molecular identification of giardia duodenalis isolates from humans, dogs, cats and cattle from the...
TRANSCRIPT

Iranian J Parasitol: Vol. 9, No. 1, Jan -Mar 2014, pp.70-78
70 Available at: http://ijpa.tums.ac.ir
Original Article
Molecular Identification of Giardia duodenalis Isolates from Fars Province, Iran
Mohammad RAYANI 1, 2
, Ngah ZASMY UNYAH 2, *Gholamreza HATAM
3
1. Department of Microbiology and Parasitology, School of Health, Bushehr University of Medical Sciences, Bushehr, Iran
2. Department of Medical Microbiology and Parasitology, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia, Malaysia
3. Basic Sciences in Infectious Diseases Research Center, Medical School, Shiraz University of Medical Sciences, Shiraz, Iran
Received 05 Aug 2013 Accepted 19 Nov 2013
Abstract Background: Giardia duodenalis is one of the most common human intestinal pro-
tozoan parasites worldwide and is endemic throughout the world with a vast range of mammalian hosts. The present study aimed to identify the prevalence of G. duo-denalis isolates and determine the most common of its assemblages in the patients referring to health centers and hospitals in Fars province, Iran that will be subjected to further molecular investigation. Methods: We collected 1000 human fecal samples from health centers and hospi-tals in Shiraz, Iran in a one year period from September 2009 to August 2010. Mi-croscopic examination for the presence of G. duodenalis cysts and trophozoites was performed by direct wet mount before and after the concentration techniques. Ex-traction of DNA was performed by Phenol-Chloroform-Isoamylalcohol (PCI). G. duodenalis-positive specimens were analyzed by PCR. A fragment of the SSU-rDNA (292 bp) gene was amplified by PCR using the forward primer RH11 and the re-verse primer RH4. Genotyping was performed using sequence analysis of G. duode-nalis glutamate dehydrogenase gene using primers GDHeF, GDHiF, and GDHiR. Results: The prevalence of Giardia infection was 10.7% (107/1000) examined based on microscopic examination. PCR identified 80% (40/50) of the samples as positive for G. duodenalis based on SSU-rDNA amplification on sucrose gradient samples. Besides, genotyping results indicated 32 isolates (80%) as assemblage AII and 8 isolates (20%) as assemblage BIII and BIV based on the DNA sequence analysis of the glutamate dehydrogenase locus of G. duodenalis. Conclusion: The findings of this study emphasize that Iran (Fars Province) is a favorable area for giardiasis with an anthroponotic infection route.
Keywords Giardia duodenalis, Microscopic, Molecular characterization,
Prevalence, Iran, SSU-rDNA
*Correspondence Email: [email protected],ir [email protected]
Iranian Society of Parasitology
http:// isp.tums.ac.ir
Iranian J Parasitol
Open access Journal at
http:// ijpa.tums.ac.ir
Tehran University of Medical
Sciences Publication
http:// tums.ac.ir

Rayani et al.: Molecular Identification of Giardia …
Available at: http://ijpa.tums.ac.ir 71
Introduction
iardia duodenalis (synonymous G. lamblia or G. intestinalis) is one of the most common intestinal parasites in
humans worldwide and is endemic throughout the world with a vast range of mammalian hosts (1). The genus Giardia is categorized as phylum Sarcomastigophora, class Zoomasti-gophorea, and a member of the order Diplo-monadida (2). Although the specific name duo-denalis is as zoological nomenclature, the names intestinalis and lamblia are often used, particularly for the isolates of human origin (3). The prevalence of this infection varies be-tween 2-5% in industrialized countries up to 20–30% in less developed countries (4). About 200 million people have symptomatic giardia-sis in Asia, Africa, and Latin America with some 500,000 new cases reported each year (5, 6). Giardiasis is among the top ten human parasitic diseases that is a widespread intestinal disease, responsible for 2.5 million diarrhea and nutritional deficiencies in children in de-veloping countries (7). Giardia grows in the small intestine and multiplies by asexual binary fission on the surface of the mucosa. Tropho-zoite and cyst are two distinct forms of Giardia. Ingestion of tetra nucleated agent is responsi-ble for transmission from one host to another (8, 3). Giardiasis has variable clinical symp-toms from the absence of symptoms to acute or chronic diarrhea, dehydration, abdominal pain, nausea, vomiting, and weight loss. The interplay between the virulence of the parasite and the developmental, nutritional, and im-munological status of the host determines the severity of the disease (9-11).
Parasitological techniques have been applied for detecting Giardia using direct wet-mount, concentration, and staining method (12, 13). Moreover, techniques of molecular detection based on PCR have been developed to detect G. duodenalis cysts in feces from isolates of hu-man and animal origin worldwide. In addition, these molecular methods allow the genotyping
of G. duodenalis cysts. Molecular genetics tech-niques have successfully provided new power-ful tools to characterize Giardia isolates and G. duodenalis specific. The majority of molecular studies of G. intestinalis have relied on the anal-ysis of the small subunit ribosomal RNA (SSU-rRNA), β-giardin (bg), glutamate dehy-drogenase (gdh), elongation factor 1-alpha (ef-1), triose phosphate isomerise (tpi), and variant surface protein (vsp) genes (14). These tools are widely used for identification of G. duode-nalis genotypes in clinical specimens as well as differentiation of Giardia at the species/asse-mblage and genotype levels. The utility of mo-lecular diagnostic tools is frequently deter-mined by the SSU-rRNA, gdh, tpi, and bg genes targeted. The usage of these loci for the geno-typing or subtyping of G. duodenalis, their re-striction fragment length poly-morphism anal-ysis, sequence characteristics and phylogenetic tree is useful to understanding of the popula-tion genetics, epidemiology, and taxonomy of giardiasis in humans and domesticated animals (8). Isolates of G. duodenalis are classified into seven assemblages based on the char-acterization of the glutamate dehydrogenase (gdh), subunit ribosomal RNA (SSU-rRNA), and triose phosphate isomerase (tpi) genes. Two assemblages (A and B) are found in both humans and animals, whereas the remaining five (C-G) are animal host-specific (15). Be-sides, DNA sequence studies on G. duodenalis have shown the existence of two subgroups in assemblage A (AI and AII) and assemblage B (BIII and BIV). Genotype AI is generally found in animals, whereas genotype AII has mainly been identified in humans. Assemblage B comprises a genetically more diverse group of predominantly human isolates although some animal genotypes have been included, as well (16,17). Information on the genotype of G. duodenalis from individual patients over time will assist understanding the course of infec-tion and the rate of reinfection following
G

Iranian J Parasitol: Vol. 9, No. 1, Jan -Mar 2014, pp.70-78
72 Available at: http://ijpa.tums.ac.ir
treatment. It will also be possible to identify those genotypes of Giardia that are more per-sistent and associated with infections of long-term duration (18). Giardiasis has been one of the most common intestinal infections in Iran. In recent years the Iranian researchers have contributed greatly to increase their knowledge about giardiasis in human and an-imals (19, 20). Reports of giardiasis prevalence ranged from 1.4% to 59.6% among different socioeconomic and geographical situation in the community. The incidence of G. duodenalis in the patients who had referred to hospitals and health care centers was reported as 25.8%, 10.1%, and 21.3% by Arani et al. (12), Haghighi et al. (21), and Taherkhani et al. (22), respectively. Also, another study indicated that the prevalence of giardiasis varied in different parts of Iran (5%-23%) (22). The findings of the studies emphasize that Giardia infection is a major public health problem. Thus, the present study aims were to determine the prevalence of G. duodenalis and assemblages or sub-assemblage in the patients referring to health centers and hospitals in Fars province based on microscopic and molecular examination that presents a base situation of giardiasis in the un-derstudy region.
Materials and Methods
Sample collection In a one year period from September 2009,
one thousand human fecal samples were col-lected from the patients with gastrointestinal discomfort, such as diarrhea, dehydration, ab-dominal pain, nausea, vomiting, referring to the health centers and hospitals in Shiraz, south of Iran. For further examination, the samples were sent to the Research Laboratory of Intes-tinal Parasites in the Department of Parasit-ology and Mycology in Faculty of Medicine, Shiraz University of Medical Sciences.
Parasitological examination Microscopic confirmation for the presence
of G. duodenalis cysts and trophozoites was
performed by examination on direct wet mount before and after the concentration techniques. Direct microscopy was done on all formed and liquid samples. Sedimentation of ethyl acetate was performed in order to max-imize the number of detectable organisms which may be too limited to be seen by direct microscopy alone. The fluid on the side of the tube was allowed to drain onto the deposit. Aliquots of the concentrated fecal samples were stored at -20 ºC. Monolayer sucrose gra-dient with specific gravity at 0.85 M was done on positive stool samples (to purify fecal sam-ples and concentrate of cysts). Fresh stool samples were homogenized 1:10 (v/v) in dis-tilled water or physiologic saline solution, fil-tered through two layer gauze to discard larger detritus, and washed two times by spin at 400 × g for 5 min in 50 ml falcon tube. Then, 5 ml of the sample suspension was gently added to 5 ml of 0.85 M sucrose solution that made two separate layers in a 15-ml glass tube. After spin at 1700 × g for 10 min in a 4°C centri-fuge, the cysts formed a thin ring liquid layer between the sucrose solution and the suspen-sion that was carefully transferred into a 15-ml clean glass tube. The resulting cysts ring solu-tion was diluted in distilled water for twice (first time 1:10 (v/v) and second time 1:5 (v/v) and centrifuged at 400 × g for 5 min. The sediment material was then placed in a clean 1.5 ml ependorf tube. Aliquots of puri-fied fecal samples were stored at 4°C and -20 ºC.
Molecular techniques DNA extraction
DNA was extracted by Phenol-Chloroform-Isoamylalcohol (PCI) on purified and concen-trated fecal by sucrose gradient and sedimen-tation of ethyl acetate samples as follows. Moreover, glass beads or freeze-thaw was used for disruption of the cyst wall on some samples. First, 200 μl of sucrose gradient or sediment concentrated cysts sample and 200 μl 3%Triton X100 were mixed and incubated in a water bath at 75 ºC for 1 hour. Then, 200

Rayani et al.: Molecular Identification of Giardia …
Available at: http://ijpa.tums.ac.ir 73
μl of lysis buffer and 10 μl of proteinase K were added to 200 μl of homogenate and in-cubated at 37 ºC overnight. Phenol-chloroform extraction method (PCI) was used for extraction of genomic DNA from the cysts. PCI was then added to the solution and centrifuged at 15000 rpm at 25 ºC for 10 min. The supernatant was precipitated with ethanol absolute (2 or 4 equal) in a new tube at -20 ºC for 24 hours. The solution was centrifuged at 15000 rpm at 4 ºC for 10 min. After air dry sediment, 100 DDW or TE (100 mMTris, and 25 mM EDTA, pH= 8.0) was added to the sediment in the tube and stored at -20°C until PCR analysis.
Molecular identification PCR Amplification of SSU-rDNA
A fragment of the SSU-rDNA (292 bp) gene was amplified by PCR using the forward pri-mer RH11 (5′-CATCCGGTCGATCCTGCC-3′) and the reverse primer RH4 (5′-AG-TCGAAC CCTGATTCTCCGCCCAGG-3′) as previously described (23, 24) with some slight modification. The PCR mixture in the 25 µL reaction volume contained 1×PCR buffer, 1.5 mM MgCl2, 0.2 mM each dNTP, 0.5 µM for each forward and reverse primer, and 2 U Taq. The PCR was performed under the following conditions: An initial hot start at 96 ºC for 2 min followed by 30 cycles each including 96 ºC for 45 s (denaturing), 58 ºC for 30 s (annealing), 72 ºC for 45 s (extension), and a final extension step at 72 ºC for 4 min using (Corbett Research Thermal Cycler, Aus-tralia). The primers were checked by a positive and a negative control. The PCR products were electrophoresized on a 1.5% agarose gel with 5 μl of the reaction solution and visual-ized by staining the gel with ethidium bromide.
PCR Amplification and sequencing of the glutamate dehydrogenase gene (gdh)
A semi-nested PCR was performed using the primers GDHeF, GDHiF, and GDHiR to amplify a 432 bp fragment of Giardia gluta-mate dehydrogenase gene (gdh) (25) with some
slight modifications. Genotyping was per-formed using sequence analysis on 40 PCR products of G. duodenalis based on gdh amplifi-cation in both directions. DNA sequencing reactions were performed by First BASE La-boratories in Malaysia (http://www.base-asia.com) to sequencing commercially in both forward and reverse directions using primers GDHiF and GDHiR for GDH gene fragment.
Results
In this study, microscopy and molecular technique were performed directly on fecal samples. The results indicated that 107/1000 (10.7%) samples were positive for G. duodenalis based on microscopy. Parasite load for micro-scopically positive G. duodenalis samples were different in the quantity of cysts from low (less than 2 cysts), moderate (2 to 10 cysts) and high (more than 10 cysts) per 40X field of view. One step monolayer sucrose gradient successfully was done on positive stool sam-ples to purify fecal samples and concentrate G. duodenalis cysts (Fig.1 A-B).
Fig. 1: A and B- Microscopic slides of concen-trated and purified G. duodenalis cysts observing after 1 step sucrose gradient in the stool samples. (Original picture 100 X, 1000 X)
1000 X
100 X
Iranian J Parasitol: Vol. 9, No. 1, Jan -Mar 2014, pp.70-78
74 Available at: http://ijpa.tums.ac.ir
Pretreatment to disrupt the cysts wall was suc-cessfully performed before extraction of DNA using 3% Triton X100 (Fig. 2 A-B).
Fig. 2: A and B: Cyst wall disruption after using 3% TritonX100 on fresh sucrose gradient samples. (Original picture 100 X)
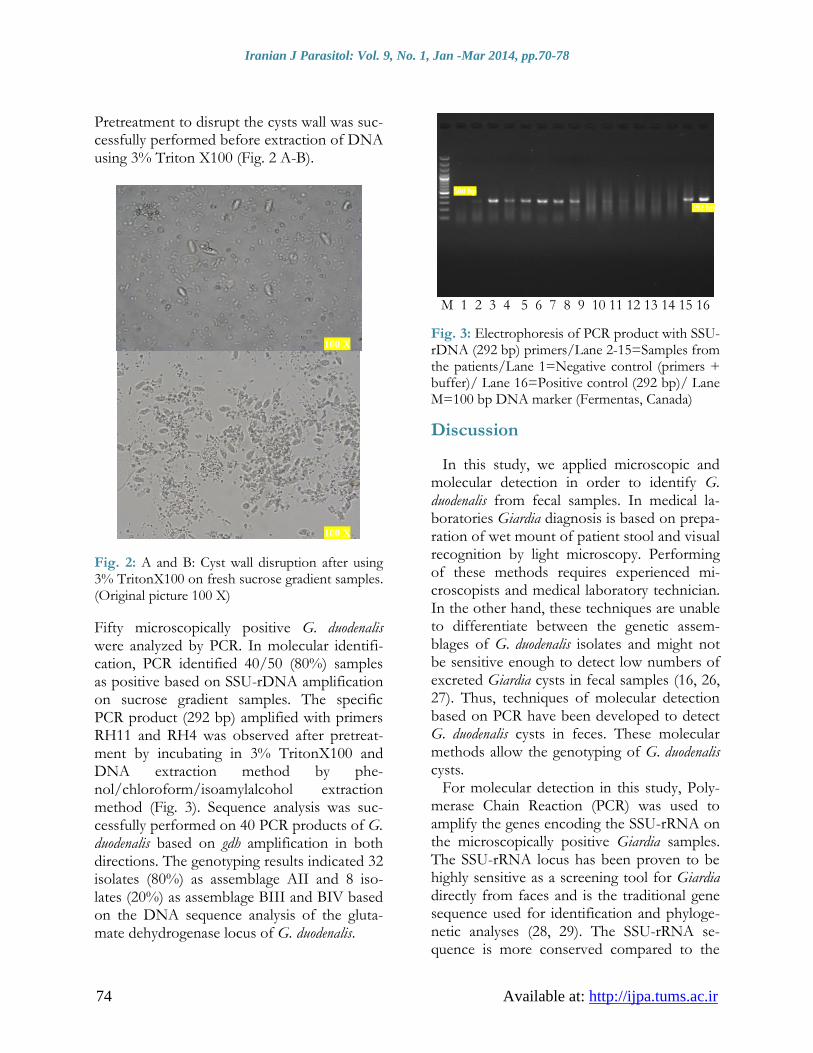
Fifty microscopically positive G. duodenalis were analyzed by PCR. In molecular identifi-cation, PCR identified 40/50 (80%) samples as positive based on SSU-rDNA amplification on sucrose gradient samples. The specific PCR product (292 bp) amplified with primers RH11 and RH4 was observed after pretreat-ment by incubating in 3% TritonX100 and DNA extraction method by phe-nol/chloroform/isoamylalcohol extraction method (Fig. 3). Sequence analysis was suc-cessfully performed on 40 PCR products of G. duodenalis based on gdh amplification in both directions. The genotyping results indicated 32 isolates (80%) as assemblage AII and 8 iso-lates (20%) as assemblage BIII and BIV based on the DNA sequence analysis of the gluta-mate dehydrogenase locus of G. duodenalis.
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Fig. 3: Electrophoresis of PCR product with SSU-rDNA (292 bp) primers/Lane 2-15=Samples from the patients/Lane 1=Negative control (primers + buffer)/ Lane 16=Positive control (292 bp)/ Lane M=100 bp DNA marker (Fermentas, Canada)
Discussion
In this study, we applied microscopic and molecular detection in order to identify G. duodenalis from fecal samples. In medical la-boratories Giardia diagnosis is based on prepa-ration of wet mount of patient stool and visual recognition by light microscopy. Performing of these methods requires experienced mi-croscopists and medical laboratory technician. In the other hand, these techniques are unable to differentiate between the genetic assem-blages of G. duodenalis isolates and might not be sensitive enough to detect low numbers of excreted Giardia cysts in fecal samples (16, 26, 27). Thus, techniques of molecular detection based on PCR have been developed to detect G. duodenalis cysts in feces. These molecular methods allow the genotyping of G. duodenalis cysts.
For molecular detection in this study, Poly-merase Chain Reaction (PCR) was used to amplify the genes encoding the SSU-rRNA on the microscopically positive Giardia samples. The SSU-rRNA locus has been proven to be highly sensitive as a screening tool for Giardia directly from faces and is the traditional gene sequence used for identification and phyloge-netic analyses (28, 29). The SSU-rRNA se-quence is more conserved compared to the
292 bp
100 X
100 X
500 bp

Rayani et al.: Molecular Identification of Giardia …
Available at: http://ijpa.tums.ac.ir 75
other regions that were used as templates. This could explain why the primers amplifying the SSU-rRNA gene worked better (26). The conserved SSU-rRNA gene is a commonly used marker for the species and assemblage differentiation (mostly genotyping) of Giardia (8). For molecular characterization, we used the glutamate dehydrogenase (gdh) marker that allows distinguishing between the subgroups of the assemblages A and B. The variable gdh locus has been used for successfully genotyp-ing and subtyping the isolates of G. duodenalis from a range of vertebrate hosts (30, 31, 32). An effective DNA extraction method is need-ed to overcome the problems of inadequate yield of extracted DNA and improve molecu-lar detection apart from the sensitive amplifi-cation systems for gene identification.
We had chosen the Phenol-Chloroform pro-tocol for DNA extraction of G. duodenalis from fecal samples with pretreatment using Triton X100. Our results showed that this method was the most efficient DNA extract-ing method for the samples was purified by sucrose gradient. Also, all steps of DNA ex-traction and amplification by PCR were per-formed in a short time. A high percentage of false-negative results may occur in the PCR assay in clinical specimens, foods, and envi-ronmental samples (27). In our experiment, the false negative PCR reaction could have occurred due to some reasons: 1- Parasite load: the low intensity of infection in naturally infected host samples is the problem in using PCR related to DNA extraction from fecal samples (33). 2- Storage of stool specimen: long storage of stool specimens could have caused a false negative PCR reaction (34).
The prevalence of Giardia infection was 10.7% of 1000 patients examined in the area under study. Islamic Republic of Iran is a fa-vorite area for giardiasis. The prevalence of giardiasis varies in different parts of Iran (5%-23%) (22). In a national survey of the preva-lence of intestinal parasitic infections in the community, G. duodenalis was the most com-mon infection (10.9%) (35), which is highly
consistent with the results of other similar study (10.2%) (36). The incidence of G. duode-nalis in the patients referring to hospitals and health centers were reported as 25.8%, 10.1%, and 21.3% by Arani et al. (12), Haghighi et al. (21), and Taherkhani et al. (22), respectively. In all these studies, confirmation of the infec-tion was based on microscopic detection and parasitological techniques. In our study, the prevalence of G. duodenalis among the patients was 10.7% (107/1000) which is in agreement with other researchers' findings in Iran. Over-all, health promotion, public education, im-proving sanitation conditions, and improving clean drinking water and food are important strategies which can be used for control and prevention of the giardiasis infection. Studies performed on DNA extracted directly from fecal samples from various countries confirm that only Giardia assemblages A and B are as-sociated with human infections. The preva-lence of each assemblage varies from country to country and sometimes even within the same country. The reasons behind the geo-graphic variations in the distribution of the G. duodenalis assemblages are still unclear. In our study, DNA sequence showed that the assem-blage A was the most prevalent (80%) geno-type and 20% of the positive samples be-longed to assemblage B in humans living in the area. The predominance of assemblage A in this study is similar to the previous studies in Brazil (37), Iran (38), Italy (39), New Zea-land (40), and Thailand (41). However, it was in contrast to some other studies showing the predominance of assemblage B from Australia (6), Bangladesh (42), Belgium (43), England (44), Nicaragua (45), and Philippine (46). In this study, subtyping analysis showed that all the 32 assemblage A isolates were identical to sub-genotype AII that has also been shown in Australia (6), Brazil (37), Iran (38), Nicaragua (45) and Peru (47).
Although assemblage AII is most likely transmitted from human to human, assem-blage AI is most often responsible for zoono-tic transmission with a wide range of animals

Iranian J Parasitol: Vol. 9, No. 1, Jan -Mar 2014, pp.70-78
76 Available at: http://ijpa.tums.ac.ir
acting as the reservoir hosts. Moreover assem-blage B appears to be largely human specific, it has been reported in some animals and may represent a zoonotic potential. The predomi-nance of assemblage AII and lack of assem-blage AI in our study may be due to the geo-graphical differences among the countries re-garding the prevalence of infection that might be influenced by the regional epidemic or local dynamics of the transmission related to host fauna, infection sources, and socioeconomic factors. Despite, the findings of this study sug-gested anthroponotic transmission from hu-man to human, it is difficult to compare hu-man and animal isolates in this context simply because there was no investigation or study conducted in this area to genotype assemblage A and B from animals isolates thus setting as one of the limitation of the study. Therefore, current result was not enough to demonstrate the role of anthroponotic and zoonotic of gi-ardiasis infections. Of course, further molecu-lar genotypes and epidemiological information about the distribution and prevalence of G. duodenalis assemblages and subtypes within human and animal populations will be attend-ed in future studies. We hope to include the prevalence of assemblage A or B in domestic and pet animals that may serve as a potential reservoir for human in zoonotic giardiasis. Also a thorough more molecular characteriza-tion at multiple loci shall be performed in fu-ture on G. duodenalis isolates ranges from hu-mans, animals and the environment.
Conclusion
G. duodenalis is a flagellated protozoan para-site found in the small intestine of many dif-ferent mammalian species including man and animals. The clinical manifestation of G. duo-denalis infection varies from asymptomatic to acute diarrhea, flatulence, abdominal pain, fa-tigue, and anorexia or chronic disease. G. duo-denalis is one of the most common intestinal parasites in Iran. Important factors, such as socioeconomic, geographical, sanitary and hy-
gienic, cultural, and nutritional factors, con-tribute to the prevalence of the parasite. The present study showed that G. duodenalis sub-assemblage AII was the predominant assem-blage in Fars Province, which indicates an an-throponotic transmission in this population.
Acknowledgments
This is a common project between Shiraz University of Medical Sciences in Iran and University Putra Malaysia. It is a part of the Ph.D. thesis of Mohammad Rayani. We ap-preciate Vice Chancellor of Research, Shiraz University of Medical Sciences, Shiraz, Iran for the collaboration and financial support of this research. The authors declare that there is no conflict of interests.
References
1. Johnston AR, Gillespie, TR, Rwego, IB, McLachlan, TLT, Kent AD, Goldberg TL. Molecular epidemiology of cross-species Giardia duodenalis transmission in western Uganda. PLoS Negl Trop Dis. 2010; 4(5), 683.
2. Adam RD. The biology of Giardia spp. Microbiological Reviews. 1991; 55(4), 706-732.
3. Thompson, R., Monis, P. Variation in Giardia: implications for taxonomy and epidemiology. Adv Parasitol. 2004; 58, 69-137.
4. Volotao A, Costa-Macedo L, Haddad F, Brandao A, Peralta J, Fernandes O. Genotyping of Giardia duodenalis from human and animal samples from Brazil using [beta]-giardin gene: A phylogenetic analysis. Acta Trop. 2007; 102(1), 10-19.
5. Kosuwin R, Putaporntip C, Pattanawong U, Jongwutiwes S. Clonal diversity in Giardia duodenalis isolates from Thailand: Evidences for intragenic recombination and purifying selection at the beta giardin locus. Gene. 2010; 449(1), 1-8.
6. Yang R, Lee J, Ng J, Ryan U. High prevalence Giardia duodenalis assemblage B and potentially zoonotic subtypes in sporadic human cases in Western Australia. Int J Parasitol. 2010; 40(3), 293-297.
7. Gelanew T, Lalle M, Hailu A, Pozio E, Caccio SM. Molecular characterization of human isolates

Rayani et al.: Molecular Identification of Giardia …
Available at: http://ijpa.tums.ac.ir 77
of Giardia duodenalis from Ethiopia. Acta Trop. 2007; 102(2), 92-99.
8. Feng Y, Xiao L. Zoonotic potential and molecular epidemiology of Giardia species and giardiasis. Clin Microbiol Rev. 2011; 24(1), 110-140.
9. Berrilli F, D'Alfonso R, Giangaspero A, Marangi M, Brandonisio O, Kaboré Y, et al. Giardia duodenalis genotypes and Cryptosporidium species in humans and domestic animals in Cote d’Ivoire: occurrence and evidence for environmental contamination. Trans R Soc Trop Med Hyg. 2012; 106(3), 191-195.
10. Read C, Walters J, Robertson ID, Thompson RC. Correlation between genotype of Giardia duodenalis and diarrhoea. Int J Parasitol. 2002; 32(2), 229-31.
11. Tungtrongchitr A, Sookrung N, Indrawattana N, Kwangsi S, Ongrotchanakun J, Chaicumpa W. Giardia intestinalis in Thailand: identification of genotypes. J Health Popul Nutr. 2010; 28(1), 42-52.
12. Arani AS, Alaghehbandan R, Akhlaghi L, Shahi M, Lari AR. Prevalence of intestinal parasites in a population in south of Tehran, Iran. Rev Inst Med Trop Sao Paulo. 2008; 50(3), 145-149.
13. Mohammed Mahdy A, Surin J, Wan K, Mohd-Adnan A, Al-Mekhlafi M, Lim Y. Giardia intestinalis genotypes: Risk factors and correlation with clinical symptoms. Acta Trop. 2009; 112(1), 67-70.
14. Caccio SM, Ryan U. Molecular epidemiology of giardiasis. Mol Biochem Parasitol. 2008; 160(2), 75-80.
15. Wang R, Zhang X, Zhu H, Zhang L, Feng Y, Jian F, et al. Genetic characterizations of Cryptosporidium spp. and Giardia duodenalis in humans in Henan, China. Exp Parasitol. 2011; 127(1), 42-45.
16. Bertrand I, Albertini L, Schwartzbrod J. Comparison of two target genes for detection and genotyping of Giardia lamblia in human feces by PCR and PCR-restriction fragment length polymorphism. J Clin Microbiol. 2005; 43(12), 5940-5944.
17. Sousa M, Morais J, Machado J, Poiares Da Silva J. Genotyping of Giardia lamblia Human Isolates from Portugal by PCR RFLP and Sequencing. J Eukaryot Microbiol. 2006; 53(S1), S174-S176.
18. Thompson RCA. Towards a better unders-tanding of host specificity and the transmission of
Giardia: the impact of molecular epidemiology. In: Olson, B.E., Olson, M.E. and Wallis, P.M., (eds.) Giardia the cosmopolitan parasite. CABI Publishing, Wallingford, 2002. p. 55-69.
19. Sarkari B, Ashrafmansori A, Hatam GR, Mota-zedian MH, Asgari Q, Mohamma-dpour I. Genotyping of Giardia lamblia isolates from human in southern Iran. Trop Biomed. 2012; 29 (3), 366-371.
20. Shoorijeh SJ, Sadjjadi SM, Asheri A, Eraghi K. Giardia Spp. And Sarcocystis Spp. Status In Pet Dogs Of Shiraz, Southern Part Of Iran. Trop Bi-omed. 2008; 25 (2), 154-159.
21. Haghighi A, Khorashad AS, Mojarad EN, Kazemi B, Nejad MR, Rasti S. Frequency of enteric protozoan parasites among patients with gastrointestinal complaints in medical centers of Zahedan, Iran. Trans R Soc Trop Med Hyg. 2009; 103(5), 452-454.
22. Taherkhani H, Shariati S, Abdolahi N, Rosha-ndel G. Clinical manifestations of giardiasis in Iran. J Clin Diagn Res. 2009; 3, 1416-1418.
23. Hopkins RM, Meloni BP, Groth DM, Wether-all JD, Reynoldson JA, Thompson RC. Ribo-somal RNA sequencing reveals differences between the genotypes of Giardia isolates recovered from humans and dogs living in the same locality. J Parasitol. 1997; 44-51.
24. Santín M, Trout JM, Vecino JA, Dubey JP, Fayer R. Cryptosporidium, Giardia and Enter-ocytozoon bieneusi in cats from Bogota (Colombia) and genotyping of isolates. Vet Parasitol. 2006; 141(3), 334-339.
25. Read CM, Monis PT, Thompson RC. Discrimination of all genotypes of Giardia duodenalis at the glutamate dehydrogenase locus using PCR-RFLP. Infect Genet Evol. 2004; 4(2), 125-130.
26. Nantavisai K, Mungthin M, Tan-ariya P, Ran-gsin R, Naaglor T, Leelayoova S. Evaluation of the sensitivities of DNA extraction and PCR methods for detection of Giardia duodenalis in stool specimens. J Clin Microbiol. 2007; 45(2), 581-583.
27. Orlandi PA, Lampel KA. Extraction-free, filter-based template preparation for rapid and sensitive PCR detection of pathogenic parasitic protozoa. J Clin Microbiol. 2000; 38(6), 2271-2277.
28. Traub RJ, Monis PT, Robertson I, Irwin P, Mencke N, Thompson RC. Epidemiological and molecular evidence supports the zoonotic transmission of Giardia among humans and dogs

Iranian J Parasitol: Vol. 9, No. 1, Jan -Mar 2014, pp.70-78
78 Available at: http://ijpa.tums.ac.ir
living in the same community. Parasitology. 2004; 128(03), 253-262.
29. Wielinga CM, Thompson RC. Comparative evaluation of Giardia duodenalis sequence data. Parasitology. 2007; 134(12), 1795-821.
30. Ey PL, Mansouri M, Kulda J, Nohýnková E, Monis PT, Andrews RH, et al. Genetic Analysis of Giardia from Hoofed Farm Animals Reveals Artiodactyl-Specific and Potentially Zoonotic Genotypes. J Eukaryot Microbiol. 1997; 44(6), 626-635.
31. Monis PT, Mayrhofer G, Andrews RH, Homan WL, Limper L, Ey PL. Molecular genetic analysis of Giardia intestinalis isolates at the glutamate dehydrogenase locus. Parasitology. 1996; 112, 1-12.
32. Monis PT, Andrews RH, Mayrhofer G, Mackrill J, Kulda J, Isaac-Renton JL, et al. Novel lineages of Giardia intestinalis identified by genetic analysis of organisms isolated from dogs in Australia. Parasitology. 1998; 116(1), 7-19.
33. Bajer A. Cryptosporidium and Giardia spp. Infect-ions in humans, animals and the environment in Poland. Parasitol Res. 2008; 104(1), 1-17.
34. Boontanom P, Siripattanapipong S, Mungthin M, Tan-ariya P, Leelayoova S. Improved sensitivity of PCR amplification of glutamate dehydrogenase gene for detection and genotyping of Giardia duodenalis in stool specimen. Southeast Asian J Trop Med Public Health. 2010; 41(2), 280-284
35. Sayyari AA, Imanzadeh F, Bagheri Yazdi SA, Karami H, Yaghoobi M. Prevalence of intestinal parasitic infections in the Islamic Republic of Iran. East Mediterr Health J. 2005; 11(3). 377-83.
36. Kia E, Hosseini M, Nilforoushan M, Meamar A, Rezaeian M. Study of intestinal protozoan parasites in rural inhabitants of Mazandaran province, Northern Iran. Iran J Parasitol. . 2008; 3(1), 21-25.
37. Souza SL, Gennari SM, Richtzenhain LJ, Pena HF, Funada MR, Cortez A, et al. Molecular identification of Giardia duodenalis isolates from humans, dogs, cats and cattle from the state of Sao Paulo, Brazil, by sequence analysis of fragments of glutamate dehydrogenase (gdh) coding gene. Vet Parasitol. 2007; 149(3-4), 258-264.
38. Babaei Z, Oormazdi H, Akhlaghi L, Rezaie S, Razmjou E, Soltani-Arabshahi S, et al. Molecular
characterization of the Iranian isolates of Giardia lamblia: application of the glutamate dehydro-genase gene. Iranian J Publ Health. 2008; 37(2), 75-82.
39. Caccio SM, De Giacomo M, Pozio E. Sequence analysis of the beta-giardin gene and develo-pment of a polymerase chain reaction-restriction fragment length polymorphism assay to geno-type Giardia duodenalis cysts from human faecal samples. Int J Parasitol. 2002; 32(8), 1023-1030.
40. Winkworth CL, Learmonth JJ, Matthaei CD, Townsend CR. Molecular characterization of Giardia isolates from calves and humans in a region in which dairy farming has recently intensified. Appl Environ Microbiol. 2008; 74(16), 5100-5105.
41. Traub RJ., Inpankaew T, Reid SA, Sutthikornchai C, Sukthana Y, Robertson ID, et al. Transmission cycles of Giardia duodenalis in dogs and humans in Temple communities in Bangkok-a critical evalu-ation of its prevalence using three diagnostic tests in the field in the absence of a gold standard. Acta Trop. 2009; 111(2), 125-132.
42. Haque R, Roy S, Kabir M, Stroup SE, Mondal D, Houpt ER. Giardia assemblage A infection and diarrhea in Bangladesh. J Infect Dis . 2005; 192(12), 2171-2173.
43. Geurden T, Levecke B, Cacció SM, Visser A, De Groote G, Casaert S, et al. Multilocus genotyping of Cryptosporidium and Giardia in non-outbreak related cases of diarrhoea in human patients in Belgium. Parasitology. 2009; 136(10), 1161-1168.
44. Breathnach AS, McHugh TD, Butcher PD. Prevalence and clinical correlations of genetic subtypes of Giardia lamblia in an urban setting. Epidemiol Infect. 2010; 138(10), 1459-1467.
45. Lebbad M, Ankarklev J, Tellez A, Leiv-a B, Andersson JO, Svard S. Dominance of Giardia assemblage B in Leon, Nicaragua. Acta Trop. 2008; 106(1), 44-53.
46. Yason JA, Rivera WL. Genotyping of Giardia duodenalis isolates among residents of slum area in Manila, Philippines. Parasitol Res. 2007; 101(3), 681-687.
47. Cooper MA, Sterling CR, Gilman RH, Cama V, Ortega Y, Adam RD. Molecular analysis of household transmission of Giardia lamblia in a region of high endemicity in Peru. J Infect Dis . 2010; 202(11), 1713-1721.