ichnofacies and microbial build-ups on late miocene rocky shores from menorca (balearic islands),...
TRANSCRIPT

Facies
DOI 10.1007/s10347-010-0240-2ORIGINAL ARTICLE
Ichnofacies and microbial build-ups on Late Miocene rocky shores from Menorca (Balearic Islands), Spain
Markes E. Johnson · B. Gudveig Baarli · Ana Santos · Eduardo Mayoral
Received: 10 April 2010 / Accepted: 25 September 2010© Springer-Verlag 2010
Abstract Angular unconformites between Jurassic andMiocene strata are exposed in the sea cliVs of Cala Cigonyaon the northwest coast of Menorca in the Balearic Islandsof Spain. The geological discontinuities represent rockyshores on opposite sides of a former headland with 15 m oftopographic relief. On the east Xank, Jurassic dolomite isoverlain by Upper Miocene (Tortonian to Messinian) brec-cia and laminated limestone. Here, a partially exhumeddolomite surface records Miocene bivalve borings of theichnospecies Gastrochaenolites torpedo and G. lapidicusthat achieved a density of >1,000 borings/m2. Other associ-ated traces include sponge borings (Entobia isp.) and poly-chaete borings (Caulostrepsis isp.). A breccia deposit 0.8 mthick was derived from the underlying dolomite and angu-lar clasts still retain evidence of bivalve borings. Above fol-lows a succession of laminated limestone beds more than5 m thick, including some levels with dome-shaped stro-matolites and other horizons with reworked dolomite clasts.Thin-section analysis of the laminated limestone revealsdark and light couplets 0.2 mm thick consistent with micro-bial origins. In contrast, the west Xank was buried by coarsesandstone and laminated sediments. Here, dwelling struc-tures of regular echinoids (Circolites kotoncensis) are thedominant traces preserved on the dolomite surface, reach-ing a maximum density of 66 borings/m2. Associatedborings include Entobia geometrica as well as rare traces of
Gastrochaenolites isp. and Trypanites isp. Notable for theabsence of a basal Miocene breccia, the west Xank is inter-preted as a sheltered rocky shore coeval with an exposedrocky shore on the east Xank. Today, heavy surf on thenorth coast of Menorca is related to the Tramontana windsthat blow out of Spain during winter months. Similar atmo-spheric circulation patterns must have prevailed during theLate Miocene, but the replacement of ichnofacies by micro-bial build-ups resulted from increased salinity during theMessinian crisis.
Keywords Bioerosion · Coastal dynamics · Mediterranean Sea · Tortonian and Messinian stages (Upper Miocene) · Stromatolites
Introduction
The Mediterranean shores of southern Europe are 63%dominated by rocky shores (Johnson 1988a, cf. Bird andSchwartz 1985). Many Mediterranean islands exhibit evenlarger proportions of rocky shores. Mallorca, the largest ofSpain’s Balearic Islands for example, is famous for itsbeaches but modern rocky shores account for about 90% ofits 550-km coastline (Gomila 1999a). The neighboringisland of Menorca with a 216-km coastline, has much thesame breakdown between contemporary sandy and rockyshores (Gomila 1999b). Extensive Mesozoic and Cenozoiclimestone strata form prominent sea cliVs around bothislands. Patterns of regional tectonics and sea-level changesrecorded in these strata show ample evidence for the occur-rence of former rocky shorelines. Much research has beendevoted to the wide carbonate ramps and reefs that accruedduring Late Miocene time most extensively along thesouthern margins of older, tectonically deformed Paleozoic
M. E. Johnson (&) · B. G. BaarliDepartment of Geosciences, Williams College, Williamstown, MA 01267, USAe-mail: [email protected]
A. Santos · E. MayoralDepartamento de Geodinámica y Paleontología, Facultad de Ciencias Experimentales, Universidad de Huelva, Campus de El Carmen, Avd. 3 de Marzo, s/n, 21071 Huelva, Spain
123

Facies
and Mesozoic core rocks on Mallorca and Menorca (Pomar2001; Pomar and Ward 1994; Pomar et al. 2002; 2004;Obrador et al. 1992; Brandano et al. 2005; Mateu-Vicenset al. 2008). In contrast, the northern margins of Mallorcaand Menorca are poorly understood with regard to paleo-shores.
Many studies on Miocene carbonate platforms in theBalearic Islands (as cited, above) Wnd Lower Tortonianramps and a widely developed reef complex marking thetransition between the Lower and Upper Tortonian stages.Younger Messinian strata record the persistence of suchreefs, but also demonstrate facies with stromatolites,oolites, and marl. Stromatolitic facies of Messinian age aresaid to occur on all the other Balearic Islands, but not Men-orca (Asprion et al. 2009, p. 39). Mats made by bacterialorganisms (tapissos de microorganisms bacterians) likenedto stromatolites are, however, described by Rosell-Sanuyand Llompart-Diaz (2002, pp. 271–272) from Upper Mio-cene (Messinian) carbonates on Cape Menorca o Bajoli inthe northwestern corner of the island.
The goal of this paper is to portray all aspects of theUpper Miocene rocky-shore deposits seated on a pro-nounced angular unconformity with Jurassic dolomite atCala Cigonya near the great sea arch at Pont d’en Gil. Threedistinct parts to the present study concern: (1) Miocene bioe-rosion of the unconformity surfaces on which the UpperMiocene succession follows, (2) the physical characteristicsof basal Miocene deposits that deWne them as accumulationswith contrasting degrees of wave activity, and (3) variationsin the shape and style of microbial build-ups that completethe Miocene sequence. Microbes that thrived under a widerange of physical conditions have formed sedimentary-organic structures commonly called stromatolites. Thesestructures have an extensive record in marine and fresh-water carbonate rocks through geologic time (Riding 2000),but are rarely associated with rocky shorelines post-datingProterozoic time (Johnson 2006). Hence, the Late Miocenerocky shores of northern Menorca are of special interest.
Geological and geographic setting
Located in the western Mediterranean Sea (Fig. 1a), theBalearic Islands of Spain were aVected by a complex seriesof events from about 54–5.5 million years ago related to thetectonic closure of the Tethys seaway between North Africaand southern Europe (Gueguen et al. 1998; Rosenbaumet al. 2002). Mallorca and Menorca underwent deformationand uplift as a result of NW–SE compression, culminatingabout 30 million years ago (Middle Oligocene) when theislands were linked to the Iberian mainland by a high pla-teau. This area began to rift apart to form the ValencianTrough about 24 million years ago (Early Miocene), and
was Xooded by seawater to create a water barrier betweenthe islands and the mainland. From 18 to 5.5 million yearsago (Middle to Late Miocene), ongoing migration of theApenninic arc and associated back-arc basins led to a east-moving peripheral bulge that increasingly obstructed thewestern opening of the Tethys with the North AtlanticOcean. Culminating at the end of the Miocene 5.33 millionyears ago, the Messinian crisis entailed full seclusion of theTethys, transforming it into the vestigial body of water wecall the Mediterranean Sea. Restricted from replenishmentby normal seawater and isolated under a desert climate, theyoung Mediterranean grew increasingly saline and partiallydried out (Krijgsman et al. 1999; Duggen et al. 2003). Cata-strophic Xooding of the western Mediterranean Sea(Caracuel et al. 2004) may be linked to the tectonic collapseof the land barrier that formerly sealed oV the North Atlan-tic at the Straits of Gibraltar (Doblas and Oyarzun 1989;Platt and Vissers 1989) or to local non-tectonic factors suchas regressive Xuvial erosion (Loget and Van Den Driessche2006).
With an area of 702 km2 (Gomila 1999b), Menorca isthe second largest in the Balearic Islands. The basic geol-ogy of Menorca is represented by a broad Miocene rampthat wraps around the west and south margins of a smallercore of tectonically disturbed Mesozoic rocks (Fig. 1b).The Mesozoic core rocks rise above the rolling countrysideas a morphologically distinct upland with a central
Fig. 1 Location of the study site. a Map of Western Europe and NorthAfrica. b Geological map of Menorca in the Balearic Islands (afterObrador et al. 1992, Fig. 1). c Map of the Menorcan northeast coastshowing geological details around Cala Cigonya (after Rosell-Sanuyand Llompart-Diaz 2002, Fig. 9.1)
Iberian Peninsla
A
B
Menorca
study site
North Africa
Miocene
Jurassic and Cretaceous
Triassic
Paleocene
Ciutadella
Mahon
0 5 10 km
NJurassic
Miocene
dolomite
brecciacalcareniteshelly calcarenite
C1 km
CalaCigonya
123

Facies
maximum elevation of 358 m at Monte Toro (Rosell-Sanuyand Llompart-Diaz 2002).
Sailing directions for Menorca warn of the dangerousnorth coast with its high rocky shores subject to strongwinds, particularly during the winter season whenTramontana winds blow hard oV the Iberian mainland(Gomila 1999b). This is a signiWcant aspect of the coastaldynamics that has possible ramiWcations for Miocene con-ditions in the same region relevant to the adjacent Iberianmainland.
The study site for this project is located 5 km northwestof Ciutadella at Cala Cigonya (Fig. 1b). At this locality,Miocene strata come into direct contact with Jurassic stratawith an angular unconformity exposed in the sea cliVs(Figs. 1c, 2a, b). This particular area was described for itsgeneral geology by Rosell-Sanuy and Llompart-Diaz(2002, pp. 269–274) in their treatment of the great sea archat Pont d’en Gil. Our emphasis is on the ichnological recordof Miocene borings preserved on two related unconformitysurfaces, a topic not previously considered at this localityor elsewhere on Menorca. We provide, also, a more exten-
sive treatment of microbial build-ups conserved in Miocenestrata near the unconformity.
Methods
Multiple criteria (after Johnson 1988b) were used to recog-nize the presence of former rocky shores in the study area:(1) exposure of an unconformity showing a history of sub-aerial exposure and subsequent marine onlap, (2) develop-ment of a basal conglomerate, (3) clast lithology in the basalconglomerate strictly limited by available variation in thelithology of the unconformity surface, (4) joints or Wssureseroded in the unconformity Wlled by the basal conglomerate,(5) occurrence of fossils representing organisms or traces oforganisms specialized for life in a high-energy marine set-ting (borings and/or body fossils associated with the uncon-formity surface and clasts from the overlying conglomerate).
Direct observations on Miocene trace fossils were regis-tered using a 50 £ 50-cm grid on irregular unconformitysurfaces eroded from Jurassic dolomite. Bioerosion structures
Fig. 2 Coastal proWle of the geology exposed in sea cliVs at Cala Cigonya (see Fig. 1c for location). a General perspective looking south over thebay from the sea arch at Pont d’en Gil. b Geological reconstruction showing the folded Jurassic dolomites dipping towards the WSW
123
Facies
were identiWed to ichnospecies where possible, and quanti-Wed. A Jacob’s staV was used to measure original topo-graphic relief on the Miocene rocky shore. Orientations anddip angles were recorded on unconformities representingformer sea cliVs.
An extensive photographic record of fossil borings wasmade. Small hand samples of the Wnely laminated Miocenelimestone were collected for preparation of standard petro-graphic thin sections (4.5 £ 2.5 cm).
Paleotopography and stratigraphy
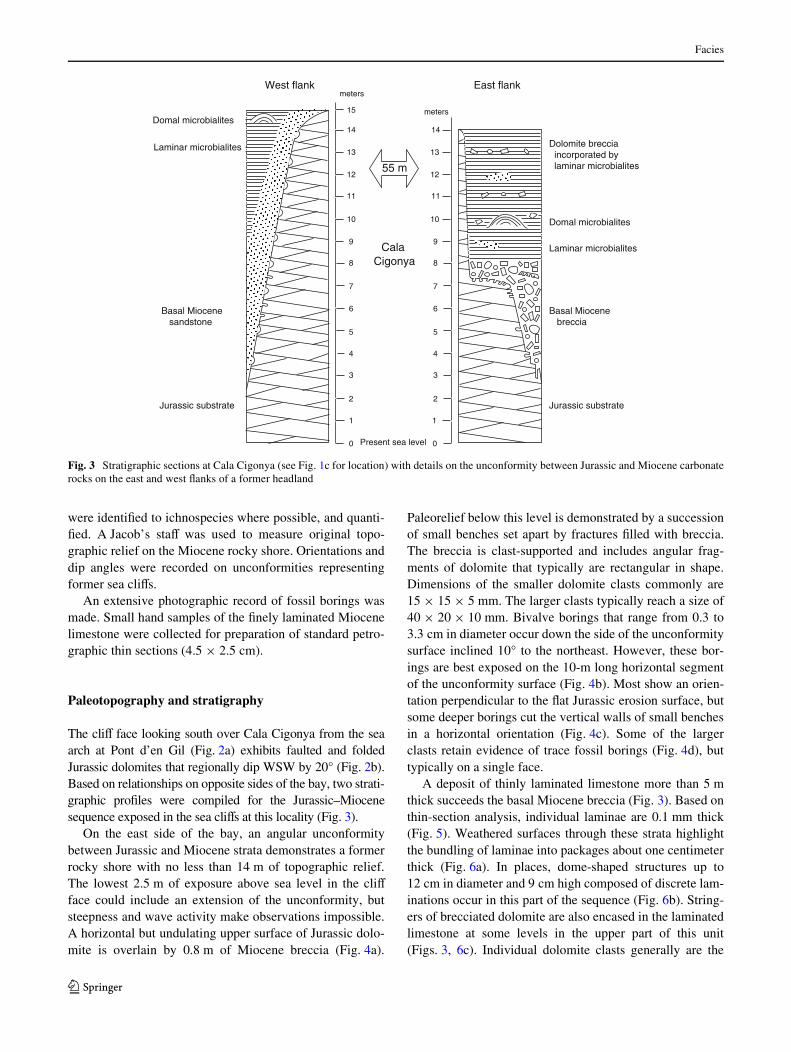
The cliV face looking south over Cala Cigonya from the seaarch at Pont d’en Gil (Fig. 2a) exhibits faulted and foldedJurassic dolomites that regionally dip WSW by 20° (Fig. 2b).Based on relationships on opposite sides of the bay, two strati-graphic proWles were compiled for the Jurassic–Miocenesequence exposed in the sea cliVs at this locality (Fig. 3).
On the east side of the bay, an angular unconformitybetween Jurassic and Miocene strata demonstrates a formerrocky shore with no less than 14 m of topographic relief.The lowest 2.5 m of exposure above sea level in the cliVface could include an extension of the unconformity, butsteepness and wave activity make observations impossible.A horizontal but undulating upper surface of Jurassic dolo-mite is overlain by 0.8 m of Miocene breccia (Fig. 4a).
Paleorelief below this level is demonstrated by a successionof small benches set apart by fractures Wlled with breccia.The breccia is clast-supported and includes angular frag-ments of dolomite that typically are rectangular in shape.Dimensions of the smaller dolomite clasts commonly are15 £ 15 £ 5 mm. The larger clasts typically reach a size of40 £ 20 £ 10 mm. Bivalve borings that range from 0.3 to3.3 cm in diameter occur down the side of the unconformitysurface inclined 10° to the northeast. However, these bor-ings are best exposed on the 10-m long horizontal segmentof the unconformity surface (Fig. 4b). Most show an orien-tation perpendicular to the Xat Jurassic erosion surface, butsome deeper borings cut the vertical walls of small benchesin a horizontal orientation (Fig. 4c). Some of the largerclasts retain evidence of trace fossil borings (Fig. 4d), buttypically on a single face.
A deposit of thinly laminated limestone more than 5 mthick succeeds the basal Miocene breccia (Fig. 3). Based onthin-section analysis, individual laminae are 0.1 mm thick(Fig. 5). Weathered surfaces through these strata highlightthe bundling of laminae into packages about one centimeterthick (Fig. 6a). In places, dome-shaped structures up to12 cm in diameter and 9 cm high composed of discrete lam-inations occur in this part of the sequence (Fig. 6b). String-ers of brecciated dolomite are also encased in the laminatedlimestone at some levels in the upper part of this unit(Figs. 3, 6c). Individual dolomite clasts generally are the
Fig. 3 Stratigraphic sections at Cala Cigonya (see Fig. 1c for location) with details on the unconformity between Jurassic and Miocene carbonaterocks on the east and west Xanks of a former headland
meters
Present sea level
Jurassic substrate
Basal Miocene breccia
Laminar microbialites
Domal microbialites
Dolomite breccia incorporated by laminar microbialites
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
meters
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
Jurassic substrate
Basal Miocene sandstone
Domal microbialites
Laminar microbialites
East flankWest flank
55 m
Cala Cigonya
123

Facies
same size as found in the basal breccia, but rarely come intocontact with one another and appear to Xoat in the lami-nated limestone.
On the opposite side of the bay, the dolomite surfaceinclines southwest with a variable dip ranging from 30 to40°. The entire exhumed surface shows evidence of exten-sive bioerosion by echinoids, although the lowest 2.5 abovesea level is impossible to examine. The west Xank of the
headland is buried by very coarse-grained sandstone 0.7 mthick, on average. The sandstone is poorly sorted and con-tains subangular grains that include lithic fragments oflimestone or dolomite (roughly 30%), feldspar (30%), andquartz (40%). The composition of this unit changes upwardto register an increase in quartz (roughly 70%) and decreasein the lithic constituents. A stromatolitic unit about 35 cmthick succeeds the sandstone, but is most obvious at the top
Fig. 4 Jurassic–Miocene unconformity exposed in the sea cliV on theeast side of Cala Cigonya. a General perspective showing the contactbetween Jurassic dolomite and overlying Miocene breccia with clastsderived from the dolomite (dotted line traces the unconformity sur-face). b Closer view showing the exposed dolomite surface exten-
sively penetrated by Miocene bivalve borings dominated byGastrochaenolites lapidicus (9-cm-long pocket knife for scale).c Overlapping structures belonging to G. torpedo (left) and the morebulbous G. lapidicus (right). d Remains of similar borings in a brecci-ated clast of dolomite (same scale)
123

Facies
of the former rocky shore. The measurable extent of topo-graphic relief on the exhumed Jurassic-Miocene surface isno less than 15 m.
Ichnofossil assemblages
The overall diversity of borings found at Cala Cigonyaincludes Wve identiWable ichnospecies and three indetermi-nate ichnospecies belonging to four ichnogenera. Theseillustrate structures produced by polychaete annelids(Caulostrepsis isp., Trypanites isp.), demosponges (Ento-bia geometrica Bromley and D’Alessandro 1984, Entobiaisp.), echinoids (Circolites kotoncensis Mikulán 1992), andendolithic bivalves (Gastrochaenolites torpedo Kelly andBromley 1984, G. lapidicus Kelly and Bromley 1984). Allthe bioerosion structures correspond to the boring activityof endolithic organisms. From an ethological point of view,only dwelling structures (domichnia) are present.
East Xank
Gastrochaenolites is the most abundant ichnotaxon of therock-boring assemblage from the east side of the Mioceneheadland. Traces of this structure are irregularly distributed
Fig. 5 Petrographic thin section through laminated limestone showscouplets of light and dark material approximately 0.2 mm thick
Fig. 6 Details from the upper part of the stratigraphic section on theeast side of Cala Cigonya (see Fig. 2 for relationships). a Thinlylaminated limestone in mat form. b Dome-shaped laminations. c Bits
of reworked dolomite breccia bound by laminated limestone. A 9-cm-long pocket knife serves as scale in each photo
123

Facies
over the Jurassic dolomite surface. Most Gastrochaenolitesperforations on the study surface show partial to nearlycomplete erosion of the original boring (Fig. 4b). Someborings are oriented perpendicular to the bedding plane(Gastrochaenolites lapidicus), while others (Fig. 4c) aresub-parallel to it (Gastrochaenolites torpedo and G. lapidicus).
G. lapidicus is abundant and evenly distributed on thesurface. The structures are represented by the lower part ofthe clavate-shaped boring, showing a circular outline with arounded base in plan view. Diameters of the borings arebetween 2 and 18 mm, with an average value of 10 mm.The density of G. lapidicus structures was approximately1,100 individuals/m2.
G. torpedo borings are less common and only appear ashorizontal structures that penetrate the vertical surface ofrock benches. The borings show a mostly wide elongatedoutline in longitudinal section subparallel to the beddingplane (Fig. 4c). The structures vary in dimension: 110 mmin maximum length and 29 mm in maximum width.
The area between bivalve borings is occupied by spongeborings Entobia isp., which have been eroded in most areasto expose the chambers. Caulostrepsis worm borings (withthe typical section in a Wgure-of-eight pattern) were alsoidentiWed, but are rare.
On average, 40% of the dolomite clasts belonging to theMiocene breccia in any given square meter are bored, andin some cases, 20% of their exposed surface is bioeroded.Borings are common on the upper surfaces of the largerclasts and less on lateral surfaces. In general, these bioero-sive structures correspond to the boring activity of endo-lithic bivalves. None of the clasts carry an epilithozoanfauna.
West Xank
The most abundant ichnofossil of the rock-boring assem-blage found over the Jurassic dolomite substrate exhumedon the west side of Cala Cigonya are concave, subcircularpits 25–60 mm in diameter and 10 to 18 mm in depth(Fig. 7a). These are attributed to the dwelling structures ofregular echinoids (Circolites kotoncensis) and they occurover an original slope on the west margin of the outcrop at adensity of approximately 66 structures/m2 (Fig. 7b). TheWrst structures appear near the bottom of the sea cliV (2.5 mabove present sea level) and continue until a height 15 mabove present sea level.
Occasionally, Entobia isp. (Fig. 7c) and small Caulo-strepsis isp. appear inside the concave surface of the
Fig. 7 Details of the Jurassic–Miocene unconformity exposed in thesea cliVs on the west side of Cala Cigonya. a Steeply dipping uncon-formity surface of Jurassic dolomite overlain by 0.70-m-thick basalMiocene sandstone. b Evidence of Miocene bioerosion on the Jurassic
surface shown by dwelling structures of regular echinoids (Circoliteskotoncensis). c Close-up view of an echinoid dwelling structure sec-ondarily colonized by a boring sponge (Entobia isp.). d Close-up viewof Entobia geometrica on the surface between Circolites borings
123

Facies
Circolites dwelling structures. The areas around the echi-noid borings are occupied by Entobia geometrica, whichhas been eroded in most areas to expose chambers andcanals (Fig. 7d). Gastrochaenolites isp. and Trypanites isp.also are present but rare. Diameters of the borings arebetween 5 and 10 mm, with an average value of 7 mm. Try-panites usually is less than 20 mm long with an averagediameter of 2 mm.
Discussion
SigniWcance of the trace fossil assemblages
Based on marine borings on unconformity surfaces withsubstantial paleorelief, a sustained marine transgressionXooded the headlands of the dolomite Jurassic Complex inthe northwest part of Menorca (Pont d’en Gil) during LateMiocene (Tortonian) time. The ichnofossil assemblagefrom the east side of the Miocene headland was preservedunder the inXuence of intertidal to shallow subtidal condi-tions, reXecting a pattern of zonation that corresponds tothe Entobia Ichnofacies Wrst described by Bromley andAsgaard (1993). Sequential generations of borings wereresponsible for the high densities achieved. Examples ofG. torpedo are oriented horizontal as found only on the ver-tical sides of the rocky-shore benches near the base of theformer sea cliV. G. lapidicus and other borings are orientedperpendicular to the unconformity surface where it Xattensout, above.
The erosion surface on the west side of the Miocene head-land records an intertidal to shallow subtidal environment,whereas the endolithic bivalves, sponges and worms, on theeast side, represent a slightly deeper environment. Althoughprimarily subtidal, regular echinoids typically inhabit wetareas of the intertidal zone beneath ledges or in tidal pools, asrepresented on the west side of the outcrop. These organismsare active borers at and below the low-tide line, commonly inhigh-energy settings (Asgaard and Bromley 2008).
This is consistent with the habits of Recent rocky-shoresea urchins. Today, in the Mediterranean Sea, regular echi-noids (Paracentrotus lividus Lamarck 1816) responsiblefor Circolites borings mainly inhabit extremely shallowrockgrounds between zero and ¡2 m (Martinell 1981;Frantzis et al. 1992). The upper limit for this species is con-trolled by desiccation (Boudouresque and Verlaque 2001).Therefore, the presence of the trace fossil Circolites at CalaCigonya records the highest shoreline position. Also, bor-ing bivalves (Lithophaga lithophaga Linné 1758), responsi-ble for G. torpedo, are restricted to shallow marineenvironments and are found today both subtidally and inthe intertidal zone at depths down to ¡10 m (Bromley andAsgaard 1993).
The absence of epilithobiont remains on this Miocenerocky shore is clearly related to the high-energy environ-ment, as shown by the degree of truncation of Gastro-chaenolites borings.
Bioerosion on opposite sides of the Jurassic core rocks atCala Cigonya supports a hypothesis for the colonizationsequence of the Miocene headland. Initial Xooding in the lit-toral zone allowed rapid colonization by pioneer echinoids(producing Circolites) on the sheltered west side of the head-land. On the exposed windward shore (east side), a diVerentbioerosional activity was established immediately below thelittoral zone by a dense population of lithophagous bivalvesand endolithic sponges. The ancient rocky shore ichnoassem-blage of Cala Cigonya is especially interesting because itrecords distinct vertical zonation patterns adopted by organ-isms of variable life styles and ecological tolerances. Thetrace fossil Circolites occupies the highest part of the inter-tidal zone (highest shoreline position) where the trace makerendured the longest subaerial exposure time in a protectedarea, while all other recorded ichnospecies occupy the lowerpart of this zone in a more hydrodynamic area.
Deposition of basal Miocene breccia
The accumulation of Miocene breccia directly above theJurassic–Miocene unconformity on the east side of CalaCigonya is attributed to physical erosion caused by succes-sive storms. Relatively thin partings in the Jurassic dolo-mite layers were susceptible to wave-driven hydraulicpressure that wedged layers apart and detached dolomiteclasts from the parent dolomite body. Little or no furtherboring by bivalves took place on the clasts once they wereseparated from the physically eroded surface. Only 40% ofthe clasts with a surface exposure show traces of bivalveborings. As a test of the hypothesis that those borings weremade prior to exfoliation from the bedrock, individualclasts with borings on the exposed surface were broken freefrom the deposit in order to examine the hidden surface onthe underside. Invariably, no trace of borings was found onthe underside. Arguably, wave activity was suYcient toturn the clasts from time to time, but individual clasts weretoo small or unstable to support continued bioerosion. It isalso possible that clasts were moved out of the zone whereborings could be made. The Upper Miocene breccia fromthe northwest side of Menorca is related to a former rockyshoreline and distinct from the Upper Miocene brecciaassociated with ramp deposits on the south side of theisland (Obrador et al. 1992).
Deposition of breccia stringers
Ongoing physical erosion of the parent Jurassic dolomiteby storm activity after the development of microbial life at
123

Facies
Cala Cigonya during Messinian time is supported by theoccurrence of breccia stringers incorporated into the lami-nated limestone deposits well above the level of the basalMiocene breccia. Individual clasts are not in contact withone another and are bound in place by the laminar growth.The bound clasts are very angular, and some show evidenceof small borings (Fig. 6c). These borings were likely tohave been made earlier in Tortonian time prior to erosion ofthe clasts from the parent dolomite body. The heightenedsalinity of the Mediterranean Sea during Messinian timesshould have prohibited normal marine life. If true, thismeans that the bored Tortonian surface of the Jurassic dolo-mite continued to be physically abraded by wave shockduring Messinian time. The energy needed to transporteroded dolomite clasts to a place where they were depositedas stringers demonstrates that the enclosing microbial matswere suYciently robust to withstand a physical beating inclose proximity to an active rocky shore.
Deposition of basal Miocene sandstone
Several lines of evidence demonstrate that the basal Mio-cene sandstone covering the erosion surface on the westXank of the former headland had a terrestrial source andwas little altered by wave activity after deposition in amarine setting. Foremost are the facts that the sandstone ispoorly sorted and dominated by subangular grains. Themajor compositional elements are quartz and plagioclase,which have no source in the dolomite Jurassic Complex ofMenorca. Lithic fragments that are carbonate or dolomiticin nature, however, may have been eroded from these rocksin a more inland zone. No grains in the basal sandstone arelarger than 2 mm in diameter (very coarse sand), whichconstrains the input of erosive energy. Although dolomitebreccia and laminated limestone characterize deposits onthe opposite east Xank of the Miocene headland, it shouldbe noted that pockets of sandstone also occur there. Thus,stream erosion due to runoV from heavy rainfall was likelyto have caused the introduction of clastic sediments to
small embayments on the northern shores of Menorca.Widespread Triassic rocks in the central elevations of Men-orca (Fig. 1b) are the probable source for sediments rich insilica and plagioclase, in particular the Buntsandstein of theLower Triassic (Rosell-Sanuy and Llompart-Diaz 2002).
Comparisons with other rocky-shore microbial deposits
Stromatolitic facies are well represented on former rockyshores in Archean and Proterozoic rocks, but show a steadydecline in Paleozoic, Mesozoic, and Cenozoic rocks (John-son 2006). This trend may be due to the accelerated evolu-tion of highly competitive rocky-shore biotas in Mesozoicand Cenozoic times. Johnson and Baarli (1999), Fig. 3)refer to stromatolites as the principal component of anArchaic rocky-shore biota that managed to maintain a slimpresence in the post-Proterozoic world.
A representative sample of references on rocky-shorestromatolites from the survey by Johnson (2006) is pro-vided in Table 1. Emphasis is given to examples from thepost-Proterozoic record. Undoubted stromatolites builtmound-like accumulations directly on hard substrates likegranite during Proterozoic time (Goodwin 1960, p. 49) inthe GunXint Formation of Canada. Thick stromatoliticcrusts of a similar nature can be traced to the Lower Ordov-ician of the Czech Republic, where encrustation on a chertsubstrate is documented (Mergl 1983, plate 4). Thickmicrobial build-ups of Devonian age constructed on graniteand gneiss inliers are known from the western OrkneyIslands (Fannin 1969), but depositional conditions areunclear and may have entailed a lacustrine setting.
Thin stromatolitic crusts are reported on the karst surfaceof Jurassic limestone in Spain (García-Hernández et al.1988). Stratiform and columnar stromatolitic crusts of earli-est Paleocene age are described as covering a bored lime-stone hardground from the Upper Cretaceous of Greece(Pomoni-Papaioannou and Solakius 1991). Mineralizationsuggests unusual conditions, at least in the case of thePaleocene stromatolites from Greece. The only other
Table 1 Some microbial build-ups associated with rocky shore settings (from Johnson 2006)
Geologic age Country Substrate Author(s)
Miocene Egypt PC basement Burchette (1988)
Paleocene Greece Limestone Pomoni-Papaioannou and Solakiu (1991)
Late Jurassic Spain Limestone García-Hernández et al. (1988)
Devonian Scotland Granite/gneiss Fannin (1969)
Ordovician Czech Republic Chert Mergl (1983)
Cambrian Spain Andesite Vennin et al. (2003)
Proterozoic Canada Granite Goodwin (1960)
Archean Australia Greenstone Allwood et al. (2006)
Archean S. Africa Basalt Byerly et al. (1986)
123

Facies
Miocene example comes from Egypt on the Gulf of Suez,where stromatolitic sheets and mounds formed directly onPrecambrian basement rocks associated with coastal faultblocks (Burchette 1988). By any comparison, the sheetedand dome-shaped build-ups of microbialites from the Mio-cene of northwest Menorca (Figs. 3, 6) are far more sub-stantial in thickness than any other examples associatedwith rocky-shore settings. The special circumstances underwhich deposition occurred in close proximity to a rockyshoreline at Cala Cigonya does not exclude rough-waterconditions, but was abetted by elevated levels of salinity inthe Mediterranean Sea during the Messinian crisis.
Conclusions
The north coast of Menorca in the Balearic Islands of Spainfeatures many places where steep rocky shores are formedby carbonate strata that receive heavy surf from Tramon-tana winds during the winter months. In Late Miocene(Tortonian) times, headlands shaped from the dolomiteJurassic Complex along this shore were selectively colo-nized and bioeroded by assemblages of boring organismscapable of withstanding wave activity in varying intensi-ties. During the Messinian crisis at the end of the Miocene,endolithic invertebrates on rocky shores were replaced byextensive microbial build-ups tolerant of increased salinity.The following conclusions relate to the development of ich-nofacies on Miocene rocky shores and the accumulation ofsedimentary facies that buried hard substrates at Cala Cigo-nya in northwestern Menorca.
• Cala Cigonya was occupied by a narrow, northwest-directed headland composed of Jurassic dolomite with aminimum paleorelief of 15 m in Tortonian and Messin-ian times. The east Xank of the headland developed rockyshores on a gentle slope from 10 to 14° with incised stepsor benches, while the opposite Xank maintained steeperslopes inclined to the southwest between 30 and 40°.
• Ichnofacies from the rocky shores on the east Xank of theMiocene headland are dominated by Gastrochaenolitestorpedo and G. lapidicus (boring bivalve traces) withminor representations by Entobia isp. (boring spongetraces) and Caulostrepsis isp. (boring polychaete traces).In contrast, the ichnofacies from the west Xank are domi-nated by Circolites kotencensis (boring echinoid traces)with lesser contributions from Entobia geometrica,Caulostrepsis isp, Gastrochaenolites isp., and Trypanitesisp.
• The lithofacies on the erosion surface from the east Xankof the headland is an Upper Miocene (Tortonian) brecciaderived from the underlying Jurassic dolomite. Clasts arerectangular and plate-like in shape with some borings
typically present only on one face. In contrast, thelithofacies that buried the erosion surface on the headland’swest Xank consists of very coarse sandstone with compo-nents of silica, plagioclase, and other lithic elements.This facies is terrestrial in origin and probably related tothe inland erosion of Triassic sandstone.
• Both the east and west Xanks of the Miocene headlandfeature laminated deposits overlying basal Tortonianfacies. Only on the east Xank are clasts of Jurassic brec-cia incorporated into these deposits. The laminationsshow Wne couplets of light and dark sediments consistentwith a microbial origin. Stromatolitic facies are not com-monly associated with a rocky-shore setting, but thesedeveloped during the Messinian crisis under conditionsof elevated salinity in the Mediterranean Sea.
• DiVerences in ichno- and sedimentary facies on opposingsides of the same Miocene headland can be explained interms of wave energy stimulated by seasonal Tramon-tana winds oV the Iberian mainland. The east side of theheadland was exposed to higher levels of wave energythat led to exfoliation of the parent Jurassic substrate andproduction of a basal Miocene breccia. The west sidewas the leeward Xank, protected from strong waveshock. Echinoids responsible for bioerosion of Circuliteskotencensis traces thrived under these more shelteredconditions. The same conditions supported the accumu-lation of a localized sandy facies for the basal Miocenelittle inXuenced by the physical erosion of the Jurassicsurface.
Future research on comparative ichno- and sedimentaryfacies from the Upper Miocene on north Menorcan shoresis expected to provide evidence for similar relationshipsaround other former headlands associated with the wide-spread dolomite Jurassic Complex on the island.
Acknowledgments Fieldwork on Menorca was conducted in March2005 and March 2010. Johnson was supported by a travel grants fromthe Class of 1945 Faculty World Fellowship during sabbatical leavesfrom Williams College. Santos received Wnancial support for her par-ticipation in 2010 from the Ministry of Science and Technology ofSpain in the form of a Juan de la Cierva contract (Refª JCI-2008-2431).Financial support was also provided by the Junta de Andalucía (Span-ish government) to the Research Group RNM316 (Tectonics and Pal-aeontology) and by the Spanish DGI Project CGL 2007-60507/BTE.We thank an anonymous referee for helpful comments and referencesthat improved the quality of the manuscript.
References
Allwood AC, Walter MR, Kamber BS, Marshall CP, Burch IW (2006)Stromatolite reef from the early Archaean era of Australia. Nature441:714–718
Asgaard U, Bromley RG (2008) Echinometrid sea urchins, theirtrophic styles and corresponding bioerosion. In: Wisshak M,Tapanila L (eds) Current development s in bioerosion. Erlangen
123

Facies
earth conference series. Springer, Berlin Heidelberg New York,pp 279–303
Asprion U, Westphal H, Nieman M, Pomar L (2009) Extrapolation ofdepositional geometries of the Menorcan Miocene carbonateramp with ground-penetrating radar. Facies 55:37–46
Bird CF, Schwartz ML (eds) (1985) The world’s coastline. VanNostrand Reinhold, New York, p 1,071
Boudouresque CF, Verlaque M (2001) Ecology of Paracentrotus livi-dus. Dev Aquac Fish Sci 32:177–216
Brandano M, Vannucci G, Pomar L, Obrador A (2005) Rhodolithassemblages from the lower Tortonian carbonate ramp of Menor-ca (Spain): environmental and paleoclimatic implications. Palae-ogeogr Palaeoclimatol Palaeoecol 226:307–323
Bromley RG, Asgaard U (1993) Endolithic community replacement ona Pliocene rocky coast. Ichnos 2:93–116
Bromley RG, D’Alessandro A (1984) The ichnogenus Entobia fromthe Miocene, Pliocene, and Pleistocene of southern Italy. Riv ItalPaleontol Stratig 90:227–296
Burchette TP (1988) Tectonic control on carbonate platform facies dis-tribution and sequence development: Miocene, Gulf of Suez. SedGeol 59:179–204
Byerly GR, Lowe DR, Walsh MM (1986) Stromatolites from the3,300–3,500-Myr Swaziland supergroup, Barberton MountainLand, South Africa. Nature 319:489–491
Caracuel JE, Soria JM, Yébenes A (2004) Early Pliocene transgressivecoastal lags (Bajo Segura Basin, Spain): a marker of the Xoodingafter the Messinian salinity crisis. Sediment Geol 169:121–128
Doblas M, Oyarzun R (1989) Neogene extensional collapse in thewestern Mediterranean (Betic-Rif Alpine orogenic belt): implica-tions for the genesis of the Gibraltar arc arc and magmatic activ-ity. Geology 17:430–433
Duggen S, Hoernle K, van den Bogaard P, Rüpke L, Morgan JP (2003)Deep roots of the Messinian salinity crisis. Nature 422:602–606
Fannin NGT (1969) Stromatolites from the middle old red sandstoneof western Orkney. Geol Mag 106:77–88
Frantzis A, Gremare A, Vetion G (1992) Growth rates and RNA: DNAratios in Paracentrotus lividus (Echinodermata: Echinoidea) fedon benthic macrophytes. J Exp Mar Biol Ecol 156:125–138
García-Hernández M, Más JR, Molina JM, Ruiz-Ortiz PA, Vera JA(1988) Episodio de karstiWcaión en litorales insulares del JurásicoSuperior (Fm. Ammonítico Rosso, Subbético Externo, Provinciade Córdoba). III Coloquio de Estratigrafía y Paleogeografía delJurásico de España., Logroño, pp 32–35
Gomila BA (1999a) Mallorca coastline aeroguide. Editorial Planeta,SA, Barcelona, p 230
Gomila BA (1999b) Menorca coastline aeroguide. Editorial Planeta,SA, Barcelona, p 105
Goodwin AM (1960) GunXint iron formation of the WhiteWsh Lakearea, district of Thunder Bay. Ont Dep Mines 69(7):41–67
Gueguen E, Doglioni C, Fernandez M (1998) On the post-25 Ma geo-dynamic evolution of the western Mediterranean. Tectonophysics298:259–269
Johnson ME (1988a) Why are ancient rocky shores so uncommon?J Geol 96:469–480
Johnson ME (1988b) Hunting for ancient rocky shores. J Geol Educ36:147–154
Johnson ME (2006) Uniformitarianism as a guide to rocky-shore eco-systems in the geological record. Can J Earth Sci 43:1119–1147
Johnson ME, Baarli BG (1999) DiversiWcation of rocky-shore biota’sthrough geologic time. Geobios 32:257–273
Kelly P, Bromley RG (1984) Ichnological nomenclature of clavateborings. Palaeontology 2(4):793–807
Krijgsman W, Hilgen FJ, RaY I, Sierro FJ, Wilson DS (1999) Chro-nology, causes and progression of the Messinian salinity crisis.Nature 400:652–655
Lamarck JBPA (1816) Histoire naturelle des animaux sans vertèbres,présentant les caractères, généraux et particuliers de ces animaux,leurs distribution, leurs classes, leurs familles, leurs genres et lacitation synonymique des principales espèces qui s’y rapportent.Paris, vol 3, 586 pp
Linné C (1758) Systema naturae, vol 1, 10th edn. Laurentii Salvii,Holmiae, 824 pp
Loget N, Van Den Driessche J (2006) On the origin of the strait ofGibraltar. Sed Geol 188–189:341–358
Martinell J (1981) Actividad erosiva the Paracentrotus lividus (Lmk.)(Echinodermata, Echinoidea) en el litoral gerundense. Oecologiaaquatica 5:219–225
Mateu-Vicens G, Hallock P, Brandano M (2008) A depositional modeland paleoecological reconstruction of the lower Tortonian distallysteepened ramp of Menorca (Balearic Islands, Spain). Palaios23:465–481
Mergl M (1983) Rocky-bottom fauna of Ordovician age in Bohemia(Arenigian; Prague Basin, Barrandian area). Vestnik Ustrednihoustavu geologickeho 58:333–339
Mikulán R (1992) Early cretaceous borings from Stramberk (Czecho-slovakia). Casopis pro mineralogii a geologii 37(4):297–312
Obrador A, Pomar L, Taberner C (1992) Late Miocene breccia of Men-orca (Balearic Islands): a basis for the interpretation of a neogeneramp deposit. Sed Geol 79:203–223
Platt JP, Vissers RLM (1989) Extensional collapse of thickened conti-nental lithosphere: a working hypothesis for the Alboran Sea andGibraltar arc. Geology 17:540–543
Pomar L (2001) Ecological control of sedimentary accommodation:evolution from a carbonate ramp to rimmed shelf, Upper Mio-cene, Balearic Islands. Palaeogeogr Palaeoclimatol Palaeoecol175:249–272
Pomar L, Ward WC (1994) Response of a Late Miocene Mediterra-nean reef platform to high-frequency eustasy. Geology 22:131–134
Pomar L, Obrador A, Westhal H (2002) Sub-wavebase cross-beddedgrainstones on a distally steepened carbonate ramp, Upper Mio-cene, Menorca, Spain. Sedimentology 49:139–169
Pomar L, Brandano M, Westphal H (2004) Environmental factorsinXuencing skeletal grain sediment associations: a critical reviewof Miocene examples from the western Mediterranean. Sedimen-tology 51:627–651
Pomoni-Papaioannou F, Solakiu N (1991) Phosphatic hardgrounds andstromatolites from the limestone/shale boundary section atProssilion (Maastrichtian-Paleocene) in Parnassus-Ghiona zone,Central Greece. Palaeogeogr Palaeoclimatol Palaeoecol 86:243–254
Riding R (2000) Microbial carbonates: the geological record of calci-Wed bacterial-algal mats and bioWlms. Sedimentology 47:179–214
Rosell-Sanuy J, Llompart-Diaz C (2002) El Naixement d’una IllaMenorca: Guía de Geología Práctica. Dacs, Indústria GráWca, SA,Montcada i Reixac, p 279
Rosenbaum G, Lister GS, Duboz C (2002) Reconstruction of the tec-tonic evolution of the western Mediterranean since the Oligocene.J Virtual Explor 8:107–130
Vennin E, Álvaro JJ, Moreno-Eiris E, Perejón A (2003) Early Cam-brian coelobiontic communities in tectonically unstable crevicesdeveloped in neoproterozoic andesites, Ossa-Morena, southernSpain. Lethaia 36:53–65
123