molecular and biochemical characterization of trichoderma isolates inhibiting a phytopathogenic...
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Molecular and biochemical characterization of Trichoderma isolatesinhibiting a phytopathogenic fungi Aspergillus niger Van Tieghem
H.P. Gajera*, D.N. VakhariaDepartment of Biotechnology, College of Agriculture, Junagadh Agricultural University, Junagadh-362 001, Gujarat, India
a r t i c l e i n f o
Article history:Accepted 7 April 2010
Keywords:Trichoderma isolatesAspergillus nigerAntagonismCell wall degrading enzymesMolecular markers
a b s t r a c t
Random Amplified Polymorphic DNA (RAPD) was used to examine the genetic variability among twelveisolates of Trichoderma representing three species and their ability to antagonize Aspergillus niger VanTieghem causing collar rot in peanut using dual culture assay for correlation among RAPD products andtheir hardness to A. niger. One hundred and three of the 108 bands, using random decamer fungalprimers, were polymorphic with an average frequency of 11.4 bands. The calculated PolymorphismInformation Content (PIC) values for RAPD markers ranged from 0.172 to 0.401 and RAPD primer index(RPI) ranged from 0.99 to 6.01. RPI showed that RFu C-5 gave best results of polymorphism among theprimer used in the experiment. RAPD analysis showed 10 marker loci for diagnosis of Trichoderma viride60 and/or Trichoderma harzianum 2J, first two highest inhibitory acting antagonists. A UPGMA dendro-gram constructed on the basis of Jaccard’s similarity coefficient using NTSYS 2.2 program which illus-trated two distinct clusters of 12 isolates of Trichoderma and A. niger pathogen, and shared only 19%similarity. However, the in vitro highest A. niger growth inhibitory Trichoderma isolates e T. viride 60(86.2%) and T. harzianum 2J (80.4%) were in same out group and shared 63% similarity. Relationship wasfound between the polymorphism showed by the Trichoderma isolates and their hardness to A. niger, interms of in vitro production of cell wall degrading enzymes- chitinase, b-1,3 glucanase and protease,during antagonism.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Aspergillus niger Van Tieghem causes a disease called black moldon certain fruits and vegetables such as grapes, onions, and peanut,and is a common contaminant of food. Some strains of A. niger havebeen reported to produce potentmycotoxins called ochratoxins [1].The A. niger causing collar rot disease on peanut seedlings was firstreported by Jochem [2]. A. nigermay cause an average 5 per cent lossin yield but in some parts itmay cause as high as 40 per cent losses inPeanut. Collar rot is more serious problem in sandy soil [3,4]. Themeasures preconized for its control, such as rotation of crops, use ofresistant varieties and treatment of seeds and/or soil with fungicides,a lot of times become unsuitable or not effective, mainly due to thegenetic variability presented by the pathogen to the hosts range,capacity to survive in the soil and in the seeds, and physiologicflexibility to infect different hosts [5].
The use of antagonistic microorganisms to A. niger hasbeen investigated as an alternative control method, species ofTrichoderma, applied as treatment of seeds or soil, have been
demonstrated to control the pathogen in a variety of cultures inthe greenhouse and field studies [6e9]. Knowledge concerningthe behavior of these fungi as antagonists is essential for theireffective use since they can act against target organisms in severalways. Species of Trichoderma can produce extracellular enzymesand antifungal antibiotics, but they may also be competitors tofungal pathogens, promote plant growth, and induce resistance inplants [8]. The commercial use of Trichoderma must be precededby precise identification, adequate formulation, and studies aboutthe synergistic effects of their mechanism of bio-control [10]. Theability of Trichoderma to control A. niger varies considerably, and itis possible to improve its biological control efficiency by theselection of isolates with high antagonistic potential and adaptedto certain ecological or geographical areas [11].
Fujimori and Okuda [12] examined 74 strains of Trichoderma byRAPD profiles and the results were consistent with the morpho-logical, physiological and ecological data of these strains, whatsuggests that the technique can aid to eliminate strains duplicatedin a program for microbial selection. Using RAPD, Schlick et al. [13]analyzed strains of Trichoderma harzianum andmutants induced bygamma radiation originated from one wild isolate, verifying thatwith RAPD it was possible to differentiate all themutants strains for
* Corresponding author.E-mail address: [email protected] (H.P. Gajera).
Contents lists available at ScienceDirect
Physiological and Molecular Plant Pathology
journal homepage: www.elsevier .com/locate/pmpp
0885-5765/$ e see front matter � 2010 Elsevier Ltd. All rights reserved.doi:10.1016/j.pmpp.2010.04.005
Physiological and Molecular Plant Pathology 74 (2010) 274e282

Author's personal copy
at least one primer and concluding that the method was valuablefor identification and fast differentiation of strains.
Molecular markers are important to develop tools that monitorthe genetic and environmental fate of bio-control agents-Trichodermaas more fungal bio-control agents are registered as alternativesto chemical pesticides. Molecular characterization provides animmense source of data that can assist to the scientists in the study ofidentity, relatedness, diversity and selection of proper candidates forbiological control.
Our goal was to document the genetic variability of twelveisolates of three Trichoderma species (Trichoderma virens, Tricho-derma viride, T. harzianum) using the RAPD technique and theirantagonistic potential against A. niger. Our observations provideinformation about the relationship between antagonistic capacityof Trichoderma isolates, production of cell wall degrading enzymesduring antagonism and RAPD markers.
2. Materials and methods
2.1. Collection and isolation of the fungus
Twelve isolates of Trichoderma (six of T. harzianum, five ofT. viride, one of T. virens) were isolated by serial dilution techniquefrom different soil rhizosphere of Saurashtra region (Gujarat, India)and sent for species identification at IARI, New Delhi. Peanutseedlings which showed typical symptoms of collar rot were cutinto small bits with the help of sterilized blade and the pure cultureof pathogen (A. niger) wasmade by hyphal tip isolationmethod [14]on the solidified PDA medium in petri plates. All microbes weremaintained throughout the study by periodical transfers on PDAmedium under aseptic condition to keep the culture fresh andviable.
2.2. Fungal growth condition and DNA extraction
Cultures were maintained on PDA at 28 �C � 2 �C were grown inpotato dextrose broth (PDB) for 48 h. Hyphae were collected onfilter paper in a Buchner funnel, washed with distilled water,frozen, and were used for DNA extraction. Total genomic DNA wasextracted from the acetone dried mycelium by a modified sodiumdodisylsulphate (SDS) method [15]. The genomic DNAwas stored in50 ml TE buffer at �20 �C for further use.
2.3. Random amplified polymorphic DNA (RAPD)
The Polymerase chain reactions (PCR) were carried out ina 200 ml thin walled PCR tubes. The PCR for RAPD were carried outin a 25 ml of reactionmixture as described by Abbasi et al. [16]. A setof 10 RAPD fungal primers was tested across twelve Trichodermaisolates and pathogen A. niger. RAPD primers and PCR relatedchemicals were obtained from Bangalore genei, India. Amplifica-tion reactions were carried out in a total volume of 25 ml containing,10� PCR buffer with 1.5 mM MgCl2 and 10 mM Tris, 2.25 U Taqpolymerase, 0.20 mM each dNTPs, 1 mM primer and 50 ng templateDNA. Amplification was carried out on Bio-Rad Thermal-Mycyclerwith the following programme as described by Shalini et al. [17].The samples were initially heated to 94 �C for 5 min and thensubjected to 45 cycles of denaturation at 94 �C for 1 min, annealingat 37 �C for 1 min and 30 s, and extension at 72 �C for 2 min fol-lowed by a final extension for 10 min. After the process, samples of15 ml of the amplification products were assayed by electrophoresisin 1.5% agarose gel containing ethidium bromide, running with TBEbuffer. The electrophoresis was carried out at 80 V (constant) forabout 100 min using Bio-Rad submerge gel electrophoresis system.The gel was viewed under UV trans-illuminator for visualizing
separated bands, and photographed. The low range ruler (100 bp to3 kb) was used as molecular weight size markers.
Amplifications were repeated once more for each RAPD primerand only consistent bands were considered for scoring. The NTSYS.PC (Numerical Taxonomy System Applied Biostatistics, Setauket,New York) system version 2.2 by Exeter Software was used for dataanalysis [18]. The data (band presence or absence) were introducedin the form of a binary matrix and a pair wise similarity matrix wasconstructed using the Jaccard’s coefficient. The SIMQUALK pro-gramme was used to calculate Jaccard’s similarity coefficient anda graphical phenogram (dendrogram) of the genetic relatednessamong the different isolates was produced by means of theunweighted pair group method with arithmetic average (UPGMA)analysis [19]. Size of specific bands of DNA was determined usingsoftware Alphaimager 2200 manufactured by Alpha Ease FC, USA.
A polymorphic information index (PIC) for RAPD profile wascalculated as PIC ¼ 1 � p2 � q2, where, p is band frequency and q isno band frequency [20]. PIC values were than used to calculatea RAPD primer index (RPI), which was generated by adding up thePIC values of all the markers amplified by the same primer.
2.4. Antagonistic activity
The dual culture technique described by Dennis and Webster[21] was used to test the antagonistic ability of Trichoderma againstA. niger. The test fungus and twelve Trichoderma isolates weregrown on PDA for a week at 28 � 2 �C in incubator. Disk of 5 mm ofthe target fungus (A. niger) cut from the periphery were transferredto the petri plate with PDA. Trichoderma was also transferredaseptically in the same plate. Each plate received two disks, onewith Trichoderma mycelium and another with A. niger, placed ata distance of 7 cm away from each other. The experiment wasconducted in five replications for each antagonist. The plates wereincubated at 28� 2 �C temperatures and observed after six days forgrowth of antagonist and test fungus. Index of antagonism as percent growth inhibition of A. niger was determined by following themethod of Watanabe [22].
2.5. Extraction of cell wall degrading enzymesfrom antagonistic petri plates
Cell wall degrading enzymes was extracted from petri platescontaining both fungal pathogen and Trichoderma along with thetest fungus alone as a control (C) at 6 days after inoculation (DAI).For that, 25 ml of 100 mM phosphate buffer (pH-5.5) containing50 mM sodium chloride was added to each petri plate and trans-ferred mycelia mat to conical flask. For enzyme induction, 1% ofeither carboxymethyl cellulose (CMC), sodium polypectate, chitin,laminarin or casein was added into culture medium of conical flaskand pH adjusted to 5.5 [23,24]. Cultures were then shaken well inorbital shaker at 120 rpm at 28 �C for about 6 h [25]. After that,it was transferred to 50 ml centrifuge tubes and centrifuged at14 000 rpm for 10 min. Supernatant was collected and stored at�20 �C until it used for assay of enzymes activities (cellulase, polygalacturonase (PG), chitinase, b-1, 3 glucanase, protease). Themethod of FolineLowry [26] was used to estimate the proteincontent in culture supernatant and used for calculating specificactivity of the enzyme.
2.6. Enzymes assays
Cellulase activity (EC 3.2.1.21) was determined bymeasuring theamount glucose formed from CMC [27]. Reactions were conductedfor 15min at 55 �C [28]. Poly galacturonase (PG) activity (EC 3.2.1.15)was measured at 37 �C using sodium polypectate as the substrate
H.P. Gajera, D.N. Vakharia / Physiological and Molecular Plant Pathology 74 (2010) 274e282 275

Author's personal copy
[28]. The galacturonic acid released was measured by the DNSAmethod [27]. Chitinase activity (EC 3.2.1.14) was assayed at 50 �C[29] monitoring the amount of sugar N-acetylglucosamine fromacetylated chitin, measured by DMABmethod [30]. b-1, 3 glucanaseactivity (EC 3.2.1.39) was assayed on the release of glucose fromlaminarin by DNSAmethod [27] Reactions were carried out at 37 �Cfor 10 min [31].
Specific activity of cellulase, PG, chitinase and b-1,3 glucanasewere expressed as Unit mg�1 protein. However, Unit activity wasdefined as the amount of enzyme necessary to produce one mM ofcorresponding reducing sugar in 1 min of one ml culture super-natants. Non enzymatic controls were also performed using boiledenzymes and were subtracted from the enzymatic values.
Protease activity (EC 3.4.21.4) was measured using casein assubstrate as described by Malik and Singh [32]. Blank was treatedas zero time incubation. The amount of released total free aminoacids was estimated by ninhydrin method [33]. Specific activity ofprotease was expressed as Unit mg�1protein and one unit ofprotease activity was defined as the amount of protein necessaryto produce mg free amino acids in 1 min of one ml culturesupernatant.
3. Results
3.1. Primer selection
Amplified products were observed when the genomic DNA ofmicrobes (Trichoderma isolates and A. niger) was subjected to RAPDanalysis using 10 randomdecamer primers. These 10 random fungalprimers were obtained from Bangalore Genei, India. Initially,10 primers were examined and of these, one primer (RFu C-8) failedto give any amplified products of DNA. Possibly this may be due toabsence of complementary sequence in the genomic DNA. Thus, 9out of 10 fungal primers were selected for evaluating moleculardifferences existing in 12 isolates of Trichodermawhich belongs to 3species for inhibiting the growth of fungal pathogen A. niger.However, molecular characterization of pathogen A. niger alsocarried out to identify genetic variation and diversity, and comparedit with various isolates of Trichoderma. The accession number ofeach primer is given in Table 1.
3.2. DNA polymorphism analysis of the Trichodermaisolates and A. niger
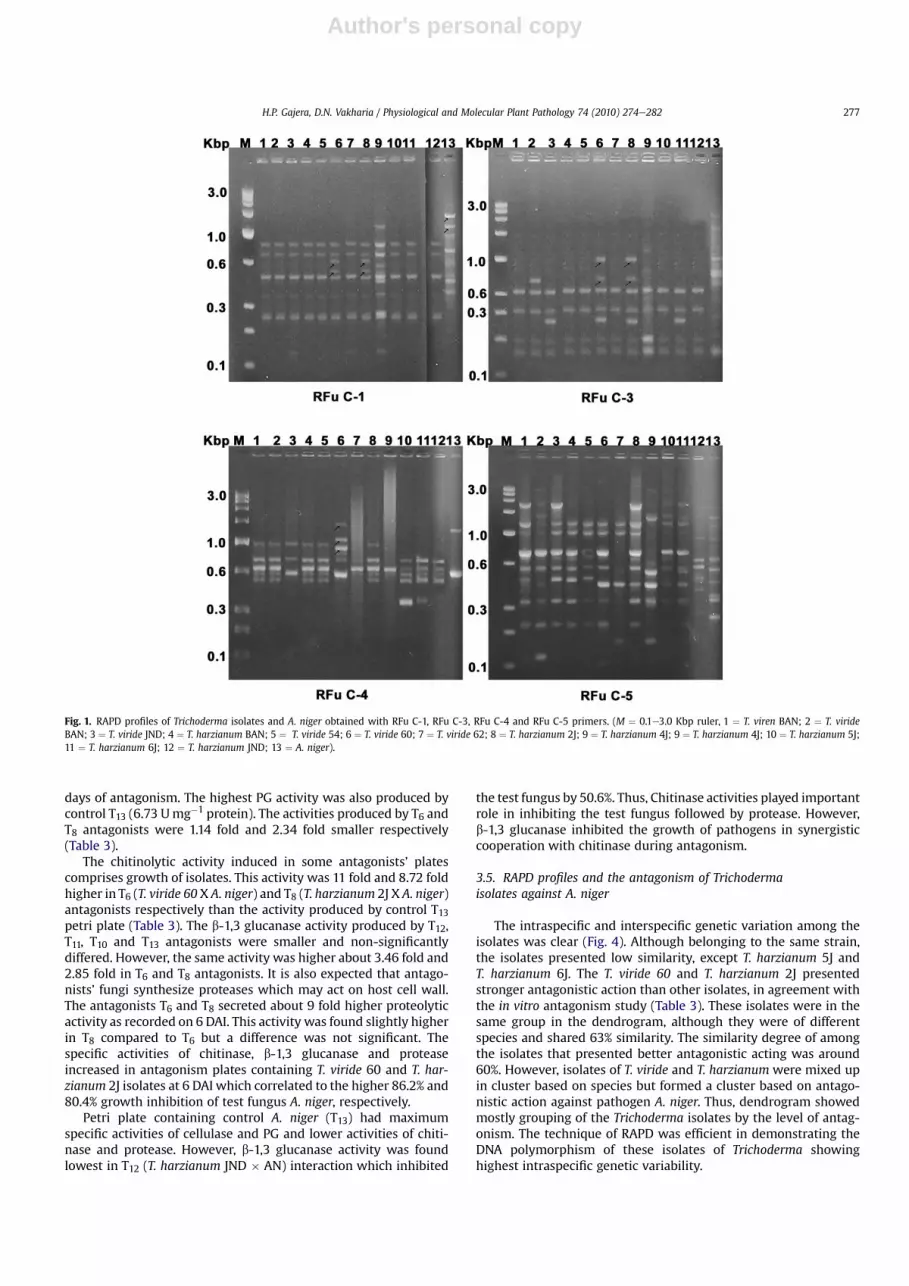
A total 108 bands were produced by 9 RAPD primers with anaverage frequency of 12 bands per primer (Figs. 1 and 2). Total 103polymorphic bands were evident out of which 66 were poly-morphic and shared between at least two individuals, and 37 bands
were polymorphic and unique while 5 bands were monomorphic(Table 1). Four (RFu C-1, RFu C-3, RFu C-4, RFu C-6) of the 9 RAPDprimers produced monomorphic profiles. Primer RFu C-6 gener-ated the maximum 19 bands, whereas RFu C-9 generated thelowest with 5 bands. The per cent polymorphism furnished by eachprimer ranged between 84.6 and 100 (Table 1). The calculated PICvalues for RAPDmarkers were ranged from 0.172 to 0.401 and RAPDprimer index (RPI) ranged from 0.99 to 6.01 (Table 1). The lowestPIC and RPI values obtained by RFu C-2 and highest was with RFuC-5. Thus, RPI showed that RFu C-5 (Fig. 1) gave best results amongthe primer used here.
Dendrogram was constructed using the similarity matrix. Thesimilarity coefficient ranged from 0.19 to 0.80 (Fig. 4). The dendro-gram obtained indicates that there was a major cluster consisting of12 isolates of Trichoderma of total 13 microbes, whereas one fungalpathogen (A. niger) were found to be different from the rest of themicrobese Trichoderma. Themajor cluster A and cluster B consistedof 12 isolates of Trichoderma and 1pathogen A. niger and shared19%similarity (Fig. 4). Total 5 out of 9 primers generated 8 uniquemarker loci for A. niger. Primer RFu C-2 amplified DNA of pathogenA. niger only and produced two highly dense unique markers (1100,786 bp)whichmay be useful to identify pathogen from Trichodermaisolates. However, primer RFu C-1 generated 1675 and 1381 bpRAPD markers from DNA of A. niger. Similarly, RFu C-10 associatedwith 1023 and 565 bp marker for A. niger. RAPD markers linked toantagonism of 12 isolates of Trichodermawith pathogen A. nigerwastabulated in Table 2.
3.3. In vitro growth inhibition of A. niger by Trichoderma isolates
Per cent growth inhibition of pathogen (A. niger) was signifi-cantly higher in T6 (86.2%) antagonist followed by T8 (80.4%), T3(74.3%), T2 (71.9%), T1 (60.9%) and T12 (50.6%) at 6 DAI (Table 3).However, other antagonists were recorded below 30% growthinhibition of fungal pathogen. Thus, it was observed that T6antagonist (i.e., interaction between T. viride 60 and pathogenA. niger) have a better growth of inhibition of test fungus A. nigercompared to other bio-control agents (Fig. 3).
3.4. Production of cell wall degrading enzymes during antagonism
Twelve Trichoderma isolates, tested for antagonism with fungalpathogen, produced and secreted on induction substantial amountsof various cell wall degrading enzymes in comparison to control T13(A. niger alone). Maximal specific activity (2.64 U mg�1 protein) ofcellulase was produced by control T13 (A. niger alone). This activitywas 2.43 fold and 1.36 fold higher than those determined for T6(T. viride 60 X A. niger) and T8 (T. harzianum 2J X A. niger) during 6
Table 1Polymorphism obtained with different RAPD primers generated from twelve isolates of Trichoderma and pathogen A. niger.
Sr. No. Name ofprimer
Accession no. Polymorphic bands Mono morph.bands
Total bands Polymor.(%)
PIC RPI value
S U T
1 RFu C-1 AM 911695 9 4 13 1 14 92.8 0.189 2.652 RFu C-2 AM 911696 0 7 7 0 7 100.0 0.172 0.993 RFu C-3 AM 911697 7 4 11 2 13 84.6 0.198 2.584 RFu C-4 AM 773320 5 4 09 1 10 90.0 0.251 2.515 RFu C-5 AM 911698 15 0 15 0 15 100.0 0.401 6.016 RFu C-6 AM 765822 10 8 18 1 19 94.7 0.260 4.807 RFu C-7 AM 911699 6 3 9 0 9 100.0 0.260 2.348 RFu C-8 AM 773321 e e e e e e e e
9 RFu C-9 AM 773779 4 1 05 0 5 100.0 0.265 1.3310 RFu C-10 AM 765832 10 6 16 0 16 100.0 0.281 4.50Total 66 37 103 5 108 95.4
S ¼ Shared; U ¼ Unique; T ¼ Total Polymorphic Bands; PIC ¼ Polymorphism Information Content; RPI ¼ RAPD Primer Index.
H.P. Gajera, D.N. Vakharia / Physiological and Molecular Plant Pathology 74 (2010) 274e282276
Author's personal copy
days of antagonism. The highest PG activity was also produced bycontrol T13 (6.73 Umg�1 protein). The activities produced by T6 andT8 antagonists were 1.14 fold and 2.34 fold smaller respectively(Table 3).
The chitinolytic activity induced in some antagonists’ platescomprises growth of isolates. This activity was 11 fold and 8.72 foldhigher in T6 (T. viride 60 X A. niger) and T8 (T. harzianum 2J X A. niger)antagonists respectively than the activity produced by control T13petri plate (Table 3). The b-1,3 glucanase activity produced by T12,T11, T10 and T13 antagonists were smaller and non-significantlydiffered. However, the same activity was higher about 3.46 fold and2.85 fold in T6 and T8 antagonists. It is also expected that antago-nists’ fungi synthesize proteases which may act on host cell wall.The antagonists T6 and T8 secreted about 9 fold higher proteolyticactivity as recorded on 6 DAI. This activity was found slightly higherin T8 compared to T6 but a difference was not significant. Thespecific activities of chitinase, b-1,3 glucanase and proteaseincreased in antagonism plates containing T. viride 60 and T. har-zianum 2J isolates at 6 DAI which correlated to the higher 86.2% and80.4% growth inhibition of test fungus A. niger, respectively.
Petri plate containing control A. niger (T13) had maximumspecific activities of cellulase and PG and lower activities of chiti-nase and protease. However, b-1,3 glucanase activity was foundlowest in T12 (T. harzianum JND � AN) interaction which inhibited
the test fungus by 50.6%. Thus, Chitinase activities played importantrole in inhibiting the test fungus followed by protease. However,b-1,3 glucanase inhibited the growth of pathogens in synergisticcooperation with chitinase during antagonism.
3.5. RAPD profiles and the antagonism of Trichodermaisolates against A. niger
The intraspecific and interspecific genetic variation among theisolates was clear (Fig. 4). Although belonging to the same strain,the isolates presented low similarity, except T. harzianum 5J andT. harzianum 6J. The T. viride 60 and T. harzianum 2J presentedstronger antagonistic action than other isolates, in agreement withthe in vitro antagonism study (Table 3). These isolates were in thesame group in the dendrogram, although they were of differentspecies and shared 63% similarity. The similarity degree of amongthe isolates that presented better antagonistic acting was around60%. However, isolates of T. viride and T. harzianum were mixed upin cluster based on species but formed a cluster based on antago-nistic action against pathogen A. niger. Thus, dendrogram showedmostly grouping of the Trichoderma isolates by the level of antag-onism. The technique of RAPD was efficient in demonstrating theDNA polymorphism of these isolates of Trichoderma showinghighest intraspecific genetic variability.
Fig. 1. RAPD profiles of Trichoderma isolates and A. niger obtained with RFu C-1, RFu C-3, RFu C-4 and RFu C-5 primers. (M ¼ 0.1e3.0 Kbp ruler, 1 ¼ T. viren BAN; 2 ¼ T. virideBAN; 3 ¼ T. viride JND; 4 ¼ T. harzianum BAN; 5 ¼ T. viride 54; 6 ¼ T. viride 60; 7 ¼ T. viride 62; 8 ¼ T. harzianum 2J; 9 ¼ T. harzianum 4J; 9 ¼ T. harzianum 4J; 10 ¼ T. harzianum 5J;11 ¼ T. harzianum 6J; 12 ¼ T. harzianum JND; 13 ¼ A. niger).
H.P. Gajera, D.N. Vakharia / Physiological and Molecular Plant Pathology 74 (2010) 274e282 277

Author's personal copy
Fig. 2. RAPD profiles of Trichoderma isolates and A. niger obtained with RFu C-6, RFu C-7, RFu C-9 and RFu C-10 primers. (M ¼ 0.1e3.0 Kbp ruler, 1 ¼ T. viren BAN; 2 ¼ T. virideBAN; 3 ¼ T. viride JND; 4 ¼ T. harzianum BAN; 5 ¼ T. viride 54; 6 ¼ T. viride 60; 7 ¼ T. viride 62; 8 ¼ T. harzianum 2J; 9 ¼ T. harzianum 4J; 9 ¼ T. harzianum 4J; 10 ¼ T. harzianum 5J;11 ¼ T. harzianum 6J; 12 ¼ T. harzianum JND; 13 ¼ A. niger).
Table 2RAPD markers associated with characterization of twelve isolates of three Trichoderma species and pathogen A. niger.
Sr. No. Name ofPrimer
Molecular markers (bp)
Trichoderma isolates AN
1 2 3 4 5 6 7 8 9 10 11 12 13
1. RFu C-1 e e e e e 712580
e 712580
1381484
e e e 16751381484
2. RFu C-2 e e e e e e e e e e e e 1100786
3. RFu C-3 e 670 305 e e 1015703305
e 1015703305
e e 305 e 747
4. RFu C-4 e e e e e 15281100916
e e e 296 296 e 1340
5. RFu C-5 1899 114 1899 e e e 151 1833 151 e e 688 6886. RFu C-6 1032 e e e e 2002
1789681
1032 1032 1032 e 1032 e
7. RFu C-7 639 929 e 735 e 951 e 908780
e e 185 e e
8. RFu C-9 e 159 e e e 427 e 427 e e e e e
9. RFu C-10 e 980 899 e e 879254
e e e e e 16981115367
1023565
Total No. 3 5 3 1 e 15 1 10 4 2 3 5 10
1. T. virens BAN; 2. T. viride BAN; 3. T. viride JND; 4. T. Harzianum BAN; 5. T. viride 54; 6. T. viride 60; 7. T. viride 62; 8. T. harzianum 2J; 9. T. harzianum 4J; 10. T. harzianum 5J;11. T. harzianum 6J; 12. T. harzianum JND; 13. A. niger (AN).

Author's personal copy
The highest (86.2%) fungal growth inhibition (A. niger) wasachieved by T. viride 60 during in vitro antagonism followed byT. harzianum 2J (80.4%). RAPD markers linked to this trait werenoticed with some fungal primers. Primer RFu C-4 gave threeamplified products (1528, 1100 and 916 bp) for T. viride 60 whichwere not yielded by any other Trichoderma Isolates. Similarly, RFuC-6 detected 2002, 1789 and 681 bp markers associated withT. viride 60. However, T. viride 60 and T. harzianum 2J (first twohigherinhibitory acting antagonists) generated common RAPD markers(Table 2). These were 712 and 580 bp by primer RFu C-1, and 1015and 703 bp by RFu C-3. Interestingly, these higher inhibitory actingantagonists also elevated production of cell wall degrading enzymeslike chitinase, b-1,3 glucanase and protease in the culture mediumof antagonism. So, the markers loci for T. viride 60 and T. harzianum2J may have relationship with the production of cell wall degradingenzymes (chitinase, b-1,3 glucanase and protease).
4. Discussion
Antagonism of 12 isolates of 3 Trichoderma species (T. virens,T. viride and T. harzianum) with A. niger Van Tieghem, causing collarrot in peanut, during in vitro interaction indicated maximum(86.2%) growth inhibition of test pathogen with T. viride 60 fol-lowed by T. harzianum 2J (80.4%) at 6 DAI (Table 3, Fig. 3).
The antagonistic effect of Trichoderma spp. and Pseudomonasfluorescence against isolates of Fusarium oxysporum f. sp. carthamithat causes wilt disease in sunflower was studied by Prameala et al.[34]. Among three antagonists tested, T. viridewas found to be moreeffective than T. harzianum and P. fluorescens which confirm thepresent experimental results that T. viride was the best antagonistthanT. harzianum and T. virens. Seventy Trichoderma isolates collectedfrom different regions of Morocco were tested for their capacity toinhibit in vitromycelial growth of Sclerotium rolfsii [35]. Four of theseisolates (Nz, Kb2, Kb3 and Kf1) showed good antagonistic activityagainst S. rolfsii and were also highly competitive in natural soil.These isolates would therefore be candidates for development inbiological control. However, Shalini and Kotasthane [36] screenedseventeen Trichoderma strains against Rhizoctonia solani in vitro. Allstrains including T. harzianum, T. viride and Trichoderma aureoviridewere more or less inhibited the growth of R. solani.
It was observed that when T. viride 60 (T6) interact with A. nigerand in vitro antagonism revealed the highest production of cell wall
degrading enzymes- chitinase and b-1,3-glucanase (Table 3), inaddition to protease followed by T. harzianum 2J and other subse-quent antagonists of various isolates of Trichoderma. However,levels of cellulase and PG were found to be the lowest in T6antagonist and it was recorded highest in control e T1 (A. niger).Pectinase and cellulase had broken down pectin and cellulose, thetwo major polymers that maintain the firmness and structure ofhost cell walls. Production of cellulase and PG determined thepathogenicity of necrotrophic pathogen [37].
There are several mechanisms involved in Trichoderma antago-nism namely antibiosis whereby the antagonic fungus showsproduction of antibiotics, competition for nutrients. In case ofmycoparasitism, Trichoderma directly attacks the plant pathogen byexcreting lytic enzymes such as chitinases, b-1,3 glucanases andproteases [38]. Because of the skeleton of pathogenic fungi cellwalls contains chitin, glucan and proteins, enzymes that hydrolyzethese components have to be present in a successful antagonist inorder to play a significant role in cell wall lysis of the pathogen[39,40].
In present study, percent growth inhibition of test fungus and cellwall degrading enzymese chitinases, b-1,3-glucanase and proteasein the culture medium of antagonist treatment established a rela-tionship to inhibit growth of fungal pathogen by increasing thelevels of these lytic enzymes (Table 3, Fig. 3).
Most of the bio-controlagents are known to produce chitinaseand b-1,3-glucanases enzymes which could degrade the cell wallleading to the lysis of hyphae of the pathogen [41]. The pathogencell wall and chitin induce nag1 gene, but it is only triggered whenthere is contact with the pathogen [40,42,43]. Chit36 inhibited theBotrytis cinerea spore germination and the growth of both S. rolfsiiand F. oxysporum [44]. Other genes homologous to chit36 have beencloned from T. harzianum TM, Trichoderma atroviride P1 and Tri-choderma asperellum Te203 [44].
Present findings showed higher specific activity of enzymes-chitinases, b-1,3 glucanase and protease during antagonism ofsome T. isolates with fungal plant pathogen. Activity of theseenzymes varied with various isolates of Trichoderma species. Thismay be due to the expression of certain gene in Trichoderma spp.during antagonism as Chit33 is expressed only during the contactphase and not before overgrowing R. solani [45]. However, chit36Ydoes not need the direct contact of the pathogen to be expressed.Chit33, chit42 and chit36 have been over expressed in Trichoderma
Table 3Specific activity of cell wall degrading enzymes during in-vitro antagonism of Trichoderma isolates with A. niger at 6 days after inoculation (DAI).
No. Antagonists % Inhibitionof A. niger
Cell wall degrading enzymes (U mg�1protein)
Cellulase Poly galacturonase Chitinase b-1,3 Glucanase Protease
1 2 3 4a 5a 6a 7a 8a
T1 T. virens BAN X AN 60.9 1.59 3.93 0.72 1.87 3.21T2 T. viride BAN X AN 71.9 1.47 3.64 0.70 2.15 3.68T3 T. viride JND X AN 74.3 1.50 3.60 0.90 1.95 3.43T4 T. harzianum BAN X AN 26.1 2.26 4.81 0.35 1.78 2.63T5 T. viride 54 X AN 42.4 1.96 5.89 0.44 1.72 2.75T6 T. viride 60 X AN 86.3 0.93 3.14 1.31 3.48 5.24T7 T. viride 62 X AN 23.7 1.90 5.64 0.30 1.09 1.57T8 T. harzianum 2J X AN 81.6 1.35 2.01 1.07 3.01 5.63T9 T. harzianum 4J X AN 32.1 2.71 5.31 0.44 1.64 3.04T10 T. harzianum 5J X AN 40.2 2.33 6.19 0.41 0.42 2.50T11 T. harzianum 6J X AN 9.81 2.64 5.00 0.65 0.63 4.27T12 T. harzianum JND X AN 50.6 2.03 4.62 0.14 0.40 3.56T13 Control e A. Niger (AN) e 3.19 6.73 0.11 0.78 0.55S.Em. 0.90 0.05 0.15 0.04 0.10 0.11C.D. at 5% 2.60 0.16 0.43 0.11 0.29 0.33C.V. % 3.35 4.74 5.52 10.90 10.73 6.07
a U mg�1protein; 4. U ¼ mM glucose min�1 ml�1; 5. U ¼ mM galacturonic acid min�1 ml�1; 6 . U ¼ mM N acetylglucosamine min�1 ml�1; 7. U ¼ mM glucose min�1 ml�1; 8.U ¼ mg free amino acids min�1 ml�1.
H.P. Gajera, D.N. Vakharia / Physiological and Molecular Plant Pathology 74 (2010) 274e282 279

Author's personal copy
spp. in order to test the role of these chitinases in mycoparasitism,and the 42-kDa chitinase is believed to be a key enzyme [43].
Another enzymatic system that is involved in cell wall degrada-tion by an antagonistic organism is b-glucan degrading enzymes. Theproduction of b-1,3 glucanase was reported as an important enzy-matic activity in bio-control microorganisms because b-1,3 glucan isa structural component of fungal cell walls. b-1,3 glucanase inhibitedspore germination or the growth of pathogens in synergistic coop-eration with chitinases [46,47]. Many b-1,3-glucanases have beenisolated, but only a few genes have been cloned, e.g. bgn13.1 [46].
Transformants overexpressing bgn13.1 have been reported to inhibitthe growth of B. cinerea, R. solani and Phytophthora citrophthora. Ourstudy also showed some molecular markers from Trichodermaisolates liked to higher antagonist activity against A. niger andproduction of cell wall degrading enzymes (chitinases, b-1,3 gluca-nase and protease) during antagonism.
In addition to chitin and glucan, filamentous fungi cell wallcontains proteins. Some antagonists like T. viride 60 and T. harzianum2J secreted about 9 fold higher proteolytic activity during antagonismcompared to control as recorded on 6 DAI. The production of
Fig. 3. Antagonism between Trichoderma isolates and A. niger at 6 DAI (Antagonists petri dish (T1 to T12) have Trichoderma isolates at the top and pathogen A. niger at the bottom).(T1 ¼ T. viren BAN � AN; T2 ¼ T. viride BAN � AN; T3 ¼ T. viride JND � AN; T4 ¼ T. harzianum BAN � AN; T5 ¼ T. viride 54; T6 ¼ T. viride 60 � AN; T7 ¼ T. viride 62 � AN;T8 ¼ T. harzianum 2J � AN; T9 ¼ T. harzianum 4J � AN; T10 ¼ T. harzianum 5J � AN; T11 ¼ T. harzianum 6J � AN; T12 ¼ T. harzianum JND � AN; T13 ¼ Control-A. niger(AN)).
H.P. Gajera, D.N. Vakharia / Physiological and Molecular Plant Pathology 74 (2010) 274e282280

Author's personal copy
proteasesmay play a role in antagonism [48]. The role of proteases inmycoparasitism has been reinforced with the isolation of newprotease-overproducing strains of T. harzianum [49]. Proteasesinvolved in the degradation of heterologous proteins have beencharacterized by Delgado-Jarana et al. [50]. Marco et al. [24]demonstrated that antagonistic fungi Trichoderma synthesizedproteases which may act on the host cell wall of pathogens in vitro.
Identification of Trichoderma isolates based on morphologicaldata, which used as a taxonomic tool have been confusing. Therefore,re-identification of isolates of Thichoderma using molecular tools(RAPD) was important [51]. In present study, the highest growthinhibitionof pathogenA. nigerwas86.2% and 80.4% by T. viride60 andT. harzianum 2J respectively during in vitro study, which were alsoin same cluster and shared 63% similarity (Fig. 4). However, isolatespecific markers for various Trichoderma were generated withspecific primers (Table 2). The results showed that RAPD can be usedfor evaluating the molecular variation existing within a group ofgenotypes (intraspecific genetic variability). Several workers haveused RAPD markers to detect genetic variations among the variousTrichoderma isolates [16,52,53]. Similar to our study, Goes et al. [54]also used RAPD technique to examine the genetic variability among14 isolates of Trichoderma and their ability to antagonize R. solaniusing a dual culture assay for correlation among RAPD products andtheir hardness to R. solani. They found that belonging to the samespecies or strains, the isolates presented low similarity around 55%which support present findings. Shalini et al. [17] carried out char-acterization of 17 bio-control strains identified as Trichoderma, andfrom R. solani using RAPD. They found 85 out of 102 bands poly-morphic using 17 random primers.
Fourteen strains of Trichoderma were studied by Santos [55],using molecular markers of RAPD among other techniques, and itwas possible to verify the natural genetic variability and to dividethe strains in similarity groups, as well as differentiating the orig-inal strains of different regions. Zimand et al. [56] used RAPDmarkers to distinguish strains of Trichoderma. Ten of the strainsidentified as T. harzianum exhibited similarities, and it was possibleto distinguish the isolate T-39, used commercially as bio-controlagent of B. cinerea. For efficient selection of species of Trichodermawith taxonomic finalities, this may support our results.
The results reported in this work clearly indicated that the RAPDmarkers linked to antagonistic action are observed with some fungalprimers. T. viride 60 and T. harzianum 2J (first two higher inhibitoryacting antagonists), elevated production of cell wall degradingenzymes - chitinase, b-1,3 glucanase and protease in culturemediumduring in vitro antagonismwith test fungus A. niger, generated RAPDmarkers. This showed 10 marker loci for diagnosis of T. viride 60
and/or T. harzianum 2J viz., RFu C-4 (1528, 1100 and 916 bp), RFu C-6(2002, 1789, 681 bp), RFu C-1 (712, 580 bp), RFu C-3 (1015, 703 bp).Primer RFu C-2 amplified DNA of pathogen A. niger onlyand produced 1100, 786 bp markers which may be useful to identifypathogen from Trichoderma isolates.
Among the 12 isolates of Trichoderma, T. viride 60 was the bestagent to inhibit the growth of fungal A. niger on PDAmedia. Further,T. viride 60 isolate was tested in pot culture study. Five varieties ofpeanut seeds (J-11, GG-2, GAUG-10, GG-13 and GG-20) treated withT. viride 60 (microbial load 1.83 � 106 cfu/g talc powder) and shownin A. niger infested potting mixture (microbial load 1.5 � 107 cfu/gsoil) were resulted to reduce the collar rot incidence of peanut byabout 58% under sick soil condition as recorded on 15 days aftersowing. Thus, T. viride 60might have a significant role in the controlof collar rot disease by reducing the virulence of A. niger in thepeanut rhizosphere.
Appendix. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.pmpp.2010.04.005.
References
[1] Abarca M, Bragulat M, Castellá G, Cabañes F. Ochratoxin- a production bystrains of Aspergillus niger var. niger. Appl Environ Microbiol 1994;60:2650e2.
[2] Jochem SCJ. Aspergillus niger on groundnut. Indisch Culturen (Teysmannia)1926;11:325e6.
[3] Gibson IAS. Crown rot seedling diseases of groundnut caused by A. niger II.Anomalour effect of orange mercurial seed dressings. Trans Br Mycol Soc1953;36:324e34.
[4] Chohan JS. Collar rot of groundnut caused by A. niger in the Punjab. J Res PAU1965;3:25e33.
[5] Leach LD, Garber RH. Control of Rhizoctonia solani. In: Parmeter JR, editor.Rhizoctonia solani: biologyand pathology. Berkeley: The University of Cal-ifornia Press; 1970. p. 189e99.
[6] Melo S. Potencialidade da utilização de Trichoderma spp., no controle biológicode doenças de plantas (Org.). In: BettiolW, editor. Controle biológico de doençasde plantas. Jaguariúna:EMBRAPA-CNPDA; 1991. p. 135e56 (EMBRAPA-CNPDA.Documentos; 15).
[7] Nelson EB. Current limits to biological control of fungal phytopathogens. In:Arora DK, Rai B, Mukerji KG, Knudsen G, editors. Handbook of appliedmycology-soil and plants, 1. New York: Marcell Dekker; 1991. p. 327e55.
[8] Papavizas GC. Trichoderma and Gliocladium: biology and potential forbiocontrol. Annual Rev Phytopathol 1985;23:23e54.
[9] Papavizas GC. Biological control of selected soil borne plant pathogens withGliocladium and Trichoderma. In: Tjamos EC, Papavizas GC, Cook RJ, editors.Biological control of plant diseases. , New York: Plenum Press; 1992. p. 223e30.
[10] Grondona I, Hermosa MR, Tejada M, Gomis MD, Mateos PF, Bridge P, et al.Physiological and biochemical characterization of Trichoderma harzianum,a biological control agent against soil borne fungal plant pathogens. ApplEnviron Microbiol 1997;63:3189e98.
[11] Noronha MA, Sobrinho SA, Silveira NSS, Michereff SJ, Mariano RLR,Maranhão E. Seleção de isolados de Trichoderma spp. para o controle deRhizoctonia solani em feijoeiro. Summa Phytopathologica 1996;22:156e62.
[12] Fugimori F, Okuda T. Application of the random amplified polymorphic DNAusing the polymerase chain reaction for efficient elimination of duplicatestrains in microbial screening. J Antibiot 1993;47:173e82.
[13] Schlick A, Kuhls K, Meyer W, Lieckfeldt E, Börner T, Messner K. Fingerprintingreveals gamma-ray induced mutations in fungal DNA: implications for iden-tification of patent strains of Trichoderma harzianum. Curr Genet 1994;26:74e8.
[14] Sinclair JB, Dhingra OD. Basic plant pathology method. MW Boca Rotam,Florida: CRC Press, Inc. Corporate Blud; 1985. 295e315.
[15] Punekar NS, Suresh Kumar SV, Jayashri TN, Anuradha R. Isolation of genomicDNA from acetone dried Aspergillus mycelia. Fungal Genet Newsl 2006;50:15e6.
[16] Abbasi PA, Miller SA, Mealia T, Hoitink HAJ, Kim JM. Precise detection andtracing of Trichoderma hamatum 382 in compost amended potting mixes byusing molecular markers. Appl Environ Microbiol 1999;12:5421e6.
[17] Shalini, Narayan KP, Lata, Kotasthane AS. Genetic related ness among Tri-choderma isolates inhibiting a pathogenic fungi Rhizoctonia Solini. AfrJ Biotech 2006;5:580e4.
[18] Rohlf FJ. NTSYS-pe numerical taxonomy and multivariate analysis systemversion 2.2, Exeter software, Applied biostatics. New York; 2004.
[19] Sneath PHA, Sokal RR. Numerical taxonomy. The principle and practice ofnumerical classification. San Francisco: W.H. Freemon and Company; 1973.
Jaccard's Coefficient0.19 0.34 0.49 0.65 0.80
T.virenB
T.harziaB
T.viride54
T.virideL
T.virideB
T.harzia5J
T.harzia6J
T.viride60
T.harzia2J
T.harziaL
T.viride62
T.harzia4J
A.niger
J
J
A
B
Fig. 4. UPGMA dendrogram depicting the genetic relationships among the twelveTrichoderma isolates and pathogen A. niger based on the RAPD data.
H.P. Gajera, D.N. Vakharia / Physiological and Molecular Plant Pathology 74 (2010) 274e282 281

Author's personal copy
[20] Ghislain M, Zhang D, Fajardo D, Huamann Z, Hijmans RH. Marker assistedsampling of the cultivated Andean potato Solanum phureja collection usingRAPD markers. Genet Resour Crop Evol 1999;46:547e55.
[21] Dennis CJ, Webster J. Antagonism properties of species groups of Tricho-derma, III hyphal interaction. Trans Br Mycol Soc 1971;57:363e9.
[22] Watanabe N. Antagonism by various kind of Trichoderma fungi to soil bornplant pathogen. Maiji University, Japan. Bulletin of Faculty of Agriculture1984;66:45e50.
[23] Kishore GK, Pande S, Podile AR. Pseudomonas aeruginosa GSE 18 inhibits thecell wall degrading enzymes of Aspergillus niger and activates defence-relatedenzymes of groundnut in control of collar rot disease. Aust Pl Pathol 2006;35:259e63.
[24] Marco JLD, Valadares-Inglis MC, Felix CR. Production of hydrolytic enzymes byTrichoderma isolates with antagonistic activity against Crinipellis perniciosathe causal agent of witches broom of cocoa. Braz J Microbiol 2003;34:33e8.
[25] Sivan A, Chet I. Integrated control of media on growth and interactionsbetween a range of soil borne glasshouse pathogens and antagonistic fungi.Phytopathology 1993;10:127e42.
[26] Lowry OW, Rosebrough NJ, Farr AC, Randall RJ. Protein measurements withfolin-phenol reagent. J Biol Chem 1951;193:255e7.
[27] Sadasivam S, Manickam K. Biochemical method for agricultural sciences.Coimbatore: Wiley Estern Limited; 1992.
[28] Collmer A, Reid JL, Mount MS. Assay methods for pectic enzymes. MethEnzymol 1988;161:329e35.
[29] Boller T, Mauch F. Colorimetric assay of chitinase. Meth Enzymol 1988;161:431e5.
[30] Reissig JL, Strominger JL, Lefloir LF. A modified colorimetric method for theestimation of N-acetyl amino sugars. J Biol Chem 1955;217:959e66.
[31] Kauffman S, Legrand M, Geoffory P, Fritig B. Biological functions of ‘pathogen-esis related’proteins four PR proteins of tobacco have b�1,3�glucanaseactivity.EMBO J 1987;6:3209e12.
[32] Malik CP, Singh SP. Plant enzymology and histoenzymology. Ludhiana:Kalyani Publishers; 1980.
[33] Lee YP, Takahashi T. An improved colorimetric determination of amino acidswith the use of ninhydrin. Anal Biochem 1966;14:71e3.
[34] Prameala M, Rajeswari B, Prasad RD, Reddy DRR. Bioefficacy of antagonistagainst Fusarium oxysporum f. sp. Carthami isolates inciting sunflower wilt.J Mycol Plant Pathol 2005;35:2.
[35] Khattabi N, Ezzahiri B, Louali L, Oihabi A. Antagonistic activity of Trichodermaisolates against Sclerotium rolfsii: screening of efficient isolates from Moroccosoils for biological control. Phytopathol Mediterr 2004;43:332e40.
[36] Shalini S, Kotasthane AS. Parasitism of Rhizoctonia solani by strains of Tri-choderma spp. EJEAF Chem 2007;6:2272e81.
[37] Collmer A, Keen NT. The role of pectic enzymes in plant pathogenesis. AnnuRev Phytopathol 1986;24:383e409.
[38] Haran S, Schickler H, Chet I. Molecular mechanisms of lytic enzymes involvedin the biocontrol activity of Trichoderma harzianum. Microbiology 1996;142:2321e31.
[39] Lorito M, Hayes CK, Di Pietro A, Woo SL, Harman GE. Purification, characteriza-tion and synergistic activity of a glucan 1,3-b-glucosidase and an N-acetylglu-cosaminidase from T. harzianum. Phytopathol 1994;84:398e405.
[40] Carsolio C, Benhamou N, Haran S, Cortés C, Gutiérrez A. Chet Herrera-EstrellaA. Role of the Trichoderma harzianum endochitinase gene, ech42, in myco-parasitism. Appl Environ Microbiol 1999;65:929e35.
[41] Sangle UR, Bambawale OM. New strains of Trichoderma spp. Strongly antag-onistic against Fusarium oxysporum f. sp. Sesami. J Mycol Plant Pathol 2004;34:1.
[42] Harman GE, Howell CR, Viterbo A, Chet I, Lorito M. Trichoderma species-opportunistic, avirulent plant symbionts. Nat Rev 2004;2:43e56.
[43] Howell CR. Mechanisms employed by Trichoderma species in the biologicalcontrol of plant diseases: the history and evolution of current concepts. PlantDis 2003;87:4e10.
[44] Viterbo A, Haran S, Friesem D, Ramot O, Chet I. Anti-fungal activity of a novelendochitinase gene (chit36) from T. harzianum Rifai TM. FEMS Microbiol Lett2001;200:169e74.
[45] Dana MM, Limón MC, Mejías R, Mach RL, Benítez T, Pintor-Toro JA, et al.Regulation of chitinase 33 (chit33) gene expression in Trichoderma harzianum.Curr Genet 2001;38:335e42.
[46] Benítez T, Delgado-Jarana J, Rincón AM, Rey M, Limón MC. Biofungicides: tri-choderma as a biocontrol agent against phytopathogenic fungi. In: Pandalai SG,editor. Recent research developments microbiology, vol. 2. Trivandrum:Research Signpost; 1998. p. 129e50.
[47] El-Katatny MH, Gudelj M, Robra KH, Elnaghy MA, Gubitz GM. Characterizationof a chitinase and an endo-b�1,3�glucanasefrom Trichoderma harzianum RifaiT24 involved in control of the phytopathogen Sclerotium rolfsii. Appl MicrobiolBiotechnol 2001;56:137e43.
[48] Flores A, Chet I, Herrera-Estrella A. Improved biocontrol activity of T. harzia-num strains by overexpression of the proteinase encoding gene prb1. CurrGenet 1997;31:30e7.
[49] Szekeres A, Kredics L, Antal Z, Kevei F, Manczinger L. Isolation and charac-terization of protease overproducing mutants of Trichoderma harzianum.FEMS Microbiol Lett 2004;233:215e22.
[50] Delgado-Jarana J, Pintor-Toro JA, Benítez T. Overproduction of â-1,6-glucanasein Trichoderma harzianum is controlled by extracellular acidic proteases andpH. Biochim Biophys Acta 2000;1481:289e96.
[51] Ranganath HR, Shyam Prasad G, Sheeba. PCR-fingerprinting of some Tricho-derma isolates from two Indian type culture collections e a need for re-identification of these economically important fungi. Curr Sci 2002;83:372e4.
[52] Lieckfeldt E, Samuels GJ, Nirenberg HI, Petrint O. A morphological andmolecular perspective of Trichoderma viride: is it one or two species? ApplEnviron Microbiol 1999;65:2418e28.
[53] Hermosa MR, Grondona I, Iturriaga EA, Diaz-minguez JM, Castro C, Monte E,et al. Molecular characterization and identification of biocontrol isolates ofTrichoderma spp. Appl Environ Microbiol 2000;66:1890e8.
[54] Goes LB, Costa ABL, Freire LLC, Oliveira NT. Randomly amplified polymorphicDNA of Trichoderma isolates and antagonism against Rhizoctonia solani. BrazArch Biol Technol 2002;45:151e60.
[55] Santos IC. Avaliacao da variabilidade genética de Trichoderma por isoenzimas,RFLP e RAPD.Piracicaba-SP., Thesis- São Paulo University, Escola Superior deAgricultura “Luiz de Queiroz” (ESALQ), Brazil; 1992.
[56] Zimand G, Valinsky L, Elad Y, Chet I, Manulis S. Use of the RAPD procedure forthe identification of Trichoderma strains. Mycol Res 1994;98:531e4.
H.P. Gajera, D.N. Vakharia / Physiological and Molecular Plant Pathology 74 (2010) 274e282282