adaptive mechanisms that preserve cardiac function in mice without myoglobin
TRANSCRIPT

Adaptive Mechanisms That Preserve Cardiac Function inMice Without Myoglobin
Annette P. Meeson, Nina Radford, John M. Shelton, Pradeep P.A. Mammen, J. Michael DiMaio,Kelley Hutcheson, Yanfeng Kong, Joel Elterman, R. Sanders Williams, Daniel J. Garry
Abstract—Mice lacking myoglobin survive to adulthood and meet the circulatory demands of exercise and pregnancywithout cardiac decompensation. In the present study, we show that many myoglobin-deficient embryos die in utero atmidgestation with signs of cardiac failure. Fetal mice that survive to gestational day 12.5, however, suffer no subsequentexcess mortality. Survival in the absence of myoglobin is associated with increased vascularity and the induction ofgenes encoding the hypoxia-inducible transcription factors 1a and 2, stress proteins such as heat shock protein 27, andvascular endothelial growth factor. These adaptations are evident in late fetal life, persist into adulthood, and aresufficient to maintain normal myocardial oxygen consumption during stressed conditions. These data reveal thatmyoglobin is necessary to support cardiac function during development, but adaptive responses evoked in some animalscan fully compensate for the defect in cellular oxygen transport resulting from the loss of myoglobin.(Circ Res. 2001;88:713-720.)
Key Words: myoglobin n transgenic micen metabolismn hypoxia n vasculature
Myoglobin is a monomeric cytoplasmic hemoprotein thatis restricted to cardiomyocytes and oxidative skeletal
myofibers.1–3 Acute chemical inhibition of oxymyoglobinformation demonstrated that myoglobin is an essential proteinin the delivery of oxygen from the erythrocyte to mitochon-dria during periods of high metabolic demand.1–7 Surpris-ingly, we observed that mice with a complete deficiency ofmyoglobin are viable and have preserved cardiac and skeletalmuscle function over a wide range of oxygen levels.8 Fur-thermore, animals lacking myoglobin are fertile and capableof surviving the hemodynamic stress of pregnancy. We foundno evidence of spontaneous deaths or congestive heart failurewithin a large colony of adult mice (.100 animals) devoid ofmyoglobin.8
These observations suggested either that previous conceptsof myoglobin function in the heart are incorrect or that micedeveloping in the absence of myoglobin adapt to this defi-ciency in a manner that maintains respiratory function in theface of an otherwise catastrophic decline in oxygen transport.In the present article, we present new evidence to support thelatter viewpoint. In the absence of myoglobin, most embryossuccumb to heart failure at midgestation. Those embryos thatsurvive do so by mounting a complex compensatory responsethat involves extensive reprogramming of cardiac gene ex-pression and serves to maintain myocardial oxygen consump-tion at normal levels.
Materials and MethodsAnimalsHomozygous myoglobin-deficient (Mb–/–) mice were generated us-ing gene disruption technology as previously described.8 A completeabsence of myoglobin mRNA and protein was previously document-ed.8 Offspring of heterozygote intercrosses (Mb1/– 3 Mb1/–) fromtimed pregnancies were harvested at 3 periods during embryogenesis(gestational days 8 to 9 [E8 to E9], E9.5 to E10.5, and E11 to E12.5)and genotyped by Southern blot assays. Viable progeny from theheterozygote intercrosses were genotyped and analyzed as describedbelow.
Histological and Immunohistochemical Analysisand Vascular QuantitationAll embryos and embryonic tissues were staged, harvested, fixed in4% paraformaldehyde overnight, either paraffin-processed or quick-frozen in liquid nitrogen, and cryosectioned as previously described.9
The staging of embryos was performed by counting the presence ofthe vaginal plug as day 0.5 after conception and by the number ofsomites. Tissue for genotyping the embryos was obtained from theyolk sac. Immunohistochemistry was performed as previously de-scribed.2,9 The whole-mount immunohistochemical staining protocolincluded overnight fixation of the embryonic hearts in 4% parafor-maldehyde and overnight incubation with both the primary (rabbitanti-CD31 or platelet-endothelial cell adhesion molecule 1[PECAM-1]; 1:50, Pharmingen) and secondary antiserum (horserad-ish peroxidase–conjugated goat anti-rat serum; 1:100, Vector Labs).The embryonic hearts were then incubated with dimethylaminoazo-benzene/NiCl2 solution, postfixed overnight in 4% paraformalde-hyde/0.1% glutaraldehyde, and photographed using an OlympusSZH stereomicroscope.
Original received March 13, 2000; resubmission received January 16, 2001; revised resubmission received March 7, 2001; accepted March 7, 2001.From the Departments of Internal Medicine (A.P.M., N.R., J.M.S., P.P.A.M., Y.K., J.E., R.S.W., D.J.G.), Thoracic and Cardiovascular Surgery (J.M.D.,
K.H.), and Molecular Biology (R.S.W., D.J.G.), University of Texas Southwestern Medical Center, Dallas.Correspondence to Daniel J. Garry, UT Southwestern Medical Center at Dallas, 5323 Harry Hines Blvd., NB11.200, Dallas, TX 75390-8573. E-mail
[email protected]© 2001 American Heart Association, Inc.
Circulation Researchis available at http://www.circresaha.org
713
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2016http://circres.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2016http://circres.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2016http://circres.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2016http://circres.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2016http://circres.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from

In addition to whole-mount immunohistochemical staining withPECAM-1, we used a second technique to identify the endothelialcells of blood vessels in the developing and adult wild-type andmutant hearts. Embryonic (E12.5 to E16.5) or adult male Mb1/1 andMb–/– hearts were isolated, immersion-fixed overnight in methyl-Carnoy’s fixative, embedded in paraffin, sectioned, and stained usingthe biotinylated lectin Bandiera simplicifolia lectin B4 (10mg/mL;Vector labs).10 Substrate was developed with dimethylaminoazoben-zene, and slides were either cover-slipped or lightly counterstainedwith hematoxylin and analyzed microscopically (magnification of203 to 1003). For each embryo (n53 for each genotype), the entiresection was analyzed at 3 separate levels. For each adult animal, 30fields were examined at 3 separate levels (n53 for each genotype).The vasculature was quantified by 2 blinded investigators. Statisticalanalysis was performed with a Student’s pairedt test.
RNA Isolation and Reverse Transcription–Polymerase Chain ReactionTotal RNA was isolated from individually dissected embryonic oradult hearts or whole embryos using the Tripure isolation kit(Boehringer Mannheim). Four micrograms of total RNA were usedin each reverse transcription (RT) reaction (Retro-script, Ambion).Complementary DNA (2mL) was then used as a template for thepolymerase chain reaction (PCR) in a 20-mL reaction volumeincluding 100 ng of each primer, 2 mmol/L MgCl2, Taq buffer, and1 U of Taq polymerase (GIBCO/BRL). Fifteen microliters of eachPCR reaction were loaded on a 2% agarose gel, as previouslydescribed.9 For myoglobin, a single forward primer was positionedto include exon 1, and amplification by PCR (25 cycles) was doneusing reverse primers complementary to sequences from exon 2.Semiquantitative RT-PCR using RNA isolated from individuallydissected embryonic (E12.5) or adult (3-month-old) hearts wasperformed as previously described.9 For each primer pair, PCRconditions were used to establish linearity of the output signalrelative to the input template DNA.9 PCR primer pairs used for thisstudy are included in the online data supplement (located athttp://www.circresaha.org).
Image AnalysisStained sections were examined with a Leitz Laborlux-S microscopeequipped with an Optronics VI-470 CCD camera and Scion Image
1.62 analysis software. Image processing was completed usingAdobe Photoshop 5.0 and printed using a Kodak XLS 8600 PSprinter.
Hemodynamic Studies (WorkingHeart Preparation)Mice (25 to 30 g) were euthanized, and their thoracic contents wereexcised and placed in a 4°C perfusate. The heart and aorta weredissected free and perfused in the Langendorff mode at 37.5°C witha (retrograde) perfusion pressure of 100 cm H2O. The preparationwas then converted to a working mode with a left atrial pressure of15 cm H2O and an anterograde perfusion pressure of 80 cm H2O11,12
(online data supplement).
13C Metabolic StudiesHearts were perfused in the working mode as described above. Afterstabilization with a standard Krebs-Henseleit buffer, the buffer waschanged to the following mixture of physiological substrates: 3-13Clactate (1.2 mmol/L), 1,3-13C acetoacetic acid (0.17 mmol/L), 3-13Cpyruvate (0.12 mmol/L), BSA (0.75%), U-13C labeled fatty acids(0.5 mmol/L; consisting of 50% palmitic acid, 23% oleic acid, 15%linoleic acid, 11% palmitoleic acid, and 1% stearic acid), andunlabeled glucose (8 mmol/L).13 After 30 minutes of perfusion,hearts were freeze-clamped and extracted in perchloric acid, neutral-ized with KOH, reconstituted in D2O, and analyzed by nuclearmagnetic resonance (NMR) spectroscopy.14 Proton-decoupled13CNMR spectra were obtained at 150.87 MHz using a Varian Inovaspectrometer with a broadband probe, a 45-degree observe pulse, aninterpulse delay of 1.5 s, 43 488 data points, and 4000 scans.
Isoproterenol-Induced CardiacHypertrophic ChallengeVentricular hypertrophy was induced in age- and sex-matched adultMb1/1 and Mb–/– mice by continuous administration of isoproterenol(0.028 g/mL at a rate of 1.0mL/h), as described previously.15,16After7 days of continuous infusion, the mice were weighed and eutha-nized. The hearts were excised, weighed, and either fixed in 4%paraformaldehyde or frozen for isolation of total RNA.2 Cardiachypertrophy was determined by the comparison of heart weight tobody weight, histological analysis, and gene expression.
Ischemic Injury/Cardiomyopathic ChallengeAfter continuous anesthesia with 2.0% isoflurane, age- and sex-matched adult Mb1/1 and Mb–/– mice underwent a left thoracotomy/pericardiotomy to expose the left ventricle, and the left anteriordescending coronary artery was ligated with 8-0 prolene sutures. Theresidual pneumothorax was evacuated to restore negative pressure,and buprenorphine (0.05 mg/kg) was administered for postoperativepain control. Two months after ischemic injury, transthoracic echo-cardiography was performed under isoflurane anesthesia. The micewere weighed and euthanized, and their hearts were excised,weighed, fixed in 4% paraformaldehyde, and processed for histolog-ical analysis.
An expanded Materials and Methods section can be found in anonline data supplement available at http://www.circresaha.org.
ResultsMorphogical Phenotype of the MyoglobinMutant EmbryosAnalysis of a large group of embryos from 16 male and 16female heterozygote intercrosses at sequential stages of em-bryonic development demonstrated a Mendelian distributionof genotypes (1:2:1 ratio) only in embryos at E9.0 days oryounger (online data supplement). Between days E9.5 andE10.5, the majority of embryos homozygous for the mutatedmyoglobin allele (Mb–/–) exhibited a number of severe de-fects. These included congestive heart failure manifested by
Figure 1. Morphological appearance of Mb–/– embryos. Littersfrom myoglobin E9.5 to E10.5 heterozygote intercrosses werecollected and analyzed. The gross morphological appearance ofan E9.5 Mb–/– embryo (A) reveals a prominent pericardial effu-sion (arrow) compared with the normal appearance of a wild-type littermate (B). From a second E9.5 litter, the gross appear-ance of the Mb–/– embryo (C) revealed evidence of congestion(arrow) compared with the wild-type littermate (E). D and F arehistological sections of the Mb–/– and Mb1/1 embryos in C andE, respectively. Evidence of cardiac congestion and myocardialthinning (arrows mark myocardium) are observed in the heart ofthe Mb–/– embryo (D) compared with its wild-type littermate (F).Bar550 mm in D and F; bar5500 mm in A, B, C, and E.
714 Circulation Research April 13, 2001
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from

prominent pericardial effusion and vascular insufficiencymanifested by diffuse hemorrhages, a generalized develop-mental delay, and reduced size (Figure 1). Similar defectswere observed more rarely in animals bearing a single copy ofthe myoglobin-null allele (Mb1/–). Of the myoglobin-nullembryos that developed beyond this stage (E11.0 days orolder), there was no evidence of further fetal loss or structuralabnormalities (online data supplement). These findings werenoted in 2 strains of mice (C57BL/6 and Sv 129).
To further define the morphological defects associatedwith reduced expression of myoglobin, histological sectionsof Mb1/1, Mb1/– (data not shown), and Mb–/– embryos werecompared. Histological analysis of runted mutant embryosrevealed evidence of myocardial thinning, congestive heartfailure, and vascular insufficiency with peripheral hemor-rhage (Figure 1). Other than a generalized developmentaldelay, morphological defects in Mb–/– embryos were limitedto the cardiovascular system, and other major developmentalevents were not noted until the time of death. Among the 12failing Mb–/– embryos at E9.5 to E10.5, 4 had a severepericardial effusion consistent with congestive heart failureand 8 had extensive hemorrhages. Ultrastructural analysis ofphenotypically abnormal but viable Mb–/– hearts at E10.5revealed evidence of edema in cells of both the endocardialand myocardial layers, increased cytoplasmic vacuoles, and aless well-developed sarcomeric myosin actin contractile ap-paratus compared with normal wild-type littermates (data notshown). The TdT-mediated dUTP nick-end labeling assayshowed no evidence of an apoptotic process associated withthe hearts of failing Mb–/– embryos (data not shown), which isconsistent with an oncotic process associated with the demiseof the mutant embryos.
Increased Vascularity in Surviving Mb–/– Embryosand Adult MiceHearts were dissected free from Mb–/– and Mb1/1 embryosthat survived to E12.5. Immunostaining to detect PECAM-1,a marker for endothelial cells, demonstrated hypervascularityin the viable Mb–/– heart (n53) compared with wild-type(Mb1/1) hearts (n53) (Figure 2). These results were con-firmed using a specific lectin that identifies the endothelialcells. Further, we observed that the formation of myocardialvascularization was established earlier during cardiac devel-opment in the absence of myoglobin (Figure 2). There was a48% increase in capillary density in the ventricles of Mb–/–
mice at E13.5 (Figure 2).Adult Mb–/– mice also exhibited an increased vascular supply
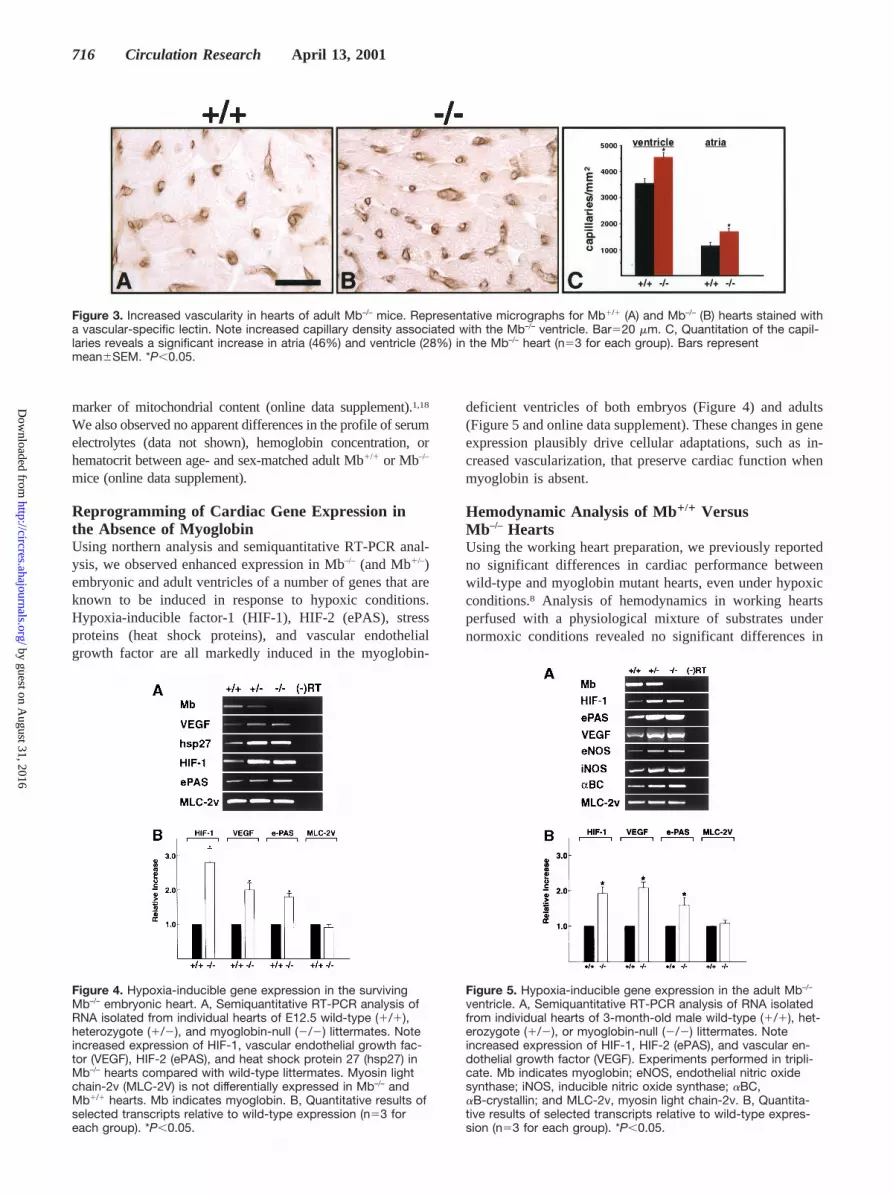
in the atria (46% increase) and ventricles (28% increase)compared with wild-type controls (Figure 3). These findings arein agreement with those of Godecke et al,17 who used ultrastruc-tural morphometric techniques and reported a 31% increase incapillaries in the myoglobin mutant ventricle. There were noapparent differences in the ultrastructural morphology or thehistochemical staining of succinate dehydrogenase activity, a
Figure 2. Increased vascularity in hearts of viable Mb–/– embryos.Individually dissected embryonic hearts from genotyped E12.5 lit-termates were immunostained with anti-CD31 (PECAM-1) serum.Note an increase in the vascular network in the mutant (B) com-pared with the wild-type (A) embryonic heart (n53). Representativemicrographs for E12.5 Mb1/1 (C, E) and Mb–/– (D, F) hearts andE13.5 Mb1/1 (G, I) and Mb–/– (H, J) hearts were stained with avascular-specific lectin. Note increased capillary density associatedwith the Mb–/– ventricular septum at E12.5 (F) and E13.5 (J). K,Quantitation of the capillaries reveals a 48% increase in E13.5
Mb–/– hearts (561610 capillaries per mm2; n53) compared withthe age-matched Mb1/1 hearts (380611 capillaries per mm2;n53). Bars represent mean6SEM. *P,0.05. Bars5400 mm in A,B, C, D, G, and H; bars5200 mm in E, F, I, and J.
Meeson et al Adaptations in Myoglobin Knockout Mice 715
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from
marker of mitochondrial content (online data supplement).1,18
We also observed no apparent differences in the profile of serumelectrolytes (data not shown), hemoglobin concentration, orhematocrit between age- and sex-matched adult Mb1/1 or Mb–/–
mice (online data supplement).
Reprogramming of Cardiac Gene Expression inthe Absence of MyoglobinUsing northern analysis and semiquantitative RT-PCR anal-ysis, we observed enhanced expression in Mb–/– (and Mb1/–)embryonic and adult ventricles of a number of genes that areknown to be induced in response to hypoxic conditions.Hypoxia-inducible factor-1 (HIF-1), HIF-2 (ePAS), stressproteins (heat shock proteins), and vascular endothelialgrowth factor are all markedly induced in the myoglobin-
deficient ventricles of both embryos (Figure 4) and adults(Figure 5 and online data supplement). These changes in geneexpression plausibly drive cellular adaptations, such as in-creased vascularization, that preserve cardiac function whenmyoglobin is absent.
Hemodynamic Analysis of Mb1/1 VersusMb–/– HeartsUsing the working heart preparation, we previously reportedno significant differences in cardiac performance betweenwild-type and myoglobin mutant hearts, even under hypoxicconditions.8 Analysis of hemodynamics in working heartsperfused with a physiological mixture of substrates undernormoxic conditions revealed no significant differences in
Figure 3. Increased vascularity in hearts of adult Mb–/– mice. Representative micrographs for Mb1/1 (A) and Mb–/– (B) hearts stained witha vascular-specific lectin. Note increased capillary density associated with the Mb–/– ventricle. Bar520 mm. C, Quantitation of the capil-laries reveals a significant increase in atria (46%) and ventricle (28%) in the Mb–/– heart (n53 for each group). Bars representmean6SEM. *P,0.05.
Figure 4. Hypoxia-inducible gene expression in the survivingMb–/– embryonic heart. A, Semiquantitative RT-PCR analysis ofRNA isolated from individual hearts of E12.5 wild-type (1/1),heterozygote (1/2), and myoglobin-null (2/2) littermates. Noteincreased expression of HIF-1, vascular endothelial growth fac-tor (VEGF), HIF-2 (ePAS), and heat shock protein 27 (hsp27) inMb–/– hearts compared with wild-type littermates. Myosin lightchain-2v (MLC-2V) is not differentially expressed in Mb–/– andMb1/1 hearts. Mb indicates myoglobin. B, Quantitative results ofselected transcripts relative to wild-type expression (n53 foreach group). *P,0.05.
Figure 5. Hypoxia-inducible gene expression in the adult Mb–/–
ventricle. A, Semiquantitative RT-PCR analysis of RNA isolatedfrom individual hearts of 3-month-old male wild-type (1/1), het-erozygote (1/2), or myoglobin-null (2/2) littermates. Noteincreased expression of HIF-1, HIF-2 (ePAS), and vascular en-dothelial growth factor (VEGF). Experiments performed in tripli-cate. Mb indicates myoglobin; eNOS, endothelial nitric oxidesynthase; iNOS, inducible nitric oxide synthase; aBC,aB-crystallin; and MLC-2v, myosin light chain-2v. B, Quantita-tive results of selected transcripts relative to wild-type expres-sion (n53 for each group). *P,0.05.
716 Circulation Research April 13, 2001
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from

coronary flow, cardiac output, or oxygen consumption inMb–/– and Mb1/1 hearts (online data supplement and the Tableabove). The results obtained in the present study agree closelywith the hemodynamic measurements obtained by Grupp etal12 using the working heart preparation in wild-type mice.
Myocardial oxygen consumption has a number of determi-nants, including heart rate, contractility, systolic wall tension,and substrate utilization.1,13,14 Under physiological condi-tions, the adult heart generates energy or ATP primarily fromthe oxidative phosphorylation of fatty acids as the preferredsubstrate. To define the preferred myocardial substrate foroxidative phosphorylation in the absence of myoglobin,working heart preparations were perfused with13C-labeledsubstrates and then analyzed using NMR spectroscopy.13,14
We found that fatty acids were the preferred substrate in bothMb–/– and wild-type hearts. The relative use of fatty acids andacetoacetate for the production of acetyl-coenzyme A unitsfeeding into the Krebs cycle was similar in the 2 groups, butthere was a modestly greater use of13C-labeled lactate in thehearts from Mb–/– animals (Table). The use of these13C-labeled substrates in the present study agrees with previouslypublished results using this technique in the rat heart.14
Working heart preparations were then challenged by theadministration of theb-adrenergic agonist isoproterenol.Despite a 20% increase in heart rate, no differences wereobserved between Mb1/1 and Mb–/– hearts with respect tomyocardial oxygen consumption in response to isoproter-enol stimulation (Figure 6). These results support theconclusion that cardiac function in response to short-termadrenergic stimulation is preserved in the absence ofmyoglobin.
Isoproterenol-Induced Cardiac HypertrophyTo assess the response to long-term adrenergic stimulation inthis knockout model, ventricular hypertrophy was induced bya 7-day continuous administration of isoproterenol.15,16Usingthis well-characterized model of cardiac hypertrophy, wedetermined the functional and structural adaptations in re-sponse to adrenergic stimulation in mice that lacked myoglo-bin. No significant differences in heart weight were notedbetween age- and sex-matched wild-type and myoglobinmutant mice. Knockout and wild-type mice that were sub-jected to long-term isoproterenol infusion had a 34%(P,0.005) and 22% (P,0.05) increase, respectively, in theheart weight to body weight ratio (online data supplement)and an induction of atrial natriuretic factor expression in theventricle compared with their respective controls (Figure 6).
There was no evidence of congestive heart failure in eitherMb1/1 or Mb–/– mice in response to this stimulus. Histologicalanalysis (Figure 6) after long-term isoproterenol infusionrevealed the presence of focal regions of myocardial fibrosis,
Figure 6. Analysis of cardiac function in response to acute andchronic stimulation of isoproterenol. Using the working heart prep-aration, no differences were noted in oxygen consumption (MV̇O2;A) or heart rate (B) under baseline conditions (Pre) or after stimula-tion with isoproterenol (1310–6 mol/L; Post) between Mb1/1 (n55)and Mb–/– (n56) hearts. Histograms depict mean6SEM. Followinga 7-day continuous infusion of isoproterenol in adult Mb1/1 (E) andMb–/– (F) mice, the hearts were excised and noted to be increasedin size compared to the wild-type (C) and knockout (D) controls.No significant histological differences were noted between chroni-cally stimulated Mb1/1 (G) and Mb–/– (H) hearts using a Massontrichrome stain. I, Northern blot analysis was undertaken usingtotal RNA isolated from the ventricles of Mb1/1 and Mb–/– mice inthe absence (2) of or following a 7-day continuous infusion of iso-proterenol (1). There was a similar induction of atrial natriureticfactor (ANF) expression in wild-type and Mb–/– ventricles after iso-proterenol stimulation. Mb indicates myoglobin; GAPDH, glyceral-dehyde phosphate dehydrogenase. Bar52.5 mm in C, D, E and F;bar550 mm in G and H.
Oxygen Consumption and Substrate Use in Mb1/1 andMb2/2 Hearts
Mb1/1 MB2/2
n 4 5
M.VO2, mmol z min21 z g wet weight21 1.6560.17 1.4460.07
FA utilization, % 59.067 54.062.4
Lactate utilization, % 4.861.3* 14.762.7*
Acetoacetate utilization, % 22.164.8 19.963.5
Values are mean6SEM. FA indicates fatty acid.*P,0.05
Meeson et al Adaptations in Myoglobin Knockout Mice 717
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from

but the frequency of such abnormalities did not seem to differbetween Mb1/1 and Mb–/– animals.
Ischemic Injury and CardiomyopathyAs a further challenge, ischemic injury was induced surgi-cally in wild-type and myoglobin mutant mice. We observedno differences in survival during the perioperative period orduring a 2-month survival period between wild-type orknockout mice. Furthermore, 2 months after the ischemicinsult, wild-type and Mb–/– mice had comparable increases inheart size and decreases in left ventricular function (Figure 7).Using transthoracic echocardiography, fractional shorteningwas depressed in wild-type (wild-type control [n55],0.6060.02; wild-type with coronary ligation [n53],0.3460.05;P,0.001) and myoglobin mutant (Mb–/– control[n55], 0.5960.03; Mb–/– with coronary ligation [n53],0.3460.05;P,0.001] mice after coronary artery ligation.Although myoglobin-deficient mice survived this ischemicchallenge, these data do not exclude the possibility thatmyoglobin may function in oxygen metabolism during theearly phase(s) of ischemic injury.
DiscussionWe observed that a majority of embryos lacking myoglobindie, apparently of congestive heart failure and vascularinsufficiency, within the brief period (E9.5 to E10.5) ofgestation that corresponds to the initial expression of myo-globin in wild-type embryos. Increased lethality was alsoobserved in embryos (E9.5 to E10.5) who were haploinsuf-ficient for myoglobin, suggesting that myoglobin content wasimportant for viability during this critical period of cardio-vascular development. However, a proportion of embryoswithout myoglobin survive past this critical developmentalstage, and surviving animals exhibit grossly normal cardio-vascular function as adults.8,17
These new data establish that myoglobin is importantduring cardiac development at a stage (E9.5 to 10.5 days)associated with the onset of rhythmic cardiac contractionsand increasing metabolic demands that precedes the estab-lishment of a functional coronary vasculature.19–22 It islogical to surmise that the rapid growth of the heart thatoccurs during this period of development, particularly theexpansion of the ventricular wall, increases the distanceoxygen must diffuse from the ventricular cavity to reach cellscloser to the epicardial surface of the heart.22 The establish-ment of the coronary vasculature is ultimately required tosupply oxygen to the expanding myocardium, and even subtledifferences in the timing of vasculogenesis and cardiacgrowth may be sufficient to increase the dependence ofworking myocytes on myoglobin to facilitate oxygen deliv-ery. Remarkably, the requirement for myoglobin at this stage
Figure 7. Morphological assessment of nonligated and ligatedleft anterior descending coronary arteries in Mb1/1 and Mb–/–
hearts. After a 2-month period, echocardiography was per-formed and mice were euthanized. A, Both wild-type and myo-
globin knockout hearts were larger 2 months after ischemicinjury compared with their respective controls. No significanthistological differences were noted between chronically ligatedMb1/1 and Mb–/– hearts using a Masson trichrome stain (datanot shown). B and C, Echocardiography revealed left ventriculardysfunction in both wild-type and knockout hearts 2 monthsafter ischemic injury. *P,0.001.
718 Circulation Research April 13, 2001
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from

of development is not absolute, and some animals mount anadaptive response that sustains myocardial function.
Our current findings suggest that the ability to survivewithout myoglobin is based on increased vasculogenesis,driven by intracellular hypoxia, and ultimately establishes anew steady-state in which oxygen transfer is preserved in theabsence of myoglobin. This conclusion is supported by theincreased expression of hypoxia-responsive genes, includingthose encoding angiogenic growth factors, and morphologicalevidence of hypervascularity in Mb–/– embryos that survivepast E12.5 and into adult life.
The observation that embryos surviving beyond E12.5suffer little or no subsequent mortality and maintain relativelynormal circulatory function as adults is consistent with eitherof two conclusions with respect to the importance of myo-globin in the adult heart. Fetal death at E9.5 to E10.5 mayresult from a developmental bottleneck at this stage that isbased on a critical requirement for myoglobin unique to theperiod that precedes the development of a coronary circula-tion,19–22 and myoglobin may have lesser physiological im-portance at later stages. Alternatively, the ability to survivethe fetal stage may select for those animals that establish andmaintain powerful adaptive mechanisms that remain active asthe heart matures in postnatal life. Our data argue in favor ofthe latter viewpoint.
In adult mice, we showed that cardiac function is preservedunder baseline conditions and in response to adrenergicstimulation in mice without myoglobin. Furthermore,myoglobin-deficient mice survived ischemic injury and suf-fered left ventricular dysfunction comparable to that ofwild-type animals exposed to the same coronary ligationprotocol. We observed no abnormalities with respect to serumelectrolytes, hemoglobin concentration, or hematocrit inmyoglobin-mutant mice. A comprehensive hemodynamicanalysis showed normal cardiac output, stroke volume, heartrate, coronary flow, and myocardial oxygen consumption inMb–/– hearts. Substrate utilization was relatively preserved inthe absence of myoglobin. Although fatty acids remained thepreferred metabolic substrate, there was a modest increase inlactate utilization in the myoglobin mutant heart. Futurestudies will further examine substrate utilization in themyoglobin-deficient heart in response to fatiguing exerciseand environmentally challenging conditions.
In contrast to knockout mice that lack either desmin23 orcreatine phosphokinase,24,25 we observed no pathologicalchanges in mitochondria or the sarcomeric ultrastructure ofcardiomyocytes from mice that lack myoglobin. Furthermore,myoglobin mutant mice are capable of developing cardiachypertrophy, induced by long-termb-adrenergic stimulation,without apparent hemodynamic compromise.
Our current findings also indicate that survival in theabsence of myoglobin is attributable to reprogramming geneexpression within the myocardium and compensatory cellularresponses that include increased vasculogenesis. Intracellularhypoxia is likely to be an inciting stimulus to these adapta-tions, through mechanisms that include the induction ofHIF-1, a basic-helix-loop-helix-PAS protein.26 Target genesinduced by HIF-1 increase oxygen delivery (eg, angiogenicgrowth factors and nitric oxide synthase) or facilitate ATP
production in the absence of oxygen (eg, glucose transportersand glycolytic enzymes).26,27 Disruption of the HIF-1 generesults in embryonic lethality at midgestation in associationwith developmental arrest and cardiovascular malforma-tions.28,29Even haploinsufficiency at the HIF-1 locus impairscardiovascular responses to long-term hypoxia.30 These re-sults collectively support an essential role for HIF-1 as amaster regulator of oxygen homeostasis.
Other results of the present study are largely in agreementwith those of Godecke et al,17 who confirmed our initialobservation of preserved cardiac function in mice withoutmyoglobin. They observed increased vascular density in thehearts of Mb–/– mice. Our present report provides the firstdescription of fetal death in embryos without myoglobin andis the first to define specific changes in gene expression thatare likely to drive the adaptive changes in vascular supply.
In summary, we demonstrate that myoglobin plays animportant role during cardiac development. In the absence ofmyoglobin, heart failure and circulatory insufficiency lead todeath in the majority of embryos. This requirement formyoglobin is not absolute, and a minority of animals mountan adaptive response that reprograms gene expression andincreases myocardial vascularity; this adaption is sufficient tomaintain oxygen transfer to cardiomyocytes and to preservecirculatory function. A more complete definition of thecellular, physiological, and molecular mechanisms of thispowerful adaptive response could conceivably lead to ad-vances in therapy for patients with ischemic heart disease.
AcknowledgmentsThis work was supported by grants from the Texas Affiliate of theAmerican Heart Association, the National Institutes of Health (AR40849,HL54794, and HL06296), and the D.W. Reynolds Foundation.
References1. Wittenberg BA, Wittenberg JB. Transport of oxygen in muscle.Ann Rev
Physiol. 1989;51:857–878.2. Garry DJ, Bassel-Duby RS, Richardson JA, Grayson J, Neuffer PD,
Williams RS. Postnatal development and plasticity of specialized musclefiber characteristics in the hindlimb.Dev Genet. 1996;19:146–156.
3. Parsons WJ, Richardson JA, Graves KH, Williams RS, Moreadith RW.Gradients of transgene expression directed by the human myoglobinpromoter in the developing mouse heart.Proc Natl Acad Sci U S A.1993;90:1726–1730.
4. Hochachka PW. The metabolic implications of intracellular circulation.Proc Natl Acad Sci U S A. 1999;96:22:12233–12239.
5. Kreutzer U, Jue T. Critical intracellular O2 in myocardium as determinedby1H nuclear magnetic resonance signal of myoglobin.Am J Physiol.1995;268:H1675–H1681.
6. Doeller JE, Wittenberg BA. Myoglobin function and energy metabolismof isolated cardiac myocytes: effect of sodium nitrite.Am J Physiol.1991;261:H53–H62.
7. Conley KE, Jones C. Myoglobin content and oxygen diffusion: modelanalysis of horse and steer muscle.Am J Physiol. 1996;271:C2027–C2036.
8. Garry DJ, Ordway GA, Lorenz JN, Radford NB, Chin ER, Grange RW,Bassel-Duby RS, Williams RS. Mice without myoglobin.Nature. 1998;395:905–908.
9. Garry DJ, Yang Q, Bassel-Duby R, Williams RS. Persistent expression ofMNF identifies myogenic stem cells in postnatal muscles.Dev Biol.1997;188:280–294.
10. Coffin JD, Harrison J, Schwartz S, Heimark R. Angioblast differentiationand morphogenesis of the vascular endothelium in the mouse embryo.Dev Biol. 1991;148:51–62.
Meeson et al Adaptations in Myoglobin Knockout Mice 719
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from

11. Bittner HB, Chen EP, Peterseim DS, Van Trigt P. A work-performingheart preparation for myocardial performance analysis in murine hearts.J Surg Res. 1996;64:57–62.
12. Grupp IL, Subramaniam A, Hewett TE, Robbins J, Grupp G. Comparisonof normal, hypodynamic, and hyperdynamic mouse hearts using isolatedwork-performing heart preparations.Am J Physiol. 1993;265:H1401–H1410.
13. Malloy CR, Sherry AD, Jeffrey MH. Analysis of tricarboxylic acid cycleof the heart using 13C isotope isomers.Am J Physiol. 1990;259:H987–H995.
14. Malloy CR, Sherry AD, Jeffrey FMH. Evaluation of carbon flux andsubstrate selection through alternate pathways involving the citric acidcycle of the heart by 13C NMR spectroscopy.J Biol Chem. 1988;263:6964–6971.
15. Soonpaa MH, Field LJ. Assessment of cardiomyocyte DNA synthesisduring hypertrophy in adult mice.Am J Physiol. 1994;266:H1439–H1445.
16. Kim KK, Soonpaa MH, Daud AI, Koh GY, Kim JS, Field LJ. Tumorsuppressor gene expression during normal and pathologic myocardialgrowth.J Biol Chem. 1994;269:36:22607–22613.
17. Godecke A, Flogel U, Zanger K, Ding Z, Hirchenhain J, Decking U,Schrader J. Disruption of myoglobin in mice induces multiple compen-satory mechanisms.Proc Natl Acad Sci U S A. 1999;96:10495–10500.
18. Chalmers GR, Roy RR, Edgerton R. Motoneuron and muscle fiber suc-cinate dehydrogenase activity in control and overloaded plantaris.J ApplPhysiol. 1991;71:4:1589–1592.
19. Keller BB, MacLennan M, Tinney J, Yoshigi M. In vivo assessment ofembryonic cardiovascular dimensions and function in day-10.5 to 14.5mouse embryos.Circ Res. 1996;79:247–255.
20. Raddatz E, Servin M, Kucera P. Oxygen uptake during early cardio-genesis of the chick.Am J Physiol. 1992;31:H1224–H1230.
21. Loiselle DS, Gibbs CL. Species differences in cardiac energetics.Am JPhysiol. 1979;237:H90–H98.
22. Maltepe E, Simor MC. Oxygen, genes, and development: an analysis ofthe role of hypoxic gene regulation during murine vascular development.J Mol Med. 1998;76:391–401.
23. Li Z, Colucci-Guyon E, Pincon-Raymond M, Mericskay M, Pournin S,Paulin D, Babinet C. Cardiovascular lesions and skeletal myopathy inmice lacking desmin.Dev Biol. 1996;175:362–366.
24. Venturaclapier R, Kuznetsov AV, Dalbis A, Vandeursen J, Wieringa B,Veksler VI. Muscle creatine kinase-deficient mice.J Biol Chem. 1995;270:34:19914–19920.
25. Saupe KW, Spindler M, Tian R, Ingwall JS. Impaired cardiac energeticsin mice lacking muscle-specific isoenzymes of creatine kinase.Circ Res.1998;82:898–907.
26. Semenza GL. Perspectives on oxygen sensing.Cell. 1999;98:281–284.27. Semenza GL, Jiang BH, Leung SW, Passantino R, Concordet JP, Maire
P, Giallongo A. Hypoxia response elements in the aldolase A, enolase 1and lactate dehydrogenase A gene promoters contain essential bindingsites for hypoxia-inducible factor 1.J Biol Chem. 1996;271:51:32529–32537.
28. Ryan HE, Lo J, Johnson RS. HIF-1a is required for solid tumor formationand embryonic vascularization.EMBO J. 1998;17:11:3005–3015.
29. Iyer NV, Kotch LE, Agani F, Leung SW, Laughner E, Wenger RH,Gassmann M, Gearhart JD, Lawler AM, Yu AY, Semenza GL. Cellularand developmental control of O2 homeostasis by hypoxia-induciblefactor 1a. Genes Dev. 1998;12:149–162.
30. Yu AY, Shimoda LA, Iyer NV, Huso DL, Sun X, McWilliams R, BeatyT, Sham JSK, Wiener CM, Sylvester JT, Semenza GL. Impaired physi-ological responses to chronic hypoxia in mice partially deficient forhypoxia-inducible factor1a. J Clin Invest. 1999;103:5:691–696.
720 Circulation Research April 13, 2001
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from

GarryDiMaio, Kelley Hutcheson, Yanfeng Kong, Joel Elterman, R. Sanders Williams and Daniel J.
Annette P. Meeson, Nina Radford, John M. Shelton, Pradeep P. A. Mammen, J. MichaelAdaptive Mechanisms That Preserve Cardiac Function in Mice Without Myoglobin
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2001 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/hh0701.0897532001;88:713-720; originally published online March 30, 2001;Circ Res.
http://circres.ahajournals.org/content/88/7/713World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2001/04/03/hh0701.089753.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on August 31, 2016
http://circres.ahajournals.org/D
ownloaded from








