lara etal2014 postharvest bio technol
TRANSCRIPT

R
T
Ia
b
A
a
ARA
KCDDFWW
1
praml
itcbiaitmvc
0h
Postharvest Biology and Technology 87 (2014) 103–112
Contents lists available at ScienceDirect
Postharvest Biology and Technology
jou rn al h om epage: www.elsev ier .com/ locate /postharvbio
eview
he fruit cuticle as a modulator of postharvest quality
sabel Laraa,∗, Burcu Belgea, Luis F. Goulaob
Departament de Química, Unitat de Postcollita-XaRTA, Universitat de Lleida, Rovira Roure 191, 25198 Lleida, SpainAgri4Safe/BioTrop, Instituto de Investigac ão Científica Tropical (IICT), Polo Mendes Ferrão–Pavilhão de Agro-Indústrias e Agronomia Tropical, Tapada dajuda, 1349-017 Lisboa, Portugal
r t i c l e i n f o
rticle history:eceived 2 May 2013ccepted 10 August 2013
eywords:utinecayisordersirmness
a b s t r a c t
The composition and structure of fruit surface tissues have a noticeable influence on the postharveststorage potential of fruit, inasmuch as they behave as a barrier against drying, chemical attack, mechanicalinjuries and microbial infection. The cuticle is made of cutin, a biological insoluble polyester, embedded inan impermeable wax complex, and its inner side interacts intimately with the underlying epidermal cellwalls. The cuticle plays a decisive role in plant development, being the first communication system withthe surrounding biotic and abiotic environment. Published reports on the composition and biosynthesisof fruit cuticles are comparatively scarce, and many knowledge gaps exist on the part cuticles play inquality determination and postharvest performance. This review aims at collecting available information
ater lossax
in relation to the role of the fruit cuticle as a determinant factor of some important traits related topostharvest quality, including water loss, susceptibility to physical and biological stresses, and decreasedfruit firmness. To the best of our knowledge, this is the first published work focusing on the fruit cuticleas a major modulator of postharvest quality and interlinking existing dispersed literature on this topic.A deeper understanding of cuticle structure and function will be of help in understanding postharvestbiology and in designing new technological solutions.
. Introduction
Metabolic events that occur during maturation, ripening andostharvest development in fruit cause significant changes thateduce the marketable volume of the harvested product, the visualnd organoleptic appeal to the consumer, the level of available vita-ins and antioxidants, and storage ability. These occurrences cause
arge economic losses to the fruit crop sector.The fruit cuticle has been largely disregarded with respect to
ts putative influence in modulating fruit development, and in par-icular, fruit ripening and postharvest performance. However, theuticle is synthesised by, and covers, the epidermis of the fruit,eing the first barrier against abiotic and biotic conditions in which
t develops (Domínguez et al., 2011a). The main function tradition-lly attributed to fruit cuticles is to minimise water loss, althought also limits the loss of substances from internal tissues, pro-ects against physical, chemical and biological attack, and provides
echanical support to maintain plant organ integrity. The preser-
ation of all these functions requires structural integrity of theuticle throughout fruit expansion and development.∗ Corresponding author. Tel.: +34 973 702526; fax: +34 973 238264.E-mail address: [email protected] (I. Lara).
925-5214/$ – see front matter © 2013 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.postharvbio.2013.08.012
© 2013 Elsevier B.V. All rights reserved.
Cuticles contribute to traits such as maximum strain, breakingstress or elastic modulus, which affect the mechanical behaviour offruit and other plant organs. The viscoelastic and strain-hardeningproperties of cuticles contribute to reinforcing the epidermal cellwalls, while simultaneously conferring extensibility (Domínguezet al., 2011a). In turn, such biophysical properties are dependentto some extent upon external conditions such as temperature andrelative humidity (Edelmann et al., 2005; Matas et al., 2005). Cuti-cle strength and rigidity decrease when temperature increases,the structure exhibiting a phase transition at a given temperature.The effect of this transition temperature on the elastic modulus isdependent on relative humidity, as water is known to plasticise theplant cuticle (reviewed in Domínguez et al., 2011b). These two fac-tors are thus among the most important ones in devising storagestrategies for postharvest preservation. This review focuses specifi-cally on the current available knowledge on the relevance of cuticlecomposition and properties for fruit quality during the postharvestperiod (see Fig. 1).
2. A brief overview of cuticle composition in fruit
Cuticles are lipidic layers mainly composed by cutin, a polyesterpolymer rich in hydroxylated and epoxy-hydroxylated C16 andC18 fatty acids. This cutin matrix is embedded with amorphouswaxes and a minor fraction of phenolics, while the plant surface is

104 I. Lara et al. / Postharvest Biology and Technology 87 (2014) 103–112
harac
ctpwemRca2cp2eicYav
cmhotadncshbcpWtcts
3r
ds
Fig. 1. Major unanswered questions on the interactions between cuticle c
overed by epicuticular waxes, both amorphous and crystalline. Onhe inner side of the cuticle, cutin is mixed with pectin and glucanolysaccharides from the epidermal cell walls, the composition ofhich closely resembles that of primary cell walls (López-Casado
t al., 2007). The cuticle also contains cutan, a non-ester networkade of aliphatic compounds assembled mainly by ether bonds.
ecent reviews have summarised available information on theomposition and biosynthesis of cutin and cuticular waxes (Kunstnd Samuels, 2003, 2009; Nawrath and Poirier, 2008; Pollard et al.,008; Samuels et al., 2008). Some studies have shown that cuti-le composition differs significantly in different organs of the samelant, both regarding cutin (Espelie et al., 1979, 1980; Marga et al.,001; Järvinen et al., 2010) and waxes (Radler, 1965, 1970; Bakert al., 1975). In order to shed light on the possible roles of the cuticlen fruit quality and postharvest performance, therefore, it would beonvenient to have data on the specific composition of fruit cuticles.et published reports on cuticle composition of fruit are compar-tively scarce, research efforts having been focused primarily onegetative tissues.
Currently available information on cutin and cuticular waxomposition in different fruit types is summarised in Table 1. Inany cases, the composition of either cutin or cuticular waxes
as been reported separately, and thus for some fruit types theverview of cuticle constituents is still incomplete. Regardless ofhis, this survey shows that fruit cuticles display substantial vari-bility according to species, genotypes within a given species, andevelopmental stage. In most of the reports summarised in Table 1,-alkanes and triterpenoids have been identified as prominentomponents of cuticular waxes, in some cases accompanied byignificant amounts of aldehydes. Among the n-alkanes, the C29ydrocarbon n-nonacosane is regularly cited as a major compound,eing the most abundant in cuticles of apple, citrus fruit and sweetherry, while the C31 n-hentriacontane has been identified as theredominant alkane in other species such as pepper and tomato.ith respect to the triterpenoid components of cuticular waxes,
he triterpene ursolic and oleanoic acids dominate cuticular waxomposition of apple, grape, peach and sweet cherry, whereasriterpenoid alcohols such as amyrins are predominant in citruspecies, Asian pear, pepper and tomato.
. Cuticle biosynthesis during fruit maturation andipening
Some studies have addressed cuticle biosynthesis during fruitevelopment, mainly from a morphological or quantitative per-pective. Cuticle deposition reportedly ceases early during fruit
teristics and fruit quality attributes impacting postharvest management.
development, prior to the onset of the ripening process and fre-quently before the fruit has attained maximum size, resulting indecreased amounts of cuticle per surface area and thus in reducedcuticle thickness in ripe fruit (Rosenquist and Morrison, 1988;Comménil et al., 1997; Belding et al., 1998; Dong et al., 2012; Liuet al., 2012). In some cases, early-arrested deposition of cuticularcomponents and the associated decline in cuticle thickness causesmicrocracks as surface strain increases when fruit expand (Salaet al., 1992; Knoche et al., 2004; Knoche and Peschel, 2007; Peschelet al., 2007; Khanal et al., 2011; Becker and Knoche, 2012). A notableexception to this generally observed trend is tomato, for which acontinuous increase in cuticular waxes and cutin monomers hasbeen found during fruit development (Kosma et al., 2010).
In contrast, the compositional change of specific cuticular com-ponents during fruit maturation has been reported for only a fewspecies, including grape berries (Comménil et al., 1997), apple(Belding et al., 1998; Dong et al., 2012), sweet cherry (Peschel et al.,2007), tomato (Saladié et al., 2007; Kosma et al., 2010) and orange(Liu et al., 2012). This has revealed substantial differences in thetime-course changes of particular wax and cutin constituents foreach fruit type, thus illustrating the need to undertake such studieson a case-by-case basis.
In spite of the important implications for fruit quality andpostharvest performance, few efforts have been devoted to thestudy of cuticle formation in fruit, particularly from the biochemi-cal and molecular perspectives, although some information existsfor tomato (Vogg et al., 2004; Leide et al., 2007; Mintz-Oron et al.,2008; Isaacson et al., 2009; Nadakuduti et al., 2012; Yeats et al.,2012a; Shi et al., 2013), sweet cherry (Alkio et al., 2012), and apple(Albert et al., 2013). These studies have allowed the identificationof genes potentially involved in cuticular wax or cutin biosynthe-sis in fruit surfaces, which should facilitate further research on theformation of this important outer layer.
For tomato, a very long-chain fatty acid �-ketoacyl-CoA syn-thase (LeCER6), required for the biosynthesis of very long-chain(>C30) n-alkanes and aldehydes, has been identified and charac-terised (Vogg et al., 2004), and its loss-of-function mutant fruitdisplayed altered cuticle permeability and transpiration properties(Leide et al., 2007). Accordingly, Mintz-Oron et al. (2008) reportedprogressively increased expression of SlCER6 (referred to as LeCER6in the previous papers by Vogg et al., 2004, and Leide et al., 2007)throughout maturation and ripening of tomato fruit. Some other
cuticle-related genes have been also identified in tomato whichare involved in cutin deposition (Isaacson et al., 2009; Nadakudutiet al., 2012; Yeats et al., 2012b; Shi et al., 2013), as well as a tran-scription factor that regulates fruit cuticle formation and epidermal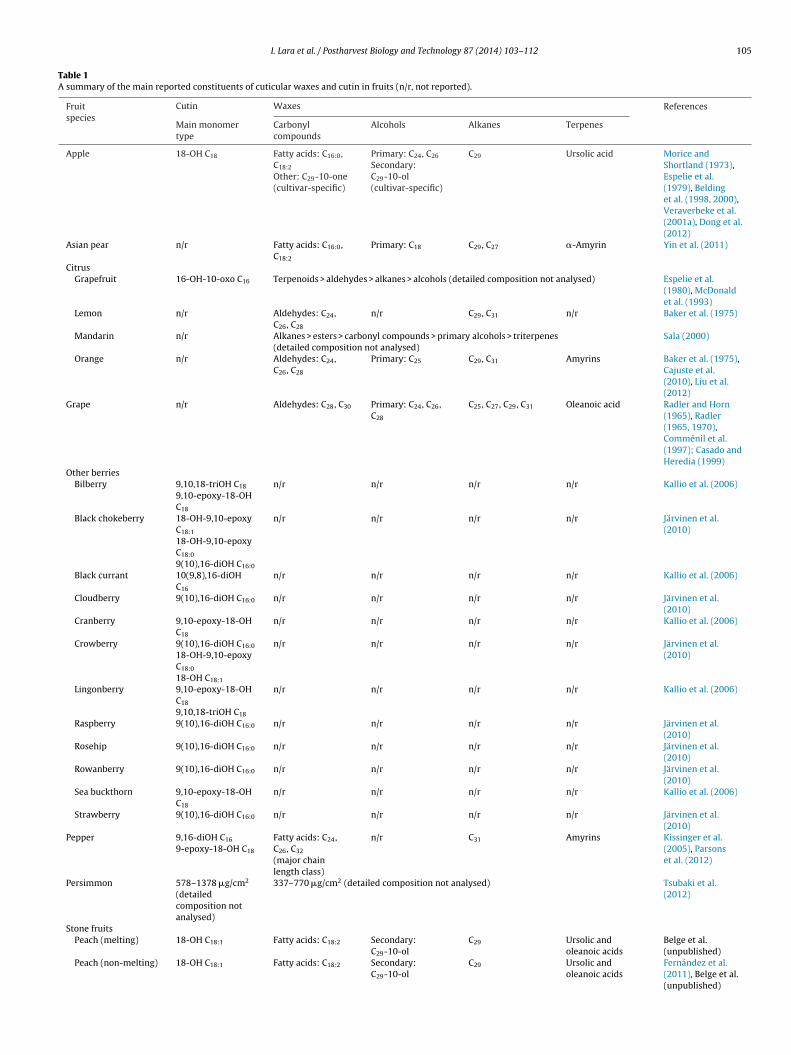
I. Lara et al. / Postharvest Biology and Technology 87 (2014) 103–112 105
Table 1A summary of the main reported constituents of cuticular waxes and cutin in fruits (n/r, not reported).
Fruitspecies
Cutin Waxes References
Main monomertype
Carbonylcompounds
Alcohols Alkanes Terpenes
Apple 18-OH C18 Fatty acids: C16:0,C18:2
Other: C29-10-one(cultivar-specific)
Primary: C24, C26
Secondary:C29-10-ol(cultivar-specific)
C29 Ursolic acid Morice andShortland (1973),Espelie et al.(1979), Beldinget al. (1998, 2000),Veraverbeke et al.(2001a), Dong et al.(2012)
Asian pear n/r Fatty acids: C16:0,C18:2
Primary: C18 C29, C27 �-Amyrin Yin et al. (2011)
CitrusGrapefruit 16-OH-10-oxo C16 Terpenoids > aldehydes > alkanes > alcohols (detailed composition not analysed) Espelie et al.
(1980), McDonaldet al. (1993)
Lemon n/r Aldehydes: C24,C26, C28
n/r C29, C31 n/r Baker et al. (1975)
Mandarin n/r Alkanes > esters > carbonyl compounds > primary alcohols > triterpenes(detailed composition not analysed)
Sala (2000)
Orange n/r Aldehydes: C24,C26, C28
Primary: C25 C29, C31 Amyrins Baker et al. (1975),Cajuste et al.(2010), Liu et al.(2012)
Grape n/r Aldehydes: C28, C30 Primary: C24, C26,C28
C25, C27, C29, C31 Oleanoic acid Radler and Horn(1965), Radler(1965, 1970),Comménil et al.(1997); Casado andHeredia (1999)
Other berriesBilberry 9,10,18-triOH C18
9,10-epoxy-18-OHC18
n/r n/r n/r n/r Kallio et al. (2006)
Black chokeberry 18-OH-9,10-epoxyC18:1
18-OH-9,10-epoxyC18:0
9(10),16-diOH C16:0
n/r n/r n/r n/r Järvinen et al.(2010)
Black currant 10(9,8),16-diOHC16
n/r n/r n/r n/r Kallio et al. (2006)
Cloudberry 9(10),16-diOH C16:0 n/r n/r n/r n/r Järvinen et al.(2010)
Cranberry 9,10-epoxy-18-OHC18
n/r n/r n/r n/r Kallio et al. (2006)
Crowberry 9(10),16-diOH C16:0
18-OH-9,10-epoxyC18:0
18-OH C18:1
n/r n/r n/r n/r Järvinen et al.(2010)
Lingonberry 9,10-epoxy-18-OHC18
9,10,18-triOH C18
n/r n/r n/r n/r Kallio et al. (2006)
Raspberry 9(10),16-diOH C16:0 n/r n/r n/r n/r Järvinen et al.(2010)
Rosehip 9(10),16-diOH C16:0 n/r n/r n/r n/r Järvinen et al.(2010)
Rowanberry 9(10),16-diOH C16:0 n/r n/r n/r n/r Järvinen et al.(2010)
Sea buckthorn 9,10-epoxy-18-OHC18
n/r n/r n/r n/r Kallio et al. (2006)
Strawberry 9(10),16-diOH C16:0 n/r n/r n/r n/r Järvinen et al.(2010)
Pepper 9,16-diOH C16
9-epoxy-18-OH C18
Fatty acids: C24,C26, C32
(major chainlength class)
n/r C31 Amyrins Kissinger et al.(2005), Parsonset al. (2012)
Persimmon 578–1378 �g/cm2
(detailedcomposition notanalysed)
337–770 �g/cm2 (detailed composition not analysed) Tsubaki et al.(2012)
Stone fruitsPeach (melting) 18-OH C18:1 Fatty acids: C18:2 Secondary:
C29-10-olC29 Ursolic and
oleanoic acidsBelge et al.(unpublished)
Peach (non-melting) 18-OH C18:1 Fatty acids: C18:2 Secondary:C29-10-ol
C29 Ursolic andoleanoic acids
Fernández et al.(2011), Belge et al.(unpublished)

106 I. Lara et al. / Postharvest Biology and Technology 87 (2014) 103–112
Table 1 (Continued)
Fruitspecies
Cutin Waxes References
Main monomertype
Carbonylcompounds
Alcohols Alkanes Terpenes
Sweet cherry 9(10),16-diOH C16:0
9,10,18-triOH C18:0
(cultivar-specific)
Fatty acids: C18:2 Primary: C23, C30
Secondary:C29-10-ol
C29, C27 Ursolic andoleanoic acids
Peschel et al.(2007); Belge et al.(unpublished)
Tomato 10,16-diOH C16:0 Fatty acids: C30, C32 Primary: C32 C31 (C29, C33) Amyrins Vogg et al. (2004),Saladié et al.(2007), Hovav et al.(2007), Leide et al.(2007), Mintz-Oronet al. (2008),Isaacson et al.(2009), Kosma et al.(2010), Nadakudutiet al. (2012), Yeatset al. (2012a), Shiet al. (2013)
C22-C3
specifi
pa(tfba
4
cpfMaaSit‘ccectc1f
iancftoosIaiui
Wild tomato Mid-chainepoxy-triOH C18
(cultivar-specific)
Fatty acids: C16-C28
(cultivar-specific)Primary:
(cultivar-
atterning (Shi et al., 2013). Little information is currently avail-ble for fruit other than tomato, but recent studies on sweet cherryAlkio et al., 2012) and apple (Albert et al., 2013) have also allowedhe identification of a range of genes potentially involved in cuticleormation during fruit development, including some related to theiosynthesis of wax and cutin components, transcription factorsnd cuticular lipid transporters.
. Changes in fruit cuticle composition after harvest
Limited research attention has been focused on changes in fruituticle composition during storage or in response to particularostharvest procedures, but some published studies are availableor apple, which reveal considerable cultivar-associated variability.
orice and Shortland (1973) did not observe any changes in waxmount or composition of ‘Sturmer’ apple fruit throughout storaget 3 ◦C for up to nine months, while total wax amounts in ‘Grannymith’ and ‘Dougherty’ samples increased to different extents dur-ng that time. Changes in fatty acid composition were also observed,he most notable being an 11-fold rise in linolenic acid found forGranny Smith’ fruit. Controlled atmosphere (CA) storage and, espe-ially, shelf-life thereafter, affected wax properties, structure andhemical composition in fruit of three apple cultivars (Veraverbeket al., 2001a), particularly for extended storage periods. The mainhanges in wax components referred to the alkane and ester frac-ions, apparently due to ester hydrolysis which led to augmentedontents of free fatty acids and in the secondary alcohol nonacosan-0-ol. The presence of an alcohol instead of an alkane can accountor wax glossiness, which is correlated with alcohol content.
Total wax amount decreased sharply in ‘Red Fuji’ apples dur-ng storage at 0 ◦C for seven months, and wax composition waslso modified (Dong et al., 2012). The alkanes n-nonacosane,-heptacosane and n-nonacosene, prominent among the hydro-arbons present in surface waxes of this cultivar and requiredor the formation of crystalline wax structures, decreased overhe storage period. In contrast, the content of nonacosan-10-l, essential for the development of cuticular wax crystals, andf nonacosan-10-one, decreased over the first months of coldtorage followed by a sharp increase for longer storage periods.nterestingly, 1-methylcyclopropene (1-MCP) treatment largely
ttenuated both the decline in n-nonacosane and the increasen nonacosan-10-ol and nonacosan-10-one levels observed forntreated fruit, indicating that wax composition of ‘Red Fuji’ appless an ethylene-dependent attribute. 1-MCP treatment also delayed
0
c)C29, C31
(cultivar-specific)Amyrins Leide et al. (2007),
Yeats et al. (2012a)
the development of certain wax constituents during cold storage forup to six months of ‘Autumn Gold’ and ‘Royal Gala’ fruit, althoughonly a few of them were unequivocally identified (Curry, 2008),among them nonacosan-10-ol, the increase in which after long-term storage was partially suppressed in 1-MCP-treated fruit. Theseobservations agree with recent work on ‘Navelate’ orange fruit,where ethylene treatment (2 �L/L) increased both total and softepicuticular waxes and induced structural changes in surface waxesduring shelf-life at 22 ◦C for up to three weeks (Cajuste et al., 2010).
5. The cuticle as a determinant of postharvest qualityattributes of fruits
Fruit cuticles influence postharvest shelf-life, and are associatedwith some important quality disorders such as cracking (Edelmannet al., 2005; Matas et al., 2005). Published research on the rela-tionship between cuticle properties or composition and qualityattributes of fruit commodities has reported on three main aspects,namely water permeability and loss, susceptibility to infections,and physiological disorders. Of late, though, some new evidencesuggests a possible important role of cuticle composition and/orproperties on postharvest firmness changes, which is worthy offurther consideration.
Given the hydrophobic nature of cuticle components and itscapacity to function as a barrier, it has generally been assumedthat this thin, outer layer of aerial organs of plants serves mainlyprotective purposes. For instance, intracuticular waxes have adominant role in minimising non-stomatal water loss across thecuticle. Cutin is involved in waterproofing leaves and fruit of higherplants, in regulating nutrient flow, and in minimising pathogenattack (Heredia, 2003). Some fungi infecting important cropsproduce cutinases, which aid infection through their esterase activ-ity (Sweigard et al., 1992). Plants with altered cutin depositionshow a wide range of phenotypes, including defective develop-ment and growth, altered morphology, permeability and stomatastructure, as well as increased sensitivity to stress and infections(Pollard et al., 2008). Evidence also indicates that cuticle struc-ture and links among its components must be a central aspectof cuticular functionality as well. For example, Li et al. (2007)overexpressed simultaneously GPAT5 and CYP86A1 in Arabidopsis,
respectively a glycerol-3-phosphate acyltransferase and a fatty acid�-hydroxylase required for cutin assembly, and observed increasedpermeability of cuticles in spite of increased cutin and unchangedwax accumulation.
y and
rsmcbmiafa1ccbcua(
5
dgasiacncMe
ofia(c1gottarwrftwb
gicdtmmucm‘
I. Lara et al. / Postharvest Biolog
If we focus specifically on fruit, cuticle properties are veryelevant from a physiological point of view, and often have con-iderable economic importance. Apart from its general role ininimising water loss (Riederer and Schreiber, 2001), the cuti-
le limits solute loss from the inner tissues and protects againstiotic and abiotic factors in the surrounding environment. Yet theechanical properties of fruit cuticles change with relative humid-
ty and temperature (Edelmann et al., 2005; Matas et al., 2005),nd this observation is relevant for postharvest performance ofruit. Riederer and Schreiber (2001) found that cuticular perme-bility to water increases by a factor of 2 in the temperature range5 to 35 ◦C and relates directly to solute transport through theuticle, in agreement with the general observation that heatinghemical solutions in many instances improves their absorptiony fresh fruit. Cuticular permeability is also impacted by organicompounds of both biogenic and anthropogenic (for instance, thosesed in pesticides) origin, which may increase cuticular perme-bility apparently through a plasticising action on wax structurereviewed in Riederer and Schreiber, 2001).
.1. Water loss
Postharvest water loss of fruit results from fruit respiration andiffusion through the fruit cuticle. Although the water potentialradient from inside to outside the fruit varies throughout stor-ge according to temperature and relative humidity, considerablepecies- and cultivar-related variability exists in water loss, whichndicates differences in cuticular permeability to water. This vari-bility arises from a combination of fruit maturity, environmentalonditions, and genetic differences in factors such as cuticle thick-ess, presence or pores or cracks, and quantity, distribution orhemical nature of cuticular components (Lownds et al., 1994;aguire et al., 1999; Knoche et al., 2000; Peschel et al., 2003; Gibert
t al., 2010).Contradictory results have been reported as to the relevance
f cuticle thickness for weight loss during storage. Significant dif-erences in wax deposition and cuticle thickness were observedn ozone-treated ‘Casselman’ plums as compared to control fruit,nd suggested to have contributed to differences in weight lossCrisosto et al., 1993a). Pepper fruit with high epicuticular waxontents also lost weight at significantly lower rates (Lownds et al.,993). In other cases, the positive effects on weight loss reduction ofiven postharvest treatments have been shown to be independentf cuticle thickness. For example, decreased weight loss in heat-reated cactus pears was suggested to arise from rearrangement ofhe epicuticular wax layers, which minimised cracks and fissures,s no treatment effects were found on cuticle thickness or respi-ation rates of fruit (López-Castaneda et al., 2010). Indeed, whenater permeability values of cuticles from 61 plant species were
eviewed in a survey which included mostly leaf, but also a fewruit cuticles (Riederer and Schreiber, 2001), it was concluded thathis property was not correlated to thickness or wax coverage, mostater apparently diffusing as single molecules across a lipophilic
arrier.The thickness of the cuticular wax layer is usually estimated
ravimetrically by extracting in organic solvent, drying and weigh-ng the wax, and then relating it to the surface units of skin oruticular membrane used for the extraction. However, this proce-ure implies tissue destruction and damage to the wax layer, andhus does not allow for assessing changes in cuticle structure, which
ay prove more relevant than thickness or amount for water per-eability properties of cuticle. Instead, Veraverbeke et al. (2001b)
sed confocal laser scanning microscopy (CLSM) to visualise thehanges in cuticle structure and thickness during long-term (nineonths) ultra-low oxygen (ULO) storage of ‘Elstar’, ‘Jonagold’ and
Jonagored’ apple fruit. This approach allowed the use of the same
Technology 87 (2014) 103–112 107
fruit throughout the experimental time, and showed profound dif-ferences in the wax layer according to cultivar and storage period,likely related to water loss rates of each particular cultivar. Duringstorage of all three cultivars, the wax layer smoothed to cover thecracks, possibly as a protection against moisture loss, and the waxpenetrated into the cutin layer. Further research on the same applecultivars demonstrated that the diffusion coefficient of wax was atleast a hundred times smaller than that of cutin (Veraverbeke et al.,2003a), and revealed considerable cultivar-related differences indiffusion properties of the cuticle. Based on these investigations,a model for the prediction of moisture loss across the cuticle ofapple during storage was developed (Veraverbeke et al., 2003b)and partially validated (Veraverbeke et al., 2003c).
In addition to the significance of cuticle thickness in weight lossas discussed above, experimental evidence also exists on the roleof particular cuticle components in cuticular permeability, waxeshaving been generally shown as the major cuticular componentsaccounting for its function as an impermeability barrier. Removalof surface waxes of ‘Calrico’ peaches resulted in significantly higherwater loss rates (Fernández et al., 2011). The peel of the glossy stickypeel tomato mutant is more elastic and shows increased cuticu-lar permeability than the wild type, apparently associated witha lower proportion of long-chain alkanes in epicuticular waxes(Nadakuduti et al., 2012). Similarly, biochemical characterisation ofthe cuticular waxes of the LeCER6 loss-of-function tomato mutantshowed decreased content in very long-chain n-alkanes along withelevated levels of amyrins, with an associated 3- to 8-fold increasein water loss per fruit surface area in comparison with the wild-typefruit, in spite of the fact that the mutant has thicker cuticles (Vogget al., 2004; Leide et al., 2007). Because the difference in water lossbetween the mutant and the wild type was abolished by chloro-form extraction of total waxes, but not when only the epicuticularwaxes were removed (Vogg et al., 2004), it was concluded that themain portion of the transpiration barrier is located in the intracutic-ular wax layer, whose core properties are defined by the n-alkanesbut modulated by the presence of triterpenoids. The physiologi-cal and biochemical characterisation of three cutin deficient tomatomutants (cd1, cd2 and cd3) has provided interesting clues as to thepossible role of each type of cuticle component on cuticle function(Isaacson et al., 2009). For instance, despite severely impaired cutindeposition in all three mutant lines, only cd1 fruit show dramati-cally increased water loss during the postharvest period, suggestingthat the cutin matrix does not play a central role in limiting waterloss and that waxes are the primary barrier against dehydration.Interestingly, the total amount of waxes in cuticles of wild typeand cd fruit is similar, but wax composition differs in each cd line.It was concluded that reduced wax alkanes and enhanced triter-penoids would lead to an increase in the amorphous portion of thecuticular waxes, thus impairing cuticle function as a barrier againstwater loss. This model questions a role for cutin as a significanthydrophobic barrier, and rather attributes it the function of pro-viding a framework into which the intracuticular wax compoundscan adopt a functional structure for restricting water movement.
Total cuticular wax amounts were likewise directly related topostharvest water loss during storage of pepper fruit from 10 cul-tivars, while no association was found to total cuticle amount,total cutin or to the amount of individual cutin or wax con-stituents (Kissinger et al., 2005). In contrast, Parsons et al. (2012)reported significant correlations of water loss in Capsicum sp.fruit not only with wax, but also with cutin monomer compo-sition. Simple straight-chain aliphatic cuticle components weremore closely related to the formation of impermeable barriers
than isoprenoid compounds, the ratio of alkanes to triterpenoidsplus sterols being negatively correlated with dehydration in accor-dance with previous observations for tomato (Isaacson et al.,2009). Moreover, water loss rates were associated to total cutin
1 y and
cTeatfg
tHginwmnairma
5
scfafiittvrsmaoapctPsci
n2bam2oattcttppFi
08 I. Lara et al. / Postharvest Biolog
ontent, to total C16 monomers, and to 16-hydroxypalmitic acid.he analytical procedures in both cases were similar, involvingnzymatic isolation of cuticles, chloroform/methanol dewaxingnd N,O,-bis(trimethylsilyl)trifluoroacetamide (BSTFA) derivatisa-ion of cutin monomers. These discrepancies may thus have arisenrom the different Capsicum genotypes used in each case, and sug-est relevant cultivar-related differences in cuticle functionality.
However, cuticle properties and their effect on fruit characteris-ics may be only partially related to cuticular chemical components.ovav et al. (2007) developed a novel tomato genotype via intro-ression from the wild tomato species Solanum habrochaites, whichs prone to cuticle microfissuring and fruit dehydration. This phe-otype was demonstrated to be inherited as a single gene (cuticularater permeability; Cwp1) which is expressed mainly in the epider-al tissue and codes for a protein of unidentified function. Because
o significant differences were observed between the wild-typend the introgressed genotypes in either wax or cutin components,t was hypothesised that the CWP1 protein is likely to affect theheological properties of the skin by a yet unclear mechanism, oray impact on other components of epidermal development such
s cell division or expansion.
.2. Susceptibility to infections
Disease resistance of fruit declines with ripeness stage and withtorage period. In some cases, hyphae of the pathogen organisman infect the fruit by direct penetration of the cuticle, sometimesorming highly specialised structures. For example, Colletotrichumcutatum penetrates the cuticle layer of Capsicum spp. fruit byorming a highly branched, thick-walled hypha within 24 h post-noculation (Liao et al., 2012). This structure has only been foundn the cuticle layer of Capsicum fruit, and it was not observed whenhe pathogen was inoculated onto pepper petals, mango leaves, oromato and eggplant fruit. Although many other common posthar-est pathogens are unable to penetrate directly the host cuticle andequire surface injuries or natural openings such as lenticels andtomata (Coates and Johnson, 1997), fruit become progressivelyore susceptible to wounds and mechanical damages in general
fter long-term storage, thus opening a route for the developmentf infection. Some of these injuries can be microscopic such as thoserising from surface microcracking to which some fruit species arearticularly prone, and which is directly dependent on the amount,omposition and mechanical properties of the cuticle. The pro-ective effect of ethylene treatment against disease incidence byenicilium digitatum in ‘Navelate’ oranges has been ascribed to theynthesis of new waxes in treated fruit, which may cover stomata,racks or areas lacking wax that are generated during storage, thusmposing a physical barrier to infection (Cajuste et al., 2010).
Relationships between infection susceptibility and cuticle thick-ess and structure have been suggested for apple fruit (Konarska,012). Lower incidence of rots and longer postharvest life in cran-erries has also been attributed to a thicker cuticle and waxccumulation at the calyx end, which would retard the entry oficroorganisms into the fruit during wet harvest (Özgen et al.,
002), a harvesting practice in which the fields are flooded inrder to facilitate gathering of fruit. Similarly, cuticle thicknessnd wax contents correlated positively with resistance to Botry-is cinerea in up to 42 genetically diverse cultivars and selections ofable grapes (Marois et al., 1986; Mlikota Gabler et al., 2003). Cuti-le density and thickness have been shown likewise to be relatedo stone fruit susceptibility to infection by Monilia fructicola, yethis could only partially explain the differences in fruit predis-
osition to this rot (Crisosto et al., 1997), suggesting a role forarticular cuticular components in modulating disease resistance.or instance, when ‘Thomson Seedless’ grape berries were dippedn chloroform to remove epicuticular waxes, a significant increaseTechnology 87 (2014) 103–112
in susceptibility to infection by B. cinerea was observed (Maroiset al., 1985). In contrast, the cutin content of the cuticle, rather thanwaxes, and cuticle thickness, were suggested as indicators of sus-ceptibility to B. cinerea infection in ‘Pinot noir’ grapes (Comménilet al., 1997). A major role for cutin in determining susceptibilityto infections is also supported by observations that epicuticularwax content and appearance were unchanged in grapes of fourseedless cultivars after dipping in hot ethanol, despite the factthat dipped fruit were more susceptible to subsequent infection(Mlikota Gabler et al., 2005). No apparent relationship was foundeither between the severity of the sooty blotch disease caused byPeltaster fructicola and Leptodontidium elatius and the major epicu-ticular wax components of five different apple cultivars (Beldinget al., 2000).
The relative importance of particular cutin and wax compo-nents to resistance against infection is poorly understood, but someresearch efforts have been focused on this aspect. Bostock et al.(1999) studied the basis for the high resistance of ‘Bolinha’ peachto infection by the brown rot fungus M. fructicola, and found thatthe phenolic chlorogenic and caffeic acids, otherwise quantitativelyminor cuticular components, were present in high amounts on thesurface of the resistant fruit and suppressed cutinase activity lev-els in cultures of M. fructicola. A series of cinnamic and benzoic acidderivatives also suppressed the activity of cutinase, an enzyme thatpathogenic fungi can use to aid penetration of the cuticle. Like-wise, in vitro assays demonstrated inhibiting effects of n-alkanesand triterpenoids on spore germination and mycelial growth ofAlternaria alternata, a causal agent of Alternaria rot of pears, sug-gesting a major contribution to the antifungal properties of thecuticular waxes of Asian pear (Yin et al., 2011).
Recent efforts to characterise genes involved in fruit cuticledevelopment will allow identification or confirmation of key ele-ments for specific cuticle functions. For instance, all three cdtomato mutants (Isaacson et al., 2009) have as little as only 5–10%cutin as compared to the wild-type, with an associated increasein pathogen susceptibility. Similarly, silencing of the transcrip-tion factor SlSHN3 and the fatty acid �-hydroxylase SlCYP86A69in tomato led to increased predisposition to infection by thefungus Colletotrichum coccodes and postharvest water loss (Shiet al., 2013). Biochemical characterisation of cuticles of mutantfruit showed that higher susceptibility to infection was related tosharply decreased contents of specific cutin monomers, particularlyof the C16 class and, for the mature green stage, also of the phenoliccompounds benzoic acid and trans-coumaric acid.
5.3. Physiological disorders
The outer surface layers of fruit, including cuticles as well asepidermal and subepidermal tissues, constitute the skin and repre-sent the main structures conferring mechanical strength to fruit.The skin surrounds a mass of more deformable material; whenskin failure occurs, cracks develop (Matas et al., 2005; Becker andKnoche, 2012). This cuticular cracking appears mainly in ripe fruitand, when severe enough to be visible, it represents an importantproblem for many commodities, causing large economic losses dueto negative effects on fruit appearance, acceleration of water lossand creation of an entry for pathogens. Cracks may also help accel-erate oxidative processes, thus favouring browning alterations. Onthe other hand, microcracks on the surface can be an importantpathway for the uptake of minerals such as calcium. Transport ofexogenously applied Ca2+ through apple cuticles has been shown tooccur mainly through lenticels and cracks in the cuticle (Harker and
Ferguson, 1988), and epicuticular wax cracks become wider anddeeper as storage period increases, thus intensifying Ca2+ uptakeby fruit (Roy et al., 1999). Heat-treated ‘Golden Delicious’ applesabsorbed significantly less calcium than untreated fruit (Roy et al.,
y and
1ii
cbtseKqbstewitmttbistdo
sGchaccech1taBtcacu
imawt(
tptwpfwbiat
I. Lara et al. / Postharvest Biolog
994; Lurie et al., 1996), this effect being attributed to wax soften-ng leading to smoothing or elimination of the deep cracks presentn epicuticular waxes of non-heated controls.
Cracking typically occurs when maturation and harvest coin-ide with a period of high humidity, causing water movement fromranches and leaves to the fruit owing to a large difference inheir water potentials. The cracking disorder has been especiallytudied in sweet cherry fruit (Sekse, 1995, 1998, 2008; Knochet al., 2000, 2001, 2002; Knoche and Peschel, 2002; Peschel andnoche, 2005). Removal of epicuticular wax had no effect on the fre-uency of water-induced microcracks in seven different cultivars,ut hydration increased fracture strain while decreasing fracturetress and elasticity modulus (Knoche and Peschel, 2006). No rela-ionship was found either between cracking susceptibility and skinlasticity, cuticle thickness or the content of different minerals,ith the exception of calcium, the levels of which were higher
n the cracking-resistant cultivar ‘Sue’ (Lane et al., 2000). Whilehe physical properties of the epidermal and subepidermal cells
ight be relevant for cracking susceptibility, which would explainhe observed impact of calcium, the cessation of cuticle deposi-ion during early sweet cherry development has been reported toe associated with down-regulation of several genes putatively
nvolved in cuticle formation (Alkio et al., 2012). The increase intrain imposed on the surface as the fruit expands would lead tohe formation of microcracks. The identification of these candi-ate genes will allow cause/effect studies on cuticle compositionr properties and the development of this disorder.
Cuticle density has been related to the development of disordersuch as flesh browning during storage of apples (Jobling, 2002).as diffusion through fruit tissues can be limited under certainontrolled atmosphere conditions, resulting in too low O2 and tooigh CO2 levels, which cause browning damage in the cortex. Somepple varieties are particularly susceptible to russeting and lenti-el breakdown, originated when the cuticle surrounding lenticelsracks under conditions of rapid fruit growth and that are wors-ned during storage (Curry, 2003). In turn, the severity of thoseracks is related to the structure of surface waxes, and impacted byumidity conditions during the growing season (Faust and Shear,972). Cuticular free phenolic compounds are correlated nega-ively with the formation of conjugated trienes during storage ofpples, and thus with the development of superficial scald (Ju andramlage, 2000), possibly mediated by the antioxidant activity ofhese compounds. The skin spot disorder, an important physiologi-al alteration in ‘Elstar’ apple fruit after removal from controlledtmosphere storage, is associated with microcracks in the cuti-le, apparently arising from oxidative damage through those crackspon removal from storage (Grimm et al., 2012).
Other alterations are likewise related apparently to the loss ofntegrity of the cuticle structure and functionality. The develop-
ent of skin discolouration in stone fruit species such as peachnd nectarine has been ascribed to abrasion injuries in the cuticle,hich expose the epidermal cells to alterations in pH or con-
ent of metallic ions, thus inducing changes in anthocyanin colourCrisosto et al., 1993b).
In relation to citrus fruit, epicuticular wax morphology is relatedo water permeability of cuticles in ‘Fortune’ mandarins, higherermeability values coinciding with higher incidence of peel pit-ing (Vercher et al., 1994). Morphological changes in the surfaceaxes might also underlie the protective role of ethylene againsteel pitting in oranges, although no satisfactory relationship wasound without water stress or with compositional changes inaxes (Cajuste et al., 2010). No correlation was observed either
etween epicuticular wax content and susceptibility to rindstain-ng of orange fruit (Sala et al., 1992), suggesting that compositionallterations rather than total amount of waxes are relevant forhe development of the alteration. This view is supported by the
Technology 87 (2014) 103–112 109
finding that higher incidence of rindstaining in ‘Fortune’ mandarinsis associated with lower content of n-alkanes and esters, and withhigher proportion of ketones and fatty acids (Sala, 2000), possiblyin relation to the severe damage observed in the structure of theepicuticular wax layer and resulting increase in flavedo water per-meability and water loss. Finally, greater incidence of chilling injuryin grapefruit after cold storage may be mediated by differences inepicuticular wax morphology and composition, specifically by anincrease in the content of n-alkanes (McDonald et al., 1993).
5.4. A role in postharvest firmness changes?
Mechanical factors, chiefly tissue firmness, have the most influ-ence on fruit texture. Firmness changes of fruit during ripening andafter harvest have been the object of intensive research efforts formore than 40 years. The primary focus of research on fruit softeninghas been placed on cell wall metabolism (reviewed in Goulao andOliveira, 2008) and has generally overlooked other possible phys-iological processes involved. Genetically suppressed expression ofseveral ripening-related cell wall-modifying proteins has generallyfailed to counteract firmness loss to a significant extent (Rose et al.,2003), leading to the view that ripening-related cell wall disassem-bly results from the cooperative action of many different activities.In this context, some reports suggest that cuticle composition andarchitecture may also play a key role in firmness changes of fruit.
The cuticle provides structural support for fruit lacking hardinternal tissue. It may function as an external structural elementthat adds mechanical support for tissue integrity. Cuticular waxcomponents of grape berries are tightly bound and highly orderedat the molecular level, and the resulting barrier structure protectsbut also imparts consistency to fruit (Casado and Heredia, 1999).The biomechanics of tomato fruit skin and isolated cuticles havebeen investigated for three cultivars differing in cracking suscep-tibility and fruit shape, and it was concluded that the cuticle is amechanically important component of the tomato fruit (Bargel andNeinhuis, 2005). Stiffness of skin and cuticle increased during mat-uration and ripening, and failure stress and failure strain decreased,although some cultivar-related discrepancies were observed. Themodifications in these properties at the final stages of ripeninghave implications for handling and storage potential. Mechanicalproperties of the cuticle are altered according to storage conditionssuch as temperature and relative humidity (Edelmann et al., 2005;Matas et al., 2005); this can also be of relevance for changes in firm-ness. Despite that, the structure-function relationships betweencuticle composition/structure and biomechanical properties havenot been intensively explored. Bargel et al. (2006) hypothesisedthat the amount of phenolic compounds is correlated with rigidityof the cutin matrix at full maturity. Accordingly, the mechanicalcharacterisation of cuticles isolated from mature green and redripe ‘Cascada’ tomato fruit revealed that phenolics present in thecutin network are likely candidates to account for rigidity, whereasthe polysaccharide components provide the elastic behaviour ofthe whole cuticle (López-Casado et al., 2007), which suggests arole in determining at least some fruit textural attributes. Theseattributes include firmness as well as other physical characteris-tics related to deformation or fracture. A survey of the chemicalcomposition, density and tensile properties of fruit cuticles in 27persimmon varieties revealed that density and polysaccharide con-stituents contribute to cuticle strength and elasticity, while cutinis related to viscoelasticity (Tsubaki et al., 2012), and these proper-ties were related to the mechanical characteristics specific to eachof the cultivars considered in the study.
Some studies have shown parallels between firmness loss andchanges in cell turgor, which were apparently under cellular con-trol and not simply a consequence of loss of membrane integrity(Shackel et al., 1991). Solutes accumulate in the apoplast during

1 y and
rcotbotaTmf
hmTtdot2maslSntcbs
6
tgdahqttnftpditipbeguFacowaToit
10 I. Lara et al. / Postharvest Biolog
ipening of tomato (Almeida and Huber, 1999; Shackel et al., 1991),oncomitant with transpiration-related water loss favouring effluxf water from the fruit. Moisture loss has been identified recently ashe major cause of firmness changes during postharvest storage oflueberry fruit (Paniagua et al., 2013), which otherwise displaysnly small changes in cell walls after harvest. A relationship ofurgor loss with mechanical properties has also been observed forpples and nectarines (Lin and Pitt, 1986; Heyes and Sealey, 1996).his points to the regulation of water transpiration as a possibleechanism involved in firmness loss, and thus also suggests a role
or the fruit cuticle in the process.Further insight on a possible role of cuticle in firmness changes
as been provided by the characterisation of the otherwise nor-ally ripening tomato mutant DFD (‘delayed fruit deterioration’).
hese fruit remain firm for at least six months and show high resis-ance to opportunistic pathogens although the degree of cell wallisassembly, the reduction in cell-to-cell adhesion and the ratesf gene expression related to cell wall degradation are similar tohose in the normally softening cultivar ‘Ailsa Craig’ (Saladié et al.,007). Detailed characterisation of this mutant demonstrated mini-al transpiration water loss and substantially enhanced cell turgor
rising from remarkable differences in fruit cuticle composition andtructure. These results suggest that turgor and transpiration wateross are important determinants in the softening process of tomato.ince cuticle properties help modulation of water loss, this exter-al structure may contribute significantly to fruit texture. However,his may be restricted to fruit displaying thick and well developeduticles, while the role of this outer layer of the fruit surface mighte considerably less in species characterised by weaker cuticlesuch as strawberries.
. Conclusions
Biological surrounding structures represent the first contact ofhe plant cells and organs with the environment in which theyrow, triggering signalling pathways that stand for the first line ofefense and adaptation against biotic and abiotic stresses. Bioticnd abiotic conditions keep modulating fruit development afterarvest, and are important determinants of storage potential anduality of produce with vital impacts on economic revenues. Whilehe importance of the cell wall is widely recognised and has beenhe centre of attention in studies on postharvest biology and tech-ology of fruit produce, the role of the cuticle, that surrounds the
ruit, has received much less attention. Major questions relatedo the interactions between fruit quality attributes and the com-osition and physiological roles of fruit cuticles, which demandetailed explanation, are illustrated in Fig. 1. From this review,
t can be concluded that much more emphasis should be giveno investigating fruit cuticles, under a multidisciplinary approachncluding developmental biology, fruit physiology, physics andostharvest technology. On the other hand, knowledge of theackground of each specific genotype is needed to understand gen-ral and specific postharvest behaviour. Even so, many knowledgeaps exist regarding the specific cuticle composition in individ-al fruit species and varieties (Table 1) that need to be addressed.rom the technological point of view, huge benefits may alsorise from a more comprehensive knowledge of the fruit cuticleomposition and physiological role. A more detailed knowledgef the fine structure and role of cuticle biology and biophysicsould be crucial to optimise tailor-made postharvest strategies
imed at improving quality maintenance and storage capability.
o the best of our knowledge, this review is the first focusingn fruit cuticles as major factors modulating postharvest qual-ty, and gathering and interlinking dispersed literature on thisopic.Technology 87 (2014) 103–112
Acknowledgments
B. Belge is the recipient of a FI-DGR grant from AGAUR (Gen-eralitat de Catalunya). Financial support through the AGL2010-14801/ALI project, granted by the Ministerio de Ciencia eInnovación (MICINN) of Spain, is gratefully acknowledged.
References
Albert, Z., Ivanics, B., Molnár, A., Miskó, A., Tóth, M., Papp, I., 2013. Candidate genes ofcuticle formation show characteristic expression in the fruit skin of apple. PlantGrowth Regul. 70, 71–78.
Alkio, M., Jonas, U., Sprink, T., van Nocker, S., Knoche, M., 2012. Identification ofputative candidate genes involved in cuticle formation in Prunus avium (sweetcherry) fruit. Ann. Bot. 110, 101–112.
Almeida, D.P.F., Huber, D.J., 1999. Apoplastic pH and inorganic ion levels in tomatofruit: a potential means for regulation of cell wall metabolism during ripening.Physiol. Plant. 105, 506–512.
Baker, E.A., Procopiou, J., Hunt, G.M., 1975. The cuticles of Citrus species: compositionof leaf and fruit waxes. J. Sci. Food Agric. 26, 1093–1101.
Bargel, H., Neinhuis, C., 2005. Tomato (Lycopersicon esculentum Mill.) fruit growthand ripening as related to the biomechanical properties of fruit skin and isolatedcuticle. J. Exp. Bot. 56, 1049–1060.
Bargel, H., Koch, K., Cerman, Z., Neinhuis, C., 2006. Structure-function relationshipsof the plant cuticle and cuticular waxes: a smart material? Funct. Plant Biol. 33,893–910.
Becker, T., Knoche, M., 2012. Deposition, strain, and microcracking of the cuticle inthe developing ‘Riesling’ grape berries. Vitis 51, 1–6.
Belding, R.D., Blankenship, S.M., Young, E., Leidy, R.B., 1998. Composition and vari-ability of epicuticular waxes in apple cultivars. J. Am. Soc. Hortic. Sci. 123,348–356.
Belding, R.D., Sutton, T.B., Blankenship, S.M., Young, E., 2000. Relationship betweenapple fruit epicuticular wax and growth of Peltaster fructicola and Leptodontidiumelatius, two fungi that cause sooty blotch disease. Plant Dis. 84, 767–772.
Bostock, R.M., Wilcox, S.M., Wang, G., Adaskaweg, J.E., 1999. Suppression of Moniliniafructicola cutinase production by peach fruit surface phenolic acids. Physiol. Mol.Plant Pathol. 54, 37–50.
Cajuste, J.F., González-Candelas, L., Veyrat, A., García-Breijo, F.J., Reig-Arminana, J.,Lafuente, M.T., 2010. Epicuticular wax content and morphology as related toethylene and storage performance of ‘Navelate’ orange fruit. Postharvest Biol.Technol. 55, 29–35.
Casado, C.G., Heredia, A., 1999. Structure and dynamics of reconstituted cuticularwaxes of grape berry cuticle (Vitis vinifera L.). J. Exp. Bot. 50, 175–182.
Coates, L.M., Johnson, G.I., 1997. Chapter 33: postharvest diseases of fruit and vegeta-bles. In: Brown, J., Ogle, H. (Eds.), Plant Pathogens and Plant Diseases. RockvalePublications, Cambridge, UK, pp. 533–547.
Comménil, P., Brunet, L., Audran, J.C., 1997. The development of the grape berry cuti-cle in relation to susceptibility to bunch rot disease. J. Exp. Bot. 48, 1599–1607.
Crisosto, C.H., Retzlaff, W.A., Williams, L.E., DeJong, T.M., Zoffoli, J.P., 1993a. Posthar-vest performance evaluation of plum (Prunus salicina Lindel., ‘Casselman’) fruitgrown under three ozone concentrations. J. Am. Soc. Hortic. Sci. 118, 497–502.
Crisosto, C.H., Johnson, R.S., Luza, J., Day, K., 1993b. Incidence of physical damage onpeach and nectarine skin discoloration development: anatomical studies. J. Am.Soc. Hortic. Sci. 118, 796–800.
Crisosto, C.H., Johnson, R.S., DeJong, T., Day, K.R., 1997. Orchard factors affectingpostharvest stone fruit quality. HortScience 32, 820–823.
Curry, E., 2003. Factors Associated with Apple Lenticel Breakdown.Washington State University, Postharvest Information Network,http://postharvest.tfrec.wsu.edu/REP2003B.pdf
Curry, E., 2008. Effects of 1-MCP applied postharvest on epicuticular wax of apples(Malus domestica Borkh.) during storage. J. Sci. Food Agric. 88, 996–1006.
Domínguez, E., Cuartero, J., Heredia, A., 2011a. An overview of plant cuticle biome-chanics. Plant Sci. 181, 77–84.
Domínguez, E., Heredia-Guerrero, J.A., Heredia, A., 2011b. The biophysical design ofplant cuticles: an overview. New Phytol. 189, 938–949.
Dong, X., Rao, J., Huber, D.J., Chang, X., Xin, F., 2012. Wax composition of ‘Red Fuji’apple fruit during development and during storage after 1-methylcyclopropanetreatment. Hortic. Environ. Biotechnol. 53, 288–297.
Edelmann, H.G., Neinhuis, C., Bargel, H., 2005. Influence of hydration and temper-ature on the rheological properties of plant cuticles and their impact on plantorgan integrity. J. Plant Growth Regul. 24, 116–126.
Espelie, K.E., Dean, B.B., Kolattukudy, P.E., 1979. Composition of lipid-derived poly-mers from different anatomical regions of several plant species. Plant Physiol.64, 1089–1093.
Espelie, K.E., Davis, R.W., Kolattukudy, P.E., 1980. Composition, ultrastructure andfunction of the cutin- and suberin-containing layers in the leaf, fruit peel,juice-sac and inner seed coat of grapefruit (Citrus paradisi Macfed.). Planta 149,
498–511.Faust, M., Shear, C.B., 1972. Fine structure of the fruit surface of three apple cultivars.J. Am. Soc. Hortic. Sci. 97, 351–355.
Fernández, V., Khayet, M., Montero-Prado, P., Heredia-Guerrero, J.A., Liakopoulos,G., Karabourniotis, G., del Río, V., Domínguez, E., Tacchini, I., Nerín, C., Val, J.,

y and
G
G
G
H
H
H
H
I
J
J
J
K
K
K
K
K
K
K
K
K
K
K
K
K
K
L
L
L
L
L
L
I. Lara et al. / Postharvest Biolog
Heredia, A., 2011. New insights into the properties of pubescent surfaces: peachfruit as a model. Plant Physiol. 156, 2098–2108.
ibert, C., Génard, M., Vercambre, G., Lescouret, F., 2010. Quantification and mod-elling of the stomatal, cuticular and crack components of peach fruit surfaceconductance. Funct. Plant Biol. 37, 264–274.
oulao, L.F., Oliveira, C.M., 2008. Cell wall modifications during fruit ripening: whena fruit is not the fruit. Trends Food Sci. Technol. 19, 4–25.
rimm, E., Khanal, B.P., Winkler, A., Knoche, M., Köpcke, D., 2012. Structural andphysiological changes associated with the skin spot disorder in apple. Posthar-vest Biol. Technol. 64, 111–118.
arker, F.R., Ferguson, I.B., 1988. Transport of calcium across cuticles isolated fromapple. Sci. Hortic. 36, 205–217.
eredia, A., 2003. Biophysical and biochemical properties of cutin, a plant barrierbiopolymer. Biochim. Biophys. Acta 1620, 1–7.
eyes, J.A., Sealey, D.F., 1996. Textural changes during nectarine (Prunus persica)development and ripening. Sci. Hortic. 65, 49–58.
ovav, R., Chehanovsky, N., Moy, M., Jetter, R., Schaffer, A.A., 2007. The identifica-tion of a gene (Cwp1) silenced during Solanum evolution, which causes cuticlemicrofissuring and dehydration when expressed in tomato fruit. Plant J. 52,627–639.
saacson, T., Kosma, D.K., Matas, A.J., Buda, G.J., He, Y., Yu, B., Pravitasari, A., Batteas,J.D., Stark, R.E., Jenks, M.A., Rose, J.K.C., 2009. Cutin deficiency in the tomato fruitcuticle consistently affects resistance to microbial infection and biomechanicalproperties, but not transpirational water loss. Plant J. 60, 363–377.
ärvinen, R., Kaimainen, M., Kallio, H., 2010. Cutin composition of selected northernberries and seeds. Food Chem. 122, 137–144.
obling, J., 2002. Understanding Flesh Browning in Pink Lady Apples. Sydney Posthar-vest Laboratory Information Sheet, www.postharvest.com.au
u, Z., Bramlage, W.J., 2000. Cuticular phenolics and scald development in ‘Delicious’apples. J. Am. Soc. Hortic. Sci. 125, 498–504.
allio, H., Nieminen, R., Tuomasjukka, S., Hakala, M., 2006. Cutin composition of fiveFinnish berries. J. Agric. Food Chem. 54, 457–462.
hanal, B.P., Grimm, E., Knoche, M., 2011. Fruit growth, cuticle deposition, wateruptake, and fruit cracking in jostaberry, gooseberry, and black currant. Sci. Hor-tic. 128, 289–296.
issinger, M., Tuvia-Alkalai, S., Shalom, Y., Fallik, E., Elkind, Y., Jenks, M.A., Goodwin,M.S., 2005. Characterization of physiological and biochemical factors associatedwith postharvest water loss in ripe pepper fruit during storage. J. Am. Soc. Hortic.Sci. 130, 735–741.
noche, M., Peschel, S., Hinz, M., Bukovac, M.J., 2000. Studies on water transportthrough the sweet cherry fruit surface: characterizing conductance of the cutic-ular membrane using pericarp segments. Planta 212, 127–135.
noche, M., Peschel, S., Hinz, M., Bukovac, M.J., 2001. Studies on water transportthrough the sweet cherry fruit surface: II. Conductance of the cuticle in relationto fruit development. Planta 213, 927–936.
noche, M., Peschel, S., Hinz, M., 2002. Studies on water transport through thesweet cherry fruit surface: III. Conductance of the cuticle in relation to fruitsize. Physiol. Plant. 114, 414–421.
noche, M., Peschel, S., 2002. Studies on water transport through the sweet cherryfruit surface: VI. Effect of hydrostatic pressure on water uptake. J. Hortic. Sci.Biotechnol. 77, 609–614.
noche, M., Beyer, M., Peschel, S., Oparlakov, B., Bukovac, M.J., 2004. Changes instrain and deposition of cuticle in developing sweet cherry fruit. Physiol. Plant.120, 667–677.
noche, M., Peschel, S., 2006. Water on the surface aggravates microscopic crackingof the sweet cherry fruit cuticle. J. Am. Soc. Hortic. Sci. 131, 192–200.
noche, M., Peschel, S., 2007. Deposition and strain of the cuticle of developingEuropean plum fruit. J. Am. Soc. Hortic. Sci. 132, 597–602.
onarska, A., 2012. Differences in the fruit peel structures between two apple culti-vars during storage. Acta Sci. Pol., Hortorum Cultus 11, 105–116.
osma, D.K., Parsons, E.P., Isaacson, T., Lü, S., Rose, J.K.C., Jenks, M.A., 2010. Fruit cuti-cle lipid composition during development in tomato ripening mutants. Physiol.Plant. 139, 107–117.
unst, L., Samuels, A.L., 2003. Biosynthesis and secretion of plant cuticular wax. Prog.Lipid Res. 42, 51–80.
unst, L., Samuels, A.L., 2009. Plant cuticles shine: advances in wax biosynthesis andexport. Curr. Opin. Plant Biol. 12, 721–727.
ane, W.D., Meheriuk, M., McKenzie, D.L., 2000. Fruit cracking of a susceptible,an intermediate, and a resistant sweet cherry cultivar. HortScience 35, 239–242.
eide, J., Hildebrandt, U., Reussing, K., Riederer, M., Vogg, G., 2007. The develop-mental pattern of tomato fruit wax accumulation and its impact on cuticulartranspiration barrier properties: effects of a deficiency in a �-ketoacyl-coenzyme A synthase (LeCER6). Plant Physiol. 144, 1667–1679.
i, Y., Beisson, F., Koo, A.J.K., Molina, I., Pollanrd, M., Ohlrogge, J., 2007. Identificationof acyltransferases required for cutin biosynthesis and production of cutin withsuberin-like monomers. PNAS 104, 18339–18344.
iao, C.Y., Chen, M.Y., Chen, Y.K., Kuo, K.C., Chung, K.R., Lee, M.H., 2012. Formationof highly branched hyphae by Colletotrichum acutatum within the fruit cuticlesof Capsicum spp. Plant Pathol. 61, 262–270.
in, T.T., Pitt, R.E., 1986. Rheology of apple and potato tissue as affected by cell turgor
pressure. J. Texture Stud. 17, 291–313.iu, D.C., Zeng, Q., Ji, Q.X., Liu, C.F., Liu, S.B., Liu, Y., 2012. A comparison of the ultra-structure and composition of fruits’ cuticular wax from the wild-type ‘Newhall’navel orange (Citrus sinensis [L.] Osbeck cv. Newhall) and its glossy mutant. PlantCell Rep. 31, 2239–2246.
Technology 87 (2014) 103–112 111
López-Casado, G., Matas, A.J., Domínguez, E., Cuartero, J., Heredia, A., 2007. Biome-chanics of isolated tomato (Solanum lycopersicum L.) fruit cuticles: the role ofthe cutin matrix and polysaccharides. J. Exp. Bot. 58, 3875–3883.
López-Castaneda, J., Corrales-García, J., Terrazas-Salgado, T., Colinas-León, T., 2010.Effect of saturated air heat treatments on weight loss reduction and epicuticularchanges in six varieties of cactus pear fruit. J. PACD 12, 37–47.
Lownds, N.K., Banaras, M., Bosland, P.W., 1993. Relationships between posthar-vest water loss and physical properties of pepper fruit (Capsicum annuum L.).HortScience 28, 1182–1184.
Lownds, N.K., Banaras, M., Bosland, P.W., 1994. Postharvest water loss and storagequality of nine pepper (Capsicum) cultivars. HortScience 29, 191–193.
Lurie, S., Fallik, E., Klein, J.D., 1996. The effect of heat treatment on apple epicuticularwax and calcium uptake. Postharvest Biol. Technol. 8, 271–277.
Marga, F., Pesacreta, T.C., Hasenstein, K.H., 2001. Biochemical analysis of elastic andrigid cuticles of Cirsium horridulum. Planta 213, 841–848.
Maguire, K.M., Lang, A., Banks, N.H., Hall, A., Hopcroft, D., Bennett, R., 1999. Rela-tionship between water vapour permeance of apples and micro-cracking of thecuticle. Postharvest Biol. Technol. 17, 89–96.
Marois, J.J., Bledsoe, A.M., Gubler, W., 1985. Effects of surfactants on epicuticu-lar wax and infection of grape berries by Botrytis cinerea. Phytopathology 75,1329.
Marois, J.J., Nelson, J.K., Morrison, J.C., Lile, L.S., Bledsoe, A.M., 1986. The influence ofberry contact within grape clusters on the development of Botrytis cinerea andepicuticular wax. Am. J. Enol. Vitic. 37, 293–296.
Matas, A.J., López-Casado, G., Cuartero, J., Heredia, A., 2005. Relative humidity andtemperature modify the mechanical properties of isolated tomato fruit cuticles.Am. J. Bot. 92, 462–468.
McDonald, R.E., Nordby, H.E., McCollum, T.G., 1993. Epicuticular wax morphol-ogy and composition are related to grapefruit chilling injury. HortScience 28,311–312.
Mintz-Oron, S., Mandel, T., Rogachev, I., Feldberg, L., Lotan, O., Yativ, M., Wang, Z.,Jetter, R., Venger, I., Adato, A., Aharoni, A., 2008. Gene expression and metabolismin tomato fruit surface tissues. Plant Physiol. 147, 823–851.
Mlikota Gabler, F., Smilanick, J.L., Mansour, M., Ramming, D.W., Mackey, B.E.,2003. Correlations of morphological, anatomical, and chemical featuresof grape berries with resistance to Botrytis cinerea. Phytopathology 93,1263–1273.
Mlikota Gabler, F., Ghosoph, J.M., Margosan, D.A., 2005. Impact of postharvest hotwater or ethanol treatment of table grapes on gray mold incidence, quality, andethanol content. Plant Dis. 89, 309–316.
Morice, I.M., Shortland, F.B., 1973. Composition of the surface waxes of apple fruitsand changes during storage. J. Sci. Food Agric. 24, 1331–1339.
Nadakuduti, S.S., Pollard, M., Kosma, D.K., Allen, C., Ohlrogge, J.B., Barry, C.S., 2012.Pleoiotropic phenotypes of the sticky peel mutant provide new insight into therole of CUTIN DEFICIENT2 in epidermal cell function in tomato. Plant Physiol.159, 945–960.
Nawrath, C., Poirier, Y., 2008. Pathways for the synthesis of polyesters in plants:cutin, suberin, and polyhydroxyalkanoates. In: Bioengineering and MolecularBiology of Plant Pathways, In: Bohnert, H.J., Nguyen, H., Lewis, N.G. (Eds.),Advances in Plant Biochemistry and Molecular Biology, 1. Elsevier, L&D Amster-dam.
Özgen, M., Palta, J.P., Smith, J.D., 2002. Ripeness stage at harvest influencespostharvest life of cranberry fruit: physiological and anatomical explanations.Postharvest Biol. Technol. 24, 291–299.
Paniagua, A.C., East, A.R., Hindmarsh, J.P., Heyes, J.A., 2013. Moisture loss is the majorcause of firmness change during postharvest storage of blueberry. PostharvestBiol. Technol. 79, 13–19.
Parsons, E.P., Popopvsky, S., Lohrey, G.T., Lü, S., Alkalai-Tuvia, S., Perzelan, Y., Paran, I.,Fallik, E., Jenks, M.A., 2012. Fruit cuticle lipid composition and fruit post-harvestwater loss in an advanced backcross generation of pepper (Capsicum sp.). Physiol.Plant. 146, 15–25.
Peschel, S., Beyer, M., Knoche, M., 2003. Surface characteristics of sweet cherry fruit:stomata number, distribution, functionality and surface wetting. Sci. Hortic. 97,265–278.
Peschel, S., Knoche, M., 2005. Characterization of microcracks in the cuticle of devel-oping sweet cherry fruit. J. Am. Soc. Hortic. Sci. 130, 487–495.
Peschel, S., Franke, R., Schreiber, L., Knoche, M., 2007. Composition of the cuticle ofdeveloping sweet cherry fruit. Phytochemistry 68, 1017–1025.
Pollard, M., Beisson, F., Li, Y., Ohlrogge, J.B., 2008. Building lipid barriers: biosynthesisof cutin and suberin. Trends Plant Sci. 13, 236–246.
Radler, F., 1965. The surface waxes of the sultana vine (Vitis vinifera cv. ThompsonSeedless). Aust. J. Biol. Sci. 18, 1045–1056.
Radler, F., 1970. Investigations on the cuticular wax of Vitis vinifera ssp. sylvestrisand Vitis vinifera ssp. vinifera. Angew. Bot. 44, 187–195.
Radler, F., Horn, D.H.S., 1965. The composition of grape cuticle wax. Aust. J. Chem.18, 1059–1069.
Riederer, M., Schreiber, L., 2001. Protecting against water loss: analysis of the barrierproperties of plant cuticles. J. Exp. Bot. 52, 2023–2032.
Rose, J.K.C., Català, C., González-Carranza, C.Z.H., Roberts, J.A., 2003. Plant cell walldissasembly. In: Rose, J.K.C. (Ed.), The Plant Cell Wall, 8. Blackwell PublishingLtd, Oxford, UK, pp. 264–324.
Rosenquist, J.K., Morrison, J.C., 1988. The development of the cuticle and epicuticularwax of the grape berry. Vitis 27, 63–70.
Roy, S., Conway, W.S., Watada, A.E., Sams, C.E., Erbe, E.F., Wergin, W.P., 1994. Heattreatment affects epicuticular wax structure and postharvest calcium uptake in‘Golden Delicious’ apples. HortScience 29, 1056–1058.

1 y and
R
S
S
S
S
S
S
S
S
S
S
T
12 I. Lara et al. / Postharvest Biolog
oy, S., Conway, W.S., Watada, A.E., Sams, C.E., Erbe, E.F., Wergin, W.P., 1999. Changesin the ultrastructure of the epiculticular wax and postharvest calcium uptake byfruit. HortScience 34, 121–124.
ala, J.M., 2000. Content, chemical composition and morphology of epicuticularwax of Fortune mandarin fruits in relation to peel pitting. J. Sci. Food Agric.80, 1887–1894.
ala, J.M., Lafuente, T., Cunat, P., 1992. Content and chemical composition of epi-cuticular wax of ‘Navelina’ oranges and ‘Satsuma’ mandarins as related torindstaining of fruit. J. Sci. Food Agric. 59, 489–495.
aladié, M., Matas, A.J., Isaacson, T., Jenks, M.A., Goodwin, S.M., Niklas, K.J., Xiaolin, R.,Labavitch, J.M., Shackel, K.A., Fernie, A.R., Lytovchenko, A., O’Neill, M.A., Watkins,C.B., Rose, J.K.C., 2007. A reevaluation of the key factors that influence tomatofruit softening and integrity. Plant Physiol. 144, 1012–1028.
amuels, L., Kunst, L., Jetter, R., 2008. Sealing plant surfaces: cuticular wax formationby epidermal cells. Annu. Rev. Plant Biol. 59, 683–707.
ekse, L., 1995. Fruit cracking in sweet cherries (Prunus avium L.,). Some physiologicalaspects—a mini review. Sci. Hortic. 63, 135–141.
ekse, L., 1998. Fruit cracking mechanisms in sweet cherries (Prunus avium L.,)—areview. Acta Hortic. 468, 637–648.
ekse, L., 2008. Fruit cracking in sweet cherries—some recent advances. Acta Hortic.795, 615–624.
hackel, K.A., Greve, C., Labavitch, J.M., Ahmadi, H., 1991. Cell turgor changesassociated with ripening in tomato pericarp tissue. Plant Physiol. 97,814–816.
hi, J.X., Adato, A., Alkan, N., He, Y., Lashbrooke, J., Matas, A.J., Meir, S., Malitsky, S.,Isaacson, T., Prusky, D., Leshkowitz, D., Schreiber, L., Granell, A.R., Widemann,E., Grausem, B., Pinot, F., Rose, J.K.C., Rogachev, I., Rothan, C., Aharoni, A., 2013.The tomato SlSHINE3 transcription factor regulates fruit cuticle formation andepidermal patterning. New Phytol. 197, 468–480.
weigard, J.A., Chumley, F.G., Valent, B., 1992. Cloning and analysis of CUT1,
a cutinase gene from Magnaporthe grisea. Mol. Gen. Genet. MGG 232,174–182.subaki, S., Ozaki, Y., Yonemori, K., Azuma, J., 2012. Mechanical properties of fruit-cuticular membranes isolated from 27 cultivars of Diospyros kaki Thunb. FoodChem. 132, 2135–2139.
Technology 87 (2014) 103–112
Veraverbeke, E.A., Lammertyn, J., Saevels, S., Nicolaï, B.M., 2001a. Changes in chem-ical wax composition of three different apple (Malus domestica Borkh.) cultivarsduring storage. Postharvest Biol. Technol. 23, 197–208.
Veraverbeke, E.A., Van Bruaene, N., Van Oostveldt, P., Nicolaï, B.M., 2001b. Nondestructive analysis of the wax layer of apple (Malus domestica Borkh.) by meansof confocal laser scanning microscopy. Planta 213, 525–533.
Veraverbeke, E.A., Verboven, P., Scheerlinck, N., Hoang, M.L., Nicolaï, B.M., 2003a.Determination of the diffusion coefficient of tissue, cuticle, cutin and wax ofapple. J. Food Eng. 58, 285–294.
Veraverbeke, E.A., Verboven, P., Van Oostveldt, P., Nicolaï, B.M., 2003b. Predic-tion of moisture loss across the cuticle of apple (Malus sylvestris subsp. mitis(Wallr.)) during storage Part 1. Model development and determination of diffu-sion coefficients. Postharvest Biol. Technol. 30, 75–88.
Veraverbeke, E.A., Verboven, P., Van Oostveldt, P., Nicolaï, B.M., 2003c. Prediction ofmoisture loss across the cuticle of apple (Malus sylvestris subsp. mitis (Wallr.))during storage: part 2. Model simulations and practical applications. PostharvestBiol. Technol. 30, 89–97.
Vercher, R., Tadeo, F.R., Almela, V., Zaragoza, S., Primo-Millo, E., Agustí, A., 1994. Rindstructure, epicuticular wax morphology and wáter permeability of ‘Fortune’mandarin fruits affected by peel pitting. Ann. Bot. 74, 619–625.
Vogg, G., Fischer, S., Leide, J., Emmanuel, E., Jetter, R., Levy, A.A., Riederer, M., 2004.Tomato fruit cuticular waxes and their effect on transpiration barrier properties:functional characterization of a mutant deficient in a very-long-chain fatty acid�-ketoacyl-CoA synthase. J. Exp. Bot. 55, 1404–1410.
Yeats, T.H., Buda, G.J., Wang, Z., Chehanovsky, N., Moyle, L.C., Jetter, R., Schaffer, A.A.,Rose, J.K.C., 2012a. The fruit cuticles of wild tomato species exhibit architecturaland chemical diversity, providing a new model for studying the evolution ofcuticle function. Plant J. 69, 655–666.
Yeats, T.H., Martin, L.B.B., Viart, H.M.F., Isaacson, T., He, Y., Zhao, L., Matas, A.J., Buda,G.J., Domozych, D.S., Clausen, M.H., Rose, J.K.C., 2012b. The identification of
cutin synthase: formation of the plant polyester cutin. Nat. Chem. Biol. 8, 609–611.Yin, Y., Bi, Y., Chen, S., Li, Y., Wang, Y., Ge, Y., Ding, B., Li, Y., Zhang, Z., 2011. Chemicalcomposition and antifungal activity of cuticular wax isolated from Asian pearfruit (cv Pingguoli). Sci. Hortic. 129, 577–582.