macrophage cell adhesion and inflammation cytokines on magnetostrictive nanowires
TRANSCRIPT

This article was downloaded by:[CDL Journals Account]On: 8 January 2008Access Details: [subscription number 785022370]Publisher: Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
NanotoxicologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t716100760
Macrophage cell adhesion and inflammation cytokineson magnetostrictive nanowiresKristy M. Ainslie a; Eric M. Bachelder bc; Gaurav Sharma d; Craig A. Grimes de;Michael V. Pishko adfa Department of Chemical Engineering, The Pennsylvania State University,Bethesda, Marylandb National Institutes of Health, National Institute of Allergy and Infectious Diseases,Bethesda, Marylandc Department of Chemical Engineering, University of Nebraska, Lincoln, Nebraskad Materials Science and Engineering, The Pennsylvania State University,Pennsylvania, USAe Electrical EngineeringThe Pennsylvania State University, Pennsylvania, USAf Chemistry, The Pennsylvania State University, Pennsylvania, USA
Online Publication Date: 01 December 2007To cite this Article: Ainslie, Kristy M., Bachelder, Eric M., Sharma, Gaurav, Grimes, Craig A. and Pishko, Michael V.(2007) 'Macrophage cell adhesion and inflammation cytokines on magnetostrictive nanowires', Nanotoxicology, 1:4, 279 -290To link to this article: DOI: 10.1080/17435390701781142URL: http://dx.doi.org/10.1080/17435390701781142
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08
Macrophage cell adhesion and inflammation cytokines onmagnetostrictive nanowires
KRISTY M. AINSLIE1, ERIC M. BACHELDER2,3, GAURAV SHARMA4,
CRAIG A. GRIMES4,5, & MICHAEL V. PISHKO1,4,6
1Department of Chemical Engineering, The Pennsylvania State University, 2National Institutes of Health, National Institute
of Allergy and Infectious Diseases, Bethesda, Maryland, 3University of Nebraska, Department of Chemical Engineering,
Lincoln, Nebraska, Departments of 4Materials Science and Engineering, 5Electrical Engineering, 6Chemistry, The
Pennsylvania State University, Pennsylvania, USA
AbstractA common problem with medical implants is the biofouling response which can detrimentally damage implants orprevent the implant from function properly. This response is characterized by a thick, frequently avascular, layer ofproteins and cells over the implant. To study this problem, we have examined here the adhesion of macrophages andthe subsequent expression of inflammatory cytokines on nanowire arrays. We found that the cells on the nanowirestypically occupied less area and were more circular than on a flat surface of the same material as the nanowires ortissue culture polystyrene (TCPS) in both the presence and absence of fetal bovine serum. Furthermore, this differencewas amplified by pre-coating the surfaces with collagen. The smaller area and circular shape indicated that the cellswere not thriving on the surface. Since there was potentially a high amount of cell death on the material, andbiofouling is frequently characterized as a chronic inflammation, an eighteen cytokine Luminex† panel was performedon the supernatant from macrophages on nanowires, control wafers, and TCPS. As a positive control for inflammation,lipopolysaccharide (LPS) was added to macrophages on TCPS to estimate the maximum inflammation response of themacrophages. Our results indicated that the nanowire structure results in the up-regulation of production inmacrophages of inflammatory cytokines such as IL-1a, and IFN-g and the down-regulation of IL-6, compared tocontrol wafers. In addition, the nanostructure also increased the production of IL-10 which is known as an inhibitor ofinflammation. Our results showed that the nanoarchitecture can disrupt cell adhesion and may lead to an inflammatoryresponse.
Keywords: nanomaterials, biomaterials, biocompatibility
Introduction
As an implant, or foreign body, is introduced to the
body the first event at the biofluid-material barrier is
adsorption of proteins onto the surface of the
material. The adsorbed proteins serve as an anchor
to which cells and additional proteins adhere (Bala-
subramanian et al. 1999). The results of this protein
adsorption and subsequent cell adhesion can be
detrimental to the implant, such is the case of
biofouling, an adverse host response that results
in implant encapsulation with proteins and cells
(Padera & Colton 1996). The biofouling response is
characterized by several layers of macrophages and
large multinucleated cells known as Foreign Body
Giant Cells (FBGC). These FBGCs are generally
several fused macrophages and are surrounded by
collagen, and other extracellular matrix (ECM)
proteins (Anderson 1994). ECM proteins also serve
especially as anchors for these giant cells to form
(Ruoslahti & Pierschbacher 1987; Rosales & Juliano
1995). It is these giant cells that not only serve as a
mass transfer barrier if the implant is a biosensor,
but they also have been shown to release lysosomal
enzymes and reactive oxygen intermediates which
can degrade the surface of the implant (Abramson &
Gallin 1990; Zhao et al. 1990; Adams & Hamilton
1992).
The first step in cell adhesion to implanted
devices is the adsorption of proteins. Biomolecule
adsorption has been shown to be altered by the
nanoarchitecture of the surface in vitro (Lampin
et al. 1997; Luck et al. 1998; McFarland et al.
2000; Denis et al. 2002; Suh et al. 2004; Pallandre
et al. 2005) and in vivo (Brauker et al. 1995;
Rosengren et al. 1999; Rosengren & Bjursten
Correspondence: Michael V. Pishko, 3122 TAMU, Texas A & M University, College Station, TX 77845-3122, USA. Tel:
�1(979) 845 3348. Fax: �1(979) 862 3362. E-mail: [email protected]
Nanotoxicology, December 2007; 1(4): 279�290
ISSN 1743-5390 print/ISSN 1743-5404 online # 2007 Informa UK Ltd.
DOI: 10.1080/17435390701781142

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08 2003). Suh et al. showed a decrease in protein
adsorption on silica particles with decreasing pore
size from 45�2.2 nm (Suh et al. 2004). Similarly,
Luck et al. demonstrated that the adsorption of
serum proteins was reduced on polymeric nano-
particles ranging in size from 141�61 nm, respec-
tively (Luck et al. 1998). Lampin et al. concluded
that increasing surface roughness of poly(methyl
methacrylate) (PMMA) to 0.20�3.4 mm, thus
increasing hydrophobicity, resulted in increased
protein adsorption (Lampin et al. 1997). Further-
more, we have previously shown that protein
adsorption is significantly reduced, around an order
of magnitude, on the nanoarchitectured magnetos-
trictive wires (Ainslie et al. 2005).
By disrupting the adsorption of protein on the
surface the nanoarchitecture should also disrupt
the adsorption of cells, since protein adsorption is
the first step in cell attachment. Indeed several
researchers have shown that by altering the amount
of protein adsorbed to the surface, the amount or
success of the cells on the surface is altered also (Naji
& Harmand 1990; Martin et al. 1995; Altankov et al.
1996; Kanagaraja et al. 1996; Lampin et al. 1997;
Steele et al. 1997; Balcells & Edelman 2002; Lan
et al. 2005). Furthermore, Lampin et al. show that
cell adhesion increased with roughness, most likely
due to the change in fibronectin adsorption on the
surface. This was similarly seen by Martin et al. who
observed a reduced number of osteoblasts on
roughened titanium compared to a more smoothed
surface (Martin et al. 1995). Osteoblasts and fibro-
blast adhesion was also shown to be influenced by
the surface state of cobalt chromium based alloys by
Naji and Harmand (1990). Clearly protein adsorp-
tion and cell adhesion go hand-in-hand and can be
mutually altered as a result of surface architecture or
modifications.
The influence of architecture on protein adsorp-
tion led us to hypothesize that nanofabricated
surfaces would significantly inhibit the formation of
an avascular fibrous capsule. To explore this we have
created a nanowire array comprised of Fe-Co-Ni
ternary alloy, imaged with SEM in Figure 1. The
characterization of this array is outlined in the
author’s previous work (Sharma et al. 2004).
The diameter of the wires was 75 nm, which
approaches the dimensions of several proteins (e.g.,
immunoglobin G � 45�23.5 nm, fibronectin � 45�0.6 nm and albumin � 0.4�14 nm) (Zhang et al.
1998). The author’s previous work has highlighted
the attenuation of protein adsorption on the surface
of the nanowires (Ainslie et al. 2005). The wire
length is on the order of 10 microns, as are most
mammalian cells. To explore the hypothesis that cell
adhesion will be altered as a result of the nanowire
architecture we have studied the cell shape of
macrophages and cytokines released by those macro-
phages on the nanowire, control wafer (flat surfaces
of the same materials as nanowires) and tissue
culture polystyrene (TCPS).
Figure 1. Nanowire array as visualized with FE- SEM. The deposition conditions are 15 V AC 1000 Hz for 15 min. The alumina
membrane has been partially etched in 0.2 M NaOH solution for 35 min to expose standing nanowire array. The free standing nanowires
are 4 microns in length and have 75 nm diameter.
280 K. M. Ainslie et al.

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08 Materials and methods
Materials were purchased from Sigma (St Louis,
MO, USA; Dulbecco’s Modified Eagles Medium
(DMEM), Antibiotic-antimycotic, trypsin-EDTA
solution 0.25%, Type 1 collagen from rat tail,
lipopolysaccharide (LPS) from Escherichia coli
0127:B8, and Phosphate Buffered Solution (PBS;
0.15 M NaCl, 0.001 M KH2PO4, and 0.002 M
Na2HPO4 pH 7.4)), ATCC (Manasas, VA; RAW
264.7 mouse macrophages), Hyclone (Logan, UT;
Standard Fetal Bovine Serum [FBS]), and Molecu-
lar Probes (Eugene, OR; Cell Tracker Green
CMFDA).
Scanning electron microscopy
High resolution images were collected with a field
emission SEM (JEOL 6700F) after coating the
samples with gold.
Surface fabrication
The nanowires were developed in a method identical
to that outlined by Sharma et al. (2004). The control
(Fe-Co-Ni) wafers are flat surfaces comprised of the
same material as the nanowires, but lacking the
architecture of the wires. The electrolyte solution for
Fe-Co-Ni film deposition was the same as used for
nanowire deposition (14.055 g/l of CoSO4.7H2O,
52.5718 g/l of NiSO4.6H2O, 5.56 g/l of FeS-
O4.7H2O, and 24.7328 g/l of H3BO3). The deposi-
tion of the alloy was carried out at a current density
of 10 mA/cm2 for 30 min.
Cell culture
For both cell types, cell culture media was comprised
of 10% FBS and 1% antibiotic-antimycotic in
DMEM. The cells were maintained in T-75 poly-
styrene culture flasks at 5% COB2B and 378C and
subcultured by cell scraping. For each subculturing,
cells were assigned a passage number. For all
experiments presented here, passages 4�9 were used.
For cell adhesion and cytokine expression experi-
ments, cells were suspended in the media and
separated from that media through centrifugation.
The cells were either resuspended in media with
FBS (DMEM with 1% antibiotic-antimycotic and
10% FBS) or in FBS-free media (DMEM with 1%
antibiotic-antimycotic). The cells were diluted to a
concentration of 50,000 cells per a milliliter. They
were then seeded at 0.5 ml per a 24-well plate well.
For the desired cases, nanowires or control wafers
were placed at the bottom of the well with the
surface occupying the rectangular area of the circular
well. Experimental plates were maintained until
point of imaging wherein the surface for that
particular time point was removed from the incu-
bator and imaged.
For collagen pre-treated surfaces type 1 collagen
from rat tail was suspended in accordance with
manufacturer’s direction. The collagen was diluted
to a 0.1% solution in PBS and incubated with the
surfaces overnight in the refrigerator in a 24-well
plate. Surfaces were rinsed with fresh PBS and
warmed to 378C before cells were introduced. FBS
free media was used in conjunction with the collagen
experiments.
Cell imaging
Cells were imaged at discrete time points with
fluorescent dye Cell Tracker Green CMFDA†.
Cell tracker solution was prepared and incubated
with cells in accordance with manufacturer’s direc-
tions. Samples were washed in PBS prior to imaging.
The fluorphore laden cells were imaged with a
fluorescein isothiocyanate (FITC) filter on a Zeiss
Axiovert 200M. Cell number, area, and length were
calculated by manually tracing or measuring the cell
via AxioVision LE version 4.3 (Zeiss). Circularity is
calculated by Equation 1.
Circularity�Area
pL2
4
Equation 1
where area is the area of the cell and L is the length
of the cell.
Cytokine profile with Luminex
CytokineProfilerTM Testing Service from Upstate
(Lake Placid, NY, USA) was used to measure 18
cytokines using Luminex† technology. Briefly, by
using a Luminex† 100 instrument from the Lumi-
nex† Corporation (Austin, Texas) multiple cytokines
in a single sample can be detected at simultaneously.
This is achieved by using a combination of fluores-
cent microbeads and reporter molecules. The Lumi-
nex† 100 uses two lasers to detect the fluorescent
microbead and the reporter molecules. By using
standard curves for every cytokine, the cytokines in
the samples can be determined and quantified.
Activation of macrophages
In the Luminex† cytokine profile studies it was
necessary to show the cytokine profile of activated
macrophages as a positive control. As such we could
then determine if the nanowire or control wafer
surfaces were immunogenic. To fully activate macro-
phages, lipopolysaccharide (LPS) was used. LPS is a
Macrophage cell adhesion and inflammation cytokines on magnetostrictive nanowires 281
Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08 major constituent of the cell wall of gram-negative
bacteria, is highly immunogenic, and is one of the
best activators of macrophages (Paul 1994). LPS
was added at 1 mg/ml to macrophages seeded on
TCPS to obtain the maximum production of in-
flammatory cytokines. By comparing the amount
and type of cytokines released by the LPS activated
macrophages to the nanowire or control wafer
cultured macrophages, it can be determined if
surfaces are immunostimulatory or not.
Statistical analysis
All graphical figures present the average of three or
more data points. The confidence interval of the
data set was determined with Microsoft Excel with
an alpha value of 0.05 (CI of 95%). The differences
between two groups of data were determined by a
Student’s t-test (Microsoft Excel; Redmond, WA,
USA). Differences among groups were determined
with an ANOVA (Minitab; State College, PA, USA).
A p-value less than 0.05 is considered significantly
different.
Results
Cell shape
The average area occupied by the macrophages on
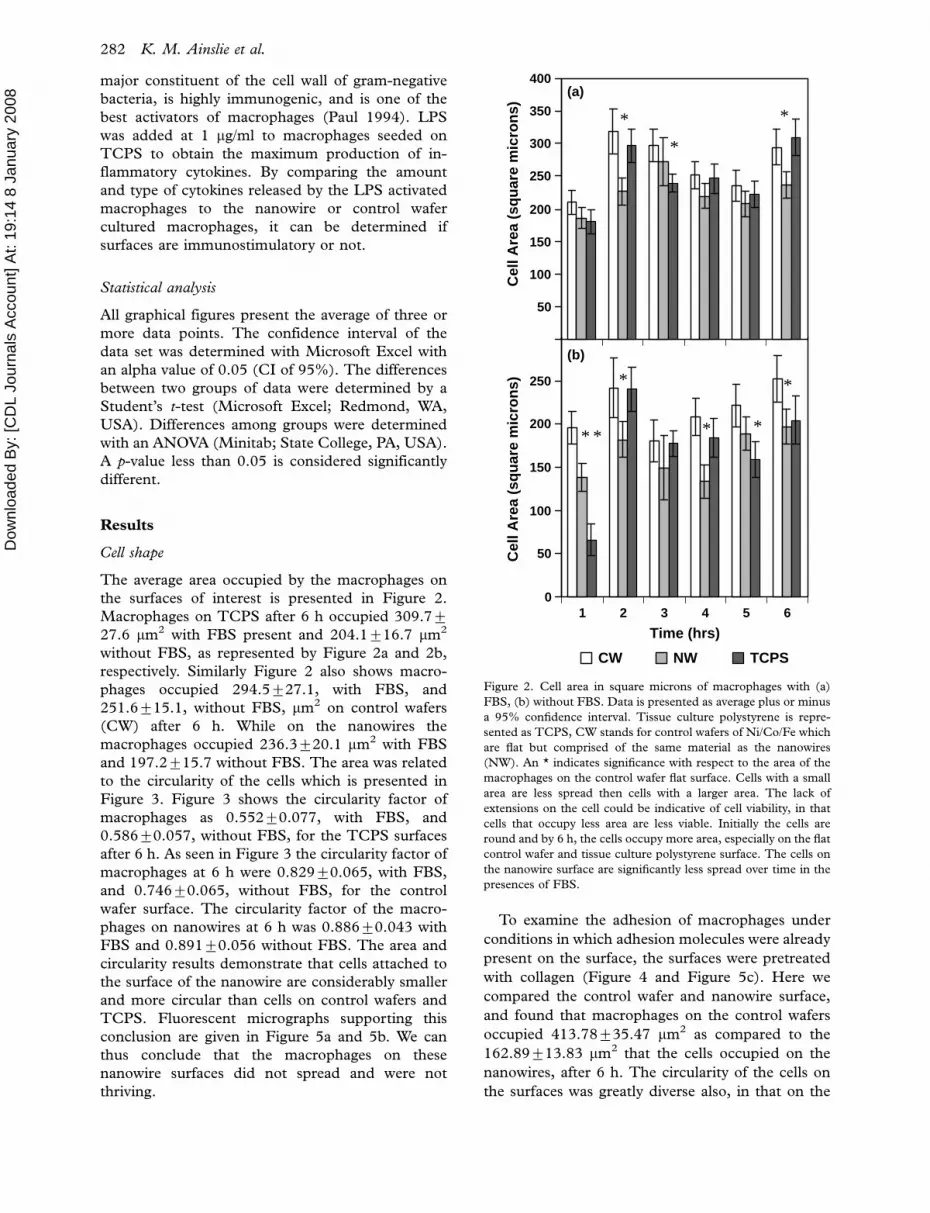
the surfaces of interest is presented in Figure 2.
Macrophages on TCPS after 6 h occupied 309.79
27.6 mm2 with FBS present and 204.1916.7 mm2
without FBS, as represented by Figure 2a and 2b,
respectively. Similarly Figure 2 also shows macro-
phages occupied 294.5927.1, with FBS, and
251.6915.1, without FBS, mm2 on control wafers
(CW) after 6 h. While on the nanowires the
macrophages occupied 236.3920.1 mm2 with FBS
and 197.2915.7 without FBS. The area was related
to the circularity of the cells which is presented in
Figure 3. Figure 3 shows the circularity factor of
macrophages as 0.55290.077, with FBS, and
0.58690.057, without FBS, for the TCPS surfaces
after 6 h. As seen in Figure 3 the circularity factor of
macrophages at 6 h were 0.82990.065, with FBS,
and 0.74690.065, without FBS, for the control
wafer surface. The circularity factor of the macro-
phages on nanowires at 6 h was 0.88690.043 with
FBS and 0.89190.056 without FBS. The area and
circularity results demonstrate that cells attached to
the surface of the nanowire are considerably smaller
and more circular than cells on control wafers and
TCPS. Fluorescent micrographs supporting this
conclusion are given in Figure 5a and 5b. We can
thus conclude that the macrophages on these
nanowire surfaces did not spread and were not
thriving.
To examine the adhesion of macrophages under
conditions in which adhesion molecules were already
present on the surface, the surfaces were pretreated
with collagen (Figure 4 and Figure 5c). Here we
compared the control wafer and nanowire surface,
and found that macrophages on the control wafers
occupied 413.78935.47 mm2 as compared to the
162.89913.83 mm2 that the cells occupied on the
nanowires, after 6 h. The circularity of the cells on
the surfaces was greatly diverse also, in that on the
50
100
150
200
250
300
350
400(a)
(b)
0
50
100
150
200
250
Time (hrs)
CW NW TCPS
*
*
*
* *
*
* *
*
Cel
l Are
a (s
qu
are
mic
ron
s)
1 2 3 4 5 6
Cel
l Are
a (s
qu
are
mic
ron
s)
Figure 2. Cell area in square microns of macrophages with (a)
FBS, (b) without FBS. Data is presented as average plus or minus
a 95% confidence interval. Tissue culture polystyrene is repre-
sented as TCPS, CW stands for control wafers of Ni/Co/Fe which
are flat but comprised of the same material as the nanowires
(NW). An * indicates significance with respect to the area of the
macrophages on the control wafer flat surface. Cells with a small
area are less spread then cells with a larger area. The lack of
extensions on the cell could be indicative of cell viability, in that
cells that occupy less area are less viable. Initially the cells are
round and by 6 h, the cells occupy more area, especially on the flat
control wafer and tissue culture polystyrene surface. The cells on
the nanowire surface are significantly less spread over time in the
presences of FBS.
282 K. M. Ainslie et al.

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08
control wafers the circularity is 0.52190.086 com-
pared to the nanowires where the circularity is
0.82890.063. The collagen pretreatment seemed
to enhance the adhesion of cells to the flat control
wafer surface, but not significantly alter the adhesion
of the macrophages to the nanowires.
Inflammation cytokines
The results of Luminix† inflammation panel are
presented in Table I. These expression levels are
reported in picograms per milliliter. Bold values
indicate where the expression level was significant
with respect to the positive LPS control at that time
Cir
cula
rity
1 2 3 4 5 6
Cir
cula
rity
0.2
0.4
0.6
0.8
1
1.2
(b)
(a)
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Time (hrs)
CW NW TCPS
**
* *
* *
*
*
* * * * * * * *
Figure 3. Circularity of macrophages with (a) FBS, (b) without
FBS. Circularity is calculated via area divided by p multiplied by
the square of one-half the cell length. A value close to 1 represents
a perfect circle, and a rectangle would be 0.57 or less. Data is
presented as average plus or minus a 95% confidence interval. An
* represents significance with respect to the circularity of the cells
on the control surface. Tissue culture polystyrene is represented as
TCPS, CW stands for control wafers of Ni/Co/Fe which are flat
but comprised of the same material as the nanowires (NW).
Overtime, the graph represents a spreading of cells on the TCPS
surface in that the circularity generally goes down, especially in the
presence of FBS. The circularity of the cells on the control wafer
surface spread somewhat over time, however the cells on the
nanowire surface have a constant circularity and do not appear to
spread.
100
200
300
400
500
600
(b)
(a)
Cel
l Are
a (s
qu
are
mic
ron
s)
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5 6
Time (hrs)
Cir
cula
rity
CW NW
*
*
**
* ** *
Figure 4. Cell area in square microns (a) and circularity (b) of
macrophages with collagen pretreatment. Circularity is calculated
via area divided by p multiplied by the square of one-half the cell
length. A value close to 1 represents a perfect circle, and a
rectangle would be 0.57 or less. Data is presented as average plus
or minus a 95% confidence interval. An * represents significance
with respect to the shape factor of the cells on the control surface.
Tissue culture polystyrene is represented as TCPS, CW stands for
control wafers of Ni/Co/Fe which are flat but comprised of the
same material as the nanowires (NW). Here an extracellular
matrix protein, collagen, was added to enhance cell adhesion.
Both figures conclude that the cells on the nanowire surface are
more circular and have less area than those on the control wafer.
The cells are more round and smaller, indicative of cell death or
apoptosis.
Macrophage cell adhesion and inflammation cytokines on magnetostrictive nanowires 283

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08
point. An * signifies significance with respect to the
expression of macrophages on the nanowires (NW).
A description of each cytokine is given in Table II.
Figure 6 shows a direct comparison between cyto-
kine expression levels between the nanowire and
control wafer surfaces.
Discussion
Cell number and shape
This study examined cell spreading in two ways:
First by monitoring its area; and second by noting
the shape of the cell and how circular it is. In general
Figure 5. Fluorescent micrographs of macrophages on the respective surfaces are presented (a) in the presences of FBS, (b) absence of FBS
and (c) with collage pretreatment. The micrographs were imaged with a 10� lens. Individual isolated cells were traced, at 40�, to obtain
area and circularity measurements. The lack of spreading and more circular nature of cells on the nanowire surface can be observed for all
the conditions.
284 K. M. Ainslie et al.

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08
the less spread a cell is, the less two dimensional area
it occupies. Linked to this is circularity which relates
how circular the cell is, the thought being that
circular cells are weakly adherent to the substrate.
If they find a viable surface on which to spread, they
will adhere and become less circular. A circular cell
(i.e., one that is spherical) will occupy the least
amount of area on a surface. In the absence of FBS
the cells appear to spread and adhere (Figure 2) less
over all and do not adhere as quickly on the TCPS
and NW surface. Although the area of the cell is
lowest at the one hour time point, more so for the
cultures without FBS, they are highest at the two
hour time point. Without FBS, the cells appear to
equilibrate to a smaller area at the three hour time
point and then spread, since all the average areas are
less at the three over six hour time point. In contrast,
the cells in the presence of FBS appear to shrink at
the five hour time point and then increase in area
again at the six hour time point. Similar trends for
the circularity are seen on the TCPS and CW
surface with respect to circularity in the FBS
Table I. Inflammation panel as measured with Lumix† for 18 cytokines. Table (a) is at the 24-hour time point and Table (b) is at the 48-
hour time point. Values are reported in picograms/milliliter and as average995% confidence interval. At least three samples were analyzed
for each surface at each time point and for DMEM. Where samples had one or more replicates detected below detection limits, the value is
reported as less than the maximum reported value or in the case of no reported value, the detection limit. Bold values indicate where the
value is significant with respect to the maximum or LPS value at that time point. The * denotes significance with respect to the nanowires
(NW). The DMEM values serve as background values for all cases and time points. Tissue culture polystyrene is represented as TCPS, CW
stands for control wafers of Ni/Co/Fe which are flat but comprised of the same material as the nanowires (NW). Except for chemokines
(MCP-1 and RANTES) and anti-inflammatory cytokines (IL-10 and IL-6), the cytokine expression of macrophages on the nanowire
surface is not significantly different from the LPS stimulated macrophages, indicating that the conditions are equally inflammatory. The
values of pro-inflammatory IFN-g, and IL-1a cytokine expression from macrophages on the control wafer surface are significantly lower at
the 48-hour time point compared to expression from macrophages on the nanowire surface, concluding that the flat surface is less
inflammatory than the nanowire surface.
(a) NW 24 hr CW 24 hr TCPS 24 hr LPS 24 hr DMEM
GM-CSF B228 B194 B216 B194 209957
IFNg 140925 B61 B145 94914 B70.9
IL-10 52950 B33 B83 324914 B34.6
IL-12p40 40915 4496 2799* 54931 B47.8
IL-12p70 65918 B58 B58 B48 B58
IL-13 87931 B122 4892* B76 4795*
IL-17 47939 B22 B49 17913 44925
IL-1a 1,4929368 98932* 27922* 308916 B15
IL-1b 115940 101970 112914 169977 133929
IL-2 490.2 B3 691* 492 490.0
IL-3 B8 B8 B8 B8 B8
IL-4 25913 32910 21917 31915 2590.1
IL-5 109926 1229114 80936 45913 121919
IL-6 55917 29995* 53919 1,8539323 1592*
KC B66 B66 B66 B66 B65
MCP-1 114932 138960 11294 813991 13195
RANTES 3599 B11 1393* 2,3189392 B51
TNFa B3 B3 B3 7719138 B3
(b) NW 48 hr CW 48 hr TCPS 48 hr LPS 48 hr
GM-CSF 86921 84938 71980 106917
IFNg 12399 62917* 50954* 60925
IL-10 61912 3595* 1999* 551947
IL-12p40 5194 B48 64931 B48
IL-12p70 53921 34923 B78 6391
IL-13 2697 19913 66930 33929
IL-17 B91 B96 38934 B14
IL-1a 931992 1409129* 37917* 757942
IL-1b 200910 230955 1899121 278935
IL-2 492 591 B6 792
IL-3 490.1 291 290 391
IL-4 32910 1295* 1696* 2895
IL-5 77915 7397 84931 67924
IL-6 127928 3259114* 189938 3,2659363
KC 1493 15910 38913 25911
MCP-1 73918 100935 1709141 2,7509161
RANTES 4398 34910 3293 3,9329347
TNFa B2 B3 B3 1,5939154
Macrophage cell adhesion and inflammation cytokines on magnetostrictive nanowires 285

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08
supplemented cultured. The circularity in the FBS
free cultures appears to deviate minimally with
respect to the individual surface at the different
time points. Overall FBS appears to alter the time
course of the spreading of the cell, with respect to
both area and circularity.
In both the presence and absence of FBS, the area
the cells occupied on the nanowire surface was
significantly less than on the control wafer surface,
as seen in Figure 2 after 6 h. Furthermore, the cells
were far more circular on the nanowire surface than
on the control wafer surface. This indicates that the
cells were not thriving on the nanostructured surface
and that this was not simply a property of the metal
oxide surface but rather the nanoarchitecture itself.
These results support the hypothesis that these
nanowire array might aid in the limiting of biofouling
on the surface of an implant.
The presence of FBS in the cell system is con-
founding. FBS has growth factors that stimulate cell
growth and proteins (cell adhesion molecules) that
can aid in cell adherence. As such, one must delineate
how growth factors or cell adhesion molecules
contributes to increase cell spreading and decrease
cell circularity. To potentially reduce the confound-
ing effects of FBS, the nanowire and control wafers
surfaces were pre-coated with 0.1% type 1 collagen
from rat tail. The aim of this experiment was to test
Table II. Cytokine definitions inflammation panel results in Table I. (Paul 1994; Al-Saffar et al. 1996).
Role(s)
GM-CSF Granulocyte/macrophage colony-
stimulating factor
A hematopoietic growth factor with a regulatory effect on the transformation of
immature macrophages into multinucleated giant cells (MNGC). Plays a role in
antibody stimulation.
IL-1 Interleukin-1 Major inflammatory mediator produced by activated macrophages. Increases local
blood flow, fever, release of other cytokines and enhanced expression of adhesion
molecules.
IL-10 Interleukin-10 Produced by macrophages and cytotoxic T-cells. Inhibits the production of
pro-inflammatory cytokines and interferences with macrophage-mediated antigen
presentation.
IL-12 Interleukin-12 Product of macrophages and B-cells that enhances the synthesis of IFN-g and
stimulates proliferation of NK, and T-cells.
IL-13 Interleukin-13 Produced by macrophages and cytotoxic T-cells. Inhibits the production of
pro-inflammatory cytokines and interferences with macrophage-mediated antigen
presentation. Aids in the differentiation of B-cells.
IL-17 Interleukin-17 Pro-inflammatory cytokine which induces stromal cell pro-inflammatory responses
and the production of hematopoietic cytokines.
IL-2 Interleukin-2 Produced by T-cells in response to antigenic or mitogenic stimulation. Also termed
a T-cell growth factor. Increases the growth and activity of macrophages, B-cells,
NK cells and other white blood cells.
IL-3 Interleukin-3 Produced in NK and T-cells to target mast and stem cells for growth and histamine
release.
IL-4 Interleukin-4 Participates in cell fusion related to granulomas and is anti-inflammatory. Also
related to allergic inflammation including stimulating of basophil development,
eosinophil chemotaxis, and expression of IgE receptors on B cells. Signals the
formation of giant or fused macrophages.
IL-5 Interleukin-5 Produced in T-helper cells and activates B-cells and the antibody response.
Proliferates and differentiates B-cells.
IL-6 Interleukin-6 Produced by T-cells, endothelial cells, monocytes and fibroblasts. Promotes
monocytes differentiations, increased number of platelets, and synthesis of
fibrinogen in T- and B-Cells and macrophages. Involved in the downregulation of
neutrophil superoxide production.
IL-8 Interleukin-8 Produced by monocytes, lymphocyte and Europhiles when stimulated with IL-1a,
IL-1b, or TNF. In neutrophils enhances chemotactic and degranulation response.
Induces an increase in the expression of cell surface adhesion molecules.
IFN-g Interferon-gamma Product of T- and natural killer (NK) cells. Increases generation of highly reactive
oxygen species (e.g., superoxide anion and hydrogen peroxide) and alters the cell
surface antigens of macrophages permitting them to engulf pathogens.
KC KC Potent neutrophil activator. Plays an important role in inflammation.
MCP-1 Monocyte chemotactic protein 1 Produced by most non-lymphocytes and plays roles in chronic inflammation,
activation of macrophages, humoral response and histamine release.
RANTES Regulated upon Activation, Normal T-cell
Expressed and Secreted
Produced in T-cells, endothelial cells and platelets. A T-cell product that promotes
mononuclear infiltrating (e.g., migration of macrophages into cancerous growth)
TNF Tumor Necrosis Factor Derived from activated macrophages. Is associated with the production of a fever
and promotes the stimulation of most other pro-inflammatory mediators. It is
primarily associated with the induction of cellular apoptosis.
286 K. M. Ainslie et al.

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08
that the presence of an adhesion promoting protein
aid in cell spreading without the presence of growth
factors. The macrophages on the control wafer sur-
face were much more spread than in the presence of
FBS, also they were much less circular. The differ-
ence between the area and circularity on the control
wafers and nanowires was more profound in the
presence of collagen. The macrophages on the
nanowires were slightly more circular, but occupied
roughly the same area as those on the control wafer
surface. There were two reasons for the potential
change of cell viability on the nanowires over the
control wafers. One, the disruption of cell adhesion
on the nanowires could be the result of the con-
formation of collagen on the surface i.e. collagen is
present in a conformation that the cells are unable to
attach. Secondly a lack of adsorbed collagen may
simply reduce cell adhesion and spreading. The
authors have previously shown that protein adsorp-
tion on the nanowires is attenuated by almost an
order of magnitude (Ainslie et al. 2005) It would
appear that despite the presence of ECM proteins,
the macrophages on the nanowires were not suffi-
ciently spread and much more circular than on the
control wafer surface. This trend was seen by the
authors with fibroblasts and endothelial cells also
(Ainslie et al. 2007). It would appear that the
nanowire surface disrupts the adherence of cells to
the surface, over a flat surface of equal material. To
look further into the ability of the surface to inhibit
the biofouling response, cytokines released from the
macrophages on these surfaces were studied.
Inflammation cytokine response
We examine cytokine release profiles on these
surfaces for two reasons. The first reason was that
an inflammatory response can occur when there is a
high amount of cell death on a surface (Matzinger
2002), which might result from rounded cells on a
surface. The second reason why we examined the
immune response is that biofouling response is
generally characterized as a chronic inflammatory
response. When an implant is placed in vivo, a high
percentage of cells bound to the surface are host
inflammatory cells Padera & Colton 1996). To
measure the cytokines produced with macrophages
cultured on nanowire surfaces we used Luminex†
Technology to measure a broad panel of expressed
cytokines. A description of the cytokines tested can
be seen in Table II. Cytokines released by macro-
phages can either stimulate (e.g., IL-1, IL-12,
1
10
100
1000
10000
IFNg IL-1a IL-4 IL-6 IL-10 RANTES
)L
m/smar
goci
p(n
oitcu
dor
Pe
nikoty
C
NW CW TCPS LPS
24 48
24 48
24 48
24 48
24 48
24 48
** ##
**
**##
** ##
* ###
* ###
# **###
*# # #
Figure 6. Cytokine data presented in Table I which highlights the cytokines that are statistically different when comparing the macrophages
on the surface of the nanowires and the other surfaces. IFN-g stands for IFN-g, IL-1a stands for IL-1a, tissue culture polystyrene is
represented as TCPS, and CW stands for control wafers of Ni/Co/Fe which are flat but comprised of the same material as the nanowires
(NW). The 24 and 48 indicators signal the data from 24- and 48-hour time points. Data is presented as average 995% confidence interval.
An * indicates significance with respect to the concentration of cytokine released from macrophages on the nanowires and a # represents
significance with respect to the LPS stimulated macrophage production. The production of pro-inflammatory cytokines IFN-g, and IL-1afrom the macrophages on the surface of the nanowires is not significantly different from LPS stimulated macrophages at any of the time
point, indicating that the conditions are equally inflammatory.
Macrophage cell adhesion and inflammation cytokines on magnetostrictive nanowires 287

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08 MCP-1 or TNF-a) or inhibit (e.g., IL-10, IL-4,
and IL-13) the inflammatory response.
IFN-g
As shown in Table I, the macrophages production of
IFN-g increased significantly on nanowire surfaces.
IFN-g is secreted by thymus derived lymphocytes
(T-cells), natural killer (NK) cells, and other im-
mune cells of the immune system. IFN-g secretion
by T-cells and NK cells is usually a strong indicator
of inflammation; however, IFN-g secretion by
macrophages has not been well studied and further
studies would need to be performed to show the
significance of the macrophages secreting IFN-gwhen cultured with the nanowires.
IL-1
IL-1 is an inflammatory cytokine of importance in
implant material pathologies. Ojo-Amaize et al. has
shown that women with breast implants have ele-
vated levels of IL-1, a common inflammatory
cytokine, circulating in their blood stream (Ojo-
Amaize et al. 1996). They concluded the IL-1 in the
bloodstream was due to a chronic inflammatory
response against the breast implant. In addition, it
has been shown that macrophages can cause bio-
corrosion on such metals as stainless steel and
implant alloys that contain cobolt (Lin & Bumgard-
ner 2004; Bailey et al. 2005). With this bio-corrosion
of metals there was a related increase in the
production of IL-1 b. We did not observe an increase
of production of IL-1 b on the control wafers or the
nanowires. Our results on the nanowires showed an
increase in IL-1a production compared to control
wafers, LPS, and TCPS at the 24- and 48-hour time
point. Even though IL-1 a and IL-1 b belong to the
same family of cytokines and have similar structure,
the regulation of production was completely differ-
ent. It is known that the release of IL-1a in vitro can
occur as a consequence of cell injury and cell death
(Paul 1994). It has been shown that 3T3 fibroblasts
cultured on the same nanowires used in this paper
resulted in significantly increased cell death, possibly
the result in the macrophages secreting high levels of
IL 1-a (Ainslie & Pishko 2007).
IL-4
At the 48-hour time point IL-4 was up-regulated on
the nanowire surface. The level of IL-4 expression in
the nanowires was higher than LPS, but not
significantly so. The up-regulation of IL-4 could be
of a concern in the formation of FBGC, which is of
concern in implanted devices. The formation of
these cells is sparked by the anchoring of ECM
proteins and the production of IL-4 and GM-CSF
(Jenney et al. 1998). Although IL-4 was up-regu-
lated, GM-CSF was not and actually less than the
negative control level in DMEM. This would
indicate that IL-4 production on the nanowire sur-
face was responsible for other cytokine actions and
not just the formation of FBGC. Like IL-10, the
presence of IL-4 could be that of an anti-inflamma-
tory response (Paul 1994).
IL-10
The amount of IL-10 produced by the macrophages
on the nanowires at the 48 hour time point was
significantly higher than the control wafers or TCPS.
This indicates that the nanoarchitecture seems to
lead to an up-regulation of anti-inflammatory IL-10.
IL-10 is a cytokine known to inhibit inflammatory
responses, especially in parts of the body that are
sensitive to inflammation such as the eye (Skelsey
et al. 2003) and the gut (Alpan et al. 2004). It has
been shown that nanofibrous PTFE (nPTFE) re-
sulted in the increased expression of IL-10 by
macrophages on the surface of this material (Ainslie
et al. 2007). This phenomenon that was found with
nPTFE may be the same phenomenon that was
found in the experiments described here.
IL-6
IL-6 seemed to be significantly up-regulated on the
control wafers over the nanowires or TCPS. Prior
research has shown IL-6 plays a key role in experi-
mental atherosclerosis. It is one of the key cytokines
that result in inflammation and plaque formation
(Schieffer et al. 2004). Also it has been shown that
IL-6 is secreted in vivo around copper implants
(Suska et al. 2005). Additionally, copper sulfate has
been shown to stimulate VEGF production in
keratinocytes (Sen et al. 2002). The nanowires
have also been shown to upregulate VEGF produc-
tion from 3T3 fibroblasts, equal to that of copper,
although no copper is in the alloy (Ainslie & Pishko
2007). Although no copper is present in the alloy
both IL-6 and VEGF were up-regulated. The IL-6
reduction in the nanowires may be a result of the
increase of IL-10. It has been shown that the
endogenous secretion of IL-10 by macrophages
results in the reduction of IL-6 (Wilson et al.
2005). This result can explain why the nanowires
have a much lower level of IL-6 compared to the
control wafers.
RANTES
RANTES is a chemokine and was elevated at the 24-
hour time point in nanowires compared to the
288 K. M. Ainslie et al.

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08 control wafers and then statistically equal to the
control wafers at the 48-hour time point. These
values at both time points were slightly higher than
the TCPS. Chemokines are a group of molecules
that are secreted by macrophages to recruit macro-
phages and other immune cells. Chemokines are
usually secreted at a site of inflammation to recruit
other cells to the site. The RANTES production is in
no way as high as the production by macrophages
stimulated by LPS, in fact it was roughly two orders
of magnitude less. Due to the small amount of
RANTES production and lack of significance at the
48-hour time point, the ability of the nanowires or
control wafers to recruit macrophages seems minor.
IL-13 and IL-2
Briefly, IL-13 and IL-2 were respectively up- and
down-regulated on the nanowire surface, compared
to TCPS, at the 12 hour time point. IL-13 typically
limits the production of inflammatory cytokines,
whereas IL-2 is termed a T-cell growth factor and
stimulates the activation of such cells. Since both of
these cytokines were no longer significant at the 48-
hour time points; their significance at the 24-hour
time point may be considered inconclusive.
MCP-1
The production of MCP-1 is significantly less on all
the surfaces compared to the LPS-stimulated control
at the 24- and 48-hour time points. Like RANTES,
MCP-1 is a chemokine that is linked with chronic
inflammation. The amount of MCP-1 expressed by
macrophages on the nanowires was on average the
lowest at the 48-hour time point, however it is not
significant. A reduced production of MCP-1 in vitro
may indicate that the surface does not stimulate a
chronic response associated with implants.
The potential for inflammation on nanowires
From the data presented with respect to IL-1a, IFN-
g, IL-10, and IL-6, in addition to the rest of the data
presented in Table II and Figure 6, one may
conclude the Ni-Co-Fe alloy that comprises the
nanowire and control wafer surface is immunogenic
and may yield an inflammatory response in vivo. The
nanoarchitecture of the surface appeared to affect
the production of cytokines, in particular the up-
regulation of IFN-g, IL-10, and IL-1a and down
regulation of IL-6. Further studies must be per-
formed to see if these nanowires would cause an
inflammatory response in vivo. The only other study
that includes the results of nanoarchitecture on
numerous inflammatory cytokine releases does not
compare a surface with and without nanoarchitec-
ture. It does, however, report that on nanofibrous
PTFE (nPTFE) there is not a significant up-regula-
tion of any of the 18 cytokines listed below after
48 h compared to TCPS (Ainslie et al. 2007, in
preparation).
Conclusion
This study compares a two Ni/Co/Fe surfaces, one
that is flat and one that is a nanowire array
comprised of nano- and micro-architecture. We
presented conclusive studies that show that na-
noarchitecture, in the form of a nanowire array,
disrupted cell adhesion by preventing cell adhesion
and spreading, and leads to the up-regulation of IL-
1a, IFN-g, IL-6 and IL-10 expression by adherent
macrophages. Furthermore, the architecture leads to
an inflammatory response above that of the flat
surface. The surface presented here has the ability to
prevent cell adhesion, as seen with biofouling, but
might create an inflammatory response in vivo.
References
Abramson SL, Gallin JI. 1990. IL-4 inhibits superoxide produc-
tion by human mononuclear phagocytes. J Immunol
144(2):625�630.
Adams DO, Hamilton TA. 1992. Macrophages as destructive cells
in host defense. In: Gallin I, Goldstein IM, Snyderman R,
editors. Inflamation: Basic principles and clinical correlates.
New York: Raven Press. p 637�662.
Ainslie KM, Bachelder EM, Borkar S, Zahr AS, Sen A, Badding
JV, Pishko MV. 2007. Cell adhesion on nanofibrous polytetra-
fluoroethylene (nPTFE). Langmuir 23(2):747�754.
Ainslie KM, Sharma G, Dyer MA, Grimes CA, Pishko MV. 2005.
Attenuation of protein adsorption on static and oscillating
magnetostrictive nanowires. Nano Lett 5(9):1852�1856.
Al-Saffar N, Khwaja HA, Kadoya Y, Revell PA. 1996. Assessment
of the role of GM-CSF in the cellular transformation and the
development of erosive lesions around orthopaedic implants.
Am J Clin Pathol 105(5):628�639.
Alpan O, Bachelder E, Isil E, Arnheiter H, Matzinger P. 2004.
‘Educated’ dendritic cells act as messengers from memory to
naive T helper cells. Nat Immunol 5(6):615�622.
Altankov G, Grinnell F, Groth T. 1996. Studies on the biocom-
patibility of materials: Fibroblast reorganization of substratum-
bound fibronectin on surfaces varying in wettability. J Biomed
Mater Res 30(3):385�391.
Anderson JM. 1994. Inflammation and the foreign body response.
Problems Gen Surg 11:147�160.
Bailey LO, Lippiatt S, Biancanello FS, Ridder SD, Washburn NR.
2005. The quantification of cellular viability and inflammatory
response to stainless steel alloys. Biomaterials 26(26):5296�302.
Balasubramanian V, Grusin NK, Bucher RW, Turitto VT, Slack
SM. 1999. Residence-time dependent changes in fibrinogen
adsorbed to polymeric biomaterials. J Biomed Mat Res
44(3):253�260.
Balcells M, Edelman ER. 2002. Effect of pre-adsorbed proteins
on attachment, proliferation, and function of endothelial cells. J
Cellular Physiol 191:155�161.
Brauker J, Carr-Brendel V, Martinson L, Crudele J, Johnston W,
Johnson R. 1995. Neovascularization of synthetic membranes
Macrophage cell adhesion and inflammation cytokines on magnetostrictive nanowires 289

Dow
nloa
ded
By:
[CD
L Jo
urna
ls A
ccou
nt] A
t: 19
:14
8 Ja
nuar
y 20
08 directed by membrane microarchitecture. J Biomed Mater Res
29:1517�1524.
Denis FA, Hanarp P, Sutherland DS, Gold J, Mustin C, Rouxhet
PG, Dufrene YF. 2002. Protein adsorption on model surfaces
with controlled nanotopography and chemistry. Langmuir
18(3):819�828.
Jenney CR, DeFife KM, Colton E, Anderson JM. 1998. Human
monocyte/macrophage adhesion, macrophage motility, and IL-
4-induced foreign body giant cell formation on silane-modified
surfaces in vitro. Student Research Award in the Master’s
Degree Candidate Category, 24th Annual Meeting of the
Society for Biomaterials, San Diego, CA, April 22�26, 1998.
J Biomed Mater Res 41(2):171�184.
Kanagaraja S, Lundstrom I, Nygren H, Tengvall P. 1996. Platelet
binding and protein adsorption to titanium and gold after short
time exposure to heparinized plasma and whole blood.
Biomaterials 17(23):2225�2232.
Lampin M, Warocquier C, Legris C, Degrange M, Sigot-Luizard
MF. 1997. Correlation between substratum roughness and
wettability, cell adhesion, and cell migration. J Biomed Mater
Res 36(1):99�108.
Lan S, Veiseh M, Zhang M. 2005. Surface modification of silicon
and gold-patterned silicon surfaces for improved biocompat-
ibility and cell patterning selectivity. Biosens Bioelectron
20(9):1697�1708.
Lin HY, Bumgardner JD. 2004. Changes in the surface oxide
composition of Co-Cr-Mo implant alloy by macrophage cells
and their released reactive chemical species. Biomaterials 25(7�8):1233�1238.
Luck M, Paulke BR, Schroder W, Blunk T, Muller RH. 1998.
Analysis of plasma protein adsorption on polymeric nanopar-
ticles with different surface characteristics. J Biomed Mat Res
39(3):478�485.
Martin JY, Schwartz Z, Hummert TW, Schraub DM, Simpson J,
Lankford J Jr, Dean DD, Cochran DL, Boyan BD. 1995. Effect
of titanium surface roughness on proliferation, differentiation,
and protein synthesis of human osteoblast-like cells (MG63). J
Biomed Mater Res 29(3):389�401.
Matzinger P. 2002. The danger model: A renewed sense of self.
Science 296(5566):301�305.
McFarland CD, Thomas CH, DeFilippis C, Steele JG, Healy KE.
2000. Protein adsorption and cell attachment to patterned
surfaces. J Biomed Mat Res 49(2):200�210.
Naji A, Harmand MF. 1990. Study of the effect of the surface
state on the cytocompatibility of a Co-Cr alloy using human
osteoblasts and fibroblasts. J Biomed Mater Res 24(7):861�871.
Ojo-Amaize EA, Lawless OJ, Peter JB. 1996. Elevated concentra-
tions of interleukin-1 beta and interleukin-1 receptor antagonist
in plasma of women with silicone breast implants. Clin Diagn
Lab Immunol 3(3):257�259.
Padera R, Colton C. 1996. Time course of membrane micro-
architecture-driven neovascularization. Biomaterials 17(3):
277�284.
Pallandre A, De Meersman B, Blondeau F, Nysten B, Jonas AM.
2005. Tuning the orientation of an antigen by adsorption onto
nanostriped templates. J Am Chem Soc 127(12):4320�4325.
Paul WE. 1994. Fundamental immunology. New York: Raven
Press.
Rosales C, Juliano RL. 1995. Signal transduction by cell adhesion
receptors in leukocytes. J Leukoc Biol 57(2):189�198.
Rosengren A, Bjursten LM. 2003. Pore size in implanted
polypropylene filters is critical for tissue organization. J Biomed
Mater Res Part A 67A(3):918�926.
Rosengren A, Bjursten LM, Danielsen N, Persson H, Kober M.
1999. Tissue reactions to polyethylene implants with different
surface topography. J Mater Sci �Mater Med 10(2):75�82.
Ruoslahti E, Pierschbacher MD. 1987. New perspectives in cell
adhesion: RGD and integrins. Science 238(4826):491�497.
Schieffer B, Selle T, Hilfiker A, Hilfiker-Kleiner D, Grote K,
Tietge UJ, Trautwein C, Luchtefeld M, Schmittkamp C,
Heeneman S, Daemen MJ, Drexler H. 2004. Impact of
interleukin-6 on plaque development and morphology in
experimental atherosclerosis. Circulation 110(22):3493�500.
Sen CK, Khanna S, Venojarvi M, Trikha P, Ellison EC, Hunt TK,
Roy S. 2002. Copper-induced vascular endothelial growth
factor expression and wound healing. Am J Physiol Heart
Circ Physiol 282(5):H1821�1827.
Sharma G, Mor GK, Varghese OK, Paulose M, Grimes CA.
2004. Synthesis and characterization of extremely uniform Fe-
Co-Ni ternary alloy nanowire arrays. J Nanosci Nanotechnol
4(7):738�743.
Skelsey ME, Mayhew E, Niederkorn JY. 2003. CD25�, inter-
leukin-10-producing CD4� T cells are required for suppressor
cell production and immune privilege in the anterior chamber
of the eye. Immunology 110(1):18�29.
Steele JG, Johnson G, Griesser HJ, Underwood PA. 1997.
Mechanism of initial attachment of corneal epithelial cells to
polymeric surfaces. Biomaterials 18(23):1541�1551.
Suh CW, Kim MY, Choo JB, Kim JK, Kim HK, Lee EK. 2004.
Analysis of protein adsorption characteristics to nano-pore
silica particles by using confocal laser scanning microscopy. J
Biotechnol 112(3):267�277.
Suska F, Gretzer C, Esposito M, Emanuelsson L, Wennerberg A,
Tengvall P, Thomsen P. 2005. In vivo cytokine secretion and
NF-kappaB activation around titanium and copper implants.
Biomaterials 26(5):519�527.
Wilson EH, Wille-Reece U, Dzierszinski F, Hunter CA. 2005. A
critical role for IL-10 in limiting inflammation during toxo-
plasmic encephalitis. J Neuroimmunol 165(1�2):63�74.
Zhang MQ, Desai T, Ferrari M. 1998. Proteins and cells on PEG
immobilized silicon surfaces. Biomaterials 19(10):953�960.
Zhao Q, Agger MP, Fitzpatrick M, Anderson JM, Hiltner A,
Stokes K, Urbanski P. 1990. Cellular interactions with bioma-
terials: In vivo cracking of pre-stressed Pellethane 2363-80A. J
Biomed Mater Res 24(5):621�637.
290 K. M. Ainslie et al.