alginate oligosaccharide postharvest treatment preserve
TRANSCRIPT

Accepted Manuscript
Alginate Oligosaccharide Postharvest Treatment Preserve Fruit Quality and In-crease Storage Life Via Abscisic Acid Signaling in Strawberry
Santosh Kumar Bose, Prianka Howlader, Jia Xiaochen, Wang Wenxia, YinHeng
PII: S0308-8146(19)30128-1DOI: https://doi.org/10.1016/j.foodchem.2019.01.060Reference: FOCH 24155
To appear in: Food Chemistry
Received Date: 26 November 2018Revised Date: 3 January 2019Accepted Date: 8 January 2019
Please cite this article as: Kumar Bose, S., Howlader, P., Xiaochen, J., Wenxia, W., Heng, Y., AlginateOligosaccharide Postharvest Treatment Preserve Fruit Quality and Increase Storage Life Via Abscisic AcidSignaling in Strawberry, Food Chemistry (2019), doi: https://doi.org/10.1016/j.foodchem.2019.01.060
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, andreview of the resulting proof before it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.

1
Alginate Oligosaccharide Postharvest Treatment Preserve Fruit Quality and Increase Storage Life Via Abscisic Acid Signaling in StrawberrySantosh Kumar Bosea, b, c, Prianka Howladera, b, c, Jia Xiaochena, d, Wang Wenxiaa, d, *, Yin Henga, d,*
a Liaoning Provincial Key Laboratory of Carbohydrates, Dalian Institute of Chemical Physics, Chinese Academy of Sciences, Dalian, 116023, China.b University of Chinese Academy of Sciences, Beijing 100049, China.c Department of Horticulture, Patuakhali Science and Technology University, Patuakhali, Bangladesh. d Dalian Engineering Research Center for Carbohydrate Agricultural Preparations, Dalian, 116023, China.
* Address of correspondence author: Natural Products and Glyco-Biotechnology Lab, Dalian Institute of
Chemical Physics, Chinese Academy of Sciences, Zhongshan Road, Dalian, China [Telephone (0411)
84379061; Fax (0411) 84379061; E-mail: [email protected], [email protected]].
Authors email address:
1. Santosh Kumar Bose: [email protected]. Prianka Howlader: [email protected]. Jia Xiaochen: [email protected]. Wang Wenxia: [email protected]. Heng Yin: [email protected]

2
Abstract
Abscisic acid (ABA) has been advocated to play substantial role on ripening of non-climacteric
fruit. Here we report that alginate oligosaccharide (AOS) postharvest treatments delayed the
accumulation of ABA and ABA-conjugates and restrained the expression of ABA signaling
genes, resulting enlarged storage life of strawberry. In addition, AOS postharvest treatments also
increased the quality and reduced the degradation of cell wall components and repressed the
expression of cell wall degradation genes. AOS treated fruits exhibited significant delays of
hardness, decay percentage, titratable acidity, pH, total soluble solids and vitamin C content
compared to untreated fruits. Moreover, AOS had a positive effect on retaining higher amount of
anthocyanin, total phenol and flavonoids contents. The finding of this study suggests that AOS
postharvest treatments are very useful for preserving fruit quality and enhancing shelf life by
delayed ABA accretion, restrained the gene expression related to ABA signaling and cell wall
degeneration.
Keywords: Alginate oligosaccharide (AOS); quality; postharvest; abscisic acid; strawberry.
Chemical compounds studied in this article
CTAB: Cetyl Trimethyl Ammonium Bromide (Pubchem CID: 5974)
EDTA: Ethylenediaminetetraacetic acid (Pubchem CID: 6049)
BHT: 2, 6-Di-tert-butyl-4-methylphenol (Pubchem CID:31404)
CDTA: Trans-1, 2-diaminocyclohexane-N,N,N',N'-tetraacetic acid (Pubchem CID: 92240)
2, 6- dichlorophenolindophenol (Pubchem CID: 23697355)
Gallic acid (Pubchem CID: 370)
Sodium Borohydride (Pubchem CID: 4311764)

3
1. Introduction
Strawberry (Fragaria × ananassa) fruits are categorized as non-climacteric, highly appreciated
worldwide due to its unique taste, distinct flavor and also for its health benefits. Strawberry fruit
exhibit very short storage life mainly due to their susceptibility to various pathogens and
mechanical damage which can rapidly reduce the quality (Vu et al., 2011). To control
postharvest decay and increase shelf life of strawberry, different fungicides are used but most of
the fungicides leave residue to the fruit that have detrimental effect on both human and
environment (Li and Yu, 2000). Globally, consumers prefer more natural, eco-friendly
minimally processed products without considerable changes in their fresh characteristics with
preserved high nutritional quality and longer shelf life. Regarding this issue, the use of natural
protectors and antimicrobials is of growing interest for promoting nutritional quality and shelf
life of strawberry. In recent times, a notable research work has been conducted on the utilization
of natural polysaccharides (chitosan and alginate) and oligosaccharide (chitosan oligosaccharide)
as postharvest treatments for increasing the storage life of different horticultural products.
Among these polysaccharides and oligosaccharides, chitosan oligosaccharides (COS) has been
considered a potential plant immunity elicitor and used on several plants (Falcon et al., 2008).
Pre-harvest spraying of strawberry fruits with COS are very effective for enhancing quality,
higher antioxidant activity and longer postharvest life (He et al., 2018). In addition, COS
postharvest treatment also had a noticeable effect on minimization of decay percentage and
ripening of strawberry fruits (Ma et al., 2014).
Alginate is a natural polysaccharide extracted from brown sea algae (Phaeophyceae) having a
strong affinity for water and is a biopolymer that has a coating function (Acevedo et al., 2012).
Alginate oligosaccharide (AOS) extracted through hydrolysis of alginate, is a water-soluble non-

4
toxic compound, plant biostimulator which can regulate plant development and innate immunity
system to enhance plant quality and act against plant diseases (Battacharyya et al., 2015). In
recent times, AOS has paid much consideration because of their uncommon properties. The
utilization of AOS in various fields including agriculture, food and pharmaceutical industry has
been studied recently. Moreover, frozen shrimp treated with AOS, decreased thawing and
cooking losses, preserved texture and myofibrillar protein content (Zhang et al., 2017). The
above report suggested that AOS might be an excellent promising food additive to maintain
quality during storage. However, the effect and mechanism of AOS on conserving fruit quality is
staying ambiguous.
Strawberry fruit softening seems to be closely associated with pectin degeneration and has been
correlated with the activity of a several cell wall hydrolyses enzymes. The hydrolysis,
solubilization and depolymerization of cell wall by different enzymes such as pectin esterase
(PE), pectin lyase (PL), polygalacturonase (PG) andendo-1,4-d-glucanase (EG) during fruit
ripening are closely associated with fruit softening (Pose et al., 2018). Khan and Singh (2007)
reported that the application of 1-MCP, repressed the activity of fruit softening enzymes (PE, EG,
exo-PG and endo-PG) during storage resulting increment the storage life of ‘Tegan Blue’ plum
fruit. In this sense, postharvest fruit treatments paid attention for retarding the activity of cell
wall degeneration enzymes which could help to increase the shelf life of quickly spoiled fruits,
like strawberry.
ABA considered to play very important role during fruit maturation and senescence. ABA
participated and played promising role in non-climacteric watermelon ripening. The ABA
contents raised in watermelon during whole period of fruit development and ripening, reaching
their highest levels at the ripening stage (Wang et al., 2017). Ji et al. (2012) noted a dramatic

5
increment of ABA content throughout development and ripening of strawberry. ABA content
progressively increases throughout fruit development and participated in ripening of strawberry
fruit (Jiang and Joyce, 2003). The increasing content of ABA suggests a potential role during
fruit ripening, but little advancement has been made regarding understanding the ABA signaling
pathway in strawberry fruits. ABA conjugates are the storage form of ABA and biologically
active ABA is released during hydrolysis of these conjugates resulting increased the
concentration of free ABA (Zhang et al., 2014; Lee et al., 2006). The most considerable forms
of ABA conjugates is ABA-glucosyl ester known as ABA-GE. ABA-GE, is also the
considerably most important storage form of ABA and to a great degree of wide spread ABA
conjugate. Zhang et al. (2014) previously reported that ABA-GE exists in red colored strawberry
fruit, which promoted the accumulation of ABA by hydrolysis of ABA conjugates, plays a role
in fruit ripening.
To understand the molecular mechanisms related to ABA-mediated ripening, ABA signaling
pathways are essential to study. Some important information regarding ABA biosynthesis and
signaling was reported in former work. FaNCED is an important ABA biosynthesis gene, the
inhibition of FaNCED1gene expression reduces ABA concentration and fruit red coloring in
strawberry (Fragaria × ananassa cv. Fugili) and the highest expression of FaNCED gene was
reported at the end of the ripening (Jia et al., 2011). The reduction of FaNCED expression
correlates with retardation of ripening in strawberry (Jia et al., 2011).
ABA action is very important in case of physiological change of strawberry and is started by
ABA perception, which generates downstream signaling cascades to induce physiological
effects. The down-regulation of FaCHLH/ABAR gene may potentially give rise to early ABA
signaling resulting inhibits fruit ripening (Jia et al., 2011). Another receptor gene FaPYR1 is

6
involved in later reddening of strawberry fruit was found by transcription analysis (Chai et al.,
2011).
An important transcription factor FaASR on ABA downstream signaling was identified in recent
years. During development and ripening of strawberry, the higher expression FaASR gene in
response to ABA treatment was observed by Chen et al. (2011), proved that FaASR gene was
also up-regulated by ABA and contributes to accelerate strawberry fruit ripening. These data
suggests that ABA is a signaling molecule that participates and regulate strawberry fruit
ripening.
However, to the best of our knowledge, the effect and mechanism of AOS postharvest treatments
has not been reported in ripe strawberry fruits. Therefore, initially the present study was
conducted to assess the postharvest application effect of AOS on decay rate, fruit quality and,
cell wall degradation of strawberry. Then, to explore the role of ABA and ABA conjugates
content during storage as well as some important genes involved in ABA biosynthesis and
signaling of strawberries has been checked thoroughly in the present study.
2. Materials and Methods
2.1 Electrospray ionization mass spectrometry (ESI-MS) analysis of AOS
AOS was produced in our lab by previously identified alginate lyase. Electrospray ionization
mass spectrometry (ESI-MS) was used for profiling the oligosaccharide composition. To discard
proteins and salts, microcrystalline cellulose column followed by a cation exchange column was
used and then evaporated, dried and liquified with 1 ml of HPLC grade methanol. The clear
aqueous solution was collected by centrifugation at 12000 rpm for 3 minutes, 2 µl of the clear
liquid was inserted to an LTQ-XL linear ion trap mass spectrometer (Thermo Fisher Scientific,
Waltham, MA, USA). The following settings were used for the detection of oligosaccharides;

7
measuring temperature 275-300 ºC, electromotive force 4.5 kV, hose lens 250 V, covering gas
30 arbitrary units (AU) and mass range 300-2000 m/z.
2.2 Plant materials
Uniform size, shape and color without any blemished commercially mature strawberry (Fragaria
× ananassa) fruits were collected from farmer’s field located at 39° 25' to 40° 12' N latitude and
122° 29' to 123° 31' E longitude (Zhuanghe city, Liaoning Province, China) on April 2017. A
total of 360 fruits were randomly selected for the experiment and fruits were divided in three
groups with 120 fruits in each treatment.
2.3 Application of AOS treatments
AOS powder was weighted with the help of a electric balance and dissolved in distilled water at
a concentration of 50 and 100 mg.L-1. Then two groups of fruit were treated with different
concentration of AOS solution for 60 second and the remaining group was used as control. The
control group was immersed in same volume of distilled water. Then the treated and untreated
fruits were kept on brown paper at room temperature for drying the solutions. After drying, the
fruits were placed in phytotron and stored at 20±2ºC with 80±2% relative humidity. From each
fruit group, 15 fruits were taken at every alternate day for measuring the hardness. After
measuring the hardness fruits were cut into small pieces and group wise packed with aluminum
foil, immediately dipped in liquid nitrogen and then stored in -80 ºC for further analysis. The
weight loss and decay percentage was calculated from three fixed groups.
2.4 Hardness, weight loss and decay percentage analyses
Digital penetrometer (Stable Micro System Ltd., Surrey UK) along with a measuring probe
(5mm diameter stainless steel) were used for hardness determination. From each treatment, 15
fruits were used and every fruit was divided into two equal parts with the help of a sharp knife.

8
Fruit hardness was measured from two opposite sides of each fruits by penetrating the probe at a
distance of 5 mm into the fruit with pre- and post-test speed 1mms-1. The hardness was
calculated as maximum penetration force reached during tissue breakage and expressed as
Newtons (N).
Weight loss % was estimated as [(Initial weight-Final weight)/Initial weight] ×100
Fruits with visible brown spot and softened area caused by fungus or any other microorganisms
infection were regarded as decayed fruit. The number of decayed fruits were counted every
alternate day from three fixed group of each treatment and calculated by the following equation:
D (%) = ND/NT×100, where ND express as the number of decay fruits and NT indicate the number
of total fruits.
2.5 Total sugar (TS) and total soluble solids (TSS) analyses
The amount of total sugars (TS) of storage strawberry fruit was determined by modified
anthrone-sulfuric acid colorimetric method as described by Jayaraman (1981). The percentage of
total sugar was calculated by using the following formula:
100samplesofWeight
obtainedsugar ofAmount sugar Total%
The TSS content of storage strawberry fruit was estimated by using hand refractometer
(Shanghai optical instrument, 2W, China). Fruits were homogenized in a kitchen blender
(Joyoung, JYL-C50T, China) for 2 minutes and filtrated through 4 layers of muslin cloth. By
using a dropper, few drops of the extract were placed on the refractometer prism glass and data
was noted from direct reading.
2.6 Titratable acidity (TA) and pH determination

9
Titratable acidity (TA) was analyzed according to the method described by Ranganna (1977)
with minor modification. The amount of TA was calculated by using following formula and
expressed as citric acid percentage.
Citric acid (%)
=Titre (mL) × NaOH normality (0.1N) × vol.made up (50 mL) × citric acid eq.weight (64 g) × 100
Vol.of sample for titrate (5 mL) × wt.of sample taken (10 g) × 1000
The remaining juice extract from TA measurement was used to measure the pH of the fruit pulp.
The pH was determined by using a glass electrode pH meter (PHS-25 Precision pH/mV Meter).
2.7 Vitamin C, total anthocyanin, total phenols and flavonoid content determination
Vitamin C content was calculated according to the 2,6-dichlorophenolindophenol titration
method as described by He et al (2018) with minor modification. Shortly, 5 g strawberry was
extracted with 50 mL of 2% oxalic acid solution and then upper aqueous phase was collected by
centrifugation at 12000 rpm for 10 min at 4 ◦C. An aliquot of 10 ml was titrated by using 0.1%
2,6-dichlorophenolindophenol to a pink color. Vitamin C concentration was calculated using
following formula and expressed as mg·100 g−1 FW.
Ascorbic acid (mg/100g) =Titre (ml) × dye factor × vol.made up (ml) × 100
Aliquot used for estimation (ml) × sample weight (g)
Extraction and quantification of total anthocyanin content was performed according to Lees and
Francis (1972) with slight alteration. Briefly, 100 g of fruit pulp was homogenized in a kitchen
blender for two minutes with 100 mL of extracting solvent (70% ethanol in water and adjusted to
pH 2.0 with 0.1% HCl) and kept it refrigerator overnight at 4 °C in dark condition. After
incubation, the fruit samples were filtered into a volumetric flask using Whatman paper and for
preventing light, the flask was covered with the aluminum foil. For completely removal of
pigments, the remained residue was washed two times with same solvent and filtered as

10
discussed above. All filtrate solutions were mixed together and adjusted to a total volume of 200
mL with the same solvent. A 2 ml extract was taken in a 100 ml volumetric flask and the volume
was adjusted with same solvent. Then solutions were placed in dark condition at room
temperature. Two hours later, absorbance was recorded at 535 nm by using spectrophotometer
for the quantification of anthocyanin content.
The amount of total phenol in storage strawberry fruit was estimated using Folin Ciocalteu
Reagent as described by Bray and Thorpe (1954), with some alteration. Concisely, 10 mL of
80% methanol was used for extracting 0.5 g fruit and the upper aqueous phase was collected by
centrifugation at 10,000 rpm for 20 min. The final concentration of collected aqueous solution
was made 1mg/ml with same extracting solution. Then 0.5 ml supernatant was mixed with 2.5 ml
10% Folin Ciocalteu Reagent and 7.5% NaHCO3. Thereafter the mixed sample was incubated in
a oven for 1 hour at 60ºC. The absorbance was read by using spectrophotometer at 765 nm and
the phenolic content was measured as gallic acid equivalent (mg of GA/100g fresh weight)
against prepared blank sample. The total flavonoid content was determined following the method
previously described by Zhishen (1999) with some changes. Total flavonoid content was
standardized against rutin and expressed as milligrams of rutin equivalent (RE) per 100 grams
fresh weight (FW).
2.8 Cell wall fractionation analyses
Cell wall components were achieved as alcohol-insoluble residue (AIR) according to d’Amour et
al. (1993) with minor alteration. Almost 100 mg of AIR was suspended in 15 ml of water and
agitated at room temperature for 12 h. The aqueous phase was collected by centrifugation at
10000g for 20 min at 4°C, the sample sediments were washed twice with 15 ml of distilled water
and collected by same way. The clear liquid and water washings were mixed together and named

11
as water-soluble fraction (WSF). The remainder was then agitated with 15 ml of 50 mM trans-
1,2-diaminocyclohexane-N,N,N′,N′-tetraacetic acid (CDTA) (pH 6.5) for 12 h at room
temperature. The mixture was centrifuged, the clear liquid was collected as above, and the
sediment was washed twice with 15 ml of CDTA solution. The buoyant and washings were
pooled and marked as CDTA soluble fraction (CSF). The CDTA-insoluble sediment was then
stirred with 15 ml of 50 mM Na2CO3 containing 20 mM NaBH4, for 1 h at 4°C. The stirred
solution was centrifuged, the supernatant was collected as above, and the residue washed twice
with 15 ml of Na2CO3 solution. The clear liquid and washings were combined together and
named as Na2CO3 soluble fraction (NSF). The remainder was again stirred with 15 ml of 4 M
KOH containing 1 mMNaBH4, for 1 h at 4 °C. The aqueous phase was collected as above, and
the remainder was washed twice with 15 ml of KOH solution. The collected clear liquid and
washings were neutralized with acetic acid and recorded as hemicellulose and the remaining
final fraction named as cellulose.
2.9 Determination of endogenous ABA and ABA conjugates from strawberry
The internal ABA content of storage strawberry fruits were measured by using ultra-performance
liquid chromatography-mass spectrometry (UPLC-MS) as explained by Symons et al. (2012)
with minor alteration. Shortly, 1.0 g powder was incorporated with 10 mL of 80 % methanol
including 1 mg of 2, 6-di-tert-butyl-4- methylphenol (BHT). The suspension was allowed to rest
12 hours at 4°C and then seep through a filter paper. The filtered was evaporated to less than 1
mL at 35 °C using rotary evaporator. The concentrate was collected by washing the tube with 3
×3 mL of 10 % methanol holding 0.4 % acetic acid and purify by passed through Sep-Pak C18
cartridge (Waters Corporation, Milford, MA, USA). The collected elutes were vacuum dried at
40 ºC, the residue was re-suspended in 500 µl HPLC grade methanol and centrifuged at 12,000g

12
for 3 min. Finally, clear aqueous phase was collected by passed through a 0.45 μm filter and
loaded into target vials for ABA analysis.
ABA-conjugates was extracted and detected by using UPLC-MS. Briefly, fruit samples were
ground as described in ABA extraction, and 1 g of powder was transferred in a 50 ml tube.
Samples were then extracted with 8 ml of 10 mM CaCl2 for 24 h at 4°C. The aqueous solutions
were acidified (pH 3.0) with 0.1M HCl and separated three times against ethyl acetate by
centrifuged at 20,000 ×g for 20 minutes. The lower aqueous fractions containing ABA
conjugates were hydrolyzed with 1M NaOH for 1 h and then pH was adjusted to 3.0 using 0.1M
HCl and partitioned again as described above. Collected aqueous phase were combined together
and injected into a Sep-Pak C18cartridge (Waters Corporation, Milford, MA, USA). After that
samples were prepared as described in ABA quantification.
For detection of ABA and ABA conjugates, used a hypercarb column (Thermo Fisher, 150 × 2.1
mm, 5 µm) at 35 °C, and 0.3% formic acid adjusted to pH 9.0 with ammonium hydroxide
(mobile phase A) and 100% acetonitrile ( mobile phase B) were used as elution gradient at a
flow rate of 0.2 mL/min up to 20 minutes. The details of gradient conditions were as follows: 0
min, 90%A+10%B; 10 min, 40%A+60%B; 14 min, 40%A+60%B; 14.1 min, 90%A+10%B, and
20 min, 90%A+10%B. Q-trap 5500 (AB SCIEX, USA) with optimized collision energy was
used for hormone detection. The mass spectrometer was conducted in the negative ion mode
with the following conditions: The voltage of ion spray: −4500 V, de-clustering potential (DP):
−80 V, entrance potential (EP): −10 V, temperature source (TEM): 120°C. Parent and fragment
ions were detected in multiple reactions monitoring (MRM) mode for mass transitions under
following settings: precursor ion (m/z): 263, production ion (m/z): 153/219 and collision energy

13
(ev): 25. Instrument control and data processing software was used for (Analyst software, AB
SCIEX, USA) calculation of data.
2.10 RNA extraction, cDNA preparation and gene expression analyses
Total RNA was extracted from 5.0g fruits by using modified CTAB methods as described by
Sambrook et al. (1989). The extracted RNA was quantified by agarose gel electrophoresis and
absorbance ratio at 260-280 nm. Genomic DNA was eradicated by the treatment with RNase-free
DNase (Promega Biotech Ibérica. Madrid, Spain), cDNA was prepared by using AMV Reverse
Transcriptase (Takara, Dalian, China) according to the manufacturer’s protocol. For RT-qPCR
analysis, 10 µl Power SYBR Green PCR Master Mix (Applied Biosystems), 0.4 µL gene specific
forward primer, 0.4 µl gene specific reverse primer, 2 µl cDNA, and 7.2 µL DEPC water were
mixed together in a final volume 20 µl. The sequences of the primer pair used for each gene are
shown in Supplementary Table 1. DNA amplification was carried out under following
thermocycling conditions: 95 ℃ for 2 min followed by 40 cycles at 95 ℃ for 5 seconds, at 58℃ for
10 seconds and at 72 ℃ for 10 seconds, using the RT-qPCR machine (qTOWER 2.2, Analytik Jena
AG, German). Actin was used as reference gene. The relative expressions of desire genes were
checked by following the 2−ΔΔCt method as explained by Livak and Schmittgen (2001).
2.11 Statistical analyses
The whole experiment was carried out following completely randomized design (CRD) with
three replications. Data were articulated as mean ± SD (n =3). The data were compiled and
analyzed by using SPSS 22.0 (IBM, New York, USA) for calculating ANOVA and the standard
deviation (SD). The level of significance difference was established at P< 0.05.
3. Results and Discussion
3.1 ESI-MS analysis of AOS

14
The AOS which we used in this study was prepared in our laboratory by enzymatic hydrolysis of
alginate. The biological activity of oligosaccharides is highly dependent on their degree of
polymerization (DP). In order to confirm the degree of polymerization (DP) of alginate
oligosaccharides, ESI-MS analysis was conducted. From the Fig. 1, it can be observed that ion
peaks that emerged at m/z showed the DP of AOS is from 2 to 7. Zhang et al. (2015) reported
that the maximum elicitor activity of alginate degradation product was observed with a DP of 7.9
and 6.5 in germinating rice seedlings. So, we validated our produced AOS DP within 2 to 7 and
used in our experiment.
3.2 Effect of AOS on hardness, weight loss and decay percentage
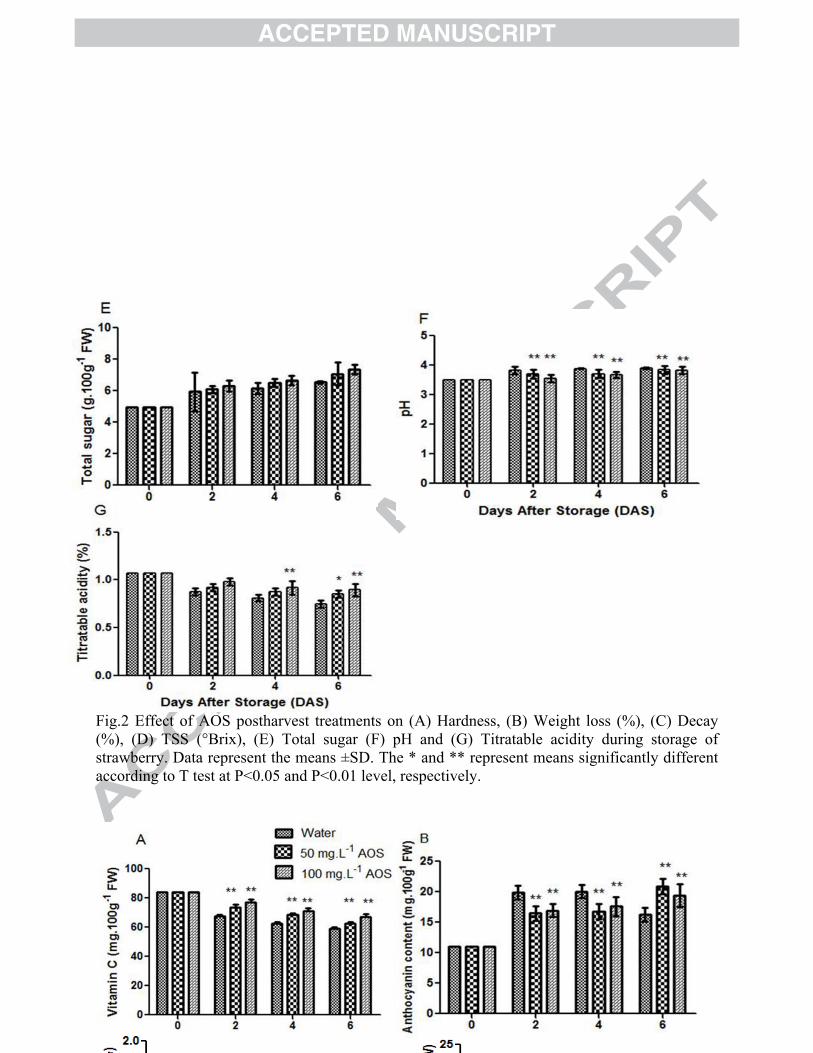
The hardness of fruit is one of the most vital indices that govern the quality of fruits during
ripening. The hardness declined entire storage period in both treated and untreated fruits and
reached the lowest values at the end of storage period (Fig. 2A). The 50 and 100mg.L-1AOS
treatment significantly delayed the decrease of strawberry hardness compared to the untreated
fruit. He et al. (2018) reported that pre-harvest spraying of COS is effective for preserving the
hardness of strawberry. During fruit ripening, hardness devaluation occurs mainly due to the
rupture of the middle lamella and the internal change of the composition and structure of
polymers existent in the primary cell wall (Brummell, 2006).
The weight loss increased during storage in case of all AOS treated and untreated strawberry
fruit (Fig.2B). AOS treatment had no significant effect but delayed the weight loss percentage
compared to untreated strawberry during storage. At the end of the storage period (6 DAS), the
highest weight loss (34.67%) was noted from untreated fruits whereas the lowest weight loss
(29.73%) was counted from 100mg.L-1AOS treated fruits. Due to the extremely thin skins,
strawberry fruits are highly perishable and susceptible to rapid water loss. The cause of weight
loss during storage of fruits mainly due to the movement of water from fruit to the atmosphere

15
(Duan et al., 2011). Our results suggest that AOS had no coating effect but reduced weight loss
compared to untreated fruit. Reduction of weight loss at 100 mg.L-1AOS treated fruits may
possibly be due to lowering respiration and metamorphic activity of fruits.
Strawberry fruits exhibited very short storage life as a result of sensitivity to mechanical damage
and storage pathogens that can quickly decline the fruit quality and market value (Vu et al.,
2011). From the results, it was observed that the decay percentage increased entire the storage
period in case of all treated and untreated fruits (Fig. 2C). However, AOS significantly (P< 0.05)
inhibited the decay percentage of storage strawberries compared to the untreated fruit. Babalar et
al. (2007) reported that SA is an important plant hormone that effectively reduced decay
percentage in ‘Selva’ strawberry fruit. Previous reports also showed that COS treatment had
noticeable effect on reduction of postharvest fruit decay (Ma et al., 2014). Our study revealed
that AOS also have similar function as SA and COS, suggest that AOS treatment on strawberry
against decay, performed good result probably due to the defense response of AOS toward
pathogen infection.
3.2 Effect of AOS on total soluble solid (TSS) and total sugar (TS) content
TSS values gradually increased in all AOS treated and untreated strawberry fruits until 4 days of
storage and then decreased (Fig. 2D). During storage, AOS treated fruits exhibited significantly
(P<0.05) higher TSS content compared to untreated fruits. In untreated fruits, TSS content
decreased from (10.07 ° Brix) to (7.11° Brix) during 4 days to 6 days of storage. Our results
validate the results sated by Velickova et al. (2013) who noted that TSS content of strawberry
declined at the end of storage.
Total sugar (TS) content of strawberry fruit was affected by AOS treatment. No significant effect
was observed between treated and untreated fruits during storage but AOS treatment enhanced

16
the TS during entire storage period and 6 days of storage 100mg.L-1treatment showed higher TS
than untreated fruits (Fig. 2E). From the above results, it was noted that TS content gradually
increased with the increasing storage period and AOS 100mg.L-1 was more effective treatment
from which we obtained the highest TS. This remarkable increase in total sugars may be
attributed to the conversion of organic acid to starch during ripening.
3.3 Effect of AOS on pH and titratable acidity (TA) content
The pH of strawberry fruits enhanced sharply throughout the storage period and significant
difference (P<0.05) was marked between treated and untreated fruits (Fig. 2F). Geransayeh et al.
(2015) also reported that postharvest SA treatment delayed the pH increase during storage of
strawberry. With the advancement of fruit maturity organic acids concentration are decreases.
The organic acids are used in the respiration process during advances of fruit from development
to ripening stage and increase the sugar content of fruit resulting enhance pH (Kafkas et al.,
2007).
The rise in pH and decrease in TA indicates that the acid concentration in fruits is declining with
maturity. TA content in strawberry fruit significantly (P<0.05) decreased during entire storage
period with lower values in untreated fruits compared to AOS treated fruits (Fig.2G).
Maftoonazad et al. (2008) reported that titratable acidity is the indicator of acidity of fleshy fruit,
is directly related to the amount of organic acid present in the fruit, during ripening the
devaluation of acidity may be due to the metabolic changes in fruit and use of organic acid in the
respiratory process of fruit.
3.4 Effect of AOS on vitamin C, total anthocyanin, total phenol, flavonoid content and antioxidant activity
Vitamin C (Vc) is only a minor constituent of fruits and vegetables but it has a vital role in
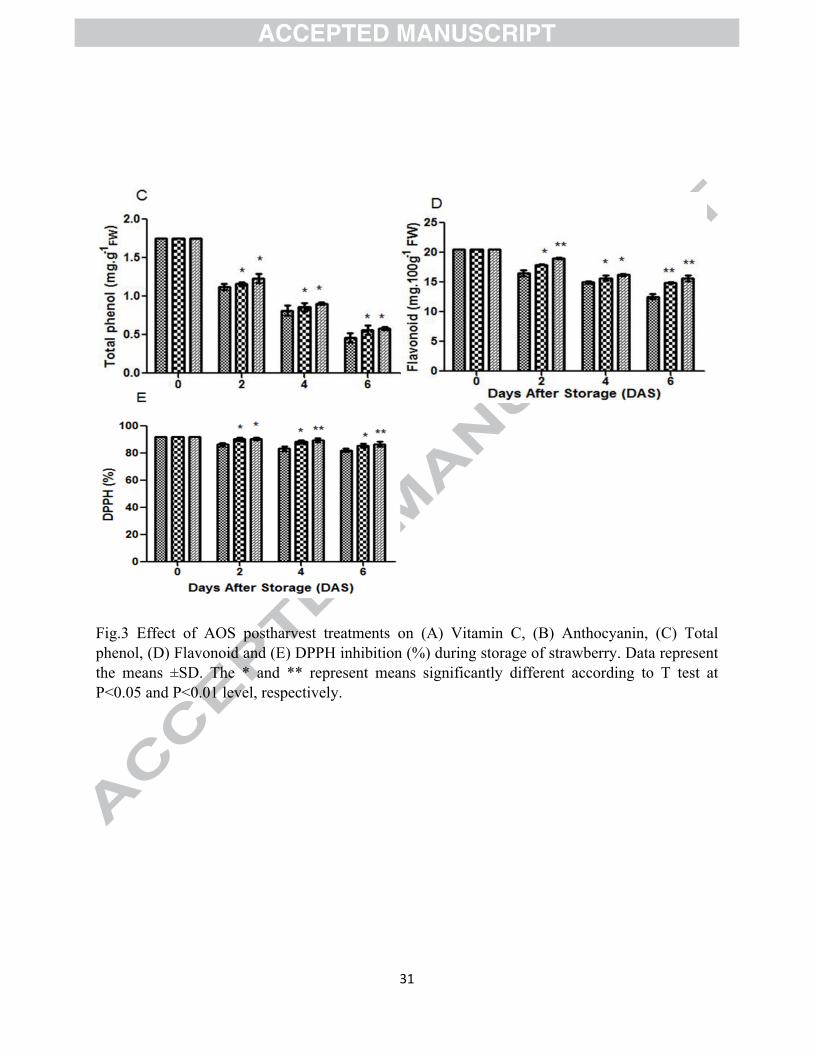
human nutrition. The content of Vc in both treated and untreated fruit decreased during prolong

17
storage time and AOS treatment significantly (P<0.05) inhibited the decrease (Fig. 3A). The
similar observations were also noted by Atress et al. (2010), who reported that Vc content of
strawberry decreased during storage. Vc decreased with the prolong storage period because it is a
water soluble compound and the rate of decrease is positively correlated with the reduction of
moisture content and weight loss of fruits. In our study, we also observed declining trend of Vc
content during prolong storage period but AOS treatment inhibited the decreasing rate may be
due to the inhibition activity of AOS against oxidation reaction of Vc and weight loss of fruits.
During storage, both treated and untreated fruits showed a significant increase in anthocyanin
content (Fig. 3B). The result of the present study demonstrated that up to 4 days of storage
anthocyanin content was higher in untreated fruit compared to AOS treated fruit but at the end of
storage (6 DAS), the highest anthocyanin content was recorded from AOS treated fruits. Atress
et al. (2010) revealed that anthocyanin plays an important role for the coloration of strawberry
and fruit become redder and darker due to synthesis of anthocyanin pigment during storage.
Anthocyanin possesses divers biological properties and are considered as secondary metabolites
with a potential health benefits.
Total phenol content in treated and untreated strawberries during storage is presented in Fig. 3C.
The total phenol content gradually decreased during entire storage period but control fruits had
significantly (P<0.05) faster decrease. During ripening and senescence of strawberry fruit, the
phenol content decrease might be due to the disruption of cell structure (Macheix et al., 1990). In
our study, we also found similar result as reported by Macheix et al. (1990). AOS treatments
significantly delayed the reduction of phenol content compared to the control may be due to the
inhibitory effect against cell wall loosening.

18
Flavonoids possess important pharmacological properties, such as antioxidant, anti-
inflammatory, antimicrobial, anticancer, and anti-diabetic activities. The flavonoid content in
both treated and untreated fruit decreased during prolong storage time and AOS treatment
significantly (P<0.05) inhibited the decrease rate (Fig. 3D).
The result of the present study demonstrated that the antioxidant activity of both treated and
untreated strawberry fruits had decreasing trend but showed significant (P<0.05) difference
among the treatments (Fig. 3E). The fruit treated with 100 mg.L-1 AOS exhibited higher
antioxidant activity compared to untreated fruits at the end of storage. From the above
discussions it can be concluded that that AOS 100 mg.L-1 postharvest treatment preserved fruit
quality by inhibiting the decreasing rate of Vc, phenolic compounds and then antioxidant
capacity.
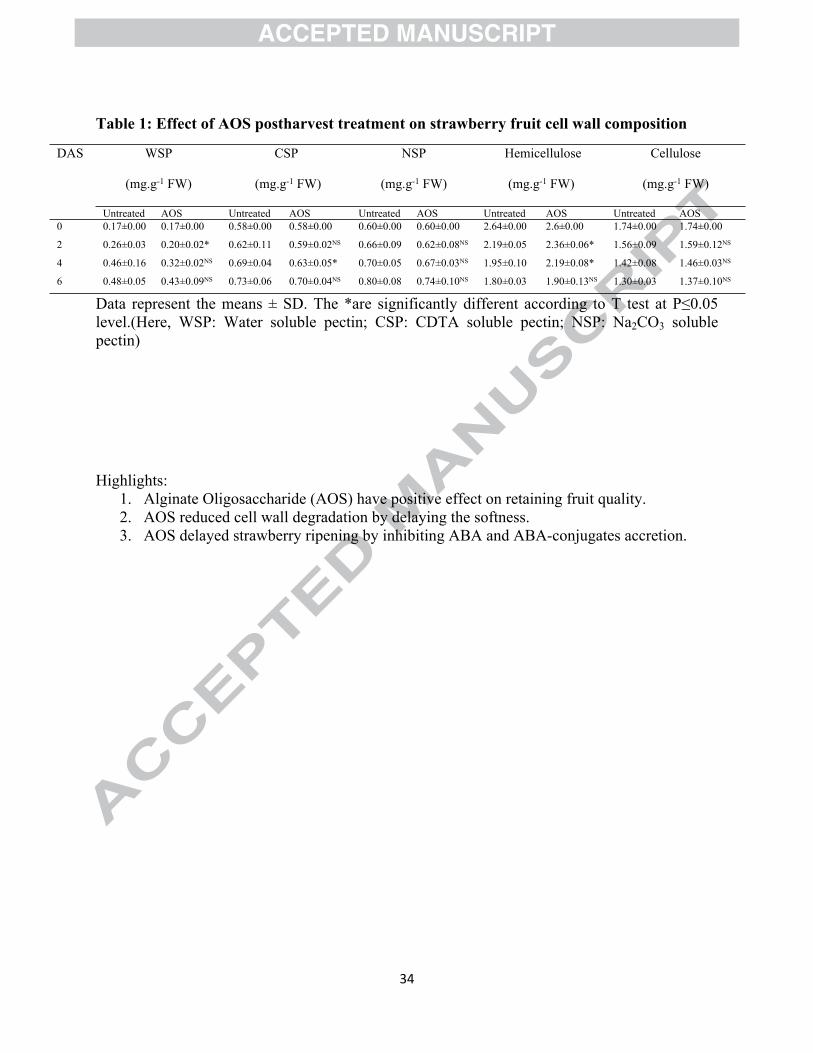
3.5 Effect of AOS on cell wall components and related gene expression
The main cell wall fractions WSP, CSP, NSP, hemicellulose and cellulose were sequentially
extracted from both treated and untreated fruits during storage. The results of the present study
showed that the cell wall components like WSP, CSP, NSP increased and hemicellulose,
cellulose are decreased during storage (Table 1). The devaluation of fruit firmness during fruit
ripening is mainly due to the disruption of the middle lamella and the internal changes of the
composition and structure of polymers present in the primary cell wall (Brummell, 2006). During
storage, the hardness of strawberry fruits gradually decreased mainly due to the disintegration
and depolymerization of pectin and hemicellulose from cell walls. Fruit softening is associated
with cell wall disassembly and during fruit softening, pectin and hemicellulose in cell walls
undergo solubilization and depolymerization which contribute to cell wall loosening (Pose et al.,

19
2018). These polysaccharides play an substantial role in fruit consistency and in its textural
changes during ripening.
Fruits softening are closely correlated with disintegration and depolymerization of different cell
wall enzymes such as pectin esterase (PE), pectin lyase (PL), polygalacturonase (PG) and endo-
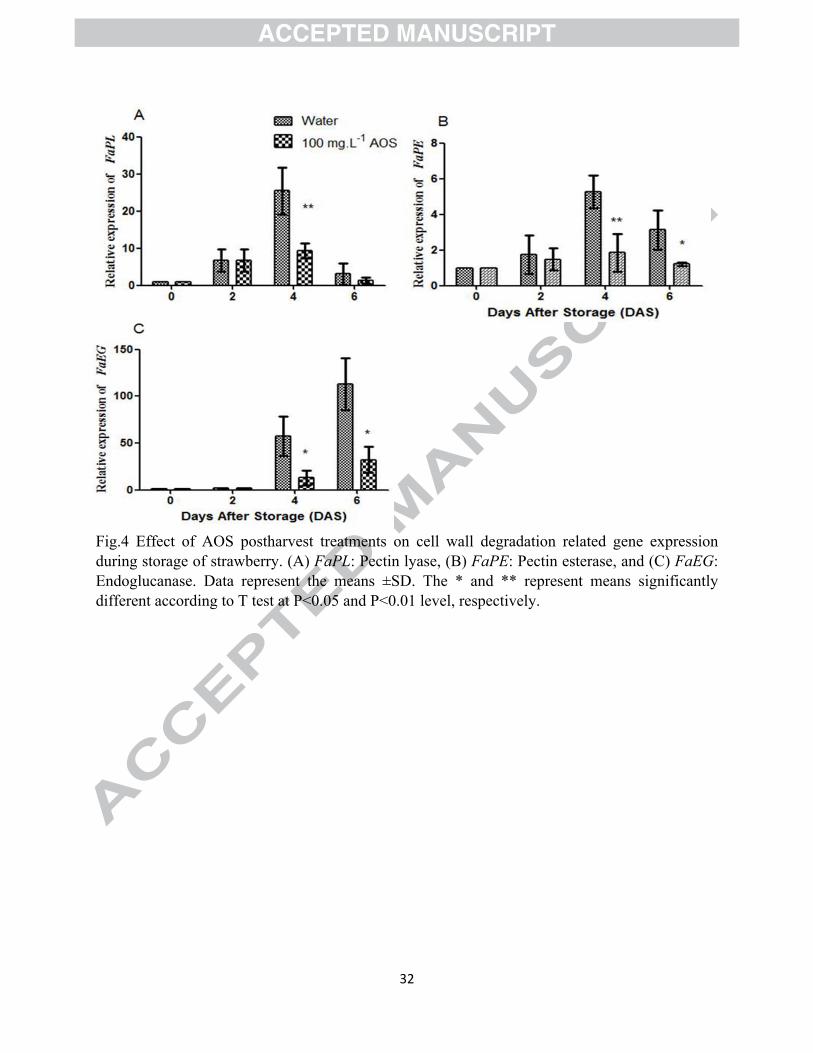
1,4-d-glucanase (EG), during ripening (Pose et al., 2018). In this study, we found that FaPL,
FaPE and FaEG genes showed very high transcript levels in a strawberry during storage (Fig. 4
A-C), which harmonized with the cell wall degradation during fruit softening, providing
evidence that cell wall modifying enzymes controlled the cell wall degradation (Ortiz et al.,
2010). These results showed that AOS could delay the degradation of cell wall by inhibiting gene
expression related to cell wall degradation enzymes, which in consistent with the results of cell
wall fractions change.
3.6 Effect of AOS on ABA and ABA conjugates content and ABA related gene expression
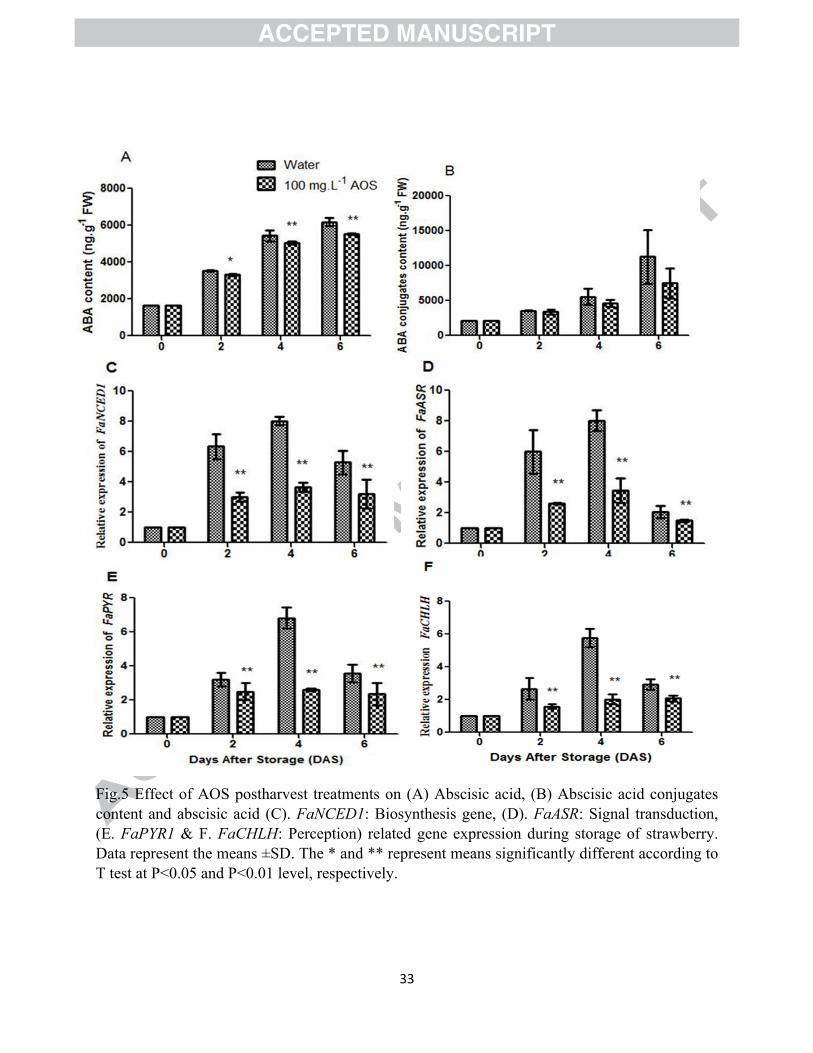
Former works remind that ABA play important role on the ripening of non-climacteric fruits. To
explore the AOS influencing effect on ABA signaling pathway and to confirm the ABA role in
strawberry fruit ripening, we first quantified ABA and ABA conjugates content and detected an
increasing trend during storage of strawberry. ABA content was significantly differed (P<0.05)
among the treatments but ABA conjugates did not show significant difference between treated
and untreated fruits (Fig. 5A-B). AOS 100 mg.L-1 treatment significantly inhibited the increment
of ABA during storage i.e untreated fruits exhibited higher ABA content compared to AOS
treated fruits during entire the storage period. Ji et al. (2012) observed dramatic increase of ABA
levels during ripening of strawberry which support our present study results. Wang et al. (2017)
also noticed that ABA is involved and plays an important role in non-climacteric watermelon
ripening, the ABA contents increased in watermelon throughout fruit development and ripening,

20
reaching their highest levels at the ripening stage. In this study, it can be observed that AOS
treatment inhibited ABA accumulation during ripening, the possible reasons of ABA
accumulation inhibition during ripening may be due to the inhibitory effect of AOS on the
activity of ABA synthesis related different enzymes during storage.
On the other hand, the most exuberant forms of ABA conjugate is an ABA-glucosyl ester
named as ABA-GE, plays an pivotal role in the regulation of ABA level and the most crucial
storage form of ABA. The ABA-GE present in red colored strawberry fruit, which promoted the
accumulation of ABA by hydrolysis of ABA conjugates, plays an important role in fruit ripening
(Zhang et al., 2014). In our study, we observed that ABA conjugates content increased
throughout the storage period which was paralleled to the ABA content of fruits, so we assumed
that ABA conjugates also have potential impact on ABA releasing and ripening of strawberry
during storage.
Previous results implied that ABA is an important hormone and playing vital role during
strawberry ripening, and ABA synthesis enzymes are the key point to regulate ABA content.
However, to confirm the role of ABA underlying strawberry fruit ripening, we examined the
expression of FaNCED1 (biosynthesis), FaASR (signal transduction), FaPYR and FaCHLH
(perception) genes. The gene expression results in both treated and untreated fruits during
storage of strawberry are shown in Fig. 5 (C-F). From the results we observed that AOS
treatment significantly (P<0.05) suppressed the ABA related gene expression that was similar
with ABA content during storage. It indicates that ABA contribute a potential role during fruit
ripening and AOS treatment delayed this ripening process by suppressing ABA related gene
expression.

21
Sweet cherry treated with ABA, elevated internal ABA biosynthesis during ripening was
reported by Ren et al. (2011). Jia et al. (2011) reported the highest expression of FaNCED1 gene
at the end of the ripening of strawberry (Fragaria × ananassa cv. Fugilia), and concluded that
this gene might be involved in the regulation of strawberry fruit ripening because during ripening
the signal molecule sucrose is increase and play a indispensable role in ABA accretion which
enhance the mRNA expression levels of FaNCED1. In our results it was revealed that the
transcription levels of FaNCED1 were congruent with ABA content as evidenced by qRT-PCR
and this result advocate that FaNCED1 might be involved in the regulation of strawberry fruit
ripening.
In addition, to validate the performance of ABA in strawberry fruit ripening, we also investigated
the expression pattern of ABA receptor genes (FaCHLH and FaPYR1) and observed that these
two genes expression were synchronized with ABA content during strawberry fruit ripening.
This result suggests that FaCHLH and FaPYR1 genes might be participated in governing the
strawberry fruit ripening. Chai et al. (2011) revealed that ABA receptor gene FaPYR1 is mainly
involved in early strawberry fruit growth and later reddening and act as a positive regulator of
strawberry fruit ripening.
In the present study we also observed the increasing trend of transduction pathway gene
expression (FaASR) which was similar with ABA content in fruits during storage. Chen et al.
(2011) reported that increment of internal ABA content inducing the FaASR gene expression at
transcriptional levels which partially contribute to the hastening strawberry fruit ripening. The
FaASR gene expression enhanced by exogenous spraying of ABA at development and ripening
stages of strawberry and higher FaASR expression is mainly related to ABA content in
strawberry (Chai et al., 2011).

22
In our study, we observed that during storage, ABA content of strawberry gradually increased
which influenced the ABA related gene expression that is in agreement with above discussions
and also Ricardo et al. (2016), who reported that ABA stimulates the perception (FaPYR1 and
FaCHLH), signal transduction pathway (FaASR) and ABA biosynthesis genes (FaNCED1)
expression. Experimental evidence indicates that, in case of non-climacteric fruits, the effects of
ABA on fruit ripening are not only dependent on ABA metabolism, but also on the presence of
an intact ABA and ABA signaling pathways (i.e., biosynthesis, perception and signal
transduction). Our results suggest that, AOS postharvest treatments delayed the accumulation of
ABA and suppressed the expression of ABA signaling genes, resulting increased the storage life
of strawberry.
Conclusions
The findings of the present study demonstrate that AOS postharvest treatment have a gainful
impact on the retention of strawberry fruit quality during storage. AOS treatments preserve
higher levels of hardness, TA, pH, TSS, Vc, anthocyanin, total phenol and flavonoids and reduce
the decay percentage. Furthermore, AOS postharvest treatments also delayed the degradation of
cell wall components and repressed the expression of cell wall degradation related genes.
Moreover, AOS also retard the accumulation of ABA content and suppressed the expression of
ABA signaling genes. In addition, the results of the present study advocate that AOS postharvest
treatments enhanced the strawberry fruit shelf life by regulating the ABA signaling. From above
findings, it can be concluded that AOS could be used as environment-friendly compounds and
partial substitution of chemical fungicides to maintain strawberry fruit quality during storage.
Acknowledgements

23
This work was supported by the National Key R & D program of China (2017YFD0201302),
NSFC (31672077, 31370811), CAS Youth Innovation Promotion Association (2015144) and
Natural Science Foundation of Liaoning Province, China(201602742). Santosh Kumar Bose and
Prianka Howlader are grateful for the award of CAS-TWAS president fellowship PhD program.
Conflict of interest: Authors declared no conflict of interest.
References
Acevedo, C. A., Lopez, D. A., Tapia, M. J., Enrione, J., Skurtys, O., Pedreschi, F., Brown, D. I.,
Creixell, W., & Osorio, F. (2012). Using RGB image processing for designating an
alginate edible film. Food and Bioprocess Technology, 5, 1511–1520.
Atress, A. S. H., El-Mogy, M. M., Aboul-Anean, H. E., & Alsaniu, B. W. (2010). Improving
strawberry fruit storability by edible coating as a carrier of thymol or calcium chloride.
Journal of Horticultural Science and Ornamental Plants, 2, 88–97.
Babalar, M., Asghari, M., Talaei, A., & Khosroshahi, A. (2007). Effect of pre and postharvest
salicylic acid treatment on ethylene production, fungal decay and overall quality of
‘Selva’ strawberry fruit. Food Chemistry, 105, 449-453.
Battacharyya, D., Babgohari, M. Z., Rathor, P., & Prithiviraj, B. (2015). Seaweed extracts as
biostimulants in horticulture. Scientia Horticulturae, 196, 39-48.
Bray, H. C., & Thorpe, W. (1954). Analysis of phenolic compounds of interest in metabolism.
Methods of Biochemical Analysis, 1, 27–52.
Brummell, D. A. (2006). Cell wall disassembly in ripening fruit. Functional Plant Biology, 33,
103-119.

24
Chai, Y. M., Jia, H. F., Li, C. L., Dong, Q. H., & Shen, Y. Y. (2011). FaPYR1 is involved in
strawberry fruit ripening. Journal of Experimental Botany, 62, 5079–5089.
Chen, J., Liu, D., Jiang, Y., Zhao, M., Shan, W., Kuang, J., & Lu, W. (2011). Molecular
characterization of a strawberry FaASR gene in relation to fruit ripening. PLoSOne, 6,
e24649.
d’Amour, J., Gosselin, C., Arul, J., Castaigne, F., & Willemot, C. (1993). Gamma-radiation
affects cell wall composition of strawberries. Journal of Food Science, 58, 182–185.
Duan, J., Wu, R., Strik, B. C., & Zhao, Y. (2011). Effects of edible coatings on the quality of
fresh blueberries (Duke and Elliott) under commercial storage conditions. Postharvest
Biology and Technology, 59, 71–79.
Falcon A. B., Cabrera J. C., Costales D., Ramirez M. A., Cabrera G., Toledo V., et al.
(2008). The effect of size and acetylation degree of chitosan derivatives on tobacco plant
protection against Phytophthora parasitica nicotianae. World Journal of Microbiology.
Biotechnology, 24, 103–112.
Geransayeh, M., Sepahvand, S., & Abdossi, V. (2015). Extending postharvest longevity and
improving quality of strawberry (Fragaria ananasa Duch cv. Gaviota) fruit by
postharvest salicylic acid treatment. Journal of Agricultural Studies, 3(2), 17-36.
Jayaraman, J. (1981). Laboratory Manual in Biochemistry. Wiley Eastern Ltd. New Delhi, India.
Ji, K., Chen, P., Sun, L., Wang, Y., Dai, S., & Li, Q., Li, P., Sun, Y., Wu, Y., Duan, C., & Leng,
P. (2012). Non-climacteric ripening in strawberry fruit is linked to ABA, FaNCED2 and
FaCYP707A1. Functional Plant Biology, 39, 351-357.

25
Jia, H. F., Chai, Y. M., Li, C. L., Lu, D., Luo, J. J., Qin, L., & Shen, Y. Y. (2011). Abscisic acid
plays an important role in the regulation of strawberry fruit ripening. Plant Physiology,
157, 188–199.
Jia, H. F., Lu, D., Sun, J. H., Li, C. L., Xing, Y., Qin, L., & Shen, Y.Y. (2013a). Type 2C
protein phosphatase ABI1 is a negative regulator of strawberry fruit ripening. Journal of
Experimental Botany, 64, 1677–1687.
Jiang, Y. M., & Joyce, D. C. (2003). ABA effects on ethylene production, PAL activity,
anthocyanin and phenolic contents of strawberry fruit. Journal of Plant Growth
Regulator, 39, 171-174.
Kafkas, E., Kosar, M., Paydas, S., Kafkas S., & Baser., K. H. C. (2007). Quality characteristics
of strawberry genotypes at different maturationstages. Food Chemistry, 100, 1229-1236.
Khan, A. & Singh, Z. (2007). 1-MCP regulates ethylene biosynthesis and fruit softening during
ripening of 'Tegan Blue' plum. Postharvest Biology and Technology, 43(3), 298-306.
Lee, K. H., Piao H. L., Kim, H. Y., Choi, S. M., Jiang, F., Hartung, W., Hwang, I., Kwak, J. M.,
Lee, I. J. & Hwang, J. (2006). Activation of glucosidase via stress-induced
polymerization rapidly increases active pools of abscisic acid. Cell, 126, 1109–1120.
Lees, D. H., & Francis, F. J. (1972). Standardization of pigment analysis in cranberries.
HortScience 7, 83–84.
Li, H., & Yu, T. (2000). Effect of chitosan on incidence of brown rot, quality and physiological
attributes of postharvest peach fruit. Journal of the Science of Food and Agriculture, 81,
269–274.
Livak, K. J., & Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-

26
time quantitative PCR and the 2(T)(-DeltaDelta C) method. Methods, 25, 402-408.
Ma, L., Cao, J., Xu, L., Zhang, X., Wang, Z., & Jiang, W. (2014). Effects of 1-
methylcyclopropene in combination with chitosan oligosaccharides on post-harvest
quality of aprium fruits. Scientia Horticulture, 179, 301-305.
Macheix, J. J., Fleuriet, A., & Bilot, J. (1990). Fruit Phenolics. CRC Press, Inc., Florida.
Maftoonazad, N., Ramaswamy, H. S., & Marcotte, M. (2008). Shelf life extension of peaches
through sodium alginate and methyl cellulose edible coatings. International Journal of
Food Science and Technology, 43, 951-957.
Ortiz, A., Seymour, G. B., Tucker, G. A., & Lara, I. (2010). Cell wall disassembly during the
melting phase of softening in 'Snow Queen' nectarines. Postharvest Biology and
Technology, 58,88-92.
Pose, S., Paniagua, C., Matas, A. J., Gunning, A. P., Morris, V. J., Quesada, M. A., & Mercado,
J. A. (2018). A nanostructural view of the cell wall disassembly process during fruit
ripening and postharvest storage by atomic force microscopy. Trends in Food Science
and Technology, Doi: 10.1016/j.tifs.2018.02.011.
Ranganna, S. (1977). Manual of Analysis of Fruit and Vegetable products. Tata McGraw-Hill
Pub. Co. Ltd., New Delhi. P. 634.
Ren, J., Sun, L., Wang, C., Zhao, S., & Leng, P. (2011). Expression analysis of the cDNA for
magnesium chelatase H subunit (CHLH) during sweet cherry fruit ripening and under
stress conditions. Plant Growth Regulator, 63, 301–307.
Ricardo, A. A., Luane, B., Carolina, W. G., Rafael, M. E., Juliana, I., & Precila, Z. L. (2016).
Abscisic acid involvement on expression of related gene and phytochemicals during

27
ripening in strawberry fruit Fragaria × anonnasa cv. Camino Real.
ScientiaHorticulturae, 203, 178-184.
Sambrook, J., & Russell, D. W. (2001). Molecular Cloning: A Laboratory Manual. Cold Spring
Harbour Laboratory Press, New York.
Sun, L., Wang, Y. P., Chen, P., Ren, J., Ji, K., Li, Q., Li, P., Dai, S. J., & Leng, P. (2011).
Transcriptional regulation of SlPYL, SlPP2C, and SlSnRK2 gene families encoding ABA
signal core components during tomato fruit development and drought stress. Journal of
Experimental Botany, 62, 5659–5669.
Symons, G. M., Ghua, Y. J., Ross, J. J., Quittenden, L. J., Davies, N. W., & Reid, J. B. (2012).
Hormonal changes during non-climateric ripening in strawberry. Journal of Experimental
Botany, 63, 4741-4751.
Velickova, E., Winkelhausen, E., Kuzmanova, S., Alves, V. D., & Moldão-Martins, M. (2013).
Impact of chitosan-beeswax edible coatings on the quality of fresh strawberries (Fragaria
ananassa cv. Camarosa) under commercial storage conditions. LWT: Food Science and
Technology, 52, 80–92.
Vu, K. D., Hollingsworth, R. G., Leroux, E., Salmieri, S., & Lacroix, M. (2011). Development of
edible bioactive coating based on modified chitosan for increasing the shelf life of
strawberries. Food Research International, 44, 198–203.
Wang, Y., Guo, S., Tian, S., Zhang, J., Ren, Y., Sun, H., Gong, G., Zhang, H., & Xu, Y. (2017).
Abscisic acid pathway involved in the regulation of watermelon fruit ripening and quality
trait evolution. PloSOne 12, e0179944.

28
He, Y., Bose, S. K., Wang, W., Jia, X., Lu, H., & Yin, H. (2018). Pre-Harvest treatment of
chitosan oligosaccharides improved strawberry fruit quality. International Journal of
Molecular Sciences, 19, 2194.
Zhang, B., Wu, H., Yang, H., Xiang, X., Li, H., Deng, S. (2017). Cryoprotective roles of
trehaloseand alginate oligosaccharides during frozen storage of peeled shrimp
(Litopenaeus vannamei). Food Chemistry, 228, 257–264.
Zhang, S. H., Sun, J. H., Dong,Y. H., Shen, Y. Y., Li,C. L., Li Y. Z., & Guo, J. X. (2014).
Enzymatic and functional analysis of β-glucosidase FaBG1 during strawberry fruit
ripening. The Journal of Horticultural Science and Biotechnology, 89(6), 733-739.
Zhang, S., Tang, W., Jiang, L., Hou,Y., Yang, F., Chen, W., & Li, X. (2015). Elicitor activity of
algino-oligosaccharide and its potential applicationin protection of rice plant
(Oryzasativa L.) against Magnaporthe grisea. Biotechnology and Biotechnological
Equipment, 29(4), 646-652,
Zhishen, J., Mengcheng, T., & Jianming, W. (1999). The determination of flavonoid contents in
mulberry and their scavenging effects on superoxide radicals. Food Chemistry, 64, 555–
559.
29
Fig.1. ESI-MS analysis showing different degree of polymerization (DP 2-7) of AOS
30
Fig.2 Effect of AOS postharvest treatments on (A) Hardness, (B) Weight loss (%), (C) Decay (%), (D) TSS (°Brix), (E) Total sugar (F) pH and (G) Titratable acidity during storage of strawberry. Data represent the means ±SD. The * and ** represent means significantly different according to T test at P<0.05 and P<0.01 level, respectively.
31
Fig.3 Effect of AOS postharvest treatments on (A) Vitamin C, (B) Anthocyanin, (C) Total phenol, (D) Flavonoid and (E) DPPH inhibition (%) during storage of strawberry. Data represent the means ±SD. The * and ** represent means significantly different according to T test at P<0.05 and P<0.01 level, respectively.
32
Fig.4 Effect of AOS postharvest treatments on cell wall degradation related gene expression during storage of strawberry. (A) FaPL: Pectin lyase, (B) FaPE: Pectin esterase, and (C) FaEG: Endoglucanase. Data represent the means ±SD. The * and ** represent means significantly different according to T test at P<0.05 and P<0.01 level, respectively.
33
Fig.5 Effect of AOS postharvest treatments on (A) Abscisic acid, (B) Abscisic acid conjugates content and abscisic acid (C). FaNCED1: Biosynthesis gene, (D). FaASR: Signal transduction, (E. FaPYR1 & F. FaCHLH: Perception) related gene expression during storage of strawberry. Data represent the means ±SD. The * and ** represent means significantly different according to T test at P<0.05 and P<0.01 level, respectively.
34
Table 1: Effect of AOS postharvest treatment on strawberry fruit cell wall composition
WSP
(mg.g-1 FW)
CSP
(mg.g-1 FW)
NSP
(mg.g-1 FW)
Hemicellulose
(mg.g-1 FW)
Cellulose
(mg.g-1 FW)
DAS
Untreated AOS Untreated AOS Untreated AOS Untreated AOS Untreated AOS0 0.17±0.00 0.17±0.00 0.58±0.00 0.58±0.00 0.60±0.00 0.60±0.00 2.64±0.00 2.6±0.00 1.74±0.00 1.74±0.00
2 0.26±0.03 0.20±0.02* 0.62±0.11 0.59±0.02NS 0.66±0.09 0.62±0.08NS 2.19±0.05 2.36±0.06* 1.56±0.09 1.59±0.12NS
4 0.46±0.16 0.32±0.02NS 0.69±0.04 0.63±0.05* 0.70±0.05 0.67±0.03NS 1.95±0.10 2.19±0.08* 1.42±0.08 1.46±0.03NS
6 0.48±0.05 0.43±0.09NS 0.73±0.06 0.70±0.04NS 0.80±0.08 0.74±0.10NS 1.80±0.03 1.90±0.13NS 1.30±0.03 1.37±0.10NS
Data represent the means ± SD. The *are significantly different according to T test at P≤0.05 level.(Here, WSP: Water soluble pectin; CSP: CDTA soluble pectin; NSP: Na2CO3 soluble pectin)
Highlights:1. Alginate Oligosaccharide (AOS) have positive effect on retaining fruit quality.2. AOS reduced cell wall degradation by delaying the softness.3. AOS delayed strawberry ripening by inhibiting ABA and ABA-conjugates accretion.

35