marine-derived dinoflagellates in antarctic saline lakes: community composition and annual dynamics
TRANSCRIPT

MARINE-DERIVED DINOFLAGELLATES IN ANTARCTIC SALINE LAKES:COMMUNITY COMPOSITION AND ANNUAL DYNAMICS1
Karin Rengefors2
Limnology, Department of Ecology, Ecology Building, S-22362 Lund, Sweden
Johanna Laybourn-Parry
Institute for the Environment, Physical Sciences and Applied Mathematics, Keele University, Keele, Staffordshire ST5 5BG, UK
Ramiro Logares
Limnology, Department of Ecology, Ecology Building, S-22362 Lund, Sweden
William A. Marshall
Environmental Science Department, Westlakes Scientific Consulting, Princess Royal Building, Westlakes Science
and Technology Park, Cumbria CA24 3LN, UK
and Gert Hansen
Department of Biology, University of Copenhagen, Øster Farimagsgade 2D, 1353 Copenhagen K., Denmark
The saline lakes of the Vestfold Hills in Antarcticaoffer a remarkable natural laboratory where theadaptation of planktonic protists to a range of evolv-ing physiochemical conditions can be investigated.This study illustrates how an ancestral marine com-munity has undergone radical simplification leavinga small number of well-adapted species. Our objec-tive was to investigate the species composition andannual dynamics of dinoflagellate communities inthree saline Antarctic lakes. We observed that dino-flagellates occur year-round despite extremely lowPAR during the southern winter, which suggests sig-nificant mixotrophic or heterotrophic activity. Onlya small number of dominant dinoflagellate specieswere found in each lake, in contrast to the species-rich Southern Ocean from which the lake communi-ties are believed to be derived. We verified that thelake species were representatives of the marinepolar dinoflagellate community, and not freshwaterspecies. Polarella glacialis Montresor, Procaccini etStoecker, a bipolar marine species, was for the firsttime described in a lake habitat and was an impor-tant phototrophic component in the higher salinitylakes. In the brackish lakes, we found a new siblingspecies to the brackish-water species Scrippsiella han-goei (J. Schiller) J. Larsen, previously observed onlyin the Baltic Sea.
Key index words: Antarctica; cold-water dinoflagel-lates; dinoflagellate dynamics; polar; Polarellaglacialis; saline lakes; Scrippsiella hangoei
Abbreviation: aff., affinis; DIC, differentialinterference contrast; DOC, dissolved organiccarbon; ITS, internal transcribed spacer; Kd, lightextinction coefficient; PCA, principal componentanalysis
The microbial communities of Antarctic lakes areof great scientific interest for at least two fundamen-tal reasons. First, they offer a system for studyingmicrobial evolution and diversity in a geographicallyisolated region. The separation of the Antarcticfrom the other southern continents and the forma-tion of the polar front occurred over 10 millionyears ago (Vincent 2000). Consequently, organismson the Antarctic continent have either been isolatedfor millions of years or have colonized the continentfrom the sea or through long-distance dispersal(Marshall 1996). Second, the Antarctic lake commu-nities make it possible to study adaptations to anextreme environment, including very low tempera-tures and high variability in salinity conditions, incombination with long periods of little or no lightavailability. Extreme environmental conditions arelikely to exert considerable abiotic selection pres-sure on the organisms present. In particular, due tothe different physicochemical characteristics of thelakes, selection will be driven in different directionsfrom lake to lake. Thus, colonizing populationsthat previously experienced a broadly similar, lessextreme environment would be exposed to an arrayof new selective conditions.
The Vestfold Hills in Eastern Antarctica containmany marine-derived chloride-dominated lakes rang-ing in salinity from slightly brackish to hypersaline
1Received 9 April 2007. Accepted 27 September 2007.2Author for correspondence: e-mail Karin.Rengefors@limnol.
lu.se.
J. Phycol. 44, ***–*** (2008)� 2008 Phycological Society of AmericaDOI: 10.1111/j.1529-8817.2008.00517.x
1

(Ferris et al. 1988). The saline lakes were formedby isostatic uplift leaving behind pockets of marinewater, which have subsequently undergone salin-ity changes due to dilution from snowmelt orconcentration by evaporation (Adamson and Pickard1986). Consequently, some lake waters have dec-lined in salinity, becoming brackish or even nonsa-line, while others have increased in salinity, by up to10 times that of seawater, becoming hypersaline(Pickard 1986). There is good paleolimnological evi-dence to show that periods of marine incursionsand successive periods of meromixis have occurred,for example in Ace Lake (Roberts and McMinn1999, Cromer et al. 2005). In these instances, mar-ine incursions are attributed to phases when eustaticsea level changes occurred more rapidly than iso-static uplift (Hodson et al. 2004). It is proposed thatmost lakes were isolated from the sea about 6500–5500 BP (Zwartz et al. 1998). Consequently, manyof these lakes have become marine relics with abiota consisting of a subset of the marine microbialspecies (Rankin et al. 1999). The planktonic foodwebs are simple and fishless, with few or nozooplankton taxa (Laybourn-Parry 1997). Some ofthe brackish lakes have been invaded by speciesfrom the many freshwater lakes in the VestfoldHills, for example rotifers, diatoms, and the cladoc-eran Daphniopsis studeri (Laybourn-Parry andMarchant 1992, Swadling et al. 2001).
The phytoplankton communities of the salinelakes in the Vestfold Hills are dominated by smallphototrophic flagellates, including prymnesiophytesand cryptophytes (Parker et al. 1982, Perriss andLaybourn-Parry 1997). However, dinoflagellates arealso present and were found in 14 out of 16 lakes ina previous study (Perriss and Laybourn-Parry 1997).Dinoflagellates are of particular interest since theyare both diverse and abundant in the coastal waterssurrounding Antarctica (McKenzie and Cox 1991,Stoecker et al. 1992, Gast et al. 2006). Dinoflagellatesinclude species with a range of nutritional modesspanning from obligate autotrophs to mixotrophicand completely heterotrophic (Stoecker 1999). Theycontain chloroplasts from a range of different ori-gins, permanent as well as kleptoplastids (Fields andRhodes 1991, Tengs et al. 2000, Gast et al. 2007). Inaddition, dinoflagellates have a number of otheradaptive strategies, including diel vertical migra-tion (Heaney and Furnass 1980), cyst formation(Anderson and Wall 1978), and toxin production(Van Dolah 2000), to deal with competition, preda-tion, and adverse environmental conditions. Thesecharacteristics probably allowed dinoflagellates tosurvive and proliferate in the extreme environmen-tal conditions of the polar regions.
The objective of this study was to investigate thespecies composition and annual dynamics of thedinoflagellate communities in three saline Antarcticlakes. Annual dynamics have not been describedfor the individual dinoflagellate species in these
Antarctic lakes, nor have the species identities beenconfirmed by SEM or DNA sequence analyses. Here,we present data on dinoflagellate dynamics in rela-tion to environmental conditions, as well as anattempt to unravel the identity of dominant speciesin three brackish-saline Vestfold Hills lakes.
MATERIALS AND METHODS
Three lakes in the Vestfold Hills (68� S, 78� E; Fig. 1),Eastern Antarctica, were sampled from November 2002 toDecember 2003. The lakes included slightly brackish HighwayLake (salinity of 5) of 0.2 km2 and a maximum depth of 15 m;brackish Pendant Lake (salinity 16–20, hereafter referred to asbrackish-saline) with a maximum depth of 12 m and an area of0.16 km2; and the saline and meromictic Ace Lake (salinity 18in the upper mixolimnion), with a maximum depth of 20 mand an area of 0.16 km2. In Ace Lake, the chemocline occurredbetween 8 and 10 m. All three lakes are ice-covered for at least10 months per year but are usually partially or completely ice-free during late summer.
Annual sampling. The three lakes were sampled on abiweekly or monthly basis, except for a period during Marchand April when the lakes were not accessible due to logistic andsea-ice conditions. Except when lakes were ice free, a hole wasdrilled at the deepest point in each lake with a Jiffy drill (Jiff IceDrills, Sheboygon Falls, WI, USA). The water column wassampled at 2 (immediately under the ice), 4, 6, and 8 m depthin all three lakes. Ace Lake was also sampled at 10 and 12 mdepth. Samples of 2.5 L were collected using a Kemmerersampler. Subsamples of 1 L were preserved with Lugol’ssolution for dinoflagellate counts, concentrated by settling,and counted in a Sedgewick–Rafter counting chamber,(Graticules Ltd., Tunbridge Wells, UK) under differential
Fig. 1. A map of the Vestfold Hills showing the larger freshwa-ter and saline lakes. The study lakes are situated on Long Penin-sula as follows: 1, Ace Lake; 2, Highway Lake; 3, Pendant Lake.
2 KARIN RENGEFORS ET AL.

interference contrast (DIC) and phase microscopy (ZeissAxioskop; Carl Zeiss, Oberkochen, Germany) at ·320. Epiflu-orescence microscopy (Zeiss Axioskop) was used on freshsamples at the time of collection to determine whetherparticular species were phototrophic or heterotrophic. Othersubsamples were analyzed for pH, conductivity, temperature,inorganic nutrients (NH4, PO4, NO3), dissolved organic carbon(DOC), and chl a. Aliquots (1 L) were filtered through ashed47 mm GF ⁄ F filters (Whatman, Maidstone, UK), and thefiltrate was used for chemical analyses of nitrate (NO3-N) andammonium (NH4-N) content according to the methods ofParsons et al. (1984), and soluble reactive phosphorus (PO4-P)using the protocol of Eisenriech et al. (1975). DOC wasmeasured using a Shimadzu TOC 5000 carbon analyzer (GMIInc., Ramsey, MN, USA). Chl a on the filters was extracted inmethanol at )20�C overnight and assayed spectrophotometri-cally according to the method of Talling (1974).
PAR profiles, using a PAR sensor (LI-COR LI-192 SA, flatcosine collector; LI-COR, Lincoln, NE, USA), were measuredon the ice surface and at 1 m depth intervals. Ice thickness andsnow cover were measured on each sampling occasion.
Dinoflagellate abundance. Dinoflagellate mean cell countswere calculated for the top 8 m of the lakes by making volume-weighted calculations. Volumes of 2 m depth layers (0–2, 2–4,4–6, 6–8) were estimated using lake area, maximum depth, andmean depth. Since the morphometry of the lakes was unknown, aconical shape for the lake was assumed. The volume of eachdepth interval was then calculated by assuming a circular surfacearea and using the formula V = mean depth · radius2 · 1.45(Wetzel 2001). The contribution (weight) of each 2 m layer inthe lake was determined by setting the smallest (6–8 m) volumeto 1. The cell counts for each 2 m layer were then averaged,taking the weight of each into account. In other words, surfacelayers with the largest volume usually accounted for 1.6· that ofthe deepest layer. Integrated cell counts were of the same orderas simple nonweighted averaging of the abundance at differentdepths and differed by 5%–25%. Physicochemical parametermeans were calculated by averaging the top 8 m.
Light climate. To compare the light climate in the threelakes, the extinction coefficient, Kd, was calculated for eachlake and each sampling occasion using the equation (Kirk1994): Ez = Eo · e)Kdz. Ez is the light intensity at depth z, and E0
at the depth above, using 2 m intervals. Kd was determinedfrom log-linear regressions using Microsoft Excel (MicrosoftCorp., Redmond, WA, USA).
Statistical analyses. To determine which environmentalfactors may have influenced the dinoflagellate abundance ineach lake, a principal component analysis (PCA) was performed.All the measured physicochemical data and Mesodiniumabundance (a dinoflagellate competitor) were included. Allvalues were natural log transformed to achieve normality.Components with Eigen values >1 were retained and tested forstatistical significance with regression analysis. The factors ofsignificant components with an absolute loading >0.5 wereconsidered to be important and were subsequently tested one byone by Spearman’s correlation analysis against dinoflagellateabundance.
Key lake parameters (pH, conductivity, temperature, nutri-ents, DOC, dinoflagellate abundance) were compared amonglakes using one-way analysis of variance (ANOVA). All statisticalanalyses were performed using SPSS v. 11 for Macintosh (SPSSInc., Chicago, IL, USA).
Establishment of single-cell cultures, DNA extraction, sequencing,and genetic analyses. In the Antarctic summer of 2004, dinofla-gellates were isolated from different lakes in the Vestfold Hillsarea. Tow samples using 10 lm plankton sieves (Swedaq, Hoor,Sweden) were taken from lakes included in this study, as well assome additional lakes. Single cells were isolated from towsamples using microcapillary pipettes. Cells were placed in f ⁄ 2
medium based on GF ⁄ C-filtered lake water. The two speciesthat survived in culture were later identified based onribosomal DNA sequences and SEM analyses.
DNA was extracted from clonal culture strains (our isolatesand the public available ones CCMP1383 ⁄ 2088) following aphenol-chloroform protocol (see Adachi et al. 1994). For thiswork, we used partial SSU rDNA. SSU PCR amplifications wereperformed using 25 ng of template genomic DNA, 0.125 mMof each nucleotide, 3.0 mM MgCl2, 1· PCR buffer, 0.4 lM ofeach primer, and 0.5 U of Taq DNA Polymerase (AmpliTaq;Applied Biosystems, Foster City, CA, USA) in 25 lL totalvolume reactions. For the SSU PCR, we used the combination ofthe universal primers 4616 (forward), 5¢-AACCTGGTTGATCC-TGCCAG-3¢, and 4618 (reverse), 5¢- TGATCCTTCTGCAGGTT-CACCTAC-3¢. The SSU PCR started with 5 min at 94�C; followedby 35 cycles of 1 min at 94�C, 1.5 min at 55�C, 2 min at 72�C; andending with 7 min at 72�C. The complete PCR product was�1,700 bp. For one strain, S. aff. hangoei, the small subunitribosomal size was 1,232 bp since we were not able to sequencethe entire fragment, although the complete PCR product wasalso �1,700 bp.
All PCR amplicons were cleaned using PCR-MTM Clean-UpSystem (Viogene, Taiwan). SSU amplicons were directlysequenced using the PCR primer 4616, plus the sequencingprimers 516F (5¢-CACATCTAAGGAAGGCAGCA-3¢), 528F(5¢-CGGTAATTCCAGCTCC-3¢), 690F (5¢-CAGAGGTGAAATT-CT-3¢), and 1055F (5¢-GGTGGTGCATGGCCG-3¢; Edvardsenet al. 2003). The sequencing reaction was carried out usingBigDye (v1.1, Applied Biosystems) chemistry, and the productswere precipitated according to the manufacturer’s instructionsand then loaded into an ABI Prism 3100 sequencer (AppliedBiosystems). The obtained sequences were edited and assem-bled by carefully analyzing the chromatograms using Bioedit(ver. 7.0.4.1; Hall 1999). Sequences were deposited in GenBankunder the accession numbers EF434275, EF417317/6/7, andEF417318. The obtained SSU sequences were manually alignedand visually analyzed, and the sequences were aligned alongwith the most similar sequences obtained by a BLAST search inGenBank (Peridinium aciculiferum Lemmermann ⁄ S. hangoei,and P. glacialis) using ClustalX (v1.8; Thompson et al.1997). The percentage of nucleotide differentiation betweenthese species was calculated.
SEM analyses. Two different fixation protocols were usedfor SEM preparation. Protocol 1, used for the species identifiedas P. glacialis (Abraxas-strain), was a slightly modified version ofParducz’s (1967) protocol: 300 lL culture was added to 300 lLfixative cocktail consisting of 200 lL saturated HgCl2 and100 lL OsO4. Cells were fixed for 30 min while settling on apoly-l-lysine covered coverslip. Protocol 2 was used for thespecies identified as S. aff. hangoei (strain High-4): Cells werefixed in 5% glutaraldehyde and stored at 5�C for nearly10 months. Cells were then concentrated onto a 5 lm Isoporefilter (Millipore, Billerica, MA, USA) for further processing.
In both protocols, the material was washed in DH2O for 1 hand dehydrated in an ethanol series: 30%, 50%, 70%, 96%, and99.9% for 10 min in each change and finally twice in 100%ethanol for 30 min in each change, achieved by the addition ofmolecular sieves to 99.9% ethanol. Critical-point-drying wasmade using a BAL-TEC CPD-030 (Boeckeler Instruments Inc.,Tucson, AZ, USA). Filters or coverslips were mounted on stubsand coated with Pt and examined using a JEOL JSM-6335F(JEOL Ltd., Tokyo, Japan) field emission scanning electronmicroscope.
RESULTS
Dinoflagellate species composition. The dinoflagellatecommunities in the saline lakes were typically domi-
DINOFLAGELLATES IN ANTARCTIC SALINE LAKES 3

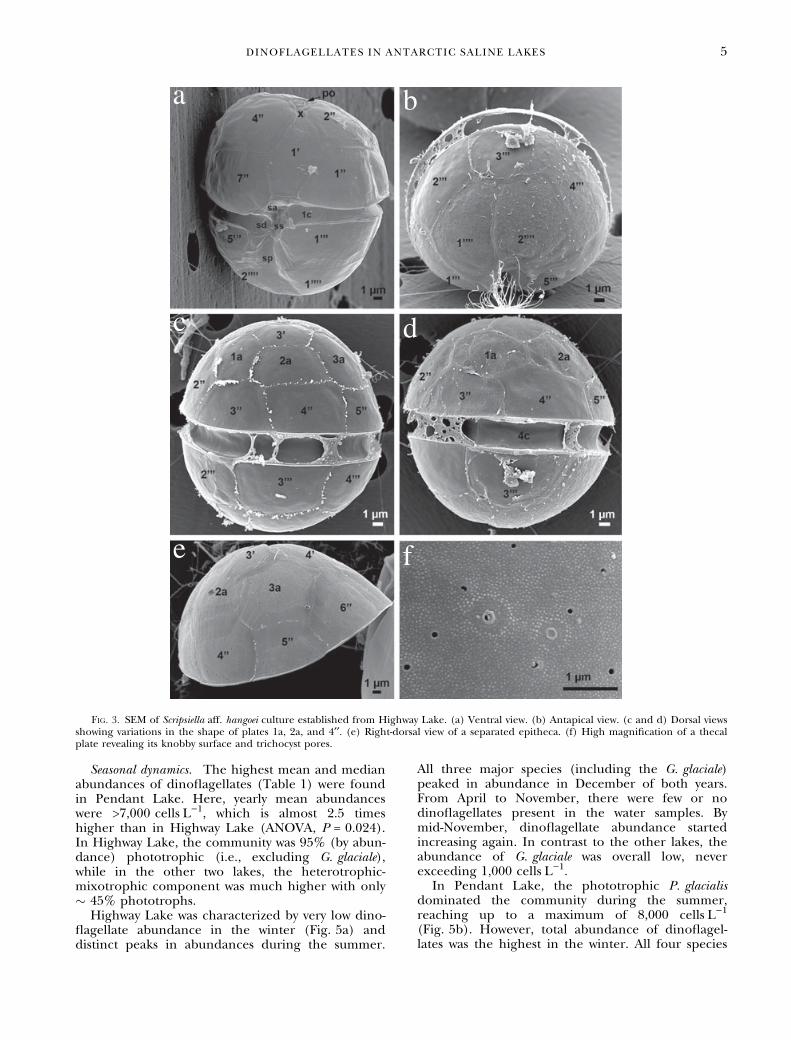
nated by only three to four dinoflagellate species,including heterotrophic taxa. In the slightly brack-ish Highway Lake (salinity �5), there were twodominant phototrophic species; one (or more)unidentified naked gymnodinoid species referred toas ‘‘Gymnodinium’’ spp. of 15–25 lm (Fig. 2a) andone thecate species. Using our single-cell isolatedcultures, the thecate species could be identified bySEM and the partial SSU sequence. The plate pat-tern corresponded to that of brackish S. hangoei.Some variations were noted in the shape of plates1a, 2a, and 4¢¢, similar to the variations observed byLarsen et al. (1995). Thus, in some cells, plate 4¢¢was almost quadrangular and separated from plates1a (penta), which was five-sided (Fig. 3c). In othercells, plate 1a was six-sided and shared a side(suture) with plate 1a. Penta and hexa configura-tions of plate 2a were also observed (not shown).When analyzed, the S. aff. hangoei partial SSUsequences turned out to be very similar (0.2% dif-ference in 1,200 bp) yet not identical to those ofthe morphospecies S. hangoei ⁄ Per. aciculiferum Lem-merm. (data not shown). Since the partial SSUsequence was not identical, and other sequences[e.g., internal transcribed spacer (ITS), Logareset al. 2008] also had discrepancies, we could notwith any certainty determine whether it is the samespecies as the Baltic S. hangoei cultures. Thus, wedecided to use the term affinis (aff.) as having affin-ity with but not identical with. The third dominantspecies was the marine heterotrophic species Gyrodi-nium glaciale Hada, which was identified by SEM
analyses (Fig. 2b) from plankton tows. In previousstudies (Perriss and Laybourn-Parry 1997, Laybourn-Parry et al. 2002), this species was referred to asG. lachryma (Meunier) Kof. et Swezy, but on thebasis of its small size and large girdle displacement,we suggest that it is a G. glaciale. Balech (1976)observed small circular chloroplasts in this species(probably kleptoplasts). Although we did notobserve these, their presence could not be ruledout.
In Pendant Lake (salinity 16–20), there were fourdominant dinoflagellate species. These includedG. glaciale, the S. aff. hangoei species also found inHighway Lake, ‘‘Gymnodinium’’ spp. (Fig. 2), andthe marine bipolar species P. glacialis (Montresoret al. 1999, 2003; Fig. 4). P. glacialis was the domi-nant phototrophic dinoflagellate in Pendant Lake.The complete SSU rDNA sequences (from culturesisolated in Ekho and Abraxas lakes since no culturesurvived from Pendant Lake) were identical to theP. glacialis clones CCMP 1383 from Antarctica(McMurdo Sound) and CCMP 2088 from the Arctic(Baffin Bay, Canada). The two CCMP sequenceswere identical to each other when we sequencedthem anew, contrary to the findings of Montresoret al. 2003. SEM of both vegetative cells and cystscorresponded to the general morphology of P. gla-cialis (Fig. 4).
In saline, meromictic Ace Lake (17–34 psu), thedinoflagellate community was dominated by theheterotrophic or possibly mixotrophic G. glaciale,‘‘Gymnodinium’’ spp., and low numbers of P. glacialis.
Fig. 2. SEM and LM images of Lugol’s preserved ‘‘Gymnodinium’’ spp. cells from Highway Lake (a–c), and SEM and LM images ofGyrodinium glaciale in natural samples from Ace Lake (d–e). SEMs courtesy of Fiona Scott, Antarctic Australian division; cells fixed inglutaraldehyde and prepared as described in Marchant and Scott (2005).
4 KARIN RENGEFORS ET AL.
Seasonal dynamics. The highest mean and medianabundances of dinoflagellates (Table 1) were foundin Pendant Lake. Here, yearly mean abundanceswere >7,000 cellsÆL)1, which is almost 2.5 timeshigher than in Highway Lake (ANOVA, P = 0.024).In Highway Lake, the community was 95% (by abun-dance) phototrophic (i.e., excluding G. glaciale),while in the other two lakes, the heterotrophic-mixotrophic component was much higher with only� 45% phototrophs.
Highway Lake was characterized by very low dino-flagellate abundance in the winter (Fig. 5a) anddistinct peaks in abundances during the summer.
All three major species (including the G. glaciale)peaked in abundance in December of both years.From April to November, there were few or nodinoflagellates present in the water samples. Bymid-November, dinoflagellate abundance startedincreasing again. In contrast to the other lakes, theabundance of G. glaciale was overall low, neverexceeding 1,000 cellsÆL)1.
In Pendant Lake, the phototrophic P. glacialisdominated the community during the summer,reaching up to a maximum of 8,000 cellsÆL)1
(Fig. 5b). However, total abundance of dinoflagel-lates was the highest in the winter. All four species
a b
c d
e f
Fig. 3. SEM of Scripsiella aff. hangoei culture established from Highway Lake. (a) Ventral view. (b) Antapical view. (c and d) Dorsal viewsshowing variations in the shape of plates 1a, 2a, and 4¢¢. (e) Right-dorsal view of a separated epitheca. (f) High magnification of a thecalplate revealing its knobby surface and trichocyst pores.
DINOFLAGELLATES IN ANTARCTIC SALINE LAKES 5

were present in the water from November until theend of June, when solar radiation was close to zero.At this time, a peak of G. glaciale up to 15,000cellsÆL)1 formed and then remained in the waterthroughout the winter. By the end of November,the other species appeared in the water samplesagain.
Ace Lake also had a low abundance of photo-trophic species (P. glacialis and ‘‘Gymnodinium’’spp.) throughout the year, which did not increasesubstantially even during summer (Fig. 5c). TheP. glacialis maximum was only �1,000 cellsÆL)1.Instead, G. glaciale dominated the community asin Pendant Lake, reaching a maximum of 15,000cellsÆL)1 in May.
In all lakes, the ciliate Mesodinium rubrum (=Myro-necta rubra) was also enumerated, as this organism isfunctionally a phototrophic and mixotrophic spe-cies, similar in size to the dinoflagellates. Thus,M. rubrum is a potential dinoflagellate competitor.Moreover, it dominated the ciliate communities ofthe lakes and could contribute significantly to phy-toplankton photosynthesis, up to 26% in Ace Lakeand 40% in Highway Lake (Laybourn-Parry and Per-riss 1995). The cell counts showed significantly
higher mean abundance (19,075 cellsÆL)1) in High-way Lake compared with Pendant and Ace lakes(1,173 and 5,845 cellsÆL)1, respectively; Table 1). Inboth Highway and Ace Lakes, Mesodinium was pres-ent in the water throughout the year (Fig. 6).
Physicochemical parameters and dinoflagellate abun-dance. The PCA analyses were generally unable toreduce the number of parameters that may affectthe variance in dinoflagellate abundance, exceptin Pendant Lake (Table 2). Dinoflagellate abun-dance in Highway Lake had a significant(P < 0.05) negative correlation with conductivity,nitrate, Kd, and DOC, while there was a positivecorrelation with pH (P < 0.01, r = 0.68) and tem-perature (P < 0.01). In contrast, dinoflagellateabundance in Pendant Lake was negatively corre-lated to both orthophosphate and nitrate (Fig. 7;P < 0.05), and no other factors. In Ace Lake,there was a positive correlation (P < 0.05) betweenlight extinction (Kd) and dinoflagellate abundance(high numbers of G. glaciale in the winter whenKd was high). Orthophosphate, DOC, and conduc-tivity were negatively correlated with dinoflagellatecell concentration (Fig. 8). No parameter wassignificant in all three lakes.
a b
Fig. 4. SEM images of Polarella glacialis culture established from Lake Abraxas (salinity similar to Pendant Lake); morphology identicalto cells from Pendant and Ace lakes. (a) Ventral view of a vegetative cell. (b) Spiny resting cyst.
Table 1. Annual mean abundance of dinoflagellates and Mesodinium.
LakeChl a, mean
(lgÆL)1)
Dinoflagellateabundance, mean
(cellsÆL)1)
Dinoflagellateabundance, median
(cellsÆL)1)
Phototrophicdinoflagellate abundance
(cellsÆL)1)Proportion phototrophic
dinoflagellates
Mesodinium rubrumabundance(cellsÆL)1)
Highway 2.2ab 2,645b 224 2,532 0.96 19,075ab
Pendant 4.4b 7,076b 4,402 3,309 0.46 1,173b
Ace 4.2a 4,829 3,682 2,057 0.42 5,845a
Significant difference (ANOVA, LSD post hoc, P < 0.001) between pairs with the same letter. Phototrophic dinoflagellatesincluded all species except Gyrodinium glaciale.
6 KARIN RENGEFORS ET AL.

The main difference among the lakes was salinity,in this study measured as conductivity (Table 3,Fig. 8). While Ace Lake had an annual mean con-ductivity (0–10 m) of 24.3 mSÆcm)1, Pendant Lakehad 18.6 mSÆcm)1, and Highway Lake only6.3 mSÆcm)1. Pendant and Ace lakes were both sig-nificantly different from Highway Lake (P < 0.001).In addition, both Pendant and Ace lakes had highersalinities in the deeper layers of the water column.The seasonal variations were small in Highway Lakebut varied between 15 and 25 in the upper mixolim-nion of meromictic Ace Lake, presumably due tothe melting of the ice cap (Fig. 9). Ace Lake had anaverage pH of 8.01, which was significantly higherthan the pH of the other two lakes, which wereclose to neutral (7.1–7.2; P < 0.01).
Another significant difference among the threelakes was their trophic status, as determined byorthophosphate concentrations. Orthophosphate inPendant and Ace lakes had an annual mean of 41.3and 43.7 lgÆL)1, respectively, and was thus higher(P < 0.05) than in Highway Lake (mean10.6 lgÆL)1).Nitrate and ammonium concentrations were low inall three lakes and did not differ significantly. Theseasonal dynamics of nitrate were very similar in alllakes, with very low levels (below 30 lgÆL)1) in thesummer months and a maximum in late winter (Sep-tember) reaching 150 lgÆL)1 (Fig. 7). Orthosphatemaxima were observed in spring ⁄ early summer in alllakes and reached high levels (>200 lgÆL)1) inPendant and Ace lakes in November ⁄ December2003.
Average annual temperature did not differ signifi-cantly among the lakes (Table 3); nevertheless,there were differences in the dynamics. While thewinter temperature in all three lakes was close to)1�C, summer temperature varied more. The maxi-mum temperature in Ace Lake was 6.5�C; in High-way Lake, 4�C; and in Pendant Lake, 1�C. Icethickness data (not shown) revealed that Highwayand Ace lakes were ice free in December and Janu-ary, while Pendant Lake retained most of its icecover, accounting for the differences in tempera-ture. There were no differences in light availabilityamong the lakes, despite differences in ice thicknessduring summer. The calculated average and medianKd values were low and similar in all lakes. DOC lev-els were also relatively low and similar for all lakes.
DISCUSSION
In this study, we investigated the species composi-tion and seasonal dynamics of dinoflagellates inthree Antarctic lakes, which differed in salinity. Tothe best of our knowledge, this is the first study on
Fig. 5. Dinoflagellate abundance (cellsÆL)1) from November2002 to January 2004, volume weighted and averaged from sam-ples from 2, 4, 6, and 8 m depth. (a) Highway Lake. (b) PendantLake. (c) Ace Lake.
Fig. 6. Mesodinium rubrum abundance (cellsÆL)1) in all threelakes, volume integrated averages from 2 to 8 m, November 2002to January 2004. Highway Lake (filled triangles), Pendant Lake(filled circles), and Ace Lake (open squares).
DINOFLAGELLATES IN ANTARCTIC SALINE LAKES 7
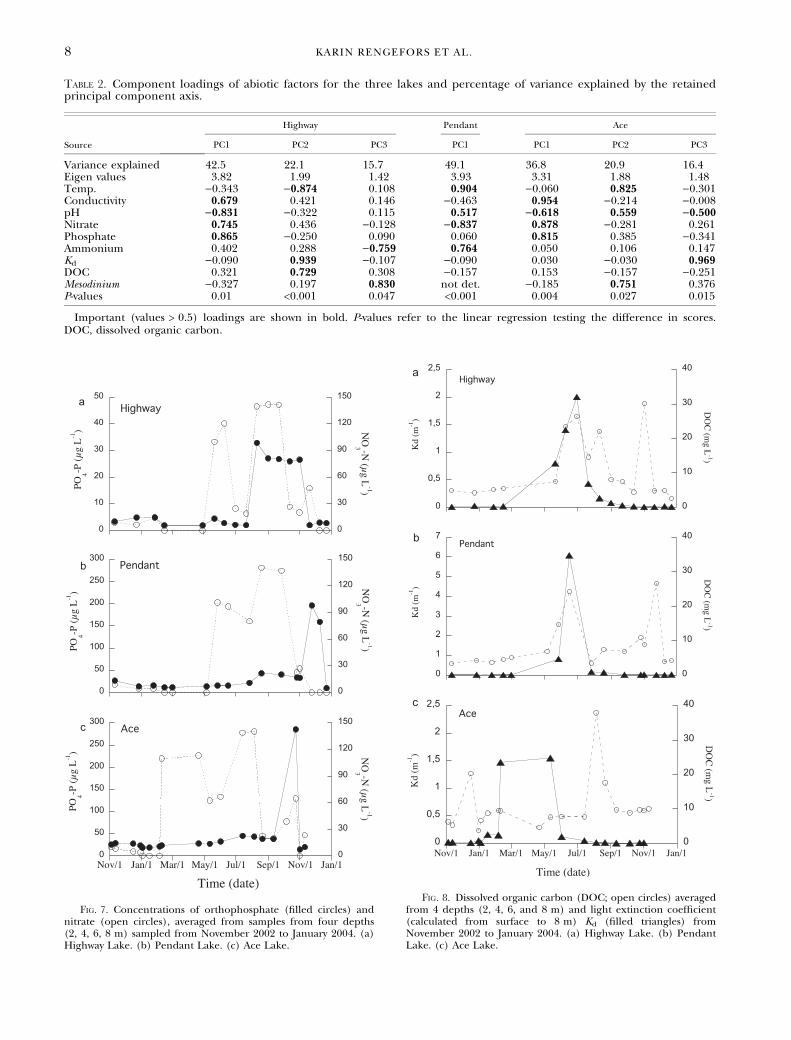
Table 2. Component loadings of abiotic factors for the three lakes and percentage of variance explained by the retainedprincipal component axis.
Source
Highway Pendant Ace
PC1 PC2 PC3 PC1 PC1 PC2 PC3
Variance explained 42.5 22.1 15.7 49.1 36.8 20.9 16.4Eigen values 3.82 1.99 1.42 3.93 3.31 1.88 1.48Temp. )0.343 )0.874 0.108 0.904 )0.060 0.825 )0.301Conductivity 0.679 0.421 0.146 )0.463 0.954 )0.214 )0.008pH )0.831 )0.322 0.115 0.517 )0.618 0.559 )0.500Nitrate 0.745 0.436 )0.128 )0.837 0.878 )0.281 0.261Phosphate 0.865 )0.250 0.090 0.060 0.815 0.385 )0.341Ammonium 0.402 0.288 )0.759 0.764 0.050 0.106 0.147Kd )0.090 0.939 )0.107 )0.090 0.030 )0.030 0.969DOC 0.321 0.729 0.308 )0.157 0.153 )0.157 )0.251Mesodinium )0.327 0.197 0.830 not det. )0.185 0.751 0.376P-values 0.01 <0.001 0.047 <0.001 0.004 0.027 0.015
Important (values > 0.5) loadings are shown in bold. P-values refer to the linear regression testing the difference in scores.DOC, dissolved organic carbon.
Fig. 7. Concentrations of orthophosphate (filled circles) andnitrate (open circles), averaged from samples from four depths(2, 4, 6, 8 m) sampled from November 2002 to January 2004. (a)Highway Lake. (b) Pendant Lake. (c) Ace Lake.
Fig. 8. Dissolved organic carbon (DOC; open circles) averagedfrom 4 depths (2, 4, 6, and 8 m) and light extinction coefficient(calculated from surface to 8 m) Kd (filled triangles) fromNovember 2002 to January 2004. (a) Highway Lake. (b) PendantLake. (c) Ace Lake.
8 KARIN RENGEFORS ET AL.

dinoflagellate community dynamics in landlockedcoastal saline lakes. In these Antarctic lakes, wefound at least two species that have not beendescribed in saline limnic habitats, one possibly anew species. All the dinoflagellate species that weidentified to the species level in the lakes were rep-resentative of the polar seawater dinoflagellate com-munity, and not freshwater species. Although
species diversity was much lower than in the polarseas, the total abundance was higher. Finally,we observed that the dinoflagellates occur year-round despite extremely low PAR during winter,suggesting significant mixotrophic or heterotrophicactivity.
The dinoflagellate communities in the salineVestfold Hills lakes had small numbers of species,which is in sharp contrast to the species-rich polarwaters. In a study of the Scotia Sea and PolarFront Zone, 63 armored dinoflagellates were identi-fied and enumerated (McKenzie and Cox 1991).McMinn and Scott (2005) listed 70 common Antarc-tic dinoflagellates plus an additional 20 species,which are rarely reported from Antarctic waters.Although we were not able to identify or enumeratea few rare species in our lakes, we concluded thatthe total dinoflagellate species richness is substan-tially reduced (10- to 20-fold) compared with thepolar marine communities. These findings corre-spond to previous surveys of these lakes’ summerplankton communities (Perriss and Laybourn-Parry1997). Moreover, although species richness waslower in the lakes studied than the nearby EllisFjord, total dinoflagellate abundance was muchhigher. In the tidal Ellis Fjord, 13 dinoflagellate spe-cies were identified and counted during winter andspring (May–October; Grey et al. 1997). The totalabundance never amounted to more than 100cellsÆL)1, which is substantially less than the thou-sands of dinoflagellates encountered during winterand spring in the studied Antarctic lakes. Likewise,a relationship between high diversity ⁄ low biomasshas been observed in Antarctic deep-sea plankton(Lopez-Garcıa et al. 2001).
The low species richness of dinoflagellates in ourlakes supports the predictions of island biogeogra-phy theory (Macarthur and Wilson 1967). This the-ory can be directly applied to the Antarctic salinelake system, where the lakes can be considered asseparate islands, and the ocean as the mainland.Island theory explains species richness as a balancebetween immigration and extinction rate, and thatthere is a positive relationship between species rich-ness and island (lake) area. Accordingly, small lakeswould be expected to hold a lower number of spe-cies than the ocean. Smith et al. (2005) recently
Table 3. Annual median and mean values of the physicochemical parameters.
Lake Annual median/mean pHTemp.(�C)
Conductivity(mSÆcm)1)
PO4-P(lgÆL)1)
NH4-N(lgÆL)1)
NO3-N(lgÆL)1)
DOC(mgÆL)1)
Kd
(m)1)
Highway Median 7.2 1.0 6.3 3.9 16.2 21.9 6.5 0.03Mean 7.2a 0.9 6.3ab 10.6ab 18.3 46.3 11.0 0.32
Pendant Median 6.9 )0.3 18.8 18.6 13.3 6.3 6.0 0.02Mean 7.1b )0.1 17.8a 41.3a 15.4 38.9 8.7 0.74
Ace Median 8.1 )0.8 24.3 27.7 10.9 20.9 8.9 0.02Mean 8.01ab 0.72 23.2b 43.7b 18.4 44.7 10.6 0.21
Statistically significant differences (ANOVA, LSD post hoc, P < 0.01 for conductivity and pH, and P < 0.05 for phosphate)between pairs with the same letter. DOC, dissolved organic carbon.
Fig. 9. (a) Conductivity (mSÆcm)1) as a proxy for salinity, aver-aged from 4 depths (2, 4, 6, 8 m) for all three lakes. (b) Temper-ature (�C) averaged from 4 depths (2, 4, 6, 8 m) for all threelakes. Highway Lake (filled triangles), Pendant Lake (filled cir-cles), and Ace Lake (open squares).
DINOFLAGELLATES IN ANTARCTIC SALINE LAKES 9

showed that phytoplankton adhere to this species-area relationship and observed that ecosystem sizehad a strong effect on local phytoplankton speciesrichness. However, the effects of local filtering mayhave restricted the Antarctic lake species pool fur-ther. These lakes represent habitats that are quitedifferent from the sea, with variable salinity, lownutrient levels (except Pendant), extreme light limi-tation, and different predators. Both nutrient levelsand predation are known to have strong effects ondiversity (Worm et al. 2002), as may island (lake)age (Borges and Brown 1999). Moreover, the factthat the lakes are young and have undergone physi-cal evolution (salinity changes) could also contrib-ute to the low species numbers, since changingconditions may have resulted in unusually highextinction rates or low colonization rates.
Another saline lake reported to contain marinedinoflagellates is the Salton Sea in California. Thislarge (975 km2) shallow lake is located in the Cali-fornian desert and is inhabited by raphidophytesand haptophytes in addition to marine dinoflagel-lates (Reifel et al. 2002). Twenty-two species ofdinoflagellates have been observed in Salton Seawaters, as well as several unidentified dinoflagellates(Reifel et al. 2002, K. Steidinger, unpublished data).Just as in the Antarctic lakes, the Salton Sea appearsto contain fewer dinoflagellate species than marineecosystems. However, the Salton Sea has a largernumber of species than the Antarctic lakes, which isconsistent with the species-area relationship, as theSalton Sea has a lake area that is almost 5,000 timeslarger than the Antarctic lakes. In contrast to theVestfold Hills lakes, the Salton Sea has never beenconnected to the ocean, has a temperature rangefrom 17�C to 21�C and a salinity of �43, and is veryyoung (100 years; Reifel et al. 2002).
Total dinoflagellate abundance was the lowest inthe brackish Highway Lake. This was also the lakewith the highest proportion of phototrophic dino-flagellates (i.e., S. aff. hangoei and ‘‘Gymnodinium’’spp.). Highway Lake is the lake that appears tohave undergone the most significant change sinceits isolation from the ocean. The salinity hasdecreased from >30 down to 5. Consequently, sig-nificant selection pressure for species tolerant oflow salinities would be expected in this lake. Salin-ity differences impose an important physiologicalbarrier to many species, and a recent study showsthat only a few dinoflagellate taxa have made theevolutionary transitions between marine and fresh-water environments (Logares et al. 2007b). Never-theless, several dinoflagellates (e.g., S. hangoei;Logares et al. 2007a) can tolerate wide salinityranges (Kremp et al. 2005). Thus, it is likely that adiversity of dinoflagellates similar to that observedtoday in Ellis Fjord was trapped in Highway Lake.Following the break of the marine connection andas salinity declined due to freshwater input, mostspecies became extinct.
The species we identified in the Vestfold Hillslakes were either identical to or similar to knownmarine species. In a previous study (Perriss andLaybourn-Parry 1997), Gonyaulax cf. tamarensis wasreported as the most common phototrophic thecatetaxa in the brackish lakes (including Highway andPendant). Although G. tamarensis has been trans-ferred to Alexandrium tamarense, we have no evidencethat this was the species observed previously, northat A. tamarense was present in the lakes. We sug-gest that ‘‘G. cf. tamarensis’’ was misidentified and isthe S. aff. hangoei species that we have described.The plate pattern of this species corresponded tothat of S. hangoei as described by Larsen et al.(1995). However, the partial SSU rDNA region dif-fered in a few base pairs (�0.2%) from the S. han-goei isolated and sequenced from the Baltic Sea(Logares et al. 2007a). Although the difference issmall, the sequences are not identical, suggestingthat these populations have not diverged veryrecently. As we have earlier observed identical SSU,LSU, and ITS sequences of S. hangoei and Per. acicu-liferum, despite different morphology, physiology,and habitat (Logares et al. 2007a), we cannot claimwith any certainty that the Baltic S. hangoei and theAntarctic S. aff. hangoei are the same species. Thetwo populations may have undergone rapid allopat-ric speciation, but this issue will be further discussedelsewhere (Logares and Rengefors, unpublisheddata).
Most likely, S. aff. hangoei in the Vestfold Hillslakes originates from a population in the polarocean (rather than long-distance dispersal from theBaltic Sea). Although we observed cells that looklike the S. aff. hangoei species in our samples fromthe sea, we were not successful in establishing thesein culture and thereby unable to confirm their iden-tity. These observations indicate that S. aff. hangoeican tolerate a salinity between 25 and 35. Despitethis, S. aff. hangoei was not observed in Ace Lakenor any of the hypersaline lakes (Perriss andLaybourn-Parry 1997), suggesting low tolerance tohigh salinities. Similarily, the sibling species S. han-goei from the brackish Baltic Sea has been shown tohave a salinity tolerance range from 0 to 30 (Log-ares et al. 2007a), yet is not found in the adjacentsaline North Sea. Possibly, it is the salinity fluctua-tions, rather than just high salinity that preventS. aff. hangoei from becoming common in Ace Lake,but further experimentation is necessary to eluci-date this point.
In the more saline lakes (Pendant and Ace), wefound that the bipolar marine species P. glacialis wasa common and abundant component of the dino-flagellate community. This species was recentlydescribed from Antarctic and Arctic seawater and ice(Montresor et al. 1999, 2003, Thomson et al. 2004).In contrast to S. aff. hangoei, the SSU sequenceswere identical for all the P. glacialis strains thatwe sequenced. Moreover, inspection of SEM
10 KARIN RENGEFORS ET AL.

micrographs of an unidentified gymnodinoid spe-cies and its cysts from McMurdo Sound, Antarctica,in Stoecker et al. (1992) suggests that this is alsoP. glacialis. The various cold-water habitats in whichP. glacialis occurs indicate that it has a wide salinitytolerance. However, in the least saline lake (High-way), P. glacialis was not observed, suggesting thatthere could be a minimum salinity requirement forthis species. However, other factors, such as competi-tion and predation, cannot be ruled out. Indeed, incontrast to the other lakes, Highway Lake is inhabitedby the large freshwater cladoceran D. studeri, a poten-tial grazer.
In the saline lakes studied, many of the dinofla-gellates were encountered year-round despite theextremely low PAR during a large portion of theyear. This finding implies that most species wereeither mixotrophic or heterotrophic. In an environ-ment with a poor light climate, phototrophic dino-flagellates have two possible strategies. They caneither form cysts and rest during the periods of lowlight and cold temperatures, as is the case for manymarine and freshwater dinoflagellates (Dale 1983,Rengefors 1997). The other option is to adoptmixotrophic nutrition and acquire carbon throughosmo- or phagotrophy (Caron et al. 1990, Hansenet al. 1994, Jacobson and Anderson 1996, Legrandand Carlsson 1998). P. glacialis cysts were observedin the water samples, and in July to October, cellnumbers were at or close to zero. The data suggestthat this species overwinters in the benthos, but it ispossible that some cells survive in the water, as ourcell counts indicate. Cysts (equivalent to thoseobserved in the field) were also formed in old clo-nal cultures. The S. hanogei–like taxon was absentfrom the water samples most dates during May–October, suggesting that this species also forms over-wintering cysts. Cysts were observed in clonal culturesof this species as well as in the Baltic S. hangoei(Kremp and Parrow 2006). Nevertheless, it is possiblethat both species have the additional potentialfor mixotrophic nutrition, utilizing osmotrophy orphagotrophy to supplement photosynthesis.
G. glaciale was the dominant heterotrophic dino-flagellate in all three lakes. We consider the speciesreferred to as G. lachryma by Laybourn-Parry (1997)and Laybourn-Parry et al. (2000, 2002) as G. glaciale,since the cells in our material were not larger than25 lm, while G. lachryma have large cells rangingfrom 60 to 135 lm long (Balech 1976). In Pendantand Ace lakes, this species forms a population peakin May–June, corresponding to early to mid-winter.The reason for the absence of this peak in HighwayLake is unknown, but it could be related to thelower salinity or higher abundance (>20,000 cellsÆL)1
during winter) of the mixotrophic ciliate M. rubrum,a potential competitor, in this lake. The ‘‘Gymnodini-um’’ spp. in the three lakes did not survive in cul-ture; their identity therefore remains uncertain andcan only be resolved by a future study.
Contrary to what one might suppose, Antarcticlakes function throughout the year. The ice usuallylacks snow cover, allowing good transmission of light.Photosynthesis starts as soon as the light returns inlate winter, and bacterial production continuesthroughout the winter (Laybourn-Parry 2002, 2003).Consequently, significant levels of dissolved organiccarbon are present throughout the year, as are bacte-ria and both heterotrophic and phototrophic flagel-lated protozoa. Mixotrophy is a commonphenomenon among some of the major phototroph-ic components in Antarctic lakes, involving theingestion of bacteria and dissolved organic carbon,thereby allowing active populations to function overthe winter period (Marshall and Laybourn-Parry2002, Laybourn-Parry et al. 2005). Moreover, there isnow evidence of high virus activity within Antarcticlakes, suggesting that viral activity may play a signifi-cant role in carbon recycling (Madan et al. 2005).
Our PCA analyses between dinoflagellate dynam-ics and physicochemical parameters revealed that allparameters measured could in part explain the vari-ation in dinoflagellate abundance. However, onlytemperature, nitrate, and pH were important in allthree lakes. Nevertheless, none of these factors wassignificantly correlated to dinoflagellate abundanceon their own. These results are not surprising, sincelight, temperature, and nutrients are all known toaffect phytoplankton growth, while phytoplanktoncan influence these parameters as well as DOC andpH. The only clear trend was that salinity (measuredas conductivity) and phosphate appeared to have asubstantial effect on both abundance and speciescomposition in our between-lake comparison.
The saline lakes of the Vestfold Hills offer aremarkable natural laboratory in which to study howplanktonic protists have adapted to a range of evolv-ing physicochemical conditions. This study illus-trates how the ancestral marine community hasundergone radical simplification, leaving a smallnumber of well-adapted species. Over the relativelyshort time since isolation from the ocean after thelast major glaciations, distinct lacustrine populationshave most probably evolved. Further work will showwhether these populations are phenotypicallyand ⁄ or genetically distinct from each other, andfrom populations of the same species in the sea.
Australian Antarctic Science Advisory Committee grant toJ. L. P., Crafoord Foundation & Swedish Research Council toK. R., Carlsberg Foundation grant to G. H. G. Cronbergkindly helped with preliminary SEM and LM photographs.We thank T. Richardson for advice on light extinction calcu-lations, and K. Olsson for help with PCA. Finally, we appreci-ate the comments of the editor and two anonymousreviewers, which greatly improved the manuscript.
Adachi, M., Sako, Y. & Ishida, Y. 1994. Restriction-fragment-length-polymorphism of ribosomal DNA internal transcribed spacerand 5.8s-regions in Japanese Alexandrium species (Dinophy-ceae). J. Phycol. 30:857–63.
DINOFLAGELLATES IN ANTARCTIC SALINE LAKES 11

Adamson, D. A. & Pickard, J. 1986. Cainozoic history of the VestfoldHills. In Pickard, J. [Ed.] Antarctic Oasis. Academic Press,Sydney, pp. 63–97.
Anderson, D. M. & Wall, D. 1978. Potential importance of benthiccysts of Gonyaulax tamarensis and G. excavata in initating toxicdinoflagellate blooms. J. Phycol. 14:224–34.
Balech, E. 1976. Clave Ilustrada de Dinoflagllados Antarticos. InstitutoAntartico Argentino, Buenos Aires, 99 pp.
Borges, P. A. V. & Brown, V. K. 1999. Effect of island geological ageon the arthropod species richness of Azorean pastures. Biol. J.Linn. Soc. 66:373–410.
Caron, D. A., Porter, K. G. & Sanders, R. W. 1990. Carbon, nitrogenand phosphorus budgets for the mixotrophic phytoflagellatePoteriochromonas malhamensis (Chrysophyceae) during bacterialgrowth. Limnol. Oceanogr. 35:433–43.
Cromer, L., Gibson, J. A. E., Swadling, K. M. & Ritz, D. A. 2005.Faunal microfossils: indicators of Holocene ecological changein a saline Antarctic lake. Palaeogeogr. Palaeoclimatol. Palaeoecol.222:83–97.
Dale, B. 1983. Dinoflagellate resting cysts: ‘‘benthic plankton.’’In Fryxell, G. A. [Ed.] Survival Strategies of the Algae. CambridgeUniversity Press, Cambridge, UK, pp. 69–137.
Edvardsen, B., Shalchian-Tabrizi, K., Jakobsen, K. S., Medlin, L. K.,Dahl, E., Brubak, S. & Paasche, E. 2003. Genetic variability andmolecular phylogeny of Dinophysis species (Dinophyceae)from Norwegian waters inferred from single cell analyses ofrDNA. J. Phycol. 39:395–408.
Eisenriech, S. J., Bannermann, R. T. & Armstrong, D. E. 1975.A simplified phosphorus analysis technique. Environ. Lett.9:45–53.
Ferris, J. M., Burton, H. J., Johnstone, G. W. & Bayly, I. A. E. 1988.Biology of the Vestfold Hills, Antarctica. Kluwer Academic Pub-lishers, Dordrecht, the Netherlands, 307 pp.
Fields, S. D. & Rhodes, R. G. 1991. Ingestion and retention ofChroomonas spp. (Cryptophyceae) by Gymnodinium acidotum(Dinophyceae). J. Phycol. 27:525–9.
Gast, R. J., Moran, D. M., Beaudoin, D. J., Blythe, J. N., Dennett, M.R. & Caron, D. A. 2006. Abundance of a novel dinoflagellatephylotype in the Ross Sea, Antarctica. J. Phycol. 42:233–42.
Gast, R. J., Moran, D. M., Dennett, M. R. & Caron, D. A. 2007.Kleptoplasty in an Antarctic dinoflagellate: caught in evolu-tionary transition? Environ. Microbiol. 9:39–45.
Grey, J., Laybourn-Parry, J., Leakey, R. J. G. & McMinn, A. 1997.Temporal patterns of protozooplankton abundance and theirfood in Ellis Fjord, Princess Elizabeth Land, Eastern Antarc-tica. Estuar. Coast. Shelf. Sci. 45:17–25.
Hall, T. A. 1999. BioEdit: a user-friendly biological sequencealignment editor and analysis program for Windows95 ⁄ 98 ⁄ NT. Nucleic Acids Symp. Ser. 41:95–8.
Hansen, B., Bjørnsen, P. K. & Hansen, P. J. 1994. The size ratiobetween planktonic predators and their prey. Limnol. Oceanogr.39:395–403.
Heaney, S. I. & Furnass, T. I. 1980. Laboratory models of diel ver-tical migration in the dinoflagellate Ceratium hirundinella.Freshw. Biol. 10:163–70.
Hodson, D. A., Doran, P. T., Roberts, D. & McMinn, A. 2004. Pal-aeolimnological studies from the Antarctic and sub-Antarcticislands. In Pienitz, R., Douglas, M. S. V. & Smol, J. P. [Eds.]Long-Term Environmental Change in Arctic and Antarctic Lakes.Springer, The Hague, the Netherlands, pp. 419–74.
Jacobson, D. M. & Anderson, D. M. 1996. Widespread phagocytosisof ciliates and other protists by marine mixotrophic and het-erotrophic thecate dinoflagellates. J. Phycol. 32:279–85.
Kirk, J. T. O. 1994. Light and Photosynthesis in Aquatic Ecosystems, 2nded. Cambridge University Press, Cambridge, UK, 509 pp.
Kremp, A., Elbrachter, M., Schweikert, M., Wolny, J. L. & Gottsch-ling, M. 2005. Woloszynskia halophila (Biecheler) comb. nov.: abloom-forming cold-water dinoflagellate co-occuring withScrippsiella hangoei (Dinophyceae) in the Baltic Sea. J. Phycol.41:629–42.
Kremp, A. & Parrow, M. W. 2006. Evidence for asexual resting cystsin the life cycle of the marine peridinoid dinoflagellate,Scrippsiella hangoei. J. Phycol. 42:400–9.
Larsen, J., Kuosa, H., Ikavalko, J., Kivi, K. & Hallfors, S. 1995. Aredescription of Scrippsiella hangoei (Schiller) comb. nov. – a‘red tide’ dinoflagellate from the northern Baltic. Phycologia34:135–44.
Laybourn-Parry, J. 1997. The microbial loop in Antarctic lakes. InHoward-Williams, C., Lyons, W. & Hawes, I. [Eds.] EcosystemProcesses in Antarctic Ice-Free Landscapes. Balkema, Rotterdam,the Netherlands, pp. 231–40.
Laybourn-Parry, J. 2002. Survival mechanisms in Antarctic lakes.Philos. Trans. R. Soc. Lond. B Biol. Sci. 357:863–9.
Laybourn-Parry, J. 2003. Polar Limnology – The Past, the Present and theFuture. Scar VIII International Biology Symposium Proceedings –Antarctic Biology in a Global Context. Backhuys Publishers,Leiden, the Netherlands, pp. 321–9.
Laybourn-Parry, J., Bell, E. M. & Roberts, E. C. 2000. Protozoangrowth rates in Antarctic lakes. Polar Biol. 23:445–51.
Laybourn-Parry, J. & Marchant, H. J. 1992. Daphniopsis studeri(Crustacea: Cladocera) in lakes of the Vestfold Hills, Antarc-tica. Polar Biol. 11:631–5.
Laybourn-Parry, J., Marshall, W. A. & Marchant, H. J. 2005. Nutri-tional versatility as a key to survival in Antarctic phytoflagel-lates in two contrasting saline lakes. Freshw. Biol. 50:830–8.
Laybourn-Parry, J. & Perriss, S. J. 1995. The role and distribution ofthe autotrophic ciliate Mesodinium rubrum (Myrionecta rubra) inthree Antarctic saline lakes. Arch. Hydrobiol. 135:179–94.
Laybourn-Parry, J., Quayle, W. & Henshaw, T. 2002. The biologyand evolution of Antarctic saline lakes in relation to salinityand trophy. Polar Biol. 25:542–52.
Legrand, C. & Carlsson, P. 1998. Uptake of high molecular weightdextran by the dinoflagellate Alexandrium catenella. Aquat.Microb. Ecol. 16:81–6.
Logares, R., Daugbjerg, N., Boltovskoy, A., Kremp, A., Laybourn-Parry, J. & Rengefors, K. 2008. Recent evolutionary diversifi-cation of a protist lineage. Environ. Mirobiol. doi: 10.1111 ⁄j.1462-2920.2007.01538.x.
Logares, R., Rengefors, K., Kremp, A., Shalchian-Tabrizi, K.,Boltovskoy, A., Tengs, T., Shurtleff, A. & Klaveness, D. 2007a.Phenotypically different microalgal morphospecies with iden-tical ribosomal DNA: a case of rapid adaptive evolution. Microb.Ecol. 53:549–61.
Logares, R., Shalchian-Tabrizi, K., Boltovskoy, A. & Rengefors, K.2007b. Extensive dinoflagellate phylogenies indicate infre-quent marine-freshwater transitions. Mol. Phylogenet. Evol.45:887–903.
Lopez-Garcıa, P., Rodrıguez-Valera, F., Pedros-Alio, C. & Moreira,D. 2001. Unexpected diversity of small eukaryotes in deep-seaAntarctic plankton. Nature 409:603–7.
Macarthur, R. H. & Wilson, E. O. 1967. The Theory of Island Bioge-ography. Princeton University Press, Princeton, New Jersey, 224pp.
Madan, N. J., Marshall, W. A. & Laybourn-Parry, J. 2005. Virus andmicrobial loop dynamics over an annual cycle in three con-trasting Antarctic lakes. Freshw. Biol. 50:1291–300.
Marchant, H. J. & Scott, F. J. 2005. Introduction. In Scott, F. J. &Marchant, H. J. [Eds.] Antarctic Marine Protists. AustralianBiological Resources Study, Canberra, Australia, pp. 1–12.
Marshall, W. A. 1996. Biological particles over Antarctica. Nature383:680.
Marshall, W. A. & Laybourn-Parry, J. 2002. The balance betweenphotosynthesis and grazing in Antarctic mixotrophic crypto-phytes. Freshw. Biol. 47:2060–70.
McKenzie, C. H. & Cox, E. H. 1991. Spatial and seasonal changes inthe species composition of armored dinoflagellates in theSouthwestern Antarctic Ocean. Polar Biol. 11:139–44.
McMinn, A. & Scott, F. J. 2005. Dinoflagellates. In Scott, F. J. &Marchant, H. J. [Eds.] Antarctic Marine Protists. Goanna Print,Canberra, Australia, pp. 202–50.
Montresor, M., Lovejoy, C., Orsini, L., Procaccini, G. & Roy, S.2003. Bipolar distribution of the cyst-forming dinoflagellatePolarella glacialis. Polar Biol. 26:186–94.
Montresor, M., Procaccini, G. & Stoecker, D. K. 1999. Polarella gla-cialis, gen. nov., sp. nov. (Dinophyceae): Suessiaceae are stillalive! J. Phycol. 35:186–97.
12 KARIN RENGEFORS ET AL.

Parducz, B. 1967. Ciliary movement and coordination in ciliates.Int. Rev. Cytol. 21:91–128.
Parker, B. C., Simmons, G. M., Seaburg, K. G., Cathey, D. D. & Allnut,F. C. T. 1982. Comparative ecology of plankton communities inseven Antarctic oasis lakes. J. Plankton Res. 4:271–86.
Parsons, R., Maita, Y. & Lalli, C. M. 1984. A Manual of Chemical andBiological Methods for Seawater Analysis. Pergamon Press, Oxford,UK, 173 pp.
Perriss, S. J. & Laybourn-Parry, J. 1997. Microbial communities insaline lakes of the Vestfold Hills (eastern Antarctica). PolarBiol. 18:135–44.
Pickard, J. 1986. Antarctic Oasis, Davis Station and the VestfoldHills. In Pickard, J. [Ed.] Antarctic Oasis. Academic Press,Sydney, pp. 1–19.
Rankin, L. M., Gibson, J. A. E., Franzmann, P. D. & Burton, H. R.1999. The chemical stratification and microbial communitiesof Ace Lake, Antarctica: a review of the characteristics of amarine-derived meromictic lake. Polarforschung 66:33–52.
Reifel, K. M., McCoy, M. P., Rocke, T. E., Tiffany, M. A., Hurlbert, S.H. & Faulkner, D. J. 2002. Possible importance of algal toxinsin the Salton Sea, California. Hydrobiologia 473:275–92.
Rengefors, K. 1997. The Role of Resting Cysts in the Survival and Suc-cession of Freshwater Dinoflagellates. Comprehensive Summaries ofUppsala Dissertations From the Faculty of Science and Technology332. Uppsala University, Uppsala, Sweden, 42 pp.
Roberts, D. & McMinn, A. 1999. A diatom-based paleosalinity his-tory of Ace Lake, Vestfold Hills, Antarctica. Holocene 9:401–8.
Smith, V. H., Foster, B. L., Grover, J. P., Holt, R. D., Leibold, M. A.& Denoyelles, F., Jr. 2005. Phytoplankton species richnessscales consistently from laboratory microcosms to the world’soceans. Proc. Natl. Acad. Sci. U.S.A. 102:4393–6.
Stoecker, D. K. 1999. Mixotrophy among dinoflagellates. J. Eukaryot.Microbiol. 46:397–401.
Stoecker, D. K., Buck, K. R. & Putt, M. 1992. Changes in the sea-icebrine community during the spring-summer transition,McMurdo Sound, Antarctica. I. Photosynthetic species. Mar.Ecol. Prog. Ser. 84:267–78.
Swadling, K. M., Darnall, H. J. G., Gibson, J. A. E., Saulnier-Talbot,E. & Vincent, W. F. 2001. Fossil rotifers and the early coloni-zation of an Antarctic Lake. Quat. Res. 55:380–4.
Talling, J. F. 1974. General outline of spectrophotometric methods.In Vollenweider, R. A. [Ed.] A Manual on Methods for MeasuringPrimary Production in Aquatic Environments. IBP Handbook No.12. Blackwell, Oxford, UK, pp. 22–4.
Tengs, T., Dahlberg, O. J., Schalchian-Tabrizi, K., Klaveness, D.,Rudi, K., Delwiche, C. F. & Jakobsen, K. S. 2000. Phylogeneticanalyses indicate that the 19’Hexanoyloxy-fucoxantin-cona-tining dinoflagellates have tertiary plastids of haptophyte ori-gin. Mol. Biol. Evol. 17:718–29.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F. &Higgins, D. G. 1997. The CLUSTAL_X windows interface:flexible strategies for multiple sequence alignment aided byquality analysis tools. Nucleic Acid Res. 25:4876–82.
Thomson, P. G., Wright, S. W., Bolch, C. J. S., Nichols, P. D.,Skerratt, J. H. & McMinn, A. 2004. Antarctic distribution,pigment and lipid composition, and molecular identificationof the brine dinoflagellate Polarella glacialis (Dinophyceae).J. Phycol. 40:867–73.
Van Dolah, F. M. 2000. Diversity of marine and freshwater algaltoxins. In Botana, L. M. [Ed.] Seafood and Freshwater Toxins:Pharmacology, Physiology, and Detection. Marcel Dekker, NewYork, pp. 19–43.
Vincent, W. 2000. Evolutionary origins of Antarctic microbiota:invasion, selection and endemism. Antarct. Sci. 12:374–85.
Wetzel, R. G. 2001. Limnology: Lake and River Ecosystems, 3rd ed. Aca-demic Press, Elsevier Science, San Diego, California, 1006 pp.
Worm, B., Lotze, H., Hillebrand, H. & Sommer, U. 2002. Consumerversus resource control of species diversity and ecosystemfunctioning. Nature 417:848–51.
Zwartz, D., Bird, M., Stone, J. & Lambeck, K. 1998. Holocene sea-level change and ice-sheet history in the Vestfold Hills, EastAntarctica. Earth Plan. Sci. Lett. 155:131–55.
DINOFLAGELLATES IN ANTARCTIC SALINE LAKES 13