l-arginine depletion in preeclampsia orients nitric oxide synthase toward oxidant species
TRANSCRIPT

L-Arginine Depletion in Preeclampsia Orients Nitric OxideSynthase Toward Oxidant Species
Marina Noris, Marta Todeschini, Paola Cassis, Fabio Pasta, Anna Cappellini, Samantha Bonazzola,Daniela Macconi, Raffaella Maucci, Francesca Porrati, Ariela Benigni,
Claudio Picciolo, Giuseppe Remuzzi
Abstract—Less nitric oxide (NO)-dependent vasodilation and excess formation of reactive oxygen species could explainpoor placenta perfusion in preeclampsia, but the pathways involved are unknown. We tested the hypothesis that reducedNO activity and increased oxidative stress in preeclamptic placenta is related to a low bioavailability of L-arginine.Placental endothelial NO synthase (ecNOS) expression (by immunoperoxidase) and activity (by diaphorase and[3H]L-citrulline formation) were comparable in normotensive pregnancy and in preeclampsia, whereas nitrotyrosinestaining, a marker of peroxynitrite, was stronger in preeclamptic villi, confirming previously reported data. Oxidativetissue damage was documented in preeclamptic villi by strong 4-hydroxynonenal-lysine staining (by immunoperoxi-dase), which closely colocalized with nitrotyrosine. Concentration of the NO precursor L-arginine (by HPLC) inumbilical blood and in villous tissue was lower in preeclampsia than in normotensive pregnancy. This was not causedby a defective L-arginine transport, because gene expression of the CAT-1, 4F2hc, and LAT-1 cationic amino acidtransporters (by real-time reverse-transcription polymerase chain reaction [RT-PCR]) was normal. Instead, geneexpression (by real-time RT-PCR) and protein tissue content (by immunoperoxidase and Western blot) of arginaseII—the enzyme that degrades arginine to ornithine—were higher in preeclamptic villi than in normotensive pregnancy.These results provide a biochemical explanation for defective NO activity and increased oxidative stress in preeclampticplacenta. In normal placenta, adequate concentration of L-arginine orients ecNOS toward NO. In preeclampsia, a lowerthan normal L-arginine concentration caused by arginase II overexpression redirects ecNOS toward peroxynitrite.(Hypertension. 2004;43:614-622.)
Key Words: arginine � nitric oxide synthase � nitrites � oxidative stress � preeclampsia
In the past few years, evidence has accumulated suggestingthat nitric oxide (NO), a potent endothelial-derived vaso-
dilator, might be implicated in gestational vasodilation.1,2 NOis synthesized from the amino acid L-arginine by a family ofenzymes, the NO synthases (NOS).3 While human placentadoes not form inducible NOS (iNOS) or neuronal NOS(nNOS) mRNAs and proteins,1 the endothelial isoform ofNOS (ecNOS) is actually detectable in healthy placenta and islocalized to the endothelium of the umbilical cord, chorionicplate, and stem villous vessels.1 Conceivably, locally formedNO serves to maintain low vascular resistance besides atten-uating the action of vasoconstrictors.1 Besides, ecNOS islocalized in villous cytotrophoblasts,1,4 the specialized epi-thelial cells that aggregate into cell columns and invade theuterine interstitium and vasculature, anchoring the fetus to themother and establishing blood flow to the placenta.5 By virtueof its unique angiogenic/vasculogenic properties,6,7 locallygenerated NO may be instrumental to promote this cytotro-
phoblast endovascular invasion that is an essential feature ofnormal placentation,5 although whether extravillous invadingtrophoblasts express ecNOS is still a controversial issue.8,9
Unlike in normal pregnancy, in preeclampsia the cytotro-phoblast fails to adopt a vascular adhesion phenotype,10
which compromises the supply of blood flow to the maternal–fetal interface. It has been hypothesized that a reducedformation of NO could account for abnormal placentalperfusion in preeclampsia.1,11 Most data indicate that pre-eclamptic placenta has a normal capacity to synthesizeecNOS as compared with normal placenta and the formationof NO is also comparable in preeclamptic and in normalplacenta.1,2 However, one study12 found that inhibiting NOsynthesis raised the perfusion pressure of isolated humancotyledon preparations from normal pregnant women, but notfrom preeclamptic women. Moreover, in preeclampsia theconcentration of the NO second messenger, cGMP, in theplacental circulation was lower than normal.1,13 Thus, pre-
Received August 25, 2003; first decision September 17, 2003; revision accepted December 24, 2003.From Mario Negri Institute for Pharmacological Research (M.N., M.T., P.C., S.B., D.M., R.M., F.Po., A.B., G.R.) Bergamo, Italy; Unit of Obstetrics
and Gynecology (F.P., A.C., C.P.), Ospedale San Gerardo, Milano, Italy; and Unit of Nephrology and Dialysis (G.R.), Azienda Ospedaliera, OspedaliRiuniti di Bergamo, Bergamo, Italy.
Correspondence to Dr Marina Noris, Mario Negri Institute for Pharmacological Research, Via Gavazzeni, 11, 24125 Bergamo, Italy. [email protected]
© 2004 American Heart Association, Inc.
Hypertension is available at http://www.hypertensionaha.org DOI: 10.1161/01.HYP.0000116220.39793.c9
614 by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from

eclampsia is a condition of normal placental expression ofecNOS and normal generation of NO, whose activity, how-ever, is abnormally reduced. This apparent inconsistency canbe reconciled considering that the relative activity of NO in agiven organ/tissue depends on the rates of synthesis anddegradation. Beside NO, NOS enzymes also generate super-oxide anion (O2
�) and the rate of NO versus O2� formation
and disposal is closely regulated by intracellular levels ofL-arginine, tetrahydrobiopterin, and superoxide dismutase(SOD).14–16 NO, by interacting with O2
�, forms peroxynitrite,a cytotoxic anion that inhibits mitochondrial electron trans-port, oxidizes proteins, initiates lipid peroxidation, and ni-trates aromatic amino acids.17 Markers of oxidative stress areincreased in placenta18 of women with preeclampsia andnitrotyrosine staining, a marker of peroxynitrite, has beenfound in the preeclamptic placenta.18,19
The present study was designed: (1) to test whether lowerplacental NO bioavailability of preeclampsia is caused moreby increased NO degradation to peroxynitrite than to lowecNOS expression or activity and (2) to investigate whetherexcessive peroxynitrite formation is related to a low bioavail-ability of the NO precursor L-arginine and to find out thebiochemical determinants of that.
MethodsPatientsPregnant women (normotensive pregnancy, NP: n�13; preeclamp-sia: n�11) were recruited among those referred to the Obstetrics andGynecology Division of the Ospedale San Gerardo in Monza.Healthy age-matched nonpregnant women (n�6) served as controls.The Institutional Review Committee approved the study and allparticipants gave informed consent to the study. Criteria for definingNP were: no history of hypertension, diastolic blood pressure (BP)�90 mm Hg, systolic BP �140 mm Hg, and no significant protein-uria. Four women underwent premature delivery for ruptured mem-branes (n�2), oligohydramnios (n�1), and fetal distress (n�1).According to previously published criteria,20 preeclampsia wasdiagnosed as increase in diastolic BP of 15 mm Hg and systolic BPof 30 mm Hg at two measurements at least 4 hours apart comparedwith BP obtained before 20 weeks of gestation, proteinuria �0.3g/24 hours in the absence of urinary tract infection, return to normalBP, resolution of proteinuria by 12 weeks postpartum, and edema.One preeclamptic pregnancy was complicated by the HELLP syn-drome and two were complicated by intrauterine growth restriction
(IUGR). The patients’ clinical characteristics are summarized inTable 1.
Tissue Collection and HistologyFor placenta tissue collection, see the online data supplementavailable at http://www.hypertensionaha.org.
Placenta pathology score was graded according to the presence (inat least 30% of the tissue) of each of the following abnormalities:intervillous thrombosis, subchorionic thrombosis, infarcts, chronichypoxia, and intima hyperplasia, according to the guidelines of theCollege of American Pathologists (Table 1).21
NADPH-Diaphorase, Immunoperoxidase, andWestern BlottingNADPH-diaphorase reaction was performed on 7-�m thick frozensections, as described.22 Immunoperoxidase for ecNOS, nitroty-rosine, HNE-lysine, and arginase was performed on 3-�m paraffinsections.22 Western blot analysis for arginase II was performed inplacenta homogenates. An expanded Methods section can be foundin online data supplement available at http://www.hypertensionaha.org.
NOS Activity and L-arginine Levels inPlacenta TissueHomogenates of total placentas or of villous and decidua tissuesfrom NP (n�9) and preeclamptic women (n�6) were used toevaluate NOS activity and L-arginine levels, respectively (see onlinedata supplement available at http://www.hypertensionaha.org).
Conjugated DienesTotal lipids extracted from frozen tissues were resuspended incyclohexane and the absorbance read at 233 nm. The extinctioncoefficient of 2.52�104 mol/L�1cm�1 was used to calculate the dieneconcentration.23
Plasma L-arginine and Nitrites/Nitrates(NO2
�/NO3�)
Plasma was obtained from maternal (antecubital vein) and fetalblood (umbilical vein) collected on heparin and was stored at �80°Cuntil assayed. Women fasted at least 8 hours before blood collection.In preliminary experiments, we found that in this condition, theinterference of dietary intake to NO2
�/NO3� plasma levels is negli-
gible (not shown). Plasma L-arginine was measured by HPLC.24
NO2�/NO3
� plasma levels were measured as reported in online datasupplement http://hypertensionaha.org.
Real-Time Quantitative RT-PCRTo analyze gene expression of the cationic amino acid transportersCAT-1,25 4F2hc,25 LAT-1,25 and arginase II,26 and of the housekeep-
TABLE 1. Major Clinical Characteristics of Women With Normotensive Pregnancyand Preeclampsia
Parameters
Normotensive Pregnancy Preeclampsia
Mean SEM (Range) Mean SEM (Range)
Age (y) 33.2 1.8 (26–42) 32.6 1.8 (27–43)
Systolic blood pressure (mm Hg) 125.4 2.7 (115–140) 157.3* 3.3 (130–170)
Diastolic blood pressure (mm Hg) 70.3 1.7 (60–83) 99.1* 1.9 (90–110)
Proteinuria (g/24 h) Absent 2.54 1.78 (0.34–5.20)
Gestational age (wk) 37.6 0.8 (32.2–41.0) 32.4† 1.3 (25.5–40.0)
Placenta pathology score 0.4 (0–3) 2‡ (1–3)
Nulliparous/multiparous 6/7 7/4
Birth weight (g) 2880 149 (1920–3650) 1683† 343 (510–3330)
Apgar score 9.7 0.2 (8–10) 8.5§ 0.4 (7–10)
*P�0.0001; †P�0.005; ‡P�0.001; §P�0.01 vs normotensive pregnancy.
Noris et al Arginine and Peroxynitrite in Preeclampsia 615
by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from

ing gene �-actin, RNA was treated with DNase and reverse transcribedto cDNA. Quantitative real-time polymerase chain reaction (PCR) wasperformed on a TaqMan ABI PRISM 5700 Sequence Detection System(PE Applied Biosystems, Monza, Italy) with SYBR Green PCR CoreReagents (Applied Biosystems), in combination with optimal primerconcentrations. Detailed methods are available in the online datasupplement http://www.hypertensionaha.org.
Statistical AnalysisResults are means�SEM. Groups were compared by 1-wayANOVA using the StatView 4.01 software. Linear regression anal-ysis was used to correlate immunoperoxidase and mRNA expressiondata with clinical and experimental parameters. The � SpearmanRank test was used to correlate immunoperoxidase data with theplacenta pathology score and Apgar score. The statistical level ofsignificance of the two-tailed test was defined as P�0.05.
ResultsPlacental NADPH-Diaphorase Staining, NOSActivity, and ecNOS Expression and LocalizationAre Comparable in NP and in PreeclampsiaComparable diaphorase staining, NOS activity, and ecNOSexpression were seen in placentas from NP and preeclampsia(online Figure I and Table 2). Detailed results and onlineFigure I are in online data supplement available athttp://www.hypertensionaha.org.
Exuberant Peroxynitrite Formationin PreeclampsiaNitrotyrosine staining was absent or very faint in villi anddecidua vessels of placentas from NP (Figure 1A, B),whereas it was intense in preeclamptic villous tissue, mainlylocalized in the syncytiotrophoblast (Figure 1C and Table 2).The endothelium of villous vessels also showed moderatenitrotyrosine staining. A faint nitrotyrosine staining was alsoseen in the vessels of the decidua in preeclamptic placenta(Figure 1D and Table 2), whereas no staining was found inthe cells of the stroma. Nitrotyrosine and ecNOS stainingcolocalized in preeclamptic villi (Figure 1E, F), indicatingthat peroxynitrite was formed within the very cells thatsynthesized NO. Moderate specific ecNOS staining was also
found in villous vascular endothelium (Figure 1G). No signalwas found in negative control (Figure 1H).
The mean gestational age of NP was significantly greaterthan in the preeclamptic group. However, we noted nodifferences in nitrotyrosine staining in placental tissue withdiffering gestational age in either group. The intensity ofnitrotyrosine staining in pre-term NP (n�4) was not differentfrom NP at term (n�9) (syncytiotrophoblast: 0.55�0.29versus 0.28�0.10; endothelium of villi: 0.38�0.38 versus0.16�0.10), but was significantly weaker (P�0.05) than inpreeclamptic villi. An inverse correlation was found betweennitrotyrosine staining in the syncytiotrophoblast and birthweight (r��0.63, P�0.01) and Apgar score (���0.47,P�0.05), whereas a positive correlation was found betweennitrotyrosine and placenta pathology score (��0.48,P�0.05).
Excess Lipid Peroxidation in thePreeclamptic PlacentaAs compared with NP, a larger amount of conjugated dieneswas extracted from preeclamptic villi (NP: 24�1; preeclamp-sia: 47�3 pmol/�g phospholipid; P�0.0001) and umbilicalcord tissue (NP: 263�27; preeclampsia: 497�54 pmol/�gphospholipid; P�0.001). Dienes concentration in preeclamp-tic deciduas was also increased, although to a lesser degree(NP: 22�2; preeclampsia: 36�4 pmol/�g phospholipid;P�0.05). Within the NP group, the amount of conjugateddienes was comparable for preterm and for term pregnancies,indicating that placental lipid peroxidation does not dependon the week of gestation.
4-HNE-lysine staining, a marker of cell oxidative stress,27
was mostly faint in the syncytiotrophoblast and villousvessels (Figure 2A) as well as in the endothelium of NPdecidua vessels (Figure 2B). A higher 4-HNE-lysine stainingintensity was found in the syncytiotrophoblast and in endo-thelial cells of preeclamptic villous vessels than in NP villi(Figure 2C, Table 2). Moderate staining was also focallyfound on preeclamptic decidua vessels (Figure 2D and Table2). 4-HNE-lysine and nitrotyrosine staining showed the same
TABLE 2. Mean Scores of ecNOS, NADPH-Diaphorase, Nitrotyrosine, 4-HNE-lysine, CuZn-SODImmunoperoxidase, and Arginase II Staining in Villi and Decidua From Normotensive Pregnancy (n�13) andPreeclampsia (n�11)
Markers
Normotensive Pregnancy Preeclampsia
Chorionic Villi Decidua Chorionic Villi Decidua
Endothelium Syncytiotrophoblast Vessels Endothelium Syncytiotrophoblast Vessels
ecNOS 1.6�0.2 1.9�0.2 1.6�0.2 1.2�0.1* 1.6�0.1 1.1�0.2
NADPH-diaphorase 2.2�0.1 2.1�0.1 1.3�0.1 2.0�0.1 2.2�0.1 1.7�0.1
Nitrotyrosine 0.3�0.1 0.4�0.1 0.2�0.1 1.0�0.1† 1.6�0.2‡ 0.7�0.2§
4-HNE-lysine 0.8�0.2 0.8�0.1 0.6�0.1 1.6�0.1§ 1.9�0.1† 1.5�0.1§
CuZnSOD 1.5�0.3 0.9�0.3 1.3�0.4 1.5�0.5 1.2�0.3 1.5�0.6
Arginase II 0.4�0.1 0.5�0.2 0.4�0.1 1.2�0.2* 1.4�0.2* 1.0�0.2
Each section was scored for intensity of immunostaining (absent, faint, moderate, intense: 0 through 3).Data are mean�SEM.ecNOS indicates endothelial nitric oxide synthase; 4-HNE-lysine, 4-hydroxynonenal-(4-HNE)-lysine; superoxide dismutase,
CuZn-SOD.*P�0.05; †P�0.001; ‡P�0.0001; §P�0.005 vs normotensive pregnancy.
616 Hypertension March 2004
by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from

cell localization (Figure 2E and F), and the intensity of4-HNE-lysine staining in the syncytiotrophoblast (r�0.79,P�0.0001) and in villous endothelium (r�0.64, P�0.005)correlated with the intensity of nitrotyrosine staining, con-firming the association between lipid peroxidation and per-oxynitrite formation. No signal was found in negative control(Figure 2G). The intensity of 4-HNE-lysine staining in villoustissue was inversely correlated with birth weight (syncytiotro-phoblast: r��0.69, P�0.001, endothelium: r��0.58,P�0.01) and Apgar score (syncytiotrophoblast: ���0.61,P�0.01, endothelium: ���0.62, P�0.01), whereas a posi-tive correlation was found between 4-HNE-lysine stainingand placenta pathology score (syncytiotrophoblast: ��0.54,P�0.05).
Immunostaining for CuZn–SOD showed no differences inintensity and localization in either villous tissue or deciduavessels between NP and preeclamptic placentas (Table 2).
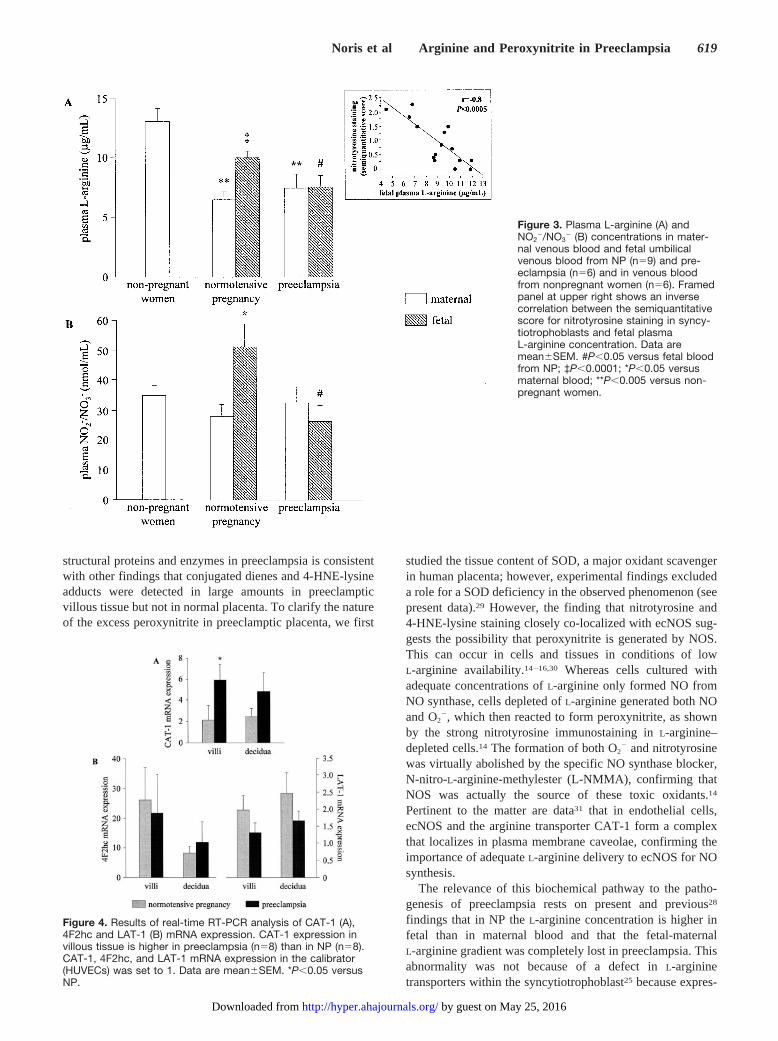
Fetal–Maternal Gradient of L-arginine andNO2
�/NO3� Is Lost in Preeclampsia
In line with previous data,28 the mean concentration ofL-arginine in maternal blood from NP was significantly lowerthan in nonpregnant women (Figure 3A). The same was truein preeclampsia (Figure 3A). No significant differences wereobserved in maternal plasma L-arginine in NP and preeclamp-
sia (Figure 3A). In NP, L-arginine was significantly higher infetal than in maternal blood (Figure 3A), which is in line withthe existence of an active placental L-arginine transport fromthe mother to the fetus.25 By contrast, in preeclampsia, fetalL-arginine concentrations were almost identical to those inmaternal blood (Figure 3A) and significantly lower than NP(Figure 3A). As previously reported,28 we found no correla-tion between maternal (r�0.03, P�0.93) or fetal (r�0.16,P�0.6) L-arginine concentrations and gestational age, thusexcluding that the lower fetal L-arginine in preeclampsiadepends on shorter gestation.
Consistent with L-arginine data, in NP the concentration ofthe NO metabolites, NO2
�/NO3�, in the fetal blood was
significantly higher than in the maternal blood (Figure 3B). Inpreeclampsia, NO2
�/NO3� concentration in maternal blood
was normal whereas levels in fetal blood were significantly(P�0.05) lower than in NP (Figure 3B). An inverse correla-tion was found between fetal blood L-arginine concentrationand the intensity of nitrotyrosine staining in the syncytiotro-phoblast (framed panel at upper right of Figure 3A) and in theendothelium of villous vessels (r��0.6, P�0.05).
Expression of Cationic Amino Acid TransportersIs Not Reduced in Preeclamptic PlacentaTo search for the possible causes of loss of fetal–maternalL-arginine and NO2
�/NO3� gradients in preeclampsia, we
Figure 1. Localization of peroxynitrite inthe placenta by immunostaining withanti-nitrotyrosine Ab. Nitrotyrosine stain-ing (dark brown) is very faint both in villi(A) and in decidua vessels (B) from NP.Intense nitrotyrosine staining is evident inpreeclamptic villi (C and E), which mainlylocalizes in syncytiotrophoblast. In NPvillous tissue (G) and in preeclamptic villi(F), intense ecNOS immunostaining(brown) is evident throughout the syncy-tiotrophoblast. D, Preeclamptic deciduasection showing a faint nitrotyrosinestaining localized in the endothelium ofvessels. No signal is evident in negativecontrol (H). v, indicates villous tissue; d,decidua; closed arrow, syncytiotropho-blast; open arrow, endothelium. Originalmagnifications: A, B, C, D, and H: �200;G, E, and F: �400.
Noris et al Arginine and Peroxynitrite in Preeclampsia 617
by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from

evaluated gene expression of two cationic amino acid trans-porters highly expressed in human placenta, CAT-1(y�system transporter)25 and 4F2hc and LAT-1 (the 2 sub-units of the y�L system transporter).25 As shown in Figure4A, CAT-1 expression in villous tissue was higher in pre-eclampsia than in NP. Expression of CAT-1 in decidua and of4F2hc and LAT-1 (Figure 4A, 4B) in villous tissue anddecidua was comparable in NP and in preeclampsia.
Excess Expression of Arginase II inPreeclamptic PlacentaWe then looked for placental expression of arginase II—theextrahepatic isoform of arginase that degrades arginine intoornithine and urea.26 Real-time RT-PCR analysis of villoustissue specimens showed a higher expression of arginase II inpreeclampsia than in NP, whereas arginase II expression indecidua tissue was comparable in the two groups (Figure 5A).The expression of arginase II in preterm NP villous tissue(3.08�1.32) was not different from arginase II in villoustissue from NP at term (4.08�1.05), indicating that placentalarginase II expression does not depend on the week ofgestation. The levels of arginase II mRNA in villous tissuewere inversely correlated with fetal L-arginine concentrations(r��0.8, P�0.005), indicating that arginase II expressed inplacenta may modulate L-arginine availability to the fetus.Immunostaining and Western blot for arginase II was per-
formed to evaluate tissue localization and protein expression.Arginase II staining was mostly faint in chorionic villi and indecidua from NP (Figure 5B, Table 2). An intense arginase IIstaining was instead found in the syncytiotrophoblast and inendothelial cells of villous vessels of preeclamptic placenta(Figure 5C, Table 2). A moderate staining was also focallyfound in preeclamptic decidua vessels (Table 2); however,mean values were not significantly different from thoserecorded in NP. No signal was found in negative control(Figure 5D). Consistent with immunostaining results, West-ern blot analysis of placenta homogenates showed a faintimmunoreactive band at 39 kDa in NP whereas a strong bandwas found in preeclampsia (Figure 5E).
To verify whether increased arginase II in the preeclampticplacenta limits L-arginine availability, L-arginine tissue levelswere evaluated. As shown in Figure 5F, L-arginine levels inpreeclamptic villi were lower (P�0.05) than levels in villifrom NP, whereas L-arginine levels in the decidua werecomparable in the two groups.
DiscussionAn excess formation of peroxynitrite was found in pre-eclamptic placentas, despite the fact that ecNOS was ex-pressed at comparable extent in NP and in preeclampsia,confirming previously reported data.19 That this can be takento reflect an excess lipid peroxidation and oxidation of
Figure 2. Tissue localization of lipid per-oxidation products by immunostainingwith anti-4-hydroxynonenal-(4-HNE)-lysine Ab. 4-HNE-lysine staining (darkbrown) is mostly faint in NP villi (A) anddecidua (B). In preeclamptic placenta,intense 4-HNE-lysine staining is evidentin syncytiotrophoblast and in the villousendothelium (C and E). F, Immunostain-ing with anti-nitrotyrosine Ab of villoustissue from the same preeclampticwoman, showing colocalization of nitro-tyrosine and 4-HNE-lysine staining. Mod-erate staining is also focally evident onvessels of preeclamptic decidua (D). Nosignal is found in the negative control(G). v, indicates villous tissue; d,decidua; closed arrow, syncytiotropho-blast; open arrow, endothelium. Originalmagnifications: A, B, C, D, and G: �200;E and F: �400.
618 Hypertension March 2004
by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from
structural proteins and enzymes in preeclampsia is consistentwith other findings that conjugated dienes and 4-HNE-lysineadducts were detected in large amounts in preeclampticvillous tissue but not in normal placenta. To clarify the natureof the excess peroxynitrite in preeclamptic placenta, we first
studied the tissue content of SOD, a major oxidant scavengerin human placenta; however, experimental findings excludeda role for a SOD deficiency in the observed phenomenon (seepresent data).29 However, the finding that nitrotyrosine and4-HNE-lysine staining closely co-localized with ecNOS sug-gests the possibility that peroxynitrite is generated by NOS.This can occur in cells and tissues in conditions of lowL-arginine availability.14–16,30 Whereas cells cultured withadequate concentrations of L-arginine only formed NO fromNO synthase, cells depleted of L-arginine generated both NOand O2
�, which then reacted to form peroxynitrite, as shownby the strong nitrotyrosine immunostaining in L-arginine–depleted cells.14 The formation of both O2
� and nitrotyrosinewas virtually abolished by the specific NO synthase blocker,N-nitro-L-arginine-methylester (L-NMMA), confirming thatNOS was actually the source of these toxic oxidants.14
Pertinent to the matter are data31 that in endothelial cells,ecNOS and the arginine transporter CAT-1 form a complexthat localizes in plasma membrane caveolae, confirming theimportance of adequate L-arginine delivery to ecNOS for NOsynthesis.
The relevance of this biochemical pathway to the patho-genesis of preeclampsia rests on present and previous28
findings that in NP the L-arginine concentration is higher infetal than in maternal blood and that the fetal-maternalL-arginine gradient was completely lost in preeclampsia. Thisabnormality was not because of a defect in L-argininetransporters within the syncytiotrophoblast25 because expres-
Figure 3. Plasma L-arginine (A) andNO2
�/NO3� (B) concentrations in mater-
nal venous blood and fetal umbilicalvenous blood from NP (n�9) and pre-eclampsia (n�6) and in venous bloodfrom nonpregnant women (n�6). Framedpanel at upper right shows an inversecorrelation between the semiquantitativescore for nitrotyrosine staining in syncy-tiotrophoblasts and fetal plasmaL-arginine concentration. Data aremean�SEM. #P�0.05 versus fetal bloodfrom NP; ‡P�0.0001; *P�0.05 versusmaternal blood; **P�0.005 versus non-pregnant women.
Figure 4. Results of real-time RT-PCR analysis of CAT-1 (A),4F2hc and LAT-1 (B) mRNA expression. CAT-1 expression invillous tissue is higher in preeclampsia (n�8) than in NP (n�8).CAT-1, 4F2hc, and LAT-1 mRNA expression in the calibrator(HUVECs) was set to 1. Data are mean�SEM. *P�0.05 versusNP.
Noris et al Arginine and Peroxynitrite in Preeclampsia 619
by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from

sion of y� and y�L system cationic transporters was normal oreven increased in preeclamptic placenta. Based on the observa-tion that L-arginine levels in preeclamptic villi were lower thanin NP villi, we then addressed the possibility that L-argininedeficiency was caused by consumption of the amino acid in thepreeclamptic villous tissue by arginase II, the extrahepaticisoform of arginase expressed in human placenta.26 Both argi-nase and NOS use arginine as common substrate and arginaseinhibits NO synthesis by reducing arginine bioavailability.Transfection of endothelial cells with arginase II increasedL-arginine consumption and reduced NO synthesis26 and in vivoadministration of arginase to experimental animals significantlydepleted plasma L-arginine.32 Arginase II mRNA expression wasmore than 4-fold higher in preeclamptic than in NP villoustissue; also, protein expression was increased, as documented byimmunostaining and Western blot data. In addition, levels ofarginase II mRNA in villous tissue inversely correlated withfetal L-arginine concentration. We hypothesize that in pre-
eclamptic placenta, higher than normal expression of arginase IIcauses less L-arginine available for ecNOS in trophoblast cellsand in the villous endothelium, which can lead to the aberrantcatalytic activity described, giving a rapid NO degradation byreactive oxygen species. This possibility is also supported byfinding in our patients that the intensity of nitrotyrosine and4-HNE-lysine staining in villous tissue and fetal L-arginineconcentration were inversely correlated. Deficiency in tetrahy-drobiopterin (BH4) can also poise ecNOS toward producingsuperoxide anion;33 however, levels of BH4 were found to benormal in preeclamptic placentas.33
Further investigation is required to clarify the mechanismresponsible for arginase II upregulation in preeclampsia.Testosterone could be a possible candidate mediator of theaforementioned phenomenon. Indeed in studies in female ratsand mice, testosterone stimulates arginase activity and pa-tients with preeclampsia have been shown to have higherlevels of testosterone than NP women.11
Figure 5. A, Results of real-time RT-PCR analysis of arginase II mRNA. Arginase II expression in preeclamptic villi (n�9) is higher thanin villi from NP (n�9). Arginase II mRNA expression in the calibrator (PBMC) was set to 1. B–D, Tissue localization of arginase II by im-munostaining with anti-arginase II Ab. Intense arginase II staining is evident in preeclamptic villous tissue (C) and faint staining is shownin NP villi (B). D, negative control. Closed arrow indicates syncytiotrophoblast; open arrow, endothelium; v, villi. Original magnification:B–D: �200. E, Western blot analysis of arginase II in homogenates from NP and preeclamptic (PE) placentas. Representative resultsfrom 2 NP and 2 preeclamptic placentas are shown. F, L-arginine levels in homogenized villi and decidua from NP (n�9) and pre-eclamptic women (n�6). Data are mean�SEM. *P�0.05 versus NP.
620 Hypertension March 2004
by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from

Previous reports1 of higher resistance in the fetal–placentalcirculation in the preeclamptic placenta are consistent withthe reduced placental NO availability we observed, becauseNO is a potent vasodilator in the vascular district.1 Inaddition, there is evidence of a major role for NO as anangiogenic and vascular remodelling factor.6–9 Exposure ofhuman endothelial cells to NO donors leads to a dose-dependent increase in endothelial cell migration and differ-entiation,34 and NO is a mediator of growth factor-inducedangiogenesis.6,34,35 That NO may be instrumental to endovas-cular invasion and vessel remodeling of developing placentais suggested by a number of experimental findings. First, NOrelease is coupled to VEGF and hepatocyte growth factor(HGF)-induced trophoblast invasion and motility.36,37 Sec-ond, in trophoblast cells, NO upregulates the expression andthe activity of the matrix-degrading proteases MMP-2 andMMP-9, which are required for invasion during embryoimplantation.38 Finally, NO causes dilation of the uteropla-cental arteries, which is another prerequisite for trophoblastinvasion and remodeling of the endothelium.1 It is tempting tospeculate that low NO availability could contribute to im-paired cytotrophoblast invasion in preeclampsia, a hypothesisthat should be formally tested in developing placenta ob-tained before 20 weeks of gestation. However, the presentstudy, as most previously published reports on preeclampticplacentas10,18,19 have been necessarily performed at the timeof disease onset, usually in the third trimester of pregnancy,represents a limitation that cannot be easily overcome.
In conclusion, our findings indicate that normal placentawith enough tissue L-arginine sustains adequate generation ofNO by ecNOS. By contrast, in preeclampsia, when theplacental L-arginine concentration is low because of exces-sive arginase II expression, activation of ecNOS leads toexcessively high generation of superoxide anion, whichreduces NO half-life by forming peroxynitrite. This maypromote microvascular oxidative damage and favor abnormalplacenta perfusion.
PerspectivesThe present findings provide a rationale for clinical trials withL-arginine or antioxidant supplementation39 that, either byproviding more substrate to ecNOS or by lowering the rate ofNO degradation to peroxynitrite, could help prevent thisdisease, which remains one of the leading causes of maternaland fetal morbidity and mortality.
AcknowledgmentsDr Paola Cassis is a recipient of a fellowship from AssociazioneRicerca Malattie Rare (ARMR, Bergamo) through the generosity ofBanco di Brescia. Dr Francesca Porrati received a fellowship inmemory of Libera Dossi Grana. This work has been partiallysupported by a grant from Farmaceutici Damor S.p.A, Napoli, Italy.
References1. Baylis C, Beinder E, Suto T, August P. Recent insights into the roles of
nitric oxide and renin-angiotensin in the pathophysiology of preeclampticpregnancy. Semin Nephrol. 1998;18:208–230.
2. Noris M, Benigni A, Remuzzi G. The role of vasoactive molecules ofendothelial origin in the pathophysiology of normal pregnancy andpregnancy-induced hypertension. Curr Opin Nephrol Hypertens. 1996;5:347–352.
3. Moncada S, Higgs A. The L-arginine-nitric oxide pathway. N Engl J Med.1993;329:2002–2012.
4. Conrad KP, Vill M, McGuire PG, Dail WG, Davis AK. Expression ofnitric oxide synthase by syncytiotrophoblast in human placental villi.FASEB J. 1993;7:1269–1276.
5. Zhou Y, Fisher SJ, Janatpour M, Genbacev O, Dejana E, Wheelock M,Damsky CH. Human cytotrophoblasts adopt a vascular phenotype as theydifferentiate. A strategy for successful endovascular invasion? J ClinInvest. 1997;99:2139–2151.
6. Shizukuda Y, Tang S, Yokota R, Ware JA. Vascular endothelial growthfactor-induced endothelial cell migration and proliferation depend on anitric oxide-mediated decrease in protein kinase Cdelta activity. Circ Res.1999;85:247–256.
7. Babaei S, Stewart DJ. Overexpression of endothelial NO synthaseinduces angiogenesis in a co-culture model. Cardiovasc Res. 2002;55:190–200.
8. Martin D, Conrad KP. Expression of endothelial nitric oxide synthase byextravillous trophoblast cells in the human placenta. Placenta. 2000;21:23–31.
9. Lyall F, Bulmer JN, Kelly H, Duffie E, Robson SC. Human trophoblastinvasion and spiral artery transformation: the role of nitric oxide. Am JPathol. 1999;154:1105–1114.
10. Zhou Y, Damsky CH, Fisher SJ. Preeclampsia is associated with failureof human cytotrophoblasts to mimic a vascular adhesion phenotype. Onecause of defective endovascular invasion in this syndrome? J Clin Invest.1997;99:2152–2164.
11. Lowe DT. Nitric oxide dysfunction in the pathophysiology of pre-eclampsia. Nitric Oxide. 2000;4:441–458.
12. Gonzalez C, Cruz MA, Gallardo V, Lagos M, Varela J, Alboroz J, SchulzC. Nitric oxide and prostaglandin system inhibition on the isolatedperfused human placenta from normal and preeclamptic pregnancies.Gynecol Obstet Invest. 1995;40:244–248.
13. Kovasc AG, Makary A, Peto J, Steinmetz G. Deficiency of cGMP levelin placental circulation in pregnancy-induced hypertensive disorders:Possibility of decreased endothelium-derived relaxing factor. HypertensPregn. 1994;13:163–169.
14. Xia Y, Dawson VL, Dawson TM, Snyder SH, Zweier JL. Nitric oxidesynthase generates superoxide and nitric oxide in arginine-depleted cellsleading to peroxynitrite-mediated cellular injury. Proc Natl Acad SciU S A. 1996;93:6770–6774.
15. Schmidt HHHW, Hofmann H, Schindler U, Shutenko ZS, CunninghamDD, Feelisch M. No NO from NO synthase. Proc Natl Acad Sci U S A.1996;93:14492–14497.
16. Vasquez-Vivar J, Kalyanaraman B, Martasek P, Hogg N, Masters BSS,Karoui H, Tordo P, Pritchard KA Jr. Superoxide generation by endothe-lial nitric oxide synthase: the influence of cofactors. Proc Natl Acad SciU S A. 1998;95:9220–9225.
17. Radi R, Beckman JS, Bush KM, Freeman BA. Peroxynitrite-inducedmembrane lipid peroxidation: the cytotoxic potential of superoxide andnitric oxide. Arch Biochem Biophys. 1991;288:481–487.
18. Many A, Hubel CA, Fisher SJ, Roberts JM, Zhou Y. Invasive cytotro-phoblasts manifest evidence of oxidative stress in preeclampsia. Am JPathol. 2000;156:321–331.
19. Myatt L, Rosenfield RB, Eis ALW, Brockman DE, Greer I, Lyall F.Nitrotyrosine residues in placenta. Evidence of peroxynitrite formationand action. Hypertension. 1996;28:488–493.
20. Redman CW. CLASP. a randomised trail of low-dose aspirin for theprevention and treatment of pre-eclampsia among 9364 pregnant women:CLASP (Collaborative Low-dose Aspirin Study in pregnancy) Collabo-rative. Lancet. 1994;343:619–629.
21. Langston C, Kapplan C, Macpherson T, Manci E, Peevy K, Clark B,Murtagh C, Cox S, Glenn G. Pratice guideline for examination of theplacenta. developed by the Placental Pathology Practice Guideline Devel-opment Task Force of the College of Am Pathologists. Arch Pathol LabMed. 1997;121:449–476.
22. Aiello S, Noris M, Todeschini M, Zappella S, Foglieni C, Benigni A,Corna D, Zoja C, Cavallotti D, Remuzzi G. Renal and systemic nitricoxide synthesis in rats with renal mass reduction. Kidney Int. 1997;52:171–181.
23. Folch J, Lees M, Stanley GHS. A single method for the isolation andpurification of total lipides from animal tissue. J Biol Chem. 1957;226:497–509.
24. Todeschini M, Macconi D, Garcia Fernandez N, Ghilardi M, Anabaya A,Binda E, Morigi M, Cattaneo D, Perticucci E, Remuzzi G, Noris M.
Noris et al Arginine and Peroxynitrite in Preeclampsia 621
by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from

Effect of acetate-free biofiltration and bicarbonate hemodialysis on neu-trophil activation. Am J Kidney Dis. 2002;40:783–793.
25. Jansson T. Amino acid transporters in the human placenta. PediatricResearch. 2001;49:141–147.
26. Li H, Meininger CJ, Hawker JR, Haynes TE, Kepka-Lenhart D, MistrySK, Morris SM Jr, Wu G. Regulatory role of arginase I and II in nitricoxide, polyamine, and proline syntheses in endothelial cells. Am JPhysiol. 2001;280:E75–E82.
27. Neale TJ, Ojha PP, Exner M, Poczewski H, Ruger B, Witztum JL, DavisP, Kerjaschki D. Proteinuria in passive heymann nephritis is associatedwith lipid peroxidation and formation of adducts on type IV collagen.J Clin Invest. 1994;94:1577–1584.
28. Cetin I, Ronzoni S, Marconi AM, Perugino G, Corbetta C, Battaglia FC,Pardi G. Maternal concentrations and fetal-maternal concentration dif-ferences of plasma amino acids in normal and intrauterine growth-restricted pregnancies. Am J Obstet Gynecol. 1996;174:1575–1583.
29. Myatt L, Eis AL, Brockman DE, Kossenjans W, Greer IA, Lyall F.Differential localization of superoxide dismutase isoforms in placentalvillous tissue of normotensive, preeclamptic, and intrauterine growth-restricted pregnancies. J Histochem Cytochem. 1997;45:1433–1438.
30. Pou S, Pou WS, Bredt DS, Snyder SH, Rosen GM. Generation ofsuperoxide by purified brain nitric oxide synthase. J Biol Chem. 1992;267:24173–24176.
31. McDonald KK, Zharikov S, Block ER, Kilberg MS. A caveolar complexbetween the cationic amino acid transporter 1 and endothelial nitric-oxidesynthase may explain the “Arginine paradox”. J Biol Chem. 1997;272:31213–31216.
32. Waddington S, Cook HT, Reaveley D, Jansen A, Cattell V. L-argininedepletion inhibits glomerular nitric oxide synthesis and exacerbates ratnephrotoxic nephritis. Kidney Int. 1996;49:1090–1096.
33. Kukor Z, Valent S, Toth M. Regulation of nitric oxide synthase activityby tetrahydrobiopterin in human placentae from normal and pre-eclamptic pregnancies. Placenta. 2000;21:763–772.
34. Bussolati B, Dunk C, Grohman M, Kontos CD, Mason J, Ahmed A.Vascular endothelial growth factor receptor-1 modulates vascular endo-thelial growth factor-mediated angiogenesis via nitric oxide. Am J Pathol.2001;159:993–1008.
35. Papapetropoulos A, Garcia-Cardena G, Madri JA, Sessa WC. Nitric oxideproduction contributes to the angiogenic properties of vascular endothe-lial growth factor in human endothelial cells. J Clin Invest. 1997;100:3131–3139.
36. Ahmed A, Dunk C, Kniss D, Wilkes M. Role of VEGF receptor-1 (Flt-1)in mediating calcium-dependent nitric oxide release and limiting DNAsynthesis in human trophoblast cells. Lab Invest. 1997;76:779–791.
37. Cartwright JE, Holden DP, Whitley GS. Hepatocyte growth factor reg-ulates human trophoblast motility and invasion: a role for nitric oxide.Br J Pharmacol. 1999;12:181–189.
38. Novaro V, Colman-Lerner A, Ortgega FV, Jawerbaum A, Paz D, LoNostro F, Pustovrh C. Regulation of metalloproteinases by nitric oxide inhuman trophoblast cells in culture. Reprod Fertil Dev. 2001;13:411–420.
39. Chappell LC, Seed PT, Briley AL, Kelly FJ, Lee R, Hunt BJ, Parmar K,Bewley SJ, Shennan AH, Steer PJ, Poston L. Effect of antioxidants on theoccurrence of pre-eclampsia in women at increased risk: a randomisedtrial. Lancet. 1999;354:810–816.
622 Hypertension March 2004
by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from

Picciolo and Giuseppe RemuzziBonazzola, Daniela Macconi, Raffaella Maucci, Francesca Porrati, Ariela Benigni, Claudio
Marina Noris, Marta Todeschini, Paola Cassis, Fabio Pasta, Anna Cappellini, SamanthaSpecies
l-Arginine Depletion in Preeclampsia Orients Nitric Oxide Synthase Toward Oxidant
Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 2004 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension doi: 10.1161/01.HYP.0000116220.39793.c9
2004;43:614-622; originally published online January 26, 2004;Hypertension.
http://hyper.ahajournals.org/content/43/3/614World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://hyper.ahajournals.org/content/suppl/2004/03/01/43.3.614.DC1.htmlData Supplement (unedited) at:
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialHypertensionin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on May 25, 2016http://hyper.ahajournals.org/Downloaded from

Plac
enta
l[3 H
]L-c
itrul
line
(nm
ol/m
g pr
otei
n)
C
(v)
B
(v)
A
(d)(v)
Figure I
normotensive pregnancy preeclampsia
0
0.2
0.4
0.6
0.8
1.0D

03-2102R Online data supplement
ONLINE DATA SUPPLEMENT
In Preeclampsia L-arginine Depletion Orients Nitric Oxide Synthase Toward
Oxidant Species
Marina Noris‡, Marta Todeschini‡, Paola Cassis‡, Fabio Pasta§, Anna Cappellini§,
Samantha Bonazzola‡, Daniela Macconi‡, Raffaella Maucci‡ , Francesca Porrati‡,
Ariela Benigni‡, Claudio Picciolo§, & Giuseppe Remuzzi¶‡
‡Mario Negri Institute for Pharmacological Research, Via Gavazzeni 11, 24125
Bergamo, Italy; §Unit of Obstetrics and Gynecology, Ospedale San Gerardo, Via
Solferino 16, 20052, Monza, Milano, Italy; ¶Unit of Nephrology and Dialysis, Azienda
Ospedaliera, Ospedali Riuniti di Bergamo, Largo Barozzi 1, 24128 Bergamo, Italy
Short title: arginine and peroxynitrite in preeclampsia
Send correspondence and reprint request to: Dr. Marina Noris "Mario Negri" Institute for Pharmacological Research Via Gavazzeni, 11 24125 Bergamo, Italy Tel. no. +39035319888 Fax no. + 39035319331

03-2102R Online data supplement
Tissue collection and histology
Placentas were obtained following vaginal delivery (5 with normotensive pregnancy, 1
with preeclampsia) or during cesarian delivery (8 with NP, 10 with preeclampsia).
Tissue blocks were taken from the fetal and maternal surfaces of the placenta to include
umbilical cord, chorionic villi (stem and terminal villi) and decidua. Some blocks were
snap-frozen in liquid nitrogen and stored at –80°C for NADPH diaphorase, conjugated
diene, Western blot and real-time RT-PCR assays, NOS activity, L-arginine levels,
others were fixed with 10% formalin and paraffin-embedded for immunoperoxidase and
conventional histological examination.
For NADPH diaphorase staining, sections were counterstained with methyl green.
Negative controls were run without NADPH or in the presence of the NO synthesis
inhibitor DPI,1 reactivity was totally NADPH-dependent and was abolished by DPI.
For immunoperoxidase, mouse monoclonal antibodies against the following antigens:
human-ecNOS (Transduction Laboratories, Exeter, UK),1 nitrotyrosine (Upstate
Biotechnology Inc, Lake Placid, NY),2 human-CuZn-SOD (Sigma Chemicals Co, St
Louis, MO) and 4-hydroxynonenal-(4-HNE)-lysine adduct (NA59, kindly provided by
Dr Witzum, The Scripps Research Institute, La Jolla, CA),3 and a rabbit polyclonal
antibody against human-arginase II (Santa Cruz Biotechnology, Inc., USA)4 were used.
After blocking aspecificities with non-immune horse serum (for ecNOS, nitrotyrosine,
CuZn-SOD, 4-HNE-lysine) or with normal goat serum (for arginase II), slides were
incubated overnight at 4°C with the primary antibody (anti-ecNOS 1:150, anti-
nitrotyrosine 1:300, anti-CuZn-SOD 1:500, anti-4-HNE-lysine 1:500, anti-arginase II
1:25) in PBS/1%-BSA, followed by the biotinylated horse anti-mouse IgG or goat anti-
rabbit IgG, ABC solution (Vector Laboratories, Burlingame, CA), and developed with

03-2102R Online data supplement
diaminobenzidine (for ecNOS) or diaminobenzidine-Nickel (for nitrotyrosine, CuZn-
SOD, 4-HNE-lysine and arginase II). For ecNOS, sections were incubated before
adding the ABC solution with a monoclonal antibody anti-biotin.1 The sections were
counterstained with Harris hematoxylin. Negative controls included the omission of the
primary antibody and replacement with a non-immune antibody on a second section on
the same slide. Each experiment randomly included samples from normotensive
pregnancy and from preeclampsia. In preliminary experiments anti-nitrotyrosine
antibody at 1:300 dilution brightly stained a monolayer of human microvascular
endothelial cells (HMEC) pre-incubated with 200 µmol/L peroxynitrite, while no signal
was found on HMEC incubated with vehicle (not shown).
Multiple sections from each patient were examined. Each section was scored for
intensity of immunostaining (absent, faint, moderate, intense: 0 through 3). At least 8-
10 fields per section were examined. The final score (S) per section was calculated as a
weighted mean:
S= N0x0+N1x1+N2x2+N3x3
(N0+N1+N2+N3)
where (N0+N1+N2+N3) is the number of villi or of decidua vessels in each category.
Two pathologists blinded to the nature of the experimental groups assigned the scores,
and the mean of the two assessments was calculated.

03-2102R Online data supplement
Western blot
For Western blotting analysis the frozen placental tissues were pulvarizated and
resuspended in 1 mL lysis buffer (50 mmol/L *-glicerolphosphate, 2 mmol/L MgCl 2, 1
mmol/L EGTA, 0.5% Triton X-100, 0.5% NP-40, 1 mmol/L DTT, 100 mL protease
inhibitor cocktail) and sonicated.
The whole lysate was stored at -80°C. Protein concentration was determined by using
the Bradford method (Biorad). The proteins (40 mg for each lane from either normal or
preeclamptic placentas) were separated on denaturating sodium dodecyl sulfate 10%
polyacrylamide gel by electrophoresis and then blotted on nitrocellulose membrane by
wet electroblotting for 90 minutes. Blots were blocked overnight at 4°C with 3% BSA,
1% goat serum in PBS at pH 7.3 (140 mmol/L NaCl, 2.7 mmol/L KCl, 10 mmol/L
Na2HPO4, 1.8 KH2PO4, 0.05% Tween 20) and then incubated for 2.5 hours with a
rabbit polyclonal antibody directed against human arginase II (1:200, Santa Cruz
Biotechnology) followed by the secondary antibody (HRP-conjugated goat anti-rabbit
IgG 1:30000, Vector Laboratories). Antibody binding was visualized by the enhanced
chemiluminescence detection system (ECL).
NOS activity in placenta tissue
Placental tissue (3-4 g) for NOS activity was finely minced and homogenized in buffer
as previously described.5 The homogenate (about 20 mg protein each) was incubated at
37°C for 24 hours with 1.2 mL Hank’s buffer containing 2 mmol/L NADPH (Sigma
Chemical Co, St Louis, MO), 0.45 mmol/L Ca2+ (1 *mol/L free calcium), 200 *mol/L
L-arginine (12 nmol L-arginine/mg protein) and 0.5 *Ci [ 3H]L-arginine.5 After

03-2102R Online data supplement
centrifugation the supernatant was extracted with 15%TCA (1:1, vol:vol) to evaluate the
conversion of [3H]L-arginine to [3H]L-citrulline.1
Measurement of L-arginine levels in placenta tissue
A portion (0.2-0.3 g) of villi or decidua was washed in 0.9% saline buffer. Tissue was
weighted and homogenized with buffer containing sucrose 0.25 mol/L, Tris 50 mmol/L,
EDTA 1 mmol/L, aprotinine 2 *g/mL, PMSF 1 *g/mL, STI 5 *g/mL, DTT 80 *g/mL,
pH 7. After centrifugation at 11000 rpm for 5 minutes at 4°C the supernatants were
used to measure levels of L-arginine as previously described.6
All results were corrected for mg protein concentration in each sample.
Measurement of NO2-/NO3- plasma levels
NO2-/NO3- plasma levels were measured using HPLC1 (MN System Gold, Beckman
Instruments Inc, Berkeley, CA). Briefly, plasma samples were treated with zincum
sulphate (60 *mol/L, final concentration) and centrifuged to eliminate proteins.
Supernatants were eluted onto a Dowex AG50WX-8 column followed by a cadmium
column, which catalysed the reduction of nitrate to nitrite (eluent: borate buffer,
pH=8.5). The post-column eluate reacted with Griess reagent (1% sulphanilic acid in
5% H3PO4, 0.1% n-[1-naphthyl]-ethylenediamine, vol:vol) (Sigma Chemical Co, St
Louis, MO) to form a purple azo dye and the color was detected at * 540nm.
Real time PCR
The amplification profile consisted of 50°C for 2 minutes, 95°C for 10 minutes and 40
cycles of 95°C for 15 minutes, then 60°C for 1 minute. All samples were run in

03-2102R Online data supplement
triplicate. To assess the overall cDNA content, *-actin served as housekeeping gene.
The following oligonucleotide primers were used: human CAT-1 (200 nmol/L): 5’-
ACACATCAGGCCGTCTCTGTT-3’ and (5’-AACCCGAAGGGCATGAATC-3’;
human 4F2hc (100 nmol/L): 5’-TGAGATTGGCCTGGATCCA-3’ and 5’-
CATGACTGGAGCCTCCATAGG-3’; human LAT-1 (600nmol/L): 5’-
TCGTTGCAGGCATTGTTAGACT-3’ and 5’-
GATGAACCCTCAAAGGAATTCTCA-3’; human arginase II (100 nmol/L): 5’-
GAAGAAATCCGTCCACTCCG-3’ and 5’-GGACCATGCTCCACTCCTTTT-3’;
human *-actin (50 nmol/L): 5’-TCACCCACACTGTGCCCATCTACGA-3’ and 5’-
CAGCGGAACCGCTCATTGCCAATGG-3’.
Similar amplification efficiencies for targets and housekeeping genes were
demonstrated by analysing serial cDNA dilutions, showing an absolute value of the slop
of log input cDNA amount versus *Ct (Ct target - Ct housekeeping gene) of <0.1. Thus,
we used the **Ct technique (user bulletin # 2, PE Applied Biosystems and reference7)
to calculate cDNA content in each sample using the cDNA expression in human
umbilical vein endothelial cells (HUVECs, for CAT-1 and 4F2hc and LAT-1) or in
peripheral blood mononuclear cells (PBMC, for arginase II) as reference (calibrator).
Melting temperature analysis evidenced no primer dimers or aspecific amplification
products. Controls consisting of ddH2O or RT- RNAs were negative for targets and
housekeeping.
Results
Placental NADPH-diaphorase staining, NOS activity and ecNOS expression and
localization are comparable in NP and in preeclampsia

03-2102R Online data supplement
Intense diaphorase staining, which detects catalytic NOS activity irrespective of
the enzyme isoform,1 was evident in the chorionic villi of placentas from NP, mainly in
the syncytiotrophoblast (Figure IA and B). Staining was very faint in the decidua
(Figure IA). The intensity and distribution of staining in villous tissue (Figure IC) and in
the decidua (Table 2) were similar in NP and in preeclampsia. Since NADPH-
diaphorase activity is not specific for NOS and enzyme involved in sex steroid
metabolism are also diaphorase enzymes, NOS activity was also evaluated by
measuring the conversion of 3HL-arginine to 3HL-citrulline in placental tissue
homogenate in the presence of 4 to 5 fold excess of the substrate L-arginine. Results
shown in Figure ID are in line with the diaphorase staining and document comparable
NOS activity in placental tissue from NP and preeclampsia.
In villous tissue ecNOS staining was intense throughout the syncytiotrophoblast in NP
and in preeclamptic women (Figure 1G and F). Moderate specific ecNOS staining was
also found in villous vascular endothelium (Figure 1G). No significant differences in
intensity and localization were seen in either villous tissue or decidua vessels from the
placentas from NP and preeclampsia (Figure 1F and Table 2). EcNOS was only slightly
reduced in the endothelium of preeclamptic villous vessels, probably depending on the
reduced number of viable cells.
Legend to Figure I:
panels A-C: Representative photomicrographs of NADPH-
diaphorase staining (blue colour) of placenta tissue from normotensive (A and B) and
preeclamptic women (C). No difference in intensity and distribution of NADPH-
diaphorase staining can be observed between normotensive pregnancy and
preeclampsia. (v): villi, (d): decidua. Original magnification: A, 100x; B and C, 400x.

03-2102R Online data supplement
(panel D): Formation of [3H]L-citrulline from [3H]L-arginine in homogenate
placental tissue from normotensive pregnancy (n=9) and preeclamptic women (n=6).
Data are mean±SEM.

03-2102R Online data supplement
References :
1. Aiello S, Noris M, Todeschini M, Zappella S, Foglieni C, Benigni A, Corna D,
Zoja C, Cavallotti D, Remuzzi G. Renal and systemic nitric oxide synthesis in rats
with renal mass reduction. Kidney Int. 1997;52:171-181.
2. Myatt L, Rosenfield RB, Eis ALW, Brockman DE, Greer I, Lyall F.
Nitrotyrosine residues in placenta. Evidence of peroxynitrite formation and
action. Hypertension. 1996;28:488-493.
3. Neale TJ, Ojha PP, Exner M, Poczewski H, Ruger B, Witztum JL, Davis P,
Kerjaschki D. Proteinuria in Passive Heymann Nephritis is associated with lipid
peroxidation and formation of adducts on type IV collagen. J Clin Invest.
1994;94:1577-1584.
4. Carraway MS, Piantadosi CA, Jenkinson CP, Huang YC. Differential expression of
arginase and iNOS in the lung in sepsis. Exp Lung Res. 1998;24:253-68.
Boccardo P, Soregaroli M, Aiello S, Noris M, Donadelli R, Loiacono A, Benigni
A. Systemic and fetal-maternal nitric oxide synthesis in normal pregnancy and pre-
eclmpsia. Br J Obstet Gynaecol. 1996;103:879-86.
5. Todeschini M, Macconi D, Garcia Fernandez N, Ghilardi M, Anabaya A,
Binda E, Morigi M, Cattaneo D, Perticucci E, Remuzzi G, Noris M. Effect of
acetate-free biofiltration and bicarbonate hemodialysis on neutrophil activation. Am J
Kidney Dis. 2002;40:783-793.
6. Caprioli J, Bettinaglio P, Zipfel PF, Amadei B, Daina E, Gamba S, Skerka C,
Marziliano N, Remuzzi G, Noris M. The molecular basis of familial hemolytic
uremic syndrome: mutation analysis of factor H gene reveals a hot spot in short
consensus repeat 20. J Am Soc Nephrol. 2001;12:297-307.

03-2102R Online data supplement
Legend to Figure I:
panels A-C: Representative photomicrographs of NADPH-diaphorase
staining (blue colour) of placenta tissue from normotensive (A and B) and
preeclamptic women (C). No difference in intensity and distribution of
NADPH-diaphorase staining can be observed between normotensive
pregnancy and preeclampsia. (v): villi, (d): decidua. Original magnification:
A, 100x; B and C, 400x.
(panel D): Formation of [3H]L-citrulline from [3H]L-arginine in homogenate
placental tissue from normotensive pregnancy (n=9) and preeclamptic
women (n=6). Data are mean±SEM.

Plac
enta
l[3 H
]L-c
itrul
line
(nm
ol/m
g pr
otei
n)
C
(v)
B
(v)
A
(d)(v)
Figure I
normotensive pregnancy preeclampsia
0
0.2
0.4
0.6
0.8
1.0D