knockdown of the cochaperone sgta results in the suppression of androgen and pi3k/akt signaling and...
TRANSCRIPT

Knockdown of the cochaperone SGTA results in thesuppression of androgen and PI3KAkt signaling and inhibitionof prostate cancer cell proliferation
Andrew P Trotta1 Eleanor F Need1 Luke A Selth2 Samarth Chopra34 Carole B Pinnock3 Damien A Leach1
Gerhard A Coetzee56 Lisa M Butler2 Wayne D Tilley2 and Grant Buchanan1
1 Cancer Biology Group Freemasons Foundation Centre for Menrsquos Health Basil Hetzel Institute for Translational Health Research Department of Medicine
University of Adelaide Adelaide Australia2 Dame Roma Mitchell Cancer Research Laboratories Department of Medicine University of AdelaideHanson Institute Adelaide Australia3 Urology Unit Repatriation General Hospital South Australia Australia4 Department of Urology St Vincentrsquos Hospital and The Garven Institute Sydney New South Wales Australia5 Department of Preventative Medicine Keck School of Medicine Norris Cancer Centre University of Southern California Los Angeles CA6 Department of Urology Keck School of Medicine Norris Cancer Centre University of Southern California Los Angeles CA
Solid tumors have an increased reliance on Hsp70Hsp90 molecular chaperones for proliferation survival and maintenance of
intracellular signaling systems An underinvestigated component of the chaperone system is the tetratricopeptide repeat
(TPR)-containing cochaperone which coordinates Hsp70Hsp90 involvement on client proteins as well as having diverse indi-
vidual actions A potentially important cochaperone in prostate cancer (PCa) is small glutamine-rich TPR-containing protein
alpha (SGTA) which interacts with the androgen receptor (AR) and other critical cancer-related client proteins In this study
the authors used small interfering RNA coupled with genome-wide expression profiling to investigate the biological signifi-
cance of SGTA in PCa and its influence on AR signaling Knockdown of SGTA for 72 hr in PCa C4-2B cells significantly altered
expression of gt1900 genes (58 decreased) and reduced cell proliferation (p lt 005) The regulation of 35 of 5a-
dihydrotestosterone (DHT) target genes was affected by SGTA knockdown with gene-specific effects on basal or DHT-induced
expression or both Pathway analysis revealed a role for SGTA in p53 generic PCa and phosphoinositol kinase (PI3K) signaling
pathways the latter evident by a reduction in PI3K subunit p100b levels and decreased phosphorylated Akt Immunohisto-
chemical analysis of 64 primary advanced PCa samples showed a significant increase in the ARSGTA ratio in cancerous
lesions compared to patient-matched benign prostatic hyperplasia tissue (p lt 002) This study not only provides insight into
the biological actions of SGTA and its effect on genome-wide AR transcriptional activity and other therapeutically targeted
intracellular signaling pathways but also provides evidence for PCa-specific alterations in SGTA expression
Molecular chaperones encompass a collection of evolutionaryconserved proteins involved in establishing monitoring andmaintaining the biological activities of diverse client mole-cules Many are thus constitutively expressed (eg Hsp90)
and coordinate with each other in processes that include pro-tein folding maturation transport and maintenance of thehomeostatic balance of the intracellular proteome12 Cellularinsult andor intracellular stress can disrupt this homeostasis
Key words tetratricopeptide repeat cochaperones androgen signaling kinase signaling
Abbreviations AR androgen receptor DAVID Database for Annotation Visualization and Integrated Discovery DHT dihydrotestos-
terone GSEA gene set enrichment analysis PCa prostate cancer PI3 kinase phosphatidylinositide 3-kinase SGTA small glutamine-
rich tetratricopeptide repeat-containing protein alpha
Additional Supporting Information may be found in the online version of this article
Grant sponsor Prostate Cancer Foundation of Australia Grant numbers IDYI02 PG2210 Grant sponsor The National Health and
Medical Research Council of Australia Grant number ID453662 Grant sponsor US Department of Defense Grant number
IDPC060443 Grant sponsor HIHNCI Grant number R01CA136924 Grant sponsors US Prostate Cancer Foundation and The
Prostate Cancer Foundation of Australia (Young Investigator Awards) Grant number YI0810 Grant sponsors The Prostate Cancer
Foundation Foundation Daw Park
DOI 101002ijc28310
History Received 22 Jan 2013 Accepted 14 May 2013 Online 6 Jun 2013
Correspondence to Andrew P Trotta Cancer Biology Group Basil Hetzel Institute for Translational Research Department of Medicine
University of Adelaide 28 Woodville Road Woodville South Adelaide 5011 Australia Tel 161-8-82227426
E-mail andrewtrottaadelaideeduau
Can
cerCellBiology
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
International Journal of Cancer
IJC
leading to protein misfolding aggregation and the inappro-priate activation of signaling pathways During the develop-ment of pathophysiological states such as cancer increasedactivity of the chaperone machinery is one mechanism thatensures cell survival in an unstable intracellular environ-ment3ndash5 Targeting the chaperone system is therefore an ave-nue of therapeutic interest in many solid tumors includingprostate cancer (PCa)6ndash8 Currently however we know rela-tively little about the molecular and proteomic consequencesof altered chaperone expression
Within the microenvironment that surrounds tumor cellscytotoxic stressors such as hypoxia and altered nutrient levelsresult in the production of intracellular free radicals leadingto DNA damage genomic instability and in some cases cel-lular senescence and apoptosis349 To combat intracellularprotein aggregation resulting from cytotoxic stress cancercells upregulate Hsp90 and other chaperone proteins1011 Asa consequence there can be selection for and accumulationof cancer cells with diverse phenotypes that can for examplestabilize and protect different oncoproteins This facilitatesclonal escape from intracellular control mechanisms thatmediate apoptosis thus allowing the survival and prolifera-tion of cancer cells in a hypoxic nutrient limiting immuno-genic or toxic environment ultimately promoting metastasisand therapy resistance4
Chaperones such as Hsp90 do not act alone but interactwith accessory cochaperone molecules in larger multichaper-one protein complexes These cochaperones are necessary forthe establishment and functionality of the Hsp90 molecularchaperone complex and for its diverse activities An impor-tant subgroup of cochaperones contain evolutionary con-served tetratricopeptide repeat (TPR) proteinndashproteininteraction domains which enable them to act as functionaladaptormodulator molecules for Hsp90 on client proteinssuch as steroid receptors and serinethreonine kinases12 Assuch cochaperone proteins can directly and indirectly affectbroad cellular processes (eg growth proliferation differen-tiation cell cycling and apoptosis) as well as being more spe-cific for key signaling pathways An example of the latter isthe murine knockout of the cochaperone Fkbp4FKBP52 inwhich male and female mice display morphological and func-tional abnormalities almost exclusively in steroid hormone-sensitive tissue including the prostate testicles penis anduterus13ndash15 Recently a study of seven TPR cochaperones [E3
ubiquitin-protein ligase CHIP cyclophilin 40 FK506-bindingprotein 51 (FKBP51) and 52 (FKBP52) protein phosphatase5 DnaJ homology subfamily C member 7 (TPR2) and arylhydrocarbon interacting protein (XAP2)] demonstrated bothoverlapping and distinct effects on the function of six steroidreceptors [androgen receptor (AR) estrogen receptor a andb progesterone receptor mineralocorticoid receptor and glu-cocorticoid receptor]16 inferring a role for TPR cochaperonesin the tissue-specific responsiveness to steroid hormones
We previously identified the Hsp90 cochaperone smallglutamine-rich TPR-containing protein alpha (SGTA) as amodulator of AR function in PCa cells and found that SGTAprotein levels were lower in metastatic PCa compared tononmalignant or primary lesions1718 Based on these find-ings it is possible that an altered Hsp90SGTA ratio contrib-utes to maintenance or reactivation of AR in a low-hormoneenvironment such as during therapeutic androgen depriva-tion for metastatic PCa The broader biological consequencesof altered SGTA expression for AR function in PCa is thusof considerable interest SGTA is nonetheless expressed in awide variety of mammalian tissues including brain liverspleen prostate testis ovary colon and small intestine1719ndash23
it also interacts with tyrosine kinase receptors such as thehuman growth hormone receptor and platelet-derived growthfactor receptor alpha (PDGFRa)2425 and SGTA depletion inHeLa cells results in metaphase arrest and apoptosis1920
Therefore the biological effects of SGTA almost certainlyextend beyond steroid hormone signaling and likely playimportant physiological roles With the above considerationsthe objectives of our study were to characterize the genome-wide transcriptional response of PCa cells to altered SGTAexpression and the subsequent effect on AR signaling Ourwork has identified novel biological pathways regulated bySGTA and has allowed us to assess the relative importance ofits multiple functions in PCa
Material and MethodsCell culture and immunoblotting
The C4-2B cell lines derived from subcutaneous LNCaP xen-ografts in castrated nude mice as described in Ref 26 weremaintained in RPMI 1640 supplemented with 5 fetal bovineserum (FBS Sigma-Aldrich Sydney Australia) at 37C in 5CO2 All assays requiring steroids were performed in phenol-red free RPMI 1640 (Sigma-Aldrich) with 5 dextran-coated
Whatrsquos new
Solid tumors have an increased reliance on Hsp70Hsp90 molecular chaperones for proliferation survival and maintenance of
intracellular signaling systems An under-studied co-chaperone is SGTA which is an inhibitor of androgen receptor transcrip-
tional activity in prostate cancer cells This study presents new data showing that SGTA acts to regulate multiple oncogenic
signaling pathways in prostate cancer Specifically SGTA acts to maintain optimal expression of genes within the androgen
and PI3 kinase pathways SGTA is also required for maximal Akt activity and overall cell viability The biological relationships
between SGTA the androgen receptor and PI3Akt may be of importance in prostate tumorigenesis
Can
cerCellBiology
Trotta et al 2813
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
charcoal (DCC)-stripped FBS (stripped serum) Immunoblotswere performed as previously described1727 The followingprimary antibody dilutions were used SGTA (11000Abnova Neihu District Taiwan) AR (11000 AR N-20Santa Cruz Dallas TX) FKBP51 (11000 Santa Cruz) b-actin (12000 Sigma-Aldrich) Hsp90 (11000 Santa Cruz)p85 (11000 Cell Signaling Danvers MA) p110b (11000Cell Signaling) Akt (1300 Santa Cruz) p53 (11000 SantaCruz) p21 (11000 BD Biosciences San Jose CA) andMdm2 (11000 Santa Cruz) Phosphorylated Akt and p53antibodies were used at the following dilutions Akt threonine308 (Akt T308 12000 Cell Signaling) Akt serine 473 (AktS473 11000 Cell Signaling) and p53 serine 15 (p53 S1511000 Cell Signaling)
Small interfering RNA transfections RNA extraction and
quantitative real-time PCR
C4-2B cell lines (200000 cells per well in six-well plate) weretransfected in serum-free RPMI 1640 medium with 5 nMnonspecific (NS) control or 167 or 5 nM SGTA-specificsmall interfering RNA (siRNA Ambion Life TechnologiesMulgrove Australia) using Oligofectamine (Life Technolo-gies) for 4 hr according to the manufacturerrsquos instructionsCells were then supplemented with 5 FBS and incubatedfor 72 hr followed by treatment with vehicle (ethanol) controlor 001ndash10 nM 5a-dihydrotestosterone (DHT) for 16 hrRNA was isolated using the RNeasy RNA isolation kit (Qia-gen Melbourne Australia) and 1 lg total RNA was synthe-sized to cDNA using the iScript cDNA synthesis kit (Bio-RadLaboratories Gladesville Australia) Samples were analyzedin triplicate by quantitative real-time PCR (qPCR) as previ-ously described18 Sequences for all primers are listed in Sup-porting Information Table S1
Microarray and pathway analysis
C4-2B cells transfected with siRNA as detailed above weregrown in stripped serum containing vehicle control or 1 nMDHT for 16 hr Three hundred nanograms of total RNAextracted from each of five replicates (vehicle treated) orpooled from five replicates (DHT treated) were analyzedusing Affymetrix Human Gene 10 ST Arrays Bioinformaticswas performed in R using Bioconductor and the Limmapackage28 Briefly array data were normalized using robustmultiarray average (RMA) and filtered by mapped probesand an arbitrary minimum expression threshold wasobtained and genes different between groups called byBenjaminindashHochberg-adjusted p-values were determinedfrom Bayesian linear regression modeling Gene pathwayanalysis was conducted using the Ingenuity Pathway Analysis(IPA) program (Ingenuity Systems Redwood City CA)Gene Set Enrichment Analysis (GSEA Broad Institute httpwwwbroadinstituteorggsea)29 and Database for AnnotationVisualization and Integrated Discovery (DAVID httpwwwdavidabccncifcrfgov)30 For GSEA a false discovery rate of
25 was used and the number of random tests or permuta-tions was set at 1000 (microarray data GEO accession num-ber GSE43521)
Cell viability assays
C4-2B cells (24000 cells per well in 24-well plates) weretransfected with 5 nM NS control siRNA or three independ-ent SGTA siRNAs at a concentration of 167 nM Cells weretransfected in medium containing either 5 DCC-FBS or 5FBS then counted via a hemocytometer and assayed withtrypan blue exclusion at the end of each time point
MTT assays
For MTT assays cells were transfected as described above inmedium containing 5 FBS At the end of each time point500 ll of serum-free PRF-RPMI containing 025 mgml MTTreagent (Sigma-Aldrich) was added to the wells and incu-bated for 2 hr The MTT reagent was aspirated and replacedwith 100 ll DMSO Cell viability was then determined bymeasuring absorbance at 570 nm using a microplate reader(Bio-Rad Laboratories) Two-way ANOVA was used to com-pare differences in means between the no siRNA (mock) NSand SGTA siRNA-transfected groups Significance was set atp lt 005
Clinical cohort and immunohistochemistry
The study sample was identified from all men undergoing atransurethral resection of the prostate (TURP) for benignprostatic hyperplasia (BPH) between 2000 and 2007 at theRepatriation General Hospital (RGH South Australia) Thosefor whom a first diagnosis of PCa was made based on pathol-ogy at TURP were eligible (n 5 105) Those patients with aGleason grading and gt5 cancer at histology (n 5 66) wereselected Areas of BPH and cancer were identified in each byhematoxylin and eosin staining and mapped onto archivedparaffin-embedded material by one pathologist All protocolswere approved by the Flinders Medical Centre AdelaideSouth Australia and the RGH ethics committee (Protocol04210) A series of six tissue microarrays were constructedin duplicate and 15-mm diameter by 3-mm deep cores wereextracted from each pathologist-defined areas of BPH andcancer and embedded across six recipient paraffin blockswith 46 samples per block Two additional PCa samples andone liver sample were included as controls for each recipientblock Immunohistochemistry was performed on the cohortwith AR (AR-N20 1100 Santa Cruz) or SGTA (11000Protein Tech Chicago IL) antisera detected with LSAB1
System HRP kit (Dako Laboratories Noble Park Australia)according to the manufacturerrsquos guidelines The immunoreac-tivity for AR and SGTA was scored in a minimum of threefields in both BPH and cancer regions independently by tworesearchers (APT and DAL) Epithelia was scored 0ndash3(none low moderate and high) whereas stroma was scored0ndash2 (none low and high) Data were collated into samplescores for epithelia (0ndash9 scale) and stroma (0ndash6) scale in
Can
cerCellBiology
2814 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
both cancer and BPH regions with the difference in scoringrange due to the less dynamic staining in the stroma in com-parison to epithelia The relationship between AR and SGTAimmunoreactivity and clinical parameters including Gleasonscore serum PSA levels percent cancer prostate size clinicalstage and cancer-related death was investigated using theWilcox Rank Sum test and Barnardrsquos Exact test The WilcoxRank Sum test was also used to determine the differencebetween medians for AR and SGTA immunoreactivity inBPH specimens compared to patient-matched cancer sam-ples Significance was set at p lt 005
ResultsKnockdown of SGTA regulates the transcriptome of C4-2B
cells
To dissect the biological actions of SGTA in C4-2B PCa cellswe examined genetic changes following siRNA knockdown of
SGTA We used a low-concentration pool of three commer-cial siRNAs targeting SGTA exons 5 8 and 9 to reduce non-specific effects associated with individual siRNAs (Fig 1a)31
Transfection of the siRNA combination (167 nM each) wasas effective as each siRNA alone (si1 si2 and si3 5nM each)in reducing SGTA steady-state protein levels at 72 hr in PCaC4-2B cells (Fig 1a) For expression profiling we extractedRNA from five independent biological replicates of C4-2Bcells transfected with the SGTA siRNA pool or a NS siRNAcontrol and treated with 1 nM DHT or vehicle control for 16hr qPCR analysis revealed an average of 825 SGTAmRNA knockdown with a variance of less than 2 betweensamples (Supporting Information Fig S1A) Global expres-sion profiling using Affymetrix microarrays demonstratedthat knockdown of SGTA significantly (p lt 005) altered theexpression of 2283 transcripts in C4-2B cells in the vehicle-treated experiment and 2156 transcripts in cells treated with
Figure 1 Knockdown of SGTA modulates global androgen signaling in conjunction with the overall expression of the C4-2B transcriptome
(a) Schematic of SGTA gene located on chromosome 19 Blue boxes represent exons separated by noncoding regions SGTA siRNA1 2 and
3 (si1 si2 and si3) and their respective target exons 5 8 and 9 are indicated Immunoblot analysis for SGTA protein levels was performed
in lysates collected from C4-2B cells that had been transfected for 72 hr with a nonspecific (NS) control siRNA one of the three SGTA siR-
NAs alone (si1 si2 and si3 5nM each) or a combination of the three SGTA siRNAs (siC each at a final concentration of 167 nM) (b) Affy-
metrix expression microarray profiling of C4-2B cells after treatment for 72 hr with the siSGTA siRNA pool compared to the NS control
(BenjaminindashHochberg-adjusted p lt 005) The Euler diagram shows the number of transcripts significantly affected in experiments with
vehicle control (VC n 5 5) versus DHT (n 5 2) The number and percentage of DHT-regulated transcripts in each set is shown in brackets
(cndashe) For each of the three Euler categories shown in (b) pie charts show the proportion of genes with increased or decreased expression
following siSGTA compared to NS siRNA The corresponding volcano plots show the degree of that effect in terms of log 2-fold change for
siSGTA compared to NS siRNA against p-value for that difference Within those categories genes are annotated by those known to be sig-
nificantly DHT regulated in C4-2B cells (red circles) or not (green circles)
Can
cerCellBiology
Trotta et al 2815
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
DHT with 950 affected in both cases (Fig 1b) The effect ofSGTA knockdown in both treatment groups was validated byqPCR in an independently generated RNA sample set (p lt005 Supporting Information Figs S2AndashS2C)
We next cross-referenced these data with DHT-regulatedgenes previously identified from microarray analysis in C4-2B cells32 Of DHT-responsive targets that satisfied aBenjaminndashHochberg-adjusted statistic of p lt 005 in that ear-lier experiment 2262 fulfilled the minimum threshold cutoffcriteria of expression on the Affymetrix arrays performedhere (12 of assessed transcripts Fig 1b and SupportingInformation Fig 1B) Of the 3489 transcripts identifiedacross the siSGTA vehicle- and DHT-treated experiments23 (2751206) are predicted as DHT targets a number thatrises to 28 (275950) of those common to vehicle controland DHT SGTA (Fig 1b) The numbers are similar whengenes rather than transcripts are considered (SupportingInformation Fig S1C) and importantly represents an approx-imate twofold enrichment for DHT-responsive genesamongst those affected by SGTA knockdown (v2 5 209two-tailed p-value lt 00001)
In assessing both the qualitative and quantitative effects ofsiSGTA it was determined that for 63 of affected tran-scripts in vehicle-treated cells 58 from DHT-treated cellsand 68 of those 950 transcripts common to both the effectof SGTA knockdown was to decrease expression (pie chartsin Figs 1cndash1e) Volcano plots of the data show that the effectof SGTA knockdown occurs on a continuum and impor-tantly that there is a higher proportion of androgen-regulatedgenes (red circles) amongst those transcripts repressed bysiRNA against SGTA (299) than those increased by theknockdown (178) These data imply that SGTA may play akey role in maintaining andor limiting the androgen-regulated program in C4-2B cells however these effects arelimited to a subset of androgen-target loci
Loss of SGTA results in subtle changes to androgen
signaling in PCa cells
We elucidated the global effect of SGTA on the DHT-regulated transcriptome in C4-2B cells Of 2385 known DHTtargets 66 (15752385) were unaffected by depletion ofSGTA (Fig 2a green circles) Of the remaining 810 expres-sion of approximately one-third was altered at basal (vehicle-treated cells red circles) DHT induced (DHT-treated cellsyellow circles) or in both states (blue circles p lt 005 Fig2a) qPCR analysis of three androgen-regulated genes(S100A10 LRRC31 and DNASE2B) from an independentlygenerated RNA sample set validated these findings (p lt 005Fig 2b)
A more detailed analysis of the array data above showedthat ablation of SGTA had very little effect on DHT regula-tion of classic AR-regulated genes such as KLK3 and FKBP5This finding suggests that SGTA may exert only a relativelysmall effect on AR signaling andor is only evident on a sub-set of AR-regulated genes as had been suggested by our
previous analysis with reporter genes18 To investigate thisfurther SGTA was knocked down in C4-2B cells and RNAwas extracted following treatment with increasing doses ofDHT With an approximate 70 depletion in SGTA acrossall samples (Fig 3a) the regulation of KLK3 or FKBP5expression by qPCR was not significantly affected at any one
Figure 2 Dichotomized effect of SGTA knockdown on androgen-
regulated genes (a) Androgen-regulated genes identified from C4-
2B cells were segregated based on the effect of SGTA knockdown
in either vehicle- or DHT-treated cells Insert Green dots (unaf-
fected) represent androgen-regulated genes unaffected by SGTA
red dots (basal only) are androgen-regulated genes significantly
affected in vehicle-treated cells only yellow dots (induced only)
represent androgen-regulated genes significantly affected by SGTA
knockdown in DHT-treated cells only and blue dots (both) repre-
sent those androgen-regulated genes significantly affected by SGTA
knockdown in both vehicle- and DHT-treated cells Data represent
pairwise comparison of log 2-fold change (siSGTAsiNS) in vehicle-
and DHT-treated cells (b) The effect of SGTA knockdown was
assessed by qPCR on androgen-regulated genes identified in (a)
Gene expression was normalized in GAPDH Data represent the
mean (6 SEM) of four biological replicates p lt 005 on Mann-
Whitney U test for difference between medians of SGTA siRNA in
comparison to NS siRNA
Can
cerCellBiology
2816 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
dose of DHT (Figs 3b and 3c) However a consistent smalleffect of SGTA knockdown on regulation across all DHTconcentrations was observed (p lt 005 Figs 3b and 3c)although no overt effect was observed at FKBP51 steady-stateprotein levels (Fig 3d) Collectively these data confirm thatSGTA exerts an influence on only a subset of androgen-regulated genes and suggests that its effects on AR proteinfolding and trafficking in the presence of fluctuating andro-gen levels may hold greater consequence for PCa cellfunction
Knockdown of SGTA modulates the expression of p53 and
phosphoinositol kinaseAkt pathways
Although the above results suggest that SGTA is a key regu-lator of a subset of AR target genes we sought to elucidateits broader biological significance in PCa cells Ingenuitypathway analysis of the microarray data identified a numberof important cellular pathways significantly altered by SGTAknockdown in vehicle- or DHT-treated cells or both (Sup-porting Information Fig S3) Similar pathways were enrichedin the gene sets obtained from both vehicle- and DHT-
treated cells implying a relatively consistent effect of SGTAthat is largely independent of exogenous hormone Amongthose pathways the most highly enriched were p53 PCa andphosphoinositol kinase (PI3K)Akt signaling pathways (p lt
005 Fig 4a) Enrichment of these pathways was confirmedfor both DHT- and vehicle-treated cells using GSEA29 (Figs4bndash4d) and DAVID30 (Supporting Information Tables S2 andS3) GSEA further implicated upregulation of PCa and PI3Kpathways by SGTA knockdown but downregulation of p53signaling in both vehicle- and DHT-treated gene sets (Figs4bndash4d and Supporting Information Fig S4) Additionally thetop four of five protein families enriched in SGTA-depletedsamples related to protein kinase or serinethreonine kinaseactivity in DAVID (Supporting Information Tables S4 andS5) suggesting that SGTA may play a role in maintainingandor modulating these activities in PCa cells IndeedSGTA has been shown to maintain receptor tyrosine kinasesignaling and Akt activity in a nonsmall cell lung cancer(NSCLC) cell line1725
The effect of SGTA knockdown on specific genes withineach of the three GSEA pathways listed above was validated
Figure 3 SGTA knockdown does not affect AR-dependent KLK3 and FKBP5 gene expression (andashc) The effect of SGTA knockdown on AR sig-
naling in C4-2B cells was assessed by qPCR Cells were transfected with 5 nM NS or SGTA siRNA for 72 hr followed by treatment with 0ndash10
nM DHT for 16 hr prior to RNA extraction qPCR was performed on SGTA KLK3 and FKBP5 in triplicate samples Error bars represent (6
SEM) of three biological replicates Statistical significance represents Friedman repeated measures test for differences in means between
SGTA siRNA and NS siRNA p lt 005 (d) Matching protein lysates to the RNA samples used in (a)ndash(c) were generated for immunoblot anal-
ysis on SGTA FKBP51 and AR steady-state protein levels Hsp90 was used as a loading control
Can
cerCellBiology
Trotta et al 2817
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
Figure 4 SGTA knockdown affects p53 PI3 kinase and prostate cancer signaling pathways (a) Ingenuity pathway analysis of canonical
pathways affected by SGTA knockdown in vehicle- and DHT-treated cells or both (bndashd) Association of SGTA knockdown vehicle-treated
datasets in prostate cancer phosphatidylinositol signaling system and p53 signaling as a pathway as assessed by GSEA Probe sets from
the SGTA knockdown arrays were collapsed to the gene level and assigned a score based on a signal-to-noise ratio algorithm and ranked-
ordered by this score29 The running enrichment score (green line) is then plotted (top panel) The enrichment of the KEGG Prostate Cancer
gene set in the ranked-ordered SGTA knockdown dataset is represented by black lines (middle panel) positive to negative value of ranking
metric and the correlation of genes within the KEGG pathway to SGTA knockdown dataset (bottom panel) The normalized enrichment score
(NES) for enriched pathways is presented in each panel qPCR validation of genes is associated with both KEGG pathways Data represent
mean (6 SEM) of four biological replicates p lt 005 and p lt 001 on Mann-Whitney U test for difference between medians of SGTA
siRNA in comparison to NS siRNA
Can
cerCellBiology
2818 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
using qPCR with some variation depending on treatmentwith vehicle or DHT Specifically analysis of PCa signalingpathway genes AR GRB2 and ERBB2 demonstrated an effectonly in the presence of DHT (p lt 005 Fig 4b) For thePI3K signaling pathway PIK3R1 and PIK3CB expressiondecreased slightly but was significantly affected with DHTtreatment but only in the knockdown samples PIK3CDexpression in contrast was affected by both vehicle and DHTtreatments (p lt 001 Fig 4c) Finally we observed upregula-tion of TP53 with SGTA knockdown but a significantdecrease in expression of the two p53 target genes MDM2and CDKN1A (also known as p21 p lt 005 Fig 4d) Theabove data implicate SGTA in a number of pathways previ-ously shown to be essential to PCa cell survival proliferationand metastasis The loss of SGTA in PCa might thus be pre-dicted to have a significant effect on disease progression
SGTA acts as a positive regulator of Akt activity and C4-2B
cell viability
We next investigated the effect of SGTA knockdown on p53and PI3 kinase pathways in C4-2B cells by assessing pathwaycomponent proteins and their downstream targets Analysisof p53 signaling by immunoblotting did not reveal anychanges in steady-state protein level of the p53 target p21whereas SGTA knockdown appears to reduce Mdm2 levelseither with or without DHT (Fig 5a) However neither totalp53 nor phosphorylated p53 (S15) levels were altered bySGTA knockdown (Fig 5a) This disconnection with themRNA expression data may indicate a specific temporalndashspa-tial protein response not captured in these studies In addi-tion SGTA knockdown did not affect PI3 kinase subunit p85(PI3KR1) levels but decreased basal p110b (PI3KCB) proteinlevels (Fig 5b) Consistent with a recent report that p110b
maintains Akt activity in PCa cells as measured by phospho-rylation of the T308 and S473 residues of Akt33 the decreasein p110b with SGTA knockdown translated to reduced levelsof phosphorylated Akt T308 and Akt S473 respectively (Fig5b) These effects were partially reversed by treatment with 1nM DHT but could not be attributed to a decrease in totalAkt The effect of SGTA knockdown on p110b protein levelsand Akt phosphorylation status was verified in LNCaP PCacells (Supporting Information Fig S5A)
Given the role of SGTA in key signaling pathways wenext investigated the role of SGTA in PCa cell growthAlthough SGTA knockdown did not affect viability of C4-2Bcells when assayed under stripped serum conditions (Fig 5c)a 524 decrease in siSGTA cell number compared to mockcontrol at Day 6 was observed when assays were performedwith normal FBS containing media (p lt 005 Fig 5d) Theeffect of SGTA knockdown on C4-2B cell proliferation in thepresence of normal FBS was validated by MTT assays (Sup-porting Information Fig S5B) Immunoblot analysis demon-strated SGTA depletion for the duration of both experiments(Figs 5e and 5f) whereas trypan-blue exclusion showed thatcell death was relatively consistent across all treatment groups
(Supporting Information Figs S5C and S5D) The role ofSGTA appears to be dependent on growth of cells in fullserum which might be consistent with activation of growthfactor signaling pathways dependent on intracellular PI3KAkt signaling
SGTA and AR protein levels in patient-matched BPH and
cancer samples
The convergence of SGTA on AR and PIK3Akt signalingpathways which have important roles in PCa etiology sug-gests that altered SGTA levels may critically influence PCaprogression Our previous study indirectly suggested this bydemonstration of lower SGTA protein levels in metastaticsamples compared to primary disease17 Immunohistochemis-try was used in our study on a clinical cohort of 66 primaryadvanced PCa samples and patient-matched BPH tissues tomore precisely define any relationship between SGTA andAR levels and clinically significant outcome parameters
AR protein was found to be predominately localized inthe nucleus of luminal epithelial cells of both BPH and PCasamples with some variable stromal staining (Fig 6a) Incontrast SGTA was mainly confined to epithelial cytoplasmalthough a small number of samples showed stromal staining(Fig 6a) Quantitative analysis identified a slightly highermean AR level in cancer samples compared to BPH andlower SGTA but these did not reach significance (p 5 0055Supporting Information Table S6) Importantly however themean and median ARSGTA ratio was significantly higher inPCa samples than in matched BPH (p 5 0017) This ratiowas also higher in cancer but not BPH of those that haddied from PCa but was not significant (Supporting Informa-tion Table S6) These data imply a cancer-specific dysregula-tion in the expression of AR and SGTA that may potentiatedisease progression
DiscussionOne of the key hallmarks of cancer is the upregulation andmaintenance of key signal transduction pathways that drivecancer cell proliferation and survival5 The molecular chaper-one and cochaperone systems facilitates this in cancer cellsby providing key structural and functional frameworksthrough which these highly proliferative signals can be sus-tained and amplified The PI3KAkt signaling pathway has awell-defined role in PCa initiation and progression3435
whereby approximately 40 of localized PCa and 70 ofmetastases contain genomic alterations within PI3 kinase sig-naling proteins36 Importantly the molecular mechanismsthat contribute to constitutive PI3 kinase signaling haveunderscored recent clinical advances in the development andadministration of therapeutic agents for the treatment ofmetastatic PCa37 The Class 1A PI3 kinase subunits are agroup of enzymes composed of regulatory (p85a p85b p55a
and p55g) subunit proteins that interact with their catalytic(p110a p110b and p110d) counterparts to form heterodi-meric complexes Activation of these complexes by tyrosine
Can
cerCellBiology
Trotta et al 2819
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
kinase receptors at the cell surface mediates subsequent signaltransduction (via Akt phosphorylation) culminating in theregulation of target genes involved in cell survival prolifera-tion growth and migration PI3 kinase subunit p110b isimportant for the maintenance of Akt phosphorylation in PCcells33 and of significance we demonstrated here that regula-tion of cellular p110b levels by SGTA may be important inmaintaining Akt phosphorylation A similar effect of SGTAknockdown on Akt phosphorylation has been observed inNSCLC H1703 cells25 suggesting that SGTA acts as an
important positive regulator of PI3KAkt signaling in multi-ple cancers and can thereby mediate cancer cell proliferationand survival
In addition to the critical role of PI3 kinase signaling inPCa development there is incontrovertible evidence demon-strating that continued AR signaling is required for diseaseprogression to a castrate-resistant state38 Although it isaccepted that regulation of AR transcriptional activity in PCacells can be achieved at multiple stages within the AR activa-tion cascade39 the role of cytoplasmic TPR-containing
Figure 5 Knockdown of SGTA affects PI3K activity and C4-2B cell viability (a and b) C4-2B cells were transfected with siRNA and treated as
described in Figure 1a Immunoblot analysis of SGTA p21 Mdm2 p53 p85 p110b and Akt protein steady-state levels Analysis of active
p53 and Akt signaling was determined by phosphorylation status of p53 serine 15 (S15) Akt threonine 308 (T308) and Akt S473 residues
Hsp90 was used as a loading control (c and d) C4-2B cells were either mock transfected or transfected with nonspecific (NS) siRNA or
SGTA-specific siRNA in serum-free medium for 4 hr and then supplemented with 5 DCS-FBS or FBS Cells were cultured for 6 days and
counted every 2 days using a hemocytometer and cell viability was assessed by Trypan blue exclusion Data are presented as the mean
(6 SEM) of triplicate wells from two independent experiments Two-way ANOVA was used to determine the difference in cell growth
between groups p lt 005 differences between mockNS transfected cells versus SGTA siRNA (e and f) Cell lysates from growth assay
were assessed for SGTA steady-state protein levels b-Actin was used as a loading control [Color figure can be viewed in the online issue
which is available at wileyonlinelibrarycom]
Can
cerCellBiology
2820 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC

leading to protein misfolding aggregation and the inappro-priate activation of signaling pathways During the develop-ment of pathophysiological states such as cancer increasedactivity of the chaperone machinery is one mechanism thatensures cell survival in an unstable intracellular environ-ment3ndash5 Targeting the chaperone system is therefore an ave-nue of therapeutic interest in many solid tumors includingprostate cancer (PCa)6ndash8 Currently however we know rela-tively little about the molecular and proteomic consequencesof altered chaperone expression
Within the microenvironment that surrounds tumor cellscytotoxic stressors such as hypoxia and altered nutrient levelsresult in the production of intracellular free radicals leadingto DNA damage genomic instability and in some cases cel-lular senescence and apoptosis349 To combat intracellularprotein aggregation resulting from cytotoxic stress cancercells upregulate Hsp90 and other chaperone proteins1011 Asa consequence there can be selection for and accumulationof cancer cells with diverse phenotypes that can for examplestabilize and protect different oncoproteins This facilitatesclonal escape from intracellular control mechanisms thatmediate apoptosis thus allowing the survival and prolifera-tion of cancer cells in a hypoxic nutrient limiting immuno-genic or toxic environment ultimately promoting metastasisand therapy resistance4
Chaperones such as Hsp90 do not act alone but interactwith accessory cochaperone molecules in larger multichaper-one protein complexes These cochaperones are necessary forthe establishment and functionality of the Hsp90 molecularchaperone complex and for its diverse activities An impor-tant subgroup of cochaperones contain evolutionary con-served tetratricopeptide repeat (TPR) proteinndashproteininteraction domains which enable them to act as functionaladaptormodulator molecules for Hsp90 on client proteinssuch as steroid receptors and serinethreonine kinases12 Assuch cochaperone proteins can directly and indirectly affectbroad cellular processes (eg growth proliferation differen-tiation cell cycling and apoptosis) as well as being more spe-cific for key signaling pathways An example of the latter isthe murine knockout of the cochaperone Fkbp4FKBP52 inwhich male and female mice display morphological and func-tional abnormalities almost exclusively in steroid hormone-sensitive tissue including the prostate testicles penis anduterus13ndash15 Recently a study of seven TPR cochaperones [E3
ubiquitin-protein ligase CHIP cyclophilin 40 FK506-bindingprotein 51 (FKBP51) and 52 (FKBP52) protein phosphatase5 DnaJ homology subfamily C member 7 (TPR2) and arylhydrocarbon interacting protein (XAP2)] demonstrated bothoverlapping and distinct effects on the function of six steroidreceptors [androgen receptor (AR) estrogen receptor a andb progesterone receptor mineralocorticoid receptor and glu-cocorticoid receptor]16 inferring a role for TPR cochaperonesin the tissue-specific responsiveness to steroid hormones
We previously identified the Hsp90 cochaperone smallglutamine-rich TPR-containing protein alpha (SGTA) as amodulator of AR function in PCa cells and found that SGTAprotein levels were lower in metastatic PCa compared tononmalignant or primary lesions1718 Based on these find-ings it is possible that an altered Hsp90SGTA ratio contrib-utes to maintenance or reactivation of AR in a low-hormoneenvironment such as during therapeutic androgen depriva-tion for metastatic PCa The broader biological consequencesof altered SGTA expression for AR function in PCa is thusof considerable interest SGTA is nonetheless expressed in awide variety of mammalian tissues including brain liverspleen prostate testis ovary colon and small intestine1719ndash23
it also interacts with tyrosine kinase receptors such as thehuman growth hormone receptor and platelet-derived growthfactor receptor alpha (PDGFRa)2425 and SGTA depletion inHeLa cells results in metaphase arrest and apoptosis1920
Therefore the biological effects of SGTA almost certainlyextend beyond steroid hormone signaling and likely playimportant physiological roles With the above considerationsthe objectives of our study were to characterize the genome-wide transcriptional response of PCa cells to altered SGTAexpression and the subsequent effect on AR signaling Ourwork has identified novel biological pathways regulated bySGTA and has allowed us to assess the relative importance ofits multiple functions in PCa
Material and MethodsCell culture and immunoblotting
The C4-2B cell lines derived from subcutaneous LNCaP xen-ografts in castrated nude mice as described in Ref 26 weremaintained in RPMI 1640 supplemented with 5 fetal bovineserum (FBS Sigma-Aldrich Sydney Australia) at 37C in 5CO2 All assays requiring steroids were performed in phenol-red free RPMI 1640 (Sigma-Aldrich) with 5 dextran-coated
Whatrsquos new
Solid tumors have an increased reliance on Hsp70Hsp90 molecular chaperones for proliferation survival and maintenance of
intracellular signaling systems An under-studied co-chaperone is SGTA which is an inhibitor of androgen receptor transcrip-
tional activity in prostate cancer cells This study presents new data showing that SGTA acts to regulate multiple oncogenic
signaling pathways in prostate cancer Specifically SGTA acts to maintain optimal expression of genes within the androgen
and PI3 kinase pathways SGTA is also required for maximal Akt activity and overall cell viability The biological relationships
between SGTA the androgen receptor and PI3Akt may be of importance in prostate tumorigenesis
Can
cerCellBiology
Trotta et al 2813
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
charcoal (DCC)-stripped FBS (stripped serum) Immunoblotswere performed as previously described1727 The followingprimary antibody dilutions were used SGTA (11000Abnova Neihu District Taiwan) AR (11000 AR N-20Santa Cruz Dallas TX) FKBP51 (11000 Santa Cruz) b-actin (12000 Sigma-Aldrich) Hsp90 (11000 Santa Cruz)p85 (11000 Cell Signaling Danvers MA) p110b (11000Cell Signaling) Akt (1300 Santa Cruz) p53 (11000 SantaCruz) p21 (11000 BD Biosciences San Jose CA) andMdm2 (11000 Santa Cruz) Phosphorylated Akt and p53antibodies were used at the following dilutions Akt threonine308 (Akt T308 12000 Cell Signaling) Akt serine 473 (AktS473 11000 Cell Signaling) and p53 serine 15 (p53 S1511000 Cell Signaling)
Small interfering RNA transfections RNA extraction and
quantitative real-time PCR
C4-2B cell lines (200000 cells per well in six-well plate) weretransfected in serum-free RPMI 1640 medium with 5 nMnonspecific (NS) control or 167 or 5 nM SGTA-specificsmall interfering RNA (siRNA Ambion Life TechnologiesMulgrove Australia) using Oligofectamine (Life Technolo-gies) for 4 hr according to the manufacturerrsquos instructionsCells were then supplemented with 5 FBS and incubatedfor 72 hr followed by treatment with vehicle (ethanol) controlor 001ndash10 nM 5a-dihydrotestosterone (DHT) for 16 hrRNA was isolated using the RNeasy RNA isolation kit (Qia-gen Melbourne Australia) and 1 lg total RNA was synthe-sized to cDNA using the iScript cDNA synthesis kit (Bio-RadLaboratories Gladesville Australia) Samples were analyzedin triplicate by quantitative real-time PCR (qPCR) as previ-ously described18 Sequences for all primers are listed in Sup-porting Information Table S1
Microarray and pathway analysis
C4-2B cells transfected with siRNA as detailed above weregrown in stripped serum containing vehicle control or 1 nMDHT for 16 hr Three hundred nanograms of total RNAextracted from each of five replicates (vehicle treated) orpooled from five replicates (DHT treated) were analyzedusing Affymetrix Human Gene 10 ST Arrays Bioinformaticswas performed in R using Bioconductor and the Limmapackage28 Briefly array data were normalized using robustmultiarray average (RMA) and filtered by mapped probesand an arbitrary minimum expression threshold wasobtained and genes different between groups called byBenjaminindashHochberg-adjusted p-values were determinedfrom Bayesian linear regression modeling Gene pathwayanalysis was conducted using the Ingenuity Pathway Analysis(IPA) program (Ingenuity Systems Redwood City CA)Gene Set Enrichment Analysis (GSEA Broad Institute httpwwwbroadinstituteorggsea)29 and Database for AnnotationVisualization and Integrated Discovery (DAVID httpwwwdavidabccncifcrfgov)30 For GSEA a false discovery rate of
25 was used and the number of random tests or permuta-tions was set at 1000 (microarray data GEO accession num-ber GSE43521)
Cell viability assays
C4-2B cells (24000 cells per well in 24-well plates) weretransfected with 5 nM NS control siRNA or three independ-ent SGTA siRNAs at a concentration of 167 nM Cells weretransfected in medium containing either 5 DCC-FBS or 5FBS then counted via a hemocytometer and assayed withtrypan blue exclusion at the end of each time point
MTT assays
For MTT assays cells were transfected as described above inmedium containing 5 FBS At the end of each time point500 ll of serum-free PRF-RPMI containing 025 mgml MTTreagent (Sigma-Aldrich) was added to the wells and incu-bated for 2 hr The MTT reagent was aspirated and replacedwith 100 ll DMSO Cell viability was then determined bymeasuring absorbance at 570 nm using a microplate reader(Bio-Rad Laboratories) Two-way ANOVA was used to com-pare differences in means between the no siRNA (mock) NSand SGTA siRNA-transfected groups Significance was set atp lt 005
Clinical cohort and immunohistochemistry
The study sample was identified from all men undergoing atransurethral resection of the prostate (TURP) for benignprostatic hyperplasia (BPH) between 2000 and 2007 at theRepatriation General Hospital (RGH South Australia) Thosefor whom a first diagnosis of PCa was made based on pathol-ogy at TURP were eligible (n 5 105) Those patients with aGleason grading and gt5 cancer at histology (n 5 66) wereselected Areas of BPH and cancer were identified in each byhematoxylin and eosin staining and mapped onto archivedparaffin-embedded material by one pathologist All protocolswere approved by the Flinders Medical Centre AdelaideSouth Australia and the RGH ethics committee (Protocol04210) A series of six tissue microarrays were constructedin duplicate and 15-mm diameter by 3-mm deep cores wereextracted from each pathologist-defined areas of BPH andcancer and embedded across six recipient paraffin blockswith 46 samples per block Two additional PCa samples andone liver sample were included as controls for each recipientblock Immunohistochemistry was performed on the cohortwith AR (AR-N20 1100 Santa Cruz) or SGTA (11000Protein Tech Chicago IL) antisera detected with LSAB1
System HRP kit (Dako Laboratories Noble Park Australia)according to the manufacturerrsquos guidelines The immunoreac-tivity for AR and SGTA was scored in a minimum of threefields in both BPH and cancer regions independently by tworesearchers (APT and DAL) Epithelia was scored 0ndash3(none low moderate and high) whereas stroma was scored0ndash2 (none low and high) Data were collated into samplescores for epithelia (0ndash9 scale) and stroma (0ndash6) scale in
Can
cerCellBiology
2814 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
both cancer and BPH regions with the difference in scoringrange due to the less dynamic staining in the stroma in com-parison to epithelia The relationship between AR and SGTAimmunoreactivity and clinical parameters including Gleasonscore serum PSA levels percent cancer prostate size clinicalstage and cancer-related death was investigated using theWilcox Rank Sum test and Barnardrsquos Exact test The WilcoxRank Sum test was also used to determine the differencebetween medians for AR and SGTA immunoreactivity inBPH specimens compared to patient-matched cancer sam-ples Significance was set at p lt 005
ResultsKnockdown of SGTA regulates the transcriptome of C4-2B
cells
To dissect the biological actions of SGTA in C4-2B PCa cellswe examined genetic changes following siRNA knockdown of
SGTA We used a low-concentration pool of three commer-cial siRNAs targeting SGTA exons 5 8 and 9 to reduce non-specific effects associated with individual siRNAs (Fig 1a)31
Transfection of the siRNA combination (167 nM each) wasas effective as each siRNA alone (si1 si2 and si3 5nM each)in reducing SGTA steady-state protein levels at 72 hr in PCaC4-2B cells (Fig 1a) For expression profiling we extractedRNA from five independent biological replicates of C4-2Bcells transfected with the SGTA siRNA pool or a NS siRNAcontrol and treated with 1 nM DHT or vehicle control for 16hr qPCR analysis revealed an average of 825 SGTAmRNA knockdown with a variance of less than 2 betweensamples (Supporting Information Fig S1A) Global expres-sion profiling using Affymetrix microarrays demonstratedthat knockdown of SGTA significantly (p lt 005) altered theexpression of 2283 transcripts in C4-2B cells in the vehicle-treated experiment and 2156 transcripts in cells treated with
Figure 1 Knockdown of SGTA modulates global androgen signaling in conjunction with the overall expression of the C4-2B transcriptome
(a) Schematic of SGTA gene located on chromosome 19 Blue boxes represent exons separated by noncoding regions SGTA siRNA1 2 and
3 (si1 si2 and si3) and their respective target exons 5 8 and 9 are indicated Immunoblot analysis for SGTA protein levels was performed
in lysates collected from C4-2B cells that had been transfected for 72 hr with a nonspecific (NS) control siRNA one of the three SGTA siR-
NAs alone (si1 si2 and si3 5nM each) or a combination of the three SGTA siRNAs (siC each at a final concentration of 167 nM) (b) Affy-
metrix expression microarray profiling of C4-2B cells after treatment for 72 hr with the siSGTA siRNA pool compared to the NS control
(BenjaminindashHochberg-adjusted p lt 005) The Euler diagram shows the number of transcripts significantly affected in experiments with
vehicle control (VC n 5 5) versus DHT (n 5 2) The number and percentage of DHT-regulated transcripts in each set is shown in brackets
(cndashe) For each of the three Euler categories shown in (b) pie charts show the proportion of genes with increased or decreased expression
following siSGTA compared to NS siRNA The corresponding volcano plots show the degree of that effect in terms of log 2-fold change for
siSGTA compared to NS siRNA against p-value for that difference Within those categories genes are annotated by those known to be sig-
nificantly DHT regulated in C4-2B cells (red circles) or not (green circles)
Can
cerCellBiology
Trotta et al 2815
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
DHT with 950 affected in both cases (Fig 1b) The effect ofSGTA knockdown in both treatment groups was validated byqPCR in an independently generated RNA sample set (p lt005 Supporting Information Figs S2AndashS2C)
We next cross-referenced these data with DHT-regulatedgenes previously identified from microarray analysis in C4-2B cells32 Of DHT-responsive targets that satisfied aBenjaminndashHochberg-adjusted statistic of p lt 005 in that ear-lier experiment 2262 fulfilled the minimum threshold cutoffcriteria of expression on the Affymetrix arrays performedhere (12 of assessed transcripts Fig 1b and SupportingInformation Fig 1B) Of the 3489 transcripts identifiedacross the siSGTA vehicle- and DHT-treated experiments23 (2751206) are predicted as DHT targets a number thatrises to 28 (275950) of those common to vehicle controland DHT SGTA (Fig 1b) The numbers are similar whengenes rather than transcripts are considered (SupportingInformation Fig S1C) and importantly represents an approx-imate twofold enrichment for DHT-responsive genesamongst those affected by SGTA knockdown (v2 5 209two-tailed p-value lt 00001)
In assessing both the qualitative and quantitative effects ofsiSGTA it was determined that for 63 of affected tran-scripts in vehicle-treated cells 58 from DHT-treated cellsand 68 of those 950 transcripts common to both the effectof SGTA knockdown was to decrease expression (pie chartsin Figs 1cndash1e) Volcano plots of the data show that the effectof SGTA knockdown occurs on a continuum and impor-tantly that there is a higher proportion of androgen-regulatedgenes (red circles) amongst those transcripts repressed bysiRNA against SGTA (299) than those increased by theknockdown (178) These data imply that SGTA may play akey role in maintaining andor limiting the androgen-regulated program in C4-2B cells however these effects arelimited to a subset of androgen-target loci
Loss of SGTA results in subtle changes to androgen
signaling in PCa cells
We elucidated the global effect of SGTA on the DHT-regulated transcriptome in C4-2B cells Of 2385 known DHTtargets 66 (15752385) were unaffected by depletion ofSGTA (Fig 2a green circles) Of the remaining 810 expres-sion of approximately one-third was altered at basal (vehicle-treated cells red circles) DHT induced (DHT-treated cellsyellow circles) or in both states (blue circles p lt 005 Fig2a) qPCR analysis of three androgen-regulated genes(S100A10 LRRC31 and DNASE2B) from an independentlygenerated RNA sample set validated these findings (p lt 005Fig 2b)
A more detailed analysis of the array data above showedthat ablation of SGTA had very little effect on DHT regula-tion of classic AR-regulated genes such as KLK3 and FKBP5This finding suggests that SGTA may exert only a relativelysmall effect on AR signaling andor is only evident on a sub-set of AR-regulated genes as had been suggested by our
previous analysis with reporter genes18 To investigate thisfurther SGTA was knocked down in C4-2B cells and RNAwas extracted following treatment with increasing doses ofDHT With an approximate 70 depletion in SGTA acrossall samples (Fig 3a) the regulation of KLK3 or FKBP5expression by qPCR was not significantly affected at any one
Figure 2 Dichotomized effect of SGTA knockdown on androgen-
regulated genes (a) Androgen-regulated genes identified from C4-
2B cells were segregated based on the effect of SGTA knockdown
in either vehicle- or DHT-treated cells Insert Green dots (unaf-
fected) represent androgen-regulated genes unaffected by SGTA
red dots (basal only) are androgen-regulated genes significantly
affected in vehicle-treated cells only yellow dots (induced only)
represent androgen-regulated genes significantly affected by SGTA
knockdown in DHT-treated cells only and blue dots (both) repre-
sent those androgen-regulated genes significantly affected by SGTA
knockdown in both vehicle- and DHT-treated cells Data represent
pairwise comparison of log 2-fold change (siSGTAsiNS) in vehicle-
and DHT-treated cells (b) The effect of SGTA knockdown was
assessed by qPCR on androgen-regulated genes identified in (a)
Gene expression was normalized in GAPDH Data represent the
mean (6 SEM) of four biological replicates p lt 005 on Mann-
Whitney U test for difference between medians of SGTA siRNA in
comparison to NS siRNA
Can
cerCellBiology
2816 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
dose of DHT (Figs 3b and 3c) However a consistent smalleffect of SGTA knockdown on regulation across all DHTconcentrations was observed (p lt 005 Figs 3b and 3c)although no overt effect was observed at FKBP51 steady-stateprotein levels (Fig 3d) Collectively these data confirm thatSGTA exerts an influence on only a subset of androgen-regulated genes and suggests that its effects on AR proteinfolding and trafficking in the presence of fluctuating andro-gen levels may hold greater consequence for PCa cellfunction
Knockdown of SGTA modulates the expression of p53 and
phosphoinositol kinaseAkt pathways
Although the above results suggest that SGTA is a key regu-lator of a subset of AR target genes we sought to elucidateits broader biological significance in PCa cells Ingenuitypathway analysis of the microarray data identified a numberof important cellular pathways significantly altered by SGTAknockdown in vehicle- or DHT-treated cells or both (Sup-porting Information Fig S3) Similar pathways were enrichedin the gene sets obtained from both vehicle- and DHT-
treated cells implying a relatively consistent effect of SGTAthat is largely independent of exogenous hormone Amongthose pathways the most highly enriched were p53 PCa andphosphoinositol kinase (PI3K)Akt signaling pathways (p lt
005 Fig 4a) Enrichment of these pathways was confirmedfor both DHT- and vehicle-treated cells using GSEA29 (Figs4bndash4d) and DAVID30 (Supporting Information Tables S2 andS3) GSEA further implicated upregulation of PCa and PI3Kpathways by SGTA knockdown but downregulation of p53signaling in both vehicle- and DHT-treated gene sets (Figs4bndash4d and Supporting Information Fig S4) Additionally thetop four of five protein families enriched in SGTA-depletedsamples related to protein kinase or serinethreonine kinaseactivity in DAVID (Supporting Information Tables S4 andS5) suggesting that SGTA may play a role in maintainingandor modulating these activities in PCa cells IndeedSGTA has been shown to maintain receptor tyrosine kinasesignaling and Akt activity in a nonsmall cell lung cancer(NSCLC) cell line1725
The effect of SGTA knockdown on specific genes withineach of the three GSEA pathways listed above was validated
Figure 3 SGTA knockdown does not affect AR-dependent KLK3 and FKBP5 gene expression (andashc) The effect of SGTA knockdown on AR sig-
naling in C4-2B cells was assessed by qPCR Cells were transfected with 5 nM NS or SGTA siRNA for 72 hr followed by treatment with 0ndash10
nM DHT for 16 hr prior to RNA extraction qPCR was performed on SGTA KLK3 and FKBP5 in triplicate samples Error bars represent (6
SEM) of three biological replicates Statistical significance represents Friedman repeated measures test for differences in means between
SGTA siRNA and NS siRNA p lt 005 (d) Matching protein lysates to the RNA samples used in (a)ndash(c) were generated for immunoblot anal-
ysis on SGTA FKBP51 and AR steady-state protein levels Hsp90 was used as a loading control
Can
cerCellBiology
Trotta et al 2817
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
Figure 4 SGTA knockdown affects p53 PI3 kinase and prostate cancer signaling pathways (a) Ingenuity pathway analysis of canonical
pathways affected by SGTA knockdown in vehicle- and DHT-treated cells or both (bndashd) Association of SGTA knockdown vehicle-treated
datasets in prostate cancer phosphatidylinositol signaling system and p53 signaling as a pathway as assessed by GSEA Probe sets from
the SGTA knockdown arrays were collapsed to the gene level and assigned a score based on a signal-to-noise ratio algorithm and ranked-
ordered by this score29 The running enrichment score (green line) is then plotted (top panel) The enrichment of the KEGG Prostate Cancer
gene set in the ranked-ordered SGTA knockdown dataset is represented by black lines (middle panel) positive to negative value of ranking
metric and the correlation of genes within the KEGG pathway to SGTA knockdown dataset (bottom panel) The normalized enrichment score
(NES) for enriched pathways is presented in each panel qPCR validation of genes is associated with both KEGG pathways Data represent
mean (6 SEM) of four biological replicates p lt 005 and p lt 001 on Mann-Whitney U test for difference between medians of SGTA
siRNA in comparison to NS siRNA
Can
cerCellBiology
2818 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
using qPCR with some variation depending on treatmentwith vehicle or DHT Specifically analysis of PCa signalingpathway genes AR GRB2 and ERBB2 demonstrated an effectonly in the presence of DHT (p lt 005 Fig 4b) For thePI3K signaling pathway PIK3R1 and PIK3CB expressiondecreased slightly but was significantly affected with DHTtreatment but only in the knockdown samples PIK3CDexpression in contrast was affected by both vehicle and DHTtreatments (p lt 001 Fig 4c) Finally we observed upregula-tion of TP53 with SGTA knockdown but a significantdecrease in expression of the two p53 target genes MDM2and CDKN1A (also known as p21 p lt 005 Fig 4d) Theabove data implicate SGTA in a number of pathways previ-ously shown to be essential to PCa cell survival proliferationand metastasis The loss of SGTA in PCa might thus be pre-dicted to have a significant effect on disease progression
SGTA acts as a positive regulator of Akt activity and C4-2B
cell viability
We next investigated the effect of SGTA knockdown on p53and PI3 kinase pathways in C4-2B cells by assessing pathwaycomponent proteins and their downstream targets Analysisof p53 signaling by immunoblotting did not reveal anychanges in steady-state protein level of the p53 target p21whereas SGTA knockdown appears to reduce Mdm2 levelseither with or without DHT (Fig 5a) However neither totalp53 nor phosphorylated p53 (S15) levels were altered bySGTA knockdown (Fig 5a) This disconnection with themRNA expression data may indicate a specific temporalndashspa-tial protein response not captured in these studies In addi-tion SGTA knockdown did not affect PI3 kinase subunit p85(PI3KR1) levels but decreased basal p110b (PI3KCB) proteinlevels (Fig 5b) Consistent with a recent report that p110b
maintains Akt activity in PCa cells as measured by phospho-rylation of the T308 and S473 residues of Akt33 the decreasein p110b with SGTA knockdown translated to reduced levelsof phosphorylated Akt T308 and Akt S473 respectively (Fig5b) These effects were partially reversed by treatment with 1nM DHT but could not be attributed to a decrease in totalAkt The effect of SGTA knockdown on p110b protein levelsand Akt phosphorylation status was verified in LNCaP PCacells (Supporting Information Fig S5A)
Given the role of SGTA in key signaling pathways wenext investigated the role of SGTA in PCa cell growthAlthough SGTA knockdown did not affect viability of C4-2Bcells when assayed under stripped serum conditions (Fig 5c)a 524 decrease in siSGTA cell number compared to mockcontrol at Day 6 was observed when assays were performedwith normal FBS containing media (p lt 005 Fig 5d) Theeffect of SGTA knockdown on C4-2B cell proliferation in thepresence of normal FBS was validated by MTT assays (Sup-porting Information Fig S5B) Immunoblot analysis demon-strated SGTA depletion for the duration of both experiments(Figs 5e and 5f) whereas trypan-blue exclusion showed thatcell death was relatively consistent across all treatment groups
(Supporting Information Figs S5C and S5D) The role ofSGTA appears to be dependent on growth of cells in fullserum which might be consistent with activation of growthfactor signaling pathways dependent on intracellular PI3KAkt signaling
SGTA and AR protein levels in patient-matched BPH and
cancer samples
The convergence of SGTA on AR and PIK3Akt signalingpathways which have important roles in PCa etiology sug-gests that altered SGTA levels may critically influence PCaprogression Our previous study indirectly suggested this bydemonstration of lower SGTA protein levels in metastaticsamples compared to primary disease17 Immunohistochemis-try was used in our study on a clinical cohort of 66 primaryadvanced PCa samples and patient-matched BPH tissues tomore precisely define any relationship between SGTA andAR levels and clinically significant outcome parameters
AR protein was found to be predominately localized inthe nucleus of luminal epithelial cells of both BPH and PCasamples with some variable stromal staining (Fig 6a) Incontrast SGTA was mainly confined to epithelial cytoplasmalthough a small number of samples showed stromal staining(Fig 6a) Quantitative analysis identified a slightly highermean AR level in cancer samples compared to BPH andlower SGTA but these did not reach significance (p 5 0055Supporting Information Table S6) Importantly however themean and median ARSGTA ratio was significantly higher inPCa samples than in matched BPH (p 5 0017) This ratiowas also higher in cancer but not BPH of those that haddied from PCa but was not significant (Supporting Informa-tion Table S6) These data imply a cancer-specific dysregula-tion in the expression of AR and SGTA that may potentiatedisease progression
DiscussionOne of the key hallmarks of cancer is the upregulation andmaintenance of key signal transduction pathways that drivecancer cell proliferation and survival5 The molecular chaper-one and cochaperone systems facilitates this in cancer cellsby providing key structural and functional frameworksthrough which these highly proliferative signals can be sus-tained and amplified The PI3KAkt signaling pathway has awell-defined role in PCa initiation and progression3435
whereby approximately 40 of localized PCa and 70 ofmetastases contain genomic alterations within PI3 kinase sig-naling proteins36 Importantly the molecular mechanismsthat contribute to constitutive PI3 kinase signaling haveunderscored recent clinical advances in the development andadministration of therapeutic agents for the treatment ofmetastatic PCa37 The Class 1A PI3 kinase subunits are agroup of enzymes composed of regulatory (p85a p85b p55a
and p55g) subunit proteins that interact with their catalytic(p110a p110b and p110d) counterparts to form heterodi-meric complexes Activation of these complexes by tyrosine
Can
cerCellBiology
Trotta et al 2819
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
kinase receptors at the cell surface mediates subsequent signaltransduction (via Akt phosphorylation) culminating in theregulation of target genes involved in cell survival prolifera-tion growth and migration PI3 kinase subunit p110b isimportant for the maintenance of Akt phosphorylation in PCcells33 and of significance we demonstrated here that regula-tion of cellular p110b levels by SGTA may be important inmaintaining Akt phosphorylation A similar effect of SGTAknockdown on Akt phosphorylation has been observed inNSCLC H1703 cells25 suggesting that SGTA acts as an
important positive regulator of PI3KAkt signaling in multi-ple cancers and can thereby mediate cancer cell proliferationand survival
In addition to the critical role of PI3 kinase signaling inPCa development there is incontrovertible evidence demon-strating that continued AR signaling is required for diseaseprogression to a castrate-resistant state38 Although it isaccepted that regulation of AR transcriptional activity in PCacells can be achieved at multiple stages within the AR activa-tion cascade39 the role of cytoplasmic TPR-containing
Figure 5 Knockdown of SGTA affects PI3K activity and C4-2B cell viability (a and b) C4-2B cells were transfected with siRNA and treated as
described in Figure 1a Immunoblot analysis of SGTA p21 Mdm2 p53 p85 p110b and Akt protein steady-state levels Analysis of active
p53 and Akt signaling was determined by phosphorylation status of p53 serine 15 (S15) Akt threonine 308 (T308) and Akt S473 residues
Hsp90 was used as a loading control (c and d) C4-2B cells were either mock transfected or transfected with nonspecific (NS) siRNA or
SGTA-specific siRNA in serum-free medium for 4 hr and then supplemented with 5 DCS-FBS or FBS Cells were cultured for 6 days and
counted every 2 days using a hemocytometer and cell viability was assessed by Trypan blue exclusion Data are presented as the mean
(6 SEM) of triplicate wells from two independent experiments Two-way ANOVA was used to determine the difference in cell growth
between groups p lt 005 differences between mockNS transfected cells versus SGTA siRNA (e and f) Cell lysates from growth assay
were assessed for SGTA steady-state protein levels b-Actin was used as a loading control [Color figure can be viewed in the online issue
which is available at wileyonlinelibrarycom]
Can
cerCellBiology
2820 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC

charcoal (DCC)-stripped FBS (stripped serum) Immunoblotswere performed as previously described1727 The followingprimary antibody dilutions were used SGTA (11000Abnova Neihu District Taiwan) AR (11000 AR N-20Santa Cruz Dallas TX) FKBP51 (11000 Santa Cruz) b-actin (12000 Sigma-Aldrich) Hsp90 (11000 Santa Cruz)p85 (11000 Cell Signaling Danvers MA) p110b (11000Cell Signaling) Akt (1300 Santa Cruz) p53 (11000 SantaCruz) p21 (11000 BD Biosciences San Jose CA) andMdm2 (11000 Santa Cruz) Phosphorylated Akt and p53antibodies were used at the following dilutions Akt threonine308 (Akt T308 12000 Cell Signaling) Akt serine 473 (AktS473 11000 Cell Signaling) and p53 serine 15 (p53 S1511000 Cell Signaling)
Small interfering RNA transfections RNA extraction and
quantitative real-time PCR
C4-2B cell lines (200000 cells per well in six-well plate) weretransfected in serum-free RPMI 1640 medium with 5 nMnonspecific (NS) control or 167 or 5 nM SGTA-specificsmall interfering RNA (siRNA Ambion Life TechnologiesMulgrove Australia) using Oligofectamine (Life Technolo-gies) for 4 hr according to the manufacturerrsquos instructionsCells were then supplemented with 5 FBS and incubatedfor 72 hr followed by treatment with vehicle (ethanol) controlor 001ndash10 nM 5a-dihydrotestosterone (DHT) for 16 hrRNA was isolated using the RNeasy RNA isolation kit (Qia-gen Melbourne Australia) and 1 lg total RNA was synthe-sized to cDNA using the iScript cDNA synthesis kit (Bio-RadLaboratories Gladesville Australia) Samples were analyzedin triplicate by quantitative real-time PCR (qPCR) as previ-ously described18 Sequences for all primers are listed in Sup-porting Information Table S1
Microarray and pathway analysis
C4-2B cells transfected with siRNA as detailed above weregrown in stripped serum containing vehicle control or 1 nMDHT for 16 hr Three hundred nanograms of total RNAextracted from each of five replicates (vehicle treated) orpooled from five replicates (DHT treated) were analyzedusing Affymetrix Human Gene 10 ST Arrays Bioinformaticswas performed in R using Bioconductor and the Limmapackage28 Briefly array data were normalized using robustmultiarray average (RMA) and filtered by mapped probesand an arbitrary minimum expression threshold wasobtained and genes different between groups called byBenjaminindashHochberg-adjusted p-values were determinedfrom Bayesian linear regression modeling Gene pathwayanalysis was conducted using the Ingenuity Pathway Analysis(IPA) program (Ingenuity Systems Redwood City CA)Gene Set Enrichment Analysis (GSEA Broad Institute httpwwwbroadinstituteorggsea)29 and Database for AnnotationVisualization and Integrated Discovery (DAVID httpwwwdavidabccncifcrfgov)30 For GSEA a false discovery rate of
25 was used and the number of random tests or permuta-tions was set at 1000 (microarray data GEO accession num-ber GSE43521)
Cell viability assays
C4-2B cells (24000 cells per well in 24-well plates) weretransfected with 5 nM NS control siRNA or three independ-ent SGTA siRNAs at a concentration of 167 nM Cells weretransfected in medium containing either 5 DCC-FBS or 5FBS then counted via a hemocytometer and assayed withtrypan blue exclusion at the end of each time point
MTT assays
For MTT assays cells were transfected as described above inmedium containing 5 FBS At the end of each time point500 ll of serum-free PRF-RPMI containing 025 mgml MTTreagent (Sigma-Aldrich) was added to the wells and incu-bated for 2 hr The MTT reagent was aspirated and replacedwith 100 ll DMSO Cell viability was then determined bymeasuring absorbance at 570 nm using a microplate reader(Bio-Rad Laboratories) Two-way ANOVA was used to com-pare differences in means between the no siRNA (mock) NSand SGTA siRNA-transfected groups Significance was set atp lt 005
Clinical cohort and immunohistochemistry
The study sample was identified from all men undergoing atransurethral resection of the prostate (TURP) for benignprostatic hyperplasia (BPH) between 2000 and 2007 at theRepatriation General Hospital (RGH South Australia) Thosefor whom a first diagnosis of PCa was made based on pathol-ogy at TURP were eligible (n 5 105) Those patients with aGleason grading and gt5 cancer at histology (n 5 66) wereselected Areas of BPH and cancer were identified in each byhematoxylin and eosin staining and mapped onto archivedparaffin-embedded material by one pathologist All protocolswere approved by the Flinders Medical Centre AdelaideSouth Australia and the RGH ethics committee (Protocol04210) A series of six tissue microarrays were constructedin duplicate and 15-mm diameter by 3-mm deep cores wereextracted from each pathologist-defined areas of BPH andcancer and embedded across six recipient paraffin blockswith 46 samples per block Two additional PCa samples andone liver sample were included as controls for each recipientblock Immunohistochemistry was performed on the cohortwith AR (AR-N20 1100 Santa Cruz) or SGTA (11000Protein Tech Chicago IL) antisera detected with LSAB1
System HRP kit (Dako Laboratories Noble Park Australia)according to the manufacturerrsquos guidelines The immunoreac-tivity for AR and SGTA was scored in a minimum of threefields in both BPH and cancer regions independently by tworesearchers (APT and DAL) Epithelia was scored 0ndash3(none low moderate and high) whereas stroma was scored0ndash2 (none low and high) Data were collated into samplescores for epithelia (0ndash9 scale) and stroma (0ndash6) scale in
Can
cerCellBiology
2814 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
both cancer and BPH regions with the difference in scoringrange due to the less dynamic staining in the stroma in com-parison to epithelia The relationship between AR and SGTAimmunoreactivity and clinical parameters including Gleasonscore serum PSA levels percent cancer prostate size clinicalstage and cancer-related death was investigated using theWilcox Rank Sum test and Barnardrsquos Exact test The WilcoxRank Sum test was also used to determine the differencebetween medians for AR and SGTA immunoreactivity inBPH specimens compared to patient-matched cancer sam-ples Significance was set at p lt 005
ResultsKnockdown of SGTA regulates the transcriptome of C4-2B
cells
To dissect the biological actions of SGTA in C4-2B PCa cellswe examined genetic changes following siRNA knockdown of
SGTA We used a low-concentration pool of three commer-cial siRNAs targeting SGTA exons 5 8 and 9 to reduce non-specific effects associated with individual siRNAs (Fig 1a)31
Transfection of the siRNA combination (167 nM each) wasas effective as each siRNA alone (si1 si2 and si3 5nM each)in reducing SGTA steady-state protein levels at 72 hr in PCaC4-2B cells (Fig 1a) For expression profiling we extractedRNA from five independent biological replicates of C4-2Bcells transfected with the SGTA siRNA pool or a NS siRNAcontrol and treated with 1 nM DHT or vehicle control for 16hr qPCR analysis revealed an average of 825 SGTAmRNA knockdown with a variance of less than 2 betweensamples (Supporting Information Fig S1A) Global expres-sion profiling using Affymetrix microarrays demonstratedthat knockdown of SGTA significantly (p lt 005) altered theexpression of 2283 transcripts in C4-2B cells in the vehicle-treated experiment and 2156 transcripts in cells treated with
Figure 1 Knockdown of SGTA modulates global androgen signaling in conjunction with the overall expression of the C4-2B transcriptome
(a) Schematic of SGTA gene located on chromosome 19 Blue boxes represent exons separated by noncoding regions SGTA siRNA1 2 and
3 (si1 si2 and si3) and their respective target exons 5 8 and 9 are indicated Immunoblot analysis for SGTA protein levels was performed
in lysates collected from C4-2B cells that had been transfected for 72 hr with a nonspecific (NS) control siRNA one of the three SGTA siR-
NAs alone (si1 si2 and si3 5nM each) or a combination of the three SGTA siRNAs (siC each at a final concentration of 167 nM) (b) Affy-
metrix expression microarray profiling of C4-2B cells after treatment for 72 hr with the siSGTA siRNA pool compared to the NS control
(BenjaminindashHochberg-adjusted p lt 005) The Euler diagram shows the number of transcripts significantly affected in experiments with
vehicle control (VC n 5 5) versus DHT (n 5 2) The number and percentage of DHT-regulated transcripts in each set is shown in brackets
(cndashe) For each of the three Euler categories shown in (b) pie charts show the proportion of genes with increased or decreased expression
following siSGTA compared to NS siRNA The corresponding volcano plots show the degree of that effect in terms of log 2-fold change for
siSGTA compared to NS siRNA against p-value for that difference Within those categories genes are annotated by those known to be sig-
nificantly DHT regulated in C4-2B cells (red circles) or not (green circles)
Can
cerCellBiology
Trotta et al 2815
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
DHT with 950 affected in both cases (Fig 1b) The effect ofSGTA knockdown in both treatment groups was validated byqPCR in an independently generated RNA sample set (p lt005 Supporting Information Figs S2AndashS2C)
We next cross-referenced these data with DHT-regulatedgenes previously identified from microarray analysis in C4-2B cells32 Of DHT-responsive targets that satisfied aBenjaminndashHochberg-adjusted statistic of p lt 005 in that ear-lier experiment 2262 fulfilled the minimum threshold cutoffcriteria of expression on the Affymetrix arrays performedhere (12 of assessed transcripts Fig 1b and SupportingInformation Fig 1B) Of the 3489 transcripts identifiedacross the siSGTA vehicle- and DHT-treated experiments23 (2751206) are predicted as DHT targets a number thatrises to 28 (275950) of those common to vehicle controland DHT SGTA (Fig 1b) The numbers are similar whengenes rather than transcripts are considered (SupportingInformation Fig S1C) and importantly represents an approx-imate twofold enrichment for DHT-responsive genesamongst those affected by SGTA knockdown (v2 5 209two-tailed p-value lt 00001)
In assessing both the qualitative and quantitative effects ofsiSGTA it was determined that for 63 of affected tran-scripts in vehicle-treated cells 58 from DHT-treated cellsand 68 of those 950 transcripts common to both the effectof SGTA knockdown was to decrease expression (pie chartsin Figs 1cndash1e) Volcano plots of the data show that the effectof SGTA knockdown occurs on a continuum and impor-tantly that there is a higher proportion of androgen-regulatedgenes (red circles) amongst those transcripts repressed bysiRNA against SGTA (299) than those increased by theknockdown (178) These data imply that SGTA may play akey role in maintaining andor limiting the androgen-regulated program in C4-2B cells however these effects arelimited to a subset of androgen-target loci
Loss of SGTA results in subtle changes to androgen
signaling in PCa cells
We elucidated the global effect of SGTA on the DHT-regulated transcriptome in C4-2B cells Of 2385 known DHTtargets 66 (15752385) were unaffected by depletion ofSGTA (Fig 2a green circles) Of the remaining 810 expres-sion of approximately one-third was altered at basal (vehicle-treated cells red circles) DHT induced (DHT-treated cellsyellow circles) or in both states (blue circles p lt 005 Fig2a) qPCR analysis of three androgen-regulated genes(S100A10 LRRC31 and DNASE2B) from an independentlygenerated RNA sample set validated these findings (p lt 005Fig 2b)
A more detailed analysis of the array data above showedthat ablation of SGTA had very little effect on DHT regula-tion of classic AR-regulated genes such as KLK3 and FKBP5This finding suggests that SGTA may exert only a relativelysmall effect on AR signaling andor is only evident on a sub-set of AR-regulated genes as had been suggested by our
previous analysis with reporter genes18 To investigate thisfurther SGTA was knocked down in C4-2B cells and RNAwas extracted following treatment with increasing doses ofDHT With an approximate 70 depletion in SGTA acrossall samples (Fig 3a) the regulation of KLK3 or FKBP5expression by qPCR was not significantly affected at any one
Figure 2 Dichotomized effect of SGTA knockdown on androgen-
regulated genes (a) Androgen-regulated genes identified from C4-
2B cells were segregated based on the effect of SGTA knockdown
in either vehicle- or DHT-treated cells Insert Green dots (unaf-
fected) represent androgen-regulated genes unaffected by SGTA
red dots (basal only) are androgen-regulated genes significantly
affected in vehicle-treated cells only yellow dots (induced only)
represent androgen-regulated genes significantly affected by SGTA
knockdown in DHT-treated cells only and blue dots (both) repre-
sent those androgen-regulated genes significantly affected by SGTA
knockdown in both vehicle- and DHT-treated cells Data represent
pairwise comparison of log 2-fold change (siSGTAsiNS) in vehicle-
and DHT-treated cells (b) The effect of SGTA knockdown was
assessed by qPCR on androgen-regulated genes identified in (a)
Gene expression was normalized in GAPDH Data represent the
mean (6 SEM) of four biological replicates p lt 005 on Mann-
Whitney U test for difference between medians of SGTA siRNA in
comparison to NS siRNA
Can
cerCellBiology
2816 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
dose of DHT (Figs 3b and 3c) However a consistent smalleffect of SGTA knockdown on regulation across all DHTconcentrations was observed (p lt 005 Figs 3b and 3c)although no overt effect was observed at FKBP51 steady-stateprotein levels (Fig 3d) Collectively these data confirm thatSGTA exerts an influence on only a subset of androgen-regulated genes and suggests that its effects on AR proteinfolding and trafficking in the presence of fluctuating andro-gen levels may hold greater consequence for PCa cellfunction
Knockdown of SGTA modulates the expression of p53 and
phosphoinositol kinaseAkt pathways
Although the above results suggest that SGTA is a key regu-lator of a subset of AR target genes we sought to elucidateits broader biological significance in PCa cells Ingenuitypathway analysis of the microarray data identified a numberof important cellular pathways significantly altered by SGTAknockdown in vehicle- or DHT-treated cells or both (Sup-porting Information Fig S3) Similar pathways were enrichedin the gene sets obtained from both vehicle- and DHT-
treated cells implying a relatively consistent effect of SGTAthat is largely independent of exogenous hormone Amongthose pathways the most highly enriched were p53 PCa andphosphoinositol kinase (PI3K)Akt signaling pathways (p lt
005 Fig 4a) Enrichment of these pathways was confirmedfor both DHT- and vehicle-treated cells using GSEA29 (Figs4bndash4d) and DAVID30 (Supporting Information Tables S2 andS3) GSEA further implicated upregulation of PCa and PI3Kpathways by SGTA knockdown but downregulation of p53signaling in both vehicle- and DHT-treated gene sets (Figs4bndash4d and Supporting Information Fig S4) Additionally thetop four of five protein families enriched in SGTA-depletedsamples related to protein kinase or serinethreonine kinaseactivity in DAVID (Supporting Information Tables S4 andS5) suggesting that SGTA may play a role in maintainingandor modulating these activities in PCa cells IndeedSGTA has been shown to maintain receptor tyrosine kinasesignaling and Akt activity in a nonsmall cell lung cancer(NSCLC) cell line1725
The effect of SGTA knockdown on specific genes withineach of the three GSEA pathways listed above was validated
Figure 3 SGTA knockdown does not affect AR-dependent KLK3 and FKBP5 gene expression (andashc) The effect of SGTA knockdown on AR sig-
naling in C4-2B cells was assessed by qPCR Cells were transfected with 5 nM NS or SGTA siRNA for 72 hr followed by treatment with 0ndash10
nM DHT for 16 hr prior to RNA extraction qPCR was performed on SGTA KLK3 and FKBP5 in triplicate samples Error bars represent (6
SEM) of three biological replicates Statistical significance represents Friedman repeated measures test for differences in means between
SGTA siRNA and NS siRNA p lt 005 (d) Matching protein lysates to the RNA samples used in (a)ndash(c) were generated for immunoblot anal-
ysis on SGTA FKBP51 and AR steady-state protein levels Hsp90 was used as a loading control
Can
cerCellBiology
Trotta et al 2817
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
Figure 4 SGTA knockdown affects p53 PI3 kinase and prostate cancer signaling pathways (a) Ingenuity pathway analysis of canonical
pathways affected by SGTA knockdown in vehicle- and DHT-treated cells or both (bndashd) Association of SGTA knockdown vehicle-treated
datasets in prostate cancer phosphatidylinositol signaling system and p53 signaling as a pathway as assessed by GSEA Probe sets from
the SGTA knockdown arrays were collapsed to the gene level and assigned a score based on a signal-to-noise ratio algorithm and ranked-
ordered by this score29 The running enrichment score (green line) is then plotted (top panel) The enrichment of the KEGG Prostate Cancer
gene set in the ranked-ordered SGTA knockdown dataset is represented by black lines (middle panel) positive to negative value of ranking
metric and the correlation of genes within the KEGG pathway to SGTA knockdown dataset (bottom panel) The normalized enrichment score
(NES) for enriched pathways is presented in each panel qPCR validation of genes is associated with both KEGG pathways Data represent
mean (6 SEM) of four biological replicates p lt 005 and p lt 001 on Mann-Whitney U test for difference between medians of SGTA
siRNA in comparison to NS siRNA
Can
cerCellBiology
2818 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
using qPCR with some variation depending on treatmentwith vehicle or DHT Specifically analysis of PCa signalingpathway genes AR GRB2 and ERBB2 demonstrated an effectonly in the presence of DHT (p lt 005 Fig 4b) For thePI3K signaling pathway PIK3R1 and PIK3CB expressiondecreased slightly but was significantly affected with DHTtreatment but only in the knockdown samples PIK3CDexpression in contrast was affected by both vehicle and DHTtreatments (p lt 001 Fig 4c) Finally we observed upregula-tion of TP53 with SGTA knockdown but a significantdecrease in expression of the two p53 target genes MDM2and CDKN1A (also known as p21 p lt 005 Fig 4d) Theabove data implicate SGTA in a number of pathways previ-ously shown to be essential to PCa cell survival proliferationand metastasis The loss of SGTA in PCa might thus be pre-dicted to have a significant effect on disease progression
SGTA acts as a positive regulator of Akt activity and C4-2B
cell viability
We next investigated the effect of SGTA knockdown on p53and PI3 kinase pathways in C4-2B cells by assessing pathwaycomponent proteins and their downstream targets Analysisof p53 signaling by immunoblotting did not reveal anychanges in steady-state protein level of the p53 target p21whereas SGTA knockdown appears to reduce Mdm2 levelseither with or without DHT (Fig 5a) However neither totalp53 nor phosphorylated p53 (S15) levels were altered bySGTA knockdown (Fig 5a) This disconnection with themRNA expression data may indicate a specific temporalndashspa-tial protein response not captured in these studies In addi-tion SGTA knockdown did not affect PI3 kinase subunit p85(PI3KR1) levels but decreased basal p110b (PI3KCB) proteinlevels (Fig 5b) Consistent with a recent report that p110b
maintains Akt activity in PCa cells as measured by phospho-rylation of the T308 and S473 residues of Akt33 the decreasein p110b with SGTA knockdown translated to reduced levelsof phosphorylated Akt T308 and Akt S473 respectively (Fig5b) These effects were partially reversed by treatment with 1nM DHT but could not be attributed to a decrease in totalAkt The effect of SGTA knockdown on p110b protein levelsand Akt phosphorylation status was verified in LNCaP PCacells (Supporting Information Fig S5A)
Given the role of SGTA in key signaling pathways wenext investigated the role of SGTA in PCa cell growthAlthough SGTA knockdown did not affect viability of C4-2Bcells when assayed under stripped serum conditions (Fig 5c)a 524 decrease in siSGTA cell number compared to mockcontrol at Day 6 was observed when assays were performedwith normal FBS containing media (p lt 005 Fig 5d) Theeffect of SGTA knockdown on C4-2B cell proliferation in thepresence of normal FBS was validated by MTT assays (Sup-porting Information Fig S5B) Immunoblot analysis demon-strated SGTA depletion for the duration of both experiments(Figs 5e and 5f) whereas trypan-blue exclusion showed thatcell death was relatively consistent across all treatment groups
(Supporting Information Figs S5C and S5D) The role ofSGTA appears to be dependent on growth of cells in fullserum which might be consistent with activation of growthfactor signaling pathways dependent on intracellular PI3KAkt signaling
SGTA and AR protein levels in patient-matched BPH and
cancer samples
The convergence of SGTA on AR and PIK3Akt signalingpathways which have important roles in PCa etiology sug-gests that altered SGTA levels may critically influence PCaprogression Our previous study indirectly suggested this bydemonstration of lower SGTA protein levels in metastaticsamples compared to primary disease17 Immunohistochemis-try was used in our study on a clinical cohort of 66 primaryadvanced PCa samples and patient-matched BPH tissues tomore precisely define any relationship between SGTA andAR levels and clinically significant outcome parameters
AR protein was found to be predominately localized inthe nucleus of luminal epithelial cells of both BPH and PCasamples with some variable stromal staining (Fig 6a) Incontrast SGTA was mainly confined to epithelial cytoplasmalthough a small number of samples showed stromal staining(Fig 6a) Quantitative analysis identified a slightly highermean AR level in cancer samples compared to BPH andlower SGTA but these did not reach significance (p 5 0055Supporting Information Table S6) Importantly however themean and median ARSGTA ratio was significantly higher inPCa samples than in matched BPH (p 5 0017) This ratiowas also higher in cancer but not BPH of those that haddied from PCa but was not significant (Supporting Informa-tion Table S6) These data imply a cancer-specific dysregula-tion in the expression of AR and SGTA that may potentiatedisease progression
DiscussionOne of the key hallmarks of cancer is the upregulation andmaintenance of key signal transduction pathways that drivecancer cell proliferation and survival5 The molecular chaper-one and cochaperone systems facilitates this in cancer cellsby providing key structural and functional frameworksthrough which these highly proliferative signals can be sus-tained and amplified The PI3KAkt signaling pathway has awell-defined role in PCa initiation and progression3435
whereby approximately 40 of localized PCa and 70 ofmetastases contain genomic alterations within PI3 kinase sig-naling proteins36 Importantly the molecular mechanismsthat contribute to constitutive PI3 kinase signaling haveunderscored recent clinical advances in the development andadministration of therapeutic agents for the treatment ofmetastatic PCa37 The Class 1A PI3 kinase subunits are agroup of enzymes composed of regulatory (p85a p85b p55a
and p55g) subunit proteins that interact with their catalytic(p110a p110b and p110d) counterparts to form heterodi-meric complexes Activation of these complexes by tyrosine
Can
cerCellBiology
Trotta et al 2819
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
kinase receptors at the cell surface mediates subsequent signaltransduction (via Akt phosphorylation) culminating in theregulation of target genes involved in cell survival prolifera-tion growth and migration PI3 kinase subunit p110b isimportant for the maintenance of Akt phosphorylation in PCcells33 and of significance we demonstrated here that regula-tion of cellular p110b levels by SGTA may be important inmaintaining Akt phosphorylation A similar effect of SGTAknockdown on Akt phosphorylation has been observed inNSCLC H1703 cells25 suggesting that SGTA acts as an
important positive regulator of PI3KAkt signaling in multi-ple cancers and can thereby mediate cancer cell proliferationand survival
In addition to the critical role of PI3 kinase signaling inPCa development there is incontrovertible evidence demon-strating that continued AR signaling is required for diseaseprogression to a castrate-resistant state38 Although it isaccepted that regulation of AR transcriptional activity in PCacells can be achieved at multiple stages within the AR activa-tion cascade39 the role of cytoplasmic TPR-containing
Figure 5 Knockdown of SGTA affects PI3K activity and C4-2B cell viability (a and b) C4-2B cells were transfected with siRNA and treated as
described in Figure 1a Immunoblot analysis of SGTA p21 Mdm2 p53 p85 p110b and Akt protein steady-state levels Analysis of active
p53 and Akt signaling was determined by phosphorylation status of p53 serine 15 (S15) Akt threonine 308 (T308) and Akt S473 residues
Hsp90 was used as a loading control (c and d) C4-2B cells were either mock transfected or transfected with nonspecific (NS) siRNA or
SGTA-specific siRNA in serum-free medium for 4 hr and then supplemented with 5 DCS-FBS or FBS Cells were cultured for 6 days and
counted every 2 days using a hemocytometer and cell viability was assessed by Trypan blue exclusion Data are presented as the mean
(6 SEM) of triplicate wells from two independent experiments Two-way ANOVA was used to determine the difference in cell growth
between groups p lt 005 differences between mockNS transfected cells versus SGTA siRNA (e and f) Cell lysates from growth assay
were assessed for SGTA steady-state protein levels b-Actin was used as a loading control [Color figure can be viewed in the online issue
which is available at wileyonlinelibrarycom]
Can
cerCellBiology
2820 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
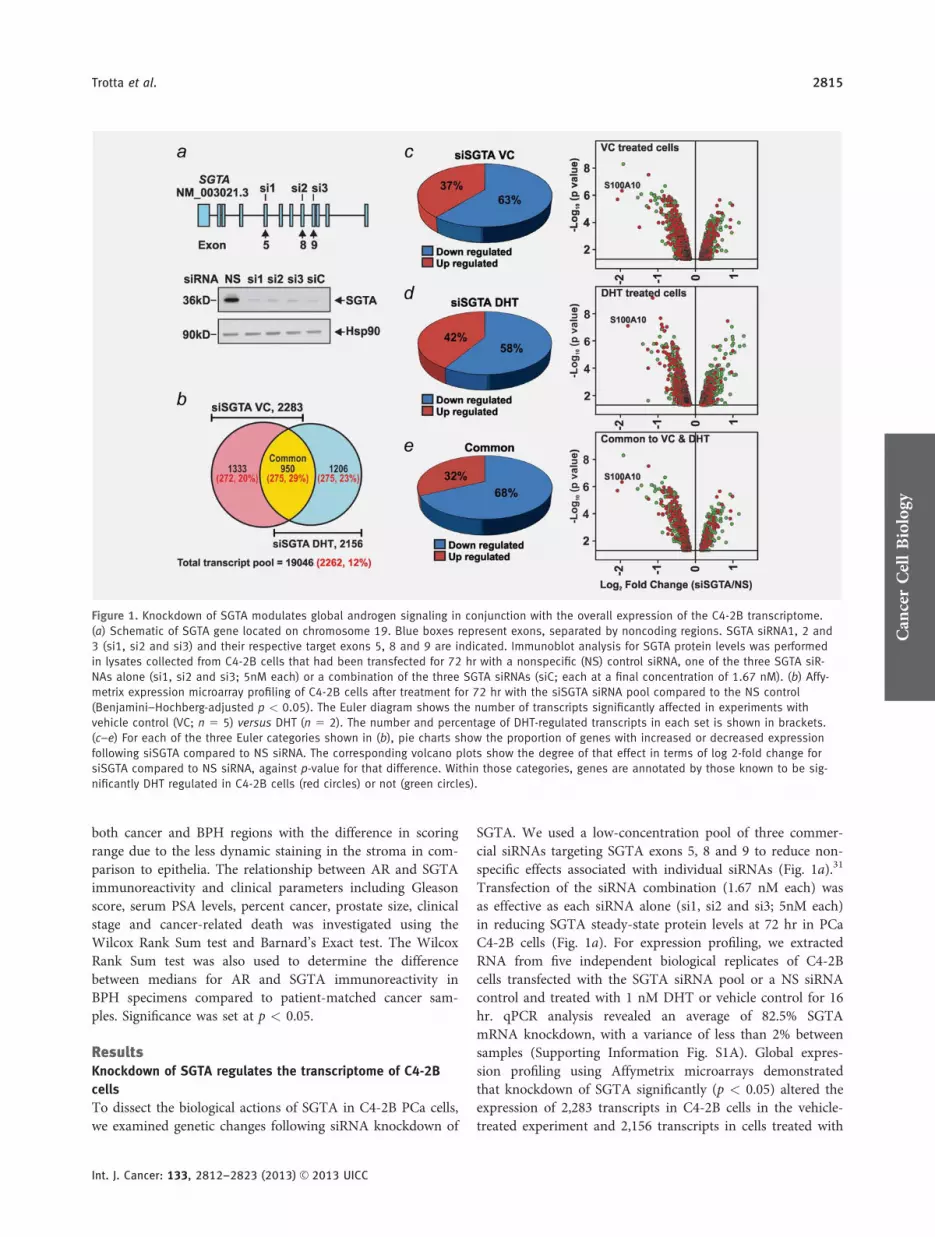
both cancer and BPH regions with the difference in scoringrange due to the less dynamic staining in the stroma in com-parison to epithelia The relationship between AR and SGTAimmunoreactivity and clinical parameters including Gleasonscore serum PSA levels percent cancer prostate size clinicalstage and cancer-related death was investigated using theWilcox Rank Sum test and Barnardrsquos Exact test The WilcoxRank Sum test was also used to determine the differencebetween medians for AR and SGTA immunoreactivity inBPH specimens compared to patient-matched cancer sam-ples Significance was set at p lt 005
ResultsKnockdown of SGTA regulates the transcriptome of C4-2B
cells
To dissect the biological actions of SGTA in C4-2B PCa cellswe examined genetic changes following siRNA knockdown of
SGTA We used a low-concentration pool of three commer-cial siRNAs targeting SGTA exons 5 8 and 9 to reduce non-specific effects associated with individual siRNAs (Fig 1a)31
Transfection of the siRNA combination (167 nM each) wasas effective as each siRNA alone (si1 si2 and si3 5nM each)in reducing SGTA steady-state protein levels at 72 hr in PCaC4-2B cells (Fig 1a) For expression profiling we extractedRNA from five independent biological replicates of C4-2Bcells transfected with the SGTA siRNA pool or a NS siRNAcontrol and treated with 1 nM DHT or vehicle control for 16hr qPCR analysis revealed an average of 825 SGTAmRNA knockdown with a variance of less than 2 betweensamples (Supporting Information Fig S1A) Global expres-sion profiling using Affymetrix microarrays demonstratedthat knockdown of SGTA significantly (p lt 005) altered theexpression of 2283 transcripts in C4-2B cells in the vehicle-treated experiment and 2156 transcripts in cells treated with
Figure 1 Knockdown of SGTA modulates global androgen signaling in conjunction with the overall expression of the C4-2B transcriptome
(a) Schematic of SGTA gene located on chromosome 19 Blue boxes represent exons separated by noncoding regions SGTA siRNA1 2 and
3 (si1 si2 and si3) and their respective target exons 5 8 and 9 are indicated Immunoblot analysis for SGTA protein levels was performed
in lysates collected from C4-2B cells that had been transfected for 72 hr with a nonspecific (NS) control siRNA one of the three SGTA siR-
NAs alone (si1 si2 and si3 5nM each) or a combination of the three SGTA siRNAs (siC each at a final concentration of 167 nM) (b) Affy-
metrix expression microarray profiling of C4-2B cells after treatment for 72 hr with the siSGTA siRNA pool compared to the NS control
(BenjaminindashHochberg-adjusted p lt 005) The Euler diagram shows the number of transcripts significantly affected in experiments with
vehicle control (VC n 5 5) versus DHT (n 5 2) The number and percentage of DHT-regulated transcripts in each set is shown in brackets
(cndashe) For each of the three Euler categories shown in (b) pie charts show the proportion of genes with increased or decreased expression
following siSGTA compared to NS siRNA The corresponding volcano plots show the degree of that effect in terms of log 2-fold change for
siSGTA compared to NS siRNA against p-value for that difference Within those categories genes are annotated by those known to be sig-
nificantly DHT regulated in C4-2B cells (red circles) or not (green circles)
Can
cerCellBiology
Trotta et al 2815
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
DHT with 950 affected in both cases (Fig 1b) The effect ofSGTA knockdown in both treatment groups was validated byqPCR in an independently generated RNA sample set (p lt005 Supporting Information Figs S2AndashS2C)
We next cross-referenced these data with DHT-regulatedgenes previously identified from microarray analysis in C4-2B cells32 Of DHT-responsive targets that satisfied aBenjaminndashHochberg-adjusted statistic of p lt 005 in that ear-lier experiment 2262 fulfilled the minimum threshold cutoffcriteria of expression on the Affymetrix arrays performedhere (12 of assessed transcripts Fig 1b and SupportingInformation Fig 1B) Of the 3489 transcripts identifiedacross the siSGTA vehicle- and DHT-treated experiments23 (2751206) are predicted as DHT targets a number thatrises to 28 (275950) of those common to vehicle controland DHT SGTA (Fig 1b) The numbers are similar whengenes rather than transcripts are considered (SupportingInformation Fig S1C) and importantly represents an approx-imate twofold enrichment for DHT-responsive genesamongst those affected by SGTA knockdown (v2 5 209two-tailed p-value lt 00001)
In assessing both the qualitative and quantitative effects ofsiSGTA it was determined that for 63 of affected tran-scripts in vehicle-treated cells 58 from DHT-treated cellsand 68 of those 950 transcripts common to both the effectof SGTA knockdown was to decrease expression (pie chartsin Figs 1cndash1e) Volcano plots of the data show that the effectof SGTA knockdown occurs on a continuum and impor-tantly that there is a higher proportion of androgen-regulatedgenes (red circles) amongst those transcripts repressed bysiRNA against SGTA (299) than those increased by theknockdown (178) These data imply that SGTA may play akey role in maintaining andor limiting the androgen-regulated program in C4-2B cells however these effects arelimited to a subset of androgen-target loci
Loss of SGTA results in subtle changes to androgen
signaling in PCa cells
We elucidated the global effect of SGTA on the DHT-regulated transcriptome in C4-2B cells Of 2385 known DHTtargets 66 (15752385) were unaffected by depletion ofSGTA (Fig 2a green circles) Of the remaining 810 expres-sion of approximately one-third was altered at basal (vehicle-treated cells red circles) DHT induced (DHT-treated cellsyellow circles) or in both states (blue circles p lt 005 Fig2a) qPCR analysis of three androgen-regulated genes(S100A10 LRRC31 and DNASE2B) from an independentlygenerated RNA sample set validated these findings (p lt 005Fig 2b)
A more detailed analysis of the array data above showedthat ablation of SGTA had very little effect on DHT regula-tion of classic AR-regulated genes such as KLK3 and FKBP5This finding suggests that SGTA may exert only a relativelysmall effect on AR signaling andor is only evident on a sub-set of AR-regulated genes as had been suggested by our
previous analysis with reporter genes18 To investigate thisfurther SGTA was knocked down in C4-2B cells and RNAwas extracted following treatment with increasing doses ofDHT With an approximate 70 depletion in SGTA acrossall samples (Fig 3a) the regulation of KLK3 or FKBP5expression by qPCR was not significantly affected at any one
Figure 2 Dichotomized effect of SGTA knockdown on androgen-
regulated genes (a) Androgen-regulated genes identified from C4-
2B cells were segregated based on the effect of SGTA knockdown
in either vehicle- or DHT-treated cells Insert Green dots (unaf-
fected) represent androgen-regulated genes unaffected by SGTA
red dots (basal only) are androgen-regulated genes significantly
affected in vehicle-treated cells only yellow dots (induced only)
represent androgen-regulated genes significantly affected by SGTA
knockdown in DHT-treated cells only and blue dots (both) repre-
sent those androgen-regulated genes significantly affected by SGTA
knockdown in both vehicle- and DHT-treated cells Data represent
pairwise comparison of log 2-fold change (siSGTAsiNS) in vehicle-
and DHT-treated cells (b) The effect of SGTA knockdown was
assessed by qPCR on androgen-regulated genes identified in (a)
Gene expression was normalized in GAPDH Data represent the
mean (6 SEM) of four biological replicates p lt 005 on Mann-
Whitney U test for difference between medians of SGTA siRNA in
comparison to NS siRNA
Can
cerCellBiology
2816 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
dose of DHT (Figs 3b and 3c) However a consistent smalleffect of SGTA knockdown on regulation across all DHTconcentrations was observed (p lt 005 Figs 3b and 3c)although no overt effect was observed at FKBP51 steady-stateprotein levels (Fig 3d) Collectively these data confirm thatSGTA exerts an influence on only a subset of androgen-regulated genes and suggests that its effects on AR proteinfolding and trafficking in the presence of fluctuating andro-gen levels may hold greater consequence for PCa cellfunction
Knockdown of SGTA modulates the expression of p53 and
phosphoinositol kinaseAkt pathways
Although the above results suggest that SGTA is a key regu-lator of a subset of AR target genes we sought to elucidateits broader biological significance in PCa cells Ingenuitypathway analysis of the microarray data identified a numberof important cellular pathways significantly altered by SGTAknockdown in vehicle- or DHT-treated cells or both (Sup-porting Information Fig S3) Similar pathways were enrichedin the gene sets obtained from both vehicle- and DHT-
treated cells implying a relatively consistent effect of SGTAthat is largely independent of exogenous hormone Amongthose pathways the most highly enriched were p53 PCa andphosphoinositol kinase (PI3K)Akt signaling pathways (p lt
005 Fig 4a) Enrichment of these pathways was confirmedfor both DHT- and vehicle-treated cells using GSEA29 (Figs4bndash4d) and DAVID30 (Supporting Information Tables S2 andS3) GSEA further implicated upregulation of PCa and PI3Kpathways by SGTA knockdown but downregulation of p53signaling in both vehicle- and DHT-treated gene sets (Figs4bndash4d and Supporting Information Fig S4) Additionally thetop four of five protein families enriched in SGTA-depletedsamples related to protein kinase or serinethreonine kinaseactivity in DAVID (Supporting Information Tables S4 andS5) suggesting that SGTA may play a role in maintainingandor modulating these activities in PCa cells IndeedSGTA has been shown to maintain receptor tyrosine kinasesignaling and Akt activity in a nonsmall cell lung cancer(NSCLC) cell line1725
The effect of SGTA knockdown on specific genes withineach of the three GSEA pathways listed above was validated
Figure 3 SGTA knockdown does not affect AR-dependent KLK3 and FKBP5 gene expression (andashc) The effect of SGTA knockdown on AR sig-
naling in C4-2B cells was assessed by qPCR Cells were transfected with 5 nM NS or SGTA siRNA for 72 hr followed by treatment with 0ndash10
nM DHT for 16 hr prior to RNA extraction qPCR was performed on SGTA KLK3 and FKBP5 in triplicate samples Error bars represent (6
SEM) of three biological replicates Statistical significance represents Friedman repeated measures test for differences in means between
SGTA siRNA and NS siRNA p lt 005 (d) Matching protein lysates to the RNA samples used in (a)ndash(c) were generated for immunoblot anal-
ysis on SGTA FKBP51 and AR steady-state protein levels Hsp90 was used as a loading control
Can
cerCellBiology
Trotta et al 2817
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
Figure 4 SGTA knockdown affects p53 PI3 kinase and prostate cancer signaling pathways (a) Ingenuity pathway analysis of canonical
pathways affected by SGTA knockdown in vehicle- and DHT-treated cells or both (bndashd) Association of SGTA knockdown vehicle-treated
datasets in prostate cancer phosphatidylinositol signaling system and p53 signaling as a pathway as assessed by GSEA Probe sets from
the SGTA knockdown arrays were collapsed to the gene level and assigned a score based on a signal-to-noise ratio algorithm and ranked-
ordered by this score29 The running enrichment score (green line) is then plotted (top panel) The enrichment of the KEGG Prostate Cancer
gene set in the ranked-ordered SGTA knockdown dataset is represented by black lines (middle panel) positive to negative value of ranking
metric and the correlation of genes within the KEGG pathway to SGTA knockdown dataset (bottom panel) The normalized enrichment score
(NES) for enriched pathways is presented in each panel qPCR validation of genes is associated with both KEGG pathways Data represent
mean (6 SEM) of four biological replicates p lt 005 and p lt 001 on Mann-Whitney U test for difference between medians of SGTA
siRNA in comparison to NS siRNA
Can
cerCellBiology
2818 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
using qPCR with some variation depending on treatmentwith vehicle or DHT Specifically analysis of PCa signalingpathway genes AR GRB2 and ERBB2 demonstrated an effectonly in the presence of DHT (p lt 005 Fig 4b) For thePI3K signaling pathway PIK3R1 and PIK3CB expressiondecreased slightly but was significantly affected with DHTtreatment but only in the knockdown samples PIK3CDexpression in contrast was affected by both vehicle and DHTtreatments (p lt 001 Fig 4c) Finally we observed upregula-tion of TP53 with SGTA knockdown but a significantdecrease in expression of the two p53 target genes MDM2and CDKN1A (also known as p21 p lt 005 Fig 4d) Theabove data implicate SGTA in a number of pathways previ-ously shown to be essential to PCa cell survival proliferationand metastasis The loss of SGTA in PCa might thus be pre-dicted to have a significant effect on disease progression
SGTA acts as a positive regulator of Akt activity and C4-2B
cell viability
We next investigated the effect of SGTA knockdown on p53and PI3 kinase pathways in C4-2B cells by assessing pathwaycomponent proteins and their downstream targets Analysisof p53 signaling by immunoblotting did not reveal anychanges in steady-state protein level of the p53 target p21whereas SGTA knockdown appears to reduce Mdm2 levelseither with or without DHT (Fig 5a) However neither totalp53 nor phosphorylated p53 (S15) levels were altered bySGTA knockdown (Fig 5a) This disconnection with themRNA expression data may indicate a specific temporalndashspa-tial protein response not captured in these studies In addi-tion SGTA knockdown did not affect PI3 kinase subunit p85(PI3KR1) levels but decreased basal p110b (PI3KCB) proteinlevels (Fig 5b) Consistent with a recent report that p110b
maintains Akt activity in PCa cells as measured by phospho-rylation of the T308 and S473 residues of Akt33 the decreasein p110b with SGTA knockdown translated to reduced levelsof phosphorylated Akt T308 and Akt S473 respectively (Fig5b) These effects were partially reversed by treatment with 1nM DHT but could not be attributed to a decrease in totalAkt The effect of SGTA knockdown on p110b protein levelsand Akt phosphorylation status was verified in LNCaP PCacells (Supporting Information Fig S5A)
Given the role of SGTA in key signaling pathways wenext investigated the role of SGTA in PCa cell growthAlthough SGTA knockdown did not affect viability of C4-2Bcells when assayed under stripped serum conditions (Fig 5c)a 524 decrease in siSGTA cell number compared to mockcontrol at Day 6 was observed when assays were performedwith normal FBS containing media (p lt 005 Fig 5d) Theeffect of SGTA knockdown on C4-2B cell proliferation in thepresence of normal FBS was validated by MTT assays (Sup-porting Information Fig S5B) Immunoblot analysis demon-strated SGTA depletion for the duration of both experiments(Figs 5e and 5f) whereas trypan-blue exclusion showed thatcell death was relatively consistent across all treatment groups
(Supporting Information Figs S5C and S5D) The role ofSGTA appears to be dependent on growth of cells in fullserum which might be consistent with activation of growthfactor signaling pathways dependent on intracellular PI3KAkt signaling
SGTA and AR protein levels in patient-matched BPH and
cancer samples
The convergence of SGTA on AR and PIK3Akt signalingpathways which have important roles in PCa etiology sug-gests that altered SGTA levels may critically influence PCaprogression Our previous study indirectly suggested this bydemonstration of lower SGTA protein levels in metastaticsamples compared to primary disease17 Immunohistochemis-try was used in our study on a clinical cohort of 66 primaryadvanced PCa samples and patient-matched BPH tissues tomore precisely define any relationship between SGTA andAR levels and clinically significant outcome parameters
AR protein was found to be predominately localized inthe nucleus of luminal epithelial cells of both BPH and PCasamples with some variable stromal staining (Fig 6a) Incontrast SGTA was mainly confined to epithelial cytoplasmalthough a small number of samples showed stromal staining(Fig 6a) Quantitative analysis identified a slightly highermean AR level in cancer samples compared to BPH andlower SGTA but these did not reach significance (p 5 0055Supporting Information Table S6) Importantly however themean and median ARSGTA ratio was significantly higher inPCa samples than in matched BPH (p 5 0017) This ratiowas also higher in cancer but not BPH of those that haddied from PCa but was not significant (Supporting Informa-tion Table S6) These data imply a cancer-specific dysregula-tion in the expression of AR and SGTA that may potentiatedisease progression
DiscussionOne of the key hallmarks of cancer is the upregulation andmaintenance of key signal transduction pathways that drivecancer cell proliferation and survival5 The molecular chaper-one and cochaperone systems facilitates this in cancer cellsby providing key structural and functional frameworksthrough which these highly proliferative signals can be sus-tained and amplified The PI3KAkt signaling pathway has awell-defined role in PCa initiation and progression3435
whereby approximately 40 of localized PCa and 70 ofmetastases contain genomic alterations within PI3 kinase sig-naling proteins36 Importantly the molecular mechanismsthat contribute to constitutive PI3 kinase signaling haveunderscored recent clinical advances in the development andadministration of therapeutic agents for the treatment ofmetastatic PCa37 The Class 1A PI3 kinase subunits are agroup of enzymes composed of regulatory (p85a p85b p55a
and p55g) subunit proteins that interact with their catalytic(p110a p110b and p110d) counterparts to form heterodi-meric complexes Activation of these complexes by tyrosine
Can
cerCellBiology
Trotta et al 2819
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
kinase receptors at the cell surface mediates subsequent signaltransduction (via Akt phosphorylation) culminating in theregulation of target genes involved in cell survival prolifera-tion growth and migration PI3 kinase subunit p110b isimportant for the maintenance of Akt phosphorylation in PCcells33 and of significance we demonstrated here that regula-tion of cellular p110b levels by SGTA may be important inmaintaining Akt phosphorylation A similar effect of SGTAknockdown on Akt phosphorylation has been observed inNSCLC H1703 cells25 suggesting that SGTA acts as an
important positive regulator of PI3KAkt signaling in multi-ple cancers and can thereby mediate cancer cell proliferationand survival
In addition to the critical role of PI3 kinase signaling inPCa development there is incontrovertible evidence demon-strating that continued AR signaling is required for diseaseprogression to a castrate-resistant state38 Although it isaccepted that regulation of AR transcriptional activity in PCacells can be achieved at multiple stages within the AR activa-tion cascade39 the role of cytoplasmic TPR-containing
Figure 5 Knockdown of SGTA affects PI3K activity and C4-2B cell viability (a and b) C4-2B cells were transfected with siRNA and treated as
described in Figure 1a Immunoblot analysis of SGTA p21 Mdm2 p53 p85 p110b and Akt protein steady-state levels Analysis of active
p53 and Akt signaling was determined by phosphorylation status of p53 serine 15 (S15) Akt threonine 308 (T308) and Akt S473 residues
Hsp90 was used as a loading control (c and d) C4-2B cells were either mock transfected or transfected with nonspecific (NS) siRNA or
SGTA-specific siRNA in serum-free medium for 4 hr and then supplemented with 5 DCS-FBS or FBS Cells were cultured for 6 days and
counted every 2 days using a hemocytometer and cell viability was assessed by Trypan blue exclusion Data are presented as the mean
(6 SEM) of triplicate wells from two independent experiments Two-way ANOVA was used to determine the difference in cell growth
between groups p lt 005 differences between mockNS transfected cells versus SGTA siRNA (e and f) Cell lysates from growth assay
were assessed for SGTA steady-state protein levels b-Actin was used as a loading control [Color figure can be viewed in the online issue
which is available at wileyonlinelibrarycom]
Can
cerCellBiology
2820 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
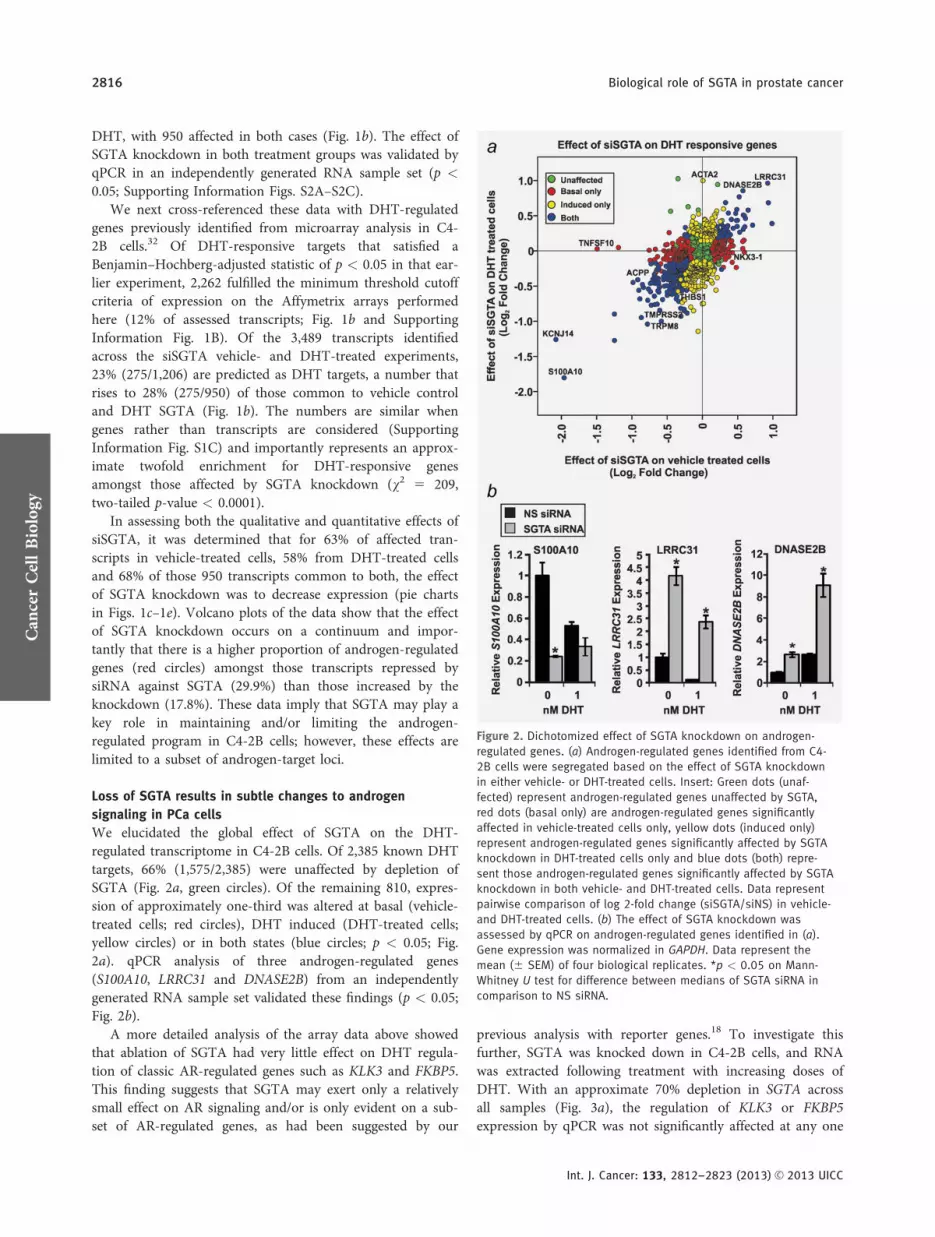
DHT with 950 affected in both cases (Fig 1b) The effect ofSGTA knockdown in both treatment groups was validated byqPCR in an independently generated RNA sample set (p lt005 Supporting Information Figs S2AndashS2C)
We next cross-referenced these data with DHT-regulatedgenes previously identified from microarray analysis in C4-2B cells32 Of DHT-responsive targets that satisfied aBenjaminndashHochberg-adjusted statistic of p lt 005 in that ear-lier experiment 2262 fulfilled the minimum threshold cutoffcriteria of expression on the Affymetrix arrays performedhere (12 of assessed transcripts Fig 1b and SupportingInformation Fig 1B) Of the 3489 transcripts identifiedacross the siSGTA vehicle- and DHT-treated experiments23 (2751206) are predicted as DHT targets a number thatrises to 28 (275950) of those common to vehicle controland DHT SGTA (Fig 1b) The numbers are similar whengenes rather than transcripts are considered (SupportingInformation Fig S1C) and importantly represents an approx-imate twofold enrichment for DHT-responsive genesamongst those affected by SGTA knockdown (v2 5 209two-tailed p-value lt 00001)
In assessing both the qualitative and quantitative effects ofsiSGTA it was determined that for 63 of affected tran-scripts in vehicle-treated cells 58 from DHT-treated cellsand 68 of those 950 transcripts common to both the effectof SGTA knockdown was to decrease expression (pie chartsin Figs 1cndash1e) Volcano plots of the data show that the effectof SGTA knockdown occurs on a continuum and impor-tantly that there is a higher proportion of androgen-regulatedgenes (red circles) amongst those transcripts repressed bysiRNA against SGTA (299) than those increased by theknockdown (178) These data imply that SGTA may play akey role in maintaining andor limiting the androgen-regulated program in C4-2B cells however these effects arelimited to a subset of androgen-target loci
Loss of SGTA results in subtle changes to androgen
signaling in PCa cells
We elucidated the global effect of SGTA on the DHT-regulated transcriptome in C4-2B cells Of 2385 known DHTtargets 66 (15752385) were unaffected by depletion ofSGTA (Fig 2a green circles) Of the remaining 810 expres-sion of approximately one-third was altered at basal (vehicle-treated cells red circles) DHT induced (DHT-treated cellsyellow circles) or in both states (blue circles p lt 005 Fig2a) qPCR analysis of three androgen-regulated genes(S100A10 LRRC31 and DNASE2B) from an independentlygenerated RNA sample set validated these findings (p lt 005Fig 2b)
A more detailed analysis of the array data above showedthat ablation of SGTA had very little effect on DHT regula-tion of classic AR-regulated genes such as KLK3 and FKBP5This finding suggests that SGTA may exert only a relativelysmall effect on AR signaling andor is only evident on a sub-set of AR-regulated genes as had been suggested by our
previous analysis with reporter genes18 To investigate thisfurther SGTA was knocked down in C4-2B cells and RNAwas extracted following treatment with increasing doses ofDHT With an approximate 70 depletion in SGTA acrossall samples (Fig 3a) the regulation of KLK3 or FKBP5expression by qPCR was not significantly affected at any one
Figure 2 Dichotomized effect of SGTA knockdown on androgen-
regulated genes (a) Androgen-regulated genes identified from C4-
2B cells were segregated based on the effect of SGTA knockdown
in either vehicle- or DHT-treated cells Insert Green dots (unaf-
fected) represent androgen-regulated genes unaffected by SGTA
red dots (basal only) are androgen-regulated genes significantly
affected in vehicle-treated cells only yellow dots (induced only)
represent androgen-regulated genes significantly affected by SGTA
knockdown in DHT-treated cells only and blue dots (both) repre-
sent those androgen-regulated genes significantly affected by SGTA
knockdown in both vehicle- and DHT-treated cells Data represent
pairwise comparison of log 2-fold change (siSGTAsiNS) in vehicle-
and DHT-treated cells (b) The effect of SGTA knockdown was
assessed by qPCR on androgen-regulated genes identified in (a)
Gene expression was normalized in GAPDH Data represent the
mean (6 SEM) of four biological replicates p lt 005 on Mann-
Whitney U test for difference between medians of SGTA siRNA in
comparison to NS siRNA
Can
cerCellBiology
2816 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
dose of DHT (Figs 3b and 3c) However a consistent smalleffect of SGTA knockdown on regulation across all DHTconcentrations was observed (p lt 005 Figs 3b and 3c)although no overt effect was observed at FKBP51 steady-stateprotein levels (Fig 3d) Collectively these data confirm thatSGTA exerts an influence on only a subset of androgen-regulated genes and suggests that its effects on AR proteinfolding and trafficking in the presence of fluctuating andro-gen levels may hold greater consequence for PCa cellfunction
Knockdown of SGTA modulates the expression of p53 and
phosphoinositol kinaseAkt pathways
Although the above results suggest that SGTA is a key regu-lator of a subset of AR target genes we sought to elucidateits broader biological significance in PCa cells Ingenuitypathway analysis of the microarray data identified a numberof important cellular pathways significantly altered by SGTAknockdown in vehicle- or DHT-treated cells or both (Sup-porting Information Fig S3) Similar pathways were enrichedin the gene sets obtained from both vehicle- and DHT-
treated cells implying a relatively consistent effect of SGTAthat is largely independent of exogenous hormone Amongthose pathways the most highly enriched were p53 PCa andphosphoinositol kinase (PI3K)Akt signaling pathways (p lt
005 Fig 4a) Enrichment of these pathways was confirmedfor both DHT- and vehicle-treated cells using GSEA29 (Figs4bndash4d) and DAVID30 (Supporting Information Tables S2 andS3) GSEA further implicated upregulation of PCa and PI3Kpathways by SGTA knockdown but downregulation of p53signaling in both vehicle- and DHT-treated gene sets (Figs4bndash4d and Supporting Information Fig S4) Additionally thetop four of five protein families enriched in SGTA-depletedsamples related to protein kinase or serinethreonine kinaseactivity in DAVID (Supporting Information Tables S4 andS5) suggesting that SGTA may play a role in maintainingandor modulating these activities in PCa cells IndeedSGTA has been shown to maintain receptor tyrosine kinasesignaling and Akt activity in a nonsmall cell lung cancer(NSCLC) cell line1725
The effect of SGTA knockdown on specific genes withineach of the three GSEA pathways listed above was validated
Figure 3 SGTA knockdown does not affect AR-dependent KLK3 and FKBP5 gene expression (andashc) The effect of SGTA knockdown on AR sig-
naling in C4-2B cells was assessed by qPCR Cells were transfected with 5 nM NS or SGTA siRNA for 72 hr followed by treatment with 0ndash10
nM DHT for 16 hr prior to RNA extraction qPCR was performed on SGTA KLK3 and FKBP5 in triplicate samples Error bars represent (6
SEM) of three biological replicates Statistical significance represents Friedman repeated measures test for differences in means between
SGTA siRNA and NS siRNA p lt 005 (d) Matching protein lysates to the RNA samples used in (a)ndash(c) were generated for immunoblot anal-
ysis on SGTA FKBP51 and AR steady-state protein levels Hsp90 was used as a loading control
Can
cerCellBiology
Trotta et al 2817
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
Figure 4 SGTA knockdown affects p53 PI3 kinase and prostate cancer signaling pathways (a) Ingenuity pathway analysis of canonical
pathways affected by SGTA knockdown in vehicle- and DHT-treated cells or both (bndashd) Association of SGTA knockdown vehicle-treated
datasets in prostate cancer phosphatidylinositol signaling system and p53 signaling as a pathway as assessed by GSEA Probe sets from
the SGTA knockdown arrays were collapsed to the gene level and assigned a score based on a signal-to-noise ratio algorithm and ranked-
ordered by this score29 The running enrichment score (green line) is then plotted (top panel) The enrichment of the KEGG Prostate Cancer
gene set in the ranked-ordered SGTA knockdown dataset is represented by black lines (middle panel) positive to negative value of ranking
metric and the correlation of genes within the KEGG pathway to SGTA knockdown dataset (bottom panel) The normalized enrichment score
(NES) for enriched pathways is presented in each panel qPCR validation of genes is associated with both KEGG pathways Data represent
mean (6 SEM) of four biological replicates p lt 005 and p lt 001 on Mann-Whitney U test for difference between medians of SGTA
siRNA in comparison to NS siRNA
Can
cerCellBiology
2818 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
using qPCR with some variation depending on treatmentwith vehicle or DHT Specifically analysis of PCa signalingpathway genes AR GRB2 and ERBB2 demonstrated an effectonly in the presence of DHT (p lt 005 Fig 4b) For thePI3K signaling pathway PIK3R1 and PIK3CB expressiondecreased slightly but was significantly affected with DHTtreatment but only in the knockdown samples PIK3CDexpression in contrast was affected by both vehicle and DHTtreatments (p lt 001 Fig 4c) Finally we observed upregula-tion of TP53 with SGTA knockdown but a significantdecrease in expression of the two p53 target genes MDM2and CDKN1A (also known as p21 p lt 005 Fig 4d) Theabove data implicate SGTA in a number of pathways previ-ously shown to be essential to PCa cell survival proliferationand metastasis The loss of SGTA in PCa might thus be pre-dicted to have a significant effect on disease progression
SGTA acts as a positive regulator of Akt activity and C4-2B
cell viability
We next investigated the effect of SGTA knockdown on p53and PI3 kinase pathways in C4-2B cells by assessing pathwaycomponent proteins and their downstream targets Analysisof p53 signaling by immunoblotting did not reveal anychanges in steady-state protein level of the p53 target p21whereas SGTA knockdown appears to reduce Mdm2 levelseither with or without DHT (Fig 5a) However neither totalp53 nor phosphorylated p53 (S15) levels were altered bySGTA knockdown (Fig 5a) This disconnection with themRNA expression data may indicate a specific temporalndashspa-tial protein response not captured in these studies In addi-tion SGTA knockdown did not affect PI3 kinase subunit p85(PI3KR1) levels but decreased basal p110b (PI3KCB) proteinlevels (Fig 5b) Consistent with a recent report that p110b
maintains Akt activity in PCa cells as measured by phospho-rylation of the T308 and S473 residues of Akt33 the decreasein p110b with SGTA knockdown translated to reduced levelsof phosphorylated Akt T308 and Akt S473 respectively (Fig5b) These effects were partially reversed by treatment with 1nM DHT but could not be attributed to a decrease in totalAkt The effect of SGTA knockdown on p110b protein levelsand Akt phosphorylation status was verified in LNCaP PCacells (Supporting Information Fig S5A)
Given the role of SGTA in key signaling pathways wenext investigated the role of SGTA in PCa cell growthAlthough SGTA knockdown did not affect viability of C4-2Bcells when assayed under stripped serum conditions (Fig 5c)a 524 decrease in siSGTA cell number compared to mockcontrol at Day 6 was observed when assays were performedwith normal FBS containing media (p lt 005 Fig 5d) Theeffect of SGTA knockdown on C4-2B cell proliferation in thepresence of normal FBS was validated by MTT assays (Sup-porting Information Fig S5B) Immunoblot analysis demon-strated SGTA depletion for the duration of both experiments(Figs 5e and 5f) whereas trypan-blue exclusion showed thatcell death was relatively consistent across all treatment groups
(Supporting Information Figs S5C and S5D) The role ofSGTA appears to be dependent on growth of cells in fullserum which might be consistent with activation of growthfactor signaling pathways dependent on intracellular PI3KAkt signaling
SGTA and AR protein levels in patient-matched BPH and
cancer samples
The convergence of SGTA on AR and PIK3Akt signalingpathways which have important roles in PCa etiology sug-gests that altered SGTA levels may critically influence PCaprogression Our previous study indirectly suggested this bydemonstration of lower SGTA protein levels in metastaticsamples compared to primary disease17 Immunohistochemis-try was used in our study on a clinical cohort of 66 primaryadvanced PCa samples and patient-matched BPH tissues tomore precisely define any relationship between SGTA andAR levels and clinically significant outcome parameters
AR protein was found to be predominately localized inthe nucleus of luminal epithelial cells of both BPH and PCasamples with some variable stromal staining (Fig 6a) Incontrast SGTA was mainly confined to epithelial cytoplasmalthough a small number of samples showed stromal staining(Fig 6a) Quantitative analysis identified a slightly highermean AR level in cancer samples compared to BPH andlower SGTA but these did not reach significance (p 5 0055Supporting Information Table S6) Importantly however themean and median ARSGTA ratio was significantly higher inPCa samples than in matched BPH (p 5 0017) This ratiowas also higher in cancer but not BPH of those that haddied from PCa but was not significant (Supporting Informa-tion Table S6) These data imply a cancer-specific dysregula-tion in the expression of AR and SGTA that may potentiatedisease progression
DiscussionOne of the key hallmarks of cancer is the upregulation andmaintenance of key signal transduction pathways that drivecancer cell proliferation and survival5 The molecular chaper-one and cochaperone systems facilitates this in cancer cellsby providing key structural and functional frameworksthrough which these highly proliferative signals can be sus-tained and amplified The PI3KAkt signaling pathway has awell-defined role in PCa initiation and progression3435
whereby approximately 40 of localized PCa and 70 ofmetastases contain genomic alterations within PI3 kinase sig-naling proteins36 Importantly the molecular mechanismsthat contribute to constitutive PI3 kinase signaling haveunderscored recent clinical advances in the development andadministration of therapeutic agents for the treatment ofmetastatic PCa37 The Class 1A PI3 kinase subunits are agroup of enzymes composed of regulatory (p85a p85b p55a
and p55g) subunit proteins that interact with their catalytic(p110a p110b and p110d) counterparts to form heterodi-meric complexes Activation of these complexes by tyrosine
Can
cerCellBiology
Trotta et al 2819
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
kinase receptors at the cell surface mediates subsequent signaltransduction (via Akt phosphorylation) culminating in theregulation of target genes involved in cell survival prolifera-tion growth and migration PI3 kinase subunit p110b isimportant for the maintenance of Akt phosphorylation in PCcells33 and of significance we demonstrated here that regula-tion of cellular p110b levels by SGTA may be important inmaintaining Akt phosphorylation A similar effect of SGTAknockdown on Akt phosphorylation has been observed inNSCLC H1703 cells25 suggesting that SGTA acts as an
important positive regulator of PI3KAkt signaling in multi-ple cancers and can thereby mediate cancer cell proliferationand survival
In addition to the critical role of PI3 kinase signaling inPCa development there is incontrovertible evidence demon-strating that continued AR signaling is required for diseaseprogression to a castrate-resistant state38 Although it isaccepted that regulation of AR transcriptional activity in PCacells can be achieved at multiple stages within the AR activa-tion cascade39 the role of cytoplasmic TPR-containing
Figure 5 Knockdown of SGTA affects PI3K activity and C4-2B cell viability (a and b) C4-2B cells were transfected with siRNA and treated as
described in Figure 1a Immunoblot analysis of SGTA p21 Mdm2 p53 p85 p110b and Akt protein steady-state levels Analysis of active
p53 and Akt signaling was determined by phosphorylation status of p53 serine 15 (S15) Akt threonine 308 (T308) and Akt S473 residues
Hsp90 was used as a loading control (c and d) C4-2B cells were either mock transfected or transfected with nonspecific (NS) siRNA or
SGTA-specific siRNA in serum-free medium for 4 hr and then supplemented with 5 DCS-FBS or FBS Cells were cultured for 6 days and
counted every 2 days using a hemocytometer and cell viability was assessed by Trypan blue exclusion Data are presented as the mean
(6 SEM) of triplicate wells from two independent experiments Two-way ANOVA was used to determine the difference in cell growth
between groups p lt 005 differences between mockNS transfected cells versus SGTA siRNA (e and f) Cell lysates from growth assay
were assessed for SGTA steady-state protein levels b-Actin was used as a loading control [Color figure can be viewed in the online issue
which is available at wileyonlinelibrarycom]
Can
cerCellBiology
2820 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
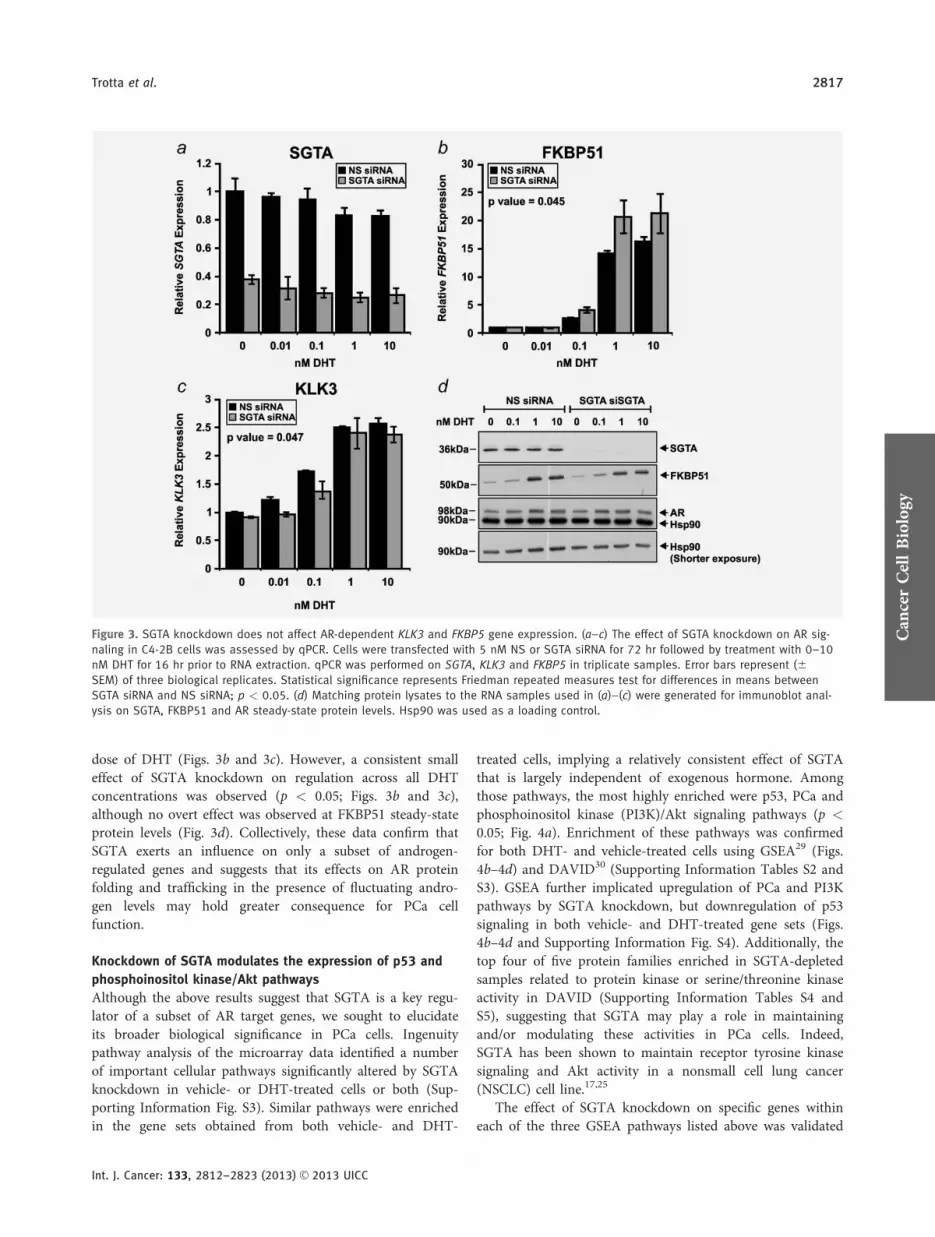
dose of DHT (Figs 3b and 3c) However a consistent smalleffect of SGTA knockdown on regulation across all DHTconcentrations was observed (p lt 005 Figs 3b and 3c)although no overt effect was observed at FKBP51 steady-stateprotein levels (Fig 3d) Collectively these data confirm thatSGTA exerts an influence on only a subset of androgen-regulated genes and suggests that its effects on AR proteinfolding and trafficking in the presence of fluctuating andro-gen levels may hold greater consequence for PCa cellfunction
Knockdown of SGTA modulates the expression of p53 and
phosphoinositol kinaseAkt pathways
Although the above results suggest that SGTA is a key regu-lator of a subset of AR target genes we sought to elucidateits broader biological significance in PCa cells Ingenuitypathway analysis of the microarray data identified a numberof important cellular pathways significantly altered by SGTAknockdown in vehicle- or DHT-treated cells or both (Sup-porting Information Fig S3) Similar pathways were enrichedin the gene sets obtained from both vehicle- and DHT-
treated cells implying a relatively consistent effect of SGTAthat is largely independent of exogenous hormone Amongthose pathways the most highly enriched were p53 PCa andphosphoinositol kinase (PI3K)Akt signaling pathways (p lt
005 Fig 4a) Enrichment of these pathways was confirmedfor both DHT- and vehicle-treated cells using GSEA29 (Figs4bndash4d) and DAVID30 (Supporting Information Tables S2 andS3) GSEA further implicated upregulation of PCa and PI3Kpathways by SGTA knockdown but downregulation of p53signaling in both vehicle- and DHT-treated gene sets (Figs4bndash4d and Supporting Information Fig S4) Additionally thetop four of five protein families enriched in SGTA-depletedsamples related to protein kinase or serinethreonine kinaseactivity in DAVID (Supporting Information Tables S4 andS5) suggesting that SGTA may play a role in maintainingandor modulating these activities in PCa cells IndeedSGTA has been shown to maintain receptor tyrosine kinasesignaling and Akt activity in a nonsmall cell lung cancer(NSCLC) cell line1725
The effect of SGTA knockdown on specific genes withineach of the three GSEA pathways listed above was validated
Figure 3 SGTA knockdown does not affect AR-dependent KLK3 and FKBP5 gene expression (andashc) The effect of SGTA knockdown on AR sig-
naling in C4-2B cells was assessed by qPCR Cells were transfected with 5 nM NS or SGTA siRNA for 72 hr followed by treatment with 0ndash10
nM DHT for 16 hr prior to RNA extraction qPCR was performed on SGTA KLK3 and FKBP5 in triplicate samples Error bars represent (6
SEM) of three biological replicates Statistical significance represents Friedman repeated measures test for differences in means between
SGTA siRNA and NS siRNA p lt 005 (d) Matching protein lysates to the RNA samples used in (a)ndash(c) were generated for immunoblot anal-
ysis on SGTA FKBP51 and AR steady-state protein levels Hsp90 was used as a loading control
Can
cerCellBiology
Trotta et al 2817
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
Figure 4 SGTA knockdown affects p53 PI3 kinase and prostate cancer signaling pathways (a) Ingenuity pathway analysis of canonical
pathways affected by SGTA knockdown in vehicle- and DHT-treated cells or both (bndashd) Association of SGTA knockdown vehicle-treated
datasets in prostate cancer phosphatidylinositol signaling system and p53 signaling as a pathway as assessed by GSEA Probe sets from
the SGTA knockdown arrays were collapsed to the gene level and assigned a score based on a signal-to-noise ratio algorithm and ranked-
ordered by this score29 The running enrichment score (green line) is then plotted (top panel) The enrichment of the KEGG Prostate Cancer
gene set in the ranked-ordered SGTA knockdown dataset is represented by black lines (middle panel) positive to negative value of ranking
metric and the correlation of genes within the KEGG pathway to SGTA knockdown dataset (bottom panel) The normalized enrichment score
(NES) for enriched pathways is presented in each panel qPCR validation of genes is associated with both KEGG pathways Data represent
mean (6 SEM) of four biological replicates p lt 005 and p lt 001 on Mann-Whitney U test for difference between medians of SGTA
siRNA in comparison to NS siRNA
Can
cerCellBiology
2818 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
using qPCR with some variation depending on treatmentwith vehicle or DHT Specifically analysis of PCa signalingpathway genes AR GRB2 and ERBB2 demonstrated an effectonly in the presence of DHT (p lt 005 Fig 4b) For thePI3K signaling pathway PIK3R1 and PIK3CB expressiondecreased slightly but was significantly affected with DHTtreatment but only in the knockdown samples PIK3CDexpression in contrast was affected by both vehicle and DHTtreatments (p lt 001 Fig 4c) Finally we observed upregula-tion of TP53 with SGTA knockdown but a significantdecrease in expression of the two p53 target genes MDM2and CDKN1A (also known as p21 p lt 005 Fig 4d) Theabove data implicate SGTA in a number of pathways previ-ously shown to be essential to PCa cell survival proliferationand metastasis The loss of SGTA in PCa might thus be pre-dicted to have a significant effect on disease progression
SGTA acts as a positive regulator of Akt activity and C4-2B
cell viability
We next investigated the effect of SGTA knockdown on p53and PI3 kinase pathways in C4-2B cells by assessing pathwaycomponent proteins and their downstream targets Analysisof p53 signaling by immunoblotting did not reveal anychanges in steady-state protein level of the p53 target p21whereas SGTA knockdown appears to reduce Mdm2 levelseither with or without DHT (Fig 5a) However neither totalp53 nor phosphorylated p53 (S15) levels were altered bySGTA knockdown (Fig 5a) This disconnection with themRNA expression data may indicate a specific temporalndashspa-tial protein response not captured in these studies In addi-tion SGTA knockdown did not affect PI3 kinase subunit p85(PI3KR1) levels but decreased basal p110b (PI3KCB) proteinlevels (Fig 5b) Consistent with a recent report that p110b
maintains Akt activity in PCa cells as measured by phospho-rylation of the T308 and S473 residues of Akt33 the decreasein p110b with SGTA knockdown translated to reduced levelsof phosphorylated Akt T308 and Akt S473 respectively (Fig5b) These effects were partially reversed by treatment with 1nM DHT but could not be attributed to a decrease in totalAkt The effect of SGTA knockdown on p110b protein levelsand Akt phosphorylation status was verified in LNCaP PCacells (Supporting Information Fig S5A)
Given the role of SGTA in key signaling pathways wenext investigated the role of SGTA in PCa cell growthAlthough SGTA knockdown did not affect viability of C4-2Bcells when assayed under stripped serum conditions (Fig 5c)a 524 decrease in siSGTA cell number compared to mockcontrol at Day 6 was observed when assays were performedwith normal FBS containing media (p lt 005 Fig 5d) Theeffect of SGTA knockdown on C4-2B cell proliferation in thepresence of normal FBS was validated by MTT assays (Sup-porting Information Fig S5B) Immunoblot analysis demon-strated SGTA depletion for the duration of both experiments(Figs 5e and 5f) whereas trypan-blue exclusion showed thatcell death was relatively consistent across all treatment groups
(Supporting Information Figs S5C and S5D) The role ofSGTA appears to be dependent on growth of cells in fullserum which might be consistent with activation of growthfactor signaling pathways dependent on intracellular PI3KAkt signaling
SGTA and AR protein levels in patient-matched BPH and
cancer samples
The convergence of SGTA on AR and PIK3Akt signalingpathways which have important roles in PCa etiology sug-gests that altered SGTA levels may critically influence PCaprogression Our previous study indirectly suggested this bydemonstration of lower SGTA protein levels in metastaticsamples compared to primary disease17 Immunohistochemis-try was used in our study on a clinical cohort of 66 primaryadvanced PCa samples and patient-matched BPH tissues tomore precisely define any relationship between SGTA andAR levels and clinically significant outcome parameters
AR protein was found to be predominately localized inthe nucleus of luminal epithelial cells of both BPH and PCasamples with some variable stromal staining (Fig 6a) Incontrast SGTA was mainly confined to epithelial cytoplasmalthough a small number of samples showed stromal staining(Fig 6a) Quantitative analysis identified a slightly highermean AR level in cancer samples compared to BPH andlower SGTA but these did not reach significance (p 5 0055Supporting Information Table S6) Importantly however themean and median ARSGTA ratio was significantly higher inPCa samples than in matched BPH (p 5 0017) This ratiowas also higher in cancer but not BPH of those that haddied from PCa but was not significant (Supporting Informa-tion Table S6) These data imply a cancer-specific dysregula-tion in the expression of AR and SGTA that may potentiatedisease progression
DiscussionOne of the key hallmarks of cancer is the upregulation andmaintenance of key signal transduction pathways that drivecancer cell proliferation and survival5 The molecular chaper-one and cochaperone systems facilitates this in cancer cellsby providing key structural and functional frameworksthrough which these highly proliferative signals can be sus-tained and amplified The PI3KAkt signaling pathway has awell-defined role in PCa initiation and progression3435
whereby approximately 40 of localized PCa and 70 ofmetastases contain genomic alterations within PI3 kinase sig-naling proteins36 Importantly the molecular mechanismsthat contribute to constitutive PI3 kinase signaling haveunderscored recent clinical advances in the development andadministration of therapeutic agents for the treatment ofmetastatic PCa37 The Class 1A PI3 kinase subunits are agroup of enzymes composed of regulatory (p85a p85b p55a
and p55g) subunit proteins that interact with their catalytic(p110a p110b and p110d) counterparts to form heterodi-meric complexes Activation of these complexes by tyrosine
Can
cerCellBiology
Trotta et al 2819
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
kinase receptors at the cell surface mediates subsequent signaltransduction (via Akt phosphorylation) culminating in theregulation of target genes involved in cell survival prolifera-tion growth and migration PI3 kinase subunit p110b isimportant for the maintenance of Akt phosphorylation in PCcells33 and of significance we demonstrated here that regula-tion of cellular p110b levels by SGTA may be important inmaintaining Akt phosphorylation A similar effect of SGTAknockdown on Akt phosphorylation has been observed inNSCLC H1703 cells25 suggesting that SGTA acts as an
important positive regulator of PI3KAkt signaling in multi-ple cancers and can thereby mediate cancer cell proliferationand survival
In addition to the critical role of PI3 kinase signaling inPCa development there is incontrovertible evidence demon-strating that continued AR signaling is required for diseaseprogression to a castrate-resistant state38 Although it isaccepted that regulation of AR transcriptional activity in PCacells can be achieved at multiple stages within the AR activa-tion cascade39 the role of cytoplasmic TPR-containing
Figure 5 Knockdown of SGTA affects PI3K activity and C4-2B cell viability (a and b) C4-2B cells were transfected with siRNA and treated as
described in Figure 1a Immunoblot analysis of SGTA p21 Mdm2 p53 p85 p110b and Akt protein steady-state levels Analysis of active
p53 and Akt signaling was determined by phosphorylation status of p53 serine 15 (S15) Akt threonine 308 (T308) and Akt S473 residues
Hsp90 was used as a loading control (c and d) C4-2B cells were either mock transfected or transfected with nonspecific (NS) siRNA or
SGTA-specific siRNA in serum-free medium for 4 hr and then supplemented with 5 DCS-FBS or FBS Cells were cultured for 6 days and
counted every 2 days using a hemocytometer and cell viability was assessed by Trypan blue exclusion Data are presented as the mean
(6 SEM) of triplicate wells from two independent experiments Two-way ANOVA was used to determine the difference in cell growth
between groups p lt 005 differences between mockNS transfected cells versus SGTA siRNA (e and f) Cell lysates from growth assay
were assessed for SGTA steady-state protein levels b-Actin was used as a loading control [Color figure can be viewed in the online issue
which is available at wileyonlinelibrarycom]
Can
cerCellBiology
2820 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
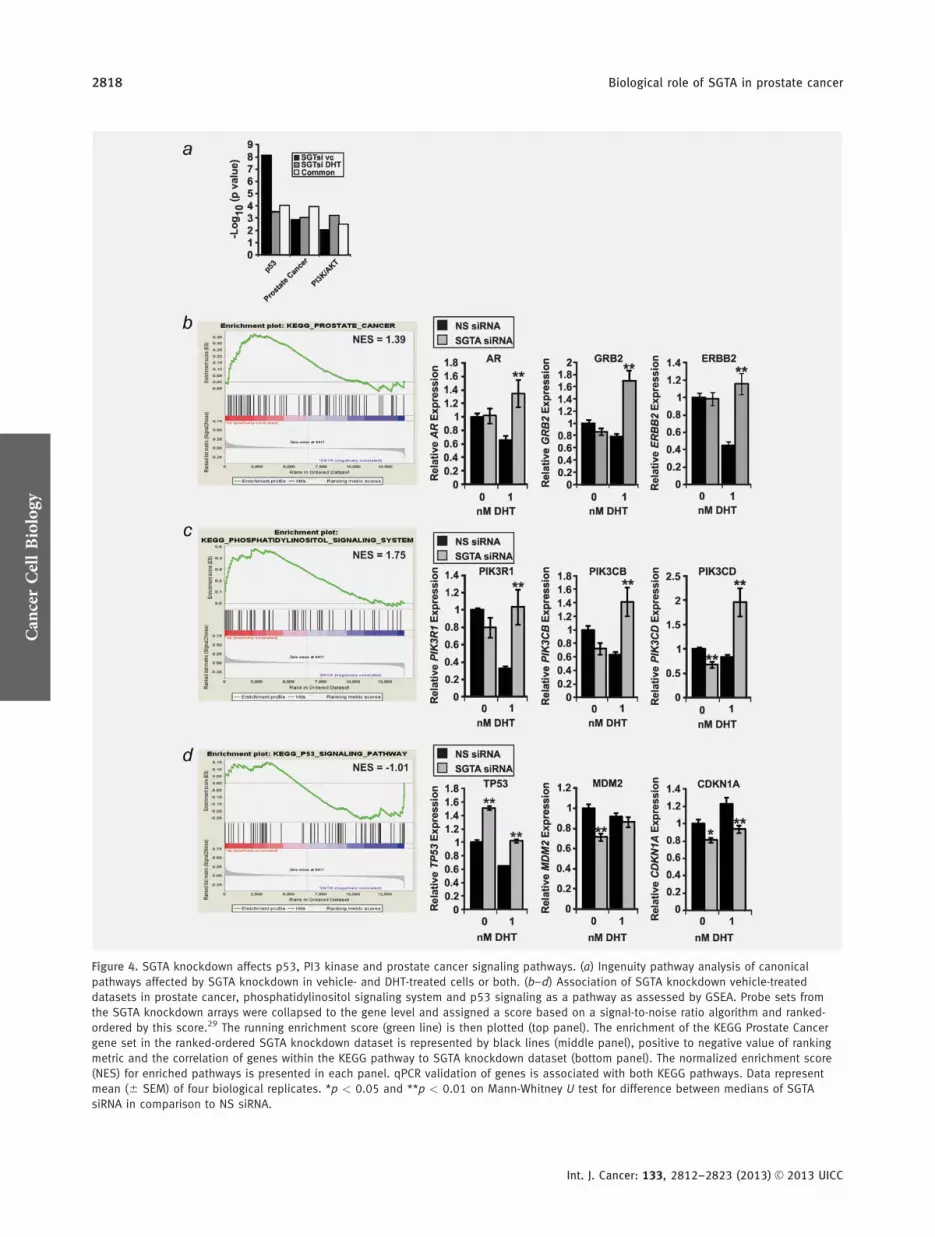
Figure 4 SGTA knockdown affects p53 PI3 kinase and prostate cancer signaling pathways (a) Ingenuity pathway analysis of canonical
pathways affected by SGTA knockdown in vehicle- and DHT-treated cells or both (bndashd) Association of SGTA knockdown vehicle-treated
datasets in prostate cancer phosphatidylinositol signaling system and p53 signaling as a pathway as assessed by GSEA Probe sets from
the SGTA knockdown arrays were collapsed to the gene level and assigned a score based on a signal-to-noise ratio algorithm and ranked-
ordered by this score29 The running enrichment score (green line) is then plotted (top panel) The enrichment of the KEGG Prostate Cancer
gene set in the ranked-ordered SGTA knockdown dataset is represented by black lines (middle panel) positive to negative value of ranking
metric and the correlation of genes within the KEGG pathway to SGTA knockdown dataset (bottom panel) The normalized enrichment score
(NES) for enriched pathways is presented in each panel qPCR validation of genes is associated with both KEGG pathways Data represent
mean (6 SEM) of four biological replicates p lt 005 and p lt 001 on Mann-Whitney U test for difference between medians of SGTA
siRNA in comparison to NS siRNA
Can
cerCellBiology
2818 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
using qPCR with some variation depending on treatmentwith vehicle or DHT Specifically analysis of PCa signalingpathway genes AR GRB2 and ERBB2 demonstrated an effectonly in the presence of DHT (p lt 005 Fig 4b) For thePI3K signaling pathway PIK3R1 and PIK3CB expressiondecreased slightly but was significantly affected with DHTtreatment but only in the knockdown samples PIK3CDexpression in contrast was affected by both vehicle and DHTtreatments (p lt 001 Fig 4c) Finally we observed upregula-tion of TP53 with SGTA knockdown but a significantdecrease in expression of the two p53 target genes MDM2and CDKN1A (also known as p21 p lt 005 Fig 4d) Theabove data implicate SGTA in a number of pathways previ-ously shown to be essential to PCa cell survival proliferationand metastasis The loss of SGTA in PCa might thus be pre-dicted to have a significant effect on disease progression
SGTA acts as a positive regulator of Akt activity and C4-2B
cell viability
We next investigated the effect of SGTA knockdown on p53and PI3 kinase pathways in C4-2B cells by assessing pathwaycomponent proteins and their downstream targets Analysisof p53 signaling by immunoblotting did not reveal anychanges in steady-state protein level of the p53 target p21whereas SGTA knockdown appears to reduce Mdm2 levelseither with or without DHT (Fig 5a) However neither totalp53 nor phosphorylated p53 (S15) levels were altered bySGTA knockdown (Fig 5a) This disconnection with themRNA expression data may indicate a specific temporalndashspa-tial protein response not captured in these studies In addi-tion SGTA knockdown did not affect PI3 kinase subunit p85(PI3KR1) levels but decreased basal p110b (PI3KCB) proteinlevels (Fig 5b) Consistent with a recent report that p110b
maintains Akt activity in PCa cells as measured by phospho-rylation of the T308 and S473 residues of Akt33 the decreasein p110b with SGTA knockdown translated to reduced levelsof phosphorylated Akt T308 and Akt S473 respectively (Fig5b) These effects were partially reversed by treatment with 1nM DHT but could not be attributed to a decrease in totalAkt The effect of SGTA knockdown on p110b protein levelsand Akt phosphorylation status was verified in LNCaP PCacells (Supporting Information Fig S5A)
Given the role of SGTA in key signaling pathways wenext investigated the role of SGTA in PCa cell growthAlthough SGTA knockdown did not affect viability of C4-2Bcells when assayed under stripped serum conditions (Fig 5c)a 524 decrease in siSGTA cell number compared to mockcontrol at Day 6 was observed when assays were performedwith normal FBS containing media (p lt 005 Fig 5d) Theeffect of SGTA knockdown on C4-2B cell proliferation in thepresence of normal FBS was validated by MTT assays (Sup-porting Information Fig S5B) Immunoblot analysis demon-strated SGTA depletion for the duration of both experiments(Figs 5e and 5f) whereas trypan-blue exclusion showed thatcell death was relatively consistent across all treatment groups
(Supporting Information Figs S5C and S5D) The role ofSGTA appears to be dependent on growth of cells in fullserum which might be consistent with activation of growthfactor signaling pathways dependent on intracellular PI3KAkt signaling
SGTA and AR protein levels in patient-matched BPH and
cancer samples
The convergence of SGTA on AR and PIK3Akt signalingpathways which have important roles in PCa etiology sug-gests that altered SGTA levels may critically influence PCaprogression Our previous study indirectly suggested this bydemonstration of lower SGTA protein levels in metastaticsamples compared to primary disease17 Immunohistochemis-try was used in our study on a clinical cohort of 66 primaryadvanced PCa samples and patient-matched BPH tissues tomore precisely define any relationship between SGTA andAR levels and clinically significant outcome parameters
AR protein was found to be predominately localized inthe nucleus of luminal epithelial cells of both BPH and PCasamples with some variable stromal staining (Fig 6a) Incontrast SGTA was mainly confined to epithelial cytoplasmalthough a small number of samples showed stromal staining(Fig 6a) Quantitative analysis identified a slightly highermean AR level in cancer samples compared to BPH andlower SGTA but these did not reach significance (p 5 0055Supporting Information Table S6) Importantly however themean and median ARSGTA ratio was significantly higher inPCa samples than in matched BPH (p 5 0017) This ratiowas also higher in cancer but not BPH of those that haddied from PCa but was not significant (Supporting Informa-tion Table S6) These data imply a cancer-specific dysregula-tion in the expression of AR and SGTA that may potentiatedisease progression
DiscussionOne of the key hallmarks of cancer is the upregulation andmaintenance of key signal transduction pathways that drivecancer cell proliferation and survival5 The molecular chaper-one and cochaperone systems facilitates this in cancer cellsby providing key structural and functional frameworksthrough which these highly proliferative signals can be sus-tained and amplified The PI3KAkt signaling pathway has awell-defined role in PCa initiation and progression3435
whereby approximately 40 of localized PCa and 70 ofmetastases contain genomic alterations within PI3 kinase sig-naling proteins36 Importantly the molecular mechanismsthat contribute to constitutive PI3 kinase signaling haveunderscored recent clinical advances in the development andadministration of therapeutic agents for the treatment ofmetastatic PCa37 The Class 1A PI3 kinase subunits are agroup of enzymes composed of regulatory (p85a p85b p55a
and p55g) subunit proteins that interact with their catalytic(p110a p110b and p110d) counterparts to form heterodi-meric complexes Activation of these complexes by tyrosine
Can
cerCellBiology
Trotta et al 2819
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
kinase receptors at the cell surface mediates subsequent signaltransduction (via Akt phosphorylation) culminating in theregulation of target genes involved in cell survival prolifera-tion growth and migration PI3 kinase subunit p110b isimportant for the maintenance of Akt phosphorylation in PCcells33 and of significance we demonstrated here that regula-tion of cellular p110b levels by SGTA may be important inmaintaining Akt phosphorylation A similar effect of SGTAknockdown on Akt phosphorylation has been observed inNSCLC H1703 cells25 suggesting that SGTA acts as an
important positive regulator of PI3KAkt signaling in multi-ple cancers and can thereby mediate cancer cell proliferationand survival
In addition to the critical role of PI3 kinase signaling inPCa development there is incontrovertible evidence demon-strating that continued AR signaling is required for diseaseprogression to a castrate-resistant state38 Although it isaccepted that regulation of AR transcriptional activity in PCacells can be achieved at multiple stages within the AR activa-tion cascade39 the role of cytoplasmic TPR-containing
Figure 5 Knockdown of SGTA affects PI3K activity and C4-2B cell viability (a and b) C4-2B cells were transfected with siRNA and treated as
described in Figure 1a Immunoblot analysis of SGTA p21 Mdm2 p53 p85 p110b and Akt protein steady-state levels Analysis of active
p53 and Akt signaling was determined by phosphorylation status of p53 serine 15 (S15) Akt threonine 308 (T308) and Akt S473 residues
Hsp90 was used as a loading control (c and d) C4-2B cells were either mock transfected or transfected with nonspecific (NS) siRNA or
SGTA-specific siRNA in serum-free medium for 4 hr and then supplemented with 5 DCS-FBS or FBS Cells were cultured for 6 days and
counted every 2 days using a hemocytometer and cell viability was assessed by Trypan blue exclusion Data are presented as the mean
(6 SEM) of triplicate wells from two independent experiments Two-way ANOVA was used to determine the difference in cell growth
between groups p lt 005 differences between mockNS transfected cells versus SGTA siRNA (e and f) Cell lysates from growth assay
were assessed for SGTA steady-state protein levels b-Actin was used as a loading control [Color figure can be viewed in the online issue
which is available at wileyonlinelibrarycom]
Can
cerCellBiology
2820 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC

using qPCR with some variation depending on treatmentwith vehicle or DHT Specifically analysis of PCa signalingpathway genes AR GRB2 and ERBB2 demonstrated an effectonly in the presence of DHT (p lt 005 Fig 4b) For thePI3K signaling pathway PIK3R1 and PIK3CB expressiondecreased slightly but was significantly affected with DHTtreatment but only in the knockdown samples PIK3CDexpression in contrast was affected by both vehicle and DHTtreatments (p lt 001 Fig 4c) Finally we observed upregula-tion of TP53 with SGTA knockdown but a significantdecrease in expression of the two p53 target genes MDM2and CDKN1A (also known as p21 p lt 005 Fig 4d) Theabove data implicate SGTA in a number of pathways previ-ously shown to be essential to PCa cell survival proliferationand metastasis The loss of SGTA in PCa might thus be pre-dicted to have a significant effect on disease progression
SGTA acts as a positive regulator of Akt activity and C4-2B
cell viability
We next investigated the effect of SGTA knockdown on p53and PI3 kinase pathways in C4-2B cells by assessing pathwaycomponent proteins and their downstream targets Analysisof p53 signaling by immunoblotting did not reveal anychanges in steady-state protein level of the p53 target p21whereas SGTA knockdown appears to reduce Mdm2 levelseither with or without DHT (Fig 5a) However neither totalp53 nor phosphorylated p53 (S15) levels were altered bySGTA knockdown (Fig 5a) This disconnection with themRNA expression data may indicate a specific temporalndashspa-tial protein response not captured in these studies In addi-tion SGTA knockdown did not affect PI3 kinase subunit p85(PI3KR1) levels but decreased basal p110b (PI3KCB) proteinlevels (Fig 5b) Consistent with a recent report that p110b
maintains Akt activity in PCa cells as measured by phospho-rylation of the T308 and S473 residues of Akt33 the decreasein p110b with SGTA knockdown translated to reduced levelsof phosphorylated Akt T308 and Akt S473 respectively (Fig5b) These effects were partially reversed by treatment with 1nM DHT but could not be attributed to a decrease in totalAkt The effect of SGTA knockdown on p110b protein levelsand Akt phosphorylation status was verified in LNCaP PCacells (Supporting Information Fig S5A)
Given the role of SGTA in key signaling pathways wenext investigated the role of SGTA in PCa cell growthAlthough SGTA knockdown did not affect viability of C4-2Bcells when assayed under stripped serum conditions (Fig 5c)a 524 decrease in siSGTA cell number compared to mockcontrol at Day 6 was observed when assays were performedwith normal FBS containing media (p lt 005 Fig 5d) Theeffect of SGTA knockdown on C4-2B cell proliferation in thepresence of normal FBS was validated by MTT assays (Sup-porting Information Fig S5B) Immunoblot analysis demon-strated SGTA depletion for the duration of both experiments(Figs 5e and 5f) whereas trypan-blue exclusion showed thatcell death was relatively consistent across all treatment groups
(Supporting Information Figs S5C and S5D) The role ofSGTA appears to be dependent on growth of cells in fullserum which might be consistent with activation of growthfactor signaling pathways dependent on intracellular PI3KAkt signaling
SGTA and AR protein levels in patient-matched BPH and
cancer samples
The convergence of SGTA on AR and PIK3Akt signalingpathways which have important roles in PCa etiology sug-gests that altered SGTA levels may critically influence PCaprogression Our previous study indirectly suggested this bydemonstration of lower SGTA protein levels in metastaticsamples compared to primary disease17 Immunohistochemis-try was used in our study on a clinical cohort of 66 primaryadvanced PCa samples and patient-matched BPH tissues tomore precisely define any relationship between SGTA andAR levels and clinically significant outcome parameters
AR protein was found to be predominately localized inthe nucleus of luminal epithelial cells of both BPH and PCasamples with some variable stromal staining (Fig 6a) Incontrast SGTA was mainly confined to epithelial cytoplasmalthough a small number of samples showed stromal staining(Fig 6a) Quantitative analysis identified a slightly highermean AR level in cancer samples compared to BPH andlower SGTA but these did not reach significance (p 5 0055Supporting Information Table S6) Importantly however themean and median ARSGTA ratio was significantly higher inPCa samples than in matched BPH (p 5 0017) This ratiowas also higher in cancer but not BPH of those that haddied from PCa but was not significant (Supporting Informa-tion Table S6) These data imply a cancer-specific dysregula-tion in the expression of AR and SGTA that may potentiatedisease progression
DiscussionOne of the key hallmarks of cancer is the upregulation andmaintenance of key signal transduction pathways that drivecancer cell proliferation and survival5 The molecular chaper-one and cochaperone systems facilitates this in cancer cellsby providing key structural and functional frameworksthrough which these highly proliferative signals can be sus-tained and amplified The PI3KAkt signaling pathway has awell-defined role in PCa initiation and progression3435
whereby approximately 40 of localized PCa and 70 ofmetastases contain genomic alterations within PI3 kinase sig-naling proteins36 Importantly the molecular mechanismsthat contribute to constitutive PI3 kinase signaling haveunderscored recent clinical advances in the development andadministration of therapeutic agents for the treatment ofmetastatic PCa37 The Class 1A PI3 kinase subunits are agroup of enzymes composed of regulatory (p85a p85b p55a
and p55g) subunit proteins that interact with their catalytic(p110a p110b and p110d) counterparts to form heterodi-meric complexes Activation of these complexes by tyrosine
Can
cerCellBiology
Trotta et al 2819
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
kinase receptors at the cell surface mediates subsequent signaltransduction (via Akt phosphorylation) culminating in theregulation of target genes involved in cell survival prolifera-tion growth and migration PI3 kinase subunit p110b isimportant for the maintenance of Akt phosphorylation in PCcells33 and of significance we demonstrated here that regula-tion of cellular p110b levels by SGTA may be important inmaintaining Akt phosphorylation A similar effect of SGTAknockdown on Akt phosphorylation has been observed inNSCLC H1703 cells25 suggesting that SGTA acts as an
important positive regulator of PI3KAkt signaling in multi-ple cancers and can thereby mediate cancer cell proliferationand survival
In addition to the critical role of PI3 kinase signaling inPCa development there is incontrovertible evidence demon-strating that continued AR signaling is required for diseaseprogression to a castrate-resistant state38 Although it isaccepted that regulation of AR transcriptional activity in PCacells can be achieved at multiple stages within the AR activa-tion cascade39 the role of cytoplasmic TPR-containing
Figure 5 Knockdown of SGTA affects PI3K activity and C4-2B cell viability (a and b) C4-2B cells were transfected with siRNA and treated as
described in Figure 1a Immunoblot analysis of SGTA p21 Mdm2 p53 p85 p110b and Akt protein steady-state levels Analysis of active
p53 and Akt signaling was determined by phosphorylation status of p53 serine 15 (S15) Akt threonine 308 (T308) and Akt S473 residues
Hsp90 was used as a loading control (c and d) C4-2B cells were either mock transfected or transfected with nonspecific (NS) siRNA or
SGTA-specific siRNA in serum-free medium for 4 hr and then supplemented with 5 DCS-FBS or FBS Cells were cultured for 6 days and
counted every 2 days using a hemocytometer and cell viability was assessed by Trypan blue exclusion Data are presented as the mean
(6 SEM) of triplicate wells from two independent experiments Two-way ANOVA was used to determine the difference in cell growth
between groups p lt 005 differences between mockNS transfected cells versus SGTA siRNA (e and f) Cell lysates from growth assay
were assessed for SGTA steady-state protein levels b-Actin was used as a loading control [Color figure can be viewed in the online issue
which is available at wileyonlinelibrarycom]
Can
cerCellBiology
2820 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
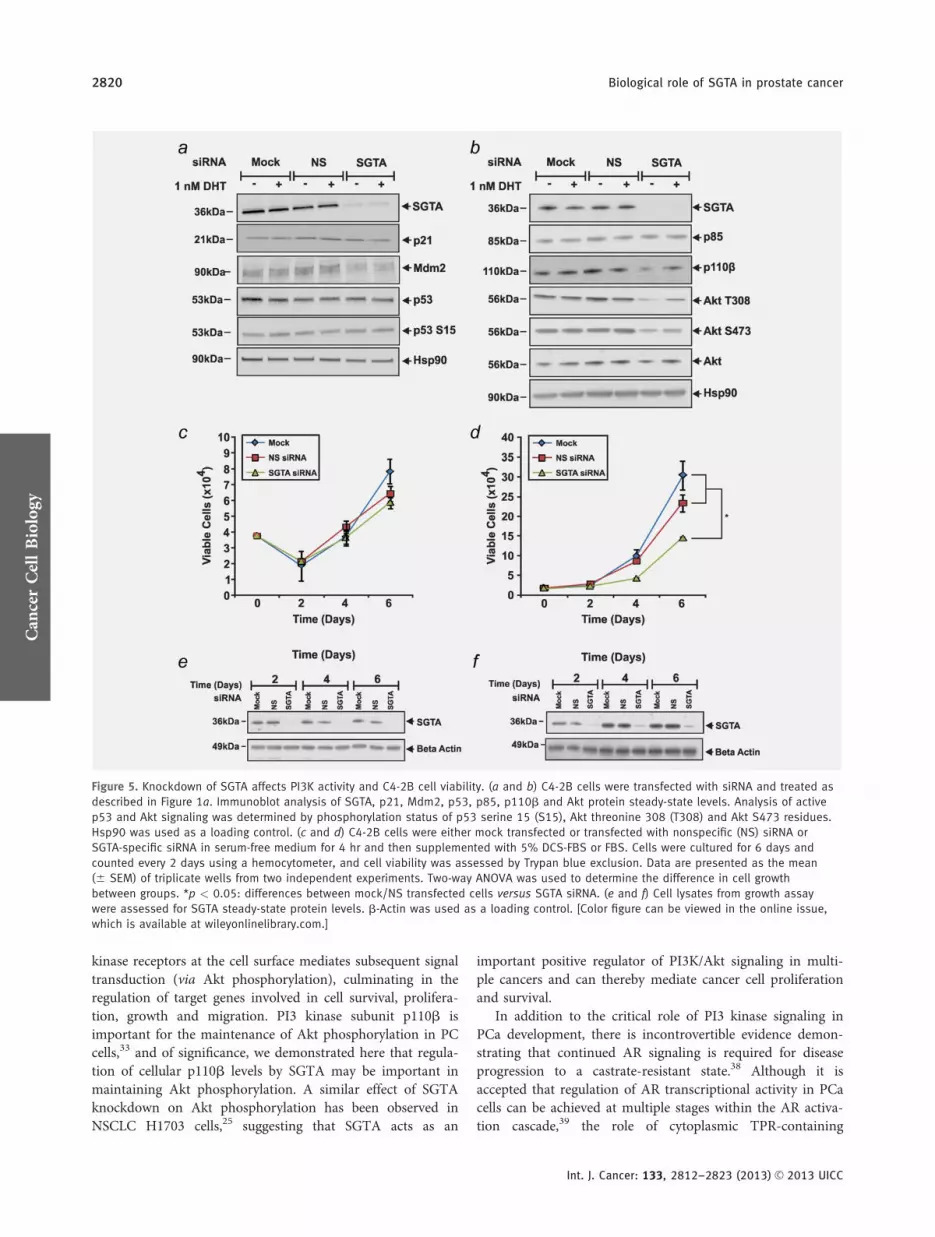
kinase receptors at the cell surface mediates subsequent signaltransduction (via Akt phosphorylation) culminating in theregulation of target genes involved in cell survival prolifera-tion growth and migration PI3 kinase subunit p110b isimportant for the maintenance of Akt phosphorylation in PCcells33 and of significance we demonstrated here that regula-tion of cellular p110b levels by SGTA may be important inmaintaining Akt phosphorylation A similar effect of SGTAknockdown on Akt phosphorylation has been observed inNSCLC H1703 cells25 suggesting that SGTA acts as an
important positive regulator of PI3KAkt signaling in multi-ple cancers and can thereby mediate cancer cell proliferationand survival
In addition to the critical role of PI3 kinase signaling inPCa development there is incontrovertible evidence demon-strating that continued AR signaling is required for diseaseprogression to a castrate-resistant state38 Although it isaccepted that regulation of AR transcriptional activity in PCacells can be achieved at multiple stages within the AR activa-tion cascade39 the role of cytoplasmic TPR-containing
Figure 5 Knockdown of SGTA affects PI3K activity and C4-2B cell viability (a and b) C4-2B cells were transfected with siRNA and treated as
described in Figure 1a Immunoblot analysis of SGTA p21 Mdm2 p53 p85 p110b and Akt protein steady-state levels Analysis of active
p53 and Akt signaling was determined by phosphorylation status of p53 serine 15 (S15) Akt threonine 308 (T308) and Akt S473 residues
Hsp90 was used as a loading control (c and d) C4-2B cells were either mock transfected or transfected with nonspecific (NS) siRNA or
SGTA-specific siRNA in serum-free medium for 4 hr and then supplemented with 5 DCS-FBS or FBS Cells were cultured for 6 days and
counted every 2 days using a hemocytometer and cell viability was assessed by Trypan blue exclusion Data are presented as the mean
(6 SEM) of triplicate wells from two independent experiments Two-way ANOVA was used to determine the difference in cell growth
between groups p lt 005 differences between mockNS transfected cells versus SGTA siRNA (e and f) Cell lysates from growth assay
were assessed for SGTA steady-state protein levels b-Actin was used as a loading control [Color figure can be viewed in the online issue
which is available at wileyonlinelibrarycom]
Can
cerCellBiology
2820 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC

cochaperones in this process particularly how they act in thestabilization of the steroid receptorligand interaction andreceptor transport to the nucleus is often overlooked Previ-ous reports from our laboratory using AR transcriptionalreporter assays suggested that SGTA acts to restrict AR tran-scriptional activity potentially by retaining the receptor inthe cytoplasm1718 Surprisingly our findings presented hereshowed that SGTA appears to play a more complex role inthe regulation of AR activity Although known AR-regulatedgenes were found to be twice as likely to be affected bySGTA knockdown compared to those genes that are unre-sponsive to androgens the effect was bidirectional and medi-ated at both basal- and ligand-induced levels of expressionNonetheless approximately two-thirds of these responseswere suppression of AR-regulated gene expression Althoughit is possible that SGTA may exert direct effects on AR tran-scriptional activity by either modulating AR protein structureandor regulating the rate of cytoplasmicndashnuclear transloca-tion17 it should be noted that secondary effects of SGTAknockdown on other critical AR cofactors such as coregula-tors or pioneer factors may also contribute to changes ingenome-wide transcriptional responses to androgens Indeedwe have previously shown that SGTA overexpression is morelikely to affect AR activity on DNA sites enriched in both ARand FOXA1 binding sites18 suggesting that specific AR DNAbinding elements and the genes they associate with may bemore or less susceptible to the effects of SGTA Neverthelessthese data suggest a model in which the relative level ofSGTA in PCa cells will shift the AR transcriptional programwith the potential clinical importance exemplified by thehigher ARSGTA protein ratio identified in cancerous lesionscompared to benign regions in tissue-matched samples
Therefore it is possible that the relationship between SGTAand AR in prostate epithelial cells is vital to the maintenanceof normal prostate function
Nevertheless it is currently uncertain whether the effectsof SGTA on AR activity are direct or via modulation of othersignaling pathways or a combination of both Indeed ligandindependent or constitutive AR signaling can be generated viacross-talk interactions with PI3KAkt40 Activated Aktincreases both AR mRNA and protein levels41 and the PI3kinase p110b subunit is required for AR DNA binding as wellas androgen-dependent and -independent AR transactiva-tion42 The finding that SGTA ablation resulted in a signifi-cant decrease in p110b protein levels and Aktphosphorylation may explain decreased overall AR transcrip-tional output Given the role of SGTA in regulating Hsc70ATPase activity43 it remains possible that the decreasedp110b protein levels following SGTA knockdown may beattributed to altered molecular chaperone activity andincreased client protein destabilization However both ourexpression microarray and qPCR results indicate that theeffect of SGTA to decrease p110b levels is mediated at themRNA level Regardless of the mechanism it is important tonote that SGTA might act as a key intermediary regulator ofboth PI3KAkt and AR signaling pathways and has importantimplications for maintenance of the homeostatic balancebetween these two signaling pathways in normal prostatecells Taken together the regulatory abilities of SGTA to affectPI3 kinase levels and activity control AR subcellular localiza-tion and restrain AR transcriptional activity signify theimportance of SGTA in dictating specific PCa cell outcomes
The key biological functions of most cochaperones are toprovide stability to clients during protein synthesis to direct
Figure 6 Expression of AR and SGTA in patient-matched clinical samples of BPH and prostate cancer (a and b) Immunohistochemistry with
SGTA or AR antisera Two examples of AR and SGTA expression in patient-matched benign prostatic hyperplasia (BPH) and prostate cancer
samples are shown Patient samples RGH004 and RGH030 were immune-stained for both AR and SGTA All images are at 340 magnifica-
tion (c) Immunoreactivity of each antisera was scored in a minimum of three independent fields by two researchers in both BPH and cancer
regions The scores were collapsed into a representative scale ranging from 0 to 9 Box plots represent the mean AR and SGTA immunore-
activity score Wilcox-ranked sum statistics were used to determine differences of the AR and SGTA mean immunoreactivity score between
BPH and cancer samples respectively
Can
cerCellBiology
Trotta et al 2821
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC

correct cellular localization andor to prevent inappropriateclient activity However it is now becoming clear that cocha-perones play an important role in directly influencing cellviability and survival Indeed the loss of one or more ofthese chaperones may therefore result in additional conse-quences for various cellular processes and thus underscoresthe concept of using the molecular chaperone systems asnovel cancer therapeutic target In our study we observed arequirement for SGTA by PCa cells for optimal proliferationIn agreement previous investigations into the biological func-tions of SGTA also demonstrate that siRNA knockdown ofSGTA mediates a significant reduction in cell viability More-over a recent report demonstrated that ablation of SGTAresults in the destabilization of PDGFRa tyrosine kinasereceptors decreased Akt phosphorylation levels decreasedcell viability and increased cell sensitivity to PI3 kinase inhib-itor inferring that an important cellular function of SGTA isto act as a positive regulator of cell proliferation andsurvival202325
Considering the recent implementations of new-generation chaperone and cochaperone inhibitors currentlybeing trialed against solid tumors444ndash46 elucidating theunderlying mechanisms of chaperonendashsteroid receptor actionsis important to our understanding of how steroid hormone-responsive cancers develop and progress to a life-threateningstage In our study we have uncovered novel biological
properties of the TPR-containing cochaperone SGTA in PCacells whereby depletion of the largely cytoplasmic SGTAcauses substantial changes to the C4-2B PCa cell transcrip-tome Specifically we have described a novel biological func-tion of SGTA in maintaining optimal expression of genes inandrogen and PI3KAkt signaling pathways both of whichhave established and interconnected oncogenic roles in PCainitiation and progression Moreover we have demonstratedthat the SGTA may act to maintain PI3 kinase signaling anda specific AR transcriptional program Alterations in SGTAexpression may thus aberrantly alter the cellular regulatorymechanisms controlling cancer cell proliferation and survival
AcknowledgementsThis study was supported by the National Health and Medical ResearchCouncil of Australia (WDT and LMB ID453662) US Department ofDefense (WDT and LMB IDPC060443) HIHNCI (GACR01CA136924) the Prostate Cancer Foundation (GAC) and the Founda-tion Daw Park (GB) LMB holds a Cancer Council South Australia SeniorResearch Fellowship LAS holds Young Investigator Awards from the USProstate Cancer Foundation and the Prostate Cancer Foundation of Aus-tralia (YI 0810) EFN holds a Hospital Research Foundation Early CareerPostdoctoral Fellowship GB holds a Freemasons Foundation Centre forMenrsquos Health Fellowship The authors thank Drs Li Jia and Baruch Frenkelfor their contribution to the C4-2B microarray data andMr Joey Puccini forhis technical assistance They also thank Dr Georgina England andMs TinaKopsaftis for their contribution to the collection diagnosis and managementof the prostate cancer patient cohort used in this study
References
1 Balch WE Morimoto RI Dillin A et al Adaptingproteostasis for disease intervention Science 2008319916ndash19
2 Taipale M Jarosz DF Lindquist S HSP90 at thehub of protein homeostasis emerging mechanis-tic insights Nat Rev Mol Cell Biol 201011515ndash28
3 Dai C Whitesell L Rogers AB et al Heat shockfactor 1 is a powerful multifaceted modifier ofcarcinogenesis Cell 20071301005ndash18
4 Trepel J Mollapour M Giaccone G et al Target-ing the dynamic HSP90 complex in cancer NatRev 201010537ndash49
5 Hanahan D Weinberg RA Hallmarks of cancerthe next generation Cell 2011144646ndash74
6 Centenera MM Gillis JL Hanson AR et al Evi-dence for efficacy of new Hsp90 inhibitorsrevealed by ex vivo culture of human prostatetumors Clin Cancer Res 2012183562ndash70
7 Kumano M Furukawa J Shiota M et al Cotar-geting stress-activated hsp27 and autophagy as acombinatorial strategy to amplify endoplasmicreticular stress in prostate cancer Mol CancerTher 2012111661ndash71
8 Rocchi P So A Kojima S et al Heat shock pro-tein 27 increases after androgen ablation andplays a cytoprotective role in hormone-refractoryprostate cancer Cancer Res 2004646595ndash602
9 Whitesell L Lindquist SL HSP90 and the chaper-oning of cancer Nat Rev 20055761ndash72
10 Cardillo MR Ippoliti F IL-6 IL-10 and HSP-90expression in tissue microarrays from humanprostate cancer assessed by computer-assistedimage analysis Anticancer Res 2006263409ndash16
11 Kamal A Thao L Sensintaffar J et al A high-affinity conformation of Hsp90 confers tumourselectivity on Hsp90 inhibitors Nature 2003425407ndash10
12 Smith DF Toft DO Minireview The intersectionof steroid receptors with molecular chaperonesobservations and questions Mol Endocrinol (Bal-timore MD) 2008222229ndash40
13 Cheung-Flynn J Prapapanich V Cox MB et alPhysiological role for the cochaperone FKBP52 inandrogen receptor signaling Mol Endocrinol (Bal-timore MD) 2005191654ndash66
14 Yang Z Wolf IM Chen H et al FK506-bindingprotein 52 is essential to uterine reproductivephysiology controlled by the progesterone recep-tor A isoform Mol Endocrinol (Baltimore MD)2006202682ndash94
15 Yong W Yang Z Periyasamy S et al Essentialrole for co-chaperone Fkbp52 but not Fkbp51 inandrogen receptor-mediated signaling and physi-ology J Biol Chem 20072825026ndash36
16 Schulke JP Wochnik GM Lang-Rollin I et alDifferential impact of tetratricopeptide repeatproteins on the steroid hormone receptors PLoSOne 20105e11717
17 Buchanan G Ricciardelli C Harris JM et alControl of androgen receptor signaling in pros-tate cancer by the cochaperone small glutaminerich tetratricopeptide repeat containing proteinalpha Cancer Res 20076710087ndash96
18 Trotta AP Need EF Butler LM et al Subdomainstructure of the co-chaperone SGTA and activityof its androgen receptor client J Mol Endocrinol20124957ndash68
19 Yin H Wang H Zong H et al SGT a Hsp90b
binding partner is accumulated in the nucleusduring cell apoptosis Biochem Biophys Res Com-mun 20063431153ndash8
20 Winnefeld M Rommelaere J Cziepluch C Thehuman small glutamine-rich TPR-containing pro-tein is required for progress through cell divisionExp Cell Res 200429343ndash57
21 Cziepluch C Kordes E Poirey R et al Identifica-tion of a novel cellular TPR-containing proteinSGT that interacts with the nonstructural proteinNS1 of parvovirus H-1 J Virol 1998724149ndash56
22 Cziepluch C Lampel S Grewenig A et al H-1parvovirus-associated replication bodies a distinctvirus-induced nuclear structure J Virol 2000744807ndash15
23 Winnefeld M Grewenig A Schnolzer M et alHuman SGT interacts with Bag-6Bat-3Scytheand cells with reduced levels of either protein dis-play persistence of few misaligned chromosomesand mitotic arrest Exp Cell Res 20063122500ndash14
24 Schantl JA Roza M De Jong AP et al Smallglutamine-rich tetratricopeptide repeat-containingprotein (SGT) interacts with the ubiquitin-dependent endocytosis (UbE) motif of the growthhormone receptor Biochem J 2003373855ndash63
25 Moritz A Li Y Guo A et al Akt-RSK-S6 kinasesignaling networks activated by oncogenic recep-tor tyrosine kinases Sci Signal 20103ra64
26 Wu HC Hsieh JT Gleave ME et al Derivationof androgen-independent human LNCaP pros-tatic cancer cell sublines role of bone stromalcells Int J Cancer 199457406ndash12
Can
cerCellBiology
2822 Biological role of SGTA in prostate cancer
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC
27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC

27 Need EF Scher HI Peters AA et al A novelandrogen receptor amino terminal region revealstwo classes of aminocarboxyl interaction-deficient variants with divergent capacity to acti-vate responsive sites in chromatin Endocrinology20091502674ndash82
28 Smyth G Limma linear models for microarraydata New York Springer 2005
29 Subramanian A Tamayo P Mootha VK et alGene set enrichment analysis a knowledge-basedapproach for interpreting genome-wide expres-sion profiles Proc Natl Acad Sci USA 200510215545ndash50
30 Huang da W Sherman BT Lempicki RA Sys-tematic and integrative analysis of large gene listsusing DAVID bioinformatics resources Nat Pro-toc 2009444ndash57
31 Jackson AL Linsley PS Noise amidst the silenceoff-target effects of siRNAs Trends Genet 200420521ndash4
32 Jia L Berman BP Jariwala U et al Genomicandrogen receptor-occupied regions with differentfunctions defined by histone acetylation coregu-lators and transcriptional capacity PLoS One20083e3645
33 Jiang X Chen S Asara JM et al Phosphoinosi-tide 3-kinase pathway activation in phosphateand tensin homolog (PTEN)-deficient prostate
cancer cells is independent of receptor tyrosinekinases and mediated by the p110b and p110d
catalytic subunits J Biol Chem 201028514980ndash934 Vivanco I Sawyers CL The phosphatidylinositol
3-kinase AKT pathway in human cancer Nat Rev20022489ndash501
35 Li L Ittmann MM Ayala G et al The emergingrole of the PI3-K-Akt pathway in prostate cancerprogression Prostate Cancer Prostatic Dis 20058108ndash18
36 Taylor BS Schultz N Hieronymus H et al Inte-grative genomic profiling of human prostate can-cer Cancer Cell 20101811ndash22
37 Sarker D Reid AH Yap TA et al Targeting thePI3KAKT pathway for the treatment of prostatecancer Clin Cancer Res 2009154799ndash805
38 Scher HI Sawyers CL Biology of progressivecastration-resistant prostate cancer directedtherapies targeting the androgen-receptor signal-ing axis J Clin Oncol 2005238253ndash61
39 Prescott J Coetzee GA Molecular chaperonesthroughout the life cycle of the androgen recep-tor Cancer Lett 200623112ndash19
40 Shen MM Abate-Shen C Pten inactivation andthe emergence of androgen-independent prostatecancer Cancer Res 2007676535ndash8
41 Ha S Ruoff R Kahoud N et al Androgenreceptor levels are upregulated by Akt in pros-tate cancer Endocr Relat Cancer 201118245ndash55
42 Zhu Q Youn H Tang J et al Phosphoinositide3-OH kinase p85a and p110b are essential forandrogen receptor transactivation and tumor pro-gression in prostate cancers Oncogene 2008274569ndash79
43 Liu FH Wu SJ Hu SM et al Specific interactionof the 70-kDa heat shock cognate protein withthe tetratricopeptide repeats J Biol Chem 199927434425ndash32
44 Periyasamy S Hinds T Jr Shemshedini L et alFKBP51 and Cyp40 are positive regulators ofandrogen-dependent prostate cancer cell growthand the targets of FK506 and cyclosporin AOncogene 2010291691ndash701
45 Periyasamy S Warrier M Tillekeratne MPet al The immunophilin ligands cyclosporin Aand FK506 suppress prostate cancer cell growthby androgen receptor-dependent and -independ-ent mechanisms Endocrinology 20071484716ndash26
46 Horibe T Kohno M Haramoto M et alDesigned hybrid TPR peptide targeting Hsp90as a novel anticancer agent J Transl Med 201198
Can
cerCellBiology
Trotta et al 2823
Int J Cancer 133 2812ndash2823 (2013) VC 2013 UICC