lecithinized superoxide dismutase: an inhibitor of human immunodeficiency virus replication
TRANSCRIPT

AIDS RESEARCH AND HUMAN RETROVIRUSESVolume 13, Number 4, 1997Mary Ann Liebert, Inc.
Lecithinized Superoxide Dismutase: An Inhibitor of HumanImmunodeficiency Virus Replication
MARIAPPAN PREMANATHAN,1 HIDEKI NAKASHIMA,1 RIE IGARASHI,2 YUTAKA MIZUSHIMA,2and KANEO YAMADA2
ABSTRACT
Superoxide dismutase (SOD) is an enzyme used in the treatment of oxygen radical-related diseases.Lecithinization of SOD enhances its pharmacological activity. Lecithinized SOD (PC-SOD) inhibits humanimmunodeficiency virus (HIV) types 1 and 2 in MT-4 cells. HIV-1-infected MT-4 cells were cultured for 5days in the presence of PC-SOD, at various concentrations. In an MTT assay, reverse transcriptase (RT) ac-
tivity of the cell extract and p24 antigen production were measured. Untreated, HIV-1-infected MT-4 cellsserved as control. PC-SOD inhibited viral replication most effectively at 2500 U/ml, a concentration that didnot affect cell viability, with an EC50 value of 718 U/ml. PC-SOD treatment inhibited RT activity and p24production in a dose-dependent manner. Western blot analysis of the HIV-1-infected MT-4 cells treated withPC-SOD at 2500 U/ml did not detect any expression of viral proteins. Failure to inhibit virus adsorption,proviral DNA and mRNA synthesis, and RT and proteinase enzyme activity suggests that the mechanism ofaction of PC-SOD is entirely different from those of the currently available anti-HIV drugs. PC-SOD showssynergistic interaction with AZT, ddl, ddC, KNI-272, and dextran sulfate. PC-SOD also inhibited the oxida-tive stress-induced depletion of sulfhydryls, which are the cause of diminished antioxidant defenses in III V
-infected patients.
Lecithinization of SOD potentiates its cell membrane affinity,cellular permeability, and pharmacological activity.4'5Currently, lecithinized SOD (PC-SOD) is undergoing clinicalevaluation for oxidative diseases. To provide a rationale forcombination therapy with PC-SOD, we have investigated theeffect of PC-SOD alone and in several combination ratios withthe currently available anti-HIV drugs and its inhibitory reac-
tivity on the radical-induced depletion of plasma sulfhydryls.
MATERIALS AND METHODS
Reagents and chemicalsThe following reagents were obtained from the indicated
companies: dextran sulfate (8 kDa) (Kowa, Tokyo); AZT,dideoxyinosine (ddl), and dideoxycytidine (ddC) (YamasaShoyu Co., Chiba, Japan); KNI-272 (Japan Energy, Tokyo);RPMI 1640 medium (GIBCO, Grand Island, NY); fetal calfserum (FCS) (Whittaker Bioproducts, Walkersville, MD); 3-
1 Department of Microbiology and Immunology, Kagoshima University School of Dentistry, Kagoshima-Shi 890, Japan,institute of Medical Science, St. Marianna University, Kawasaki, Kanagawa 216, Japan.
INTRODUCTION
Acquired Immunodeficiency Syndrome (AIDS) caused bythe human immunodeficiency virus (HIV) has remained a
health threat of global significance. Because of the limitationof currently available drugs, an extensive search for new anti-HIV agents is ongoing. Most of the currently available antivi-ral drugs show mitochondrial toxicity.1 The mitochondrial tox-icity of AZT (zidovudine) is due to mutant mitochondrial DNAcaused by oxygen radicals.2 Free radicals can increase the repli-cation of HIV and destroy immunocompetent cells such as Tcells.
Superoxide dismutase (SOD) is an enzyme that catalyzes thedismutation of the reactive and potentially harmful free radi-cals to less toxic hydrogen peroxide and molecular oxygen.Various attempts have been made to use it in the treatment ofoxygen radical-related diseases. It has been shown that HeLacells transfected with HIV tat gene markedly suppress the ex-
pression of Superoxide dismutase. Diminished antioxidase de-fenses cause the rapid depletion of plasma sulfhydryls.3
283

284 PREMANATHAN ET AL.
(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide(MTT) and 5,5'-dithio-bis(2-nitrobenzoic acid) (DTNB) (WakoPure Chemicals, Osaka, Japan); recombinant human CuZn-SOD (rhCuZn-SOD) (Ube Kosan Co., Ltd., Yamaguchi, Japan).PC-SOD, in which four molecules of a phosphatidylcholine(PC) derivative were covalently bound to each dimer ofrhCuZn-SOD,45 and potassium peroxochromate (^CrOs),6were synthesized as described earlier.
Cells and viruses
A human T lymphotropic virus type I (HTLV-I) positive Tcell line, MT-4,7 and lymphoblastoid T cell line, MOLT-4(clone No. 8),8 were subcultured twice a week at a concentra-tion of 3 X 105 cells/ml in RPMI 1640 medium supplementedwith 10% (v/v) heat-inactivated FCS. A strain of HIV-1 tub was
prepared from the culture supernatant of MOLT-4/HIV-lnmcells that were persistently infected with HIV-lnm. A strain ofHIV-2rod was prepared from the culture supernatant of MT-4cells infected with HIV-2ROd- Clinical isolates of HIV-lAoi2Band HIV-lAoi2D (AZT resistant) were also maintained in MT-4 cell cultures.
MTT assayThe inhibitory effect of the rhCuZn-SOD and PC-SOD on
HIV-1 replication was monitored by the inhibition of virus-in-duced cytopathogenicity in MT-4 cells. Briefly, MT-4 cellswere suspended at 3 X 105 cells/ml and infected with HIV-1 ata multiplicity of infection (MOI) of 0.01. The HIV-infected or
mock-infected MT-4 cells were placed in 96-well microtiterplates (200 /xl/well) and incubated at 37°C in a C02 incubatorin the presence of the compound. After 5 days, cell viabilitywas quantified by the MTT assay, as described previously,9,10from which the 50% cytotoxic concentration (CC50), 50% ef-fective concentration (EC50), and selectivity indices (SI =
CC50/EC50) were calculated.
Assay for viral expressionMT-4 cells were treated with virus for 90 min for the ad-
sorption of virus. After adsorption, the unbound virus was re-moved by repeated washing with the medium and then resus-
pended in medium containing various concentrations ofPC-SOD and incubated at 37°C. After 5 days of incubation, thenumber of viable cells was monitored by the trypan blue dyeexclusion method and HIV-1 antigen-positive cells were mon-itored by indirect immunofluorescence using serum from anAIDS patient and fluorescein isothiocyanate (FITC)-labeledanti-human IgG antibody.
Detection of HIV-1 p24gag antigenCell-free culture supernatant of MT-4 cells with or without
PC-SOD treatment was collected and the presence of p24 anti-gen was detected and quantified by the HIV-1 p24 core profileenzyme-linked immunosorbent assay (ELISA), using the methoddescribed by the manufacturer (Abbott GmbH Diagnostika,Wiesbaden-Delkenheim, Germany). Briefly, the standards were
run in the range of 12.5 to 100 pg/ml and the antigen-antibodycomplex was probed with a streptavidin-horseradish peroxide(HRP) conjugate. The end product was quantified by the inten-
sity of the color, which is directly proportional to the amount ofHIV-1 p24 core antigen captured. Color development was readat 492 nm, using a colorimeter.
Syncytium formation assayMOLT-4 cells (5 X 105) were cultured with an equal num-
ber of the MOLT-4/HIV-1 urn cells in microtiter plate wells con-
taining various concentrations of PC-SOD. After 24 hr of cocul-tivation, the number of giant cells (syncytium) was recorded bymicroscope examination and the fusion index was calculated as
described earlier.11
Virus adsorption assayThe inhibitory effect of PC-SOD on virus adsorption was
measured by an indirect immunofluorescence-laser flow cyto-fluorographic method.12 MT-4 cells were exposed to a high con-
centration of HIV-1 virions in the presence or absence of PC-SOD. The PC-SOD was added 1 min before the virus was
added. The cells were incubated for 1 hr at 37°C and washedtwice in phosphate-buffered saline (PBS) to remove the unab-sorbed virus. A high-titer polyclonal antibody derived from a
patient with AIDS-related complex (diluted 1:500 in PBS) was
then added. After 1 hr of incubation at 37°C, the cells were
washed twice with PBS. The cells were then incubated withFITC-conjugated F(ab')2 fragments of rabbit anti-human im-munoglobulin antibody (diluted 1:30 in PBS) for 1 hr at 37°C,washed twice in PBS, resuspended in 1 ml of 0.5%paraformaldehyde in PBS, and analyzed by laser flow cytoflu-orography.
Reverse transcriptase assayA reverse transcriptase (RT) assay was done by 3H-based
RT scintillation proximity, using the Quan-T-RT assay system(Amersham International pic, Buckinghamshire, England)13with recombinant HIV-1 RT enzyme (Seikagaku Co., Tokyo).Briefly, primer-template/beads, thymidine 5'-triphosphate(TTP)/[3H]TTP, recombinant RT enzyme, and PC-SOD were
mixed in an appropriate concentration and incubated at 37°C.After 1 hr, reaction was terminated by stop reagent and dilutedwith Tris-buffered saline (10 mM Tris-HCl [pH 7.4] and 0.15M NaCl) and counted by a scintillation counter.
Reverse transcriptase enzyme activity of the supernatant andcell extract of the HIV-infected MT-4 cells was assayed as de-scribed earlier.14,15 The cell cultures were harvested after 5 daysand separated from their supernatants by centrifugation. Thecell pellets were solubilized by vigorous vortexing in a lysisbuffer containing 0.2% Triton X-100 in 25 mM Tris-HCl (pH7.4). The supernatants and the solubilized samples were ana-
lyzed for RT enzyme activity as described above.
Proteinase enzyme assayThe HIV-1 proteinase enzyme assay was done by means of
a 125I-based scintillation proximity assay (SPA), using an HIVproteinase (125I)-SPA enzyme assay kit that is under develop-ment at Amersham International, with recombinant HIV-1 pro-teinase enzyme (AGMED, Inc., Bedford, MA). The 125I-labeledSPA beads, recombinant HIV proteinse enzyme, and PC-SODwere mixed in an appropriate concentration and incubated for

ANTI-HIV ACTIVITY OF LECITHINIZED SOD 285
2 hr at room temperature. The reaction was terminated by stopreagent and counted by a scintillation counter.
Western blot analysisThe presence of HIV-1 antigens in the culture supernatant
of MT-4 cells with or without PC-SOD treatment was deter-mined by Western blot analysis. MT-4 cells infected with HIV-Ihib and uninfected control cells were cultured for 6 days inthe presence of various concentrations of PC-SOD. After 6days, cell-free supernatant was centrifuged at 36,000 rpm at 4°Cfor 1 hr in an ultracentrifuge and the pellet was used for Westernblot assay. Viral proteins were denatured and separated by 12%sodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE) and transferred onto a Hybond-ECL (nitrocellu-lose) membrane filter.16 Western blot analysis was performedby the standard protocol, using an ECL Western blot kit(Amersham International) with HIV-positive serum as primaryantibody and biotinylated antibody as second antibody.17Polymerase chain reaction amplification
MT-4 cells were treated with virus and incubated for 90 minat 37°C for virus adsorption. The cells were then washed threetimes with medium and further incubated with media contain-ing different concentrations of PC-SOD. After 12 hr, totalmRNA and DNA were isolated from 107 cells. mRNA and DNAwere extracted by standard protocol, using a DYNALDynabeads mRNA direct kit (Dynal AS, Oslo, Norway) and an
IsoQuick (ORCA Research, Inc., Bothell, WA) nucleic acid ex-
traction kit, respectively. Reverse transcriptase-based poly-merase chain reaction (RT-PCR) was performed for mRNA am-
plification by use of a GeneAmp rTth reverse transcriptase RNAPCR kit (Perkin-Elmer/Roche Molecular Systems, Inc.,Branchburg, NJ) and primers SK38 and SK39 (upstream anddownstream primers of the gag gene sequence of HIV). ForDNA amplification, each reaction mixture contained 10 mMTris-HCl (pH 8.8), 50 mM KC1, 1.5 mM MgCl2, 0.1% TritonX-100, a 0.25 mM concentration of each of the four dNTPs,SK38 and SK39 primers (0.1 pM each), and 2.5 U of Taq DNApolymerase enzyme (Wako Pure Chemicals), overlaid with min-eral oil and amplified in a thermal cycler (Perkin-Elmer DNAthermal cycler).10Sulfhydryl determination
Both nonprotein and protein sulfhydryls were quantified bydisulfide exchange with DTNB at pH 8.18 Plasma (100 /¿l) was
added to 100 pi of sodium dodecyl sulfate (10%, w/v) andmixed thoroughly. Eight hundred microliters of phosphatebuffer (5 mM, pH 8.0) was added and the background absorp-tion read at 412 nm. The solution was then incubated for 1 hrat 37°C in the presence of 100 pi of DTNB (0.4 mg/ml). Theresulting thioquinone was measured in a photometer at 412 nm.
Reduced glutathione was used for calibration.
Analysis of drug combination effectThe inhibition of HIV-1 replication by combination of PC-
SOD and AZT, ddl, ddC, KNI-272, or dextran sulfate was eval-uated in experiments involving multiple concentration ratios ofthe drugs. For an experiment, different concentrations of each
drug, or drug combinations, were assayed in a checkerboardmanner. Antiviral activity in combination drug-treated HIV-in-fected MT-4 cells was determined by the protection againstHIV-induced cytopathic effect (CPE) assessed by the MTTmethod as described above. The combination indices (CIs) were
evaluated by three-dimensional analysis19 using a Macintoshcomputer (Apple Computer, Inc., Cupertino, CA) with a
Microsoft Excel spreadsheet (Microsoft Corp., Redmond, WA)and a Deltasoft graphics program (Delta Point, Inc., Monterey,CA).
RESULTS
Anti-HIV assayWhen rhCuZn-SOD and PC-SOD were evaluated for their in-
hibitory effect on the cytopathogenicity of HIV-1 in MT-4 cellsby MTT assay, rhCuZn-SOD did not show any activity (Fig.1 A). On the other hand, PC-SOD completely protected the cellsagainst virus-induced cell destruction at a concentration of 1250U/ml. It showed a dose-dependent inhibition of HIV-1 with a
mean 50% effective concentration (EC50) and EC90 values of718.1 and 1009.9 U/ml, respectively (Fig. IB). A 50% cytotox-icity (CC50) was observed at the concentration of 5217.1 U/ml.Anti-HIV activity of PC-SOD was observed with several strainsof both HIV-1 and HIV-2 in MT-4 cells. Irrespective of the cri-teria used to assess anti-HIV activity, i.e., inhibition of viral cy-topathogenicity, antigen expression, RT activity, and p24 pro-duction, PC-SOD invariably inhibited HIV-1, includingAZT-resistant strain, and HIV-2 replication within the EC50 con-
centration range of 422 to 846 U/ml (Table 1 ). For comparisonwe have also tested the anti-HIV activity of synthetic pseudoPC-SOD, which contains inactive SOD instead of active SOD,and did not observe any activity against HIV (data not shown).
PC-SOD inhibited viral expression in MT-4 cells in a dose-dependent manner as measured by immunofluorescence (IF)staining (Fig. IB) and reduced the amount of p24 in the cul-ture supernatant of MT-4 cells infected with HIV-1 (Fig. 2).The results were determined from the standard curve of knownamounts of p24 antigen and expressed in terms of nanogramsof p24 per milliliter of supernatant.
The effect of different concentrations of PC-SOD on HIV-1replication was quantitated by RT assay. Figure 3 shows theRT values from cell extracts of HIV-1-infected MT-4 cells withvarious concentrations of PC-SOD. The RT value for each sam-
ple was calculated from triplicate cultures and compared withthat of the infected control culture (100%). The EC50 was cal-culated from the graph. These results are in agreement with theMTT assay.
Virus adsorptionVarious experiments were undertaken to elucidate the mech-
anism of action of PC-SOD. First we investigated whether PC-SOD inhibited the binding of HIV particles to MT-4 cells, as
assessed by laser flow cytometry. PC-SOD inhibited HIV ad-sorption weakly, by only 35% at a concentration of 5000 U/ml.However, when we investigated the inhibitory activity againstmultinuclear giant cell (syncytium) formation in cocultures ofpersistently HIV-1-infected MOLT-4 cells (MOLT-4/HIV-
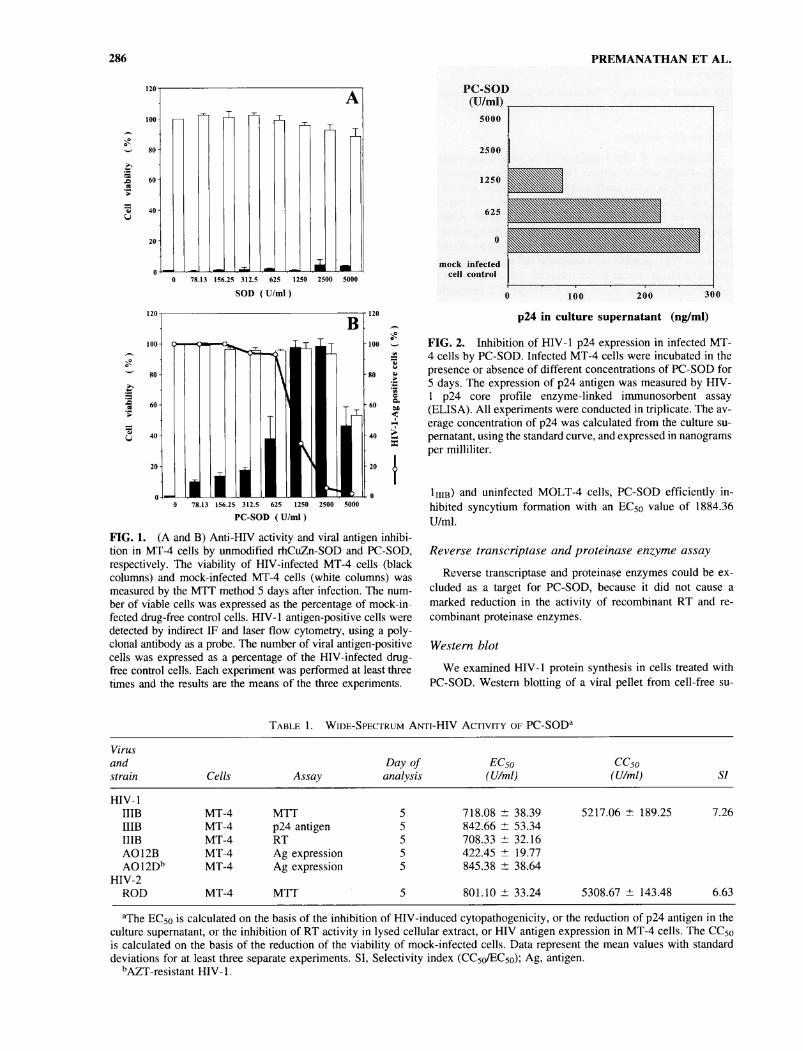
286 PREMANATHAN ET AL.
O 78.13 156.25 312.5 625 1250 2500 5000
SOD ( U/ml )
78.13 156.25 312.5 625 1250
PC-SOD (U/ml)FIG. 1. (A and B) Anti-HIV activity and viral antigen inhibi-tion in MT-4 cells by unmodified rhCuZn-SOD and PC-SOD,respectively. The viability of HIV-infected MT-4 cells (blackcolumns) and mock-infected MT-4 cells (white columns) wasmeasured by the MTT method 5 days after infection. The num-ber of viable cells was expressed as the percentage of mock-in-fected drug-free control cells. HIV-1 antigen-positive cells weredetected by indirect IF and laser flow cytometry, using a poly-clonal antibody as a probe. The number of viral antigen-positivecells was expressed as a percentage of the HIV-infected drug-free control cells. Each experiment was performed at least threerimes and the results are the means of the three experiments.
PC-SOD(U/ml)
5000
2500
1250
mock infectedcell control
0 100 200 300
p24 in culture supernatant (ng/ml)FIG. 2. Inhibition of HIV-1 p24 expression in infected MT-4 cells by PC-SOD. Infected MT-4 cells were incubated in thepresence or absence of different concentrations of PC-SOD for5 days. The expression of p24 antigen was measured by HIV-1 p24 core profile enzyme-linked immunosorbent assay(ELISA). All experiments were conducted in triplicate. The av-
erage concentration of p24 was calculated from the culture su-
pernatant, using the standard curve, and expressed in nanogramsper milliliter.
Iihb) and uninfected MOLT-4 cells, PC-SOD efficiently in-hibited syncytium formation with an EC50 value of 1884.36U/ml.
Reverse transcriptase and proteinase enzyme assayReverse transcriptase and proteinase enzymes could be ex-
cluded as a target for PC-SOD, because it did not cause a
marked reduction in the activity of recombinant RT and re-
combinant proteinase enzymes.
Western blot
We examined HIV-1 protein synthesis in cells treated withPC-SOD. Western blotting of a viral pellet from cell-free su-
Table 1. Wide-Spectrum Anti-HIV Activity of PC-SODa
Virusandstrain Cells Assay
Day ofanalysis
EC50(U/ml)
CC50(U/ml) SI
HIV-1IIIBIIIBIIIBA012BA012Db
HIV-2ROD
MT-4MT-4MT-4MT-4MT-4
MT-4
MTTp24 antigenRTAg expressionAg expressionMTT
718.08842.66708.33422.45845.38
38.3953.3432.1619.7738.64
5217.06 ± 189.25 7.26
801.10 ± 33.24 5308.67 ± 143.48 6.63
"The EC50 is calculated on the basis of the inhibition of HIV-induced cytopathogenicity, or the reduction of p24 antigen in theculture supernatant, or the inhibition of RT activity in lysed cellular extract, or HIV antigen expression in MT-4 cells. The CC50is calculated on the basis of the reduction of the viability of mock-infected cells. Data represent the mean values with standarddeviations for at least three separate experiments. SI, Selectivity index (CC5o/EC50); Ag, antigen.
bAZT-resistant HIV-1.
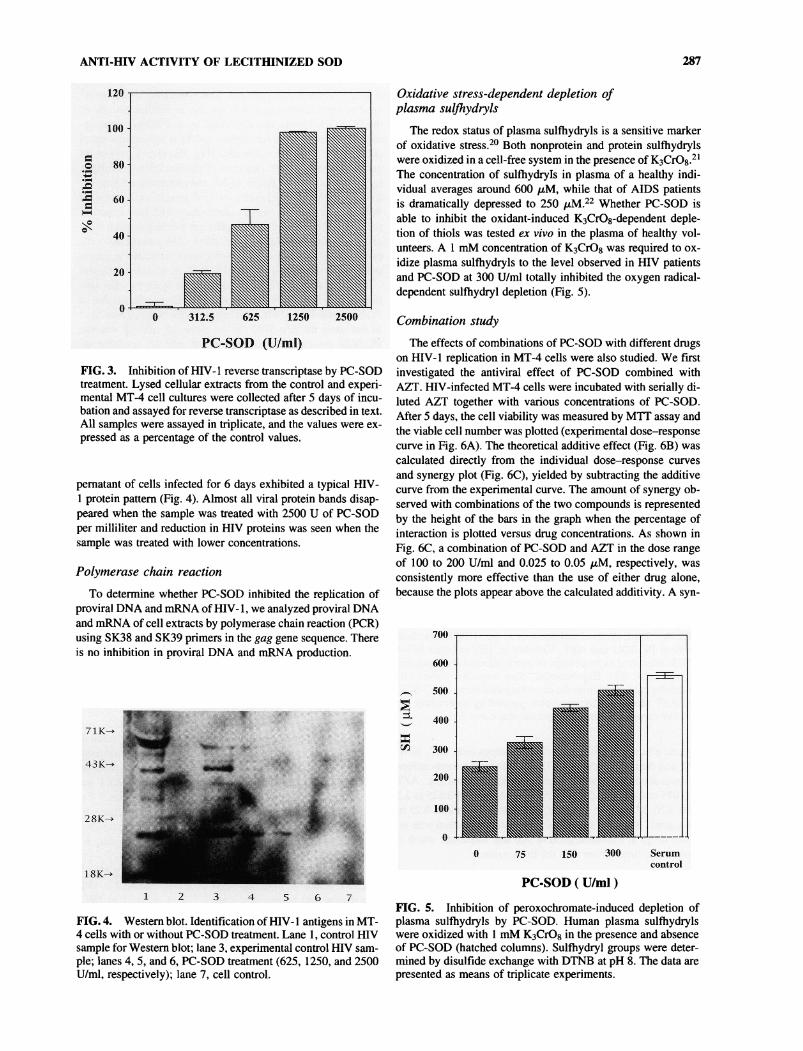
ANTI-HIV ACTIVITY OF LECITHINIZED SOD 287
120
100
ao
à?
PC-SOD (U/ml)FIG. 3. Inhibition of HIV-1 reverse transcriptase by PC-SODtreatment. Lysed cellular extracts from the control and experi-mental MT-4 cell cultures were collected after 5 days of incu-bation and assayed for reverse transcriptase as described in text.All samples were assayed in triplicate, and the values were ex-
pressed as a percentage of the control values.
pernatant of cells infected for 6 days exhibited a typical HIV-1 protein pattern (Fig. 4). Almost all viral protein bands disap-peared when the sample was treated with 2500 U of PC-SODper milliliter and reduction in HIV proteins was seen when thesample was treated with lower concentrations.
Polymerase chain reaction
To determine whether PC-SOD inhibited the replication ofproviral DNA and mRNA of HIV-1, we analyzed proviral DNAand mRNA of cell extracts by polymerase chain reaction (PCR)using SK38 and SK39 primers in the gag gene sequence. Thereis no inhibition in proviral DNA and mRNA production.
71K-
43K-
28K-
18K^~. JÊL.
i 7
FIG. 4. Western blot. Identification of HIV-1 antigens in MT-4 cells with or without PC-SOD treatment. Lane 1, control HIVsample for Western blot; lane 3, experimental control HIV sam-
ple; lanes 4, 5, and 6, PC-SOD treatment (625, 1250, and 2500U/ml, respectively); lane 7, cell control.
Oxidative stress-dependent depletion ofplasma sulfhydryls
The redox status of plasma sulfhydryls is a sensitive markerof oxidative stress.20 Both nonprotein and protein sulfhydrylswere oxidized in a cell-free system in the presence of K3Q-O8.21The concentration of sulfhydryls in plasma of a healthy indi-vidual averages around 600 pM, while that of AIDS patientsis dramatically depressed to 250 pM.22 Whether PC-SOD isable to inhibit the oxidant-induced K3Cr08-dependent deple-tion of thiols was tested ex vivo in the plasma of healthy vol-unteers. A 1 mM concentration of K3Cr08 was required to ox-
idize plasma sulfhydryls to the level observed in HIV patientsand PC-SOD at 300 U/ml totally inhibited the oxygen radical-dependent sulfhydryl depletion (Fig. 5).
Combination studyThe effects of combinations of PC-SOD with different drugs
on HIV-1 replication in MT-4 cells were also studied. We firstinvestigated the antiviral effect of PC-SOD combined withAZT. HIV-infected MT-4 cells were incubated with serially di-luted AZT together with various concentrations of PC-SOD.After 5 days, the cell viability was measured by MTT assay andthe viable cell number was plotted (experimental dose-responsecurve in Fig. 6A). The theoretical additive effect (Fig. 6B) was
calculated directly from the individual dose-response curves
and synergy plot (Fig. 6C), yielded by subtracting the additivecurve from the experimental curve. The amount of synergy ob-served with combinations of the two compounds is representedby the height of the bars in the graph when the percentage ofinteraction is plotted versus drug concentrations. As shown inFig. 6C, a combination of PC-SOD and AZT in the dose rangeof 100 to 200 U/ml and 0.025 to 0.05 pM, respectively, was
consistently more effective than the use of either drug alone,because the plots appear above the calculated additivity. A syn-
S
0 4-
75 150 300
PC-SOD ( U/ml )
Serumcontrol
FIG. 5. Inhibition of peroxochromate-induced depletion ofplasma sulfhydryls by PC-SOD. Human plasma sulfhydrylswere oxidized with 1 mM K3CrOg in the presence and absenceof PC-SOD (hatched columns). Sulfhydryl groups were deter-mined by disulfide exchange with DTNB at pH 8. The data are
presented as means of triplicate experiments.
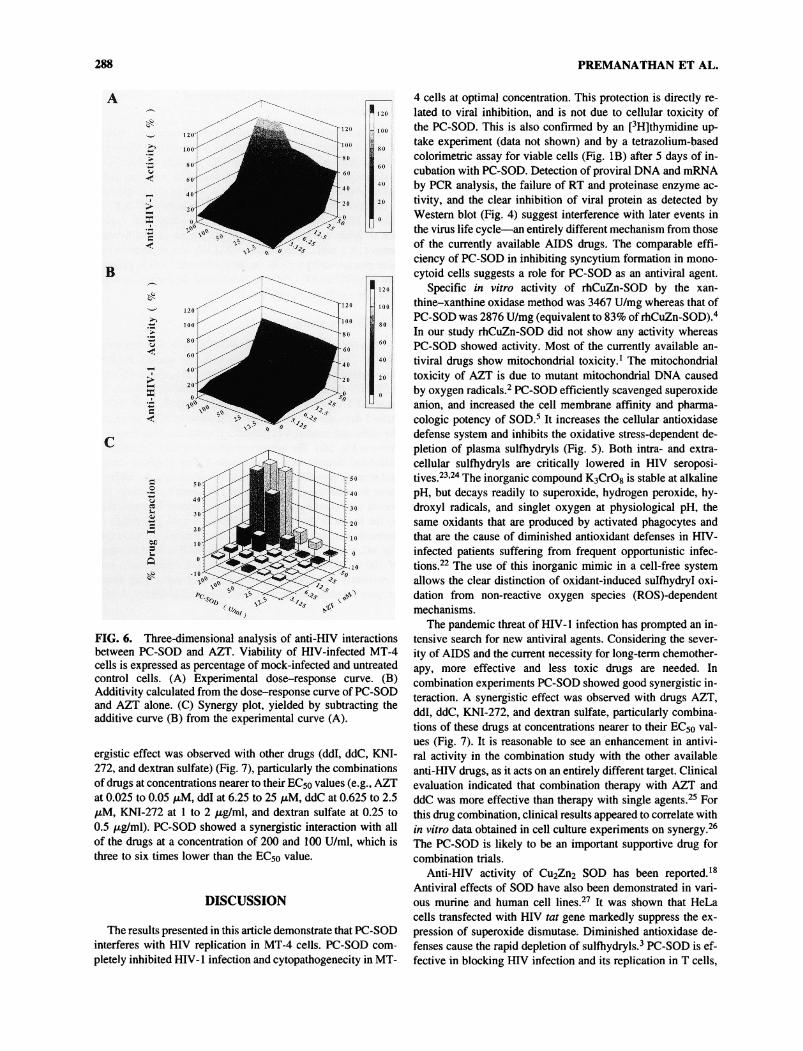
288 PREMANATHAN ET AL.
g=:
>¡Ec<
1„
O-' J*"« "^
B
5e<
a
FIG. 6. Three-dimensional analysis of anti-HIV interactionsbetween PC-SOD and AZT. Viability of HIV-infected MT-4cells is expressed as percentage of mock-infected and untreatedcontrol cells. (A) Experimental dose-response curve. (B)Additivity calculated from the dose-response curve of PC-SODand AZT alone. (C) Synergy plot, yielded by subtracting theadditive curve (B) from the experimental curve (A).
ergistic effect was observed with other drugs (ddl, ddC, KNI-272, and dextran sulfate) (Fig. 7), particularly the combinationsof drugs at concentrations nearer to their EC50 values (e.g., AZTat 0.025 to 0.05 pM, ddl at 6.25 to 25 pM, ddC at 0.625 to 2.5/xM, KNI-272 at 1 to 2 pg/ml, and dextran sulfate at 0.25 to0.5 /u,g/ml). PC-SOD showed a synergistic interaction with allof the drugs at a concentration of 200 and 100 U/ml, which isthree to six times lower than the EC50 value.
DISCUSSION
The results presented in this article demonstrate that PC-SODinterferes with HIV replication in MT-4 cells. PC-SOD com-
pletely inhibited HIV-1 infection and cytopathogenecity in MT-
4 cells at optimal concentration. This protection is directly re-
lated to viral inhibition, and is not due to cellular toxicity ofthe PC-SOD. This is also confirmed by an [3H]thymidine up-take experiment (data not shown) and by a tetrazolium-basedcolorimetric assay for viable cells (Fig. IB) after 5 days of in-cubation with PC-SOD. Detection of proviral DNA and mRNAby PCR analysis, the failure of RT and proteinase enzyme ac-
tivity, and the clear inhibition of viral protein as detected byWestern blot (Fig. 4) suggest interference with later events inthe virus life cycle—an entirely different mechanism from thoseof the currently available AIDS drugs. The comparable effi-ciency of PC-SOD in inhibiting syncytium formation in mono-
cytoid cells suggests a role for PC-SOD as an antiviral agent.Specific in vitro activity of rhCuZn-SOD by the xan-
thine-xanthine oxidase method was 3467 U/mg whereas that ofPC-SOD was 2876 U/mg (equivalent to 83% of rhCuZn-SOD).4In our study rhCuZn-SOD did not show any activity whereasPC-SOD showed activity. Most of the currently available an-
tiviral drugs show mitochondrial toxicity.1 The mitochondrialtoxicity of AZT is due to mutant mitochondrial DNA causedby oxygen radicals.2 PC-SOD efficiently scavenged Superoxideanión, and increased the cell membrane affinity and pharma-cologie potency of SOD.5 It increases the cellular antioxidasedefense system and inhibits the oxidative stress-dependent de-pletion of plasma sulfhydryls (Fig. 5). Both intra- and extra-cellular sulfhydryls are critically lowered in HIV seroposi-tives.23,24 The inorganic compound K3CrOs is stable at alkalinepH, but decays readily to Superoxide, hydrogen peroxide, hy-droxyl radicals, and singlet oxygen at physiological pH, thesame oxidants that are produced by activated phagocytes andthat are the cause of diminished antioxidant defenses in HIV-infected patients suffering from frequent opportunistic infec-tions.22 The use of this inorganic mimic in a cell-free systemallows the clear distinction of oxidant-induced sulfhydryl oxi-dation from non-reactive oxygen species (ROS)-dependentmechanisms.
The pandemic threat of HIV-1 infection has prompted an in-tensive search for new antiviral agents. Considering the sever-
ity of AIDS and the current necessity for long-term chemother-apy, more effective and less toxic drugs are needed. Incombination experiments PC-SOD showed good synergistic in-teraction. A synergistic effect was observed with drugs AZT,ddl, ddC, KNI-272, and dextran sulfate, particularly combina-tions of these drugs at concentrations nearer to their EC50 val-ues (Fig. 7). It is reasonable to see an enhancement in antivi-ral activity in the combination study with the other availableanti-HIV drugs, as it acts on an entirely different target. Clinicalevaluation indicated that combination therapy with AZT andddC was more effective than therapy with single agents.25 Forthis drug combination, clinical results appeared to correlate within vitro data obtained in cell culture experiments on synergy.26The PC-SOD is likely to be an important supportive drug forcombination trials.
Anti-HIV activity of Cu2Zn2 SOD has been reported.18Antiviral effects of SOD have also been demonstrated in vari-ous murine and human cell lines.27 It was shown that HeLacells transfected with HIV tat gene markedly suppress the ex-
pression of Superoxide dismutase. Diminished antioxidase de-fenses cause the rapid depletion of sulfhydryls.3 PC-SOD is ef-fective in blocking HIV infection and its replication in T cells,
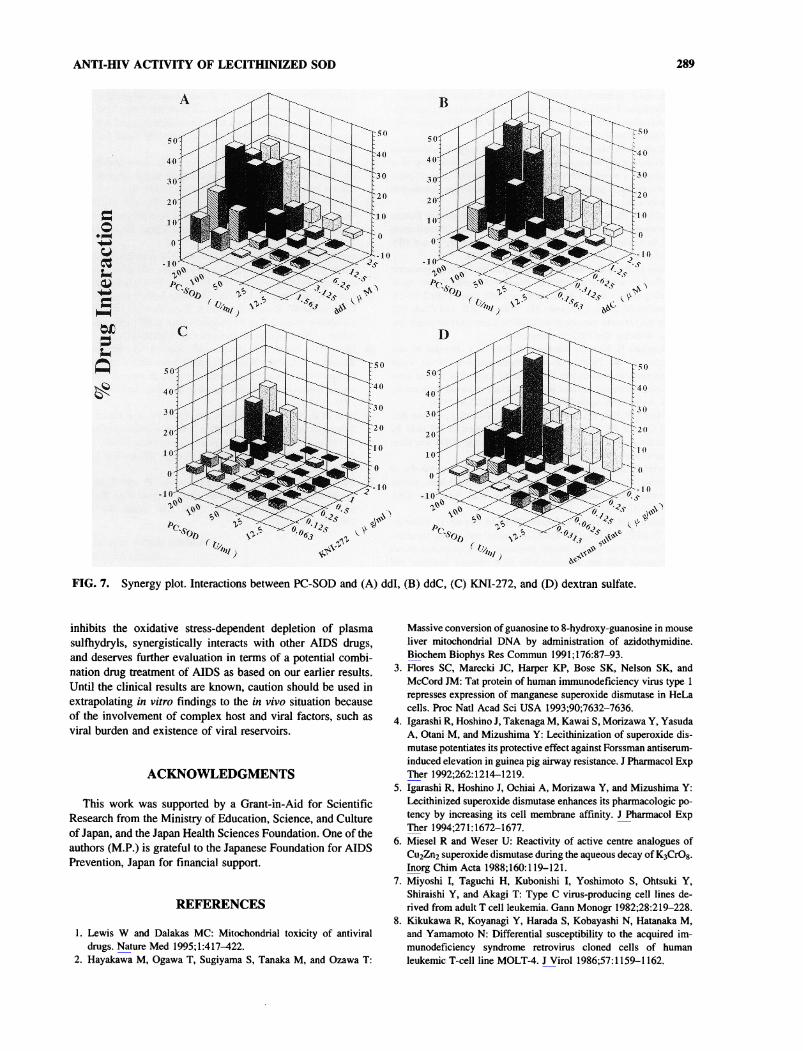
ANTI-HIV ACTIVITY OF LECITHINIZED SOD 289
CO
03-
0>
OJD
Q
"(1/ Ç» #•
FIG. 7. Synergy plot. Interactions between PC-SOD and (A) ddl, (B) ddC, (C) KNI-272, and (D) dextran sulfate.
inhibits the oxidative stress-dependent depletion of plasmasulfhydryls, synergistically interacts with other AIDS drugs,and deserves further evaluation in terms of a potential combi-nation drug treatment of AIDS as based on our earlier results.Until the clinical results are known, caution should be used inextrapolating in vitro findings to the in vivo situation becauseof the involvement of complex host and viral factors, such as
viral burden and existence of viral reservoirs.
ACKNOWLEDGMENTS
This work was supported by a Grant-in-Aid for ScientificResearch from the Ministry of Education, Science, and Cultureof Japan, and the Japan Health Sciences Foundation. One of theauthors (M.P.) is grateful to the Japanese Foundation for AIDSPrevention, Japan for financial support.
REFERENCES
1. Lewis W and Dalakas MC: Mitochondrial toxicity of antiviraldrugs. Nature Med 1995; 1:417^122.
2. Hayakawa M, Ogawa T, Sugiyama S, Tanaka M, and Ozawa T:
Massive conversion of guanosine to 8-hydroxy-guanosine in mouse
liver mitochondrial DNA by administration of azidothymidine.Biochem Biophys Res Commun 1991;176:87-93.
3. Flores SC, Marecki JC, Harper KP, Bose SK, Nelson SK, andMcCord JM: Tat protein of human immunodeficiency virus type 1represses expression of manganese Superoxide dismutase in HeLacells. Proc Nati Acad Sei USA 1993;90;7632-7636.
4. Igarashi R, Hoshino J, Takenaga M, Kawai S, Morizawa Y, YasudaA, Otani M, and Mizushima Y: Lecithinization of Superoxide dis-mutase potentiates its protective effect against Forssman antiserum-induced elevation in guinea pig airway resistance. J Pharmacol ExpTher 1992;262:1214-1219.
5. Igarashi R, Hoshino J, Ochiai A, Morizawa Y, and Mizushima Y:Lecithinized Superoxide dismutase enhances its pharmacologie po-tency by increasing its cell membrane affinity. J Pharmacol ExpTher 1994;271:1672-1677.
6. Miesel R and Weser U: Reactivity of active centre analogues ofCu2Zn2 Superoxide dismutase during the aqueous decay of ^CrOg.Inorg Chim Acta 1988;160:119-121.
7. Miyoshi I, Taguchi H, Kubonishi I, Yoshimoto S, Ohtsuki Y,Shiraishi Y, and Akagi T: Type C virus-producing cell lines de-rived from adult T cell leukemia. Gann Monogr 1982;28:219-228.
8. Kikukawa R, Koyanagi Y, Harada S, Kobayashi N, Hatanaka M,and Yamamoto N: Differential susceptibility to the acquired im-munodeficiency syndrome retrovirus cloned cells of humanleukemic T-cell line MOLT-4. J Virol 1986;57:1159-1162.

290 PREMANATHAN ET AL.
9. Pauwels R, Balzarini J, Baba M, Snoeck R, Schols D, HerdewijnP, Desmyter J, and De Clercq E: Rapid and automated tetrazolium-based colorimetric assay for the detection of anti-HIV compounds.J Virol Methods 1988;20:309-321.
10. Nakashima H, Masuda M, Murakami T, Koyanagi Y, MatsumotoA, Fujii N, and Yamamoto N: Anti-human immunodeficiency virusactivity of a novel synthetic peptide, T22 ([Tyr-5,12,Lys-7] poly-phemusin II): A possible inhibitor of virus-cell fusion. AntimicrobAgents Chemother 1992;36:1249-1255.
11. Tochikura TS, Nakashima H, Tanabe A, and Yamamoto N: Humanimmunodeficiency virus (HlV)-induced cell fusion: Quantificationand its application for the simple and rapid screening of anti-HIVsubstances in vitro. Virology 1988;164:542-546.
12. Schols D, Baba M, Pauwels R, and De Clercq E: Flow cytometricmethod to demonstrate whether anti-HIV-1 agents inhibit virionbinding to T4+ cells. J Acquir Immune Defic Syndr 1989;2:10-15.
13. Lamaitre M, Phan T, Downes MJ, Williams KB, and Cook ND:Poly r(A) reverse transcriptase (3H)SPA, a new enzyme assay sys-tem. Antiviral Res 1992;17 (Suppl.):48.
14. Nakashima H, Kido Y, Kobayashi N, Motoki Y, Neushul M, andYamamoto N: Purification and characterization of an avianmyeloblastosis and human immunodeficiency virus reverse tran-
scriptase inhibitor, sulfated polysaccharides extracted from sea al-gea. Antimicrob Agents Chemother 1987;31:1524-1528.
15. Selvam MP, Blay RA, Geyer S, Buck SM, Pollock L, Mayner RE,and Epstein JS: Inhibition of HIV-1 replication in H9 cells byNystatin-A compared with other antiviral agents. AIDS Res HumRetroviruses 1993;9:475-481.
16. Towbin H, Staehelin T, and Gordon J: Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets:Procedure and some applications. Proc Nati Acad Sei USA1979;76:4350-4354.
17. Nakashima H, Ichiyama K, Inazawa K, Ito M, Hayashi H, NishiharaY, Tsujii E, and Kino T: FR901724, a novel anti-human immuno-deficiency virus (HIV) peptide produced by streptomyces showssynergistic antiviral activities with HIV protease inhibitor and2',3'-dideoxynucleosides. Biol Pharm Bull 1996;19:405-112.
18. Miesel R, Mahmood N, and Weser U: Activity of Cu2Zn2 super-oxide dismutase against the human immunodeficiency virus type1. RedoxRep 1995;1:99-103.
19. Prichard MN and Shipman C Jr: A three-dimensional model to ana-
lyze drug-drug interactions (review). Antiviral Res 1990;14:181-206.20. Frei B, Stocker R, and Ames BN: Antioxidant defenses and lipid
peroxidation in human blood plasma. Proc Nati Acad Sei USA1988;85:9748-9752.
21. Miesel R and Haas R: Reactivity of an active centre analogue ofCu2Zn2 Superoxide dismutase in a murine model of acute andchronic inflammation. Inflammation 1993;17:595-611.
22. Eck HP, Gmunder H, Hartmann M, Petzold D, Daniel V, and DrogeW: Low concentration of acid-soluble thiol (cysteine) in the bloodplasma of HIV-1-infected patients. Biol Chem Hoppe Seyler1989;370:101-108.
23. Buhl R, Holroyd KJ, Mastrangeli A, Cantin AM, Jaffe HA, Wells FB,Saltini C, and Crystal RG: Systemic glutathione deficiency in symp-tom-free HTV-seropositive individuals. Lancet 1989;ii: 1294—1298.
24. Staal FJT, Roederer M, Isrealski DM, Bubp J, Mole LA, McshaneD, Deresinki SC, Ross W, Sussman H, Raju PA, et ai: Intracellularglutathione levels in T-cell subsets decrease in HIV-infected indi-viduals. AIDS Res Hum Retroviruses 1992;8:305-311.
25. Meng TC, Fischl MA, Boota AM, Spector SA, Bennett D,Bassiakos Y, Lai SH, Wright B, and Richman DD: Combinationtherapy with zidovudine and dideoxycytidine in patients with ad-vanced human immunodeficiency virus infection. Ann Intern Med1992;116:13-20.
26. Eron JJ Jr, Johnson VA, Merrill DP, Chou TC, and Hirsch MS:Synergistic inhibition of replication of human immunodeficiencyvirus type 1, including that of a zidovudine-resistant isolate, by zi-dovudine and 2',3'-dideoxycytidine in vitro. Antimicrob AgentsChemother 1992;36:1559-1562.
27. Pottathil R, Chandrabose KA, Cuatrecasas P, and Lang DJ:Establishment of the interferon-mediated antiviral state: Possiblerole of Superoxide dismutase. Proc Nati Acad Sei USA1981;78:3343-3347.
Address reprint requests to:Hideki Nakashima
Department of Microbiology and ImmunologyKagoshima University School of Dentistry
8-35-1 SakuragaokaKagoshima-Shi 890, Japan

This article has been cited by:
1. T. Greenwell-Wild, N. Vazquez, W. Jin, Z. Rangel, P. J. Munson, S. M. Wahl. 2009. Interleukin-27 inhibition of HIV-1involves an intermediate induction of type I interferon. Blood 114:9, 1864-1874. [CrossRef]
2. Jun Fang, Takahiro Seki, Hiroshi Maeda. 2009. Therapeutic strategies by modulating oxygen stress in cancer and inflammation.Advanced Drug Delivery Reviews 61:4, 290-302. [CrossRef]
3. M TAKENAGA, H TAMAMURA, K HIRAMATSU, N NAKAMURA, Y YAMAGUCHI, A KITAGAWA, S KAWAI,H NAKASHIMA, N FUJII, R IGARASHI. 2004. A single treatment with microcapsules containing a CXCR4 antagonistsuppresses pulmonary metastasis of murine melanoma. Biochemical and Biophysical Research Communications 320:1, 226-232.[CrossRef]
4. Mariappan Premanathan, Rieko Arakaki, Hiroyuki Izumi, Kandasamy Kathiresan, Masatoshi Nakano, Naoki Yamamoto, HidekiNakashima. 1999. Antiviral properties of a mangrove plant, Rhizophora apiculata Blume, against human immunodeficiencyvirus. Antiviral Research 44:2, 113-122. [CrossRef]
5. Hiroshi Sono, Keisuke Teshigawara, Tetsurou Sasada, Yasushi Takagi, Akira Nishiyama, Yuji Ohkubo, Yasuhiro Maeda, EijiTatsumi, Akihisa Kanamaru, Junji Yodoi. 1999. Redox Control of Epstein-Barr Virus Replication by Human Thioredoxin/ATL-Derived Factor: Differential Regulation of Lytic and Latent Infection. Antioxidants & Redox Signaling 1:2, 155-165.[Abstract] [Full Text PDF] [Full Text PDF with Links]
6. Mariappan PREMANATHAN, Kandasamy KATHIRESAN, Naoki YAMAMOTO, Hideki NAKASHIMA. 1999. In VitroAnti-Human Immunodeficiency Virus Activity of Polysaccharide from Rhizophora mucronata Poir. Bioscience, Biotechnology,and Biochemistry 63:7, 1187-1191. [CrossRef]