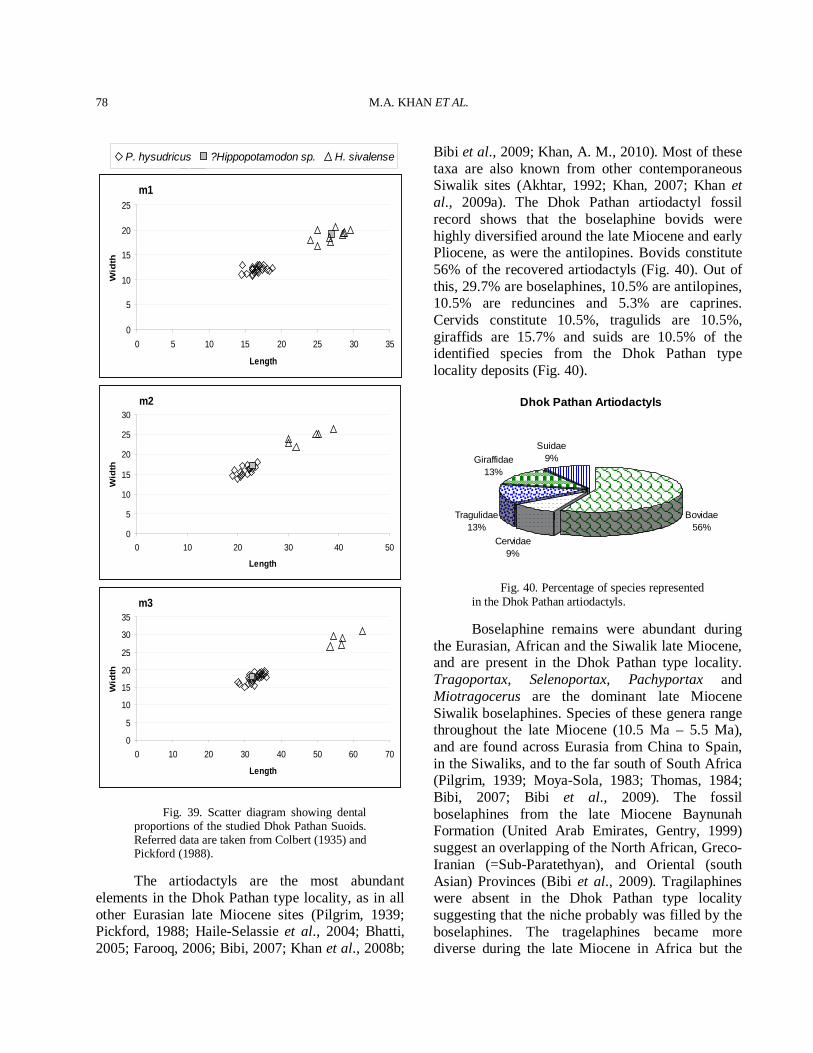
M.A. KHAN ET AL.
78
m1
0
5
10
15
20
25
0 5 10 15 20 25 30 35
Length
Wid
th
m2
0
5
10
15
20
25
30
0 10 20 30 40 50
Length
Wid
th
m3
0
5
10
15
20
25
30
35
0 10 20 30 40 50 60 70
Length
Wid
th
Fig. 39. Scatter diagram showing dental proportions of the studied Dhok Pathan Suoids. Referred data are taken from Colbert (1935) and Pickford (1988).
The artiodactyls are the most abundant elements in the Dhok Pathan type locality, as in all other Eurasian late Miocene sites (Pilgrim, 1939; Pickford, 1988; Haile-Selassie et al., 2004; Bhatti, 2005; Farooq, 2006; Bibi, 2007; Khan et al., 2008b;
Bibi et al., 2009; Khan, A. M., 2010). Most of these taxa are also known from other contemporaneous Siwalik sites (Akhtar, 1992; Khan, 2007; Khan et al., 2009a). The Dhok Pathan artiodactyl fossil record shows that the boselaphine bovids were highly diversified around the late Miocene and early Pliocene, as were the antilopines. Bovids constitute 56% of the recovered artiodactyls (Fig. 40). Out of this, 29.7% are boselaphines, 10.5% are antilopines, 10.5% are reduncines and 5.3% are caprines. Cervids constitute 10.5%, tragulids are 10.5%, giraffids are 15.7% and suids are 10.5% of the identified species from the Dhok Pathan type locality deposits (Fig. 40).
Dhok Pathan Artiodactyls
Bovidae56%
Cervidae9%
Tragulidae13%
Giraffidae13%
Suidae9%
Fig. 40. Percentage of species represented in the Dhok Pathan artiodactyls.
Boselaphine remains were abundant during the Eurasian, African and the Siwalik late Miocene, and are present in the Dhok Pathan type locality. Tragoportax, Selenoportax, Pachyportax and Miotragocerus are the dominant late Miocene Siwalik boselaphines. Species of these genera range throughout the late Miocene (10.5 Ma – 5.5 Ma), and are found across Eurasia from China to Spain, in the Siwaliks, and to the far south of South Africa (Pilgrim, 1939; Moya-Sola, 1983; Thomas, 1984; Bibi, 2007; Bibi et al., 2009). The fossil boselaphines from the late Miocene Baynunah Formation (United Arab Emirates, Gentry, 1999) suggest an overlapping of the North African, Greco-Iranian (=Sub-Paratethyan), and Oriental (south Asian) Provinces (Bibi et al., 2009). Tragilaphines were absent in the Dhok Pathan type locality suggesting that the niche probably was filled by the boselaphines. The tragelaphines became more diverse during the late Miocene in Africa but the
P. hysudricus ?Hippopotamodon sp. H. sivalense

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
79
boselaphines became extinct at the end of the Miocene in Africa (Haile-Selassie et al., 2004). Two antelope species are identified from the Dhok Pathan type locality: Prosetrpsiceros cf. vinayaki and Gazella lydekkeri. One caprine, Caprotragoides cf. potwaricus is documented from the type locality. Reduncines are represented in the Dhok Pathan type locality by two species. Four specimens are assigned to Kobus porrecticornis and one specimen is assigned to cf. Kobikeryx sp. Kobus porrecticornis considers to be an early reduncine and represents from the Dhok Pathan Formation of the Siwaliks (Pilgrim, 1939; Gentry, 1980). Kobus porrecticornis dominated as a prominent element of the Dhok Pathan type locality and it was represented by a reasonable number of specimens. Kobus aff. porrecticornis, from the Kenyan Mpesida Beds (ca. 6.5 Ma) and the Lukeino Formation (ca. 6.0 Ma), is considered to belong to the Siwalik Kobus porrecticornis (Thomas, 1979; Gentry, 1980; Vrba, 1995). The earliest reduncine records in Eurasia and in the Siwaliks date to the late Miocene (Vrba, 1995; Gentry, 1997). Late Miocene links between sub-Sahran Africa and southern Asia are also apparent in the shared presence of bovid taxa such as the reduncine Kobus porrecticornis (Vrba and Haile-Selassie, 2006; in this paper). Alcelaphines are absent in the Dhok Pathan type locality. The absence of alcelaphines may be an indication of the absence of a vast expanse of large open grasslands. The abundance of reduncines, antilopines and boselaphines indicates a mixture of woodland and grassland biomes. The absence of alcelaphines from the Dhok Pathan type locality clearly reflects ecological differences. Extant alcelaphines are known to live in arid plains, deserts, savanna, and grassland. These habitats might have not been available for the alcelaphines to flourish in the Dhok Pathan deposits. A closed habitat has been inferred for the late Miocene boselaphines (Khan et al., 2009a). Some taxa of the Dhok Pathan deposits inhabit variously drier bush and woodlands into moisture valleys and forests. What can be inferred from this observation suggesting a closed habitat in the late Miocene to a more open habitat in the Pliocene. Cervids are represented by two species from the type locality: Cervus simplicidens and Cervus
triplidens. The primitive cervine was probably tropical in distribution, it certainly inhabited woodland or open country, not closed forest, and it probably lived in eastern Eurasia or probably India (not in western Eurasia or Southern Asia) (Kahlke, 1976; Lister, 1984, 1986; Sommer et al., 2008). Tragulids are identified by two genera Dorcatherium and Dorcabune from the Dhok Pathan type locality. Dorcatherium is rather indicative of swampy-paludal habitats (Köhler, 1993; Rössner and Mörs, 2001). Dorcatherium with cervids in the artiodactyl community from the Dhok Pathan type locality reflects an environment of mainly closed forest. A significant representation of tragulids with other artiodactyls indicates humid conditions, accompanying open areas (Rossner, 2010). Dorcabune anthracotherioides were collected from the Dhok Pathan type locality and according to Barry et al. (2002) the range of Dorcabune in the Siwaliks is limited to 14.0-8.4 Ma. More recently, Dorcabune anthracotherioides recorded from the Hasnot (Farooq et al., 2007b) near the Bandar Bed which is located in the east-north of Hasnot and considered the youngest part of the Dhok Pathan Formation (Pilbeam et al., 1977; Johnson, N., et al., 1982; Barry et al., 1982). Giraffids represent three species of Hydaspitherium in the Dhok Pathan type locality fossil record. These species are also known from other contemporaneous sites in the Siwaliks and elsewhere. Hydaspitherium is the most common giraffid at the Dhok Pathan and the equivalent sites dated to between the late Miocene and early Pliocene. However, Giraffa, Bramatherium and Vishnutherium are documented from the late Miocene of the Siwaliks and Sivatherium is documented from younger deposits (Matthew, 1929; Colbert, 1935; Khan, 2007). Suid remains found in the Dhok Pathan type locality are assigned to two genera, Propotamochoerus and Hippopotamodon. The deposits exhibit the presence of the two contemporaneous and sympatric genera in the late Miocene and early Pliocene. An analogous situation occurs in Europe, where Korynochoerus and Microstonyx are common (Pickford, 1988). The known range of the genera is from about 10 Ma to about 6 Ma, but older and younger specimens might

M.A. KHAN ET AL.
80
be expected to be found. These are upper Miocene species, apparently dying out by the Pliocene. The artiodactyl assemblage from the Dhok Pathan type locality reflects a qualitative taxonomic structure well known from other communities of Eurasia and Africa in the late Miocene and early Pliocene (Pilgrim, 1939; Pickford, 1988; Gentry, 1997; Bibi, 2007; Vrba and Haile-Selassie, 2006). The artiodactyl composition from the Dhok Pathan type locality is peculiar in having a comparatively high species number. The combination of a large number of specimens at the Dhok Pathan locality means that the structure of the palaeocommunity is most probably not a product of taphonomical effects, but can be taken as real. Besides the here described twenty three species of artiodactyls, the perissodactyl fauna from the Dhok Pathan type locality comprises rhinos, chalicotheres and hipparions (Heissig 1972; Iqbal et al., 2009; Khan et al., 2009b; Khan, 2010). This number is unusual in modern world biomes, where the highest number of brachydonts and hypsodonts is represented. But, it corresponds to the composition fauna of the late Miocene of Eurasia and Africa (Gentry, 1980; Haile-Selassie et al., 2004; Kostopoulos, 2006; Bibi et al., 2009), which is associated with dry and more seasonal climate in the late Miocene. This is well in accordance with recently documented open woodlands or grassy woodlands with C4 grasses. Plant communities with predominantly C3 plants were greatly diminished after 7.0 Ma, and those with predominantly C4 plants, appeared as early as 7.4 Ma. A few paleosols may have formed under water logged, grassy woodlands, but most formed under drier conditions and closed vegetation (Barry et al., 2002). The selenodonty of the artiodactyl groups may be interpreted for fibrous foods which in most likely may have been swamp vegetation due to the depositional environment. The combined occurrence of such taxa in the locality suggests open but no grassy habitats, including some elements of fairly abrasive vegetation (Janis et al., 2002).
CONCLUSIONS
The Dhok Pathan mammalian fauna suggests a much higher diversity during the late Miocene of
the Siwaliks (Lydekker, 1878; Matthew, 1929; Colbert, 1935; Pilgrim, 1926b, 1937, 1939; Thomas, 1984; Akhtar, 1992; Bhatti, 2005; Ghaffar, 2005; Iqbal et al., 2009; Khan, 2010). On the basis of the faunal studies, it is concluded that during the late Miocene and early Pliocene the Dhok Pathan region was considerably more humid and better vegetated than it is today (Barry et al., 2002). The artiodactyl assemblages suggest a mix of heavy cover and, to somewhat lesser extent, forested environment. The presence in the area of some more open habitats can not be ruled out. The artiodactyls from the Dhok Pathan deposits suggest a mosaic of habitats ranging from grassy woodland and woodland, to riverine and forest settings. This is what one would expect in light of the diversity of the animal community and availability of diverse habitat to accommodate it. A similar mosaic palaeoenvironment is inferred for the Hasnot deposits (Khan, 2007). The autecological characterizations of all twenty three species support the reconstruction of an environment mainly shaped by water logged, grassy woodlands, formed under drier conditions and closed vegetation and are thus consistent with current environmental interpretations for the Eurasia and Africa Late Miocene. The artiodactyl community structure of the Dhok Pathan suggest the existence of a vast openland environment with alternating dry and floodseasons which forced a mosaic of ecotonal habitats with an endless number of niches and corresponding adaptations. In summary, the assemblages of the Dhok Pathan type locality represent the late Miocene to Pliocene. The Dhok Pathan type locality fossil assemblages are found in Africa and Eurasia, and they are thus of particular interest to an understanding of mammalian evolutionary processes.
ACKNOWLEDGEMENTS The publication was made possible by funding from the University of the Punjab, Lahore, Pakistan. We are grateful to Altaf Shah for his field work assistance and the hospitality during our stay in Dhok Pathan village. Sajjad Shah provided efficient help during the field work and Adeeb Babar prepared the figures.

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
81
REFERENCES AKHTAR, M., 1992. Taxonomy and Distribution of the Siwalik
Bovids. Ph.D. dissert. (unpublished), University of the Punjab, Lahore, Pakistan.
AKHTAR, M., 1995. Pachyportax giganteus, new species (Mammalia, Artiodactyla, Bovidae) from the Dhok Pathan, district Chakwal, Punjab, Pakistan. Pakistan J. Zool., 27: 337-340.
ALCALÁ, L. AND MORALES, J., 2006. Antilopinae (Bovidae, Mammalia) from the Lower Pliocene of Teruel Basin (Spain). Estud. Geológ., 62: 559-570.
ARAMBOURG, C. AND PIVETEAU, J., 1929. Dorcatherium puyhauberti sp. n. Pontian nr. Salonica. Ann. Paleont., 18: 1-34.
ARIF, M. AND RAZA, S., 1991. New findings of Cervidae (Mammalia) from the Upper Siwaliks of Potwar-Mirpur Areas, Pakistan. Proc. Pakistan Cong. Zool., 11: 275-281.
ARIF, M. AND SHAH, S. M., 1991. Cervus rewati sp. nov. (Mammalia, Cervidae) from the upper siwaliks of Pakistan. Pakistan Geolog. Sur. Mem., 17: pt.11.
ARIF, M. AND SHAH, S.M., VOS, J.D., 1991. Cervus triplidens (Mammalia, Cervidae) from the Upper Siwaliks of Pakistan. Pakistan geolog. Sur. Mem., 17: pt.11.
AZANZA, B. AND MORALES, J., 1994. Tethytragus nov. gen et Gentrytragus nov. gen. Deux nouveaux Bovidés (Artiodactyla, Mammalia) du Miocène moyen. Proc. Koninklijke Nederlandse Akademie van Wetenschappen, B97: 249-282.
AZZAROLI, A. S., 1954. Critical observations upon Siwalik deer. Proc. Linn. Soc. Lond., pp. 75-87.
BAKALOV, P. AND NIKOLOV, I., 1962. Tertiary mammals. In: Fossils of Bulgaria 10 (ed. V. Tzankov). Sofia: Acad. des Sci. de Bulg. [in Bulgarian].
BARRY, J.C., LINDSAY, E.H. AND JACOBS, L.L., 1982. A biostratigraphic zonation of the middle and upper Siwaliks of the Potwar Plateau of northern Pakistan. Palaeogeogr., Palaeoclimat., Palaeoecol., 37: 95-130.
BARRY, J.C., MORGAN, M.E., WRINKLER, A.J., FLYNN, L.J., LINDSAY, E.H., JACOBS, L.L. AND PILBEAM, D., 1991. Faunal interchange and Miocene terrestrial vertebrates of southern Asia. Paleobiology, 17: 231-245.
BARRY, J., MORGAN, M., FLYNN, L., PILBEAM, D., BEHRENSMEYER, A.K., RAZA, S., KHAN, I., BADGELY, C., HICKS, J. AND KELLEY, J., 2002. Faunal and Environmental change in the Late Miocene Siwaliks of Northern Pakistan. Paleobiology, 28: 1-72.
BARRY, J.C., COTE, S., MACLATCHY, L., LINDSAY, E.H., KITYO, R. AND RAJPAR, A.R., 2005. Oligocene and Early Miocene Ruminants (Mammalia, Artiodactyla)
from Pakistan and Uganda, Palaeont. Electron., 8: 1-29. BASU, P.K., 2004. Siwalik mammals of the Jammu Sub-
Himalaya, India: an appraisal of their diversity and habitats. Quart. Int., 117: 105-118.
BEHRENSMEYER, A. AND BARRY, J., 2005. Biostratigraphic Surveys in the Siwaliks of Pakistan. A Method for Standardized Surface Sampling of the Vertebrate Fossil Record. Palaeont. Electron., 8: 1-24.
BEHRENSMEYER, A.K., WILLIS, B.J. AND QUADE, J., 1995. Floodplains and paleosols of Pakistan Neogene and Wyoming Paleogene deposits: a comparative study. Palaeogeogr., Palaeoclimat., Palaeoecol., 115: 37–60.
BEHRENSMEYER, A.K., 1987. Miocene fluvial facies and vertebrate and vertebrate taphonomy in northern Pakistan. In: Recent developments in fluvial sedimentology (eds. F.G. Ethridge. R.M. Flores and M.D. Harvey). Society of Economic Paleontologists and Mineralogists. Spec. Public., 39: 169-176.
BEHRENSMEYER, A.K. AND TAUXE, L., 1982. Isochronous fluvial systems in Miocene deposits of northern Pakistan. Sedimentol., 29: 331-352.
BERGGERN, W.A., KENT, D.V., FLYNN, L.J. AND VAN COUVERING, J.A., 1985. Cenozoic geochronology. Geol. Soc. Am. Bull., 96: 1407-1418.
BERNOR, R.L., 1986. Mammalian biostratigraphy, geochronology, and zoogeographic relationships of the Late Miocene Maragheh fauna, Iran. J. vert. Palaeont., 6: 76-95.
BHATTI, Z. H., 2005. Taxonomy, evolutionary history and biogeography of the Siwalik Giraffids. Ph.D. dissert. (unpublished), University of the Punjab, Lahore, Pakistan.
BIBI, F., 2007. Origin, Paleoecology, and Paleobiogeography of early Bovini. Palaeogeogr., Palaeoclimat., Palaeoecol., 248: 60-72.
BIBI, F. AND GULUC, E., 2008. Bovidae (Mammalia: Artiodactyla) from the Late Miocene of Sivas, Turkey. J. vert. Palaeont., 28: 501–519.
BIBI, F., BUKHSIANIDZE, M., GENTRY, A.W., GERAADS, D., KOSTOPOULOS, D.S. AND VRBA, E. S., 2009. The Fossil Record and Evolution of Bovidae: State of the Field. Palaeont. Electron., 12: 1-11.
BLAINVILLE, H.M., 1816. Sur plusieurs espèces d animaux mammifères de 1’ order des Ruminants. Bull. Soc. Phil. Paris, pp. 73-82.
BOHLIN, B., 1941. Gazella (Protetraceros) gaudryi (Schlosser) and Gazella dorcadoides Schlosser. Bull. Geolog. Instit., University of Upsala, 28: 79–122.
BONIS, L. DE AND KOUFOS, G. D., 1999. The Miocene large mammal succession in Greece. In: Hominoid Evolution and climatic change in Europe (eds. J. Agusti, L. Rook and Andrews P), volume 1: The Evolution of Neogene Terrestrial Ecosystems in Europe. Cambridge University Press, Cambridge: pp. 205-237.

M.A. KHAN ET AL.
82
BOUVRAIN, G., 1979. Un nouveau genre de Bovide de la fin du Miocene. Bull. Soc. Geolog. Fran., Paris, 21: 507-511.
BOUVRAIN, G., 1982. Revision du genre Prostrepsiceros Major 1891. Palaontolog. Zeits., 56: 113-124.
BOUVRAIN, G., 1988. Les Tragoportax (Bovidae, Mammalia) des gisements du Miocène supérieur de Dytiko (Macédonie, Grèce). Annls. Paléontol., 74: 43-63.
BOUVRAIN, G., 1994. Un bovidé du Turolien inférieur d’Europe orientale: Tragoportax rugosifrons. Annls. Paléontol., 80: 61-87.
BOUVRAIN, G. AND BONIS, L. DE, 1983. Étude d’un Miotragocère du Miocène supérieur de Macédoine (Grèce). In: Actes du symposium paléontologique G. Cuvier (eds. E. Buffetaut, J. Mazin and E. Salmon). Montbéliard, 1982: 35-51.
BOUVRAIN, G. AND THOMAS, H., 1992. Une antilope a chevilles spiralees: Prostrepsiceros zitteli (Bovidae). Miocene superior du Jebel Hamrin en Irak. Geob., 25: 525-533.
BOUVRAIN, G., SEN, S. AND THOMAS, H., 1994. Un nouveau genre d’antilope dans le Miocène supérieur de Sinap en Turquie. Rev. Paléobiol., 13: 375-380.
BROWN, B., 1926. A new deer from the Siwalik. Am. Mus. Novit., 242: 1-6.
CANDE, S.C. AND KENT, D.V., 1995. Revised calibration of the geomagnetic polarity time scale for the Cretaceous and Cenozoic. J. Geophy. Resear., B100: 6093-6095.
CHEN, G.F., 1988. Remarks on the Oioceros species (Bovidae, Artiodactyla, Mammalia) from the Neogene of China. Vertebr. Palasiat., 26: 169-172.
CHEN, G., 1997. The genus Gazella Blainville, 1816 (Bovidae, Artiodactyla) from the Late Neogene of Yushe Basin, Shanxi, Province, China. Vertebr. Palasiat., 35: 233–249.
COLBERT, E.H., 1935. Siwalik mammals in the American Museum of Natural History. Trans. Am. Phil. Soc., N. Ser., 26: 1-401.
COOKE, H. AND WELLS, L., 1951. Fossil Remains from Chelmer, Near Bulawayo, Southern Rhodesia. S. Afr. J. Sci., 47: 205–209.
DENNELL, R.W., COARD, R. AND TURNER, A., 2008. Predators and Scavengers in Early Pleistocene southern Asia. Quat. Int., 192: 78-88.
DOWNING, K.F., LINDSAY, E.H., DOWNS, W.R. AND SPEYER, S.E., 1993. Lithostratigraphy and vertebrate biostratigraphy of the early Miocene Himalayan Foreland, Zinda Pir Dome, Pakistan. Sediment. Geol., 87: 25-37.
FAROOQ, U., 2006. Studies of evolutionary trends in dentition of the Siwalik Tragulids. Ph.D. dissertation (unpublished), University of the Punjab, Lahore, Pakistan.
FAROOQ, U., KHAN, M. A., AKHTAR, M. AND KHAN,
A.M., 2007a. Dorcatherium minus from the Siwaliks, Pakistan. J. Anim. Plan. Sci., 17: 86-89.
FAROOQ, U., KHAN, M.A., AKHTAR, M. AND KHAN, A.M., 2007b. Dorcabune anthracotherioides (Artiodactyla, Ruminantia, Tragulidae) from Hasnot, the Middle Siwaliks, Pakistan. Pakistan J. Zool., 39: 353-360.
FAROOQ, U., KHAN, M.A., AKHTAR, M. AND KHAN, A.M., 2008. Lower Dentition of Dorcatherium majus (Tragulidae, Mammalia) in the Lower and Middle Siwaliks (Miocene) of Pakistan. Turk. J. Zool., 32: 91-98.
FLYNN, L.J., 2003. Small mammal indicators of forest paleo-environment in the Siwalik deposits of the Potwar Plateau, Pakistan. Deins., 10: 183-196.
FLYNN, L.J., BARRY, J.C., MORGAN, M.E., PILBEAM, D., JACOBS, L.L. AND LINDSAY, E.H., 1995. Neogene Siwalik Mammalian Lineages: Species longevities, rates of change, and modes of speciation. Palaeogeogr., Palaeoclimat., Palaeoecol., 115: 249-264.
FLYNN, L.J. AND QI, G.Q., 1982. Age of the Lufeng, China, hominoid locality. Nat., 298: 746–747.
FRIEDMAN, R., GEE, J., TAUXE, L., DOWNING, K. AND LINDSAY, E., 1992. The magnetostratigraphy of the Chitarwata and lower Vihowa formations of the Dera Ghazi Khan area, Pakistan. Sediment. Geol., 81: 253-268.
GAUR, R., 1987. Environment and Ecology of Early Man in Northwest India. B.R. Publishing Corporation, New Delhi.
GENTRY, A. AND HEINZMANN, E.P.J., 1996. Miocene ruminants of the Central and Eastern Tethys and Paratethys. In: The evolution of Western Eurasian Neogene mammal faunas (eds. R.L. Bernor, V. Fahlbusch and H.W. Mittmann): New York, Columbia University Press, pp. 378-391.
GENTRY, A.W., 1966. Fossil Antilopini of East Africa. Bull. Br. Mus. nat. Hist. (Geol.), 12: 45-106.
GENTRY, A.W., 1970. The Bovidae (Mammalia) of the Fort Ternan fossil fauna. In: Fossil vertebrates of Africa (eds. L. S. B. Leakey and R. J. G. Savage), Vol. 2. Academic Press, London. pp. 243-323.
GENTRY, A. W., 1971. The earliest goats and other antelopes from the Samos Hipparion fauna. Bull. Br. Mus. nat. Hist. Geol. Ser., 20: 229–296.
GENTRY, A.W., 1978. Bovidae. In: Evolution of African mammals (eds. V.J. Maglio and H.B.S. Cooke),. Harvard University Press, Cambridge, Massachusetts. pp. 540-572.
GENTRY, A.W., 1980. Fossil Bovidae (Mammalia) from Langebaanweg, South Africa. Ann. S. Afr. Mus., 79: 213-337.
GENTRY, A.W., 1985. The Bovidae of the Omo Group deposits, Ethopia. In: Les faunes Plio-Pleistocenes de la Basse Vallee de l’Omo (Ethopia), tome 1,

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
83
Perissodactyles-Artiodactyles (Bovidae), CNRS, Paris, pp. 119-191.
GENTRY, A.W. AND HOOKER, J.J., 1988. The phylogeny of Artiodactyla. In: The Phylogeny and Classification of the Tetrapods, Vol. 2: Mammals, (ed. M. J. Benton), Systematics Association Special Volume No. 35B Clarendon, Oxford. pp. 235-272
GENTRY, A.W., 1990. Ruminant artiodactyls of Pasalar, Turkey. J. hum. Evol., 19: 529-550.
GENTRY, A. W., 1992. The subfamilies and tribes of Bovidae. Mamm. Rev., 22: 1-32.
GENTRY, A.W., 1994. The Miocene differentiation of Old World Pecora (Mammalia). Hist. Biol., 7: 115-158.
GENTRY, A.W., 1997. Fossil ruminants (Mammalia) from the Manonga Valley, Tanzania. In: Neogene paleontlogy of the Manonga Valley, Tanzania, (ed. T.J. Harrison). Plenum Press, New York. pp. 107-135
GENTRY, A.W., 1999. Fossil Pecorans from the Baynunah Formation, Emirate of Abu Dhabi, United Arab Emirates. Chap. 22 in Abu Dhabi pecorans, Yale University Press, New Haven, 290-316.
GENTRY, A.W., 2003. Ruminantia (Artiodactyla). In: Geology and Paleontology of the Miocene Sinap Formation, Turkey (eds. M. Fortelius, J. Kappelman, S, Sen and R. Bernor, R.). Columbia University Press, New York. pp. 332– 379.
GENTRY, A.W., ROSSNER, G.E., HEIZMANN, E.P.S., 1999. Suborder Ruminantia. In: The Miocene land mammals of Europe (eds. G.E. Rossner and K. Heissig): Munchen, Verlag Dr. Friedrich Pfeil, pp. 225-258.
GERAADS, D., SPASSOV, N. AND KOVACHEV, D., 2003. Palaeoreas lindermayeri (Wagner, 1848) from the upper Miocene of Bulgaria, and a revision of the species. Geodivers., 25: 405-415.
GERAADS, D., 1989. Vertebres fossils du Miocene superieur du Djebel Krechem el Artsouma (Tunisie centrale): Comparisons biostratigraphiques. Geob., 22: 777-801.
GHAFFAR, A., 2005. Studies on equids, cervids and Carnivora from the Siwalik Hills of Pakistan. Ph.D. dissertation (unpublished), University of the Punjab, Lahore, Pakistan.
GHAFFAR, A., AKHTAR, M., KHAN, M.A. AND NAZIR, M., 2004. Report on Cervus sivalensis from the Upper Siwaliks of Pakistan. Punjab Univ. J. Zool., 19: 83-88.
GROMOLARD, C., 1980. Une nouvelle interpretation des grands Bovidae (Artiodactyla, Mammalia) du Pliocene d’Europe occidentale classes jusqu’a present dans le genre Parabos: Parabos cordieri (de Christol) emend., ?Parabos boodon (Gervais) et Alephis lyrix n. gen., n. sp. Geob., 13: 767-775.
GUO, J., DAWSON, M.R. AND BEARD, K.C., 2000. Zhailimeryx, a new lophiomerycid artiodactyl (Mammalia) from the late middle Eocene of central China and the early evolution of ruminants. J. Mamm. Evol., 7: 239-258.
HAILE-SELASSIE, Y., WOLDE GABRIEL, G., WHITE, T., BERNOR, R.L., DEGUSTA, D., RENNE, P.R., HART, W.K., VRBA, E.S., STANLEY, A. AND HOWELL, F.C. 2004. Mio-Pliocene mammals from the Middle Awash, Ethiopia. Géob., 37: 536-552.
HAN, DE-FEN., 1974. First discovery of Dorcabune in China. Vertbr. Palasiat., 12: 217-221.
HEINTZ, E., 1971. Gazella deperdita (Gervais) 1847 (Bovidae, Artiodactyla, Mammalia) du Pontien du Mont Lubéron, Vaucluse, France. Annls. Paléontol., 57: 209–229.
HEISSIG, K., 1972. Palaontologische und geologische untersuchungen im tertiar von Pakistan. 5. Rhinocerotidae(Mamm.) aus den unteren und mittleren Siwalik- Schichten. Abh. Bayer. Akad. Wiss., Munich (NF), 152: 1-112.
IQBAL, M., LIAQAT, A., KHAN, M. A. AND AKHTAR, M., 2009. Some New Remains of Hipparion from the Dhok Pathan Type Locality, Pakistan. J. Anim. Plan. Sci., 19: 154-157.
JANIS, C. M., DAMUTH, J. AND THEODOR, J. M., 2002. The origins and evolution of the North American grassland biome: the story from the hoofed mammals. Palaeogeogr., Palaeoclimat., Palaeoecol., 177: 183–198.
JOHNSON, G.D., ZEITLER, P., NAESER, C.W., JOHNSON, N.M., SUMMERS, D.M., FROST, C.D., OPDYKE, N.D. AND TAHIRKHELI, R. A., 1982. The occurrence and fission-track ages of Late Neogene and Quaternary volcanic sediments, Siwalik group, northern Pakistan. Palaeogeogr., Palaeoclimat., Palaeoecol., 37: 63-93.
JOHNSON, N.M., OPDYKE, N.D., JOHNSON, G.D., LINDSAY, E.H. AND TAHIRKHELI, R.A., 1982. Magnetic polarity, stratigraphy and ages of Siwalik group rocks of the Potwar Plateau, Pakistan. Palaeogeogr., Palaeoclimat., Palaeoecol., 37: 17-42.
JOHNSON, N.M., STIX, J., TAUXE, L., CERVENY, P.F. AND TAHIRKHELI, R.A., 1985. Paleomagnetic chronology, fluvial processes and tectonic implications of the Siwalik deposits near Chinji Village. Pakistan J. Geol., 93: 27-40.
KAHLKE, H.D., 1976. Die Cervidenreste aus den Travertinen von Taubach. Quartarpalaontol., 2: 209–223.
KAPPELMAN, J.W., 1986. The paleoecology and chronology of the Middle Miocene hominoids from the Chinji formation of Pakistan. Ph.D. dissertation., Harvard University.
KHAN, A.M., 2010. Taxonomy and Distribution of Rhinoceros from the Siwalik Hills of Pakistan. Ph.D. dissertation (unpublished), University of the Punjab, Lahore, Pakistan.
KHAN, M. A., 2007. Taxonomic Studies on Fossil Remains of Ruminants from Tertiary Hills of Hasnot, Pakistan. Ph.D. dissertation (unpublished), University of the Punjab, Lahore, Pakistan.
KHAN, M.A., GHAFFAR, A., FAROOQ, U. AND AKHTAR,

M.A. KHAN ET AL.
84
M., 2007. New Fossil remains of Selenoportax vexillarius from the Late Miocene of Hasnot. Pakistan J. Zool., 39: 333-338.
KHAN, M.A., 2008. Fossil bovids from the late Miocene of Padri, Jhelum, Pakistan. Pakistan J. Zool., 40: 25-29.
KHAN, M.A., IQBAL, M., GHAFFAR, A., KHAN, A.M. AND AKHTAR, M. 2008a. Collection of Selenoportax from the Kundi Wala Kas, Hasnot, Pakistan. Pakistan J. Zool., 40: 303-307.
KHAN, M.A., AHMAD, Z. AND AKHTAR, M., 2008b. Selenoportax vexillarius from Dhok Pathan Formation of the Middle Siwaliks, Pakistan. Biologia (Pakistan), 54: 65-72.
KHAN, M. A., ILIOPOULOS, G. AND AKHTAR, M., 2009a. Boselaphines (Artiodactyla, Ruminantia, Bovidae) from the Middle Siwaliks of the Hasnot, Pakistan. Geob., 42: 739-753.
KHAN, M.A., IQBAL, M., AKHTAR, M. AND HASSAN, M., 2009b. Chalicotheres in the Siwaliks of Pakistan. Pakistan J. Zool., 41: 429-435.
KOHLER, M., 1987. Boviden des turkischen Miozans (Kanozoikum und Braunkohlen der Turkei 28). Paleont. Evolucio, 21: 133-246.
KOHLER, M., 1993. Skeleton and habitat of recent and fossil ruminants. Munchner Geowiss. Abh. (A) 25: 88.
KOSTOPOULOS, D.S., 2005. The Bovidae (Mammalia, Artiodactyla) from the late Miocene of Akkasdagı, Turkey. Geodivers., 27: 747–791.
KOSTOPOULOS, D.S., 2006. Late Miocene Vertebrate Locality of Perivolaky, Thessaly, Greece. Palaeontographica Abt., 276: 151–183.
KOSTOPOULOS, D.S., 2009. The late Miocene Mammal Faunas of the Mytilinii Basin, Samos Island, Greece: New Collection. 14. Bovidae. Beitr. Paläont., 31: 339–383.
KOSTOPOULOS, D.S. AND KOUFOS, G.D., 1996. Late Miocene bovids (Mammalia, Artiodactyla) from the locality “Nikiti-1” (NKT), Macedonia, Greece. Annls. Palantolo., 81: 251–300.
KRETZOI, M., 1968. New generic names for homonyms. Vertebr. Hungari., 10: 163-166.
LEHMANN, U. AND THOMAS, H., 1987. Fossil Bovidae (Mammalia) from the Mio-Pliocene of Sahabi (Libya). In: Neogene Paleontology and Geology of Sahabi, Libya (eds. N.T. Boaz, E.L. Arnauti, A.W. Gaziry and Heinzelin J. DE). Liss, New York, pp. 323-335.
LISTER, A.M., 1984. Evolutionary and ecological origins of British Deer. Proc. Roy. Soc. Edinburgh, 82B: 205–229.
LISTER, A.M., 1986. New Results on Deer from Swanscombe, and the Stratigraphical Significance of Deer Remains in the Middle and Upper Pleistocene of Europe. J. Archaeol. Sci., 13: 319–338.
LYDEKKER, R., 1876. Molar teeth and other remains of Mammalia from the India Tertiaries. Pal. Ind., 10: 19-
87. LYDEKKER, R., 1878. Indian Tertiary and Post-Tertiary
vertebrate. 3. Crania of Ruminants. Pal. Ind., 10: 88-181.
LYDEKKER, R., 1880. A sketch of the history of the fossil vertebrata of India. J. Asiat. Soc. Bengal, XLIX: 8-40, Pt.2.
LYDEKKER, R., 1884. Additional Siwalik Perissodactyla and Proboscidea. Mem. Geolog. Sur. India, 3: 1–34.
MATTHEW, W.D., 1929. Critical observations upon Siwalik Mammals. Bull. Am. Mus. Nat. Hist., 56: 435-560.
MERCERON, G., ZAZZO, A., SPASSOV, N., GERAADS, D. AND KOVACHEV, D., 2006. Bovid Palaeoecology and Palaeoenvironments from the Late Miocene of Bulgaria: Evidence from dental microwear and stable isotopes. Palaeogeogr., Palaeoclimat., Palaeoecol., 241: 637–654.
MÉTAIS, G., CHAIMANEE, Y., JAEGER, J. AND DUCROCQ, S., 2001. New remains of primitive ruminants from Thailand: evidence of the early evolution of the Ruminantia in Asia. Zoolog. Scrip., 30: 231–248.
METAIS, G., GUO, J. AND BEARD, K.C., 2004. A New Small Dichobunid Artiodactyl from Shanghuang (Middle Eocene, Eastern China): Implications for the Early Evolution of Proto-Selenodonts in Asia. Bull. Cornell. Mus. Nat. Hist., 36: 177-197.
MOYA-SOLA, S., 1988. Morphology of Lower Molars of the Ruminants (Artiodactyla, Mammalia): Phylogenetic Implication. Paleontol. Evoluc., 22: 61–70.
MOYA-SOLA, S., 1983. Les Boselaphini (Bovidae, Mammalia) del Neogeno de la Peninsula Iberica. Dissertation, Universidad Autonoma de Barcelona, Publ. Geolog., 18: 1-236.
NAKAYA, H., 1994. Faunal change of Late Miocene Africa and Eurasia: mammalian fauna from the Namurungule Formation, Samburu Hills, northern Kenya. Afr. Stud. Monogr. Supplement. Iss., 20: 1–112.
NANDA, A.C., 2008. Comments on the Pinjor Mammalian Fauna of the Siwalik Group in relation to the Post-Siwalik Faunas of Peninsular India and Indo-Gangetic Plain. Quat. Int., 192: 6-13.
OPDYKE, N.D., LINDSAY, E., JOHNSON, G.D., JOHNSON, N., TAHIRKHELI, R.A. AND MIRZA, M.A., 1979. Magnetic polarity, stratigraphy and vertebrate paleontology of the Upper Siwalik Subgroup of northern Pakistan. Palaeogeogr., Palaeoclimat., Palaeoecol., 27: 1-34.
PICKFORD, M., 1988. Revision of the Miocene Suidae of the Indian Subcontinent. Münchner Geowissenschaften Abh., 12: 1-91.
PICKFORD, M., 2001. Africa’s smallest ruminant: A new tragulid from the Miocene of Kenya and the biostratigraphy of East African Tragulidae. Geob., 34: 437–447.

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
85
PICKFORD, M., 2002. Ruminants from the Early Miocene of Napak, Uganda. Annal. de Paléontol., 88: 85−113.
PICKFORD, M., SENUT, B. AND MOURER-CHAUVIRE, C., 2004. Early Pliocene Tragulidae and peafowls in the Rift Valley, Kenya: Evidence for rainforest in East Africa. Compt. Rendus Palevol, 3: 179–189.
PILBEAM, D., BARRY, J., MEYER, G.E., SHAH, S. M.I., PICKFORD, M.H.L., BISHOP, W.W., THOMAS, H. AND JACOBS, L.L., 1977. Geology and palaeontology of Neogene strata of Pakistan. Nat., 270: 684-689.
PILGRIM, G.E., 1910. Notices of new Mammalian genera and species from the Tertieries of India-Calcutta. Rec. geolog. Sur. India, 40: 63-71.
PILGRIM, G.E., 1911. The fossil Giraffidae of India. Mem. geolog. Sur. India. Pal. Ind., N. Ser., 4: 1-29.
PILGRIM, G.E., 1913. Correlation of the Siwaliks with Mammal Horizons of Europe. Rec. geolog. Sur. India, 43: 264-326.
PILGRIM, G.E., 1915. The dentition of the Tragulid genus Dorcabune. Rec. geolog. Sur. India, 45: 226–238.
PILGRIM, G.E., 1926a. The Tertiary formations of India and the interrelation of marine and terrestrial deposits. Proc. Pan-Pacific Sci. Congr., Australia, pp. 896-931.
PILGRIM, G.E., 1926b. The fossil Suidae of India. Mem. geolog. Sur. India, N. Ser., 8: 1-65.
PILGRIM, G.E., 1937. Siwalik antelopes and oxen in the American Museum of Natural History. Bull. Am. Mus. nat. Hist., 72: 729-874.
PILGRIM, G.E., 1939. The fossil Bovidae of India. Pal. Ind., N.S., 26: 1-356.
PILGRIM, G.E. AND HOPWOOD, A.T., 1928. Catalogue of the Pontian Bovidae of Europe in the Department of Geology. Brit. Mus. (Natur. Hist.), London.
PITRA, C., FICKELA, J., MEIJAARD, E. AND GROVES, C. P. 2004. Evolution and phylogeny of old world deer. Molecul. Phylogenet. Evol., 33: 880–895.
QIU, Z.X. AND QIU, Z.D., 1995. Chronological sequence and subdivision of Chinese Neogene mammalian faunas. Palaeogeogr., Palaeoclimat., Palaeoecol., 116: 41–70.
QIU, Z. AND GU, Y., 1991. The Aragonian vertebrate fauna of Xiacaowan-8. Dorcatherium (Tragulidae, Artiodactyla). Vertebr. Palasiat., 29: 21–37. [Chinese 21–31; English 32–37].
QUADE, J. AND CERLING, T.E., 1995. Expansion of C4 grasses in the Late Miocene of Northern Pakistan: evidence from stable isotopes in paleosoles. Palaeogeogr., Palaeoclimat., Palaeoecol., 115: 91–116.
RAZA, S.M., CHEEMA, I.U., DOWNS, W.R., RAJPAR, A.R. AND WARD, S.C., 2002. Miocene stratigraphy and mammal fauna from the Sulaiman Range, Southwestern Himalayas, Pakistan. Palaeogeogr., Palaeoclimat., Palaeoecol., 186:185 –197
ROMER, A.S., 1974. Vertebrate palaeontology, III. The University of Chicago Press, Chicago Illinois, pp.1-687.
ROSSNER, G.E. AND MORS, T., 2001. A new find of the enigmatic Eurasian Miocene ruminant artiodactyls Orygotherium: its meaning for stratigraphic setting and palaeoecological interpretations. J. Vert. Paleont., 21: 591-595.
ROSSNER, G.E., 2010. Systematics and palaeoecology of Ruminantia (Artiodactyla, Mammalia) from the Miocene of Sandelzhausen (southern Germany, Northern Alpine Foreland Basin). Palaont. Zurich, 1-40. (DOI 10.1007/s12542-010-0052-2).
SACH, V. AND HEIZMANN, E.P.J., 2001. Stratigraphie und Sa¨ugetierfaunen der Brackwassermolasse in der Umgebung von Ulm (Sudwestdeutschland). Stutt. Beitr. zur Naturk., B310: 1–95.
SAVAGE, D.E. AND RUSSEL, D.E., 1983. Mammalian Paleofaunas of the World, London. Addison-Wesley, pp. 1-432.
SENYUREK, M.S., 1954. A Horn-core of Gazella capricornis Rodler and Weithofer found at Kucukyozgat. Bell., XVIII: 142–145.
SHAH, S.M.I., 1977. Stratigraphy of Pakistan. Mem. geolog. Sur. Pakistan, 12: 1-138.
SHAH, S.M.I., 1980. Stratigraphy and economic geology of Central Salt Range. Rec. geolog. Sur. Pakistan, 52: 1-104.
SOLOUNIAS, N., 1981. The Turolian fauna from the Island of Samos, Greece. Contr. vert. Evol., 6: 99-232.
SOLOUNIAS, N., BARRY, J.C., BERNOR, R.L., LINDSAY, E.H. AND RAZA, S.M., 1995. The Oldest Bovid from the Siwaliks, Pakistan. J. vert. Paleont., 15: 806-814.
SOMMER, R.S., ZACHOS, F.E., STREET, M., JORIS, O., SKOG, A. AND BENECKE, N., 2008. Late Quaternary Distribution Dynamics and Phylogeography of the Red Deer (Cervus elaphus) in Europe. Quat. Sci. Rev., 27: 714-733.
SPASSOV, N. AND GERAADS, D., 2004. Tragoportax Pilgrim, 1937 and Miotragocerus Stromer, 1928 (Mammalia, Bovidae) from the Turolian of Hadjidimovo, Bulgaria, and a revision of the late Miocene Mediterranean Boselaphini. Geodivers., 26: 339–370.
STEININGER, F.F., 1999. Chronostratigraphy, geochronology and biochronology of the Miocene “European land mammal mega-zones” (ELMMZ) and the Miocene “mammal-zones” (MN-zones). In: Rössner Gertrud, E., Heissig, K., Fahlbusch, V. (Eds.), The Miocene Land Mammals of Europe. Verlag Friedrich Pfeil, Munich, pp. 9–24.
STIX, J., 1982. Stratigraphy of the Kamlial Formation near Chinji Village, northern Pakistan. BA Dissert., Dartmouth College, Hanover, N. H.
SURESH, N., SUMIT, K., ROHTASH, G.K. AND SANGODE, S.J., 2004. Clay-mineral distribution patterns in late Neogene fluvial sediments of the Subathu sub-basin, central sector of Himalayan foreland basin: implications

M.A. KHAN ET AL.
86
for provenance and climate. Sediment. Geol., 163: 265–278.
TAUXE, L. AND OPDYKE, N.D., 1982. A time framework based on magnetostratigraphy for the Siwalik sediments of the Khaur area, northern Pakistan. Palaeogeogr., Palaeoclimat., Palaeoecol., 37: 43-61.
TEDFORD, R.H., FLYNN, L.J., ZHANXIANG, Q., OPDYKE, N.D. AND DOWNS, W.R., 1991. Yushe Basin, China: Paleomagnetically calibrated mammalian biostratigraphic standard for the late Neogene of eastern Asia. J. vert. Paleont., 11: 519–526.
THOMAS, H., 1979. Les Bovides miocenes des rifts est-africains: Implications paleobiogeogarphiques. Bull. Geol. Soc. France, 7e Series, XXI: 295-299.
THOMAS, H., 1980. Les bovides du Miocene superieur des couches de Mpesida et de la formation de Lukeino (district de Baringo, Kenya). In: Proc. 8th Pan-African Congr. Prehistory, Nairobi, 1977 (eds. R.E.F. Leakey and B.A. Ogot), Nairobi, pp. 82-91.
THOMAS, H., 1981. Les bovides miocenes de la formation de Ngorora du Basin de Baringo (Kenya), Proc. K. Nederl. Akad. Wetensch. B84: 335-409.
THOMAS, H., 1984. Les bovides ante-hipparions des Siwaliks inferieurs (plateau du Potwar), Pakistan. Mem. de la Soc. Geologique de France, Paris, 145: 1-68.
VRBA, E.S., 1995. The fossil record of African antelopes (Mammalia, Bovidae) in relation to human evolution and palaeoclimate. In: Paleoclimate and Evolution: with emphasis on human origins (eds. E.S. Vrba, G.H. Denton, T.C. Partridge and L. H. Burkle), Yale University Press, New Haven. pp. 385–424.
VRBA, E.S. AND HAILE-SELASSIE, Y., 2006. A New Antelope, Zephyreduncinus oundagaisus (Reduncini, Artiodactyla, Bovidae), from the Late Miocene of the Middle Awash, Afar Rift, Ethiopia. J. vert. Paleont., 26: 213 –218.
VRBA, E.S. AND SCHALLER, G., 2000. Phylogeny of Bovidae based on behavior, glands, skulls, and postcrania. In Antelopes, deer and relatives (eds. E.S. Vrba and G. Schaller). Yale University Press, New Haven. pp. 203-222.
WEST, R.M., 1980. A minute new species of Dorcatherium (Tragulidae, Mammalia) from the Chinji Formation near Daud Khel, Mianwali district, Pakistan. MILW. Public Mus. Contr. Biol. Geol., 3: 1-6.
WHITWORTH, T., 1958. Miocene ruminants of East Africa. Fossil Mammals of Africa. Bull. Brit. Mus. nat. Hist., 15: 1-50.
WILLIS, B.J. AND BEHRENSMEYER A.K., 1995. Fluvial systems in the Siwalik Neogene and Wyoming Paleogene. Palaeogeogr., Palaeoclimat., Palaeoecol., 114: 13-35.
WILLIS, B., 1993a. Ancient river systems in the Himalayan foredeep, Chinji village area, northern Pakistan. Sediment. Geol., 88: 1-76.
WILLIS, B., 1993b. Evolution of Miocene fluvial systems in the Himalayan foredeep through a two kilometer thick succession in northern Pakistan. Sediment. Geol., 88: 77-121.

LATE MIOCENE ARTIODACTYLS IN THE DHOK PATHAN
87
Appendix I
List of the studied artiodactyls from Dhok Pathan. Artiodactyla Bovidae Tragoportax T. punjabicus T. browni T. cf. salmontanus Miotragocerus M. large sp. M. cf. gluten Pachyportax P. latidens cf. P. nagrii Selenoportax S. cf. vexillarius Prostrepsiceros P. cf. vinayaki Gazella G. lydekkeri Kobus K. porrecticornis cf. Kobikeryx sp. Caprotragoides C. cf. potwaricus Cervidae Cervus C. cf. sivalensis C. cf. triplidens Tragulidae Dorcatherium D. majus Dorcabune D. anthracotherioides D. cf. nagrii Giraffidae Hydaspitherium H. cf. megacephalum H. cf. grande H. sp. Suidae Propotamochoerus P. hysudricus Hippopotamodon ?H. sp.
Appendix II
Studied material
Tragoportax punjabicus Skull: PUPC 86/149, opisthocranium. PUPC 86/150,
opisthocranium. Horn-core: PUPC 86/264, broken at the apex, left side. Upper dentition: PUPC 83/743, isolated right P3. PUPC 87/327, isolated left P3. PUPC 00/21, isolated left P3. PUPC 00/89, right P4-M1. PUPC 83/671, isolated right M1. PUPC 87/677, isolated right M1. PUPC 83/673, isolated left M1. PUPC 83/30, isolated left M1. PUPC 83/676, isolated right M1. PUPC 83/674, isolated right M1. PUPC 97/13, isolated right M1. PUPC 00/80, right M1-2. PUPC 85/242, isolated right M2. PUPC 97/06, isolated left M2. PUPC 83/603, isolated right M2. PUPC 83/10, isolated left M2. PUPC 83/666, isolated right M2. PUPC 00/01, isolated left M2. PUPC 83/693, isolated right M2. PUPC 08/126, palatine parts of both maxillae with right P3-M3 and left P2-M3. PUPC 83/275, right maxillary fragment with P3-M3. PUPC 97/10, right maxillary fragment with M1-M3. PUPC 83/665, isolated left M3. PUPC 97/7, isolated right M3. PUPC 83/688, isolated right M3. Lower dentition: PUPC 83/712, isolated left p2. PUPC 97/12, left mandible fragment with p2-p4. PUPC 83/204, isolated left p3. PUPC 87/324, isolated left p3. PUPC 83/711, isolated left p4. PUPC 68/237, isolated left p4. PUPC 97/3, isolated right m1. PUPC 87/282, isolated left m1. PUPC 86/26, isolated left m1. PUPC 83/700, isolated right m1. PUPC 86/25, isolated left m2. PUPC 83/617, isolated left m2. PUPC 99/28, isolated right m2. PUPC 83/701, isolated left m2. PUPC 83/680, isolated left m3. PUPC 83/767, isolated left m3. PUPC 83/277, left mandible fragment with m3.
Tragoportax browni Opisthocranium: PUPC 86/204. Horn-cores: PC-GCUF
09/11, almost complete, side indeterminate. PUPC 87/1, fragment, side indeterminate. Upper dentition: PUPC 87/281, isolated left P3. PC-GCUF 09/42, isolated right P3. PC-GCUF 09/43, isolated right P4. PUPC 83/689, isolated left M1. PUPC 86/151, left maxillary fragment with M1-M2. PUPC 86/29, left maxillary fragment with M1-M2. PUPC 83/861, left maxillary fragment with M1-M2. PUPC 97/80, isolated left M2. PUPC 83/662, isolated right M2. PUPC 87/324, right maxillary fragment with M2-M3. PUPC 83/283, right maxillary fragment with M2-M3. Lower dentition: PUPC 99/27, right mandible fragment with m1-m2. PUPC 83/265, left mandible fragment with m2-m3. PUPC 83/269, right mandible fragment with m2-m3. PUPC 97/11, left mandible fragment with m3.
Tragoportax cf. salmontanus Horn-core: PUPC 93/20, incomplete, left side.

M.A. KHAN ET AL.
88
Miotragocerus large sp. Upper dentition: PUPC 83/275, isolated right P2. PUPC
86/33, isolated right P3. PUPC 86/39, isolated left P3. PUPC 09/44, right maxillary ramus with P2-P4. PUPC 83/209, left maxillary ramus with P2-M1. PUPC 87/02 and PUPC 86/308, right maxillary ramus with DP4-M1. PUPC 87/246, left maxillary ramus with DP3-M2. PUPC 87/115, left maxillary ramus with DP4-M2. PUPC 87/116 and PUPC 87/117, right and left maxillary rami with M1-M2. Lower dentition: PUPC 88/740, left mandibular ramus with p2. PUPC 83/703, isolated left p2. PUPC 83/708 and PUPC 83/709, isolated right and left p3s. PUPC 83/742 and PUPC 87/294, isolated right and left p4s. PUPC 93/277, left mandibular ramus with p4-m1. PUPC 09/86, left mandibular ramus with p4-m3. PUPC 96/05, left mandibular ramus with p4-m1. PUPC 83/277, right mandibular ramus with m2 and broken m3.
Miotragocerus cf. gluten Horn-core: PUPC 01/01, incomplete, left side. Pachyportax latidens Horn-core: PUPC 96/18, basal fragment, right side.
Upper dentition: PUPC 80/646, isolated M2. PUPC 80/203, isolated right M2. PUPC 09/74, isolated M2. Lower dentition: PUPC 08/87, right mandibular ramus with m3 with broken hypoconulid and an anterior part of m2.
cf. Pachyportax nagrii Upper dentition: PUPC 99/50, partially damaged
isolated ?M3. Selenoportax cf. vexillarius Lower dentition: PUPC 99/47, right mandibular ramus
with m2 and roots of m1. PUPC 99/22, isolated left semierupted molar. PUPC 00/45, isolated left m2. PUPC 07/16, partially damaged molar. PUPC 97/02, isolated right m3.
Prostrepsiceros cf. vinayaki Horn-cores: PUPC 87/319, almost complete, left side.
PUPC 87/318, incomplete, probably right side. Gazella lydekkeri Horn-cores: PUPC 83/319, almost complete with a
piece of frontal, broken at the apex, left side. PC-GCUF 09/45, almost complete with supraorbital pit, left side. Lower dentition: PUPC 08/136, right isolated m2. PUPC 07/13 and PUPC 09/76, isolated left m3s.
Kobus porrecticornis Horn-cores: PUPC 87/124 A+B, left and right horn-
cores, proximally preserved fragments, perhaps same individual. PUPC 02/126 and PUPC 87/180, partial left horn-cores.
cf. Kobikeryx sp. Horn-cores: PUPC 99/49, distally preserved fragments,
side indeterminate. Caprotragoides cf. potwaricus Horn-core: PC-GCUF 09/44, right side. Cervus cf. sivalensis Lower dentition: PUPC 04/18, right mandibular ramus
with m2-3. PUPC 09/73, right mandibular ramus with m2.
Cervus cf. triplidens Upper dentition: PUPC 09/71, right maxillary ramus
with P4-M2. Dorcatherium majus Upper dentition: PUPC 03/14, isolated left M2. Lower
dentition: PUPC 01/21, isolated right m1. PUPC 01/26, left mandibular ramus with m2 and posterior portion of m1. PUPC 07/09, isolated left m2.
Dorcabune anthracotherioides Upper dentition: PUPC 99/338, isolated right M2.
Lower dentition: PUPC 04/01, right mandibular ramus with m3.
Dorcabune cf. nagrii Upper dentition: PUPC 96/45, an isolated left probably
M1. Hydaspitherium cf. megacephalum Lower dentition: PUPC 09/78, left mandibular ramus
with broken m1, complete m2 and partially erupted m3. Hydaspitherium cf. grande Lower dentition: PUPC 09/77, right mandibular ramus
with m2-3. ?Hydaspitherium sp. Lower dentition: PUPC 83/267, right mandibular ramus
with m2-3. Propotamochoerus hysudricus Lower dentition: PUPC 07/32, left mandibular ramus
with p4-m3; the p3 roots are also preserved in the alveoli. PUPC 99/02, left mandibular ramus with p4-m1. PUPC 94/65, right mandibular ramus with p4-m2. PUPC 97/15, right isolated m3.
?Hippopotamodon sp. Lower dentition: PUPC 00/14, left mandibular ramus
with broken p4 and partially complete m1. PUPC 08/26, badly crushed right mandibular ramus with damaged m1-3, probably juvenile animal.