mischaracterising density dependence biases estimated effects of coloured covariates on population...
TRANSCRIPT

ORIGINAL ARTICLE
Mischaracterising density dependence biases estimated effectsof coloured covariates on population dynamics
Andreas Linden • Mike S. Fowler • Niclas Jonzen
Received: 25 March 2012 / Accepted: 29 August 2012 / Published online: 13 November 2012
� The Society of Population Ecology and Springer Japan 2012
Abstract Environmental effects on population growth
are often quantified by coupling environmental covariates
with population time series, using statistical models that
make particular assumptions about the shape of density
dependence. We hypothesized that faulty assumptions
about the shape of density dependence can bias estimated
effect sizes of temporally autocorrelated covariates. We
investigated the presence of bias using Monte Carlo sim-
ulations based on three common per capita growth func-
tions with distinct density dependent forms (h-Ricker,
Ricker and Gompertz), autocorrelated (coloured) ‘known’
environmental covariates and uncorrelated (white)
‘unknown’ noise. Faulty assumptions about the shape of
density dependence, combined with overcompensatory
intrinsic population dynamics, can lead to strongly biased
estimated effects of coloured covariates, associated with
lower confidence interval coverage. Effects of negatively
autocorrelated (blue) environmental covariates are
overestimated, while those of positively autocorrelated
(red) covariates can be underestimated, generally to a les-
ser extent. Prewhitening the focal environmental covariate
effectively reduces the bias, at the expense of the estimate
precision. Fitting models with flexible shapes of density
dependence can also reduce bias, but increases model
complexity and potentially introduces other problems of
parameter identifiability. Model selection is a good option
if an appropriate model is included in the set of candidate
models. Under the specific and identifiable circumstances
with high risk of bias, we recommend prewhitening or
careful modelling of the shape of density dependence.
Keywords Autoregressive models � Environmental
forcing � Prewhitening � Statistical inference � Theta-Ricker
model � Time series
Introduction
Environmental forcing is recognized as an important
determinant of population fluctuations, in combination with
intrinsic factors, such as density dependence and popula-
tion structure (Turchin 1995, 1999). Statistical identifica-
tion of important environmental variables affecting
population growth, and the quantification of such envi-
ronmental effects, are crucial research questions in popu-
lation ecology (Stenseth et al. 2002; Ruokolainen et al.
2009; Knape and de Valpine 2011), including its applica-
tions to e.g., conservation, pest control, fisheries and
wildlife management. For example, rainfall has important
positive effects on hunted populations of ducks in North
America (Johnson and Grier 1988) and on red kangaroo in
Australia (McCarthy 1996), which needs to be accounted
for when making management decisions.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10144-012-0347-0) contains supplementarymaterial, which is available to authorized users.
A. Linden (&)
Department of Biology, Centre for Ecological and Evolutionary
Synthesis, University of Oslo, P.O. Box 1066, Blindern,
0316 Oslo, Norway
e-mail: [email protected]
M. S. Fowler
Population Ecology Group, Institut Mediterrani d’Estudis
Avancats (UIB–CSIC), Miquel Marques 21,
07190 Esporles, Spain
N. Jonzen
Department of Biology (Theoretical Population Ecology and
Evolution Group), Lund University, Ecology Building,
SE-223 62 Lund, Sweden
123
Popul Ecol (2013) 55:183–192
DOI 10.1007/s10144-012-0347-0

Historically the cross-correlation between environmental
variables and population density was used to detect envi-
ronmental effects, but that approach easily generates mis-
leading results (e.g., Royama 1981, 1992; Ranta et al. 2000;
Kaitala and Ranta 2001; Lundberg et al. 2002). The most
common modern approach is to fit simple phenomenolog-
ical models of population growth, including density
dependence, environmental covariates and possibly inter-
actions between those two, to time series of population
counts or density (e.g., Stenseth et al. 2002; Brannstrom and
Sumpter 2006; Knape and de Valpine 2011). Despite the
risk of oversimplification, this approach can be seen as a
step towards better estimates of a more quantitative nature.
Analyses of autocorrelated (coloured) population time
series are associated with a number of complexities,
potentially leading to spurious statistical relationships and/
or bias (Chatfield 2004; Ripa and Ives 2007; Linden and
Knape 2009). It should be emphasized that because eco-
logical time series are often short (Cazelles 2004), so are
the covariates used. Hence, even if the underlying envi-
ronmental process described by the covariate is uncorre-
lated over long time-scales, it is not uncommon that the
selected covariate has a temporal structure over shorter
temporal periods (Ruokolainen et al. 2009). Density
dependence is often modelled assuming some rather simple
functional relationship between density and population
growth (e.g., Berryman and Turchin 2001; Jonzen et al.
2002). If the estimated strength of density dependence is
biased, e.g., by ignoring observation error in population
density, the estimates of autocorrelated environmental
effects can also become biased (Linden and Knape 2009).
We hypothesize that similar bias could arise due to poor
approximation of the intrinsic population dynamic struc-
ture, e.g., the wrong shape of density dependent feedback.
Here we use a simulation approach to study whether
statistical estimates of environmental effect sizes on pop-
ulation growth are trustworthy when the true shape of
density dependence is unknown or poorly approximated.
We focus on the accuracy and precision of point estimates
for the estimated environmental effects, as well as the
confidence interval coverage. We also assess whether
identified bias can be reduced with prewhitening and/or a
flexible approach to modelling the shape of density
dependence. We highlight situations where common
practice would lead to biased estimates of environmental
effects, and discuss how existing statistical procedures can
be implemented to minimize the identified risk of bias.
Methods
To investigate the estimated effect sizes of environmental
covariates, we simulated time series data according to
stochastic generalizations of three well known, discrete
time population growth models (see section ‘Population
growth models and data simulation’ below). We varied the
strength of density compensation across a wide range of
return rates (TR) and incorporated an autocorrelated (col-
oured) stochastic environmental variable. We then applied
three statistical models (each corresponding to a specific
population model) to the data generated by each population
model, to estimate the effect sizes of the environmental
variable of interest (see section ‘Statistical estimation of
the environmental effect’ below). This approach was based
on the assumption that researchers generally do not have
perfect knowledge of the underlying processes driving
intrinsic dynamics in natural populations. Previous work
has considered systems including observation error, cou-
pled only with the correct form of density dependence
(Linden and Knape 2009), demonstrating the presence of
bias when observation error is ignored. We only considered
process error models in this study, i.e., we assumed prefect
knowledge about population density (no observation error),
to simplify the analysis and avoid overlap with that study.
Population growth models and data simulation
We simulated time series data using models of the general
form
Ntþ1 ¼ Nt exp f Ntð Þ þ bWt þ ut½ � ð1Þ
where Nt is population density at time t, f (Nt) is a density
dependent function of per capita growth that maps
expected changes in population density over consecutive
time steps, and Wt is a known, potentially autocorrelated
(coloured) environmental variable with effect size b (the
focus of this study). The term ut represents unknown
environmental variation and is i.i.d. normally distributed
(white) with zero mean and variance r2. To initiate pop-
ulations, we drew N1 from a uniform random distribution
with limits [0, 2], i.e., with a maximum of twice the
equilibrium density, N* = 1. We simulated the population
dynamics for 130 and 200 time steps, discarding the first
100 time steps in both cases to avoid transient effects due
to initial densities, using the final 30 and 100 steps (29 and
99 transitions in population size), respectively, for further
analysis.
The per capita growth functions f (Nt) investigated here
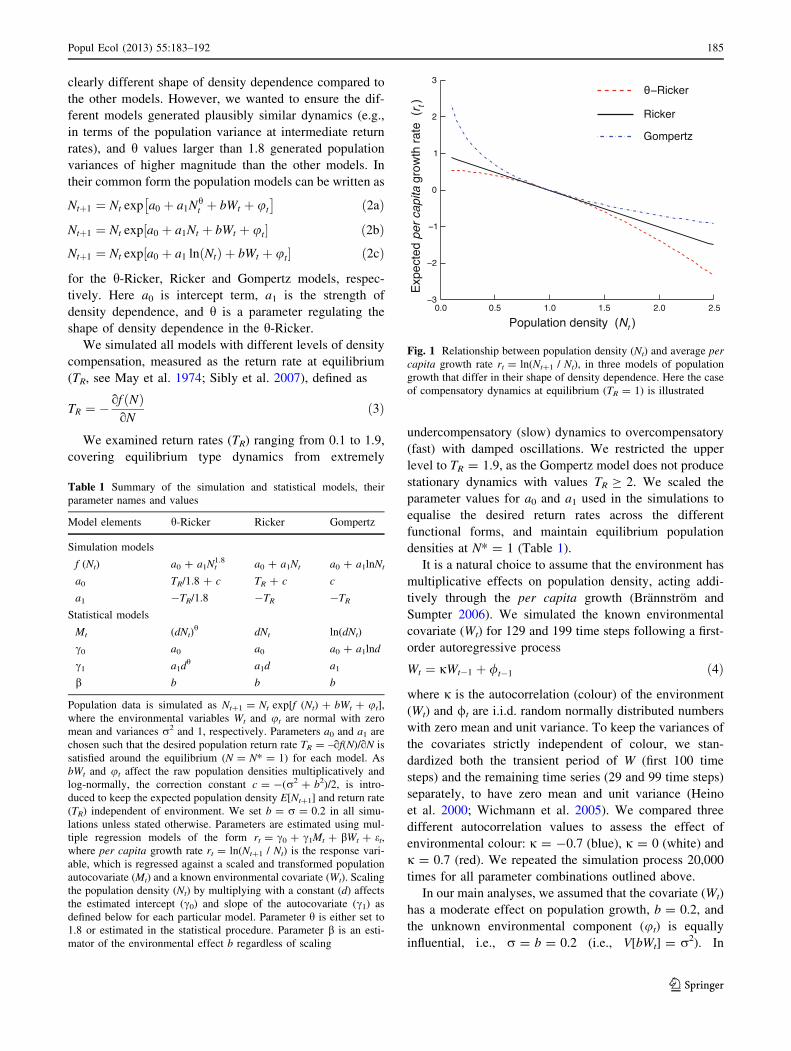
(Table 1) are generalizations of (1) the h-Ricker model
with the parameter h fixed to 1.8; (2) the Ricker model; and
(3) the Gompertz model; (Gilpin and Ayala 1973; Ricker
1954; Gompertz 1825, respectively). These models differ
here only in their shape of density dependence, showing
concave, linear and convex relationships of density to per
capita growth rate (Fig. 1). As such, the choice of
parameter h = 1.8 is somewhat arbitrary, but represents a
184 Popul Ecol (2013) 55:183–192
123
clearly different shape of density dependence compared to
the other models. However, we wanted to ensure the dif-
ferent models generated plausibly similar dynamics (e.g.,
in terms of the population variance at intermediate return
rates), and h values larger than 1.8 generated population
variances of higher magnitude than the other models. In
their common form the population models can be written as
Ntþ1 ¼ Nt exp a0 þ a1Nht þ bWt þ ut
� �ð2aÞ
Ntþ1 ¼ Nt exp a0 þ a1Nt þ bWt þ ut½ � ð2bÞNtþ1 ¼ Nt exp a0 þ a1 ln Ntð Þ þ bWt þ ut½ � ð2cÞ
for the h-Ricker, Ricker and Gompertz models, respec-
tively. Here a0 is intercept term, a1 is the strength of
density dependence, and h is a parameter regulating the
shape of density dependence in the h-Ricker.
We simulated all models with different levels of density
compensation, measured as the return rate at equilibrium
(TR, see May et al. 1974; Sibly et al. 2007), defined as
TR ¼ �of Nð ÞoN
ð3Þ
We examined return rates (TR) ranging from 0.1 to 1.9,
covering equilibrium type dynamics from extremely
undercompensatory (slow) dynamics to overcompensatory
(fast) with damped oscillations. We restricted the upper
level to TR = 1.9, as the Gompertz model does not produce
stationary dynamics with values TR C 2. We scaled the
parameter values for a0 and a1 used in the simulations to
equalise the desired return rates across the different
functional forms, and maintain equilibrium population
densities at N* = 1 (Table 1).
It is a natural choice to assume that the environment has
multiplicative effects on population density, acting addi-
tively through the per capita growth (Brannstrom and
Sumpter 2006). We simulated the known environmental
covariate (Wt) for 129 and 199 time steps following a first-
order autoregressive process
Wt ¼ jWt�1 þ /t�1 ð4Þ
where j is the autocorrelation (colour) of the environment
(Wt) and /t are i.i.d. random normally distributed numbers
with zero mean and unit variance. To keep the variances of
the covariates strictly independent of colour, we stan-
dardized both the transient period of W (first 100 time
steps) and the remaining time series (29 and 99 time steps)
separately, to have zero mean and unit variance (Heino
et al. 2000; Wichmann et al. 2005). We compared three
different autocorrelation values to assess the effect of
environmental colour: j = -0.7 (blue), j = 0 (white) and
j = 0.7 (red). We repeated the simulation process 20,000
times for all parameter combinations outlined above.
In our main analyses, we assumed that the covariate (Wt)
has a moderate effect on population growth, b = 0.2, and
the unknown environmental component (ut) is equally
influential, i.e., r = b = 0.2 (i.e., V[bWt] = r2). In
Table 1 Summary of the simulation and statistical models, their
parameter names and values
Model elements h-Ricker Ricker Gompertz
Simulation models
f (Nt) a0 ? a1Nt1.8 a0 ? a1Nt a0 ? a1lnNt
a0 TR/1.8 ? c TR ? c c
a1 -TR/1.8 -TR -TR
Statistical models
Mt (dNt)h dNt ln(dNt)
c0 a0 a0 a0 ? a1lnd
c1 a1dh a1d a1
b b b b
Population data is simulated as Nt?1 = Nt exp[f (Nt) ? bWt ? ut],
where the environmental variables Wt and ut are normal with zero
mean and variances r2 and 1, respectively. Parameters a0 and a1 are
chosen such that the desired population return rate TR = –qf(N)/qN is
satisfied around the equilibrium (N = N* = 1) for each model. As
bWt and ut affect the raw population densities multiplicatively and
log-normally, the correction constant c = -(r2 ? b2)/2, is intro-
duced to keep the expected population density E[Nt?1] and return rate
(TR) independent of environment. We set b = r = 0.2 in all simu-
lations unless stated otherwise. Parameters are estimated using mul-
tiple regression models of the form rt = c0 ? c1Mt ? bWt ? et,
where per capita growth rate rt = ln(Nt?1 / Nt) is the response vari-
able, which is regressed against a scaled and transformed population
autocovariate (Mt) and a known environmental covariate (Wt). Scaling
the population density (Nt) by multiplying with a constant (d) affects
the estimated intercept (c0) and slope of the autocovariate (c1) as
defined below for each particular model. Parameter h is either set to
1.8 or estimated in the statistical procedure. Parameter b is an esti-
mator of the environmental effect b regardless of scaling
Fig. 1 Relationship between population density (Nt) and average percapita growth rate rt = ln(Nt?1 / Nt), in three models of population
growth that differ in their shape of density dependence. Here the case
of compensatory dynamics at equilibrium (TR = 1) is illustrated
Popul Ecol (2013) 55:183–192 185
123

addition, we investigated bias with different values for
parameters b and r, in the range 0.05–0.5, fixing the other
parameter (b or r) as 0.2. This was done by simulating data
with under- and overcompensatory Ricker dynamics
(TR = 0.2, 1.8) with statistical estimation based on Gom-
pertz-type density dependence—two of the most com-
monly used approaches (see Table 1). As the magnitude of
bias is likely to be proportional to b, we presented the
relative bias in the estimate (b) here as (b - b)/b. These
results were based on 100,000 replicates.
Statistical estimation of the environmental effect
For each replicated simulation we estimated the environ-
mental effect (b) using linear multiple regression of the
general form
rt ¼ c0 þ c1Mt þ bWt þ et ð5Þ
where the realized per capita growth rate rt = ln(Nt?1/Nt)
was the response variable, while an autocovariate Mt and the
known environmental variable Wt were used as explanatory
variables. The autocovariate varied between the models,
being Mt = (dNt)1.8 in the h-Ricker, Mt = dNt in Ricker and
Mt = ln(dNt) in Gompertz. Here d is the inverse of the
sample mean of the portion of the time series used in the
regression (d = n/P
Nt; n = 29 or 99). This scaling proce-
dure was carried out to avoid numerical problems in the
fitting process, which can occasionally be present under
extreme return rates. Multiplying the population density by a
constant (d) does not alter the response variable (rt), and the
least squares solution using a scaled autocovariate still gives
the maximum likelihood estimator of b (denoted as b) for
each of the given models. However, some of the other
parameters of the models are affected by the scaling in a
manner depending on the particular model used. A summary
of the correspondence between the parameters in the simu-
lation model (Eqs. 2a, 2b, 2c) and the regression models used
for statistical estimation (Eq. 5), including consequences of
scaling, is given in Table 1.
We also fitted the h-Ricker model, with parameter h free
to be estimated (Table 1) as an example of a model with a
flexible shape of density dependence. The two first simu-
lation models represent special cases of this, with h = 1.8
and h = 1, respectively. However, the Gompertz model is
an example of a structurally different model, which can be
approximated by the free h-Ricker model (with expected
estimates of 0 \ h\ 1) to reduce possible bias. Fitting the
h-Ricker model can be viewed as a separable non-linear
least squares problem (Ruhe and Wedin 1980), which can
be solved with ordinary least squares, conditionally on the
parameter h. We used a one-dimensional golden section
search over parameter h, coupled with ordinary least
squares regression estimation of the other parameters.
Finally, the analysis was repeated with a prewhitened
version of the environmental covariate, to investigate
whether this approach reduces any bias. Prewhitening is a
collective name for methods employed to remove auto-
correlation from time series, and is typically applied to
reduce or avoid problems in time series analyses arising
from autocorrelation (Chatfield 2004). The prewhitened
covariate we used was represented by the residuals from a
first order autoregression fitted to the covariate (Wt as the
response variable, Wt-1 as the explanatory variable, for
only those time steps included in the statistical analyses,
t C 101).
We assessed accuracy (the possible bias) and precision
(the consistency of the estimation procedure) of the envi-
ronmental effect estimates (b) by examining their means
± the standard deviations under the different simulated
conditions. We calculated the 95 % confidence interval
coverage, i.e., how often the estimated confidence intervals
actually contain the true value used to generate the data,
based on profile likelihoods (e.g., Hilborn and Mangel
1997), as one minus the rejection rate of a likelihood ratio
test, with the true value as the null hypothesis. As a rough
measure of model fit we used the coefficient of determi-
nation (R2), which here also equals the proportion of var-
iance in the per capita growth between consecutive years
ln(Nt?1/Nt) explained by the model. Among the models
with fixed shape of density dependence, and hence equal
number of estimated parameters, ranking models by R2 is
equivalent to ranking by information theoretical model
selection criteria. Therefore, we did not explicitly compare
models using model selection methods.
Results
When the model used for statistical estimation corre-
sponded to the simulation model that generated the data,
the estimated effect size of the known environmental
covariate (b) was unbiased, precise, and had trustworthy
confidence intervals (results not shown). In populations
with undercompensatory dynamics (TR \ 1), with equal
strengths of known and unknown environmental variation
(b = r = 0.2), the estimation properties of b were gen-
erally very good under all combinations of simulated and
fitted models we examined [Figs. 2, 3, and Fig. S1 in
Electronic Supplementary Material (ESM)]. Varying the
autocorrelation (colour; j) of the known environmental
covariate (Wt) did not qualitatively change this aspect of
the results.
With overcompensatory population dynamics (TR [ 1),
faulty assumptions about the underlying form of density
dependence did affect statistical estimates of environmental
effects. For negatively autocorrelated (blue) environments, b
186 Popul Ecol (2013) 55:183–192
123
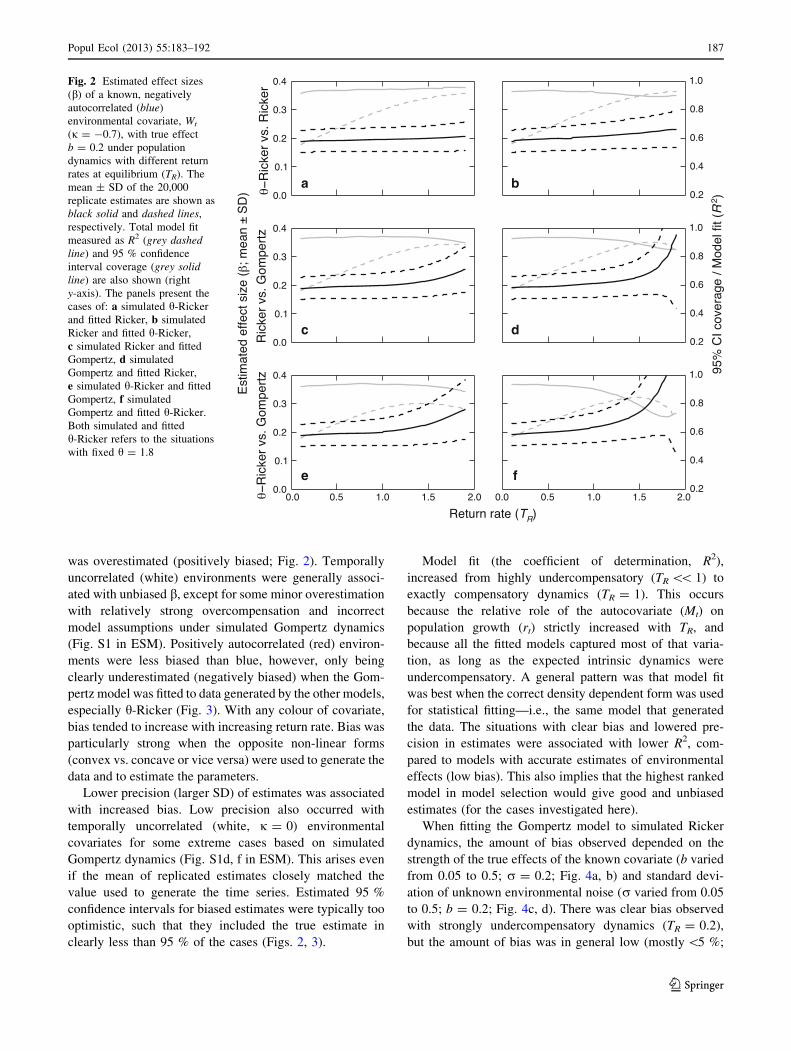
was overestimated (positively biased; Fig. 2). Temporally
uncorrelated (white) environments were generally associ-
ated with unbiased b, except for some minor overestimation
with relatively strong overcompensation and incorrect
model assumptions under simulated Gompertz dynamics
(Fig. S1 in ESM). Positively autocorrelated (red) environ-
ments were less biased than blue, however, only being
clearly underestimated (negatively biased) when the Gom-
pertz model was fitted to data generated by the other models,
especially h-Ricker (Fig. 3). With any colour of covariate,
bias tended to increase with increasing return rate. Bias was
particularly strong when the opposite non-linear forms
(convex vs. concave or vice versa) were used to generate the
data and to estimate the parameters.
Lower precision (larger SD) of estimates was associated
with increased bias. Low precision also occurred with
temporally uncorrelated (white, j = 0) environmental
covariates for some extreme cases based on simulated
Gompertz dynamics (Fig. S1d, f in ESM). This arises even
if the mean of replicated estimates closely matched the
value used to generate the time series. Estimated 95 %
confidence intervals for biased estimates were typically too
optimistic, such that they included the true estimate in
clearly less than 95 % of the cases (Figs. 2, 3).
Model fit (the coefficient of determination, R2),
increased from highly undercompensatory (TR \\ 1) to
exactly compensatory dynamics (TR = 1). This occurs
because the relative role of the autocovariate (Mt) on
population growth (rt) strictly increased with TR, and
because all the fitted models captured most of that varia-
tion, as long as the expected intrinsic dynamics were
undercompensatory. A general pattern was that model fit
was best when the correct density dependent form was used
for statistical fitting—i.e., the same model that generated
the data. The situations with clear bias and lowered pre-
cision in estimates were associated with lower R2, com-
pared to models with accurate estimates of environmental
effects (low bias). This also implies that the highest ranked
model in model selection would give good and unbiased
estimates (for the cases investigated here).
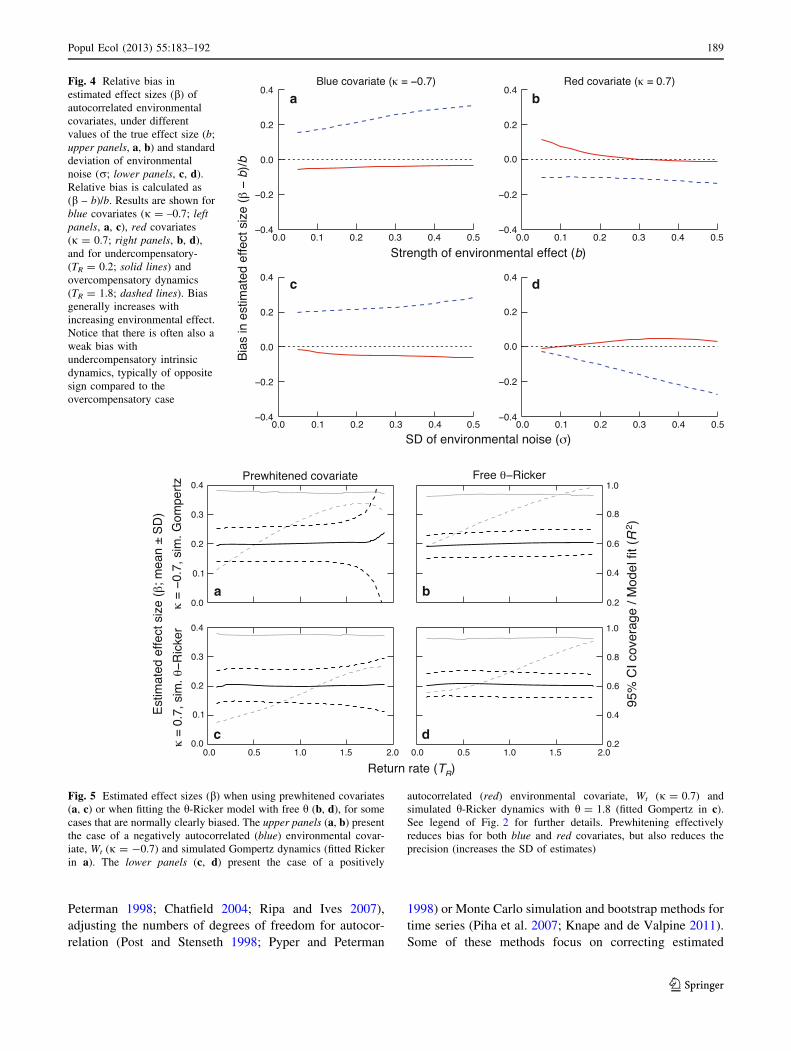
When fitting the Gompertz model to simulated Ricker
dynamics, the amount of bias observed depended on the
strength of the true effects of the known covariate (b varied
from 0.05 to 0.5; r = 0.2; Fig. 4a, b) and standard devi-
ation of unknown environmental noise (r varied from 0.05
to 0.5; b = 0.2; Fig. 4c, d). There was clear bias observed
with strongly undercompensatory dynamics (TR = 0.2),
but the amount of bias was in general low (mostly \5 %;
a b
c d
e f
Fig. 2 Estimated effect sizes
(b) of a known, negatively
autocorrelated (blue)
environmental covariate, Wt
(j = -0.7), with true effect
b = 0.2 under population
dynamics with different return
rates at equilibrium (TR). The
mean ± SD of the 20,000
replicate estimates are shown as
black solid and dashed lines,
respectively. Total model fit
measured as R2 (grey dashedline) and 95 % confidence
interval coverage (grey solidline) are also shown (right
y-axis). The panels present the
cases of: a simulated h-Ricker
and fitted Ricker, b simulated
Ricker and fitted h-Ricker,
c simulated Ricker and fitted
Gompertz, d simulated
Gompertz and fitted Ricker,
e simulated h-Ricker and fitted
Gompertz, f simulated
Gompertz and fitted h-Ricker.
Both simulated and fitted
h-Ricker refers to the situations
with fixed h = 1.8
Popul Ecol (2013) 55:183–192 187
123
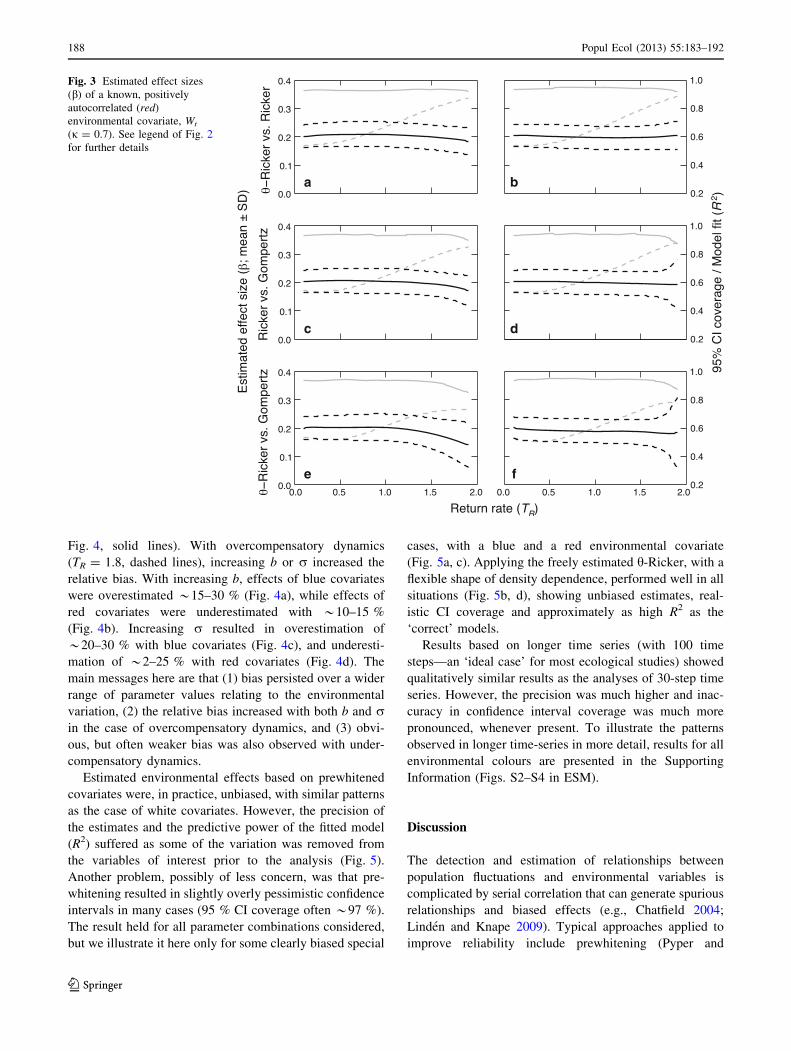
Fig. 4, solid lines). With overcompensatory dynamics
(TR = 1.8, dashed lines), increasing b or r increased the
relative bias. With increasing b, effects of blue covariates
were overestimated *15–30 % (Fig. 4a), while effects of
red covariates were underestimated with *10–15 %
(Fig. 4b). Increasing r resulted in overestimation of
*20–30 % with blue covariates (Fig. 4c), and underesti-
mation of *2–25 % with red covariates (Fig. 4d). The
main messages here are that (1) bias persisted over a wider
range of parameter values relating to the environmental
variation, (2) the relative bias increased with both b and rin the case of overcompensatory dynamics, and (3) obvi-
ous, but often weaker bias was also observed with under-
compensatory dynamics.
Estimated environmental effects based on prewhitened
covariates were, in practice, unbiased, with similar patterns
as the case of white covariates. However, the precision of
the estimates and the predictive power of the fitted model
(R2) suffered as some of the variation was removed from
the variables of interest prior to the analysis (Fig. 5).
Another problem, possibly of less concern, was that pre-
whitening resulted in slightly overly pessimistic confidence
intervals in many cases (95 % CI coverage often *97 %).
The result held for all parameter combinations considered,
but we illustrate it here only for some clearly biased special
cases, with a blue and a red environmental covariate
(Fig. 5a, c). Applying the freely estimated h-Ricker, with a
flexible shape of density dependence, performed well in all
situations (Fig. 5b, d), showing unbiased estimates, real-
istic CI coverage and approximately as high R2 as the
‘correct’ models.
Results based on longer time series (with 100 time
steps—an ‘ideal case’ for most ecological studies) showed
qualitatively similar results as the analyses of 30-step time
series. However, the precision was much higher and inac-
curacy in confidence interval coverage was much more
pronounced, whenever present. To illustrate the patterns
observed in longer time-series in more detail, results for all
environmental colours are presented in the Supporting
Information (Figs. S2–S4 in ESM).
Discussion
The detection and estimation of relationships between
population fluctuations and environmental variables is
complicated by serial correlation that can generate spurious
relationships and biased effects (e.g., Chatfield 2004;
Linden and Knape 2009). Typical approaches applied to
improve reliability include prewhitening (Pyper and
a b
c d
e f
Fig. 3 Estimated effect sizes
(b) of a known, positively
autocorrelated (red)
environmental covariate, Wt
(j = 0.7). See legend of Fig. 2
for further details
188 Popul Ecol (2013) 55:183–192
123
Peterman 1998; Chatfield 2004; Ripa and Ives 2007),
adjusting the numbers of degrees of freedom for autocor-
relation (Post and Stenseth 1998; Pyper and Peterman
1998) or Monte Carlo simulation and bootstrap methods for
time series (Piha et al. 2007; Knape and de Valpine 2011).
Some of these methods focus on correcting estimated
a b
c d
Fig. 4 Relative bias in
estimated effect sizes (b) of
autocorrelated environmental
covariates, under different
values of the true effect size (b;
upper panels, a, b) and standard
deviation of environmental
noise (r; lower panels, c, d).
Relative bias is calculated as
(b – b)/b. Results are shown for
blue covariates (j = –0.7; leftpanels, a, c), red covariates
(j = 0.7; right panels, b, d),
and for undercompensatory-
(TR = 0.2; solid lines) and
overcompensatory dynamics
(TR = 1.8; dashed lines). Bias
generally increases with
increasing environmental effect.
Notice that there is often also a
weak bias with
undercompensatory intrinsic
dynamics, typically of opposite
sign compared to the
overcompensatory case
a b
c d
Fig. 5 Estimated effect sizes (b) when using prewhitened covariates
(a, c) or when fitting the h-Ricker model with free h (b, d), for some
cases that are normally clearly biased. The upper panels (a, b) present
the case of a negatively autocorrelated (blue) environmental covar-
iate, Wt (j = -0.7) and simulated Gompertz dynamics (fitted Ricker
in a). The lower panels (c, d) present the case of a positively
autocorrelated (red) environmental covariate, Wt (j = 0.7) and
simulated h-Ricker dynamics with h = 1.8 (fitted Gompertz in c).
See legend of Fig. 2 for further details. Prewhitening effectively
reduces bias for both blue and red covariates, but also reduces the
precision (increases the SD of estimates)
Popul Ecol (2013) 55:183–192 189
123

uncertainty (e.g., statistical significance), but do not correct
for possible bias in effect sizes, and are hence of limited
help in quantitative studies. Our results indicate that there
are clear risks of bias in estimated effects of coloured
covariates on population growth (and the associated con-
fidence intervals), when the intrinsic dynamics are over-
compensatory, and the statistical model used assumes an
incorrect shape of density dependence. This is particularly
the case with negatively autocorrelated (blue) covariates,
but there are also bias problems under other conditions,
e.g., when unknown sources of variation (process error)
have higher variance than known covariates (Fig. 4). The
results presented here might also apply to other situations
where the covariate of interest is strongly autocorrelated
and the intrinsic model structure applied is incorrect or
ignores important dynamical feedbacks, such as age-
structure (Kaitala and Ranta 2001) or interspecific inter-
actions (Fowler 2009).
While problems with serial correlation are present in
simple correlation analysis between two time series, the
issue is commonly ignored in autoregressive models of the
type used here, as they specifically account for autocorre-
lation in the response variable during statistical analysis.
The results presented here show that there can still be
problems when the specific functional form of the statis-
tical model mischaracterises the autoregressive relation-
ships. A similar situation occurs when observation errors in
population density are ignored (Linden and Knape 2009),
causing strong bias in the estimated density dependence
(Freckleton et al. 2006; Knape 2008). In both cases, the
incorrect shape of modelled density dependence fails to
account for autocorrelation correctly, leaving an incorrect
autocorrelative structure of the remaining variation, rein-
troducing problems of spurious correlation. Coloured
environmental variation interacts with intrinsic dynamics,
either amplifying or dampening population fluctuations
(Roughgarden 1975; Ripa and Heino 1999; Greenman and
Benton 2005; Ruokolainen and Fowler 2008; Ruokolainen
et al. 2009). Consequently, when the intrinsic dynamics are
estimated incorrectly, assessing the role of coloured envi-
ronmental variation from the remaining variation becomes
difficult.
Stronger bias was mainly restricted to situations with
overcompensatory dynamics, while results with under-
compensatory dynamics showed in general no, or very
weak bias. The return rate (TR) is a measure of the negative
feedback strength, i.e., the effect of density on per capita
growth. With weak density effects (TR close to zero), the
shape of the density dependent function matters less, and
the relationship can be reasonably well described with any
of the functions applied. This can be illustrated by noting
that when TR = 0 all the models are identical (collapsing to
exponential growth). With overcompensatory intrinsic
dynamics (TR [ 1), bias in b increased with increasing
value of b and the standard deviation of the environmental
unknown noise r. The estimated residual variance could
potentially be used to assess when bias may be a concern in
studies of natural populations, especially for the case of red
covariates and overcompensatory dynamics (Fig. 4d). The
effect of noise on bias might be related to increased pop-
ulation variance. The population trajectory will visit a
wider range of densities with more variable (known and
unknown) environmental fluctuations. These conditions
emphasize the importance of selecting the correct shape of
density dependence, as populations will spend more time
away from equilibrium conditions, where linearization
assumptions are less appropriate.
The cases studied here should not be confused with a
situation where the unknown component of environmental
stochasticity (ut) is coloured, i.e., the part of environmental
stochasticity that is not included as a covariate in the sta-
tistical model. Coloured stochasticity can be a source of
bias, e.g., when studying interspecific interactions (Ripa
and Ives 2007). Both problems might occur at the same
time with a particular set of data. A statistical model
assuming that the residuals are autocorrelated, on a second
order autoregressive model, may provide better estimates
of statistical uncertainty for the effects of coloured envi-
ronmental covariates when density dependence is modelled
with an incorrect functional form. However, those models
do not correspond to the ‘true’ situation, and the bias
described here is still present, as confirmed by preliminary
simulations (results not shown).
Prewhitening the covariates of interest appears to
remove bias in the estimates of environmental effects when
the shape of density dependence is modelled incorrectly. It
is also likely to be a good solution when estimated effects
of coloured covariates are biased by ignoring observation
error in population density (Linden and Knape 2009).
However, prewhitening is associated with lowered preci-
sion and statistical power (Pyper and Peterman 1998),
confirmed by our results. Confidence intervals also often
become rather pessimistic following prewhitening, with
coverage[95 %. According to our prewhitening approach,
the removed part of the covariate (i.e., the fitted values of
the autoregression Wt against Wt - 1) and the prewhitened
covariate (residuals) are orthogonal, with strictly zero
correlation. That may be a necessary condition to achieve
unbiased results as observed here. For example, by simply
taking the first difference (which equals the residuals of an
autoregression with AR-coefficient fixed to one) the above
condition is not satisfied and the results may still be biased.
Fitting a model with a flexible shape of density depen-
dence, such as the h-Ricker model used here, is one effec-
tive solution for avoiding bias in estimates of environmental
effects. However, fitting the h-Ricker is associated with
190 Popul Ecol (2013) 55:183–192
123

severe problems of parameter identifiability in some cases
(Polansky et al. 2009; Clark et al. 2010), and can be mis-
leading for research questions relating to the shape of the
density dependent response. When population dynamics are
concentrated to the region close to equilibrium, the joint
likelihood of parameter h and the intrinsic growth rate (a0
here, often referred to as r) is correlated, forming a shallow
ridge (Polansky et al. 2009; Clark et al. 2010). Increasing
the strength of process noise reduces the problems of
identifiability (Clark et al. 2010). Populations visit a wider
range of their phase space, highlighting non-linear behav-
iours away form the equilibrium and increasing accuracy
and confidence in parameter estimates. Whenever these
kinds of models are applied, any conclusions should be
based on careful consideration of all plausible parameter
combinations in the h - r parameter space, e.g., as deter-
mined by the profile likelihood (Polansky et al. 2009).
Another approach is to apply model selection or multi-
model inference on a set of competing models with different
shapes of density dependence (van de Meer et al. 2000;
Williams et al. 2004; Brook and Bradshaw 2006). We
showed that high R2 is associated with unbiased estimates of
environmental effects (and reliable confidence intervals),
suggesting that this approach would work, but the risk
remains that none of the candidate models considered is
sufficient. Further, model selection between different shapes
of density dependence is similar to fitting the h-Ricker with
several arbitrarily fixed values for h, and is hence likely to be
associated with similar identifiability problems as the h-
Ricker model. Nevertheless, advocating several of the fitted
models, independently or using multimodel inference, might
offer more robust conclusions. In contrast to prewhitening
covariates, neither a flexible shape of density dependence,
nor model selection will reduce bias in the environmental
effect associated with sampling and observation error in
population density (Linden and Knape 2009).
Our results show that biased estimated effects of envi-
ronmental variables are mainly of concern when coloured
covariates are coupled with an incorrect shape of density
dependence and overcompensatory intrinsic population
dynamics. The relevance of weak bias, also present with
undercompensatory dynamics, depends on the area of
application. Bias is associated with rather specific and
somewhat recognizable conditions, which is good news for
ecologists analysing natural time series. Strongly over-
compensatory and complex single species dynamics have
been suggested to be fairly uncommon (or at least, hard to
identify) in nature (Zimmer 1999; Shelton and Mangel
2011), although by no means exclusively absent (Ellner
and Turchin 1995; Turchin 1995, 2003; Bjørnstad and
Grenfell 2001; Ives et al. 2008). Sibly et al. (2007, their
Fig. S3) demonstrate a wide range of return rates estimated
from natural populations, with approximately half showing
overcompensating patterns (TR [ 1), suggesting that
overcompensatory dynamics might be more common than
previously thought, even within the region of stable equi-
librium dynamics. These estimates remain tentative how-
ever, as Sibly et al. (2007) did not account for observation
error, nor include specific environmental covariates. Fur-
ther, terrestrial environmental variables, and commonly
used climate indices (such as the North Atlantic Oscilla-
tion) are seldom strongly autocorrelated, while marine
variables are often reddened (Vasseur and Yodzis 2004). It
is important to keep in mind that problems of bias can arise
whenever the relevant (sampled) environmental and pop-
ulation time series data appear strongly coloured (espe-
cially blue) and overcompensatory, respectively, even if
that does not reflect the long-term behaviour of the
underlying processes. It is therefore the sample autocor-
relation that is of primary importance—which makes
identifying the risk more manageable. Short time series can
show strong autocorrelation simply by chance.
In conclusion, we recommend caution when working
with covariates showing strong sample autocorrelation.
This is particularly true for negatively autocorrelated
covariates coupled with overcompensatory intrinsic popu-
lation dynamics. Severe bias was typically associated with
a lowered model fit. Hence, the conventional wisdom of
not making strong inference based on a poor model fit to
data (in all aspects) remains valid and should be a guiding
principle in applied population biology. Prewhitening the
covariates, model selection and multimodel inference of
different shapes of density dependence, and models that are
flexible in describing the intrinsic population dynamics can
all be used to reduce bias in such situations. Each of these
approaches are associated with their own problems, should
be chosen carefully, and should preferably not be applied
in cases where not needed.
Acknowledgments The work was funded by the Academy of Fin-
land (AL; grant ref. 135682), MSF received support from the Spanish
Ministry of Science (grant ref. CGL2006-04325/BOS). NJ was
financially supported by the Swedish Research Council (grant ref.
2009-5155).
References
Berryman A, Turchin P (2001) Identifying the density-dependent
structure underlying ecological time series. Oikos 92:265–270
Bjørnstad ON, Grenfell BT (2001) Noisy clockwork: time series
analysis of population fluctuations in animals. Science
293:638–643
Brannstrom A, Sumpter DJT (2006) Stochastic analogues of deter-
ministic single-species population models. Theor Popul Biol
69:442–451
Brook BW, Bradshaw CJA (2006) Strength of evidence for density
dependence in abundance time series of 1198 species. Ecology
87:1445–1451
Popul Ecol (2013) 55:183–192 191
123

Cazelles B (2004) Symbolic dynamics for identifying similarity
between rhythms of ecological time series. Ecol Lett 7:755–763
Chatfield C (2004) The analysis of time series—an introduction, 6th
edn. Chapman & Hall, London
Clark F, Brook BW, Delean S, Akcakaya HR, Bradshaw CJA (2010)
The h-logistic is unreliable for modelling most census data. Meth
Ecol Evol 1:253–262
Ellner S, Turchin P (1995) Chaos in a noisy world: new methods and
evidence from time-series analysis. Am Nat 145:343–375
Fowler MS (2009) Increasing community size and connectance can
increase stability in competitive communities. J Theor Biol
258:179–188
Freckleton RP, Watkinson AR, Green RE, Sutherland WJ (2006)
Census error and the detection of density dependence. J Anim
Ecol 75:837–851
Gilpin ME, Ayala FJ (1973) Global models of growth and compe-
tition. Proc Natl Acad Sci USA 70:3590–3593
Gompertz B (1825) On the nature of the function expressive of the
law of human mortality, and on a new mode of determining the
value of life contingencies. Philos Trans R Soc B-Biol Sci
115:513–585
Greenman JV, Benton TG (2005) The frequency spectrum of
structured discrete time population models: its properties and
their ecological implications. Oikos 110:369–389
Heino M, Ripa J, Kaitala V (2000) Extinction risk under coloured
environmental noise. Ecography 23:177–184
Hilborn R, Mangel M (1997) The ecological detective: confronting
models with data. Princeton University Press, Princeton
Ives AR, Einarsson A, Jansen VAA, Gardarsson A (2008) High-
amplitude fluctuations and alternative dynamical states of
midges in Lake Myvatn. Nature 452:84–87
Johnson DH, Grier JW (1988) Determinants of breeding distributions
of ducks. Wildl Monogr 100:1–37
Jonzen N, Hedenstrom A, Hjort C, Lindstrom A, Lundberg P,
Andersson A (2002) Climate patterns and the stochastic
dynamics of migratory birds. Oikos 97:329–336
Kaitala V, Ranta E (2001) Is the impact of environmental noise
visible in the dynamics of age-structured populations? Proc R
Soc B 268:1769–1774
Knape J (2008) Estimability of density dependence in models of time
series data. Ecology 89:2994–3000
Knape J, de Valpine P (2011) Effects of weather and climate on the
dynamics of animal population time series. Proc R Soc B
278:985–992
Linden A, Knape J (2009) Estimating environmental effects on
population dynamics: consequences of observation error. Oikos
118:675–680
Lundberg P, Ripa J, Kaitala V, Ranta E (2002) Visibility of
demography-modulating noise in population dynamics. Oikos
96:379–382
May RM, Conway GR, Hassell MP, Southwood TRE (1974) Time
delays, density-dependence and single-species oscillations.
J Anim Ecol 43:747–770
McCarthy MA (1996) Red kangaroo (Macropus rufus) dynamics:
effects of rainfall, density dependence, harvesting and environ-
mental stochasticity. J Appl Ecol 33:45–53
Piha M, Linden A, Pakkala T, Tiainen J (2007) Linking weather and
habitat to population dynamics of a migratory farmland song-
bird. Ann Zool Fenn 44:20–34
Polansky L, de Valpine P, Lloyd-Smith JO, Getz WM (2009)
Likelihood ridges and multimodality in population growth rate
models. Ecology 90:2313–2320
Post E, Stenseth NC (1998) Large-scale climatic fluctuation and
population dynamics of moose and whitetailed deer. J Anim Ecol
67:537–543
Pyper BJ, Peterman RM (1998) Comparison of methods to account
for autocorrelation in correlation analyses of fish data. Can J Fish
Aquat Sci 55:2127–2140
Ranta E, Lundberg P, Kaitala V, Laakso J (2000) Visibility of the
environmental noise modulating population dynamics. Proc R
Soc B 267:1851–1856
Ricker WE (1954) Stock and recruitment. J Fish Res Board Can
11:559–623
Ripa J, Heino M (1999) Linear analysis solves two puzzles in
population dynamics: the route to extintion and extinction in
coloured environments. Ecol Lett 2:219–222
Ripa J, Ives A (2007) Interaction assessment in correlated and
autocorrelated environments. In: Vasseur DA, McCann KS (eds)
The impact of environmental variability on ecological systems.
Springer, The Netherlands, pp 111–131
Roughgarden J (1975) A simple model for population dynamics in
stochastic environments. Am Nat 109:713–736
Royama T (1981) Fundamental concepts and methodology for the
analysis of animal population dynamics, with particular refer-
ence to univoltine species. Ecol Monogr 51:473–493
Royama T (1992) Analytical population dynamics, 1st edn. Chapman
& Hall, London
Ruhe A, Wedin PA (1980) Algorithms for separable nonlinear least
squares problems. Siam Rev 22:318–337
Ruokolainen L, Fowler MS (2008) Community extinction patterns in
coloured environments. Proc R Soc B 275:1175–1183
Ruokolainen L, Linden A, Kaitala V, Fowler MS (2009) Ecological
and evolutionary dynamics under coloured environmental var-
iation. Trends Ecol Evol 24:555–563
Shelton AO, Mangel M (2011) Fluctuation of fish population and the
magnifying effects of fishing. Proc Natl Acad Sci USA
108:7075–7080
Sibly RM, Barker D, Hone J, Pagel M (2007) On the stability of
populations of mammals, birds, fish and insects. Ecol Lett
10:970–976
Stenseth NC, Mysterud A, Ottersen G, Hurrell JW, Chan K-S, Lima
M (2002) Ecological effects of climate fluctuations. Science
297:1292–1296
Turchin P (1995) Population regulation: old arguments and a new
synthesis. In: Capuccino N, Price PW (eds) Population dynam-
ics: new approaches and synthesis, 1st edn. Academic Press, San
Diego, pp 19–40
Turchin P (1999) Population regulation: a synthetic view. Oikos
84:153–159
Turchin P (2003) Complex population dynamics: a theoretical/
empirical synthesis. Princeton University Press, Princeton
van de Meer J, Beukma JJ, Dekker R (2000) Population dynamics of
two marine polychaetes: the relative role of density dependence,
predation and winter conditions. ICES J Mar Sci 57:1488–1494
Vasseur DA, Yodzis P (2004) The color of environmental noise.
Ecology 85:1146–1152
Wichmann MC, Johst K, Schwager M, Blasius B, Jeltsch F (2005)
Extinction risk, coloured noise and the scaling of variance. Theor
Popul Biol 68:29–40
Williams ID, van de Meer J, Dekker R, Beukema JJ, Holmes SP
(2004) Exploring interactions among intertidal macrozoobenthos
of the Dutch Wadden Sea using population growth models. J Sea
Res 52:307–319
Zimmer C (1999) Life after chaos. Science 284:83–86
192 Popul Ecol (2013) 55:183–192
123