lentiviral vectors that express ugt1a1 in liver and contain mir-142 target sequences normalize...
TRANSCRIPT

LT
FSG
*J
BauabaaUpttdmpmmdmwVcwpcclnpcrvrlo
Kt
GASTROENTEROLOGY 2010;139:999–1007
entiviral Vectors That Express UGT1A1 in Liver and Contain miR-142arget Sequences Normalize Hyperbilirubinemia in Gunn Rats
RANÇOISE SCHMITT,*,‡ SÉVERINE REMY,§ ANNE DARIEL,*,‡ MAUDE FLAGEUL,* VIRGINIE PICHARD,*ÉBASTIEN BONI,* CLAIRE USAL,§ ANNE MYARA,� SOPHIE LAPLANCHE,� IGNACIO ANEGON,§ PHILIPPE LABRUNE,*,¶
UILLAUME PODEVIN,*,‡ NICOLAS FERRY,* and TUAN HUY NGUYEN*
INSERM Unité 948, ‡Service de Chirurgie Pédiatrique, and §INSERM Unité 643, CHU Hôtel Dieu, Nantes, France; �Service de Biologie, Groupe Hospitalier Saint
oseph, Paris, France; and ¶Service de Pédiatrie, Hôpital Antoine Béclère, Clamart, FranceChltlcttdtetsCcbodrcgttgtosbcntU
CgfaU
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
See editorial on page 726.
ACKGROUND & AIMS: Crigler–Najjar type 1 (CN-I) isn inherited liver disease caused by an absence of bilirubin–ridine 5=-diphosphate–glucuronosyltransferase (UGT1A1)ctivity. It results in life-threatening levels of unconjugatedilirubin, and therapeutic options are limited. We useddult Gunn rats (an animal model of the disease) to evalu-te the efficiency of lentiviral-based gene therapy to expressGT1A1 in liver. METHODS: Gunn rats were given intra-ortal injections of VSVG-pseudotyped lentiviral vectorshat encode UGT1A1 under the control of a liver-specificransthyretin promoter (mTTR.hUGT1A1); this vectoroes not contain target sequences for miR-142, aicroRNA that is expressed specifically in hemato-
oietic cells. Rats were also injected with the vectorTTR.hUGT1A1.142T, which contains 4 copies of theiR-142 target sequences; its messenger RNA should be
egraded in antigen-presenting cells. Bilirubinemia wasonitored, and the presence of transduced hepatocytesas analyzed by quantitative polymerase chain reaction.ector expression was tested in vitro in rat hematopoieticells. RESULTS: In Gunn rats, bilirubin levels normalized 2eeks after administration of mTTR.hUGT1A1. However, hy-erbilirubinemia resumed 8 weeks after vector administration,oncomitant with the induction of an immune response. Inontrast, in rats injected with mTTR-UGT1A1.142T, bilirubinevels normalized for up to 6 months and transduced cells wereot eliminated. CONCLUSIONS: Lentiviral vectors that ex-ress UGT1A1 reduce hyperbilirubinemia in immuno-ompetent Gunn rats for at least 6 months. The immuneesponse against virally expressed UGT1A1 can be circum-ented by inclusion of miR-142 target sequences, whicheduce vector expression in antigen-presenting cells. Thisentiviral-based gene therapy approach might be devel-ped to treat patients with CN-I.
eywords: Crigler–Najjar Syndrome; Gene Transfer; Len-ivirus; MicroRNA.
rigler–Najjar disease type I (CN-I) is a rare recessiveinherited metabolic disorder characterized by very
igh levels of circulating unconjugated bilirubin. It causesife-threatening neurotoxic injury, which can ultimately leado fatal encephalopathy, and the only curative alternative isiver transplantation. This genetic disease is an attractiveandidate for gene therapy for a number of reasons. First,he molecular defect is well characterized. CN-I results fromhe complete absence of hepatic activity of bilirubin–uri-ine 5=-diphosphate–glucuronosyltransferase (UGT1A1),he enzyme responsible for conjugation of a bilirubin moi-ty to 1 or 2 molecules of glucuronic acid. After conjuga-ion, the hydrophobic bilirubin molecule is rendered wateroluble and can be excreted via the bile. In patients withN-I, nonconjugated bilirubin remains in the serum. It
oncentrates in organs with high lipid content such as therain, leading to lesions described as kernicterus. The sec-nd reason why CN-I is attractive for gene therapy is thaturing the course of the disease the histology of the liveremains completely normal. The morbidity of CN-I is ex-lusively related to extrahepatic deposition of nonconju-ated bilirubin. The normality of liver architecture main-ains the accessibility of hepatocytes to blood-borne generansfer vectors and thus preserves the efficiency of in vivoene therapy. A third reason is the availability of a model ofhe disease, the Gunn rat, that mimics the human pathol-gy and facilitates preclinical evaluation of various vectorystems and delivery strategies to correct the disease. Finally,ecause CN-I is a severe disease that can only be completelyured by liver transplantation, it deserves the exploration ofew therapeutic strategies such as gene therapy. Of note,here is no need for a complete and full restoration ofGT1A1 activity in patients with CN-1 to reach a valuable
Abbreviations used in this paper: APC, antigen-presenting cells; CN-I,rigler–Najjar type 1; FACS, fluorescence-activated cell sorter; GFP,reen fluorescent protein; miRNA, microRNA; MOI, multiplicity of in-ection; PGK, phosphoglycerate kinase; qPCR, quantitative polymer-se chain reaction; Treg, regulatory CD4� T cell; TU, transducing units;GT1A1, bilirubin–uridine 5=-diphosphate–glucuronosyltransferase.
© 2010 by the AGA Institute0016-5085/$36.00
doi:10.1053/j.gastro.2010.05.008

cantggtvabtlt
smcrnapinafcLct
tmtpsmpcitmiTsItesGi
m
mawas
mbcdfya3b
Ncd(ls3(g
ocmc(stpptp
2or(FGTt
St
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1000 SCHMITT ET AL GASTROENTEROLOGY Vol. 139, No. 3
linical result, and restoration of only 5% to 10% of enzymectivity is enough to maintain bilirubin levels below theeurotoxic threshold without using phototherapy. For allhese reasons, CN-I is a paradigm for gene therapy for liverenetic diseases, and many previous studies have focused onene delivery to the Gunn rat to correct the disease pheno-ype.1 Complete and sustained correction was achieved inivo using gutless adenoviral vectors delivered to the adultnimals or using retroviral vectors delivered to the new-orn.2–4 Ex vivo gene therapy using autologous hepatocytesransduced with lentiviral vectors also achieved significantevels of correction or liver repopulation.5 These experimen-al strategies are, however, difficult to translate to the clinics.
The easiest gene therapy protocol for CN-I would entail aingle administration of a gene transfer vector via a mini-
ally invasive route with ensuing complete and definitiveure with no adverse effects. We previously showed thatetroviral vectors could achieve this goal when delivered toewborn Gunn rats.2,3 However, because diagnosis is usu-lly delayed up to 4 to 8 weeks of age, gene therapy foratients with CN-I at birth is not achievable. Moreover, the
mmaturity of the immune system at birth in rodents doesot fully compare with the human situation. Our previousttempts to treat adult Gunn rats with retroviral vectorsailed because an immune response directed against theorrected hepatocytes was triggered after vector delivery.6
ong-term survival of transduced hepatocytes in adult ratsan only be achieved by using a pharmacologic manipula-ion of the host immune system.7
Therefore, we sought to improve the design of our vec-ors to thwart the induction of immune response while
aintaining a full transduction capacity in hepatocytes. Tohis end, we took advantage of the cell-specific expressionattern of some microRNA (miRNA) as previously de-cribed.8 In this setting, an miRNA target sequence for the
iR-142-3p (miR-142) is placed downstream of the thera-eutic complementary DNA (cDNA). The miR-142 is spe-ifically expressed in cells of the hematopoietic lineage,ncluding antigen-presenting cells (APCs). After administra-ion to mice, expression in APC is turned off because the
iR-142 present in the cell will bind to its target sequencen the messenger RNA (mRNA) encoding the transgene.his will result in degradation of the mRNA if the target
equences are perfectly matched to the miR-142 sequence.n the present report, we show that addition of 4 copies ofhe miR-142 target sequences (miR-142T) to the cDNAncoding UGT1A1 allowed escape from the immune re-ponse and resulted in complete and definitive correction ofunn rats after lentivirus delivery to adult animals with no
nduction of a cytotoxic immune response.
Materials and MethodsCell Culture and TransductionHeLa and 293T cells were cultured in Dulbecco’s
odified Eagle medium and NR8383 cells in Ham’s F12 i
edium containing 10% fetal bovine serum, glutamine,nd antibiotics. Cells were transduced at 5 � 104 cells/ell (HeLa) or 1 � 105 cells/mL (NR8383) for 24 hoursnd cultured for 7 days before fluorescence-activated cellorter (FACS) analysis.
Bone marrow cells were isolated from tibias and fe-urs of Gunn rats and cultured in X-vivo medium (Cam-
rex, Paris, France) in the presence of 5 ng/mL hepato-yte growth factor (R&D Systems, Lille, France) asescribed.9 They were transduced the day of cell isolationor 16 hours and cultured for 7 days before FACS anal-sis. Bone marrow– derived dendritic cells were obtainednd cultured as described.10 They were transduced on dayat 1 � 106 cells/mL for 4 hours and cultured for 5 days
efore FACS analysis.
Animals and Vector AdministrationAnimals were housed at the animal facilities of
antes University Medical School and received humaneare according to the guidelines of the French Ministèree l’Agriculture. Male Gunn j/j rats weighed 140 to 160 g
6 to 7 weeks old) and were maintained under a 12-houright cycle and fed ad libitum. All viral vectors from aame vector batch were injected intraportally using a0-gauge syringe at a dose of 1.5 � 107 transducing unitsTU)/g. All procedures were performed under isofluraneeneral anesthesia (3% vol/vol in air).
Lentiviral VectorsThe self-inactivating mTTR.hUGT1A1 was previ-
usly described and contained the human UGT1A1DNA under control of the liver-specific transthyretinTTR promoter.3 In the mTTR.hUGT1A1.142T vector, 4
opies of perfectly matched miR-142 target sequences11
kindly provided by Dr Luigi Naldini) were added down-tream of the expression cassette in mTTR.hUGT1A1 vec-or. The pCCL.sin.cPPT.PGK.GFP.WPRE (PGK.GFP) andCCL.sin.cPPT.PGK.GFP.WPRE.142T (PGK.GFP.142T) ex-ressing green fluorescent protein (GFP) under the con-rol of phosphoglycerate kinase (PGK) promoter werereviously described.8
Vectors were produced by transient transfection into93T cells as described.12 Vector titers were determinedn HeLa cells by real-time quantitative polymerase chaineaction (qPCR) on an ABI Prism 7000 using SYBR greenMesaGreen qPCR MasterMix; Eurogentec, Angers,rance) and primers specific for vector (GAG-F,GAGCTAGAACGATTCGCAGTTA; GAG-R, GGTTG-AGCTGTCCCAGTATTTGTC) and for �-actin.13 Vector
iters were routinely 5 � 109 TU/mL.
Liver Function TestsBlood was drawn from the retro-orbital sinus.
erum total bilirubin and alanine and aspartate amino-ransferase levels were measured at the Routine Biochem-
stry Department of Nantes University Hospital. The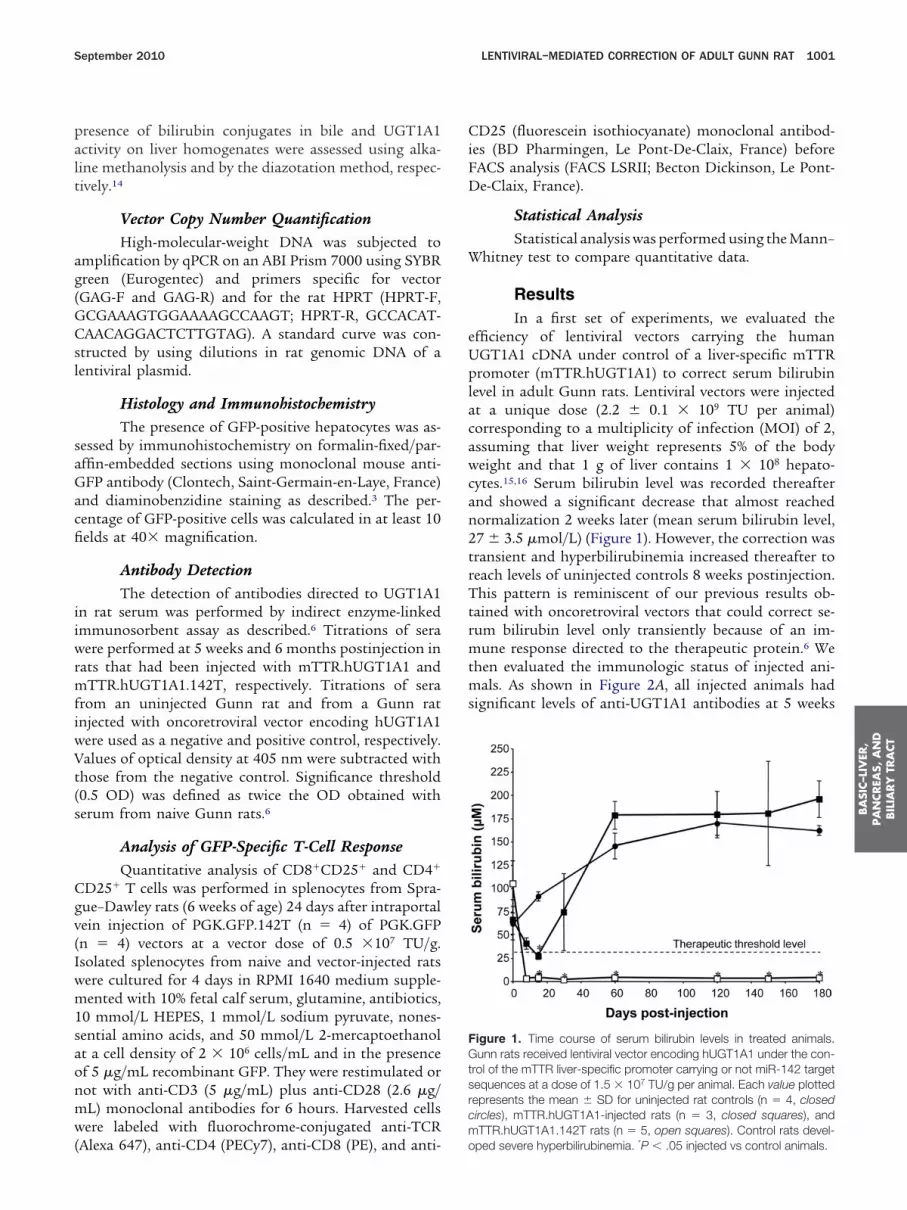
palt
ag(GCsl
saGacfi
iiwrmfiwVt(s
Cgv(Iwm1saonmw(
CiFD
W
eUplacawcan2trTtrmtms
FGtsrcm
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
September 2010 LENTIVIRAL–MEDIATED CORRECTION OF ADULT GUNN RAT 1001
resence of bilirubin conjugates in bile and UGT1A1ctivity on liver homogenates were assessed using alka-ine methanolysis and by the diazotation method, respec-ively.14
Vector Copy Number QuantificationHigh-molecular-weight DNA was subjected to
mplification by qPCR on an ABI Prism 7000 using SYBRreen (Eurogentec) and primers specific for vectorGAG-F and GAG-R) and for the rat HPRT (HPRT-F,CGAAAGTGGAAAAGCCAAGT; HPRT-R, GCCACAT-AACAGGACTCTTGTAG). A standard curve was con-
tructed by using dilutions in rat genomic DNA of aentiviral plasmid.
Histology and ImmunohistochemistryThe presence of GFP-positive hepatocytes was as-
essed by immunohistochemistry on formalin-fixed/par-ffin-embedded sections using monoclonal mouse anti-FP antibody (Clontech, Saint-Germain-en-Laye, France)
nd diaminobenzidine staining as described.3 The per-entage of GFP-positive cells was calculated in at least 10elds at 40� magnification.
Antibody DetectionThe detection of antibodies directed to UGT1A1
n rat serum was performed by indirect enzyme-linkedmmunosorbent assay as described.6 Titrations of seraere performed at 5 weeks and 6 months postinjection in
ats that had been injected with mTTR.hUGT1A1 andTTR.hUGT1A1.142T, respectively. Titrations of sera
rom an uninjected Gunn rat and from a Gunn ratnjected with oncoretroviral vector encoding hUGT1A1ere used as a negative and positive control, respectively.alues of optical density at 405 nm were subtracted with
hose from the negative control. Significance threshold0.5 OD) was defined as twice the OD obtained witherum from naive Gunn rats.6
Analysis of GFP-Specific T-Cell ResponseQuantitative analysis of CD8�CD25� and CD4�
D25� T cells was performed in splenocytes from Spra-ue–Dawley rats (6 weeks of age) 24 days after intraportalein injection of PGK.GFP.142T (n � 4) of PGK.GFPn � 4) vectors at a vector dose of 0.5 �107 TU/g.solated splenocytes from naive and vector-injected ratsere cultured for 4 days in RPMI 1640 medium supple-ented with 10% fetal calf serum, glutamine, antibiotics,
0 mmol/L HEPES, 1 mmol/L sodium pyruvate, nones-ential amino acids, and 50 mmol/L 2-mercaptoethanolt a cell density of 2 � 106 cells/mL and in the presencef 5 �g/mL recombinant GFP. They were restimulated orot with anti-CD3 (5 �g/mL) plus anti-CD28 (2.6 �g/L) monoclonal antibodies for 6 hours. Harvested cellsere labeled with fluorochrome-conjugated anti-TCR
Alexa 647), anti-CD4 (PECy7), anti-CD8 (PE), and anti- o
D25 (fluorescein isothiocyanate) monoclonal antibod-es (BD Pharmingen, Le Pont-De-Claix, France) beforeACS analysis (FACS LSRII; Becton Dickinson, Le Pont-e-Claix, France).
Statistical AnalysisStatistical analysis was performed using the Mann–
hitney test to compare quantitative data.
ResultsIn a first set of experiments, we evaluated the
fficiency of lentiviral vectors carrying the humanGT1A1 cDNA under control of a liver-specific mTTRromoter (mTTR.hUGT1A1) to correct serum bilirubin
evel in adult Gunn rats. Lentiviral vectors were injectedt a unique dose (2.2 � 0.1 � 109 TU per animal)orresponding to a multiplicity of infection (MOI) of 2,ssuming that liver weight represents 5% of the bodyeight and that 1 g of liver contains 1 � 108 hepato-
ytes.15,16 Serum bilirubin level was recorded thereafternd showed a significant decrease that almost reachedormalization 2 weeks later (mean serum bilirubin level,7 � 3.5 �mol/L) (Figure 1). However, the correction wasransient and hyperbilirubinemia increased thereafter toeach levels of uninjected controls 8 weeks postinjection.his pattern is reminiscent of our previous results ob-
ained with oncoretroviral vectors that could correct se-um bilirubin level only transiently because of an im-
une response directed to the therapeutic protein.6 Wehen evaluated the immunologic status of injected ani-
als. As shown in Figure 2A, all injected animals hadignificant levels of anti-UGT1A1 antibodies at 5 weeks
igure 1. Time course of serum bilirubin levels in treated animals.unn rats received lentiviral vector encoding hUGT1A1 under the con-
rol of the mTTR liver-specific promoter carrying or not miR-142 targetequences at a dose of 1.5 � 107 TU/g per animal. Each value plottedepresents the mean � SD for uninjected rat controls (n � 4, closedircles), mTTR.hUGT1A1-injected rats (n � 3, closed squares), andTTR.hUGT1A1.142T rats (n � 5, open squares). Control rats devel-
ped severe hyperbilirubinemia. *P � .05 injected vs control animals.
ahsrigcdiate
toaumuN1Gmalu1Pmttppi
c
teit3ttpomdcGGw
htrtTrawasCPtPtPtt
Flfim tibod
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1002 SCHMITT ET AL GASTROENTEROLOGY Vol. 139, No. 3
fter gene delivery, showing that lentivirally expressedUGT1A1 is immunogenic. Moreover, real-time PCRhowed that the vector genome was detected at a lowesidual level at 6 months postinjection when hyperbil-rubinemia had resumed (�0.001 vector copy per haploidenome). This low residual presence of transgene copiesould correspond to nonparenchymal cells, such as en-othelial cells, transduced but in which mTTR promoter
s inactive.11 These results strongly suggested that adultnimals were able to mount an immune response to theherapeutic proteins, resulting in a clearance of cellsxpressing hUGT1A1.
This induction of an immune response was likely to behe result of infection of APCs by lentiviral vectors and byff-target transgene expression in APCs despite the use ofliver-specific promoter.11 This was confirmed in vitro
sing GFP as a reporter gene driven by the mTTR pro-oter in lentiviral vectors. Using this vector, we detected
p to 2.5% of GFP-expressing cells after transduction ofR8383 rat macrophage cells (Supplementary Figure
A). A similar off-target GFP expression was observed inunn rat bone marrow– derived dendritic cells (Supple-entary Figure 1B), although the GFP expression was
bout 6-fold lower than that obtained with a PGK-GFPentiviral vector, in which GFP expression is driven by thebiquitous PGK promoter (mean of fluorescence of73 � 55 vs 1014 � 503 for mTTR and PGK, respectively;� .05; n � 3). This low level of transgene expressionay be sufficient to elicit an immune response against
he transgene product. Overall, these results clearly showhat, as suggested by others, the use of a tissue-specificromoter in lentiviral vectors is not able to blunt com-letely the expression of the transgene in APCs, resulting
n an immune clearance of the transduced cells.8
To circumvent more tightly the immune response, we
igure 2. Detection of hUGT1A1 antibodies by enzyme-linked immuninked immunosorbent assay in serum of (A) mTTR.hUGT1A1-injected anrom a Gunn rat injected with oncoretroviral vector encoding hUGT1Andividual animal. Results are expressed as arbitrary OD values with a s
TTR.hUGT1A1.142T vector did not lead to formation of hUGT1A1 an
onstructed a lentiviral vector containing the miR-142T o
o induce degradation of the mRNA in APCs. We firstvaluated the efficiency of the miR-142T in rat APCsnfected with vectors carrying GFP under the control ofhe PGK promoter (PGK.GFP.142T). As shown in FigureA and B, the proportion of GFP-positive cells was dras-ically reduced after infection with PGK.GFP.142T vec-ors at an MOI of 1. At a higher MOI, some cells ex-ressed GFP but still at a lower level than in the absencef the miR-142T. We then evaluated the potency of theiR-142T to repress GFP expression in Gunn rat den-
ritic cells. At an MOI of 20, 42% � 7.5% of rat dendriticells readily expressed GFP after incubation with PGK-FP vector (Figure 3C). After incubation with PGK.-FP.142T vector, the peak of highly positive GFP cellsas no longer present.We then determined the ability of the miR-142T to
amper in vivo a cellular immune response against aransgene product delivered to the rat liver using lentivi-al vector. After injection of PGK.GFP.142T or PGK vec-ors, splenocytes were isolated at 24 days postinjection.-cell activation was measured following exposure to
ecombinant GFP protein by FACS analyses of the CD25ctivation marker. Some GFP-stimulated T lymphocytesere also restimulated with anti-CD3 and anti-CD28ntibodies to increase the sensitivity of the assay. Ashown in Figure 3D, increased activation of CD8� andD4� T lymphocytes was detected in rats injected withGK-GFP vector. By contrast, a significant decrease inhe proportion of activated T cells was detected inGK.GFP.142T-injected rats. Interestingly, we observedhat CD4� and CD8� T lymphocytes isolated fromGK.GFP.142T-injected rats were resistant to stimula-ion with anti-CD3 and anti-CD28, as compared withhose isolated from PGK.GFP-injected and naive rats.
Thus, these results showed the cross-species efficiency
bent assay. Antibodies against hUGT1A1 were detected by enzyme-mTTR.hUGT1A1.142T-injected rats (closed squares). Titrations of serae used as a positive control (open squares). Each plot represents anance threshold of 0.5 at dilutions greater than 1:400. Administration ofies except in one animal (#138).
osord (B)
1 werignific
f the miR-142 in vitro and in vivo for repressing lenti-

vfGm
h
u�r6m
FhNitpvF � T c
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
September 2010 LENTIVIRAL–MEDIATED CORRECTION OF ADULT GUNN RAT 1003
iral gene expression from hematopoietic lineages andor down-regulating a cellular immune response againstFP, respectively. This prompted us to explore the sameiR-142T for treating Gunn rats in the long-term.A cohort of adult Gunn rats was injected with mTTR-
igure 3. Effect of addition of target sequences to miR-142 in vectoematopoietic cells were transduced with PGK.GFP.142T (PGK-mirT)R8383 cells were transduced at an MOI of 1 or 10. The percentage of
nfections were conducted with medium only (mock). (C) Gunn rat denransduction (mean � SD, n � 3) and mean intensity of fluorescence (eanels, respectively. (D) T-cell response to GFP in rats injected with PGKector-injected rats and cultured in the presence of recombinant GFPACS analysis to determine the proportion of activated CD8� and CD4
UGT1A1 vector harboring miR-142T. As shown in Fig- b
re 1, serum bilirubin level was normalized (2.8 � 1.9mol/L) as early as 1 week after lentivirus delivery and
emained stable for up to 6 months (end of the study). Atmonths, we checked by high-performance liquid chro-atography that bilirubin conjugates were present in the
kbone on transgene expression level in rat hematopoietic cells. RatPGK.GFP (PGK) lentiviral vectors. (A) Gunn rat bone marrow and (B)-positive cells is indicated in the lower right corner of each plot. Controlcells were transduced at an MOI of 20 (left panel). The percentage ofplot represents an individual animal) are shown in the middle and right.142T or PGK.GFP vectors. Splenocytes were isolated from naive anddays following restimulation or not with CD3/CD28 antibodies beforeells. Each plot corresponds to one experimental animal.
r bacandGFPdriticach.GFPfor 4
ile (Table 1), indicating that correction was actually due

twUavatqTomnclwanncTtvpsd
atcoTtles
iwatatcpaui
Ti2rvtlvlo
iaafav(gA
T
1.52
Fhfmha
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1004 SCHMITT ET AL GASTROENTEROLOGY Vol. 139, No. 3
o bilirubin conjugation. In untreated Gunn rats, thereere no bilirubin conjugates due to the absence ofGT1A1 (100% unconjugated bilirubin). We also evalu-
ted UGT1A1 activity in liver homogenates (Table 1). Thealues obtained corresponded to 44% to 372% of thectivity in the normal liver and confirmed liver transduc-ion and hUGT1A1 protein expression. We evaluated byPCR the amount of vector genome present in the liver.he results shown in Figure 4 demonstrate the stabilityf the vector content in the liver between 1 month and 6onths postinjection. The results also showed that the
umber of 0.22 vector copy/haploid genome was suffi-ient to cure the Gunn rat. Of note, the rat with theowest vector copy number in the liver at the time the ratsere killed (#92) was also the one with the highermount of unconjugated bilirubin in the bile. Unfortu-ately, antibodies directed against human UGT1A1 didot allow detection of transduced cells by immunohisto-hemistry, as has been also observed by others.17–19
herefore, to evaluate the in situ frequency and distribu-ion of transduced hepatocytes in Gunn rats, we injectedectors encoding GFP under the mTTR promoter anderformed immunohistochemical detection of GFP. Ashown in Figure 5, hepatocytes were massively trans-uced at a mean frequency of 47.67% � 6.6% at day 9
able 1. Assessment of Bilirubin Metabolism
Rat no. UGT1A1 activity (U/L) Unco
91 18.3792 25.85 293 15.36
138 26.1140 130.28
igure 4. Follow-up of vector genome content in the liver afterUGT1A1 gene transfer in cured Gunn rats. Genomic DNA was isolated
rom liver samples at 1 and 6 months after administration ofTTR.hUGT1A1.miR-142T vectors. The number of vector copies per
aploid genome (C/G) was determined by qPCR. Each plot represents
an individual animal.fter vector delivery. qPCR analysis of genomic DNA inransduced liver revealed 0.22 to 0.66 (0.41 � 0.23) vectoropy/haploid genome. These values are similar to thosebtained for mTTR.UGT1A1.142T-injected animals.ransduced hepatocytes were equally distributed be-
ween the liver lobes with a preferential localization iniver periportal areas. Of note, we did not detect GFP-xpressing nonparenchymal cells in the GFP-immuno-tained liver sections.
We evaluated the presence of anti-hUGT1A1antibodiesn the serum of treated animals. hUGT1A1 antibodiesere absent in all animals except one (#138) (Figure 2B),s compared with all animals receiving the vector lackinghe miR-142T. It is noteworthy that the presence ofntibodies at a high titer in rat #138 did not impair theherapeutic effect of the lentiviral vector. This may indi-ate that UGT1A1 expression, which is a microsomalrotein, was not affected by the presence of circulatingntibodies. Unfortunately, our repeated attempts to setp an assay in the Gunn rats to directly evaluate a cellular
mmune response were unsuccessful.In all animals, the liver histology remained normal.
he overall architecture of the parenchyma was unmod-fied as compared with controls (Supplementary Figure). Similarly, transaminase levels remained in the normalange at all time points. Finally, we analyzed by qPCR theector biodistribution. We observed that most of theransduction occurred in the liver. A lower transductionevel was also found in the spleen (representing 5% of thealue found in the liver) in all animals, as well as in theung and intestine for some animals and in the testis forne animal (#138) (Supplementary Table 1).
DiscussionCN-I is a paradigm for gene therapy for liver
nherited diseases, and Gunn rats have long been used asvaluable tool to evaluate gene transfer strategies usingpanel of gene transfer vectors. Gene therapy strategies
or the Gunn rat should circumvent immune response tochieve permanent correction of the genetic defect. Pre-ious studies have shown that adeno-associated viralAAV) or gutted adenoviral vectors could achieve thisoal. However, only partial correction was achieved withAV vectors and serum bilirubin level gradually increased
Bile derivatives (%)
ted Monoconjugates Diconjugates
40.47 45.957.03 13.140.38 54.0935.25 58.9821.97 70.13
njuga
5.820.961.362.34
fter gene delivery.18 AAV vectors are known to induce

invcrhtaawcgohtsarpoiwRtomdocsttgetchec
pawsvcmidtresroaPiwGPrttv(Ti
tddgdiilP
Fuhc ge of
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
September 2010 LENTIVIRAL–MEDIATED CORRECTION OF ADULT GUNN RAT 1005
mmune tolerance after gene delivery to the liver and areot able to infect APCs.20,21 However, the absence of AAVector integration may result in disappearance of theorrection. Gutted adenoviral vectors were shown to cor-ect completely the disease for the life of the animals.4 Aigh vector dose was required for efficient hepatocyteransduction, which is still questionable for a clinicalpplication because of the risk of acute toxic response todenoviral capsids.22 In addition, the vector copy numberas not evaluated and therefore a vector loss over time
annot be excluded.4 Indeed, similar to AAV vectors,utted adenoviral vectors do not integrate in the genomef the corrected cell and the correction may be lost whenepatocyte division is activated (eg, during acute hepati-is or during liver growth). It is therefore tempting topeculate that retroviral vectors will be able to maintain
constant level of correction whatever the subsequenteplication of corrected hepatocytes. Along this line, wereviously showed that concanavalin A–induced hepatitisr partial hepatectomy did not decrease correction level
n Gunn rats and the number of transduced hepatocyteshen using lentiviral vectors to ferry the transgene.23,24
etroviral integration increases the potential risk of vec-or genotoxicity, which was observed after transductionf hematopoietic stem cells.25–27 However, no liver tu-ors were reported after injection of human immuno-
eficiency virus 1– derived lentiviral vector by us andthers,23,28 in contrast to a non– human immunodefi-iency virus— based vector such as EIAV.29 Our previoustudies showed that one of the main issues regardingransgene delivery to the liver using retroviral vectors ishe induction of an immune response that is likely trig-ered by infection of APCs.6 Alternatively, we cannotxclude that cross-presentation of the therapeutic pro-ein engulfed by APCs by the major histocompatibilityomplex class I pathway can also trigger a cytotoxic orumoral immune response. Indeed, viral supernatants,specially when concentrated by ultracentrifugation, are
igure 5. Transduction efficacy in the rat liver after intraportal administnder the control of the mTTR promoter at a dose of 1.5 � 107 TU/g (nepatocytes in the liver was detected by immunohistochemistry. Hematopies/haploid genome (C/G) was determined by qPCR. The percenta
ontaminated by cell debris, including the therapeutic t
rotein that is expressed after transfection of the pack-ging cells. We previously showed that both pathwaysere accountable for the induction of an immune re-
ponse after in vivo delivery of �-galactosidase retroviralectors to hepatocytes.30 Gene transfer to immune in-ompetent newborns is an alternative to circumvent im-une response, but this approach is unlikely to be clin-
cally relevant for treating CN-I due to the delay iniagnosis of CN-I. Modification of the vector design ishe most promising strategy to circumvent the immuneesponse for future clinical application, particularly byxploiting the endogenous miRNA-mediated posttran-criptional pathway to induce degradation of vector-de-ived mRNA in APCs. In a seminal report, Brown et albserved strong expression of GFP limited to hepatocytesnd endothelial cells after intravenous injection ofGK.GFP.142T vectors in adult mice.8 In contrast,
n mice that received PGK.GFP vectors, GFP expressionas also detected in Kupffer cells and in splenocytes.FP expression was only sustained in mice receivingGK.GFP.142T vectors. This miR-142–regulated lentivi-al vector strategy was then successfully used to stablyreat hemophilic mice.11 Annoni et al recently showedhat systemic injection of miR-142–regulated lentiviralectors recruited and expanded regulatory CD4� T cellsTregs) in the liver with generation of GFP-specificregs.31 These Tregs were responsible for maintaining
mmunologic tolerance to GFP antigen in injected mice.In the present study, we extended this initial observa-
ion to another species and to another inherited liverisease using the same miR-142 target sequences.8 Weemonstrated in vitro a strong repression of lentiviralene expression in rat hematopoietic cells. We observedecreased GFP-specific T cell response in rats following
njection of PGK.GFP.142T, as compared with PGK.GFP-njected rats. Interestingly, we also observed that CD4� Tymphocytes and CD8� T lymphocytes isolated fromGK.GFP.142T-injected rats did not respond to stimula-
of lentiviral vectors. Gunn rats received lentiviral vector encoding GFPnd were killed at day 9 postinjection. (A) The presence of GFP-positivecounterstained. Original magnification 220�. (B) The number of vectorGFP-positive hepatocytes is also indicated for each animal.
ration� 3) aoxylin
ion with anti-CD3 and anti-CD28, in contrast to those

igTmtpdhmtterTbmpHstnbprUsi
fidwielalsevtponm
aG1
1
1
1
1
1
1
1
1
1
1
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1006 SCHMITT ET AL GASTROENTEROLOGY Vol. 139, No. 3
solated from PGK.GFP-injected and naive rats. This sug-ested (although did not prove definitely) the presence ofregs in PGK.GFP.142T-injected rats, as was shown inice.31 We next showed that the miR-142–regulated len-
iviral strategy was also valid for the delivery of a thera-eutic gene in a rat model of a human liver geneticisease. We achieved a complete and stable correction ofyperbilirubinemia in the Gunn rat injected withTTR.UGT1A1.142T. In contrast, correction was only
ransient in Gunn rats injected with mTTR.hUGT1A1 vec-or. The most likely explanation for the loss of therapeuticffect is a cytotoxic immune response against hUGT1A1esulting in immune clearance of transduced hepatocytes.his hypothesis is strengthened by the appearance of anti-odies to hUGT1A1 in all mTTR.hUGT1A1-injected ani-als. Unfortunately, we were not able to directly detect the
resence of hUGT1A1-specific T cells in injected Gunn rats.owever, our results with GFP as a surrogate marker
trongly suggest an identical mechanism. We also observedhat all but one rat treated with mTTR.UGT1A1.142T didot develop antibodies to UGT1A1. The presence of anti-odies in the mTTR.UGT1A1.142T-injected rats did notreclude a sustained and complete correction, as alsoeported by others.32 This is probably due to the fact thatGT1A1 is an intracellular protein located in the micro-
omal compartment and may be out of reach of circulat-ng antibodies.
Using miRNA target sequences to regulate the speci-city of transgene expression in various cell populationsoes not modify the endogenous amount of miRNAshen few copies of the target sequence are present. This
s indeed the case for miR-142, for which saturation ofndogenous level was only achieved with 10 copies of aentiviral vector carrying the miR-142T under control of
strong ubiquitous SFFV promoter.33 Because it is un-ikely that in vivo delivery of lentiviral vector may achieveuch a high level of transduction, we believe that usingndogenous miRNA regulation may be of clinical rele-ance for future application of lentiviral vectors for thereatment of liver genetic diseases. This might be decisiverogress toward clinical trials in patients with CN-I usingur designed miR-142–regulated lentiviral vector. We areow currently validating this strategy in nonhuman pri-ates before considering a clinical application.
Supplementary Material
Note: To access the supplementary materialccompanying this article, visit the online version ofastroenterology at www.gastrojournal.org, and at doi:0.1053/j.gastro.2010.05.008.
References
1. Nguyen TH, Ferry N. Gene therapy for liver enzyme deficiencies:what have we learned from models for Crigler-Najjar and tyrosine-
mia. Expert Rev Gastroenterol Hepatol 2007;1:155–177.2. Bellodi-Privato M, Aubert D, Pichard V, et al. Successful genetherapy of the Gunn rat by in vivo neonatal hepatic gene transferusing murine oncoretroviral vectors. Hepatology 2005;42:431–438.
3. Nguyen TH, Bellodi-Privato M, Aubert D, et al. Therapeutic lenti-virus-mediated neonatal in vivo gene therapy in hyperbiliru-binemic Gunn rats. Mol Ther 2005;12:852–859.
4. Toietta G, Mane VP, Norona WS, et al. Lifelong elimination ofhyperbilirubinemia in the Gunn rat with a single injection of helper-dependent adenoviral vector. Proc Natl Acad Sci U S A 2005;102:3930–3935.
5. Nguyen TH, Mainot S, Lainas P, et al. Ex vivo liver-directed genetherapy for the treatment of metabolic diseases: advances inhepatocyte transplantation and retroviral vectors. Curr Gene Ther2009;9:136–149.
6. Aubert D, Menoret S, Chiari E, et al. Cytotoxic immune responseblunts long-term transgene expression after efficient retroviral-mediated hepatic gene transfer in rat. Mol Ther 2002;5:388–396.
7. Puppi J, Guillonneau C, Pichard V, et al. Long term transgeneexpression by hepatocytes transduced with retroviral vectors re-quires induction of immune tolerance to the transgene. J Hepatol2004;41:222–228.
8. Brown BD, Venneri MA, Zingale A, et al. Endogenous microRNAregulation suppresses transgene expression in hematopoieticlineages and enables stable gene transfer. Nat Med 2006;12:585–591.
9. Wulf-Goldenberg A, Eckert K, Fichtner I. Cytokine-pretreatment ofCD34(�) cord blood stem cells in vitro reduces long-term cellengraftment in NOD/SCID mice. Eur J Cell Biol 2008;87:69–80.
0. Peche H, Trinite B, Martinet B, et al. Prolongation of heart allo-graft survival by immature dendritic cells generated from recipi-ent type bone marrow progenitors. Am J Transplant 2005;5:255–267.
1. Brown BD, Cantore A, Annoni A, et al. A microRNA-regulatedlentiviral vector mediates stable correction of hemophilia B mice.Blood 2007;110:4144–4152.
2. Bovia F, Salmon P, Matthes T, et al. Efficient transduction ofprimary human B lymphocytes and nondividing myeloma B cellswith HIV-1-derived lentiviral vectors. Blood 2003;101:1727–1733.
3. Nguyen TH, Birraux J, Wildhaber B, et al. Ex vivo lentivirus trans-duction and immediate transplantation of uncultured hepato-cytes for treating hyperbilirubinemic Gunn rat. Transplantation2006;82:794–803.
4. Black M, Billing BH, Heirwegh KP. Determination of bilirubinUDP-glucuronyl transferase activity in needle-biopsy specimensof human liver. Clin Chim Acta 1970;29:27–35.
5. Marcos R, Monteiro RA, Rocha E. Design-based stereologicalestimation of hepatocyte number, by combining the smooth op-tical fractionator and immunocytochemistry with anti-carcinoem-bryonic antigen polyclonal antibodies. Liver Int 2006;26:116–124.
6. Seglen PO. Preparation of rat liver cells. 3. Enzymatic require-ments for tissue dispersion. Exp Cell Res 1973;82:391–398.
7. Sauter BV, Parashar B, Chowdhury NR, et al. A replication-defi-cient rSV40 mediates liver-directed gene transfer and a long-termamelioration of jaundice in Gunn rats. Gastroenterology 2000;119:1348–1357.
8. Seppen J, van der Rijt R, Looije N, et al. Long-term correction ofbilirubin UDP-glucuronyltransferase deficiency in rats by in uterolentiviral gene transfer. Mol Ther 2003;8:593–599.
9. Thummala NR, Ghosh SS, Lee SW, et al. A non-immunogenicadenoviral vector, coexpressing CTLA4Ig and bilirubin-uridine-diphosphoglucuronateglucuronosyltransferase permits long-term,repeatable transgene expression in the Gunn rat model of Crigler-
Najjar syndrome. Gene Ther 2002;9:981–990.
2
2
2
2
2
2
2
2
2
2
3
3
3
3
R
UC
A
C
F
M
September 2010 LENTIVIRAL–MEDIATED CORRECTION OF ADULT GUNN RAT 1007
0. Loduca PA, Hoffman BE, Herzog RW. Hepatic gene transfer as ameans of tolerance induction to transgene products. Curr GeneTher 2009;9:104–114.
1. Jooss K, Yang Y, Fisher KJ, et al. Transduction of dendritic cellsby DNA viral vectors directs the immune response to transgeneproducts in muscle fibers. J Virol 1998;72:4212–4223.
2. Brunetti-Pierri N, Ng P. Progress and prospects: gene therapy forgenetic diseases with helper-dependent adenoviral vectors. GeneTher 2008;15:553–560.
3. Nguyen TH, Aubert D, Bellodi-Privato M, et al. Critical assessmentof lifelong phenotype correction in hyperbilirubinemic Gunn ratsafter retroviral mediated gene transfer. Gene Ther 2007;14:1270–1277.
4. Nguyen TH, Oberholzer J, Birraux J, et al. Highly efficient lentiviralvector-mediated transduction of nondividing, fully reimplantableprimary hepatocytes. Mol Ther 2002;6:199–209.
5. Arumugam PI, Higashimoto T, Urbinati F, et al. Genotoxic poten-tial of lineage-specific lentivirus vectors carrying the beta-globinlocus control region. Mol Ther 2009;17:1929–1937.
6. Hacein-Bey-Abina S, Garrigue A, Wang GP, et al. Insertional on-cogenesis in 4 patients after retrovirus-mediated gene therapy ofSCID-X1. J Clin Invest 2008;118:3132–3142.
7. Montini E, Cesana D, Schmidt M, et al. The genotoxic potential ofretroviral vectors is strongly modulated by vector design andintegration site selection in a mouse model of HSC gene therapy.J Clin Invest 2009;119:964–975.
8. Seppen J, van Til NP, van der Rijt R, et al. Immune response tolentiviral bilirubin UDP-glucuronosyltransferase gene transfer infetal and neonatal rats. Gene Ther 2006;13:672–677.
9. Themis M, Waddington SN, Schmidt M, et al. Oncogenesis fol-lowing delivery of a nonprimate lentiviral gene therapy vector to
fetal and neonatal mice. Mol Ther 2005;12:763–771. G0. Podevin G, Otta E, Nguyen JM, et al. Factors influencing immuneresponse after in vivo retrovirus-mediated gene transfer to theliver. J Gene Med 2004;6:16–21.
1. Annoni A, Brown BD, Cantore A, et al. In vivo delivery of amicroRNA-regulated transgene induces antigen-specific regula-tory T cells and promotes immunologic tolerance. Blood 2009;114:5152–5161.
2. Seppen J, Bakker C, de Jong B, et al. Adeno-associated virusvector serotypes mediate sustained correction of bilirubin UDPglucuronosyltransferase deficiency in rats. Mol Ther 2006;13:1085–1092.
3. Gentner B, Schira G, Giustacchini A, et al. Stable knockdown ofmicroRNA in vivo by lentiviral vectors. Nat Methods 2009;6:63–66.
Received September 14, 2009. Accepted May 11, 2010.
eprint requestsAddress requests for reprints to: Tuan Huy Nguyen, PhD, INSERM
nité 948, Biothérapies Hépatiques, CHU Hotel Dieu, 44093 Nantesedex, France. e-mail: [email protected].
cknowledgmentsN.F. and T.H.N. contributed equally to this work.
onflicts of interestThe authors disclose no conflicts.
undingSupported by grants from the Association Française contre lesyopathies (to A.M.) and the Association Francophone des
lycogénoses. M.F. received a fellowship from A.F.M.BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT

Sc1cl
1007.e1 SCHMITT ET AL GASTROENTEROLOGY Vol. 139, No. 3
upplementary Figure 1. Off-target transgene expression from a liveells and (B) Gunn rat dendritic cells were transduced with lentiviral vec0 and 20, respectively before FACS analysis. The percentage of GFP-porner of each plot. Control infections were done with medium only (
entiviral vector in transduced rat hematopoietic cells.
r-specific promoter in rat hematopioietic cells. (A) NR8383 rat macrophagetors encoding GFP under the control of the mTTR promoter at a MOI of 1 orositive cells and mean intensity of fluorescence are indicated in the lower rightmock). Significant expression of GFP was observed from the liver-specific

Sb
S
LKISTLH
N6a
September 2010 LENTIVIRAL–MEDIATED CORRECTION OF ADULT GUNN RAT 1007.e2
upplementary Figure 2. Hematoxylin and eosin stainings of paraffin-embedded liver sections of non-injected Gunn rat (left) and Gunn rat that has
een injected with mTTR.hUGT1A1 vectors (right) at month 6 post-vector delivery.upplementary Table 1. Evaluation of Vector Dissemination in Various Organs
Rat #91 #92 #93 #138 #140
iver 0.4839 0.2644 0.3944 0.7344 2.0735idney 0 0 0 0 0
ntestine 0.0027 0.0019 0 0.0035 0pleen 0.0178 0.031 0.0294 0.0976 0.0221estis 0 0 0 0.0031 0ung 0 0 0.0099 0.0428 0eart 0 0 0 0 0
OTE. Real-time quantitative PCR was performed on total DNA extracted from various organs from mTTR.hUGT1A1.142T-injected Gunn rats atmonths post-vector delivery, as described in Materials and Methods. The number of vector copies per haploid genome is shown for individual
nimals.