molecular systematics of new world gopher, bull, and pinesnakes (pituophis: colubridae), a...
TRANSCRIPT

stassWmamhmjsmtseddarrdotgtesCassdpdpf
8
Molecular Phylogenetics and EvolutionVol. 14, No. 1, January, pp. 35–50, 2000Article ID mpev.1999.0698, available online at http://www.idealibrary.com on
Molecular Systematics of New World Gopher, Bull, and Pinesnakes(Pituophis: Colubridae), a Transcontinental Species Complex
Javier A. Rodrıguez-Robles*,1 and Jose M. De Jesus-Escobar†
*Museum of Vertebrate Zoology and Department of Integrative Biology, University of California, Berkeley, California 94720-3160;and †Department of Molecular and Cell Biology, Division of Biochemistry and Molecular Biology, University of California,
Berkeley, California 94720-3204
Received October 14, 1998; revised June 17, 1999
rc
Da
ra1WiaEHW‘d1ottr(MJMa1ntcc11
Pituophis melanoleucus (gopher, bull, and pine-nakes) is among the most widely distributed poly-ypic species complexes in North America, with mostuthors recognizing from a single transcontinentalpecies (the melanoleucus complex, composed of 15ubspecies) to four (monotypic and polytypic) species.e used mitochondrial gene sequences from the twoiddle American species, P. deppei and P. lineaticollis,
nd from 13 subspecies from most of the range of theelanoleucus complex to test various phylogeneticypotheses for Pituophis. Maximum parsimony andaximum likelihood methods identified the same ma-
or clades within Pituophis and indicated that twoegments of the melanoleucus complex, the lodingi–elanoleucus–mugitus eastern pinesnake clade and
he affinis–annectens–bimaris–catenifer–deserticola–ayi–ruthveni–vertebralis clade from central and west-rn United States and northern Mexico, representivergent, allopatric lineages with no known intergra-ation zone. We recognize each of these two groupingss a different species. Our data also indicate that someuthveni are more closely related to sayi than to otheruthveni. Nonetheless, ruthveni is an allopatric taxoniagnosable from its closest relatives by a combinationf morphometric characters, and because it is likelyhat at least some of these traits are independent andenetically inherited, we interpret this as evidencehat ruthveni has attained the status of independentvolutionary lineage, despite the fact that it retainstrong genetic affinities with sayi. The endemic Bajaalifornian gopher snakes (bimaris and vertebralis)re considered by some taxonomists as a differentpecies, P. vertebralis, but we discovered that theseerpents belong to two different clades and hence weo not agree with the recognition of P. vertebralis asresently defined. In summary, we believe that threeistinct species are included in the melanoleucus com-lex, Pituophis melanoleucus (sensu stricto), P. cateni-er, and P. ruthveni, and that their recognition better
1 To whom correspondence should be addressed. Fax: (510) 643-
238. E-mail: [email protected].35
epresents the evolutionary diversity within this spe-ies complex. r 2000 Academic Press
Key Words: biogeography; Colubridae; mitochondrialNA; phylogenetics; Pituophis; snakes; species bound-ries; subspecies.
INTRODUCTION
The question as to the number of species includedin this genus [Pituophis] is a difficult one to decide.(Cope, 1900, p. 866)Boundaries between species and subspecies have
eceived considerable attention from systematists (Ballnd Avise, 1992; Cracraft, 1983; McKitrick and Zink,988; Patton and Smith, 1994; Smith et al., 1997;uster and Thorpe, 1992) and have often generated
nstructive and sometimes intense discussions (Avisend Wollenberg, 1997; Collins, 1992; Dowling, 1993;dwards, 1954; Frost and Hillis 1990; Frost et al., 1992;ighton, 1998; Inger, 1961; Wake and Schneider, 1998;iens, 1982; Wilson and Brown, 1953). During the
‘molecular systematics revolution’’ of the last threeecades (Avise, 1994; Hillis et al., 1996; Soltis et al.,998), genetic data have sometimes contradicted previ-us ideas about species integrity or taxonomic distinc-ions that were based largely on morphological descrip-ions and thus led authors to reject named subspecies,ecognize new ones, or elevate some to the species levele.g., Alcobendas et al., 1996; Cicero, 1996; Garcıa-
oreno and Fjeldså, 1999; Good and Wake, 1992;ohnson, 1995; Miththapala et al., 1996; Shaffer andcKnight, 1996; Wuster and Thorpe, 1994; Zamudio
nd Greene, 1997; Zamudio et al., 1997; Zink et al.,997). Determining which populations warrant taxo-omic recognition contributes to our understanding ofhe evolution of biodiversity and has significant, practi-al implications, e.g., for sociology (Keita, 1993) andonservation policies (Avise, 1989; Geist, 1992; Greene,994; O’Brien and Mayr, 1991; Sites and Crandall,997; Walker et al., 1998; Zhu et al., 1998).
Evolutionary relationships of transcontinental taxa1055-7903/00 $35.00Copyright r 2000 by Academic PressAll rights of reproduction in any form reserved.

hWppSacEatAso2td(SwsbcBSD
ictg
al1ppwoBcEPs
ttrmt
Clhs
36 RODRIGUEZ-ROBLES AND DE JESUS-ESCOBAR
ave rarely been studied in detail (e.g., Bernatchez andilson, 1998). Taxa with such broad geographic ranges
rovide an excellent opportunity to investigate theractical issues in naming population assemblages.nakes of the colubrid genus Pituophis Holbrook, 1842re among the most widely distributed polytypic spe-ies complexes in North America. Commonly known innglish as gopher, bull, and pinesnakes and in Spanishs ‘‘alicantes,’’ ‘‘cincuates,’’ ‘‘coralillos,’’ and ‘‘mazacua-es,’’ these serpents range from the Pacific to thetlantic coast of the United States (US) and fromouthwestern Canada to central Guatemala and the tipf the Baja California peninsula in Mexico (Figs. 1 and). Species of Pituophis are nonvenomous constrictors;hey occur in a wide variety of habitats, includingeserts, sandhills, prairies, shrublands, and forestsDegenhardt et al., 1996; Palmer and Braswell, 1995;weet and Parker, 1990); they attain body sizes in theild of up to 2.7 m; they exhibit notable variation in
nout morphology (Knight, 1986) and reproductiveiology (Fitch, 1985; Palmer and Braswell, 1995; Rei-hling, 1990; Zappalorti et al., 1983); and they may beatesian mimics of rattlesnakes (Kardong, 1980;anchez-Herrera et al., 1981; but see Sweet, 1985).
FIG. 1. Approximate distribution of the subspecies of the Pituophionant and Collins, 1991; Reichling, 1995; Stebbins, 1985; Sweet a
ocalities of the specimens included in this study. The black barypothesized Pleistocene midpeninsular seaway (after Upton and Mcale.
espite the fact that Pituophis has figured prominently i
n the anatomical, ecological, ethological, and herpeto-ulturist literature, the genus has had a contentiousaxonomic history, which we briefly review as back-round for our systematic study of these snakes.The monophyly of Pituophis is strongly supported by
t least two unique morphological characters, an en-arged epiglottal keel and a laryngeal septum (Cope,891; White, 1884; Young et al., 1995), which togetherroduce the unusual defensive hissing made by Pituo-his (Martin and Huey, 1971; Young et al., 1995), asell as by phylogenetic analyses of mtDNA sequencesf the genus and its closest relatives (i.e., Arizona,ogertophis, Cemophora, Elaphe, Lampropeltis, Rhino-
heilus, and Senticolis; Rodrıguez-Robles and De Jesus-scobar, 1999). In these analyses the monophyly ofituophis was supported by very high ($96%) boot-trap values in maximum parsimony trees.Beyond the issue of the monophyly of the genus,
here has been little agreement among different au-hors about the number of species and subspeciesecognized within Pituophis. The most common argu-ents presented can be summarized in five main
heses.(1) The most taxonomically conservative view regard-
elanoleucus complex in Canada, the United States, and Mexico (afterParker, 1990; Tennant, 1984). Numbers indicate the approximate
oss central Baja California indicates the inferred location of thehy, 1997). The four islands indicated with arrows are not drawn to
s mndacrurp
ng the number of species in the Pituophis melanoleu-

cpMleBamaCt(
KsiPseaclahsc
be
cl
itm
gcsDswoadM
pscntwspfoatnspi
N d in
37SYSTEMATICS AND BIOGEOGRAPHY OF Pituophis
us species complex (hereafter, the melanoleucus com-lex) in southwestern Canada, the US, and northernexico recognizes a single, polytypic species, P. melano-
eucus, composed of 15 subspecies, including four taxandemic to islands off the west coast of California andaja California (Conant, 1956; Dowling, 1958; Smithnd Kennedy, 1951; Fig. 1). This taxonomic arrange-ent is followed in three widely used field guides to
mphibians and reptiles from the US (Behler, 1995;onant and Collins, 1991; Stebbins, 1985), as well as in
he Catalogue of American Amphibians and ReptilesSweet and Parker, 1990).
(2) Some authors (e.g., Fugler, 1955; Holman, 1996;lauber, 1947; Wright and Wright, 1957) have at least
uggested that the melanoleucus complex is a compos-te taxonomic unit consisting of two distinct species,ituophis melanoleucus (sensu stricto, including theubspecies lodingi, melanoleucus, and mugitus) in theastern US and P. catenifer (including the subspeciesffinis, annectens, bimaris, catenifer, coronalis, deserti-ola, fuliginatus, insulanus, pumilus, sayi, and vertebra-is) in the central and western US, northern Mexico,nd offshore islands. The Louisiana pinesnake, ruthveni,as been assigned to either P. melanoleucus (sensutricto; Stull, 1940; Wright and Wright, 1957) or P.atenifer (Fugler, 1955).(3) The Cape gopher snake, vertebralis, has sometimes
een claimed to be a distinct species (Pituophis vertebralis;.g., Cope, 1900; Grismer, 1994; Stull, 1940; Sweet, 1984).(4) Based on a phenetic analysis of 14 morphometric
haracters and on its geographic distribution, Reich-
FIG. 2. Approximate distribution of Pituophis deppei and P. lineaumbers indicate the approximate localities of the specimens include
ing (1995; see also Collins, 1991) argued for recogniz- m
ng ruthveni as a distinct species, Pituophis ruthveni,herefore effectively implying the recognition of P.elanoleucus (sensu stricto) and P. catenifer as well.(5) The taxonomic status of the middle American
opher snakes (Fig. 2), Pituophis deppei and P. lineati-ollis, each with two named subspecies, has been fairlytable since Duellman’s (1960) review. Nevertheless,ixon et al. (1962) questioned the validity of the two
ubspecies of P. deppei (P. d. deppei and P. d. jani),hereas Conant (1965) suggested that intergradationr introgression occurred between P. deppei and P. m.ffinis in northern Mexico, perhaps implying that P.eppei was not a valid species, a suggestion with whichorafka (1977) agreed.The taxonomic status of the various forms of Pituo-
his thus has provided systematists with an unre-olved controversy, particularly in the melanoleucusomplex, within which different authors have recog-ized from a single transcontinental polytypic specieso four (monotypic and polytypic) species. However,ith the exception of Cope’s (1900) and Stull’s (1940)
tudies, all these taxonomic arrangements were pro-osed after examining only a subset of the describedorms of this species complex, or were suggested basedn the geographic distribution and presumed diagnos-bility of the named subspecies. Our study tests theaxonomic hypotheses presented above with phyloge-etic trees inferred from mitochondrial DNA (mtDNA)equences from 16 of the 19 described forms of Pituo-his, the greatest number of described forms includedn any previous taxonomic study of the genus. Our
llis in Mexico and Guatemala (after Duellman, 1960; Grismer, 1994).this study.
tico
ain objective is to assess evolutionary relationships

aor1t3o
T
ao(
bssaDgcMsr1CIoo
c(smDcftnutt(eZec1cDPmµ73u3mmpbvEgquai
a
38 RODRIGUEZ-ROBLES AND DE JESUS-ESCOBAR
mong the currently recognized species and subspeciesf Pituophis, assuming that our gene genealogy accu-ately reflects these relationships (see Brower et al.,996; Moore, 1995, 1997). We also compare our findingso previous phylogenetic hypotheses for Pituophis (Fig.) and discuss some of the biogeographical implicationsf our results.
MATERIALS AND METHODS
axon Sampling, DNA Isolation, and Sequencing
We obtained tissue samples from one to four individu-ls of Arizona elegans (glossy snake), Bogertophis sub-cularis (Trans Pecos ratsnake), Lampropeltis getula
FIG. 3. Evolutionary relationships proposed for Pituophis: (a)fter Stull (1940); (b) after Klauber (1947).
common kingsnake), Pituophis d. deppei (Mexican (
ullsnake), P. deppei jani (northern Mexican bull-nake), P. lineaticollis gibsoni (Guatemalan gophernake), and within the melanoleucus complex, fromffinis (Sonoran desert gopher snake), annectens (Saniego gopher snake), bimaris (northern Baja Californiaopher snake), catenifer (Pacific gopher snake), deserti-ola (Great Basin gopher snake), fuliginatus (Sanartın Island gopher snake), lodingi (black pine-
nake), pumilus (Santa Cruz Island gopher snake),uthveni, sayi (bullsnake), and vertebralis (Table 1; Fig.). We could not secure samples of coronalis (Southoronado Island gopher snake) or insulanus (Cedros
sland gopher snake) but we believe that the omissionf these two taxa does not alter the major findings ofur study.We extracted total genomic DNA from ventral scale
lips preserved in 95% ethanol or from tissue samplesblood, liver, muscle) stored frozen at 274°C using theodium dodecyl sulphate–proteinase K/phenol/RNAaseethod (Sambrook et al., 1989). Using total cellularNA as template, we amplified (with the polymerase
hain reaction, PCR [Saiki et al., 1986, 1988]) and usedor phylogenetic analyses an 893-bp fragment of mtDNAhat encompassed a 697-bp portion of the 38 end of theicotinamide adenine dinucleotide dehydrogenase sub-nit 4 (Ndh4, or ‘‘ND4’’ gene) and a 196-bp section ofhree transfer ribonucleic acid (tRNA) genes (tRNAHis,RNASer, tRNALeu), using primers labeled ND4 and LeuArevalo et al., 1994). ND4 is a reliable tracer ofvolutionary history (Russo, 1997; Russo et al., 1996;ardoya and Meyer, 1996) and a relatively fast-volving gene useful for resolving relationships amonglosely related taxa (Cracraft and Helm-Bychowski,991). PCR was carried out in a programmable thermalycler in 100-µl reactions consisting of 2 µl of templateNA (50 ng/µl), 2.5 µl of primers (40 µM), 10 µl of 103CR reaction buffer (Stratagene), 2 µl of MgCl2 (25M), 2 µl of deoxynucleoside triphosphates (10 mM), 4
l of Thermus aquaticus DNA polymerase (5 U/µl), and7.5 µl of H2O. DNA was denatured initially at 94°C formin; then 33 cycles of amplification were carried outnder the following conditions: 94°C denaturation for0 s, 55°C annealing for 30 s, and 72°C extension for 1in, followed by a final 5-min extension at 72°C. Tenicroliters of the resulting PCR product were electro-
horesed on a 1% agarose gel and stained with ethidiumromide to verify product band size. For each indi-idual, we cloned its PCR product into a phosphatasedcoRV pBluescript II SK 6 phagemid vector (Strata-ene) using Escherichia coli as the vector and se-uenced both DNA strands in an automated sequencersing the dideoxy chain-termination method (Sanger etl., 1977). The sequence of Elaphe vulpina (fox snake)ncluded in this study was provided by R. Lawson
California Academy of Sciences, San Francisco).
P
wps(r1sdNsp
taKlwab
mSFicagcttEtsr1loii
pbhooegsttmcna
tgFptrZt((r
iUcnCsdplttt1mhtps
desrpbttfodtoqpptcs
S
n
39SYSTEMATICS AND BIOGEOGRAPHY OF Pituophis
hylogenetic Analyses
Sequences from the light and heavy DNA strandsere input into the Sequence Navigator (version 1.0.1)rogram and aligned to each other and to the referenceequence of the colubrid snake Dinodon semicarinatusKumazawa et al., 1998). This initial alignment wasefined with the MacDNASIS Pro software (version.0). Pairwise comparisons of observed proportionalequence divergence ( p-distance), corrected sequenceivergence (with the Tamura–Nei model; Tamura andei, 1993), and number of transitions and transver-
ions by codon position were obtained using the com-uter program PAUP* 4.0b2a (Swofford, 1999).To estimate the phylogenetic information content of
he mtDNA character matrix, we used the g test (Hillisnd Huelsenbeck, 1992; Huelsenbeck, 1991; but seeallersjo et al., 1992) to assess the skewness of the tree
ength distribution of 100,000 trees randomly generatedith PAUP*. Probability of phylogenetic structure wasssessed using the values provided by Hillis and Huelsen-eck (1992).We used two methods of phylogenetic reconstruction:aximum parsimony (MP; Camin and Sokal, 1965;wofford et al., 1996) and maximum likelihood (ML;elsenstein, 1981; Huelsenbeck and Crandall, 1997), as
mplemented by PAUP*. For MP analyses, we used twoharacter weighting schemes: equal-weighting, in whichll nucleotide substitutions were weighted equally re-ardless of type or codon position, and differentialodon position weighting, in which we down-weightedhird position transitions (see below). Sites with inser-ion or deletion events were removed from the analyses.ach base position was treated as an unordered charac-
er with four alternative states. Ancestral charactertates were determined via outgroup comparison (Far-is, 1982; Maddison et al., 1984; Watrous and Wheeler,981). We used Arizona elegans, Bogertophis subocu-aris, Elaphe vulpina, and Lampropeltis getula asutgroups because previous molecular systematic stud-es (Rodrıguez-Robles and De Jesus-Escobar, 1999)dentified these taxa as close relatives of Pituophis.
Because the number of terminal taxa was too large toermit evaluating all trees or employing the branch-and-ound algorithm (Hendy and Penny, 1982), we usedeuristic search strategies for each tree-building meth-dology. We used 100 repeated randomized input ordersf taxa for all MP analyses to minimize the effects ofntry sequence on the topology of the resulting clado-ram(s). MP analyses were conducted without theteepest descent option and with accelerated characterransformation (ACCTRAN) optimization, tree bisec-ion–reconnection (TBR) branch swapping, save allinimal trees (MULPARS), and zero-length branches
ollapsed to yield polytomies settings in place. We usedonparametric bootstrapping (100 pseudoreplicates, 10
ddition-sequence replicates for MP, 50% majority rule) so assess the stability of internal branches in clado-rams (Berry and Gascuel, 1996; Felsenstein, 1985;elsenstein and Kishino, 1993; Sanderson, 1995). Non-arametric bootstrap values generally are a conserva-ive measure of the probability that a recovered groupepresents a true clade (Hillis and Bull, 1993; Li, 1997;harkikh and Li, 1992). Alternate topologies wereested for significance at the 95% level using the TempletonLarson, 1994; Templeton, 1983) and Kishino–HasegawaKishino and Hasegawa, 1989) tests for MP and ML trees,espectively, as implemented in PAUP*.
For ML analyses we randomly selected as the start-ng tree one of the trees found during the MP searches.sing empirical nucleotide frequencies and five rate
ategories, we fixed the probabilities of the six possibleucleotide transformations (A & C, A & G, A & T,& G, C& T, G& T), the proportion of invariable
ites u, and the a ‘‘shape’’ parameter of the gammaistribution of rate heterogeneity across nucleotideositions (Yang, 1996) to the empirical values calcu-ated from the starting tree in a search for a better MLree (a tree with a higher log-likelihood value), underhe general time-reversible model of nucleotide substi-ution (Gu et al., 1995; Swofford et al., 1996; Yang,994). In other words, we used the most parameter-richodel available to search for ML trees. When a tree of
igher likelihood was found, we reoptimized and fixedhe parameters for a subsequent ML search. We re-eated this procedure until the same tree was found inuccessive iterations (Swofford et al., 1996).Because ND4 is a protein-coding gene, we plotted p-
istance (y) versus corrected (with the Tamura–Nei model)stimates of proportional sequence divergence (x) for first,econd, and third codon positions. This was done sepa-ately for transitions and transversions to test for theossibility that some types of nucleotide substitutions haveecome saturated. Points that fall along the y 5 x line havehe same observed and estimated numbers of changes andhus have not been subjected to multiple hits. Points thatall below the y 5 x line indicate that multiple hits haveccurred; saturation is reached when observed sequenceivergence does not continue to increase, despite the facthat corrected estimates do. Conventional statistical testsf the relationship between estimated and observed se-uence divergence are not appropriate because of noninde-endence of the data points, due to the inclusion of eachoint in more than one pairwise comparison. Accordingly,he plots were used as heuristic devices to help identifylasses of changes occurring at different rates, whichhould be weighted differently in phylogenetic analyses.
RESULTS
equence Variation
There were 348 variable and 225 potentially phyloge-etically informative characters (sites with at least two
hared differences among all taxa) in the 893-bp mtDNA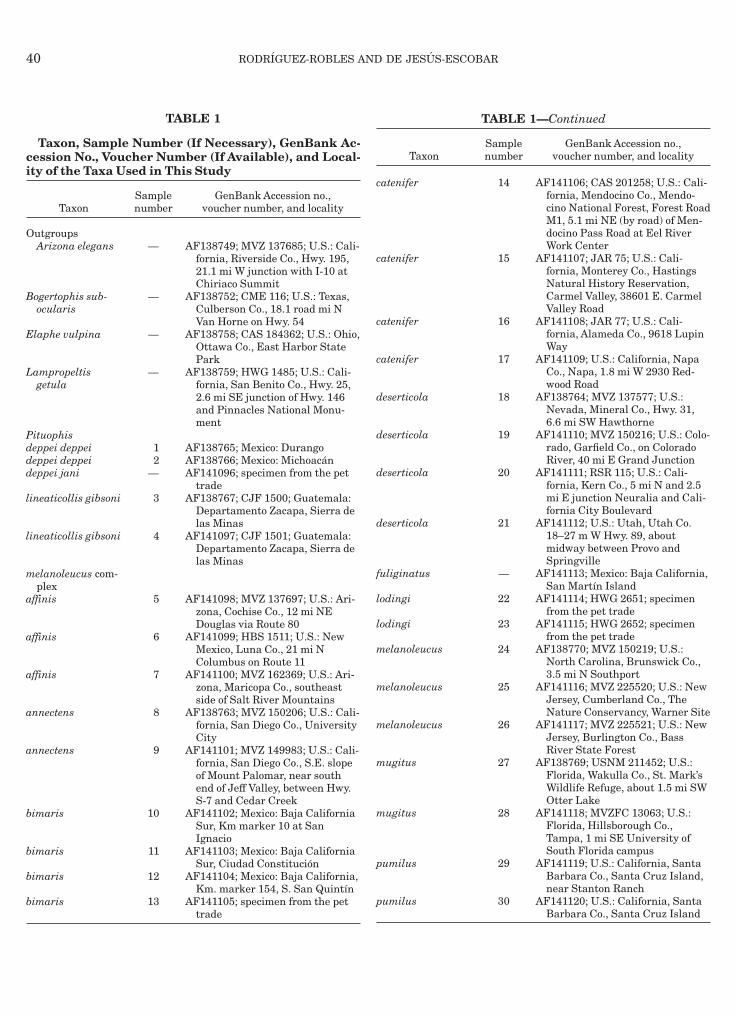
ci
O
B
E
L
Pddd
l
l
m
a
a
a
a
a
b
b
b
b
c
c
c
c
d
d
d
d
f
l
l
m
m
m
m
m
p
p
40 RODRIGUEZ-ROBLES AND DE JESUS-ESCOBAR
TABLE 1
Taxon, Sample Number (If Necessary), GenBank Ac-ession No., Voucher Number (If Available), and Local-ty of the Taxa Used in This Study
TaxonSamplenumber
GenBank Accession no.,voucher number, and locality
utgroupsArizona elegans — AF138749; MVZ 137685; U.S.: Cali-
fornia, Riverside Co., Hwy. 195,21.1 mi W junction with I-10 atChiriaco Summit
ogertophis sub-ocularis
— AF138752; CME 116; U.S.: Texas,Culberson Co., 18.1 road mi NVan Horne on Hwy. 54
laphe vulpina — AF138758; CAS 184362; U.S.: Ohio,Ottawa Co., East Harbor StatePark
ampropeltisgetula
— AF138759; HWG 1485; U.S.: Cali-fornia, San Benito Co., Hwy. 25,2.6 mi SE junction of Hwy. 146and Pinnacles National Monu-ment
ituophiseppei deppei 1 AF138765; Mexico: Durangoeppei deppei 2 AF138766; Mexico: Michoacaneppei jani — AF141096; specimen from the pet
tradeineaticollis gibsoni 3 AF138767; CJF 1500; Guatemala:
Departamento Zacapa, Sierra delas Minas
ineaticollis gibsoni 4 AF141097; CJF 1501; Guatemala:Departamento Zacapa, Sierra delas Minas
elanoleucus com-plex
ffinis 5 AF141098; MVZ 137697; U.S.: Ari-zona, Cochise Co., 12 mi NEDouglas via Route 80
ffinis 6 AF141099; HBS 1511; U.S.: NewMexico, Luna Co., 21 mi NColumbus on Route 11
ffinis 7 AF141100; MVZ 162369; U.S.: Ari-zona, Maricopa Co., southeastside of Salt River Mountains
nnectens 8 AF138763; MVZ 150206; U.S.: Cali-fornia, San Diego Co., UniversityCity
nnectens 9 AF141101; MVZ 149983; U.S.: Cali-fornia, San Diego Co., S.E. slopeof Mount Palomar, near southend of Jeff Valley, between Hwy.S-7 and Cedar Creek
imaris 10 AF141102; Mexico: Baja CaliforniaSur, Km marker 10 at SanIgnacio
imaris 11 AF141103; Mexico: Baja CaliforniaSur, Ciudad Constitucion
imaris 12 AF141104; Mexico: Baja California,Km. marker 154, S. San Quintın
imaris 13 AF141105; specimen from the pettrade
TABLE 1—Continued
TaxonSamplenumber
GenBank Accession no.,voucher number, and locality
atenifer 14 AF141106; CAS 201258; U.S.: Cali-fornia, Mendocino Co., Mendo-cino National Forest, Forest RoadM1, 5.1 mi NE (by road) of Men-docino Pass Road at Eel RiverWork Center
atenifer 15 AF141107; JAR 75; U.S.: Cali-fornia, Monterey Co., HastingsNatural History Reservation,Carmel Valley, 38601 E. CarmelValley Road
atenifer 16 AF141108; JAR 77; U.S.: Cali-fornia, Alameda Co., 9618 LupinWay
atenifer 17 AF141109; U.S.: California, NapaCo., Napa, 1.8 mi W 2930 Red-wood Road
eserticola 18 AF138764; MVZ 137577; U.S.:Nevada, Mineral Co., Hwy. 31,6.6 mi SW Hawthorne
eserticola 19 AF141110; MVZ 150216; U.S.: Colo-rado, Garfield Co., on ColoradoRiver, 40 mi E Grand Junction
eserticola 20 AF141111; RSR 115; U.S.: Cali-fornia, Kern Co., 5 mi N and 2.5mi E junction Neuralia and Cali-fornia City Boulevard
eserticola 21 AF141112; U.S.: Utah, Utah Co.18–27 m W Hwy. 89, aboutmidway between Provo andSpringville
uliginatus — AF141113; Mexico: Baja California,San Martın Island
odingi 22 AF141114; HWG 2651; specimenfrom the pet trade
odingi 23 AF141115; HWG 2652; specimenfrom the pet trade
elanoleucus 24 AF138770; MVZ 150219; U.S.:North Carolina, Brunswick Co.,3.5 mi N Southport
elanoleucus 25 AF141116; MVZ 225520; U.S.: NewJersey, Cumberland Co., TheNature Conservancy, Warner Site
elanoleucus 26 AF141117; MVZ 225521; U.S.: NewJersey, Burlington Co., BassRiver State Forest
ugitus 27 AF138769; USNM 211452; U.S.:Florida, Wakulla Co., St. Mark’sWildlife Refuge, about 1.5 mi SWOtter Lake
ugitus 28 AF141118; MVZFC 13063; U.S.:Florida, Hillsborough Co.,Tampa, 1 mi SE University ofSouth Florida campus
umilus 29 AF141119; U.S.: California, SantaBarbara Co., Santa Cruz Island,near Stanton Ranch
umilus 30 AF141120; U.S.: California, SantaBarbara Co., Santa Cruz Island

dfitPctpm1
devpprfrtpot
at1tb
P
etgw(ssroacltaPMlss
odslpo(CMfse
pasteitsnes
mmt
r
r
r
s
s
s
s
v
v
v
AZcnCG
41SYSTEMATICS AND BIOGEOGRAPHY OF Pituophis
ata matrix. Of the informative characters, 29 were atrst codon positions, 13 at second positions, 144 athird positions, and 39 at noncoding positions. Withinituophis there were 21, 9, 121, and 33 informativeharacters at first, second, third, and noncoding posi-ions, respectively. Significant phylogenetic signal wasresent in the data set (g1 5 20.6222, P ,,, 0.01;ean 6 SD tree length 5 1817.3 6 31.3, range 1625–
905); thus, inferring cladograms was justified.Scatter plots of observed versus estimated sequence
ivergences indicated that these relationships are lin-ar for first and second position transitions and trans-ersions and third position transversions (Fig. 4). Thirdosition transitions deviated greatly from a linearattern, suggesting that these mutations are satu-ated. To estimate the transition-to-transversion biasor third position transitions, we fitted a least-squaresegression line, forced through the origin, to the part ofhe curve of third position transitions versus thirdosition transversions that is roughly linear. The slopef the regression line, 0.648, is an estimate of the
TABLE 1—Continued
TaxonSamplenumber
GenBank Accession no.,voucher number, and locality
uthveni 31 AF138771; U.S.: Louisiana, Bien-ville Parish, 2 km E Kepler CreekLake Bridge
uthveni 32 AF141121; U.S.: Louisiana, Bien-ville Parish, 7 km E Kepler CreekLake Bridge
uthveni 33 AF138772; U.S.: Louisiana, Bien-ville Parish, 2 km S junction ofLA 154 and 507
ayi 34 AF141122; MVZ 150218; U.S.:Texas, Jeff Davis Co., on U.S.Route 90, 43.8 mi SE junction ofI-10 and U.S. Route 90
ayi 35 AF141123; U.S.: Colorado, Jef-ferson Co., SE junction of W. CoalMine Road and Wadsworth Bou-levard
ayi 36 AF141124; MVZ 226247; U.S.: Mis-souri, Saint Louis Co.
ayi 37 AF141125; U.S.: Oklahoma, Cleve-land Co.
ertebralis 38 AF141126; Mexico: Baja CaliforniaSur, Rancho Buena Vista
ertebralis 39 AF141127; JAR 78; Mexico: BajaCalifornia Sur, Cape region
ertebralis 40 AF141128; specimen from the pettrade
Note. Museum and collector abbreviations are: CAS, Californiacademy of Sciences, San Francisco; MVZ, Museum of Vertebrateoology, University of California, Berkeley; MVZFC, MVZ frozenollection; USNM, National Museum of Natural History, Smithso-ian Institution, Washington, DC; CJF, Carl J. Franklin; CME,urtis M. Eckerman; HBS, H. Bradley Shaffer; HWG, Harry W.reene; JAR, Javier A. Rodrıguez; RSR, Randall S. Reiserer.
ransition-to-transversion ratio (Lara et al., 1996; Moore t
nd DeFilippis, 1997). Therefore, we down-weightedhird codon transitional changes by a factor of 6 using a:1:0.17 codon position weighting (first, second, andhird codon position, respectively) to correct for theiased substitution rates at this position.
hylogenetic Relationships
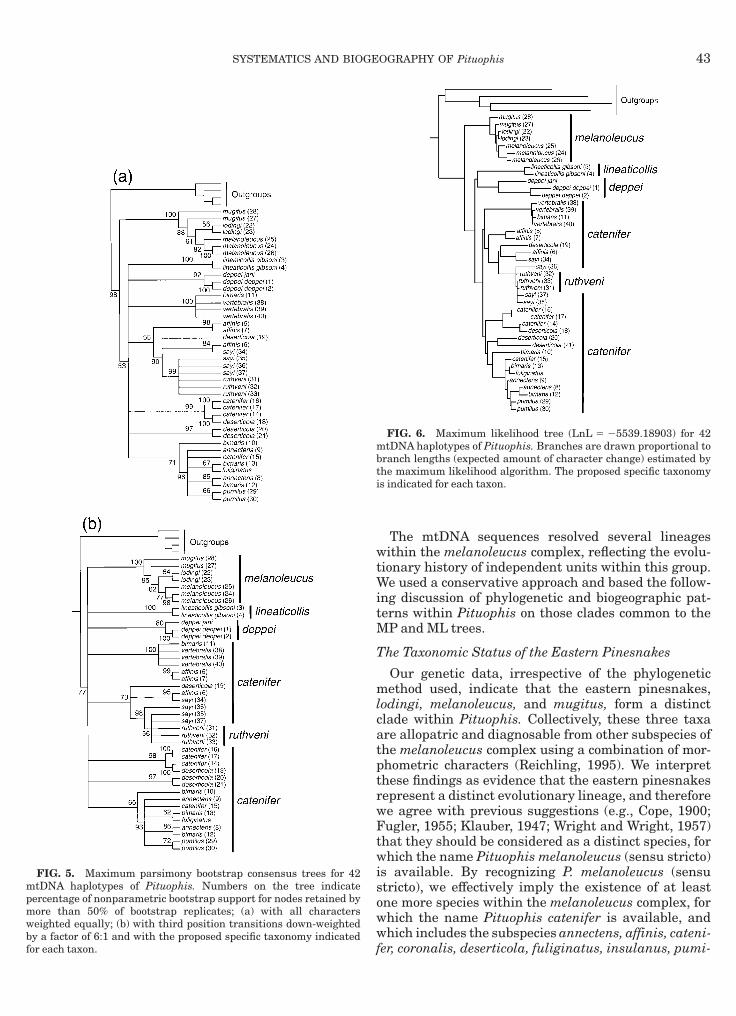
The two phylogenetic methods that we used recov-red the same major nodes (Figs. 5 and 6), regardless ofhe weighting scheme used, which suggests that theroupings represent true clades. Most of these nodesere supported by relatively high bootstrap values
$80%) in the two MP consensus trees but the relation-hips among the clades were almost completely unre-olved. Although the single ML tree obtained (Fig. 6)esolved nearly all the relationships among the cladesf Pituophis, some of the internal branches of this treere short, indicating that they are supported by fewharacters, which may explain why these nodes col-apse during the random resampling of characters thatakes place during bootstrapping. The short internodeslso suggest that the divergence of the major lineages ofituophis occurred rapidly and near simultaneously.P and ML trees constrained to include only monophy-
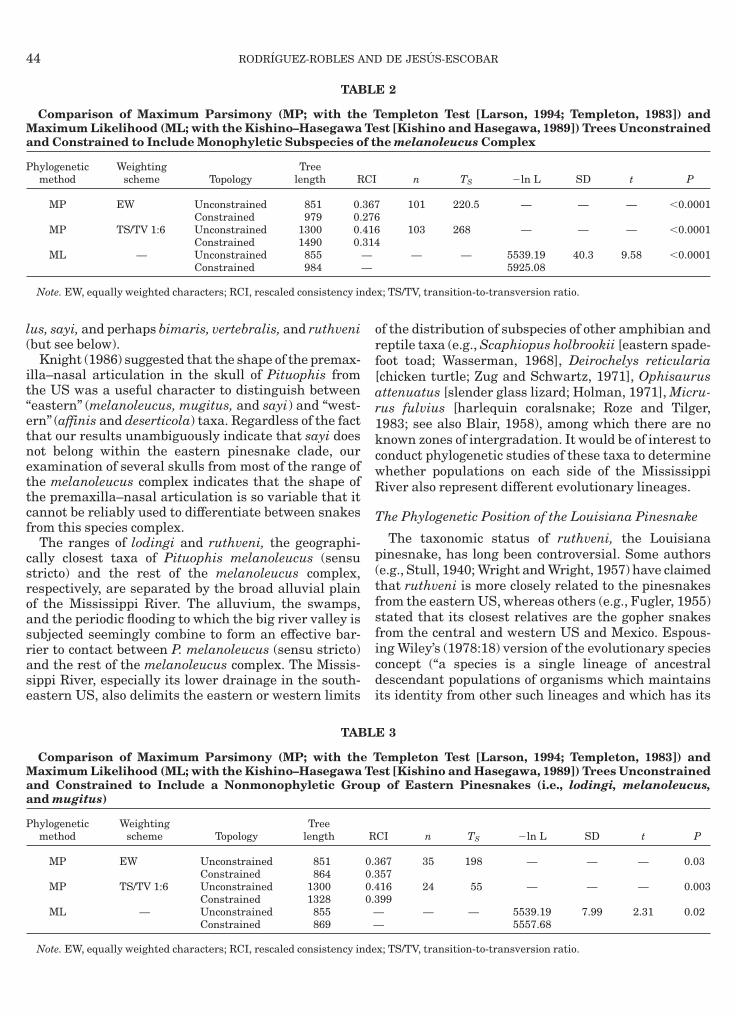
etic subspecies of the melanoleucus complex wereignificantly poorer estimates of phylogenetic relation-hips than the unconstrained trees (Table 2).MP and ML methods identified monophyletic clades
f the two species of middle American gopher snakes (P.eppei and P. lineaticollis). Nonetheless, the lack ofpecimens of affinis and sayi from Mexico and ourimited sampling of the two subspecies of P. deppeirevented us from evaluating the disputed monophylyf the latter relative to the melanoleucus complexConant, 1965; Dixon et al., 1962; Morafka, 1977).learly, further studies that include representatives ofexican affinis and sayi, of P. d. deppei and P. d. jani
rom additional localities, and of P. l. lineaticollis fromouthern Mexico are needed to better understand thevolutionary history of middle American Pituophis.The MP and ML trees also showed that the eastern
inesnakes, lodingi, melanoleucus, and mugitus, formmonophyletic clade, and these trees are significantly
horter or have a higher likelihood ratio, respectively,han those constrained to include nonmonophyleticastern pinesnakes (Table 3). The MP bootstrap treenferred with equally weighted characters and the MLree identified the eastern pinesnake lineage as theister taxon to all other Pituophis, but this topology wasot significantly different from that on which theastern pinesnakes were constrained to not be theister taxon to all other Pituophis (Table 4).All the topologies recovered by the MP and MLethods identified separate, monophyletic clades of theelanoleucus complex (Figs. 5 and 6). Within these,
here is clear geographic structuring. As mentioned,
he eastern pinesnakes formed a distinct clade, and so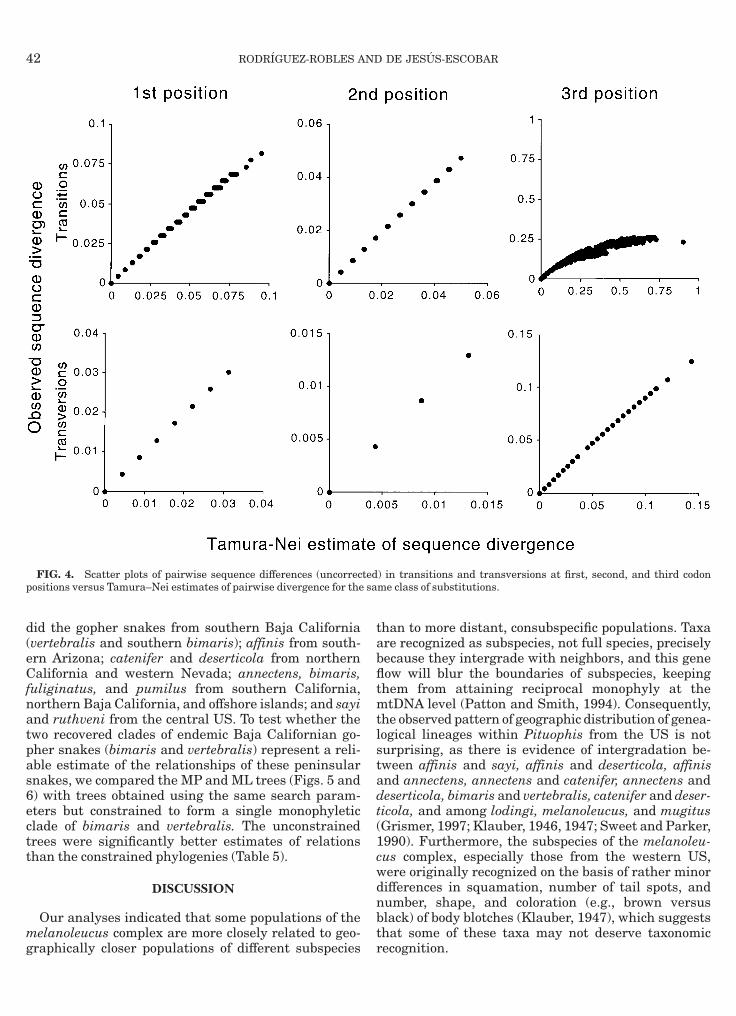
d(eCfnatpas6ectt
mg
tabfltmtlstadt(1cwdnbt
p sa
42 RODRIGUEZ-ROBLES AND DE JESUS-ESCOBAR
id the gopher snakes from southern Baja Californiavertebralis and southern bimaris); affinis from south-rn Arizona; catenifer and deserticola from northernalifornia and western Nevada; annectens, bimaris,
uliginatus, and pumilus from southern California,orthern Baja California, and offshore islands; and sayind ruthveni from the central US. To test whether thewo recovered clades of endemic Baja Californian go-her snakes (bimaris and vertebralis) represent a reli-ble estimate of the relationships of these peninsularnakes, we compared the MP and ML trees (Figs. 5 and) with trees obtained using the same search param-ters but constrained to form a single monophyleticlade of bimaris and vertebralis. The unconstrainedrees were significantly better estimates of relationshan the constrained phylogenies (Table 5).
DISCUSSION
Our analyses indicated that some populations of theelanoleucus complex are more closely related to geo-
FIG. 4. Scatter plots of pairwise sequence differences (uncorrecositions versus Tamura–Nei estimates of pairwise divergence for the
raphically closer populations of different subspecies r
han to more distant, consubspecific populations. Taxare recognized as subspecies, not full species, preciselyecause they intergrade with neighbors, and this geneow will blur the boundaries of subspecies, keepinghem from attaining reciprocal monophyly at thetDNA level (Patton and Smith, 1994). Consequently,
he observed pattern of geographic distribution of genea-ogical lineages within Pituophis from the US is noturprising, as there is evidence of intergradation be-ween affinis and sayi, affinis and deserticola, affinisnd annectens, annectens and catenifer, annectens andeserticola, bimaris and vertebralis, catenifer and deser-icola, and among lodingi, melanoleucus, and mugitusGrismer, 1997; Klauber, 1946, 1947; Sweet and Parker,990). Furthermore, the subspecies of the melanoleu-us complex, especially those from the western US,ere originally recognized on the basis of rather minorifferences in squamation, number of tail spots, andumber, shape, and coloration (e.g., brown versuslack) of body blotches (Klauber, 1947), which suggestshat some of these taxa may not deserve taxonomic
) in transitions and transversions at first, second, and third codonme class of substitutions.
ted
ecognition.
wtWitM
T
mlcatptrwFtwisowwf
mpmwbfor each taxon.
mbti
43SYSTEMATICS AND BIOGEOGRAPHY OF Pituophis
The mtDNA sequences resolved several lineagesithin the melanoleucus complex, reflecting the evolu-
ionary history of independent units within this group.e used a conservative approach and based the follow-
ng discussion of phylogenetic and biogeographic pat-erns within Pituophis on those clades common to theP and ML trees.
he Taxonomic Status of the Eastern Pinesnakes
Our genetic data, irrespective of the phylogeneticethod used, indicate that the eastern pinesnakes,
odingi, melanoleucus, and mugitus, form a distinctlade within Pituophis. Collectively, these three taxare allopatric and diagnosable from other subspecies ofhe melanoleucus complex using a combination of mor-hometric characters (Reichling, 1995). We interprethese findings as evidence that the eastern pinesnakesepresent a distinct evolutionary lineage, and thereforee agree with previous suggestions (e.g., Cope, 1900;ugler, 1955; Klauber, 1947; Wright and Wright, 1957)
hat they should be considered as a distinct species, forhich the name Pituophis melanoleucus (sensu stricto)
s available. By recognizing P. melanoleucus (sensutricto), we effectively imply the existence of at leastne more species within the melanoleucus complex, forhich the name Pituophis catenifer is available, andhich includes the subspecies annectens, affinis, cateni-
FIG. 6. Maximum likelihood tree (LnL 5 25539.18903) for 42tDNA haplotypes of Pituophis. Branches are drawn proportional to
ranch lengths (expected amount of character change) estimated byhe maximum likelihood algorithm. The proposed specific taxonomys indicated for each taxon.
FIG. 5. Maximum parsimony bootstrap consensus trees for 42tDNA haplotypes of Pituophis. Numbers on the tree indicate
ercentage of nonparametric bootstrap support for nodes retained byore than 50% of bootstrap replicates; (a) with all characterseighted equally; (b) with third position transitions down-weightedy a factor of 6:1 and with the proposed specific taxonomy indicated
er, coronalis, deserticola, fuliginatus, insulanus, pumi-
l(
it‘etnettcf
csroasrase
orf[ar1kcwR
T
p(tfsficdi
Ma
P
de
Maa
P
44 RODRIGUEZ-ROBLES AND DE JESUS-ESCOBAR
us, sayi, and perhaps bimaris, vertebralis, and ruthvenibut see below).
Knight (1986) suggested that the shape of the premax-lla–nasal articulation in the skull of Pituophis fromhe US was a useful character to distinguish between‘eastern’’ (melanoleucus, mugitus, and sayi) and ‘‘west-rn’’ (affinis and deserticola) taxa. Regardless of the facthat our results unambiguously indicate that sayi doesot belong within the eastern pinesnake clade, ourxamination of several skulls from most of the range ofhe melanoleucus complex indicates that the shape ofhe premaxilla–nasal articulation is so variable that itannot be reliably used to differentiate between snakesrom this species complex.
The ranges of lodingi and ruthveni, the geographi-ally closest taxa of Pituophis melanoleucus (sensutricto) and the rest of the melanoleucus complex,espectively, are separated by the broad alluvial plainf the Mississippi River. The alluvium, the swamps,nd the periodic flooding to which the big river valley isubjected seemingly combine to form an effective bar-ier to contact between P. melanoleucus (sensu stricto)nd the rest of the melanoleucus complex. The Missis-ippi River, especially its lower drainage in the south-astern US, also delimits the eastern or western limits
TAB
Comparison of Maximum Parsimony (MP; with theaximum Likelihood (ML; with the Kishino–Hasegawa
nd Constrained to Include Monophyletic Subspecies o
hylogeneticmethod
Weightingscheme Topology
Treelength R
MP EW Unconstrained 851 0.Constrained 979 0.
MP TS/TV 1:6 Unconstrained 1300 0.Constrained 1490 0.
ML — Unconstrained 855Constrained 984
Note. EW, equally weighted characters; RCI, rescaled consistency in
TAB
Comparison of Maximum Parsimony (MP; with theaximum Likelihood (ML; with the Kishino–Hasegawa
nd Constrained to Include a Nonmonophyletic Grond mugitus)
hylogeneticmethod
Weightingscheme Topology
Treelength
MP EW Unconstrained 851Constrained 864
MP TS/TV 1:6 Unconstrained 1300Constrained 1328
ML — Unconstrained 855Constrained 869
Note. EW, equally weighted characters; RCI, rescaled consistency inde
f the distribution of subspecies of other amphibian andeptile taxa (e.g., Scaphiopus holbrookii [eastern spade-oot toad; Wasserman, 1968], Deirochelys reticulariachicken turtle; Zug and Schwartz, 1971], Ophisaurusttenuatus [slender glass lizard; Holman, 1971], Micru-us fulvius [harlequin coralsnake; Roze and Tilger,983; see also Blair, 1958), among which there are nonown zones of intergradation. It would be of interest toonduct phylogenetic studies of these taxa to determinehether populations on each side of the Mississippiiver also represent different evolutionary lineages.
he Phylogenetic Position of the Louisiana Pinesnake
The taxonomic status of ruthveni, the Louisianainesnake, has long been controversial. Some authorse.g., Stull, 1940; Wright and Wright, 1957) have claimedhat ruthveni is more closely related to the pinesnakesrom the eastern US, whereas others (e.g., Fugler, 1955)tated that its closest relatives are the gopher snakesrom the central and western US and Mexico. Espous-ng Wiley’s (1978:18) version of the evolutionary speciesoncept (‘‘a species is a single lineage of ancestralescendant populations of organisms which maintainsts identity from other such lineages and which has its
E 2
empleton Test [Larson, 1994; Templeton, 1983]) andst [Kishino and Hasegawa, 1989]) Trees Unconstrainedhe melanoleucus Complex
n TS 2ln L SD t P
101 220.5 — — — ,0.0001
103 268 — — — ,0.0001
— — 5539.19 40.3 9.58 ,0.00015925.08
x; TS/TV, transition-to-transversion ratio.
E 3
empleton Test [Larson, 1994; Templeton, 1983]) andst [Kishino and Hasegawa, 1989]) Trees Unconstrained
of Eastern Pinesnakes (i.e., lodingi, melanoleucus,
CI n TS 2ln L SD t P
67 35 198 — — — 0.035716 24 55 — — — 0.00399
— — 5539.19 7.99 2.31 0.025557.68
L
TTef t
CI
367276416314——
L
TTeup
R
0.30.30.40.3
——
x; TS/TV, transition-to-transversion ratio.
oR
attewssr1uh1asgnawtrt
ga
qwsteoraMsdwasehcnrc1
Mam
P
Maa
P
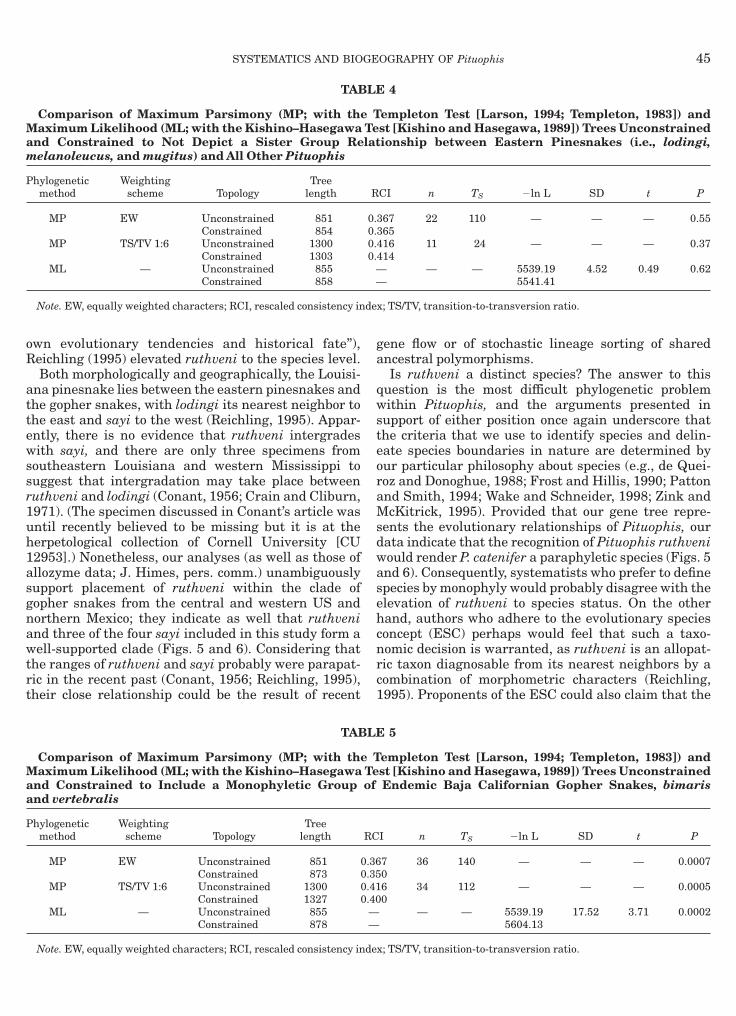
45SYSTEMATICS AND BIOGEOGRAPHY OF Pituophis
wn evolutionary tendencies and historical fate’’),eichling (1995) elevated ruthveni to the species level.Both morphologically and geographically, the Louisi-
na pinesnake lies between the eastern pinesnakes andhe gopher snakes, with lodingi its nearest neighbor tohe east and sayi to the west (Reichling, 1995). Appar-ntly, there is no evidence that ruthveni intergradesith sayi, and there are only three specimens from
outheastern Louisiana and western Mississippi touggest that intergradation may take place betweenuthveni and lodingi (Conant, 1956; Crain and Cliburn,971). (The specimen discussed in Conant’s article wasntil recently believed to be missing but it is at theerpetological collection of Cornell University [CU2953].) Nonetheless, our analyses (as well as those ofllozyme data; J. Himes, pers. comm.) unambiguouslyupport placement of ruthveni within the clade ofopher snakes from the central and western US andorthern Mexico; they indicate as well that ruthvenind three of the four sayi included in this study form aell-supported clade (Figs. 5 and 6). Considering that
he ranges of ruthveni and sayi probably were parapat-ic in the recent past (Conant, 1956; Reichling, 1995),heir close relationship could be the result of recent
TAB
Comparison of Maximum Parsimony (MP; with theaximum Likelihood (ML; with the Kishino–Hasegawa
nd Constrained to Not Depict a Sister Group Reelanoleucus, and mugitus) and All Other Pituophis
hylogeneticmethod
Weightingscheme Topology
Treelength
MP EW Unconstrained 851Constrained 854
MP TS/TV 1:6 Unconstrained 1300Constrained 1303
ML — Unconstrained 855Constrained 858
Note. EW, equally weighted characters; RCI, rescaled consistency in
TAB
Comparison of Maximum Parsimony (MP; with theaximum Likelihood (ML; with the Kishino–Hasegawa
nd Constrained to Include a Monophyletic Groupnd vertebralis
hylogeneticmethod
Weightingscheme Topology
Treelength
MP EW Unconstrained 851Constrained 873
MP TS/TV 1:6 Unconstrained 1300Constrained 1327
ML — Unconstrained 855Constrained 878
Note. EW, equally weighted characters; RCI, rescaled consistency inde
ene flow or of stochastic lineage sorting of sharedncestral polymorphisms.Is ruthveni a distinct species? The answer to this
uestion is the most difficult phylogenetic problemithin Pituophis, and the arguments presented in
upport of either position once again underscore thathe criteria that we use to identify species and delin-ate species boundaries in nature are determined byur particular philosophy about species (e.g., de Quei-oz and Donoghue, 1988; Frost and Hillis, 1990; Pattonnd Smith, 1994; Wake and Schneider, 1998; Zink andcKitrick, 1995). Provided that our gene tree repre-
ents the evolutionary relationships of Pituophis, ourata indicate that the recognition of Pituophis ruthveniould render P. catenifer a paraphyletic species (Figs. 5nd 6). Consequently, systematists who prefer to definepecies by monophyly would probably disagree with thelevation of ruthveni to species status. On the otherand, authors who adhere to the evolutionary speciesoncept (ESC) perhaps would feel that such a taxo-omic decision is warranted, as ruthveni is an allopat-ic taxon diagnosable from its nearest neighbors by aombination of morphometric characters (Reichling,995). Proponents of the ESC could also claim that the
4
empleton Test [Larson, 1994; Templeton, 1983]) andst [Kishino and Hasegawa, 1989]) Trees Unconstrainedionship between Eastern Pinesnakes (i.e., lodingi,
CI n TS 2ln L SD t P
367 22 110 — — — 0.55365416 11 24 — — — 0.37414
— — 5539.19 4.52 0.49 0.625541.41
x; TS/TV, transition-to-transversion ratio.
5
empleton Test [Larson, 1994; Templeton, 1983]) andst [Kishino and Hasegawa, 1989]) Trees UnconstrainedEndemic Baja Californian Gopher Snakes, bimaris
I n TS 2ln L SD t P
7 36 140 — — — 0.000706 34 112 — — — 0.00050
— — 5539.19 17.52 3.71 0.00025604.13
LE
TTelat
R
0.0.0.0.
——
de
LE
TTeof
RC
0.360.350.410.40
——
x; TS/TV, transition-to-transversion ratio.

pscscstuL
aossaaai(dcr
oovgrwmaLmccbatiRCssttGgoqgip
osbd
sdust
T
trrgnop(bsbtpca1aBevMeTP
sms(gtstacoaotTgfmcdfG
46 RODRIGUEZ-ROBLES AND DE JESUS-ESCOBAR
attern of genetic relationship between ruthveni andayi revealed by our analyses simply reflects the re-ency of their divergence and lack of time for lineageorting to result in reciprocal monophyly. However,ritics might still contend that Reichling’s taxonomictudy of ruthveni did not include all the subspecies ofhe melanoleucus complex and accordingly cannot besed to argue in favor of or against recognizing theouisiana pinesnake as a distinct species.Two arguments are relevant when considering the
forementioned views regarding the taxonomic statusf ruthveni. First, the theoretical basis for the empha-is on naming monophyletic species is not clear, as anypecies has evolved from a preexisting species, and thisncestral species necessarily ceased to be monophyletict the moment of speciation (Baum, 1992; de Queiroznd Donoghue, 1988, 1990; Graybeal, 1995). Hence, its impossible to avoid recognizing paraphyletic species.The practice of assigning new species status to allaughters of a cladogenetic event is simply a taxonomiconvention and does not necessarily indicate biologicaleality.)Second, although Reichling’s (1995) taxonomic study
f ruthveni did not include the western-most subspeciesf the melanoleucus complex (i.e., annectens, bimaris,ertebralis, and the four island taxa), it included theeographically and phenotypically closer relatives ofuthveni (lodingi, melanoleucus, mugitus, and sayi), asell as affinis, deserticola, and catenifer. Hence, weaintain that Reichling’s findings should be taken into
ccount when judging the taxonomic status of theouisiana pinesnake and that not doing so is tanta-ount to ignoring available evidence. Therefore, be-
ause it is likely that at least some of the morphometricharacters shown by Reichling to collectively and unam-iguously distinguish ruthveni from its closer relativesre independent and genetically inherited, we believehat the Louisiana pinesnake has attained the status ofndependent evolutionary lineage. We then agree witheichling’s (1995) suggestion (see also Collins, 1991;onant, 1956) of recognizing ruthveni as a distinctpecies, Pituophis ruthveni. One reason why our mtDNAequence data do not yet reflect the independent evolu-ionary trajectory of P. ruthveni may be that popula-ions of this snake constitute an example of whatraybeal (1995) called ‘‘ferespecies,’’ interbreedingroups of organisms that do not yet possess exclusivityf descent. Nevertheless, it is possible that DNA se-uences of other genetic markers (e.g., from the nuclearenome or even other mitochondrial genes) wouldndicate a higher degree of genetic cohesiveness amongopulations of ruthveni.Despite the above arguments, the taxonomic status
f the Louisiana pinesnake is likely to remain controver-ial, not so much due to lack of relevant data, but ratherecause of different interpretations of available evi-
ence, and although we recognize these snakes at full mpecies rank, we understand why other workers mayisagree with our position. If Louisiana pinesnakes arendergoing ‘‘the speciation process,’’ their debatabletatus illustrates the continuing difficulty in makingaxonomic assignments in such cases.
he Taxonomic Status and Biogeography of theEndemic Baja Californian Gopher Snakes
The results of our phylogenetic analyses regardinghe endemic gopher snakes from Baja California, bima-is and vertebralis, are in disagreement with the cur-ent taxonomy of these serpents. Traditionally, theopher snakes from central and southern Baja Califor-ia were assigned to bimaris, with snakes from theutskirts of the town of La Paz south to the tip of theeninsula (the Cape region) assigned to vertebralisFig. 1). Recently, Grismer (1994, 1997) synonymizedimaris with vertebralis and elevated the latter topecies level. We discovered that the southernmostimaris included in our study was more closely relatedo vertebralis (sensu stricto) than to more northernopulations of bimaris, which nested within a separatelade composed of two island subspecies (fuliginatusnd pumilus), and of annectens and catenifer (sample5) from southern California (Figs. 5 and 6). Addition-lly, the two distinct clades that include the endemicaja Californian gopher snakes are not sister taxa toach other, and when we constrained all bimaris andertebralis to form a monophyletic group, the resultingP and ML trees were significantly poorer estimates of
volutionary relationships within Pituophis (Table 2).hese findings led us to reject the recognition ofituophis vertebralis as defined by Grismer.The notable genetic break between northern and
outhern Baja Californian populations of Pituophisirrors the phylogeographic pattern displayed by Uta
tansburiana (side-blotched lizard) on the peninsulaUpton and Murphy, 1997). Upton and Murphy sug-ested that the disruption of mtDNAhaplotype distribu-ion between side-blotched lizards from northern andouthern Baja California was consistent with the exis-ence of a temporary midpeninsular seaway (Durhamnd Allison, 1960; Fig. 1). Presumably, this barrieraused the cessation of gene flow between populationsn the northern and southern portions of the peninsula,llowing them to evolve independently. Based on vari-us assumptions, Upton and Murphy hypothesizedhat the seaway existed one million years ago (Mya).he smallest uncorrected percentage sequence diver-ence between specimens of bimaris and vertebralisrom the northern and southern clades is 6.2%. Esti-ates of mtDNA sequence divergence for reptile spe-
ies for which branching events have been confidentlyated using fossil records or geological events rangerom 0.47 to 1.32% per million years (Zamudio andreene, 1997). Relying on these lower and upper esti-
ates, the two clades of endemic Baja Californian
gfioe
gCgsaLpCgfMBrm(PsActtg
C
cihbivrp3sHpcabscv
sbwcmr
ecdtctbaotwhivtaSIMfmlCwta
PfgmmavStiLddlwPbfitr
BL
47SYSTEMATICS AND BIOGEOGRAPHY OF Pituophis
opher snakes diverged 13.2–4.7 Mya. This estimate isor the divergence of the mtDNA lineages, not necessar-ly of the populations, and consequently it indicatesnly a maximum age for the split of the two clades ofndemic Baja Californian gopher snakes.Nevertheless, the above estimate suggests that the
enetic breakup between northern and southern Bajaalifornian gopher snakes was caused by a physiogeo-raphic event older than the formation of the midpenin-ular seaway. Elaborating on an idea apparently firstdvanced by Klauber (1947) and later restated byeviton and Tanner (1960), and relying on updatedaleogeographic evidence of the tectonic origin of Bajaalifornia, Murphy (1975, 1983a,b) proposed a ‘‘trans-ulfian vicariance model’’ for the origin of the herpeto-auna of the peninsula (see also Grismer, 1994). In
urphy’s scenario, several taxa migrated along withaja California when the area that became the Capeegion started rifting away from the western coast ofainland Mexico beginning approximately 11 Mya
late Miocene). If Murphy’s vicariance model applies toituophis, the endemic Baja Californian gopher snakeshould be the sister group of at least one of the middlemerican species of Pituophis, P. deppei and P. lineati-
ollis, a prediction not supported by our data. Hence,he historical events responsible for the genetic discon-inuity between northern and southern Baja Californiaopher snake populations remain unknown.
omparisons with Previous Hypothesesof Relationships within Pituophis
Although phylogenetic hypotheses do not depict an-estor–descendant relationships among the taxa stud-ed, our findings agree in part with two previousypotheses of relationships within Pituophis (Fig. 3),oth of which were based on morphological data andnferred using noncladistic methods. In Stull’s (1940)iew, sayi evolved from an affinis stock and in turn gaveise to ruthveni, which was the ancestor of the easterninesnakes (lodingi, melanoleucus, and mugitus; Fig.a). Our data support a close relationship betweenome affinis and sayi and between sayi and ruthveni.owever, contrary to Stull’s hypothesis, the easterninesnakes form a monophyletic group that is notlosely related to ruthveni. The mtDNA sequence datalso agree with Stull’s suggestion of a close relationshipetween affinis and deserticola, between catenifer andome deserticola, and between P. deppei and P. lineati-ollis but do not support a clade composed of affinis andertebralis, nor of annectens and deserticola.Klauber (1947) presented a hypothesis of relation-
hips within Pituophis (Fig. 3b) similar to Stull’s (1940)ut thought that the most primitive forms of the genusere P. deppei and P. lineaticollis. As Stull did, Klauber
onsidered affinis the most primitive taxon of theelanoleucus complex, from which he believed sayi,
uthveni, and the eastern pinesnakes evolved. In Klaub- D
r’s view, a second major offshoot of affinis was deserti-ola, from which first catenifer and then annectens wereerived. Our results support a close relationship be-ween some affinis and some sayi and between someatenifer and some deserticola but not between annec-ens and deserticola. Although Klauber considered thatimaris and vertebralis evolved from an affinis-likencestor that crossed the area now occupied by the Gulff California, he admitted that the derivation of thesewo forms was the most difficult phylogenetic problemithin Pituophis. As discussed above, the evolutionaryistory of the endemic Baja Californian gopher snakes
s indeed complex, and the two clades of bimaris andertebralis identified in our analyses are closely relatedo affinis, annectens, catenifer, and deserticola. Klauberlso believed that the insular forms pumilus (fromanta Cruz Island) and fuliginatus (from San Martınsland) evolved from nearby mainland forms, and ourP and ML trees indicate that these two taxa are in
act closely related to mainland populations of theelanoleucus complex geographically close to the is-
ands. Although specimens of coronalis (from Southoronado Island) and insulanus (from Cedros Island)ere not available for this study, we hypothesize that
hey are also closely related to populations on thedjacent mainland.
CONCLUSION
The view of a single, widely distributed, polytypicituophis melanoleucus is inconsistent with the in-
erred evolutionary history of these snakes. Our phylo-enetic analyses indicated that two segments of theelanoleucus complex, the lodingi–melanoleucus–ugitus eastern pinesnake clade and the affinis–
nnectens–bimaris–catenifer–deserticola–sayi–ruthveni–ertebralis clade from central and western Unitedtates and northern Mexico, represent divergent, allopa-ric lineages with no known intergradation zone. Wenterpret the available evidence as suggesting that theouisiana pinesnake has attained the status of indepen-ent evolutionary lineage. We therefore recognize threeistinct species in the melanoleucus complex, P. melano-eucus (sensu stricto), P. catenifer, and P. ruthveni;hether additional species should be recognized within. catenifer remains questionable. The quotation at theeginning of this paper was fitting in 1900, and thendings and arguments herein presented demonstratehat, despite several additional taxonomic studies, itemains so a century later.
ACKNOWLEDGMENTS
We thank H. W. Greene, D. L. Hardy, Sr., J. Gee, L. L. Grismer,. D. Hollingsworth, the late B. E. Dial, J. A. Campbell, M. Sasa,. M. Hardy, J. G. Himes, H. K. Reinert, G. M. Bolen, T. Benson, S. M.
eban, A. de Queiroz, W. I. Lutterschmidt, R. E. Jones, M. D. Matocq,
TCsaHRJMdaoNMaGaaH
A
A
A
A
A
B
B
B
B
B
B
B
C
C
C
C
C
C
C
C
C
C
C
C
D
d
d
D
D
D
D
D
E
F
F
F
F
F
F
F
F
G
G
48 RODRIGUEZ-ROBLES AND DE JESUS-ESCOBAR
. Moise, D. A. Creer, R. S. Reiserer, K. R. Zamudio, J. V. Vindum, D.hiszar, H. M. Smith, and C. J. Bell for kindly providing all thepecimens or tissue samples used in this study; R. M. Harland forllowing us to use his laboratory facilities; K. R. Zamudio and M.aggart for providing the primers; C. D. Ortiz, G. M. Bolen, and Y.odrıguez for assistance; H. W. Greene, J. L. Patton, K. R. Zamudio,. P. Huelsenbeck, M. Garcıa-Parıs, M. J. Mahoney, A. Graybeal, A.eyer, D. Parks, and S. K. Davis for innumerable, enlightening
iscussions on phylogenetic analyses; and H. W. Greene, J. L. Patton,nd D. B. Wake for valuable suggestions to improve previous versionsf the manuscript. This study was supported by a grant from theorthern California Chapter of the ARCS Foundation, by an Annie. Alexander Fellowship from the Museum of Vertebrate Zoology,
nd by a Mentored Research Award from the Office of the Dean of theraduate Division, University of California at Berkeley, to J.A.R.; byNational Science Foundation predoctoral fellowship to J.M.D.J.;
nd by a National Institutes of Health grant (GM 49346) to R. M.arland.
REFERENCES
lcobendas, M., Dopazo, H., and Alberch, P. (1996). Geographicvariation in allozymes of populations of Salamandra salamandra(Amphibia: Urodela) exhibiting distinct reproductive modes. J.Evol. Biol. 9: 83–102.
revalo, E., Davis, S. K., and Sites, J. W., Jr. (1994). MitochondrialDNA sequence divergence and phylogenetic relationships amongeight chromosome races of the Sceloporus grammicus complex(Phrynosomatidae) in central Mexico. Syst. Biol. 43: 387–418.
vise, J. C. (1989). A role for molecular genetics in the recognition andconservation of endangered species. Trends Ecol. Evol. 5: 279–281.
vise, J. C. (1994). ‘‘Molecular Markers, Natural History and Evolu-tion,’’ Chapman & Hall, New York.
vise, J. C., and Wollenberg, K. (1997). Phylogenetics and the originof species. Proc. Natl. Acad. Sci. USA 77: 7748–7755.
all, R. M., Jr., and Avise, J. C. (1992). Mitochondrial DNA phylogeo-graphic differentiation among avian populations and the evolution-ary significance of subspecies. Auk 109: 626–636.aum, D. (1992). Phylogenetic species concepts. Trends Ecol. Evol. 7:1–2.ehler, J. L. (1995). ‘‘National Audubon Society Field Guide to NorthAmerican Reptiles and Amphibians,’’ Knopf, New York.
ernatchez, L., and Wilson, C. C. (1998). Comparative phylogeogra-phy of Nearctic and Palearctic fishes. Mol. Ecol. 7: 431–452.
erry, V., and Gascuel, O. (1996). On the interpretation of bootstraptrees: Appropriate threshold of clade selection and induced gain.Mol. Biol. Evol. 13: 999–1011.
lair, W. F. (1958). Distributional patterns of vertebrates in thesouthern United States in relation to past and present environ-ments. In ‘‘Zoogeography’’ (C. L. Hubbs, Ed.), pp. 433–468. Am.Assoc. Adv. Sci., Washington, DC.rower, A. V. Z., DeSalle, R., and Vogler, A. (1996). Gene trees, speciestrees, and systematics: A cladistic perspective. Annu. Rev. Ecol.Syst. 27: 423–450.
amin, J. H., and Sokal, R. R. (1965). A method for deducingbranching sequences in phylogeny. Evolution 19: 311–326.icero, C. (1996). Sibling species of titmice in the Parus inornatuscomplex (Aves: Paridae). Univ. California Publ. Zool. 128: 1–217.ollins, J. T. (1991). Viewpoint: A new taxonomic arrangement forsome North American amphibians and reptiles. Herpetol. Rev. 22:42–43.ollins, J. T. (1992). The evolutionary species concept: A reply to VanDevender et al. and Montanucci. Herpetol. Rev. 23: 43–46.onant, R. (1956). A review of two rare pine snakes from the Gulf
Coastal Plain. Am. Mus. Novit. 1781: 1–31. Gonant, R. (1965). Miscellaneous notes and comments on toads,lizards, and snakes from Mexico. Am. Mus. Novit. 2205: 1–38.
onant, R., and Collins, J. T. (1991). ‘‘A Field Guide to Reptiles andAmphibians of Eastern and Central North America,’’ 3rd ed.,Houghton Mifflin, Boston.ope, E. D. (1891). The epiglottis in colubrine snakes. Am. Nat. 25:156–157.
ope, E. D. (1900). ‘‘The Crocodilians, Lizards, and Snakes of NorthAmerica. Report of the U. S. National Museum for 1898,’’ pp.153–1270, Government Printing Office, Washington, DC.racraft, J. (1983). Species concepts and speciation analysis. In‘‘Current Ornithology’’ (R. F. Johnston, Ed.), Vol. 1, pp. 159–187.Plenum, New York.
racraft, J., and Helm-Bychowski, K. (1991). Parsimony and phyloge-netic inference using DNA sequences: Some methodological strate-gies. In ‘‘Phylogenetic Analysis of DNA Sequences’’ (M. M. Miya-moto and J. Cracraft, Eds.), pp. 184–220. Oxford Univ. Press,Oxford.
rain, J. L., and Cliburn, J. W. (1971). Pituophis melanoleucuslodingi from the western part of its range. Southwest. Nat. 15:496–497.egenhardt, W. G., Painter, C. W., and Price, A. H. (1996). ‘‘Amphib-ians and Reptiles of New Mexico,’’ Univ. of New Mexico Press,Albuquerque.
e Queiroz, K., and Donoghue, M. J. (1988). Phylogenetic systematicsand the species problem. Cladistics 4: 317–338.
e Queiroz, K., and Donoghue, M. J. (1990). Phylogenetic systematicsand species revisited. Cladistics 6: 83–90.ixon, J. R., Sabbath, M., and Worthington, R. (1962). Comments onsnakes from central and western Mexico. Herpetologica 18: 91–100.owling, H. G. (1958). Pleistocene snakes of the Ozark Plateau. Am.Mus. Novit. 1882: 1–9.owling, H. G. (1993). Viewpoint: A reply to Collins. Herpetol. Rev.24: 11–13.uellman, W. E. (1960). A taxonomic study of the Middle Americansnake, Pituophis deppei. Univ. Kansas Publ. Mus. Nat. Hist. 10:601–610.urham, J. W., and Allison, E. C. (1960). The geologic history of BajaCalifornia and its marine faunas. Syst. Zool. 9: 47–91.dwards, J. G. (1954). A new approach to infraspecific categories.Syst. Zool. 3: 1–20.
arris, J. S. (1982). Outgroups and parsimony. Syst. Zool. 31:328–334.
elsenstein, J. (1981). Evolutionary trees from DNA sequences: Amaximum likelihood approach. J. Mol. Evol. 17: 368–376.
elsenstein, J. (1985). Confidence limits on phylogenies: An approachusing the bootstrap. Evolution 39: 783–791.
elsenstein, J., and Kishino, H. (1993). Is there something wrongwith the bootstrap on phylogenies? A reply to Hillis and Bull. Syst.Biol. 42: 193–200.
itch, H. S. (1985). Variation in clutch and litter size in New Worldreptiles. Univ. Kansas. Mus. Nat. Hist. Misc. Publ. 76: 1–76.
rost, D. R., and Hillis, D. M. (1990). Species in concept and practice:Herpetological applications. Herpetologica 46: 87–104.
rost, D. R., Kluge, A. G., and Hillis, D. M. (1992). Species incontemporary herpetology: Comments on phylogenetic inferenceand taxonomy. Herpetol. Rev. 23: 46–54.
ugler, C. M. (1955). New locality records for the Louisiana pinesnake, Pituophis catenifer ruthveni Stull. Herpetologica 11: 24.arcıa-Moreno, J., and Fjeldså, J. (1999). Re-evaluation of specieslimits in the genus Atlapetes based on mtDNA sequence data. Ibis141: 199–207.eist, V. (1992). Endangered species and the law. Nature 357:274–276.
ood, D. A., and Wake, D. B. (1992). Geographic variation and
GG
G
G
G
H
H
H
H
H
H
H
H
H
I
J
K
K
K
K
K
K
K
K
L
L
L
LM
M
M
M
M
M
M
M
M
M
M
O
P
P
R
R
R
R
49SYSTEMATICS AND BIOGEOGRAPHY OF Pituophis
speciation in the torrent salamanders of the genus Rhyacotriton(Caudata: Rhyacotritonidae). Univ. California Publ. Zool. 126:1–91.raybeal, A. (1995). Naming species. Syst. Biol. 44: 237–250.reene, H. W. (1994). Systematics and natural history, foundationsfor understanding and conserving biodiversity. Am. Zool. 34:48–56.rismer, L. L. (1994). The origin and evolution of the peninsularherpetofauna of Baja California, Mexico. Herpetol. Nat. Hist. 2:51–106.rismer, L. L. (1997). The distribution of Pituophis melanoleucus andP. vertebralis in northern Baja California, Mexico. Herpetol. Rev.28: 68–70.u, X., Fu, Y.-X., and Li, W.-H. (1995). Maximum likelihood estima-tion of the heterogeneity of substitution rate among nucleotidesites. Mol. Biol. Evol. 12: 546–557.endy, M. D., and Penny, D. (1982). Branch and bound algorithms todetermine minimal evolutionary trees. Math. Biosci. 59: 277–290.ighton, R. (1998). Is Ensatina eschscholtzii a ring species? Herpeto-logica 54: 254–278.illis, D. M., and Bull, J. J. (1993). An empirical test of bootstrappingas a method for assessing confidence in phylogenetic analysis. Syst.Biol. 42: 182–192.illis, D. M., and Huelsenbeck, J. P. (1992). Signal, noise, andreliability in molecular phylogenetic analysis. J. Hered. 83: 189–195.illis, D. M., Moritz, C., and Mable, B. K., Eds. (1996). ‘‘MolecularSystematics,’’ 2nd ed., Sinauer, Sunderland, MA.olman, J. A. (1971). Ophisaurus attenuatus. Cat. Am. Amphib. Rept.111.1–111.3.olman, J. A. (1996). The large Pleistocene (Sangamonian) herpeto-fauna of the Williston IIIA site, north-central Florida. Herpetol.Nat. Hist. 4: 35–47.uelsenbeck, J. P. (1991). Tree-length distribution skewness: Anindicator of phylogenetic information. Syst. Zool. 40: 257–270.uelsenbeck, J. P., and Crandall, K. A. (1997). Phylogeny estimationand hypothesis testing using maximum likelihood. Annu. Rev.Ecol. Syst. 28: 437–466.
nger, R. F. (1961). Problems in the application of the subspeciesconcept in vertebrate taxonomy. In ‘‘Vertebrate Speciation’’ (W. F.Blair, Ed.), pp. 262–285. Univ. of Texas Press, Austin.
ohnson, N. K. (1995). Speciation in vireos. I. Macrogeographicpatterns of allozymic variation in the Vireo solitarius complex inthe contiguous United States. Condor 97: 903–919.allersjo, M., Farris, J. S., Kluge, A. G., and Bult, C. (1992).Skewness and permutation. Cladistics 8: 275–287.ardong, K. V. (1980). Gopher snakes and rattlesnakes: PresumptiveBatesian mimicry. Northwest Sci. 54: 1–4.eita, S. O. Y. (1993). The subspecies concept in zoology andanthropology—A brief historical review and test of a classificationscheme. J. Black Stud. 23: 416–445.ishino, H., and Hasegawa, M. (1989). Evaluation of the maximumlikelihood estimate of the evolutionary tree topologies from DNAsequence data, and the branching order in Hominoidea. J. Mol.Evol. 29: 170–179.lauber, L. M. (1946). The gopher snakes of Baja California, withdescriptions of new subspecies of Pituophis catenifer. Trans. SanDiego Soc. Nat. Hist. 11: 1–40.lauber, L. M. (1947). Classification and ranges of the gopher snakesof the genus Pituophis in the western United States. Bull. Zool.Soc. San Diego 22: 1–81.night, J. L. (1986). Variation in snout morphology in the NorthAmerican snake Pituophis melanoleucus (Serpentes: Colubridae).J. Herpetol. 20: 77–79.umazawa, Y., Ota, H., Nishida, M., and Ozawa, T. (1998). The
complete nucleotide sequence of a snake (Dinodon semicarinatus) Rmitochondrial genome with two identical control regions. Genetics150: 313–329.
ara, M. C., Patton, J. L., and Da Silva, M. N. F. (1996). Thesimultaneous diversification of South American echimyid rodents(Hystricognathi) based on complete cytochrome b sequences. Mol.Phylogenet. Evol. 5: 403–413.
arson, A. (1994). The comparison of morphological and moleculardata in phylogenetic systematics. In ‘‘Molecular Ecology and Evolu-tion: Approaches and Applications’’ (B. Schierwater, B. Streit, G. P.Wagner, and R. DeSalle, Eds.), pp. 371–390. Birkhauser, Basel.
eviton, A. E., and Tanner, W. W. (1960). The generic allocation ofHypsiglena slevini Tanner (Serpentes: Colubridae). Occ. Pap. Cali-fornia Acad. Sci. 27: 1–7.
i, W.-H. (1997). ‘‘Molecular Evolution,’’ Sinauer, Sunderland, MA.addison, W. P., Donoghue, M. J., and Maddison, D. R. (1984).Outgroup analysis and parsimony. Syst. Zool. 33: 83–103.artin, W. F., and Huey, R. B. (1971). The function of the epiglottis insound production (hissing) of Pituophis melanoleucus. Copeia1971: 752–754.cKitrick, M. C., and Zink, R. M. (1988). Species concepts inornithology. Condor 90: 1–14.iththapala, S., Seidensticker, J., and O’Brien, S. J. (1996). Phylogeo-graphic subspecies recognition in leopards (Panthera pardus):Molecular genetic variation. Conserv. Biol. 10: 1115–1132.oore, W. S. (1995). Inferring phylogenies from mtDNA variation:Mitochondrial-gene trees versus nuclear-gene trees. Evolution 49:718–726.oore, W. S. (1997). Mitochondrial-gene trees versus nuclear-genetrees, a reply to Hoelzer. Evolution 51: 627–629.oore, W. S., and DeFilippis, V. R. (1997). The window of taxonomicresolution for phylogenies based on mitochondrial cytochrome b. In‘‘Avian Molecular Evolution and Systematics’’ (D. P. Mindell, Ed.),pp. 83–119. Academic Press, San Diego.orafka, D. (1977). ‘‘Biogeographical Analysis of the ChihuahuanDesert Through its Herpetofauna,’’ Junk, The Hague.urphy, R. W. (1975). Two new blind snakes (Serpentes: Leptotyphlo-pidae) from Baja California, Mexico with a contribution to thebiogeography of peninsular and insular herpetofauna. Proc. Califor-nia Acad. Sci. 40: 93–107.urphy, R. W. (1983a). Paleobiogeography and genetic differentia-tion of the Baja California herpetofauna. Occ. Pap. CaliforniaAcad. Sci. 137: 1–48.urphy, R. W. (1983b). The reptiles: Origins and evolution. In ‘‘IslandBiogeography in the Sea of Cortez’’ (T. J. Case and M. L. Cody,Eds.), pp. 130–158. Univ. of California Press, Berkeley.’Brien, S. J., and Mayr, E. (1991). Bureaucratic mischief: Recogniz-ing endangered species and subspecies. Science 251: 1187–1188.
almer, W. M., and Braswell, A. L. (1995). ‘‘Reptiles of NorthCarolina,’’ Univ. of North Carolina Press, Chapel Hill.
atton, J. L., and Smith, M. F. (1994). Paraphyly, polyphyly, and thenature of species boundaries in pocket gophers (genus Thomomys).Syst. Biol. 43: 11–26.eichling, S. B. (1990). Reproductive traits of the Louisiana pinesnake, Pituophis melanoleucus ruthveni (Serpentes: Colubridae).Southwest. Nat. 35: 221–222.
eichling, S. B. (1995). The taxonomic status of the Louisiana pinesnake (Pituophis melanoleucus ruthveni) and its relevance to theevolutionary species concept. J. Herpetol. 29: 186–198.
odrıguez-Robles, J. A., and De Jesus-Escobar, J. M. (1999). Molecu-lar systematics of New World lampropeltinine snakes (Colubridae):Implications for biogeography and evolution of food habits. Biol. J.Linn. Soc. 68, in press.oze, J. A., and Tilger, G. M. (1983). Micrurus fulvius. Cat. Am.Amphib. Rept. 316.1–316.4.
usso, C. A. M. (1997). Efficiencies of different statistical tests in
R
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
T
T
T
U
W
W
W
W
W
W
W
W
W
W
W
Y
Y
Y
Z
Z
Z
Z
Z
Z
Z
Z
Z
50 RODRIGUEZ-ROBLES AND DE JESUS-ESCOBAR
supporting a known vertebrate phylogeny. Mol. Biol. Evol. 14:1078–1080.
usso, C. A. M., Takezaki, N., and Nei, M. (1996). Efficiencies ofdifferent genes and different tree-building methods in recovering aknown vertebrate phylogeny. Mol. Biol. Evol. 13: 525–536.
aiki, R. K., Bugawan, T. L., Horn, G. T., Mullis, K. B., and Erlich, H.A. (1986). Analyses of enzymatically amplified b-globin and HLA-DQa DNA with allele-specific oligonucleotide probes. Nature 324:163–166.
aiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi, R.,Horn, G. T., Mullis, K. B., and Erlich, H. A. (1988). Primer-directedenzymatic amplification of DNA with a thermostable DNA polymer-ase. Science 239: 487–491.
ambrook, J., Fritsch, E. F., and Maniatis, T. (1989). ‘‘MolecularCloning: A Laboratory Manual,’’ 2nd ed., Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY.
anchez-Herrera, O., Smith, H. M., and Chiszar, D. (1981). Anothersuggested case of ophidian deceptive mimicry. Trans. Kansas Acad.Sci. 84: 121–127.
anderson, M. J. (1995). Objections to bootstrapping phylogenies: Acritique. Syst. Biol. 44: 299–320.
anger, F., Nicklen, S., and Coulson, A. R. (1977). DNA sequencingwith chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463–5467.
haffer, H. B., and McKnight, M. L. (1996). The polytypic speciesrevisited: Genetic differentiation and molecular phylogenetics ofthe tiger salamander Ambystoma tigrinum (Amphibia: Caudata)complex. Evolution 50: 417–433.
ites, J. W., Jr., and Crandall, K. (1997). Testing species boundariesin biodiversity studies. Conserv. Biol. 11: 1289–1297.
mith, H. M., Chiszar, D., and Montanucci, R. R. (1997). Subspeciesand classification. Herpetol. Rev. 27: 13–16.
mith, H. M., and Kennedy, J. P. (1951). Pituophis melanoleucusruthveni in eastern Texas and its bearing on the status of Pituophiscatenifer. Herpetologica 7: 93–96.
oltis, D. E., Soltis, P. S., and Doyle, J. J., Eds. (1998). ‘‘MolecularSystematics of Plants II: DNA Sequencing,’’ Kluwer Academic, Boston.
tebbins, R. C. (1985). ‘‘A Field Guide to Western Reptiles andAmphibians,’’ 2nd ed., Houghton Mifflin, Boston.
tull, O. G. (1940). Variations and relationships in the snakes of thegenus Pituophis. Bull. U. S. Natl. Mus. Smith. Inst. WashingtonDC 175: 1–225.
weet, S. S. (1984). Species groups within Pituophis melanoleucus:Evidence from morphology. ASIH/HL/SSAR Meeting Program.Norman, Oklahoma, p. 200. [Abstract].
weet, S. S. (1985). Geographic variation, convergent crypsis andmimicry in gopher snakes (Pituophis melanoleucus) and westernrattlesnakes (Crotalus viridis). J. Herpetol. 19: 55–67.
weet, S. S., and Parker, W. S. (1990). Pituophis melanoleucus(Daudin). Pine, bull, and gopher snakes. Cat. Am. Amphib. Rept.474.1–474.8.
wofford, D. L. (1999). ‘‘PAUP*: Phylogenetic Analysis Using Parsi-mony (* and Other Methods),’’ Vers. 4. Sinauer, Sunderland, MA.
wofford, D. L., Olsen, G. J., Waddell, P. J., and Hillis, D. M. (1996).Phylogenetic inference. In ‘‘Molecular Systematics’’ (D. M. Hillis, C.Moritz, and B. K. Mable, Eds.), 2nd ed., pp. 407–514. Sinauer,Sunderland, MA.
amura, K., and Nei, M. (1993). Estimation of the number ofnucleotide substitutions in the control region of mitochondrialDNA in humans and chimpanzees. Mol. Biol. Evol. 10: 512–526.
empleton, A. R. (1983). Phylogenetic inference from restrictionendonuclease cleavage site maps with particular reference to theevolution of humans and the apes. Evolution 37: 221–244.
ennant, A. (1984). ‘‘The Snakes of Texas,’’ Texas Monthly Press,
Austin.pton, D. E., and Murphy, R. W. (1997). Phylogeny of the side-blotched lizards (Phrynosomatidae: Uta) based on mtDNA se-quences: Support for a midpeninsular seaway in Baja California.Mol. Phylogenet. Evol. 8: 104–113.ake, D. B., and Schneider, C. J. (1998). Taxonomy of the plethodon-tid salamander genus Ensatina. Herpetologica 54: 279–298.alker, D., Ortı, G., and Avise, J. C. (1998). Phylogenetic distinctive-ness of a threatened aquatic turtle (Sternotherus depressus).Conserv. Biol. 12: 639–645.asserman, A. O. (1968). Scaphiopus holbrookii. Cat. Am. Amphib.Rept. 70.1–70.4.atrous, L. E., and Wheeler, Q. D. (1981). The out-group comparisonmethod of character analysis. Syst. Zool. 30: 1–11.hite, C. A. (1884). On the character and function of the epiglottis inthe bull-snake (Pityophis). Am. Nat. 18: 19–21.iens, J. A., Ed. (1982). Forum: Avian subspecies in the 1980’s. Auk99: 593–615.iley, E. O. (1978). The evolutionary species concept reconsidered.Syst. Zool. 27: 17–26.ilson, E. O., and Brown, W. L., Jr. (1953). The subspecies conceptand its taxonomic application. Syst. Zool. 2: 97–111.right, A. H., and Wright, A. A. (1957). ‘‘Handbook of Snakes of theUnited States and Canada,’’ Vol. II, Comstock Publishing Associ-ates, Ithaca, NY.uster, W., and Thorpe, R. S. (1992). Asiatic cobras: Populationsystematics of the Naja naja species complex (Serpentes: Elapidae)in India and central Asia. Herpetologica 48: 69–85.uster, W., and Thorpe, R. S. (1994). Naja siamensis, a crypticspecies of venomous snake revealed by mtDNA sequencing. Experi-entia 50: 75–79.
ang, Z. (1994). Estimating the pattern of nucleotide substitution. J.Mol. Evol. 39: 105–111.
ang, Z. (1996). Among-site rate variation and its impact on phyloge-netic analyses. Trends Ecol. Evol. 11: 367–372.
oung, B. A., Sheft, S., and Yost, W. (1995). Sound production inPituophis melanoleucus (Serpentes: Colubridae) with the firstdescription of a vocal cord in snakes. J. Exp. Zool. 273: 472–481.
amudio, K. R., and Greene, H. W. (1997). Phylogeography of thebushmaster (Lachesis muta: Viperidae): Implications for neotropi-cal biogeography, systematics, and conservation. Biol. J. Linn. Soc.62: 421–442.
amudio, K. R., Jones, K. B., and Ward, R. H. (1997). Molecularsystematics of short-horned lizards: Biogeography and taxonomy ofa widespread species complex. Syst. Biol. 46: 284–305.
appalorti, R. T., Johnson, E. W., and Leszczynski, Z. (1983). Theecology of the northern pine snake (Pituophis melanoleucus),(Daudin) (Reptilia, Serpentes, Colubridae), in southern New Jer-sey, with special notes on habitat and nesting behavior. Bull.Chicago Herpetol. Soc. 18: 57–72.
ardoya, R., and Meyer, A. (1996). Phylogenetic performance ofmitochondrial protein-coding genes in resolving relationshipsamong vertebrates. Mol. Biol. Evol. 13: 933–942.
harkikh, A., and Li, W.-H. (1992). Statistical properties of bootstrapestimation of phylogenetic variability from nucleotide sequences. I.Four taxa with a molecular clock. Mol. Biol. Evol. 9: 1119–1147.
hu, D., Degnan, S., and Moritz, C. (1998). Evolutionary distinctive-ness and status of the endangered Lake Eacham rainbowfish(Melanotaenia eachamensis). Conserv. Biol. 12: 80–93.
ink, R. M., and McKitrick, M. C. (1995). The debate over speciesconcepts and its implications for ornithology. Auk 112: 701–719.
ink, R. M., Blackwell, R. C., and Rojas-Soto, O. (1997). Species limitsin the Le Conte’s thrasher. Condor 99: 132–138.
ug, G. R., and Schwartz, A. (1971). Deirochelys, D. reticularia. Cat.
Am. Amphib. Rept. 107.1–107.3.