isolation and characterization of melanized fungi from limestone formations in mallorca
TRANSCRIPT

ock surfaces can be considered as extreme environ-ments that are exposed to high solar radiation, ele-vated temperatures, low nutrient availability, high
electrolyte concentration, and low relative humidity, wheremoisture may be only sporadically available. Rock surfacesare colonized by unique species of stress-tolerant fungi.Among these stress-tolerant fungi are black, yeast-like and de-matiaceous mycelial fungi that have been isolated from va-rious rock surfaces, stone monuments, and building surfaces(GORBUSHINA et al. 1993; WOLLENZIEN et al. 1995; STERFLIN-GER & KRUMBEIN 1997; STERFLINGER & PRILLINGER 2001; BO-GOMOLOVA & MINTER 2003; DE LEO, URZÍ & DE HOOG 2003).These fungi are a phylogenetically diverse assemblage of non-lichenized ascomycetes and often share high melanin pro-duction, resistance to high temperatures and UV radiation, andrestricted, meristematic growth morphology. The latter traithas led to the name of “microcolonial fungi” (STALEY, PAL-MER & ADAMS 1982) or “meristematic fungi” for these orga-nisms. Some of these fungi, including some newly described
species from the Mediterranean basin, have been implicatedin the staining, pitting and biodeterioration of marble and lime-stone monuments (STERFLINGER et al. 1997; WOLLENZIEN etal. 1997; DE LEO, URZÍ & DE HOOG 1999).
For more than a decade, our laboratory has explored prac-tical methods of providing large numbers of unique fungal iso-lates for use in natural products discovery. The laboratory hasmeasured and modeled the biodiversity of several kinds offungal communities in order to understand and improve thelogistics and processes in handling large numbers of fungalstrains. The fungi associated with rock surfaces are potentiallyan attractive group of organisms for a microbial screening pro-gram. Such fungi are apparently ubiquitous and relatively easyto collect and isolate into culture. At least some species are re-lated to other ascomycetous fungi that are known to possessmultiple secondary metabolite biosynthetic pathways (KRO-KEN et al. 2003). The melanized cell walls of these fungi in-dicate that at least melanin polyketide biosynthetic pathwaysare operational.
However, questions remain unanswered about the feasi-bility of using these fungi in an industrial microbial screeningprogram. In relatively few cases have the fungal communitieson rock surfaces been inventoried exhaustively, and the ex-tent of species diversity at a local scale is largely unknown.
Isolation and characterization of melanized fungi from limestone
formations in Mallorca
Constantino RUIBAL1, Gonzalo PLATAS1, and Gerald F. BILLS1,*
Melanized fungi were isolated from limestone surfaces in upland and coastal environments in the Mediterranean island ofMallorca. One hundred seventeen isolates were recovered from two topographically distinct sites. Due to the difficulty indistinguishing among isolates based on morphological criteria, microsatellite-primed PCR techniques were used to groupisolates into genotypes that were assumed to represent species. Seventeen genotypes were characterized from one site andtwenty six from the other, with four genotypes common to both. Classical and molecular methods were used to identifyrepresentative strains. Morphological methods rarely provided a reliable identification; only three isolates, Hortaea werneckii,Trimmatostroma abietis and Aureobasidium pullulans were identified with certainty, and the identification was confirmedby molecular data. Morphological characters that were widespread among the isolates included scarce micronematous co-nidial states or non-sporulating sterile mycelia, mature mycelia with dark olive green or black hyphae, mycelia with toruloushyphae, whose cells developed one or more transverse septa. In many of these fungi, the cells of mature hyphae disarticu-lated, suggesting asexual reproduction by a thallic micronematous conidiogenesis or by simple propagative fragmentation.Sequencing of the internal transcribed spacers (ITS1, ITS2) and 5.8S ribosomal gene, as well as the 18S rDNA ribosomalgene were employed to investigate the isolates’ phylogenetic affinities. The majority of the isolates could be grouped in twomain classes of Ascomycetes, Dothideomycetes and Chaetothyriomycetes, although many others did not correspond withany sequence deposited in public databases, suggesting they could be of unknown genera that did not correspond with anywell-defined Ascomycete order.
Keywords: Black fungi, Chaetothyriales, diversity, Dothideales, extremophile, ITS sequence, 18S sequence, rock
Mycological Progress 4(1): 23–38, February 2005 23
1 Centro de Investigación Básica, Merck, Sharp & Dohme de España,
S.A., Josefa Valcárcel 38, E-28027, Madrid, Spain* Corresponding author: Tel.: 34913210504; Fax: 34913210614; E-
mail: [email protected]
© DGfM 2005
R

24 RUIBAL, PLATAS & BILLS: Melanized fungi from Mallorcan limestone
© DGfM 2005
Still relatively few species have been described and characte-rized phylogenetically. Most of these studies have focused thesystematics and autoecology of species in manmade surfaces;much less is known about fungi from natural rock formations.
For a first consideration of these basic questions, we madea preliminary assessment of the numbers and kinds of mela-nized fungi associated with the surfaces of limestone forma-tions in Mallorca. Limestone of this island was chosen becauseseveral reports have described novel taxa from rock and mo-nument surfaces in the Mediterranean region. We describe thenumbers of different fungi that we were able to recover fromtwo different limestone formations, comment on their culturalfeatures, and present a preliminary evaluation of their phylo-genetic diversity based on rDNA ITS1-5.8S-ITS2 and 5´endof 18S gene sequences.
Material and methods
Study area, environmental and geological features. Thestudy sites, Manut II and Cala Sant Vicenç, are limestone out-crops sculpted by karstic solutional features, called karren(GINÉS & GINÉS 1995; GINÉS 1998). They are located in thenorthernmost sector of the main Mallorcan mountain range,the Serra de Tramuntana. This part of the island is mainly builtof Mesozoic carbonate rocks that were folded and overthrust-ed as a result of Alpine tectonic movements. The sampledrocks are very pure calcitic limestone. From a textural pointof view, the limestone matrix and the clasts where the fungiwere sampled are hard and fine grained limestone. They arenearly formed exclusively of calcium carbonate and have nosignificant amounts of magnesium carbonate. Limestone fromthe Manut II site is Jurassic in age, but the age of the lime-stone of Cala Sant Vicenç is unknown. The Manut II site waslocated along Highway C-710, between Lluc and Pollença,about 500 m south of the Manut II recreation area, in a northernand shady exposure at approximately 500 m above sea level.The area receives about 1000 mm rainfall distributed evenlyduring the year; 2) Cala Sant Vicenç site is on the southfacingcliffs of the cove, a few meters above the sea level near thesea’s edge. The site is sunny and semiarid, affected by seaspray, and receives slightly less than 800 mm of rainfall a year.Both outcrops show evidences at the meso- and microscale oftheir local environmental factors: for instance, the dominantkarren forms in Manut II are characterized by conspicuousfurrowing related to the interception and channelizing of rainsand storms; furrows are rare in Cala Sant Vicenç, being sub-stituted by irregular etching and pitting as well as by the do-minance of microrills.
Isolation of melanized rock-inhabiting fungi. Limestonesamples were taken from clean, lichen-free, vertical surfacesat the end of the dry season in November, 2001. At each site,2–5 rock faces separated by 20 to 100 m and in different ori-entations were sampled. A cold chisel was inserted into anexisting crack and the pieces of superficial rock were lifted
off the surface with a blow from a hammer. Limestone pieceswere packed in new clean paper bags and transported to thelab at ambient temperature. Samples were processed for iso-lation within one week.
In the lab, three different points from Cala Sant Vicençand two points from Manut II were processed. The rock sur-face was disinfected with 96 % ethanol to reduce the influ-ence of dust and airborne spores. Samples for isolation plateswere prepared by chipping or scraping with a sterile knife andchisel blade to remove small superficial fragments from 10 to20 cm2 area from the outer layer of the field-collected rockand penetrating up to 2 to 3 mm deep into the surface. Thefragments were further pulverized in a sterile mortar. About2–3 cm3 of pulverized rock were added to a sterile centrifugetube and washed once with a weak detergent solution (aqueousTween 20, Sigma-Aldrich, Inc., St. Louis, MO, USA), follow-ed by three washes with distilled water, and two final washeswith sterile water. Washed samples were resuspended in 40 mlof sterile water and aliquots were spread homogenously ontoisolation media plates. For each rock sample, aliquots of 500,300, 200, 100 and 50 µl, were spread per triplicate to one plateeach of dichloran-rose bengal agar base (DRBC) (Oxoid Ltd.,Basingstoke, Hampshire, UK); and dichloran-glycerol agarbase (DG18) (Oxoid Ltd.). Media were supplemented withstreptomycin sulfate and terramycin (50 µg/ml each) to eli-minate bacteria.
Plates were incubated for 2–3 weeks with 12 hr fluores-cent light at 22 ºC. Within a few days of the first appearanceof colonies on the plates, all colonies that had dark hyphaewere removed, while fast growing hyaline mycelia were eli-minated. Continual removal of rapidly extending colonies per-mitted more time and space for development of slower grow-ing pigmented colonies. Plates were examined for about amonth until no more new colonies appeared.
Strain characterization. The darkly pigmented isolates ob-tained from isolation plates were grown in duplicate on po-tato dextrose agar (PDA) (Difco Laboratories, Detroit, MI,USA) plates at 22 ºC for 2 weeks. We attempted to group iso-lates into gross morphological types by colony color, texture,margin type, and radial extension, but it was recognized thatthe crude grouping was inadequate to discriminate numbersand species. To recognize numbers of unique genotypes andwhich isolates were genotypically identical, we used a micro-satellite-primed PCR (MP-PCR) fingerprinting technique(LONGATO & BONFANTE 1997). Band patterns in agarose gelswere used to group isolates. The groupings based on MP-PCRwere corroborated by the macroscopic morphology of isolatesin PDA plates. Once isolates were grouped into genotypes, atleast one representative member of each group was processedand analyzed. Our underlying assumption was that this isolatewas representative of a single species and that the rest of iso-lates of the group were the same species. Although we reco-gnized that minor differences in ITS1-5.8-ITS2 rDNA se-quences by themselves may not effectively delimit species
Mycological Progress 4(1) / 2005 25
© DGfM 2005
(TAYLOR et al. 2000), we relied on these sequence to opera-tionally classify strains into taxonomic units. At least one, andsometimes more than one isolate of each group were preserv-ed when morphological deviants were observed, or if multiplestrains were sequenced. Isolates (Tab. 1) are maintained asfrozen mycelia in 10 % glycerol at –80 ºC at Merck Sharp &Dohme de España, S.A.
The representative isolates were cultured in triplicate in fivedifferent media to study their macro- and microscopic charac-ters. The set of media used for characterization was based ontheir previous efficiency in the induction of sexual or asexualstates, and included: potato carrot agar (PCA) (GAMS et al.1987); malt extract agar (MEA) (Difco Laboratories), Czapek-Dox agar (CZA) (Difco Laboratories); oatmeal agar (OA) (Dif-co Laboratories); water agar supplemented with 0.2 % yeastextract (AI) (Difco Laboratories). Colony diameter, texture,pigmentation, margin appearance, exudates, and colors in thedescriptions were recorded after 2 weeks at 22 ºC.
Microscopic features were evaluated by 1) observing hy-phae and other structures mounted in 5 % KOH; 2) growingfungal colonies on cover glasses immersed in corn meal agar(Difco Laboratories) and malt-agar (0.5 % malt extract, 1.5 %agar) and supported on 2 % water agar. Three-week-old co-lonies on cover glasses were fixed in 95 % ethanol, mountedin cotton blue-lactophenol, and photographed.
DNA extraction, PCR amplification, and DNA sequen-
cing. DNA extraction was performed following previouslydescribed methods (PELAEZ et al. 1996). A strategy that com-bined microsatellite-primed PCR (MP-PCR) and macromor-phology was used to group and recognize similar isolates. TheMP-PCR was performed using (GTG)5 primer (LONGATO &BONFANTE 1997). Reactions volumes (25 µl) were carried outin the following conditions: 0.2 mM of each dNTPs (gene-Amp dNTPs, Perkin Elmer, Norwalk, CT, USA), primer at afinal concentration of 0.4 µM, 5 µl of fungal DNA solution(1/100 dilution of the extraction solution with a concentrationbetween 0.1 and 0.01 µg/ml) and 0.2 U. of Taq polymerase(QBiogene, Inc., Carlsbad, CA, USA) in a reaction buffer. Thesteps of the PCR reactions were: 40 cycles of 30 s at 95 ºC,30 s at 53 ºC and 2 min at 72 ºC and the process was complet-ed with a final step of 10 min at 72 ºC for the polymerization
of incomplete fragments. Reactions were performed with TaqDNA polymerase (QBiogene, Inc.) following manufacturer’srecommendations. The amplification products were checkedby electrophoresis in agarose gels 1.2 % (Hispanlab, S.A.,Alcobendas, Madrid, Spain) in a Pharmacia LKB-GPS 200electrophoresis apparatus at 120 V (Amersham BiosciencesLtd., Buckinghamshire, UK).
To analyze and compare the band patterns from MP-PCR,fragments were separated electrophoretically in prefabricatedpolyacrylamide (12.5 %) gels (GeneCel Excel 12.5/24 Kit,Amersham Biosciences Ltd.) in a GenePhor unit (AmershamBiosciences Ltd.) at 600V and visualized by silver staining(PLUS ONE DNA Silver Staining Kit, 17-6000-36, AmershamBiosciences Ltd.) following the manufacturer’s procedures.Gel band patterns were scanned and used to produce pheneticgrouping with Gel Compar 4.1 software package (AppliedBiomaths, Kortajk, BE). The combined data of all fingerprintsfor each isolate were compared in a pairwise manner usingJaccard’s index (SNEATH & SOKAL 1997) to generate a matrixof similarities. A dendrogram showing the relative similarityamong the different isolates was produced by UPGMA(SNEATH & SOKAL 1997).
Amplification of the internal transcribed spacers (ITS1 andITS2) and the 5.8S ribosomal gene of the isolates were per-formed with the following primers for the ITS1-5.8S-ITS2fragment: ITS1F (GARDES & BURNS 1993), ITS4A (LARENA
et al. 1999), ITS5 (WHITE et al. 1990), NL1R (reverse of NL1)(O`DONNELL 1993) and 18S3: 5´-GAT GCC CTT AGA TGTTCT GGG G-3´ (designed in the laboratory). For the rDNAregion containing the initial sequence of 18S gene, amplifi-cations were performed using primers NS1 and NS2 (WHITE
et al. 1990). The specific primers applied for amplification ofeach isolate are listed in Table 2.
PCR reactions were performed following standard proce-dures (5 minutes at 93 ºC followed by 40 cycles of 30 s at93 ºC, 30 s at 53 ºC and 2 min at 72 ºC) with Taq DNA poly-merase (QBiogene, Inc.) following the procedures recommend-ed by the manufacturer. The amplification products (0.10 µg/ml)were sequenced using the Bigdye Terminators version 1.1 (Per-kin Elmer) following the manufacturer’s recommendations.Each strand of all the amplification products was sequencedwith the same primers used for the initial amplification.
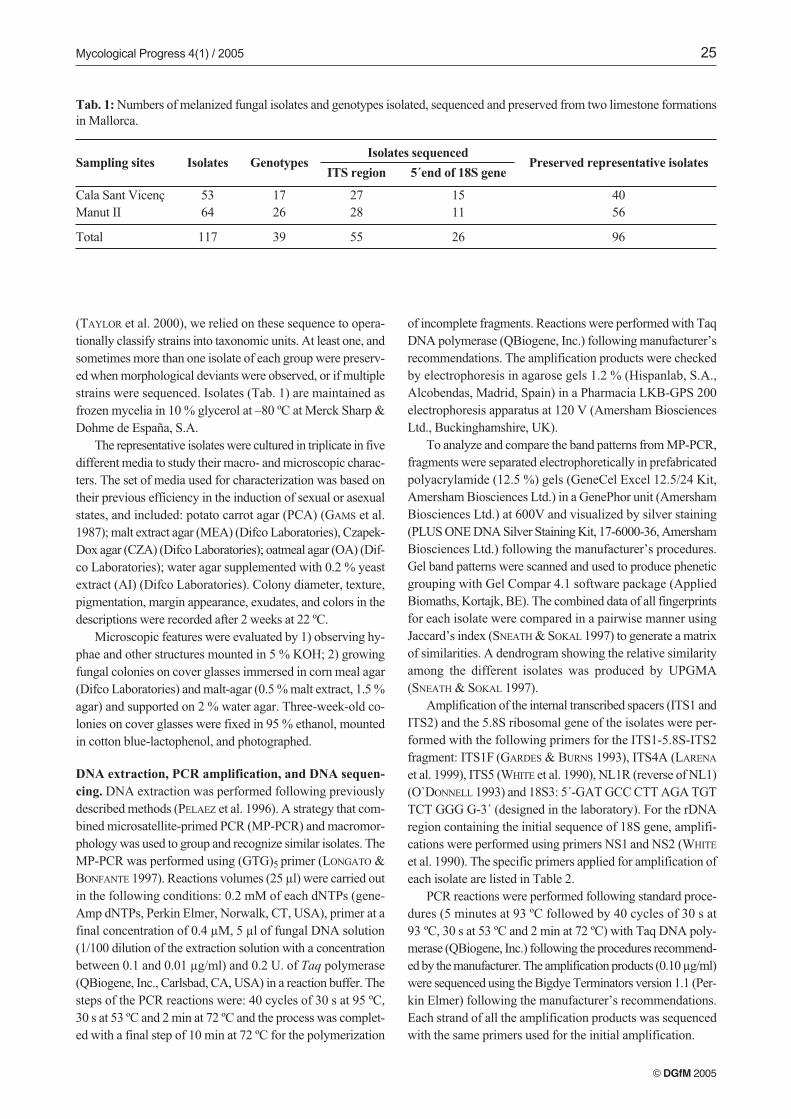
Tab. 1: Numbers of melanized fungal isolates and genotypes isolated, sequenced and preserved from two limestone formationsin Mallorca.
Preserved representative isolatesIsolates sequenced
ITS region 5´end of 18S geneSampling sites Isolates Genotypes
Cala Sant Vicenç 53 17Manut II 64 26
Total 117 39
4056
96
27 1528 11
55 26
26 RUIBAL, PLATAS & BILLS: Melanized fungi from Mallorcan limestone
© DGfM 2005
Sequence data. The following sequences were obtained fromGenBank for the ITS sequence analyses: Rhizoctonia sp. Eab-S4 (AJ242884), Phaeoramularia hachijoensis Matshus.(AY251068), Mycocalicium victoriae (C. Knight ex F. Wil-son) Tibell (AJ312123), Trimmatostroma abietis Butin & Pehl(AJ244249), Hortaea werneckii (Horta) Nishimura & Miyaji(AJ244249), Trimmatostroma sp. (AJ244268), Capnodium
coffeae Patouillard (AJ244239), Coniosporium apollinis Sterf-linger et al. (AJ244272), Aureobasidium pullulans (de Bary)G. Arnaud (AF013229), Sarcinomyces crustaceus Lindner(AJ244258), Sarcinomyces petricola U. Wollenzien & deHoog (AJ244275), Phaeococcomyces catenatus (de Hoog &Hermanides-Nijhof) de Hoog (AF050277), Exophiala salmo-
nis Carmichael (AF050274), Sarcinomyces phaeomuriformis
Matsumoto et al. (AJ244259), Dactylaria sp. P7 (AY265338).For the 18S gene analyses, the the following sequences
were obtained: Rhizoctonia solani J.G. Kühn AG-1 CS-Gi(E17097), Hortaea werneckii (Horta) Nishimura & Miyaji (ex-neotype) (Y18693), Microxyphium citri (Berk.& Desm.) Ba-tista & Ciferri (= Capnodium citri Berk. & Desm.)(AY016340), Capnodium dematum (V.A.M. Mill. & Bonar)D.R. Reynolds (AF006724), Aureobasidium pullulans
(AY030322), Dothidea hippophaëos (Pass.) Fuckel (U42475),Dothidea insculpta Wallr. (U42474), Dothidea ribesia Fr.(AY016343), Cephaliophora muscicola G.L. Barron, C. Mo-rik. & Saikawa (AB001108), Coniosporium sp. (Y11712), Cla-
dophialophora modesta McGinnis et al. (= Cladophialophora
sp. CBS 985.96) (AJ232953), Exophiala dermatitidis (Kano)de Hoog (AF397191), Exophiala sp. (AJ301704), Exophiala
nigra (Issatsch.) Haase & de Hoog (type strain) (X91896),Cladophialophora boppii (Borelli) de Hoog et al. (AJ232946),Capronia dactylotricha Untereiner, Candoussau & Samuels(AJ232943), Capronia pilosella (Karsten) E. Müller et al.(AJ232940), Sarcinomyces petricola U. Wollenzien & deHoog (Y18702), Cucurbidothis pityophila Petrak (U42480),Cucurbitaria berberidis (Pers.) S.F. Gray (U42481), Herpo-
trichia diffusa (Schwein.) Ellis & Everhart (U42484), Cocco-
dinium bartschii A. Massal. (U77668), Phaeoramularia ha-
chijoensis (AY251100), Capnobotryella sp. (AJ301706),Capnobotryella renispora J. Sugiyama (AY220614).
Sequence analyses. Partial sequences obtained in sequencingreactions were assembled using GCG software, generating aconsensus sequence for each fungal strain. Consensus sequen-ces were aligned by CLUSTAL W (THOMPSON et al. 1997)and the program-generated multiple alignments were visuallyadjusted with GeneDoc 2.5 software (NICHOLAS & NICHOLAS
1997). Maximum-parsimony analyses were performed withPAUP 4 (SWOFFORD 1998). Data were resampled with 1000bootstrap replicates (FELSENSTEIN 1985) by using the heuristicsearch option of PAUP. The percentage of bootstrap replica-tes that yielded greater than 50% for each group was used asa measure of statistical confidence. Sequence matching withpublic or proprietary databases was performed with BLAST2N and FastA applications (GCG WISCONSIN PACKAGE,Version 10.3-UNIX, Accelrys Inc., San Diego, CA, USA).
Results
Numbers of isolates and efficiency of the isolation method.
Fifty three isolates were obtained from Cala Sant Vicenç, re-presenting 13 genotypes exclusive to the site. From Manut II,64 isolates were recovered, representing 22 genotypes exclu-sive to the site. Four additional genotypes occurred at both si-tes (Tab. 1). The representative isolates chosen for sequencingare listed in Tab. 2.
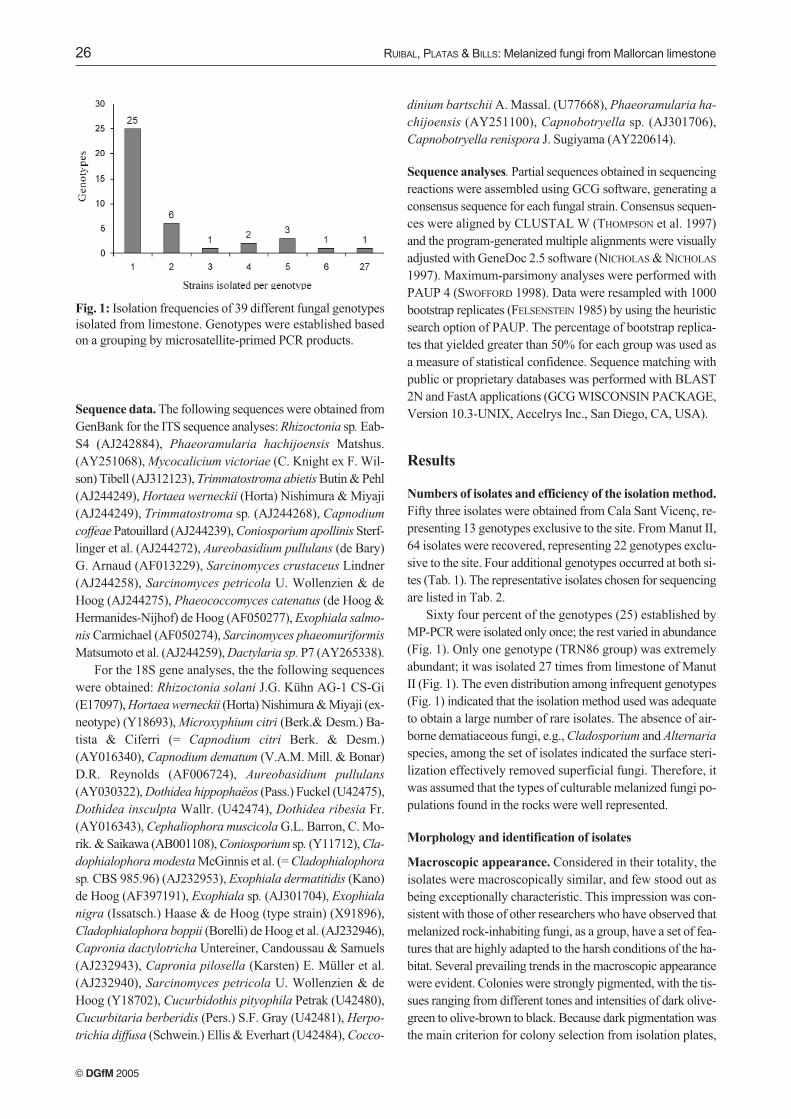
Sixty four percent of the genotypes (25) established byMP-PCR were isolated only once; the rest varied in abundance(Fig. 1). Only one genotype (TRN86 group) was extremelyabundant; it was isolated 27 times from limestone of ManutII (Fig. 1). The even distribution among infrequent genotypes(Fig. 1) indicated that the isolation method used was adequateto obtain a large number of rare isolates. The absence of air-borne dematiaceous fungi, e.g., Cladosporium and Alternaria
species, among the set of isolates indicated the surface steri-lization effectively removed superficial fungi. Therefore, itwas assumed that the types of culturable melanized fungi po-pulations found in the rocks were well represented.
Morphology and identification of isolates
Macroscopic appearance. Considered in their totality, theisolates were macroscopically similar, and few stood out asbeing exceptionally characteristic. This impression was con-sistent with those of other researchers who have observed thatmelanized rock-inhabiting fungi, as a group, have a set of fea-tures that are highly adapted to the harsh conditions of the ha-bitat. Several prevailing trends in the macroscopic appearancewere evident. Colonies were strongly pigmented, with the tis-sues ranging from different tones and intensities of dark olive-green to olive-brown to black. Because dark pigmentation wasthe main criterion for colony selection from isolation plates,
Fig. 1: Isolation frequencies of 39 different fungal genotypesisolated from limestone. Genotypes were established basedon a grouping by microsatellite-primed PCR products.
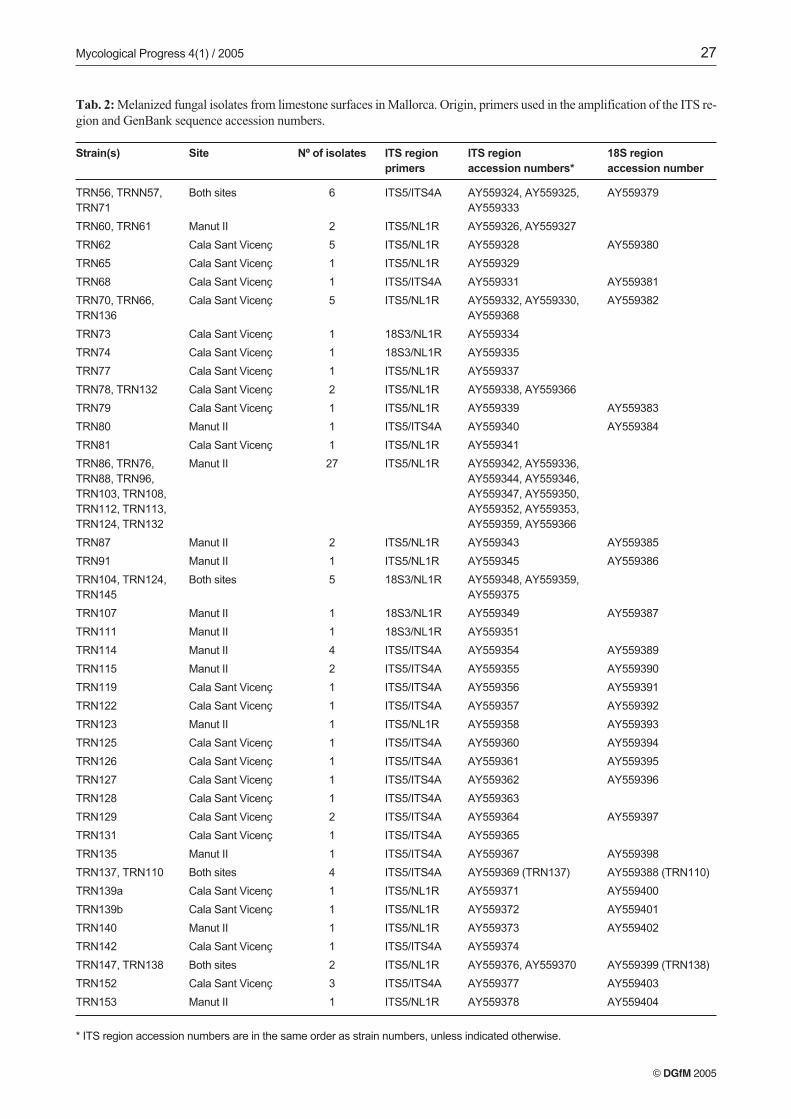
Strain(s) Site Nº of isolates ITS region ITS region 18S region
primers accession numbers* accession number
TRN56, TRNN57, Both sites 6 ITS5/ITS4A AY559324, AY559325, AY559379
TRN71 AY559333
TRN60, TRN61 Manut II 2 ITS5/NL1R AY559326, AY559327
TRN62 Cala Sant Vicenç 5 ITS5/NL1R AY559328 AY559380
TRN65 Cala Sant Vicenç 1 ITS5/NL1R AY559329
TRN68 Cala Sant Vicenç 1 ITS5/ITS4A AY559331 AY559381
TRN70, TRN66, Cala Sant Vicenç 5 ITS5/NL1R AY559332, AY559330, AY559382
TRN136 AY559368
TRN73 Cala Sant Vicenç 1 18S3/NL1R AY559334
TRN74 Cala Sant Vicenç 1 18S3/NL1R AY559335
TRN77 Cala Sant Vicenç 1 ITS5/NL1R AY559337
TRN78, TRN132 Cala Sant Vicenç 2 ITS5/NL1R AY559338, AY559366
TRN79 Cala Sant Vicenç 1 ITS5/NL1R AY559339 AY559383
TRN80 Manut II 1 ITS5/ITS4A AY559340 AY559384
TRN81 Cala Sant Vicenç 1 ITS5/NL1R AY559341
TRN86, TRN76, Manut II 27 ITS5/NL1R AY559342, AY559336,
TRN88, TRN96, AY559344, AY559346,
TRN103, TRN108, AY559347, AY559350,
TRN112, TRN113, AY559352, AY559353,
TRN124, TRN132 AY559359, AY559366
TRN87 Manut II 2 ITS5/NL1R AY559343 AY559385
TRN91 Manut II 1 ITS5/NL1R AY559345 AY559386
TRN104, TRN124, Both sites 5 18S3/NL1R AY559348, AY559359,
TRN145 AY559375
TRN107 Manut II 1 18S3/NL1R AY559349 AY559387
TRN111 Manut II 1 18S3/NL1R AY559351
TRN114 Manut II 4 ITS5/ITS4A AY559354 AY559389
TRN115 Manut II 2 ITS5/ITS4A AY559355 AY559390
TRN119 Cala Sant Vicenç 1 ITS5/ITS4A AY559356 AY559391
TRN122 Cala Sant Vicenç 1 ITS5/ITS4A AY559357 AY559392
TRN123 Manut II 1 ITS5/NL1R AY559358 AY559393
TRN125 Cala Sant Vicenç 1 ITS5/ITS4A AY559360 AY559394
TRN126 Cala Sant Vicenç 1 ITS5/ITS4A AY559361 AY559395
TRN127 Cala Sant Vicenç 1 ITS5/ITS4A AY559362 AY559396
TRN128 Cala Sant Vicenç 1 ITS5/ITS4A AY559363
TRN129 Cala Sant Vicenç 2 ITS5/ITS4A AY559364 AY559397
TRN131 Cala Sant Vicenç 1 ITS5/ITS4A AY559365
TRN135 Manut II 1 ITS5/ITS4A AY559367 AY559398
TRN137, TRN110 Both sites 4 ITS5/ITS4A AY559369 (TRN137) AY559388 (TRN110)
TRN139a Cala Sant Vicenç 1 ITS5/NL1R AY559371 AY559400
TRN139b Cala Sant Vicenç 1 ITS5/NL1R AY559372 AY559401
TRN140 Manut II 1 ITS5/NL1R AY559373 AY559402
TRN142 Cala Sant Vicenç 1 ITS5/ITS4A AY559374
TRN147, TRN138 Both sites 2 ITS5/NL1R AY559376, AY559370 AY559399 (TRN138)
TRN152 Cala Sant Vicenç 3 ITS5/ITS4A AY559377 AY559403
TRN153 Manut II 1 ITS5/NL1R AY559378 AY559404
* ITS region accession numbers are in the same order as strain numbers, unless indicated otherwise.
Tab. 2: Melanized fungal isolates from limestone surfaces in Mallorca. Origin, primers used in the amplification of the ITS re-gion and GenBank sequence accession numbers.
© DGfM 2005
Mycological Progress 4(1) / 2005 27

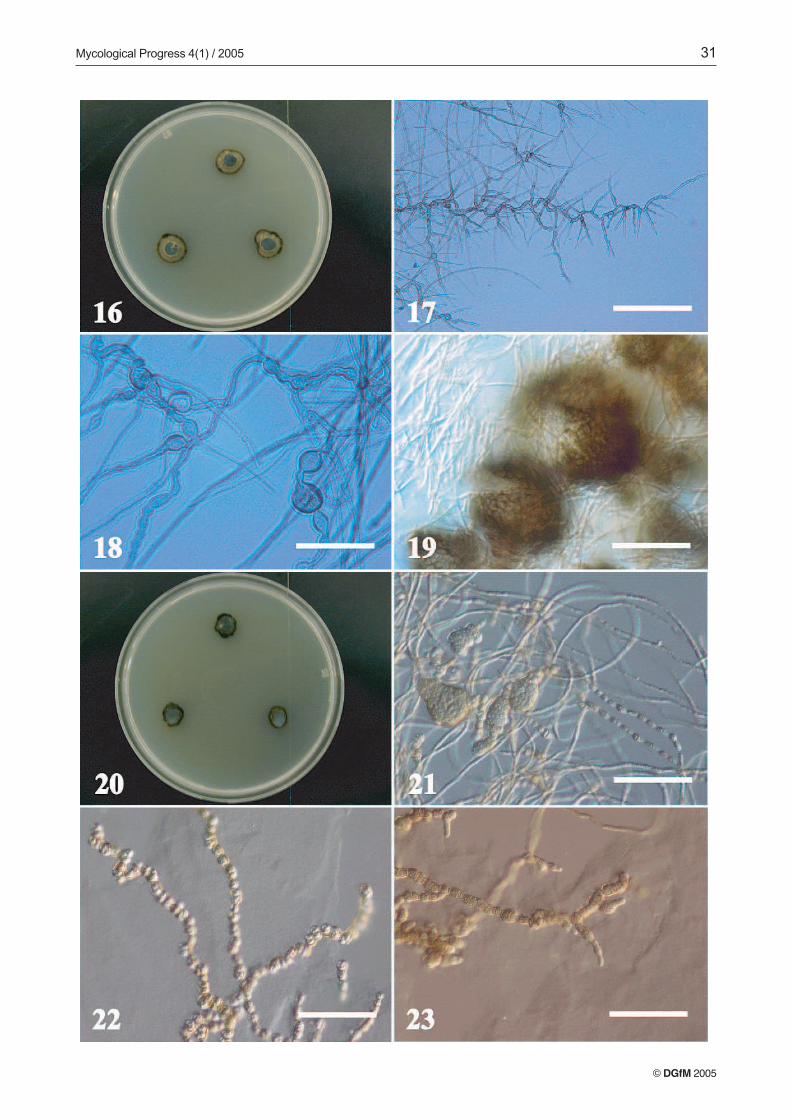
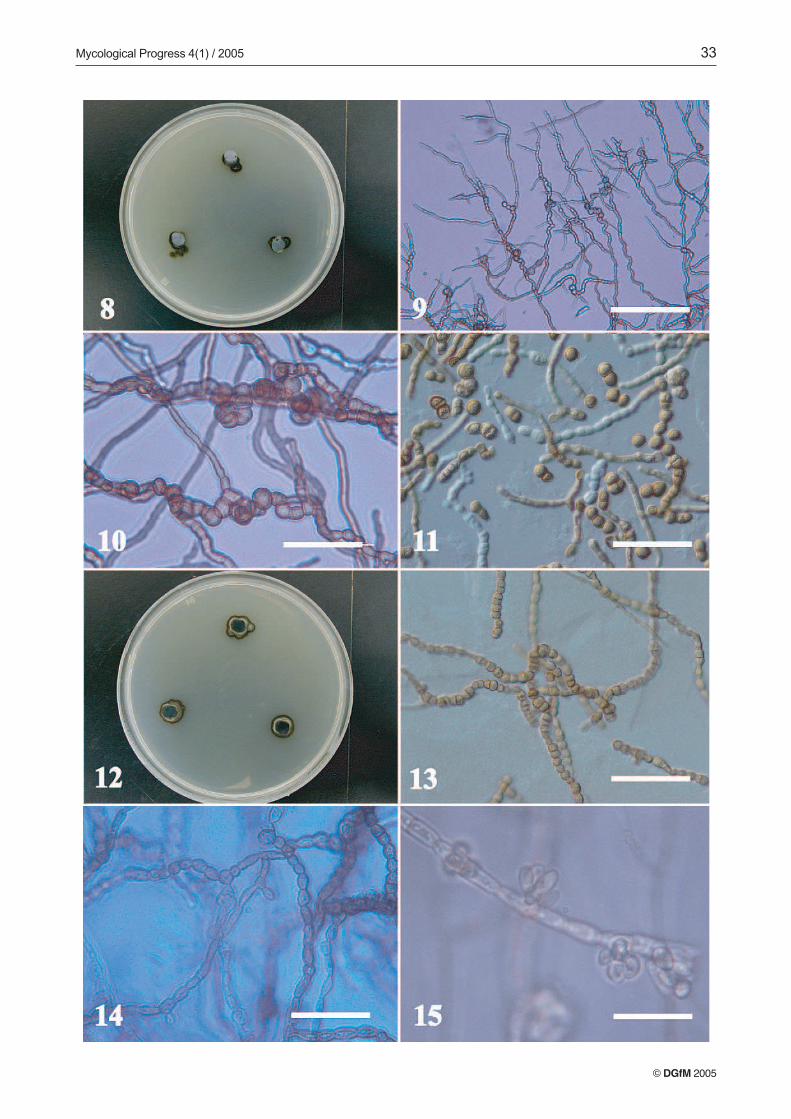
it was not helpful for discrimination among the isolates. Theisolates, for the most part, exhibited slow radial growth. Themean diameter of two-week-old colonies of all isolates (n =96) on the five different media was 10.5 mm, with maximumcolony diameter of 36 mm. The mycelia of a considerableproportion of isolates scarcely exhibited measurable radialgrowth (e.g., Figs. 2, 8 and 20). Compact and dense colonieswith a dry, granulose-crustose colony texture were evident ina high percentage of the isolates (Figs. 2, 8, 12, 16 and 20).Filamentous colony types were also quite common. Blackyeast-like colonies were nearly absent among our isolates, al-though some meristematic fungi can present yeast-like growthpatterns in culture (DE HOOG et al. 1999; STERFLINGER, DE
HOOG & HAASE 1999). Absence of yeast-like forms could bedue to the isolation and/or culture conditions. Mycelial exu-dates or soluble pigments in the agar were generally absentamong the isolates.
Microscopic appearance. The identification of the isolateswas problematic. Due to their often scant mycelium and hard,granulose texture and strong pigmentation, the preparation ofisolates for their microscopic study was difficult. In most cases,only the vegetative mycelia and their characteristics could bedescribed. With some exceptions, the isolates were poorly dif-ferentiated, with few specialized structures and tissues, e.g.,sexual or asexual sporulating states. Common features observ-ed were: hyphae with densely melanized, thick-walled cells andtorulous hyphae that developed transverse septa or muriformcells. As a result, the strong convergence and intergradationsamong vegetative features prevented confident grouping ofisolates into morphological types.
Ninety-six isolates were examined microscopically; 84 ofthem did not exhibit any conidial or sexual state on the mediaset, and were considered sterile mycelia. In 13 isolates, an ob-vious or suspected conidial state was observed (isolatesTRN122, TRN127, TRN137, TRN138, TRN56, TRN63,TRN64, TRN87, TRN126, TRN128, TRN129, TRN135 andTRN81). Only three isolates were identified unequivocally:TRN122 as Hortaea werneckii, TRN127 as Trimmatostroma
abietis and TRN135 as Aureobasidium pullulans. In the fol-lowing section, we describe some selected isolates to illustratethe range of organisms that we recovered. Isolate TRN125formed stipitate pycnidial structures, and we tentatively iden-tified it as the capnodiaceous anamorph genus Leptoxyphium.
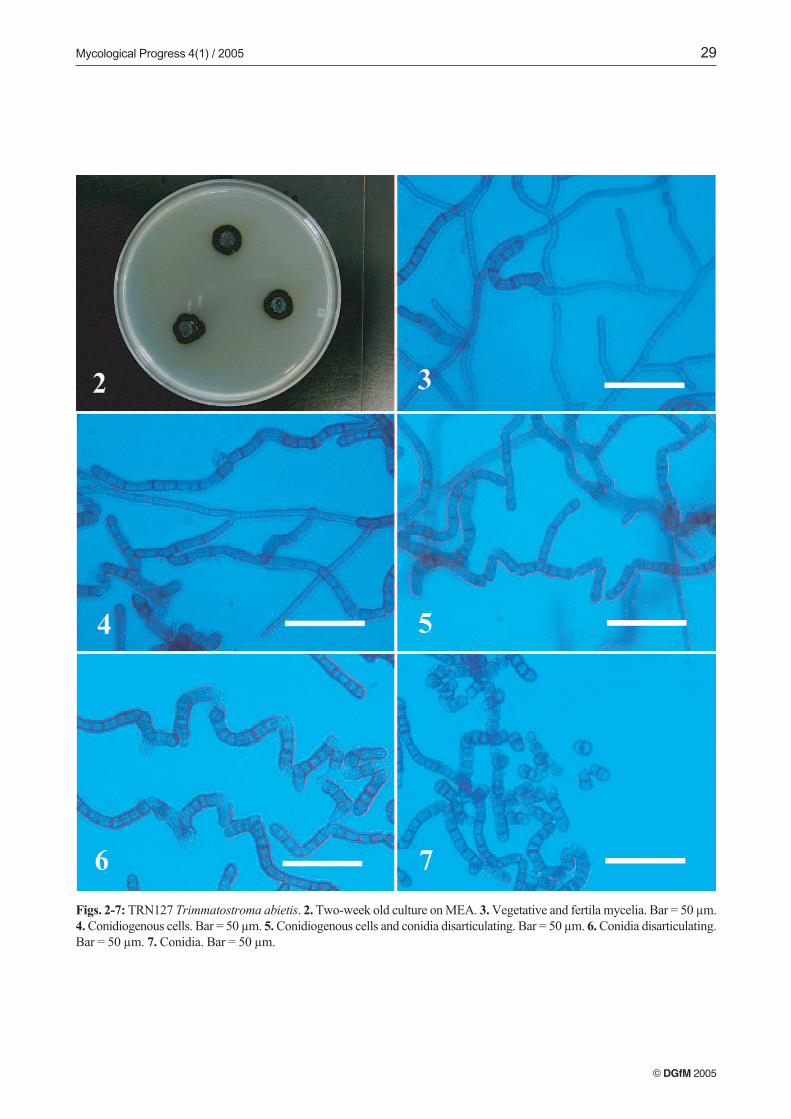
Description and comments on selected isolates. Of the iden-tified isolates we only describe TRN127 (Trimmatostroma
abietis) on the basis of the interest of its controversial natu-re and undefined taxonomic positioning.
TRN127 (Trimmatostroma abietis), (Figs. 2–7). Colonieson MEA attaining a diameter of 13 mm, with margin sub-merged, irregular, dark olive, with little aerial mycelia, sub-felty, olive grey, and agar raised at the center. Reverse darkolive to black. Colonies on PCA attaining a diameter of 15 mm,margin submerged, almost even, dark olive to black, with
aerial mycelia crustose, dark olive to black. Reverse black.Colonies on OTM attaining a diameter of 14 mm. Margin ap-pressed, submerged at the very edge, irregular, shiny blackwith aerial mycelia crustose, dark olive to black. Reverse verydark green. Colonies on CZA attaining a diameter of 7 mm,with margin raised, fringed. Closely zonate, with submerged,even, dark olive zones alternating with zones of crustose, olivegreen aerial mycelia. Reverse black to dark green. Colonieson AI attaining a diameter of 9 mm, with margin submerged,fimbriate, brownish olive with scarce aerial mycelia, crustose.Reverse dark brownish grey.
Vegetative hyphae pale green, smooth, thin-walled, sep-tate, with nearly rectangular cells, 3–5 µm wide; regularlybranched, with branch angles near 45º for vegetative hyphae,and 90º for fertile hyphae (Fig. 3). Fertile hyphae graduallydeveloping from undifferentiated, vegetative hyphae, arisinglaterally and becoming rough- and thick-walled, with heavilymelanized, thickened cells (7–9 µm wide) (Figs. 3, 4). Thallicconidia originate by intercalary dilatation of fertile hyphae.Enlarged and darkened-septa hyphal fragments formation ob-served along fertile hyphae. Hyaline cells grow terminallyfrom these fragments (Figs. 5, 6). Conidia ovoid to cylindrical,1–5 septate, olivaceous brown to black, thick- and rough-wall-ed, forming rounded, smooth, pale terminal cells after dehis-cence. Conidia may remain joined by remnants of their formermelanized walls, forming folded conidial chains (Figs. 6, 7).
Most of the isolates developed only vegetative myceliumin culture, however a few formed unrecognizable conidialstates on some media. TRN138 and TRN137 are described asexamples of two unrecognizable isolates we encountered.Other isolates developed what appeared to be immature pyc-nidial conidiomata, such as TRN152, or strongly differentiatedtorulose to moniliform hyphal patterns that may correspondto vegetative reproduction of the mycelia, such as in TRN153.
TRN138 (Figs. 8–11). Colonies on MEA attaining a dia-meter of 8 mm, with margin submerged to appressed, fringed,shiny, and even, pale olive to dark olive, with scant aerial my-celium, subfelty, pale olive. Reverse dark grey to black. Colo-nies on PCA attaining a diameter of 8 mm, with margin sub-merged, fringed, radially striated, very dark olive grey to black,with aerial mycelium only at the center, subfelty, pale olivegrey with a reverse diffuse, dark grey. Colonies on OTM at-taining a diameter of 9 mm, margin submerged, wide, even,very dark olive grey to black, with aerial mycelium woolly,pale brownish grey. Reverse diffuse, dark brownish grey toblack, agar raised at the center. Colonies on CZA with scantygrowth, with margin appressed, fringed, pale olive to darkolive, with little aerial mycelium, farinaceous-subfelty, paleolive. Reverse dark grey to black. Colonies on AI attaining adiameter of 7 mm, margin appressed, almost submerged,shiny, even and black, no aerial mycelium. Reverse dark greyto black.
Vegetative hyphae subhyaline to pale green, smooth, thin-walled, slightly flexuous, regularly and abundantly branchedwith angles less than 45º, septate, with nearly rectangular cells
© DGfM 2005
28 RUIBAL, PLATAS & BILLS: Melanized fungi from Mallorcan limestone
Mycological Progress 4(1) / 2005 29
© DGfM 2005
Figs. 2-7: TRN127 Trimmatostroma abietis. 2. Two-week old culture on MEA. 3. Vegetative and fertila mycelia. Bar = 50 µm.4. Conidiogenous cells. Bar = 50 µm. 5. Conidiogenous cells and conidia disarticulating. Bar = 50 µm. 6. Conidia disarticulating.Bar = 50 µm. 7. Conidia. Bar = 50 µm.

30 RUIBAL, PLATAS & BILLS: Melanized fungi from Mallorcan limestone
3–5 µm wide, slightly constricted at the septa. Myceliumtorulous, smooth, thick-walled, dark olive to black, with thewall pigmentation varying along the hypha, often with swol-len cells, consistently with transverse septa, sometimes formoblique septa (Figs. 9, 10). Conidium-like hyphal fragmentsdevelop by frequent disarticulation of torulous hyphae, withhyphal fragments one or more celled (Fig. 11).
TRN137 (Figs. 12–15). Colonies on MEA attaining a dia-meter of 8 mm, with margin submerged, fringed, pale olivewhite at the margin, and appressed, even, black at the center.Aerial mycelium farinaceous to subfelty, with shiny, clearexudates, pale olive. Reverse dark grey to black, agar slightlyraised at the center. Colonies on PCA attaining a diameter of9 mm, with margin submerged, irregular, zonate, very darkolive grey to black, with peripheral aerial mycelium farinace-ous, exuding clear droplets, at the center denser, almost vel-vety, pale olive grey. Reverse dark grey to black. Colonies onOTM attaining a diameter of 9 mm, margin submerged, wide,even, very dark olive grey to black, with aerial mycelium den-se, woolly, pale brownish grey. Reverse diffuse, dark brown-ish grey to black, agar raised at the center. No growth on CZA.Colonies on AI attaining a diameter of 7 mm, margin appress-ed almost submerged, bright, even, olive to dark olive, withaerial mycelium very scarce, pale brownish grey. Reverse darkgrey to black, agar slightly raised at the center.
Young hyphae subhyaline to pale green, smooth, thin-walled, flexuous, irregularly and abundantly branched withangles near 45º, septate, with cells nearly rectangular, up to5 µm wide. Mature hyphae densely septate and strongly me-lanized, olive to dark olive, with walls smooth, thick-walled.In age, developing a torulous appearance, irregularly undulat-ing hyphae consisting of irregular ovoid to subspherical cells,7–10 µm wide, (Figs. 13, 14), occasionally developing oneor more transverse septa. Sometimes oblique septa formed,rarely developing lateral conidia-like cells by sympodial coni-diogenesis (Fig. 15).
TRN152 (Figs. 16–19). Colonies on MEA attaining a dia-meter of 13 mm, with margin appressed, even to variable, olive-yellow. Aerial mycelia farinaceous-velvety, olive-yellow.Reverse dark olive grey, agar slightly raised at the centerof the colony. Colonies on PCA attaining a diameter of 12 mm,zonate, with margin submerged, even, brownish olive. Aerialmycelia farinaceous towards the edge, subfelty towards the in-terior, olive-yellow, with a reverse black. Colonies on CZA at-taining a diameter of 11 mm, with the margin, submerged atthe very edge, even, olive, with aerial mycelia farinaceous-vel-vety, grayish olive, with a reverse olive to black, with the agar
slightly buckled beneath the surface. Colonies on AI attaininga diameter of 11 mm, with the margin appressed, wavy, even;olive-grey. Aerial mycelia almost absent, yellow-ish grey witha reverse black, agar slightly raised at the colony center.
Young vegetative hyphae subhyaline, straight, rarelybranched, with nearly rectangular cells, 3–5 µm wide, withage hyphae become septate, pale olive, smooth, thick walled,flexuous, irregularly to frequently branched, with angles near45º. Mycelia with lengths that are torulose or strongly bent(Fig. 17), consisting of cells of irregular width and shape andincluding intercalary swollen cells (Fig. 18), with lateral andtransverse septa abundant. In some zones of the mature my-celium, short torulose branches (10–30 µm long) develop atnear-90º angles that may correspond with developing coni-diophores. On PCA, forming immature or aborted pycnidialconidiomata (Fig. 19), dark olive to black, solitary or in smallgroups, subspherical to ovoid (40–100 µm) with a peridiumpseudoparenchimatous, conidia not observed.
TRN153 (Figs. 20–23). Colonies on MEA attaining a dia-meter of 10 mm. Margin submerged, even, dark olive to black,or fimbriate, olive yellow, with aerial mycelia farinaceous,dark olive-yellow. Reverse dark olive-grey, agar slightly rais-ed at the center of the colony. Colonies on PCA attaining adiameter of 9 mm, with a margin submerged, fimbriated, darkbrownish olive, with aerial mycelia farinaceous, light brown-ish grey, with a reverse black. Colonies on OTM attaining adiameter of 11 mm, with a margin submerged, even, irregular,dark brown, with aerial mycelium very scarce, farinaceous,light brownish grey, with a reverse black. Colonies on CZAattaining a diameter of 10 mm, with the margin raised, olive-grey, with aerial mycelia crustose, olive-grey, with a reverseblack and the agar splitting beneath the colony. Colonies onAI attaining a diameter of 9 mm, with the margin appressed,to fimbriate, irregular, greyish olive, aerial mycelia absent.Reverse dark olive-grey.
Mycelium septate, subhyaline to pale red, smooth, thick-walled, with young vegetative hyphae straight, scarcely branch-ed, with nearly rectangular cells, 3–5 µm wide, in age deve-loping abundant, muriform to multicellular structures of va-riable size and shape, that may correspond to immature coni-diomata (Fig. 21). Terminal lengths of vegetative myceliumdeveloping a torulose to moniliform pattern, with rounded, re-gular cells (Fig. 22), lateral and transverse septa frequent, insome areas the mature, moniliform mycelium seemingly dis-articulates, indicating vegetative reproduction from the my-celia (Fig. 22). Short, torulose, lateral hyphal length may alsocorrespond to developing conidiophores (Fig. 23).
© DGfM 2005
Figs. 8-15: Microscopic features of two unidentified fungi from limestone. Figs 8-11: TRN138. 8. Two-week old culture onMEA 9. Vegetative and fertile mycelia. Bar = 200 µm. 10. Vegetative and fertile mycelia. Bar = 50 µm. 11. Conidiogenouscells and conidia. Bar = 60 µm. Figs. 12-15: TRN137. 12. Two-week old culture on MEA. 13. Vegetative and fertile mycelia.Bar = 50 µm. 14. Fertile mycelium and conidiogenous cells. Bar = 50 µm. 15. Conidiogenous cells and conidia. Bar = 5 µm.
Mycological Progress 4(1) / 2005 31
© DGfM 2005

32 RUIBAL, PLATAS & BILLS: Melanized fungi from Mallorcan limestone
Grouping and identification of isolates based on
molecular biology techniques.
Recognition of genotypes by microsatellite-primed PCR
(MP-PCR). Cluster analysis of MP-PCR products of the 117initial isolates indicated that there were 39 different genotypes.At least one example of each genotype was ITS-sequenced.In several cases, more than one representative of each geno-type was ITS-sequenced to verify the assumption that a groupof isolates from a common MP-PCR pattern represented asingle species (Tab. 2). Therefore, ITS sequences were obtain-ed from a total of 55 isolates (Tab. 1).
Amplification and sequencing of the ITS region of rDNA.
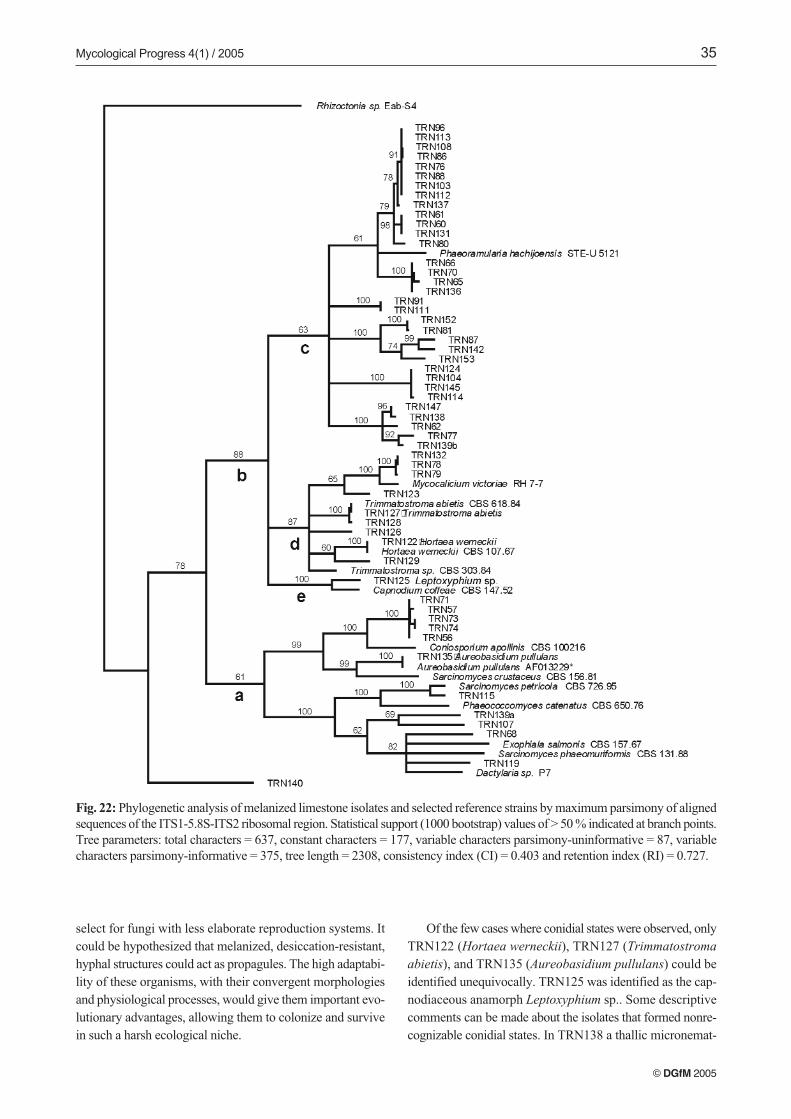
The ITS sequences of the 55 isolates (sequences lengths vary-ing from 456 to 579 bp) were aligned with sequences fromGenBank, which were selected based on sequence homologies.A phylogenetic tree was generated by the maximum parsi-mony method, and sequences of reference strains were incor-porated into the tree (Fig. 22). To estimate the relative phylo-genetic relationships among the unknown isolates and thereference strains, a basidiomycete was chosen as outgroup(Rhizoctonia sp. Eab-S4) because FASTA analysis indicatedthe isolates spanned a broad spectrum of the phylum Asco-mycota. Needless to say, the alignment of a heterogeneous as-semblage of sequences was rather deficient and ambiguousamong the isolates, resulting in many regions with low boot-strap support and an overall disorganized topology with a lowconsistency index (Fig. 22). Mycelial isolate, TRN140, wasthe most disparate ITS sequence and was not clearly associatedwith any other grouping of strains (Fig. 22). The remainder ofthe tree was divided in two main branches (Fig. 22, branchesa and b). The first branch (Fig. 22, a), with low bootstrap sup-port included reference sequences belonging to the classes Do-thideomycetes and/or Chaetothyriomycetes, including mem-bers of Herpotrichiellaceae and Dothioraceae, and several mis-cellaneous mitosporic ascomycetes. Many isolates in thisbranch could not be associated with any known sequences,except in the case of TRN135 that was microscopically iden-tified as Aureobasidium pullulans.
The other main branch (Fig. 22, b) included the majorityof the isolates and was well-supported. It was further dividedinto three sub-branches (Fig. 22, branches c, d and e) two ofthem (branches d and e) with quite good support. Branch d(Fig. 22, d) included the isolates TRN122 (Hortaea werneckii)and TRN127 (Trimmatostroma abietis) belonging to the fa-mily Mycosphaerellaceae (class Dothideomycetes) and other
ascomycetes of uncertain affinities. Again, only isolates thatwere identified microscopically matched with previous se-quences. Branch e seemed to correspond with the order Cap-nodiales, although there were insufficient data to identifyTRN125, the one isolate that grouped in this branch. The lastbranch (Fig. 22, c) included the majority of isolates. It was fur-ther subdivided into well-supported groups that only showeda certain homology with Phaeoramularia hachijoensis in theMycosphaerellaceae (BRAUN et al. 2003).
Amplification and sequencing of the 18S gene in the rDNA.
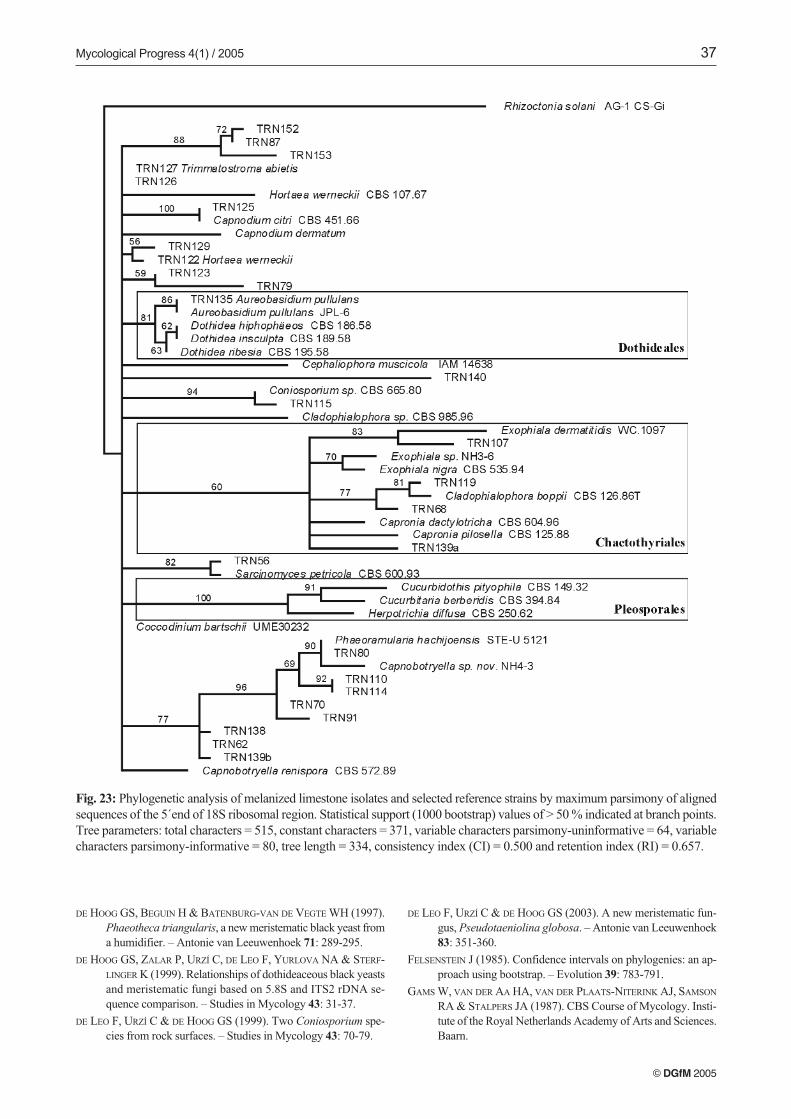
Because of the lack of ITS sequences in GenBank that wereclosely homologous with our isolates, their taxonomic positi-ons remained difficult to elucidate. Therefore, we selected 26isolates, that covered the range of different ITS genotypesrepresented among the groups appearing in the ITS tree (Fig.22). The 5’ end of the 18S gene of these 26 isolates was se-quenced (Tab. 1), sequences lengths varied from 495 to 515 bp,and those sequences were compared with others depositedin GenBank (Fig. 23). Several sequences of known fungi wereselected, based on their homology, or in other cases, samplinga limited range of strains from orders that potentially could berelated. An alignment and a phylogenetic tree were generatedby the maximum parsimony method (Fig. 23). The tree wasrooted using the basidiomycete Rhizoctonia solani E17097.
The 18S tree identified three main monophyletic groupscorresponding to three orders. The first one, corresponding tothe Pleosporales, did not encompass any isolate. This branch,with its monophyletic structure and good statistical supportconfirmed the structural quality of the tree, grouping threewell-known members of that order. Similarly, the order Do-thideales was also well delimited, with the branch includingonly one isolate, TRN135, identified as Aureobasidium pullu-
lans. The last order identified was the Chaetothyriales, althoughwith only moderate support. This was the most complexbranch and corresponded mainly with the family Herpotri-chiellaceae, including reference sequences of Exophiala, Cap-
ronia and Cladophialophora species, and included some lime-stone isolates that did not match with any known fungus. Therest of the tree showed little organized structure, with little si-milarity between any reference sequence and the rest of ourisolates, except in the case of TRN125 Leptoxyphium sp., thatshowed high homology with Capnodium citri. Nevertheless,these two sequences did not group with other reference se-quences of Capnodiales (e.g., Capnodium dematum, Capno-
botryella renispora and Capnobotryella sp.) and a monophy-letic grouping of Capnodiales was not evident as in previous
© DGfM 2005
Figs. 16-23: Microscopic features of two unidentified fungi from limestone. Figs. 16-19: TRN152. 16. Two-week old cultureon MEA. 17. Vegetative mycelia. Bar = 200 µm. 18. Vegetative mycelia and chlamydospore-like structures. Bar = 50 µm.19. Immature or aborted pycnidia-like structures. Bar = 200 µm. Figs. 20-23. TRN153. 20. Two-week old culture on MEA. 21.
Vegetative mycelia and muriform structures. Bar = 50 µm. 22. Fragmenting torulose mycelium. Bar = 50 µm. 23. Toruloselateral hyphae. Bar = 50 µm.
Mycological Progress 4(1) / 2005 33
© DGfM 2005

34 RUIBAL, PLATAS & BILLS: Melanized fungi from Mallorcan limestone
studies (REYNOLDS 1998). The 18S gene sequence of the ex-neotype strain Hortaea werneckii CBS 107.67 was divergentfrom the 18S gene sequence of the same reference strain, al-though the ITS sequence of the isolate TRN122 was identicalto the ITS sequence of the same strain. This divergence is at-tributable to 16 nucleotidic positions different in an alignmentof 514 nucleotides, and two positions indeterminate in the se-quence of the 18S gene of Hortaea werneckii CBS 107.67.Two reference strains of Hortaea werneckii, CBS 107.67 andCBS 359.66, have idenical ITS sequences and were consider-ed conspecific (DE HOOG et al. 1999). Therefore we think ouridentification of the isolate based on the results of the ITS treeis correct.
Database searches with 18S rDNA sequences of the lime-stone isolates yielded few close matches, once again makingdifficult taxonomic positioning of isolates. However, the sear-ches remained inconclusive, because 18S gene sequences da-tabases are still not extensive, and some of our sequences seemto be the first of their kind to be included in a public database.An example of this lack of data is the case of TRN122 andTRN127, which were identified by their morphology and ITSsequences; however, it was not possible to ascertain their po-sitions based on the 18S gene sequences because a lack of cor-responding 18S sequence data.
Discussion
Several new dematiaceous fungi isolated from natural andman-made rock surfaces have been described in recent years(STALEY, PALMER & ADAMS 1982; DE HOOG, BEGUIN & BA-TENBURG-VAN DE VEGTE 1997; STERFLINGER et al. 1997; STER-FLINGER & KRUMBEIN 1997; WOLLENZIEN et al. 1997; DE LEO,URZÍ & DE HOOG 1999; ONOFRI et al. 1999; DE LEO, URZÍ &DE HOOG 2003). In this study, limestone surfaces were con-firmed once again to be an important source of these orga-nisms. Although the literature indicates that novel fungal taxawere to be expected in limestone and marble surfaces (STER-FLINGER et al. 1997), especially in arid or Mediterranean cli-mates, the abundance and variety of isolates we encounteredsurpassed our expectations. Our isolation technique was si-milar to those used by others studying melanized fungi in man-made rock surfaces (DE LEO, URZÍ & DE HOOG 2003). The hightaxonomic diversity found indicated that our isolation proce-dures were reliable and adequate, and that the novelty of thespecies were consistent with previous investigations (BRAAMS
1992; STERFLINGER & KRUMBEIN 1997; STERFLINGER & PRIL-LINGER 2001; GORBUSHINA et al. 2002). The isolation and cul-ture methods possibly could have been further improved andexpanded, mainly by varying the ranges of culture tempera-tures and osmotic concentrations of isolation plates. Perhapsby mimicking the oligotrophic conditions that the fungi en-counter in their natural environment, carrying out isolationsat extreme temperatures and solute concentrations, and expos-ing isolation plates to ultraviolet radiation, the number andkinds of isolates would have been further extended.
A typical approach to grouping large numbers of unknownstrains into operational taxonomic units or morphological isto use visual clues, especially mycelial growth rate, pigmen-tation, and texture, expressed on sets of standard media(BILLS et al. 2004). Because most of the limestone isolateswere similarly pigmented and very slow growing, visually dis-criminating features were generally absent among strains. Thecomparison and grouping strategy based on MP-PCR, com-plemented by the macromorphological appearance of the iso-lates, turned out to be a useful method for the sorting and group-ing of isolates, allowing us to sequence and analyze fewernumbers of isolates without losing diversity. Sequenc was use-ful for relative phylogenetic positioning and indicating no-velty, although was less helpful for definitive identification.In summary, the analysis revealed that a remarkable diversityof ascomycetous fungi, many apparently unknown, can befound on limestone surfaces. Several very common genotypeswere identified among our isolates and some of these geno-types are very similar to those found in other rock types (BUTIN
et al. 1996; STERFLINGER & KRUMBEIN 1997; STERFLINGER &PRILLINGER 2001) or sampled by us in other regions of Spain(results not shown).
Our preliminary evaluation did not reveal any genotypicspecificity for the types of rock or the sites sampled, as evi-denced by common genotypes between both isolation sites.However, we only observed fungi from two arbitrarily select-ed small areas; a more extensive survey that mapped occur-rence on a fine spatial scale might detect preferential coloni-zation patterns. On the other hand, melanized fungi on rocksurfaces may only be transient inhabitants, with their distri-bution being largely determined by chance dispersal. Rocksurfaces may merely provide mechanical support and protec-tion, and the mineral composition may produce minimal ef-fect on fungal nutritional metabolism. These rock-inhabitingorganisms would use exogenous sources of nutrients onlywhen available, in a classic oligotrophic pattern. That is, wa-ter from rain, fog, and surface ran off carrying dissolved orsuspended organic and/or inorganic nutrients to the vicinityof fungal cells.
The general morphological features of our set of isolatesseemed to correspond with adaptations of these fungi to theextreme environmental conditions of rock surfaces (GORBU-SHINA 2003), especially the shared aspect of melanin pigments,presumably related to protection against desiccation and ul-traviolet radiation. The slow rates and granular meristematicgrowth could be related with energy conservation, necessaryfor survival on substrates with a very scarce nutrient input. Atthe microscopic level, a general lack of specialized reproduc-tive structures was observed. The lack of sporulation could bean artifact of the cultural conditions in many cases. However,if nonsporulation in situ on rock surfaces was the prevailingmycelial condition, it could represent also an adaptation toconserve energy. Alternatively, the scarcity of nutrients andwater in the habitat and the considerable energetic costs re-quired to produce such complex reproductive structures might
© DGfM 2005
Mycological Progress 4(1) / 2005 35
© DGfM 2005
Fig. 22: Phylogenetic analysis of melanized limestone isolates and selected reference strains by maximum parsimony of alignedsequences of the ITS1-5.8S-ITS2 ribosomal region. Statistical support (1000 bootstrap) values of > 50 % indicated at branch points.Tree parameters: total characters = 637, constant characters = 177, variable characters parsimony-uninformative = 87, variablecharacters parsimony-informative = 375, tree length = 2308, consistency index (CI) = 0.403 and retention index (RI) = 0.727.
select for fungi with less elaborate reproduction systems. Itcould be hypothesized that melanized, desiccation-resistant,hyphal structures could act as propagules. The high adaptabi-lity of these organisms, with their convergent morphologiesand physiological processes, would give them important evo-lutionary advantages, allowing them to colonize and survivein such a harsh ecological niche.
Of the few cases where conidial states were observed, onlyTRN122 (Hortaea werneckii), TRN127 (Trimmatostroma
abietis), and TRN135 (Aureobasidium pullulans) could beidentified unequivocally. TRN125 was identified as the cap-nodiaceous anamorph Leptoxyphium sp.. Some descriptivecomments can be made about the isolates that formed nonre-cognizable conidial states. In TRN138 a thallic micronemat-

36 RUIBAL, PLATAS & BILLS: Melanized fungi from Mallorcan limestone
© DGfM 2005
ous conversion of torulous hyphae was observed (Figs. 8–11).Conidia may develop terminally or intercalarly, initially beingunicellular and hyaline, but becoming pigmented during ma-turation and differentiation. Later, hyphal cells developed lon-gitudinal and transverse septa, becoming muriform in appea-rance. Finally, conidium-like cells or cell aggregates, whichwere released by hyphal disarticulation, presumably acting asvegetative propagules (Fig. 11). Such disarticulation of me-lanized, torulous hyphae, whose cells had developed one ormore transverse septa, was also observed in other predomi-nantly mycelial isolates, suggesting that thallic micronema-tous conidiogenesis or simple reproduction by propagativefragmentation. Nevertheless, other conidial forms were evi-dent that should be taken into account. For example, in thecase of TRN137 (Figs. 12–15), a scant, incomplete blastic co-nidiogenesis was observed. In other isolates, a poorly deve-loped Cladosporium-like conidial state was infrequently ob-served, although not identifiable as any particular species. Insome cases, we potentially could have related some of thesefungi to anamorph form genera characterized by fragmenta-tion of torulose mycelia, or muriform and meristematic cellaggregations, e.g., Coniosporium, Sclerococcum, Taeniolel-
la, and Trimmatostroma. Most the species in these genera areonly known from specimens on plants and lichens; little, ifanything, is known of their behaviour in agar culture. There-fore, we preferred not assign them to form genera.
The majority of the isolates were extremely difficult to de-scribe in classical morphological terms because of the scarcityof diagnostic features. Therefore it was necessary to resort tomolecular data in order to reveal information relevant for theiridentification and classification. The rDNA sequences offeredsurprising data. Sequences of ITS1-5.8S-ITS2 region of themajority of the isolates differed from sequences of fungi pre-viously found on stone, and most isolates differed greatly fromsequences stored in GenBank. Nevertheless, the majoritycould be loosely grouped in two main classes of Ascomyce-tes: Dothideomycetes and Chaetothyriomycetes. Many otherisolates did not correspond with any sequence from GenBank,suggesting we recovered unknown genera that did not corre-spond well with any well-defined order or family.
The aligned sequences of the 5´end of the 18S of the lime-stone isolates did not produce a well-structured tree. Thissituation was caused by the scarcity of sequences from thatregion in GenBank that corresponded to the isolates. Onlygeneral information about the taxonomic positioning of the iso-lates could be inferred from the analysis of 18S gene sequen-ces. Nonetheless, results from the 18S gene were in agreementwith the conclusions drawn from sequences of the ITS region.Many isolates once again could be loosely grouped among theDothideomycetes and Chaetothyriomycetes. Our results withfungi of the Capnodiales appeared equivocal. Some fungi pre-viously associated with the order, e.g., Capnobotryella sp. andC. renispora (YOSHIDA et al. 1996; HAMBLETON, TSUNEDA &CURRAH 2003), did not form a monophyletic clade with mem-bers sensu stricto of the order, e.g., Capnodium citri and C.
dematum. Again many isolates did not group with any se-quence of GenBank, and even they did not group with eachother, suggesting, not only a scarcity of 18S sequences in Gen-Bank related to the kinds of fungi we recovered, but also verydiverse origins of the isolates.
Sampling of the limestone in this study was arbitrary. Theonly criterion used was to select rock samples that would berelatively clean and free from influence of soil and lichens.No effort was made to enrich the samples by choosing stain-ed or biologically deteriorated surfaces or surfaces that wereobviously colonized by black fungal structures. Even with ar-bitrary selection of rock substrata, a set of isolates was obtain-ed that exhibited a combination of cryptic morphology andphylogenetic affinities agreeing with previous studies on rock-inhabiting fungi of the Mediterranean. The behavior of the iso-lates was consistent with the strong habitat specialization andcomplex relations maintained through a highly modified phy-siology and metabolism (STERFLINGER & KRUMBEIN 1997;STERFLINGER 2000; GORBUSHINA 2003). The high degree ofnovelty and large variety of the fungi isolated from only a li-mited examination of rock surfaces points to a very deep anddiverse pool of Ascomycete species in this habitat. Thereforefungi of rock surfaces of the Mediterranean region appear tobe an attractive target for natural products screening for newfungal metabolites.
Acknowledgements
We are grateful to Angel Ginés (Museu Balear de CiènciesNaturals) for providing detailed geological information andto Enrique Descals (Institut Mediterrani d'Estudis Avançats)for guiding selection of sampling sites. Sybren de Hoog, Enri-que Descals, and anonymous reviewers made helpful com-ments and corrections to the manuscript. Ana Pérez providedexpert technical assistance. This work is a portion of a doc-toral dissertation submitted to the Faculty of Science, Univer-sidad Autónoma de Madrid.
References
BILLS GF, CHRISTENSEN M, POWELL M & THORN G (2004). Sapro-bic soil fungi. Biodiversity of fungi. Inventory and monitoringmethods: 271-302. G. M. Mueller, G. F. Bills & M. S. Foster.Elsevier Academic Press. Burlington, MA, USA.
BOGOMOLOVA EV & MINTER DW (2003). A new microcolonial rock-inhabiting fungus from marble in Chersonesos (Crimea, Ukrai-ne). – Mycotaxon 86: 195-204.
BRAAMS J (1992) Ecological studies of the fungal microflora inha-biting historical sandstone monuments. PhD Thesis, Univer-sity of Oldenburg.
BRAUN U, CROUS PW, DUGAN F, GROENEWALD JZ & DE HOOG GS(2003). Phylogeny and taxonomy of Cladosporium-like hy-phomycetes, including Davidiella gen. nov., the teleomorphof Cladosporium s. str. – Mycological Progress 2: 3-18.
BUTIN H, PEHL L, DE HOOG GS & WOLLENZIEN U (1996). Trimmato-
stroma abietis sp. nov. (hyphomycetes) and related species. –Antonie van Leeuwenhoek 69: 203-209.
Mycological Progress 4(1) / 2005 37
© DGfM 2005
Fig. 23: Phylogenetic analysis of melanized limestone isolates and selected reference strains by maximum parsimony of alignedsequences of the 5´end of 18S ribosomal region. Statistical support (1000 bootstrap) values of > 50 % indicated at branch points.Tree parameters: total characters = 515, constant characters = 371, variable characters parsimony-uninformative = 64, variablecharacters parsimony-informative = 80, tree length = 334, consistency index (CI) = 0.500 and retention index (RI) = 0.657.
DE HOOG GS, BEGUIN H & BATENBURG-VAN DE VEGTE WH (1997).Phaeotheca triangularis, a new meristematic black yeast froma humidifier. – Antonie van Leeuwenhoek 71: 289-295.
DE HOOG GS, ZALAR P, URZÍ C, DE LEO F, YURLOVA NA & STERF-LINGER K (1999). Relationships of dothideaceous black yeastsand meristematic fungi based on 5.8S and ITS2 rDNA se-quence comparison. – Studies in Mycology 43: 31-37.
DE LEO F, URZÍ C & DE HOOG GS (1999). Two Coniosporium spe-cies from rock surfaces. – Studies in Mycology 43: 70-79.
DE LEO F, URZÍ C & DE HOOG GS (2003). A new meristematic fun-gus, Pseudotaeniolina globosa. – Antonie van Leeuwenhoek83: 351-360.
FELSENSTEIN J (1985). Confidence intervals on phylogenies: an ap-proach using bootstrap. – Evolution 39: 783-791.
GAMS W, VAN DER AA HA, VAN DER PLAATS-NITERINK AJ, SAMSON
RA & STALPERS JA (1987). CBS Course of Mycology. Insti-tute of the Royal Netherlands Academy of Arts and Sciences.Baarn.

© DGfM 2005
38 RUIBAL, PLATAS & BILLS: Melanized fungi from Mallorcan limestone
GARDES M & BURNS TD (1993). ITS primers with enhanced speci-ficity for basidiomycetes - Application to the identification ofmycorrhizae and rust. – Molecular Ecology 2: 113-118.
GINÉS A (1998). L´exokarst de la serra de Tramuntana de Mallor-ca. Aspectes geológics de les Balears: 361-389. U. I. Balears.Fornós, J. J.
GINÉS A & GINÉS J (1995). The exokarstic landforms of Majorca is-land. – ENDINS nº 20. Monogr. Soc. Hist. Nat. Balears 3: 59-70.
GORBUSHINA AA (2003). Methodologies and techniques for detect-ing extraterrestrial (microbial) life. Microcolonial fungi: sur-vival potential of terrestrial vegetative structures. – Astrobio-logy 3: 543-554.
GORBUSHINA AA, KRUMBEIN WE, HAMMAN CH, PANINA L, SOUK-HARJEVSKI S & WOLLENZIEN U (1993). Role of black fungiin color change and biodeterioration of antique marbles. –Geomicrobiology Journal 11: 205-221.
GORBUSHINA AA, LYALIKOVA NN, VLASOV DY & KHIZHNYAK TV(2002). Microbial communities on the monuments of Mos-cow and St. Petersburg: biodiversity and trophic relations. –Microbiology 71: 350-356.
HAMBLETON S, TSUNEDA A & CURRAH RS (2003). Comparative mor-phology and phylogenetic placement of two microsclerotialblack fungi from Sphagnum. – Mycologia 95: 959-975.
KROKEN S, GLASS NL, TAYLOR J, YODER OC & TURGEON B (2003).Phylogenomic analysis of type I polyketide synthase genes inpathogenic and saprobic ascomycetes. – Proceedings of theNational Academy of Sciences, USA 100: 15670-15675.
LARENA I, SALAZAR O, GONZÁLEZ V, JULIAN MC & RUBIO V (1999).Design of a primer for ribosomal DNA internal trascribed spa-cer with enhanced specificity for ascomycetes. – Journal ofBiotechnology 75: 187-194.
LONGATO S & BONFANTE P (1997). Molecular identification of my-corrhizal fungi by direct amplification of microsatellite regi-ons. – Mycological Research 101: 425-432.
NICHOLAS KB & NICHOLAS HB (1997). GeneDoc, a tool for editingand annotating multiple sequence alignments. Software dis-tributed by the author.
O`DONNELL K (1993). Fusarium and its near relatives in the fungalholomorph: mitotic, meiotic and pleomorphic speciation infungal systematics. CABInternational Wallingford.
ONOFRI S, PAGANO S, ZUCCONI L & TOSI S (1999). Friedman-
niomyces endolithicus (Fungi, Hyphomyces), anam.-gen. andsp. nov., from continental Antarctica. – Nova Hedwigia 68:175-181.
PELAEZ F, PLATAS G, COLLADO J & DÍEZ MT (1996). Infraspecificvariation in two species of aquatic hyphomycetes, assessed byRAPD analysis. – Mycological Research 100: 831-837.
REYNOLDS DR (1998). Capnodiaceous sooty mold phylogeny. – Ca-nadian Journal of Botany 76: 2125-2130.
SNEATH PHA & SOKAL RR (1997). Numerical taxonomy. W.H. Free-man & Co. San Francisco.
STALEY JT, PALMER FE & ADAMS JB (1982). Microcolonial fungi:Common inhabitants on desert rocks? – Science 215: 1093-1095.
STERFLINGER K (2000). Fungi as geologic agents. – GeomicrobiologyJournal 17: 97-124.
STERFLINGER K, DE BAERE R, DE HOOG GS, DE WACHTER R, KRUM-BEIN WE & HAASE G (1997). Coniosporium perforans and C.
apollinis, two new rock-inhabiting fungi isolated from mar-ble in the Sanctuary of Delos (Cyclades, Greece). – Antonievan Leeuwenhoek 72: 349-363.
STERFLINGER K, DE HOOG GS & HAASE G (1999). Phylogeny andecology of meristematic ascomycetes. – Studies in Mycology43: 5-22.
STERFLINGER K & KRUMBEIN WE (1997). Dematiaceous fungi as amajor agent for biopitting on mediterranean marbles and li-mestones. – Geomicrobiology Journal 14: 219-230.
STERFLINGER K & PRILLINGER H (2001). Molecular taxonomy andbiodiversity of rock fungal communities in an urban enviro-ment. – Antonie van Leeuwenhoek 80: 275-286.
SWOFFORD DL (1998). PAUP*. Phylogenetic Analysis Using Parsi-mony and Other Methods. Version 4. Sinauer Associates. Sun-derland, Massachusetts, U.S.A.
TAYLOR JW, JACOBSON DJ, KROKEN S, KASUGA T, GEISER DM, HIB-BETT DS & FISCHER MC (2000). Phylogenetic species reco-gnition and species concepts in fungi. – Fungal Genetics andBiology 31: 21-32.
THOMPSON JD, GIBSON TJ, PLEWNIAK F, JEANMOUGIN F & HIGGINS
DG (1997). The ClustalX windows interface: flexible strate-gies for multiple sequence alignment aided by quality analy-sis tools. – Nucleic Acids Research 24: 4876-4882.
WHITE TJ, BRUNS T, LEE S & TAYLOR J (1990). Amplification anddirect sequencing of fungal ribosomal RNA genes for phylo-genetics. In PCR Protocols: A Guide to Methods and Ampli-fications. Academic Press. New York.
WOLLENZIEN U, DE HOOG GS, KRUMBEIN WE & UIJTHOF JMJ (1997).Sarcinomyces petricola, a new microcolonial fungus frommarble in the Mediterranean basin. – Antonie van Leeuwen-hoek 71: 281-288.
WOLLENZIEN U, DE HOOG GS, KRUMBEIN WE & URZÍ C (1995). Onthe isolation of microcolonial fungi occurring on and in mar-ble and other calcareous rocks. – The Science of the Total En-viroment 167: 287-294.
YOSHIDA S, TAKEO K, DE HOOG GS, NISHIMURA K & MIYAJI M(1996). A new type of growth exhibited by Trimmatostroma
abietis. – Antonie van Leeuwenhoek 69: 211-215.
Accepted: 14.7.2004