introduced herbivores and the allee effect in animal- pollinated plants
TRANSCRIPT

Junio de 2007 ALLEE EFFECT IN ANIMAL-POLLINATED PLANTS
Sección especial
Recibido: 14 de enero de 2007; Fin de arbitraje: 10 de abrilde 2007; Revisión recibida: 9 de mayo de 2007; Aceptado:11 de mayo de 2007
* Instituto Argentino de Investigaciones de lasZonas Áridas, Centro Regional de InvestigacionesCientíficas y Tecnológicas, CONICET, CC 507, 5500Mendoza, [email protected]
Ecología Austral 17:21-36. Junio 2007Asociación Argentina de Ecología
Introduced herbivores and the Allee effect in animal-pollinated plants
DIEGO P VÁZQUEZ *
Instituto Argentino de Investigaciones de las Zonas Áridas, Centro Regional de Investigaciones Científicas yTecnológicas, CONICET, Mendoza, Argentina
ABSTRACT. Interspecific interactions can be modulated by the community context in which theyoccur. In this paper I focus on plant–pollinator interactions and their modulation by introducedherbivores through changes in plant population density. I first develop a mathematical modelthat relates the population density of an animal-pollinated plant species with pollinator visitationfrequency, pollination, reproduction and per capita growth rate. This model allows me toinvestigate how several pollination-related parameters—including relative abundance of thefocal plant species, pollinator specialization, pollination quantity and quality and pollen limitationof reproduction—influence the relationship between per capita growth rate and population density.I then use this theoretical framework to discuss two case studies that have addressed the density-mediated effects of introduced herbivores on plant reproduction. This exercise allows me topoint out several limitations of previous studies and to suggest avenues for future research.
[Keywords: abundance, Allee effect, density dependence, introduced herbivores, mutualism,plant reproduction, pollination, pollinator specialization and generalization, population dynamics]
RESUMEN. Herbívoros introducidos y el efecto Allee en plantas polinizadas por animales: Lasinteracciones interespecíficas pueden ser moduladas por el contexto comunitario en el que ocurren.Este artículo estudia las interacciones planta-polinizador y su modulación por herbívorosintroducidos mediante cambios en la densidad poblacional de las plantas. En primer lugardesarrollo un modelo matemático que relaciona la densidad poblacional de una planta polinizadapor animales con la frecuencia de visitas por polinizadores, la polinización, la reproducción y latasa per capita de crecimiento poblacional. Este modelo me permite investigar la influencia devarios parámetros relacionados con la polinización—incluyendo la abundancia relativa de laespecie focal, la especialización de los polinizadores, la cantidad y la calidad de la polinización yla limitación por polen de la reproducción—sobre la relación entre la tasa per capita de crecimientoy la densidad poblacional. Luego utilizo este marco teórico para discutir dos estudios de caso quehan evaluado los efectos mediados por la densidad de los herbívoros introducidos sobre lareproducción de las plantas. Este ejercicio me permite señalar algunas limitaciones de los estudiosprevios y sugerir direcciones para investigaciones futuras.
[Palabras clave: abundancia, efecto Allee, densodependencia, herbíviros introducidos,mutualismo, reproducción de plantas, polinización, especialización y generalización depolinizadores, dinámica poblacional]
Sección especial

DP VÁZQUEZ Ecología Austral 17:21-36
Sección especial
INTRODUCTION
One of the basic principles of ecology is thedensodependent regulation of populations(considered a “law” by some; Lawton 1999;Turchin 2001). Population regulation resultsfrom the density dependence of demographicparameters (Hixon et al. 2002), imposingbounds on population abundance and allow-ing long-term persistence. On one extreme, anupper bound may be imposed by competitionfor resources and other biological interactionsthat limit population abundance, so that thereis a negative correlation between populationdensity or size and individual fitness. This kindof negative density-dependent regulation hasreceived much attention since the origin ofecology as a discipline. For example, popula-tion regulation was the basis of Malthus’ ar-gument in his essay on the limits of humanpopulation growth (Malthus 1798), which waslater used by Darwin (1859) as an inspirationfor his idea of the struggle for survival as themechanism underlying natural selection. Like-wise, Verhulst (1845) incorporated density-dependent regulation in his now classic logis-tic model of population growth.
On the other extreme of the population abun-dance continuum, several mechanisms mayimpose a lower bound to population abun-dance, under which the population is unableto sustain itself and goes extinct. For manypopulations it is unrealistic that there will benegative density-dependence at low densitiesif there is facilitation among conspecifics, in-cluding mating in sexually reproducing organ-isms and cooperation in colonial or social or-ganisms. In these cases, there may be an in-verse density dependence at low densities (i.e.,a positive correlation between population den-sity or size and individual fitness), usuallytermed Allee effect (after W. C. Allee, who firstnoted the phenomenon; Allee et al. 1949; Den-nis 1989; McCarthy 1997; Courchamp et al.1999; Stephens & Sutherland 1999).
Animal-pollinated plants are particularlylikely to suffer from Allee effects. At low popu-lation density or size, pollinators may visitindividual plants less frequently and bringlower-quality pollen than at higher densities;
lower pollination performance can in turn leadto lower reproductive success, particularlywhen reproduction is pollen limited (Rathcke1983; Kunin 1993; Ghazoul 2005). Consider-ation of Allee effects is particularly importantwhen investigating the population ecology ofrare plants, either naturally so or made rare byhuman impact to ecosystems including habi-tat fragmentation, habitat degradation such asthat produced by fire or livestock grazing, bio-logical invasions and selective harvesting(Bond 1994; Ghazoul 2005; Aizen & Vázquez2006; Traveset & Richardson 2006). However,the Allee effect is not a necessary consequenceof decreased population density, and itsstrength may vary substantially among spe-cies; this variation will depend on several pol-lination-related factors, including the relativeabundance of the focal plant species, pollina-tor specialization, the density dependence ofpollination quality, the strength of pollen limi-tation of reproduction and the breeding system.
One situation in which plants can suffer fromAllee effects is when their abundance is de-creased following the introduction of exoticherbivores. Introduced herbivores, particularlylarge mammalian species, can lead to funda-mental changes in species composition, habi-tat structure and ecosystem processes in thehabitats they invade (de Vos et al. 1956;Ebenhard 1988; Vázquez 2002). By directlyaffecting plant population abundance, herbi-vores can also affect indirectly plant pollina-tion and reproductive success through Alleeeffects (Vázquez & Simberloff 2004; Traveset& Richardson 2006).
Here I develop a mathematical model thatincludes a mechanistic Allee effect incorporat-ing pollinator visitation, pollination and re-production into a logistic model of populationdynamics. I use the model heuristically, as atool to generate qualitative hypotheses abouthow different aspects of plant–pollinator in-teractions and plant reproductive biology af-fect the density dependent regulation of ani-mal-pollinated plants. I then exemplify the useof the model by applying it to two case studies.Although I have developed this model in thecontext of one type of ecosystem perturbation,introduced herbivores, it can be applied toother types of perturbations that operate
22

Junio de 2007 ALLEE EFFECT IN ANIMAL-POLLINATED PLANTS
Sección especial
through increased mortality; this would in-clude, for instance, harvesting and fire, butwould exclude habitat fragmentation (pro-vided that it does not increase mortality of theplant). Even in the latter case, if any condi-tions of the ecosystem change with fragmenta-tion (e.g., relative abundance or the degree ofspecialization of pollinator; see Aizen &Feinsinger 1994a 1994b) in spite of no addedmortality, the model could be used to comparethese contrasting scenarios. Using this kind ofmechanistic model could thus be useful tounderstand the varying responses of differentplant species to decreased population abun-dance (Ghazoul 2005).
MODELLING THE ALLEE EFFECT INANIMAL-POLLINATED PLANTS
Prolegomena
I start with the discrete-time logistic equationto model the population dynamics of an ani-mal-pollinated plant species,
Nt+1=Nt+rNt(1- Nt/K ) , (1)
where Nt and Nt+1 are population abundancesat time intervals t and t+1, r is the intrinsincpopulation growth rate and K is the carryingcapacity (Kot, 2001). The per capita growth rate(the number of adults produced per adult) isthen a linearly decreasing function of popula-tion density
Nt+1/Nt =1+r(1- Nt/K ) . (2)
The discrete-time logistic model is appropri-ate for unitary (non-modular) organisms withdiscrete, non-overlapping generations and noage structure. Although many plant speciesare not represented by this type demography,the simplicity of the model allows me to use itheuristically to explore how different pollina-tion and reproductive parameters modulate thedensity-mediated effects of introduced herbi-vores on the population dynamics of animal-pollinated plants. More complexity can beadded, for example by incorporating a trans-formation for modularity (Crawley 1997) or agestructure (Caswell 2001).
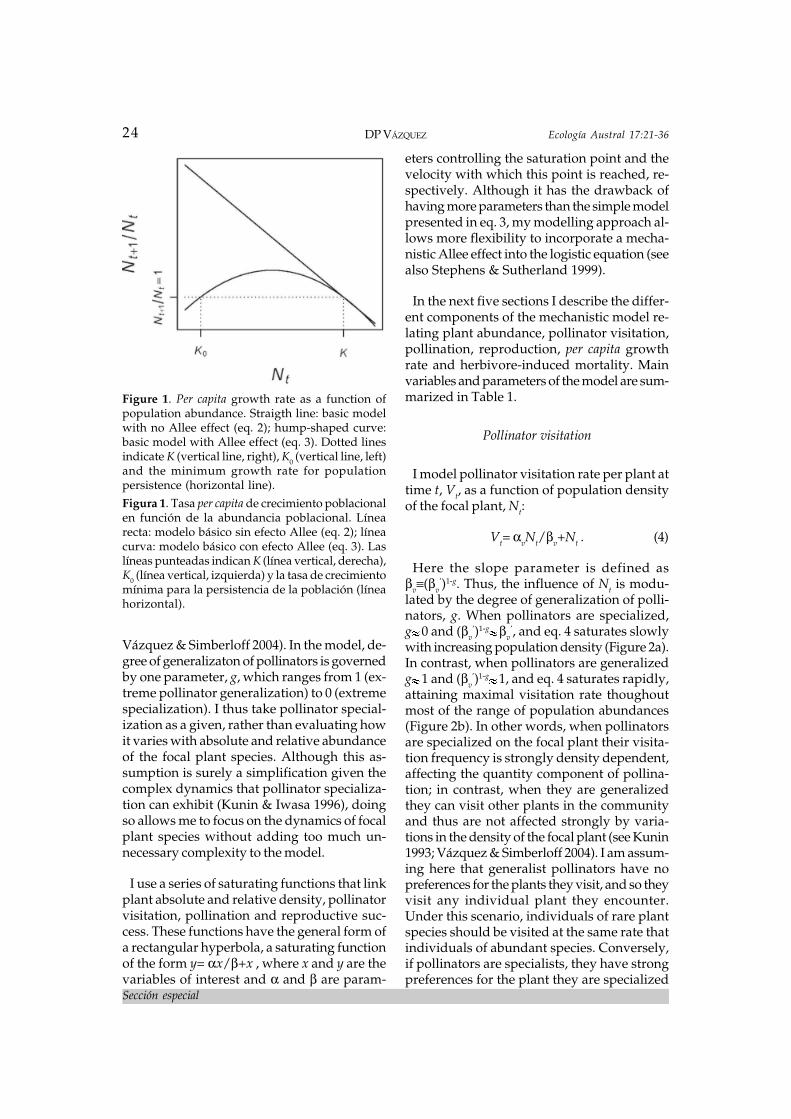
The basic model described by eq. 1 assumesthat per capita reproductive rate is a linearlydecreasing function of population size (thestraight line in Figure 1). Although this formof density dependence may be a reasonableassumption when conspecific individualscompete strongly for resources and there is nofacilitation, it is not appropriate when indi-viduals facilitate each other’s reproduction atlow densities, as may occur with animal-pol-linated plants. The most commonly used formof the logistic equation incorporating the Alleeeffect is
Nt+1=Nt+rNt( Nt-K0/K )(1- Nt/K ) , (3)
where K0 is a critical density below which thepopulation cannot reproduce (Courchamp etal. 1999; Kot 2001; Stephens & Sutherland1999). The term Nt-K0/K increases linearlywith density and causes the per capita growthrate to increase at low densities, peaking atintermediate densities (Figure 1). Albeit simple,this form of Allee effect has several limitations,particularly the lack of flexibility in the shapeof the per-capita growth rate curve (it is sym-metric) and the fact that it is not mechanistic,in the sense that it does not specify how theincrease in reproductive success at low densi-ties occurs (Stephens & Sutherland 1999).Hence, here I develop a different form of Alleeeffect, one which varies non-linearly with den-sity and which allows me to incorporatemechanistically the density-mediated effectsof herbivores on pollination and reproduction.
I assume the focal plant species is pollinatedmainly by one pollinator species (or by severalspecies that have similar characteristics). Al-though many plant species are quite general-ized in their interactions with pollinators(Waser et al. 1996; Waser & Ollerton 2006), weknow that pollinator effectiveness varieswidely among pollinator species, and usuallyone or a few species provide a large propor-tion of the total pollination service (Morris 2003;Vázquez et al. 2005); thus, the simplifying as-sumption that the focal plant is pollinated byone pollinator species seems reasonable. Pol-linators can themselves vary in their degree ofspecialization on plant species, which haveimportant implications for pollination quan-tity and quality (Rathcke 1983; Kunin 1993;
23
DP VÁZQUEZ Ecología Austral 17:21-36
Sección especial
Vázquez & Simberloff 2004). In the model, de-gree of generalizaton of pollinators is governedby one parameter, g, which ranges from 1 (ex-treme pollinator generalization) to 0 (extremespecialization). I thus take pollinator special-ization as a given, rather than evaluating howit varies with absolute and relative abundanceof the focal plant species. Although this as-sumption is surely a simplification given thecomplex dynamics that pollinator specializa-tion can exhibit (Kunin & Iwasa 1996), doingso allows me to focus on the dynamics of focalplant species without adding too much un-necessary complexity to the model.
I use a series of saturating functions that linkplant absolute and relative density, pollinatorvisitation, pollination and reproductive suc-cess. These functions have the general form ofa rectangular hyperbola, a saturating functionof the form y= αx/β+x , where x and y are thevariables of interest and α and β are param-
eters controlling the saturation point and thevelocity with which this point is reached, re-spectively. Although it has the drawback ofhaving more parameters than the simple modelpresented in eq. 3, my modelling approach al-lows more flexibility to incorporate a mecha-nistic Allee effect into the logistic equation (seealso Stephens & Sutherland 1999).
In the next five sections I describe the differ-ent components of the mechanistic model re-lating plant abundance, pollinator visitation,pollination, reproduction, per capita growthrate and herbivore-induced mortality. Mainvariables and parameters of the model are sum-marized in Table 1.
Pollinator visitation
I model pollinator visitation rate per plant attime t, Vt, as a function of population densityof the focal plant, Nt:
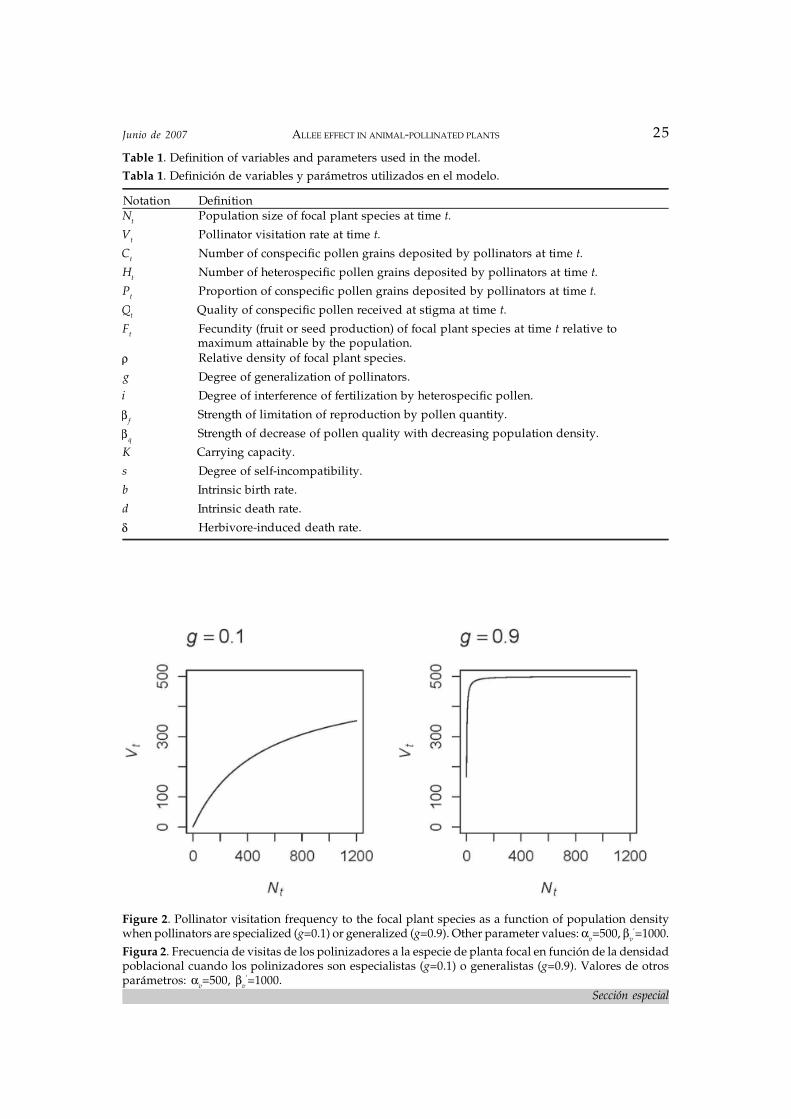
Vt= αvNt/βv+Nt . (4)
Here the slope parameter is defined asβv≡(βv
’)1-g. Thus, the influence of Nt is modu-lated by the degree of generalization of polli-nators, g. When pollinators are specialized,g 0 and (βv
’)1-g βv’, and eq. 4 saturates slowly
with increasing population density (Figure 2a).In contrast, when pollinators are generalizedg 1 and (βv
’)1-g 1, and eq. 4 saturates rapidly,attaining maximal visitation rate thoughoutmost of the range of population abundances(Figure 2b). In other words, when pollinatorsare specialized on the focal plant their visita-tion frequency is strongly density dependent,affecting the quantity component of pollina-tion; in contrast, when they are generalizedthey can visit other plants in the communityand thus are not affected strongly by varia-tions in the density of the focal plant (see Kunin1993; Vázquez & Simberloff 2004). I am assum-ing here that generalist pollinators have nopreferences for the plants they visit, and so theyvisit any individual plant they encounter.Under this scenario, individuals of rare plantspecies should be visited at the same rate thatindividuals of abundant species. Conversely,if pollinators are specialists, they have strongpreferences for the plant they are specialized
Figure 1. Per capita growth rate as a function ofpopulation abundance. Straigth line: basic modelwith no Allee effect (eq. 2); hump-shaped curve:basic model with Allee effect (eq. 3). Dotted linesindicate K (vertical line, right), K0 (vertical line, left)and the minimum growth rate for populationpersistence (horizontal line).Figura 1. Tasa per capita de crecimiento poblacionalen función de la abundancia poblacional. Línearecta: modelo básico sin efecto Allee (eq. 2); líneacurva: modelo básico con efecto Allee (eq. 3). Laslíneas punteadas indican K (línea vertical, derecha),K0 (línea vertical, izquierda) y la tasa de crecimientomínima para la persistencia de la población (líneahorizontal).
24
Junio de 2007 ALLEE EFFECT IN ANIMAL-POLLINATED PLANTS
Sección especial
Table 1. Definition of variables and parameters used in the model.Tabla 1. Definición de variables y parámetros utilizados en el modelo.
Notation DefinitionNt Population size of focal plant species at time t.
Vt Pollinator visitation rate at time t.
Ct Number of conspecific pollen grains deposited by pollinators at time t.
Ht Number of heterospecific pollen grains deposited by pollinators at time t.
Pt Proportion of conspecific pollen grains deposited by pollinators at time t.
Qt Quality of conspecific pollen received at stigma at time t.
Ft Fecundity (fruit or seed production) of focal plant species at time t relative tomaximum attainable by the population.
ρ Relative density of focal plant species.
g Degree of generalization of pollinators.
i Degree of interference of fertilization by heterospecific pollen.
β fStrength of limitation of reproduction by pollen quantity.
βqStrength of decrease of pollen quality with decreasing population density.
K Carrying capacity.
s Degree of self-incompatibility.
b Intrinsic birth rate.
d Intrinsic death rate.
δ Herbivore-induced death rate.
Figure 2. Pollinator visitation frequency to the focal plant species as a function of population densitywhen pollinators are specialized (g=0.1) or generalized (g=0.9). Other parameter values: αv=500, βv
’=1000.Figura 2. Frecuencia de visitas de los polinizadores a la especie de planta focal en función de la densidadpoblacional cuando los polinizadores son especialistas (g=0.1) o generalistas (g=0.9). Valores de otrosparámetros: αv=500, βv
’=1000.
25

DP VÁZQUEZ Ecología Austral 17:21-36
Sección especial
on, but at low densities they may be unable toencounter them, so they may leave the patch.
Pollination
Deposition of conspecific and heterospecificpollen (Ct and Ht, respectively) are modelledas functions of both visitation rate and rela-tive abundance of the focal plant. Conspecificpollen deposition is described by
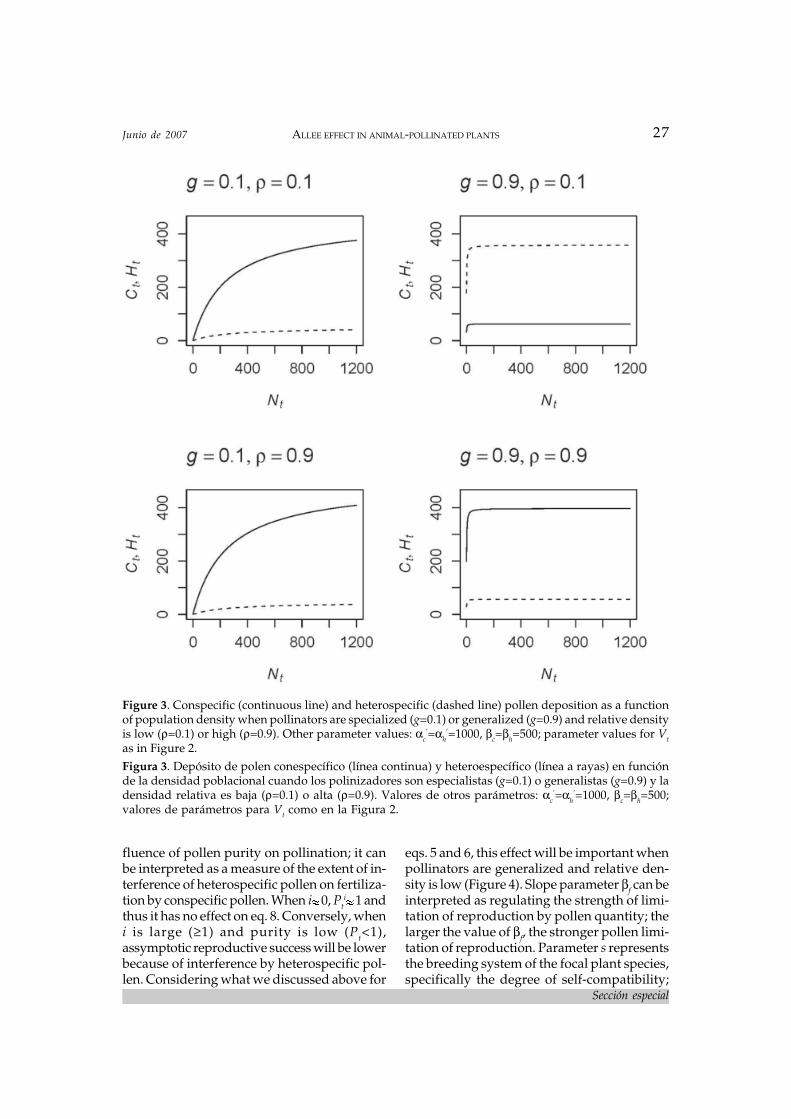
Ct= αcVt/βc+Vt , (5)
where αc and βc are saturation and slope pa-rameters as in eq. 4. Saturation point param-eter is defined as αc≡αc
’(1-g)1-ρ, whereρ=Nt/(Nt+Ot) is relative abundance (i.e., abun-dance of the focal plant Nt relative to all othercoflowering species in the community, Ot).Notice that because I do not model the dynam-ics of other species in the community, I con-sider ρ as a parameter rather than a variable,evaluating how different values of r influencethe behavior of the model. The influence of satu-ration point parameter αc is modulated by(1-g)1-ρ. Thus, when pollinators are specialized,g 0, (1-g)1-ρ 1 and αc αc
’, independently ofrelative abundance. In contrast, when polli-nators are generalists, g 1 (not exactly equalto 1 but slightly lower), and relative abundanceparameter ρ will have a strong influence onthe value of αc. At high relative abundances,ρ 1, (1-g)1-ρ 1 and αc αc
’. However, at low rela-tive abundances ρ will be close to 0 (but notexactly so) and αc<<αc
’. Thus, all else beingequal, when pollinators are generalized andrelative population density of the focal plantis low, the amount of conspecific pollen de-posited at stigmas will be substantially lowerthan when relative abundance is high and/orpollinators are specialized (Figure 3).
Likewise, heterospecific pollen deposition isgiven by
Ht= αhVt/βh+Vt , (6)
where αh and βh are parameters as in eq. 4.Saturation point parameter is defined asαh≡αh
’g(1-g)ρ. Here, when pollinators are spe-cialized, g 0 and thus αh 0, independentlyof relative abundance. Therefore, heterospe-
cific pollen deposition is always low when pol-linators are specialized. In contrast, when pol-linators are generalists, g 1 (not exactly equalto 1 but, slightly lower), and relative abun-dance parameter ρ will have a strong influ-ence on the value of αh. At high relative abun-dances, ρ 1, (1-g)ρ 0 and αh 0. Thus, again,heterospecific pollen deposition is low whenpollinators are generalized and the focal plantpopulation is at high relative density. In con-trast, at low relative abundances, ρ 0, (1-g)ρ 1and αh αh
’ (Figure 3). Thus, all else being equal,when pollinators are generalized and relativepopulation density of the focal plant speciesis low, the amount of heterspecific pollen de-posited on stigmas will be substantially higherthan when relative abundance is high and/orpollinators are specialized (Figure 3).
We can also use eqs. 5 and 6 to study thedensity dependence of pollen purity, definedas the proportion of conspecific pollen depos-ited at stigmas,
Pt=Ct/(Ct+Ht) . (7)
When pollinators are specialized, Pt is highthroughout the entire range of densities. Like-wise, when pollinators are generalized andrelative abundance is high, Pt is also high. Incontrast, when pollinators are generalized andrelative abundance is low, Pt is low (Figure 3).Thus, at low relative densities, generalist pol-linators are bringing a greater proportion ofheterospecific pollen. This is the quality com-ponent of pollination (see Rathcke 1983; Kunin1993, 1997a).
Reproduction
I model reproductive success as the numberof seeds produced per individual relative tothe maximum attainable by the population(i.e., relative fitness). This relative reproduc-tive success, Ft, is a function of the quantityand quality of pollen deposition:
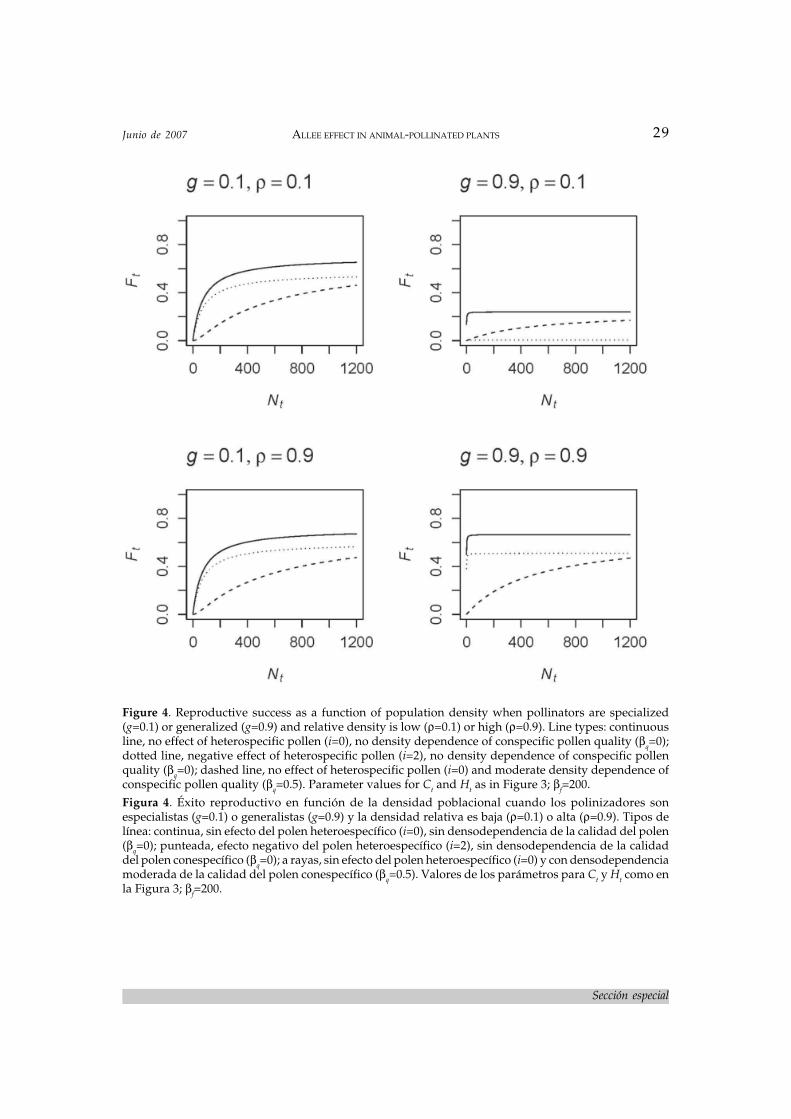
Ft=( αfCt/βf+Ct )s , (8)
where αf and βf are parameters as in previousequations. Saturation point parameter is de-fined as αf≡Pt
iQt. Parameter i controls the in-
26
Junio de 2007 ALLEE EFFECT IN ANIMAL-POLLINATED PLANTS
Sección especial
fluence of pollen purity on pollination; it canbe interpreted as a measure of the extent of in-terference of heterospecific pollen on fertiliza-tion by conspecific pollen. When i 0, Pt
i 1 andthus it has no effect on eq. 8. Conversely, wheni is large (≥1) and purity is low (Pt<1),assymptotic reproductive success will be lowerbecause of interference by heterospecific pol-len. Considering what we discussed above for
eqs. 5 and 6, this effect will be important whenpollinators are generalized and relative den-sity is low (Figure 4). Slope parameter βf can beinterpreted as regulating the strength of limi-tation of reproduction by pollen quantity; thelarger the value of βf, the stronger pollen limi-tation of reproduction. Parameter s representsthe breeding system of the focal plant species,specifically the degree of self-compatibility;
Figure 3. Conspecific (continuous line) and heterospecific (dashed line) pollen deposition as a functionof population density when pollinators are specialized (g=0.1) or generalized (g=0.9) and relative densityis low (ρ=0.1) or high (ρ=0.9). Other parameter values: αc
’=αh’=1000, βc=βh=500; parameter values for Vt
as in Figure 2.Figura 3. Depósito de polen conespecífico (línea continua) y heteroespecífico (línea a rayas) en funciónde la densidad poblacional cuando los polinizadores son especialistas (g=0.1) o generalistas (g=0.9) y ladensidad relativa es baja (ρ=0.1) o alta (ρ=0.9). Valores de otros parámetros: αc
’=αh’=1000, βc=βh=500;
valores de parámetros para Vt como en la Figura 2.
27

DP VÁZQUEZ Ecología Austral 17:21-36
Sección especial
when s=1 the plant is strictly self incompat-ible (or dioecious) and reproductive successdepends strongly on pollen deposition; on theother extreme of the spectrum, when s=0 theplant is apomictic and reproductive success isindependent of pollen deposition. For simplic-ity, I assume s=1 throughout the paper. Finally,because not only pollination quantity but alsoquality may determine reproduction (Aizen &Harder 2007), I include a component Qt thatcontrols the quality of conspecific pollen andis itself a function of density
Qt= Xt/βq+Xt , (9)
where Xt=Nt/K, and βq is a saturation param-eter as in previous equations. Thus, qualityvaries between 0 at low densites to 1 at highdensities in a saturating fashion, so that lowdensities mean more self pollination and thuslower quality of conspecific pollen. When βq=0quality of conspecific pollen is always high(i.e., Qt=1) regardless of population density;when βq>0 quality of pollen decreases at lowabsolute densities of the focal plant species(Figure 4).
Population dynamics
Finally, I model population dynamics as
Nt+1=Nt+bNtFt(1- Nt/K )-dNt , (10)
and per capita growth rate as
Nt+1/Nt =1+bFt(1- Nt/K )-d . (11)
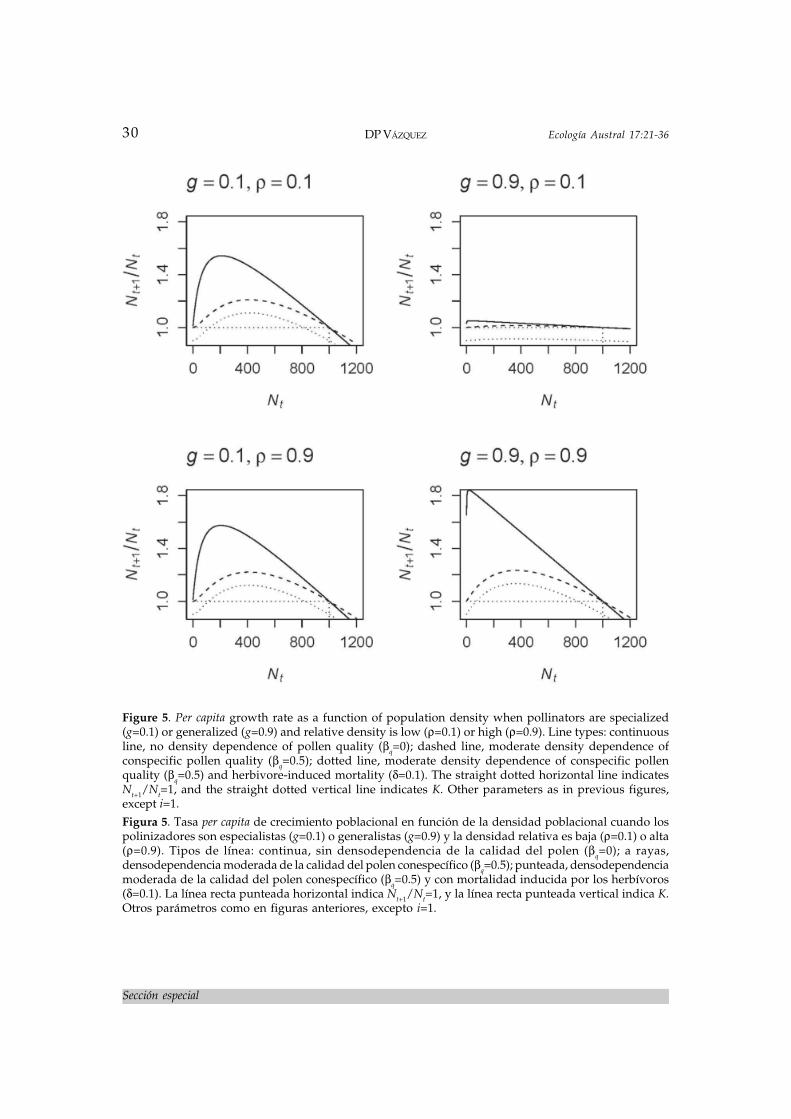
where b is the intrinsic birth rate and d is theintrinsic death rate (i.e., I have decomposedthe intrinsinc population growth rate, r, intoits birth and death components). Here, Ft (eq.8) produces the mechanistic Allee effect wewanted to model (Figure 5). As before, the in-fluence of Ft on per capita growth rate dependson the degree of pollinator specialization andthe relative abundance of the focal plant popu-lation (Figure 5). It also depends on the strengthof density dependence of pollen quality (Qt,eq. 9). When there is no density dependence ofpollen quality (continuous line in Figure 5) andpollinators are generalists, there is virtuallyno Allee effect, and per capita growth rate is
high at low densities and decreases linearlywith increasing density. Conversely, whenthere is density dependence of pollen quality(dashed line in Figure 5) and pollinators aregeneralists, there is a stronger Allee effect, sothat per capita growth rate increases at low den-sities up to a maximum and then decreasesgradually as density continues to increase.When pollinators are specialists, there is al-ways an Allee effect, but per capita growth rateis always higher when there is no density de-pendence of pollen quality.
It is important to point out here that manystudies of density dependence of reproductionand growth rate in animal-pollinated plants(e.g., Kunin 1993; Aizen & Feinsinger 1994a;Kunin 1997b; Aguilar & Galetto 2004; Vázquez& Simberloff 2004; see Ghazoul 2005 for re-view) estimate seed production as a measureof plant reproductive success. However, be-cause pollination quality can affect seed vi-ability and vigor even if it does not affect seedproduction, these studies may be missing theAllee effects for reproductive success and percapita growth rate shown in Figures 4 and 5,wrongly concluding that there is no effect ofdensity on reproductive success.
Adding herbivore-induced mortality
We can add the effect of herbivores to themodel by incorporating a term that removes aportion of the population due to herbivore-in-duced mortality:
Nt+1=Nt+bFtNt(1- Nt/K )-dNt-δNt . (12)
So now per capita growth rate becomes
Nt+1/Nt =1+bFt(1- Nt/K )-(d+δ) , (13)
where δ is the proportion of the population thatis removed by herbivores. The addition of her-bivore-induced mortality results in per capitagrowth rate now exhibiting a higher thresh-old density, under which the population can-not replace itself and goes extinct; the equilib-rium density is also lower than K, and themaximum per capita growth rate is substan-tially lower than in the absence of herbivores(dotted line in Figure 5).
28
Junio de 2007 ALLEE EFFECT IN ANIMAL-POLLINATED PLANTS
Sección especial
Figure 4. Reproductive success as a function of population density when pollinators are specialized(g=0.1) or generalized (g=0.9) and relative density is low (ρ=0.1) or high (ρ=0.9). Line types: continuousline, no effect of heterospecific pollen (i=0), no density dependence of conspecific pollen quality (βq=0);dotted line, negative effect of heterospecific pollen (i=2), no density dependence of conspecific pollenquality (βq=0); dashed line, no effect of heterospecific pollen (i=0) and moderate density dependence ofconspecific pollen quality (βq=0.5). Parameter values for Ct and Ht as in Figure 3; βf=200.Figura 4. Éxito reproductivo en función de la densidad poblacional cuando los polinizadores sonespecialistas (g=0.1) o generalistas (g=0.9) y la densidad relativa es baja (ρ=0.1) o alta (ρ=0.9). Tipos delínea: continua, sin efecto del polen heteroespecífico (i=0), sin densodependencia de la calidad del polen(βq=0); punteada, efecto negativo del polen heteroespecífico (i=2), sin densodependencia de la calidaddel polen conespecífico (βq=0); a rayas, sin efecto del polen heteroespecífico (i=0) y con densodependenciamoderada de la calidad del polen conespecífico (βq=0.5). Valores de los parámetros para Ct y Ht como enla Figura 3; βf=200.
29
DP VÁZQUEZ Ecología Austral 17:21-36
Sección especial
Figure 5. Per capita growth rate as a function of population density when pollinators are specialized(g=0.1) or generalized (g=0.9) and relative density is low (ρ=0.1) or high (ρ=0.9). Line types: continuousline, no density dependence of pollen quality (βq=0); dashed line, moderate density dependence ofconspecific pollen quality (βq=0.5); dotted line, moderate density dependence of conspecific pollenquality (βq=0.5) and herbivore-induced mortality (δ=0.1). The straight dotted horizontal line indicatesNt+1/Nt=1, and the straight dotted vertical line indicates K. Other parameters as in previous figures,except i=1.Figura 5. Tasa per capita de crecimiento poblacional en función de la densidad poblacional cuando lospolinizadores son especialistas (g=0.1) o generalistas (g=0.9) y la densidad relativa es baja (ρ=0.1) o alta(ρ=0.9). Tipos de línea: continua, sin densodependencia de la calidad del polen (βq=0); a rayas,densodependencia moderada de la calidad del polen conespecífico (βq=0.5); punteada, densodependenciamoderada de la calidad del polen conespecífico (βq=0.5) y con mortalidad inducida por los herbívoros(δ=0.1). La línea recta punteada horizontal indica Nt+1/Nt=1, y la línea recta punteada vertical indica K.Otros parámetros como en figuras anteriores, excepto i=1.
30

Junio de 2007 ALLEE EFFECT IN ANIMAL-POLLINATED PLANTS
Sección especial
Notice, however, that the threshold is notequally high in all cases (Figure 5). When pol-linators are specialized, the threshold forpopulation persistence is relatively high. Thisthreshold increases with the severity of the ef-fect of herbivores. In contrast, when pollina-tors are generalized and there is no densitydependence of pollen quality, there is virtu-ally no threshold for population persistence.Finally, when there is density dependence ofpollen quality (βq>0) there is a threshold evenif pollinators are generalists. This findingsheds some light on Ghazoul’s (2005) conjec-ture, “the existence of thresholds remains un-certain for most species, but where they occurthey will probably be mediated by the type ofpollinator involved”. The present model sug-gests that degree of pollinator specializationon the plant can determine the severity of suchthreshold.
In the next section I illustrate the use of themodel by applying it to two case studies.
Case studies
I consider two case studies that have ad-dressed the density-mediated effects of intro-duced herbivores on plant pollination and re-production. These two case studies are con-trasting in terms of the details of the ecology ofplant-pollinator interactions, and serve toshow how the model can be used to generatemechanistic predictions about the effects ofintroduced herbivores.
Alstroemeria aurea in Nahuel Huapi
The first case study I will discuss isAlstroemeria aurea (Alstroemeriaceae) inNahuel Huapi National Park, in the temper-ate forest of the southern Argentine Andes. A.aurea is a summer-flowering perennial herbfrom this region. It dominates the understoryin some forest types (Aizen 1997; 2001;Vázquez & Simberloff 2004). Although it is selfcompatible, pollination and seed productionare pollinator-dependent because of the plant’spronounced protandry (Aizen & Basilio 1995).Its main pollinator is the native bumblebeeBombus dahlbomii and, increasingly, the exotic
bumblebee B. ruderatus (Aizen 2001; Vázquez& Simberloff 2002; Morales 2007); both bumble-bee species are highly generalized (Morales &Aizen 2002; Vázquez & Simberloff 2002). A.aurea is highly susceptible to trampling byungulates, and both its absolute and relativedensities decrease sharply in native forestsites with cattle or deer, compared to ungu-late-free sites (Vázquez & Simberloff 2004). Thiseffect on density does not have a detectableeffect on pollinator visitation frequency, but itdoes affect pollination, with decreased depo-sition of conspecific pollen and increaseddeposition of heterospecific pollen in grazedsites (pollinators appear to bring a lower pro-portion of conspecific pollen grains per visit);this effect on pollination in turn results in de-creased reproductive success in the presenceof herbivores (Vázquez & Simberloff 2004).
Let’s now parameterize the model, to seewhat it can tell us about the effect of cattle onA. aurea. The parameter values I will use arenot quantitative estimates, but only “qualita-tive guesses” based on what we know aboutthis species; but these guesses will be enoughto allow us to make some qualitative predic-tions. First, given it is pollinated by highly gen-eralized pollinators, we set g=0.9. There is onlyweak density dependence of pollen quality,with high levels of heterozigocity a few metersaway from individual ramets (Souto et al.2002); so we set βq=0.1. We don’t know howmuch heterospecific pollen grains interferewith pollination; I assume one heterospecificpollen grain excludes one conspecific grain,and so we set i=1. We know cattle affects thisspecies disproportionately to other species; sowe will compare a situation of high relativeabundance in the absence of cattle (ρ=0.9) withone with moderate relative abundance whencattle are present (ρ=0.5). Finally, we assumethat herbivores kill 10% of the population whenit is at K; thus, δ=0.1 in the presence of cattle,and δ=0 in the absence of them.
The model tells us that when the focal plantspecies is pollinated by generalists, decreasedabsolute density will not lead to decreasedpollinator visitation (Figure 2). It also tells usthat if relative abundance decreases, deposi-tion of conspecific pollen will decrease andthat of heterospecific pollen will increase (Fig-
31

DP VÁZQUEZ Ecología Austral 17:21-36
Sección especial
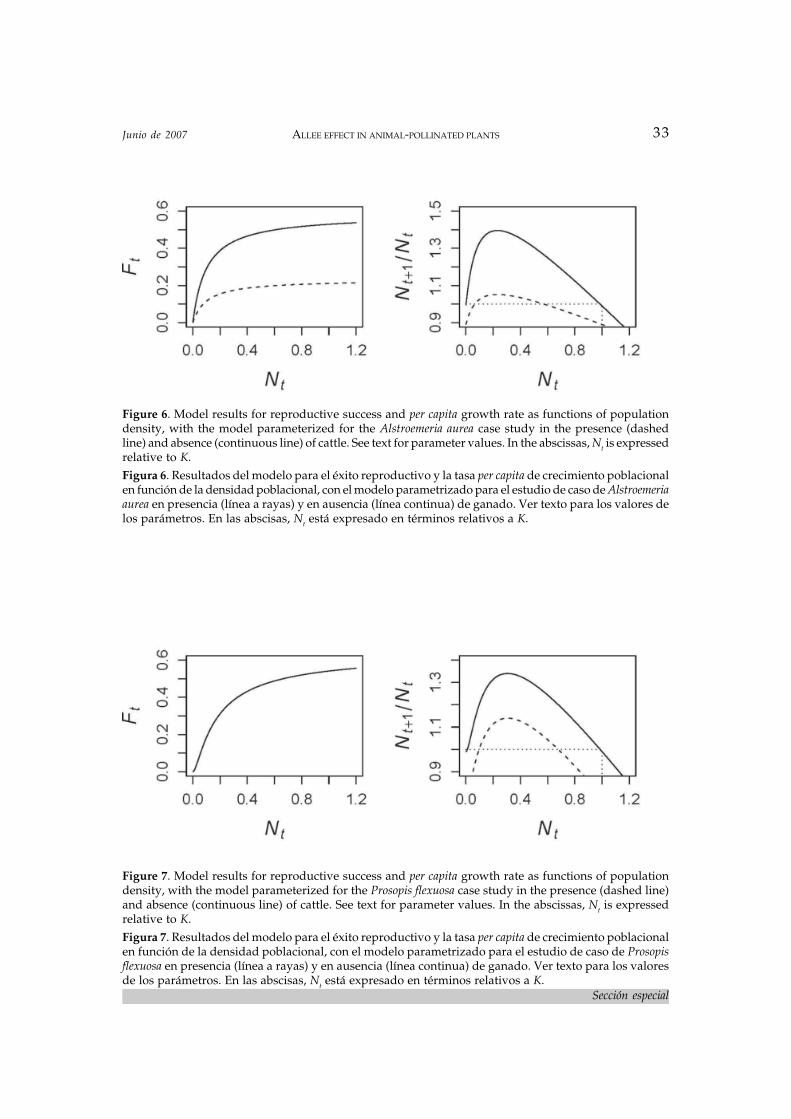
ure 3), leading to decreased pollen purity andthus lower pollination quality. Lower pollina-tion quality in turn results in lower reproduc-tive performance (Figure 4). All this agrees withthe findings of Vázquez and Simberloff (2004)described above. The model also allows us tomake a prediction about what would happenwith per capita growth rate; although we haveno estimates of intrinsic birth rates b and d forthis species and thus won’t be able to contrastthe model with reality, qualitative predictionswill be unaffected by the specific value of theseparameters used. Per capita growth rate willexhibit a threshold density in the presence ofcattle, under which the population cannot re-place itself and goes extinct (Figure 6). The equi-librium density will be lower than K, and themaximum per capita growth rate will be sub-stantially lower than in the absence of distur-bance. The existence of a threshold for per capitagrowth rate suggests that populations of A.aurea in areas with cattle may be doomed tolocal extinction, even though there may still besome seed production. The likelihood of thisscenario will depend on the severity of thethreshold and the strength of other factors notconsidered in the model that could affect smallpopulations, such as demographic stochasti-city or inbreeding depression (Lande 1988).
Prosopis flexuosa in Ñacuñán
The second case study contrasts with the pre-vious one in some crucial aspects of the polli-nation ecology of the focal species. Prosopisflexuosa (Leguminosae) is the dominant treespecies in the Central Monte Desert ofMendoza, Argentina; it is functionally impor-tant in this ecosystem (Campos & Ojeda 1997;Rossi & Villagra 2003; Milesi & López deCasenave 2004). It represents an importantsource of wood and its pods are a rich foodsource for both man or cattle. Its wood has beenoverexploited during the first half of the 20thcentury. It is self-incompatible (Masuelli &Balboa 1989), and is pollinated by several in-sect species that are mostly restricted to P.flexuosa during its flowering season (thus be-having as functionally specialized; V. Ascheroand D. P. Vázquez, unpubl. data). The perim-eter of Ñacuñán Biosphere Reserve was fencedin the early 1970s, thus excluding cattle ranch-
ing and wood and pod harvesting; althoughwood harvesting is currently banned by pro-vincial authorities, ranching and pod harvest-ing continue to date in the private land sur-rounding the reserve. Thus although adulttrees are protected, human activities still havethe potential to affect seedling and saplingsurvivorship and adult fecundity. Working insix pairs of 1 ha plots inside and outside thereserve, V. Aschero and I (unpubl. ms.) havefound that absolute density of P. flexuosa, pol-linator visitation frequency and seed produc-tion all tend to be lower outside the reserve.
As in the previous case study, to parameter-ize the model we need to make some guesses,but that will not affect the general qualitativepredictions. First, based on what we knowabout the flower visitors, we will assume thepollinator is relatively specialized; so we setg=0.1. Because pollinators are specialized, thevalue of parameter i (heterospecific pollen in-terference) is irrelevant (see eq. 8); so we set itagain at the arbitrary value of 1. We also as-sume there is a moderate density dependenceof pollen quality; thus βq=0.1. Again becausethe pollinator is relatively specialized, its rela-tive density is irrelevant (see eqs. 5 and 6); sowe set r at the arbitrary value of 0.5. Finally,we assume that disturbance kills 20% of thepopulation when it is at K; thus, δ=0.2 in thepresence of cattle and δ=0 in the absence ofthem. Per capita growth rate again exhibits athreshold density, under which the popula-tion cannot replace itself and goes extinct, andthe equilibrium density is also lower than K(Figure 7). However, in this case the effect re-sults from entirely different mechanisms thanthe previous case study: it results solely fromthe increased mortality caused by cattle, notby a change in the functional relationship be-tween Ft and Nt; in fact, this relationship isunchanged by cattle. The presence of cattlesimply keeps the population at a lower den-sity, and this lower density results in lowervisitation frequency, pollination quantity, repro-ductive success and lower per capita growth rate.
So what?
You may be asking yourself: so what? Argu-ably, many of the qualitative predictions com-
32
Junio de 2007 ALLEE EFFECT IN ANIMAL-POLLINATED PLANTS
Sección especial
Figure 6. Model results for reproductive success and per capita growth rate as functions of populationdensity, with the model parameterized for the Alstroemeria aurea case study in the presence (dashedline) and absence (continuous line) of cattle. See text for parameter values. In the abscissas, Nt is expressedrelative to K.Figura 6. Resultados del modelo para el éxito reproductivo y la tasa per capita de crecimiento poblacionalen función de la densidad poblacional, con el modelo parametrizado para el estudio de caso de Alstroemeriaaurea en presencia (línea a rayas) y en ausencia (línea continua) de ganado. Ver texto para los valores delos parámetros. En las abscisas, Nt está expresado en términos relativos a K.
Figure 7. Model results for reproductive success and per capita growth rate as functions of populationdensity, with the model parameterized for the Prosopis flexuosa case study in the presence (dashed line)and absence (continuous line) of cattle. See text for parameter values. In the abscissas, Nt is expressedrelative to K.Figura 7. Resultados del modelo para el éxito reproductivo y la tasa per capita de crecimiento poblacionalen función de la densidad poblacional, con el modelo parametrizado para el estudio de caso de Prosopisflexuosa en presencia (línea a rayas) y en ausencia (línea continua) de ganado. Ver texto para los valoresde los parámetros. En las abscisas, Nt está expresado en términos relativos a K.
33

DP VÁZQUEZ Ecología Austral 17:21-36
Sección especial
ing from the model I have developed abovecould be made without it (and actually hadbeen made repeatedly in the literature). How-ever, putting together all the informationneeded to make such predictions is not simple,and the model can be useful as a tool to for-malize them. How strong will be the Allee ef-fect generated by introduced herbivores on aparticular plant species if, for example, theplant is pollinated by a generalized pollina-tor, is self-compatible, there is no density de-pendence of conspecific pollen quality, and itsrelative abundance decreases sharply in thepresence of herbivores? What if, all else beingequal to the previous case, there is now mod-erate density dependence of conspecific pol-len quality and relatively abundance does notchange? The model can aid us to investigatethese scenarios and provide testable predic-tions that can be evaluated in field observa-tional or experimental studies.
Predicting Allee effects of introduced herbi-vores on animal-pollinated plants requiresknowledge on several aspects of the pollina-tion ecology and reproductive biology of thefocal plant species, some of which are not avail-able in the case studies discussed above (thisapplies, in fact, to the majority of studies ofAllee effects in animal-pollinated plants). Forexample, we need not only knowledge of therichness and relative importance of pollina-tors, but also of the degree of specialization ofpollinators on the focal plant species, some-thing that is not available in many studies (seealso Ashworth et al. 2004). In cases when pol-linators are not specialized on the focal plantspecies, we also need to know what happenswith both the absolute and relative densitiesof the focal plant population. Likewise, weshould not only evaluate reproductive successas seed production but also in terms of seedviability an germination potential. The modeldeveloped here could be used to generate achecklist of the variables to be measured instudies of Allee effects in animal-pollinatedplants.
As I argued in the introduction, this modelcould be applied to other types of perturba-tions besides introduced herbivores that oper-ate through increased mortality. This kind of
modelling approach can thus provide a quan-titative framework that complements the con-ceptual framework proposed recently by Aizen& Vázquez (2006). Together, these tools can beuseful guides for the design of process andmechanism-oriented studies of the effects ofanthropogenic habitat disturbance on plantpollination and reproduction.
ACKNOWLEDGMENTS
I thank the editors of this special section forinviting me to write this paper. I am a careerresearcher with CONICET, and was supportedby grants from BBVA Foundation (BIOCON03-162) and FONCyT-ANAPCyT (PICT 16-20805). I thank my colleagues V. Aschero andD. Simberloff, with whom I have conductedthe case studies discussed here. N. Chacoff, J.Dorado, J. M. Morales, B. Morris and an anony-mous reviewer made useful comments on themanuscript.
REFERENCES
AGUILAR, R & L GALETTO. 2004. Effects of forestfragmentation on male and female reproductivesuccess in Cestrum parqui (Solanaceae). Oecologia,138:513–520.
AIZEN, MA. 1997. Influence of local floral densityand sex ratio on pollen receipt and seed output:empirical and experimental results indichogamous Alstroemeria aurea (Alstroeme-riaceae). Oecologia, 111: 404–412.
AIZEN, MA. 2001. Flower sex ratio, pollinatorabundance, and the seasonal pollination dyna-mics of a protandrous plant. Ecology, 82: 127–144.
AIZEN, MA & A BASILIO. 1995. Within and amongflower sex-phase distribution in Alstroemeriaaurea (Alstroemeriaceae). Canadian Journal ofBotany, 73:1986–1994.
AIZEN, MA & P FEINSINGER. 1994a. Forest fragmen-tation, pollination, and plant reproduction in aChaco dry forest, Argentina. Ecology, 75: 330–351.
AIZEN, MA & P FEINSINGER. 1994b. Habitatfragmentation, native insect pollinators, and feralhoney bees in Argentine Chaco Serrano.Ecological Applications, 4:378–392.
AIZEN, MA & LD HARDER. 2007. Expanding the limitsof the pollen-limitation concept: effects of pollenquantity and quality. Ecology, 88:271–281.
34

Junio de 2007 ALLEE EFFECT IN ANIMAL-POLLINATED PLANTS
Sección especial
AIZEN, MA & DP VÁZQUEZ. 2006. Flower perfor-mance in human-altered habitats. Pp. 159–179in: LD Harder & SCH Barrett (eds). Ecology andEvolution of Flowers. Oxford University Press.
ALLEE, WC; AE EMERSON & O PARK. 1949. Princples ofAnimal Ecology. W.B. Saunders Co., Philadelphia.
ASHWORTH, L; R AGUILAR; L GALETTO & MA AIZEN.2004. Why do pollination generalist and specialistplant species show similar reproductivesusceptibility to habitat fragmentation? Journalof Ecology, 92:717–719.
BOND, WJ. 1994. Do mutualisms matter? Assessingthe impact of pollinator and disperser disruptionon plant extinction. Philosophical Transactions ofthe Royal Society of London Series B, 344:83–90.
CAMPOS, CM & RA OJEDA. 1997. Dispersal andgermination of Prosopis flexuosa (Fabaceae) seedsby desert mammals in Argentina. Journal of AridEnvironments, 35:707–714.
CASWELL, H. 2001. Matrix Population Models:Construction, Analysis, and Interpretation. SinauerAssociates, Suntherland, MA.
COURCHAMP, F; T CLUTTON-BROCK & B GRENFELL. 1999.Inverse density dependence and the Allee effect.Trends in Ecology & Evolution, 14:405–410.
CRAWLEY, MJ (ed.). 1997. Plant Ecology. Second ed.Blackwell Science, Oxford.
DARWIN, C. 1859. The Origin of Species by Means ofNatural Selection. Bantam, New York, U.S.A.
DE VOS, A; RH MANVILLE & RG VAN GELDER. 1956.Introduced mammals and their influence onnative biota. Zoologica (New York Zoological Society),41:163–194.
DENNIS, B. 1989. Allee effects: population growth,critical density, and the chance of extinction.Natural Resource Modeling, 3:481–538.
EBENHARD, T. 1988. Introduced birds and mammalsand their ecological effects. Swedish WildlifeResearch (Viltrevy), 13:1–107.
GHAZOUL, J. 2005. Pollen and seed dispersal amongdispersed plants. Biological Reviews, 80:1–31.
HIXON, MA; SW PACALA & SA SANDIN. 2002.Population regulation: historical context andcontemporary challenges of open vs. closedsystems. Ecology, 83:1490–1508.
KOT, M. 2001. Elements of Mathematical Ecology.Cambridge University Press.
KUNIN, W & Y IWASA. 1996. Pollinator foragingstrategies in mixed floral arrays: density effectsand floral constancy. Theoretical Population Biology,49:232–263.
KUNIN, WE. 1993. Sex and the single mustard:population density and pollinator behavior effectson seed-set. Ecology, 74:2145–2160.
KUNIN, WE. 1997a. Population biology of rarity:on the complexity of density dependence ininsect-plant interactions. Pp. 150–173 in: WEKunin & KJ Gaston (eds). The Biology of Rarity.Chapman and Hall, London.
KUNIN, WE. 1997b. Population size and densityeffects in pollination: pollinator foraging andplant reproductive success in experimental arraysof Brassica kaber. Journal of Ecology, 85:225–234.
LANDE, R. 1988. Genetics and demography inbiological conservation. Science, 241:1455–1459.
LAWTON, JH. 1999. Are there general laws inecology? Oikos, 84:177–192.
MALTHUS, TR. 1798. An essay on the Principle ofPopulation, as it affects the future improvement ofsociety, with remarks on the speculations of Mr.Godwin, M. Concorcet, and other writers. J. Johnson.
MASUELLI, R & O BALBOA. 1989. Self-incompatibilityin Prosopis flexuosa D.C. Plant Cell ImcompatibilityNewsletter, 21: 44–48.
MCCARTHY, M. 1997. The Allee effect, finding matesand theoretical models. Ecological Modelling,103:99–102.
MILESI, FA & J LÓPEZ DE CASENAVE. 2004. Unexpectedrelationships and valuable mistakes: nonmyrme-cochorous Prosopis dispersed by messy leaf-cutting ants in harvesting their seeds. AustralEcology, 29: 558–567.
MORALES, CL. 2007. Introducción de abejorros(Bombus) no nativos: causas, consecuencias ecoló-gicas y perspectivas. Ecología Austral, 17: 51-65.
MORALES, CL & MA AIZEN. 2002. Does invasion ofexotic plants promote invasion of exotic flowervisitors? A case study from the temperate forestsof the southern Andes. Biological Invasions,4: 87–100.
MORRIS, WF. 2003. Which mutualists are mostessential? Buffering of plant reproduction againstthe extinction of pollinators. Pp. 260–280 in: PKareiva & SA Levin (eds). The Importance ofSpecies: Perspectives on Expendability and Triage.Princeton University Press, Princeton.
RATHCKE, B. 1983. Competition and facilitiationamong plants for pollination. Pp. 305–329 in: L Real(ed). Pollination Biology. Academic Press, London.
ROSSI, BE & PE VILLAGRA. 2003. Effects of Prosopisflexuosa on soil properties and the spatial patternof understorey species in arid Argentina. Journalof Vegetation Science, 14:543–550. Article.
SOUTO, CP; MA AIZEN & AC PREMOLI. 2002. Effectsof crossing distance and genetic relatedness onpollen performance in Alstroemeria aurea(Alstroemeriaceae). American Journal of Botany,89:427–432.
STEPHENS, PA & WJ SUTHERLAND. 1999. Consequences
35

DP VÁZQUEZ Ecología Austral 17:21-36
Sección especial
of the Allee effect for behaviour, ecology andconservation. Trends in Ecology & Evolution,14:401–405.
TRAVESET, A & DM RICHARDSON. 2006. Biological inva-sions as disruptors of plant reproductive mutua-lisms. Trends in Ecology & Evolution, 21: 208–216.
TURCHIN, P. 2001. Does population ecology havegeneral laws? Oikos, 94:17–26.
VERHULST, PF. 1845. Recherches mathématiques surla loi d’accroissement de la population. NouveauxMémoires de l’Académie Royale des Sciences et Belles-Lettres de Bruxelles, 18:3–38.
VÁZQUEZ, DP. 2002. Multiple effects of introducedmammalian herbivores in a temperate forest.Biological Invasions, 4:175–191.
VÁZQUEZ, DP; WF MORRIS & P JORDANO. 2005.
Interaction frequency as a surrogate for the totaleffect of animal mutualists on plants. EcologyLetters, 8:1088–1094.
VÁZQUEZ, DP & D SIMBERLOFF. 2002. Ecologicalspecialization and susceptibility to disturbance:conjectures and refutations. American Naturalist,159:606–623.
VÁZQUEZ, DP & D SIMBERLOFF. 2004. Indirect effectsof an introduced ungulate on pollination andreproduction. Ecological Monographs, 74:281–308.
WASER, NM; L CHITTKA; MV PRICE; NM WILLIAMS & JOLLERTON. 1996. Generalization in pollination sys-tems, and why it matters. Ecology, 77:1043–1060.
WASER, NM & J OLLERTON (eds.). 2006. Plant–Pollinator Interactions: From Specialization toGeneralization. University of Chicago Press.
36