intestinal inflammation caused by magnesium deficiency alters basal and oxidative stress-induced...
TRANSCRIPT

Intestinal inflammation caused by magnesium deficiency altersbasal and oxidative stress-induced intestinal function
Bradford J. Scanlan Æ Blaine Tuft Æ Justin E. Elfrey ÆAllen Smith Æ Aiping Zhao Æ Motoko Morimoto ÆJoanna J. Chmielinska Æ Maria Isabel Tejero-Taldo ÆIu Tong Mak Æ William B. Weglicki Æ Terez Shea-Donohue
Received: 13 April 2007 / Accepted: 12 July 2007 / Published online: 27 July 2007
� Springer Science+Business Media, LLC 2007
Abstract The aim of this study was to determine the
effect of magnesium deficiency on small intestinal mor-
phology and function. Rats were assigned to 4 groups and
placed on magnesium sufficient or deficient diet for 1 or
3 weeks. Infiltration of neutrophils and mucosal injury
were assessed in stained sections of small intestine. Mag-
nesium deficiency alone induced a significant increase in
neutrophil infiltration and increased vascular ICAM-1
expression, in the absence of changes in mucosal injury or
expression of proinflammatory mediators. Magnesium
deficiency was associated with hyposecretory epithelial
cell responses and vascular macromolecular leak in the
small intestine and lung, which was attributed partly to
reduced expression of NOS-3. To determine the effect of
hypomagnesmia on the intestinal responses to a known
oxidative stress, groups of rats were randomized to either
sham operation or superior mesenteric artery occlusion for
10 (non-injurious) or 30 (injurious) minutes followed by a
1- or 4-hour reperfusion period. In response to mesenteric
ischemia/reperfusion, deficient rats showed exaggerated
PMN influx, but similar mucosal injury. Intestinal ischemia
in sufficient animals induced vascular macromolecular leak
in the small intestine and lung at 4 hours of reperfusion,
with levels similar to those observed in untreated deficient
rats. Acute magnesium repletion of deficient rats 24 h
before surgery attenuated the exaggerated inflammation in
deficient rats. These data show that magnesium deficiency
induced a subclinical inflammation in the small intestine in
the absence of mucosal injury, but with significant func-
tional changes in local and remote organs and increased
sensitivity to oxidative stress.
Keywords Hypomagnesemia � Oxidative stress �Neutrophil � Inflammation � Nitric oxide
Introduction
Hypomagnesemia is encountered frequently in hospitalized
patients and is seen most often in patients admitted to
intensive care units and is a primary cause of significant
morbidity and mortality [1]. Clinically, magnesium defi-
ciency (MgD) occurs often as a consequence of restricted
dietary intake (e.g. alcoholism), Mg2+-wasting drug thera-
pies, or diseases that impair intestinal absorption including
inflammatory bowel disease and celiac disease [2]. There is
The opinions contained herein are those of the authors and are not to
be construed as official policy or reflecting the views of the
Department of Defense
B. J. Scanlan
Department of Surgery, Walter Reed Army Medical Center,
Washington, DC 20307, USA
B. Tuft
Department of Pediatrics, Walter Reed Army Medical Center,
Washington, DC 20307, USA
J. E. Elfrey � A. Zhao � M. Morimoto � T. Shea-Donohue (&)
Department of Medicine & The Mucosal Biology Research
Center, University of Maryland, School of Medicine, Baltimore,
MD 21201, USA
e-mail: [email protected]
A. Smith
Beltsville Human Nutrition Research Center, Nutrient
Requirements and Functions Laboratory, ARS, USDA,
Beltsville, MD 20705, USA
J. J. Chmielinska � M. I. Tejero-Taldo � I. T. Mak �W. B. Weglicki
Division of Experimental Medicine, Department of
Biochemistry and Molecular Biology, George Washington
University Medical Center, Washington, DC 20037, USA
123
Mol Cell Biochem (2007) 306:59–69
DOI 10.1007/s11010-007-9554-y

a renewed interest in Mg2+ since deficiency is proposed to
be a novel factor in the pathogenesis of the complications
of diabetes. In diabetes, Mg2+ is not only an outcome, but is
now considered also to be a potential risk factor in the
actual development of the disease [3]. Cardiac and other
surgeries are associated with an increased incidence of
MgD. Indeed, the cardiovascular complications associated
with MgD are often the driving force behind Mg2+ reple-
tion in hospitalized patients [4]. In rodents, dietary
restriction of Mg2+ for 3 weeks was associated with the
production of reactive oxygen species and the development
of cardiomyopathy that may result from a synergism or
cascade of inflammatory events expressed by these tissues
[5–8]. The effects of MgD on cardiac pathology in these
animal models are well-documented, yet there is little
information on the impact of MgD on the intestine.
Tissue injury due to ischemia and subsequent reperfu-
sion events (IR) is a common event associated with mul-
tiple clinical pathologies. The mesenteric circulation is
especially sensitive to systemic hypoperfusion and while
hypoxia alone is detrimental, reperfusion of the affected
area dramatically exacerbates the damage [9]. Significant
neutrophil (PMN) infiltration and mucosal injury are clas-
sic features of the oxidative stress induced by mesenteric
IR [10–13]. The recruitment of leukocytes to the affected
tissue is a critical step in the inflammatory response and
involves a complex series of adhesive interactions and
signaling events [13–15]. Leukocytes also play a prominent
role in the events that link the post-ischemic gut to acute
lung injury, the earliest manifestation of ARDS. The
recruitment of leukocytes into an area of inflammation
begins with the activation of PMNs and/or endothelial
cells, followed by the bonding of white blood cells to the
endothelium and their subsequent transmigration into tis-
sues [16]. These processes are governed by the interaction
between adhesion molecules on the surface of both
leukocytes (CD11/CD18, L-selectin) and endothelial cells
(ICAM-1, E-selectin, P-selectin) [17].
The aims of the present study, therefore, were to deter-
mine (1) the effect of MgD on intestinal morphology and
mucosal function; (2) the effect of MgD on the intestinal
responses to an oxidative stress; and (3) the impact of acute
magnesium repletion (MgR) on MgD-induced changes in
the basal state and in response to an oxidative stress.
Materials and methods
Animal preparation
All experiments were conducted in accordance to the
principles for the care and use of laboratory animals as
recommended by the US Department of Health and Human
Services and approved by the Institutional Animal Care
and Use Committee. Male Sprague–Dawley rats (250–
350 gm) were kept under 12:12 h light:dark cycle, food
and distilled water were provided ad libitum. Animals were
fed either a modified diet (Teklad Laboratory, Madison,
WI) containing 2 mmole Mg2+/kg diet (Mg2+ -deficient
group [MgD], 9% Recommended Daily Allowance
[RDA]); or the same diet supplemented with 25 mmoles of
magnesium/diet (Mg2+-sufficient group [MgS], 100%
RDA). Rats were fed the MgD or MgS diet for 1 or
3 weeks.
On the day of study, rats from both diet groups were
anesthetized with ketamine (80 mg/kg im) and xylazine
(16 mg/kg im), weighed, and randomly assigned to one of
the following treatment groups: sham operation (sham),
10 min of ischemia followed by 1 h of reperfusion (IR-10/
1), or 30 min of ischemia followed by 1 h of reperfusion
(IR-30/1). Groups of rats fed the MgD or MgS diet for
3 weeks were subjected also to 30 min of ischemia
followed by 4 h of reperfusion (IR-30/4).
In all animals, a midline laparotomy was performed and
the superior mesenteric artery (SMA) was carefully
isolated as it originated from the abdominal aorta. After a
30-min stabilization period, the animals underwent their
assigned treatment. Prior to euthanasia samples of jejunum,
10 cms from the ligament of Treitz, were harvested for
microscopic evaluation. Serum levels of Mg2+, calcium
(Ca2+), and potassium (K+) were measured by atomic
absorption spectroscopy. Sham operations, consisting of a
midline laparotomy without occlusion of the SMA, were
performed in all dietary groups.
Measurement of epithelial cell function
Four 1 cm segments of mucosa were stripped of muscle
and mounted in Ussing chambers that exposed 0.126 cm2
to 10 ml Krebs’ buffer. Agar-salt bridges and electrodes
were used to measure potential difference. Concentration-
dependent changes in short-circuit current (Isc) were
determined for the cumulative addition of the neurotrans-
mitter, acetylcholine (ACH) added to the serosal side, as an
index of MgD effect on secretory function and to glucose,
added to the mucosal side, as an index of MgD effects on
mucosal absorption.
Acute Mg2+ replacement
About 24 h prior to study, some MgD animals were
assigned randomly to receive acute Mg2+ replacement.
Animals assigned to the MgR protocol were placed on the
Mg2+-supplemented diet 24 h prior to study and received 3
60 Mol Cell Biochem (2007) 306:59–69
123

doses of 10% magnesium sulfate (50 mg/kg im) 18, 6, and
2 h prior to surgery. This paradigm is based on the current
practice of repletion of magnesium deficiency in critically
ill patients prior to surgery. Serum Mg2+ levels from the
rats undergoing replacement therapy were determined to
confirm normal serum levels.
RBC glutathione determination
Blood samples were drawn from the inferior vena cava into
Na-citrate (1%) solution. Red blood cells (RBC) were
centrifuged at 200 · g for 10 min, washed twice with
Dulbecco’s PBS, and pelleted at 200 · g for 10 min. The
packed RBCs were diluted 50-fold, hemolyzed in 10%
PBS, and acidified by 5% 5-sulfosalicyclic acid immedi-
ately to preserve the reduced glutathione (GSH). Total
glutathione [unoxidized glutathione (GSH) + ½ oxidized
glutathione (GSSG)] was determined by the ‘enzymatic
recycling method’ involving GSSG reductase-catalyzed
cycling of GSH with 5,5¢-dithiobis(2-nitrobenzoic acid) as
described [18, 19]. GSSG was determined by the same
procedure except for the prior masking of GSH by 1%
vinyl pyridine. RBC hemoglobin was determined by the
cyanmethemoglobin technique using Sigma Diagnostics
hemoglobin reagents (Sigma, St. Louis, MO).
Histology
Intestinal sections were rinsed in cold saline and fixed in
4% paraformaldehyde. Tissues were embedded in paraffin
and 5 lm transverse sections were stained with H&E and
Giemsa for light microscopic evaluation of tissue archi-
tecture and differentiation of leukocytes to clearly identify
neutrophilic infiltration, respectively. Under 10· magnifi-
cation, two blinded examiners graded the mucosal injury of
each slide based on a previously established six-tiered scale
[0 (normal)-5 (severe)]. Villous height was also obtained
using a digital micrometer and reported in micrometers.
Under 40· magnification, the number of PMNs was
counted in 20 separate fields per slide immediately superior
to the muscularis mucosae. The mean number of PMNs per
high-power field (HPF) was determined for each animal.
Immunohistochemistry
Frozen tissue sections (5–10 lm) were air-dried, washed
3 times with phosphate buffered saline (PBS), and trans-
ferred to the blocking reagent 10% goat serum, 1% BSA,
and 0.03% Triton ·100 in PBS for 60 min to reduce non-
specific binding. Sections were incubated overnight at 4�C
with primary antibody, diluted in blocking buffer, washed
3 times, and incubated with secondary antibody conjugated
with FITC or Texas Red (CD11b) for 1 h at room
temperature. Tissues were protected by mounting medium
and glass cover slips. Multiple digital images were ob-
tained at different magnifications for further analysis and
fluorescence intensity and area were measured (Nikon).
CD11b, ICAM-1, and NOS-3 staining were evaluated
using this technique and appropriate primary and second-
ary antibodies (Santa Cruz Biotechnology, Inc., Santa
Cruz, CA).
Vascular macromolecular leak
In some animals, Evan’s blue dye (0.2 ml, 1.5%) was in-
jected intravenously through the tail vein 1 h prior to
harvest to allow for determination of systemic vascular
leak. At euthanasia, small bowel lavage (SBL) was per-
formed by irrigating the terminal 20 cm of small intestine
with 5 ml of normal saline and collecting the effluent. The
heart and lungs were removed en bloc via median stern-
otomy and a brocheoalveolar lavage (BAL) was performed
by infusing 5 ml of normal saline via tracheotomy, waiting
30 s, and then aspirating the effluent. Prior to the extirpa-
tion of the heart and lungs, a blood sample was obtained
from the right ventricle by cardiac puncture. All collected
samples were centrifuged and serum samples were diluted
20 fold. The concentration of dye was assessed spectro-
photometrically (Eflab, Titerrek, Multiscan Plus, Helsinki,
Finland) at a wavelength of 630 nm. SBL to serum and
BAL to serum AB630 ratios were calculated as measures of
in vivo intestinal macromolecular capillary and pulmonary
leak. Baseline leak was defined as the mean leak in sham-
operated animals.
RNA extraction and cDNA synthesis
Full thickness sections of mid jejunum were placed
immediately in TRIzol reagent (Invitrogen, Grand Island,
NY). Total RNA was extracted as per the manufacture’s
instructions and RNA quantity was assessed using the
Beckman Coulter DU530 UV/Vis Sprectrophotometer
(Beckman Coulter, Fullerton, CA). RNA samples (2 lg)
were reverse-transcribed to cDNA using the First Strand
cDNA Synthase Kit (MBI Fermentas, Hanover, MD) with
random hexamer primer.
Real-time quantitative PCR
Real-time PCR was performed with a BioRad iCycler se-
quence detection system (Hercules, CA). Primer sequences
were designed using computer program Beacon Designer
4.00 (Premier Biosoft International, Palo Alto, CA), and
synthesized by The Biopolymer Lab (University of Mary-
land, Dept. of Microbiology/Immunology, Baltimore, MD).
PCR was performed in a 25-ll volume using BioRad iQ
Mol Cell Biochem (2007) 306:59–69 61
123
SYBR Green Supermix (BioRad Hercules, CA). Assays
were optimized as follows: primer concentrations were
varied until highest signal intensity and lowest threshold of
detection were achieved; cDNA templates were diluted to
determine linearity of optimum primer and probe concen-
tration. Amplification conditions were: 50�C for 2 min,
95�C for 10 min, 50 cycles of 95�C for 15s and 60�C for
1 min. The fold-changes in mRNA expression for NOS-2,
NOS-3, NFjB, and COX-2 were relative to the respective
vehicle groups of mice after normalization to 18s rRNA
(Applied Biosystems, Foster City, CA).
Statistical analysis
Results are expressed as means ± SEM. Statistical analysis
was performed using a paired t-test or one-way ANOVA
followed by Neuman–Keuls tests for determining differ-
ences between individual means using GraphPad Prism
(v4, Graphpad Software Inc., San Diego, CA). A value of
p < 0.05 was considered significant.
Results
MgD effects on serum electrolyte levels
Rats fed the deficient diet for 3 weeks had significantly
lower levels of serum Mg2+ than sham controls (Table 1).
There were no differences in survival among the treatment
groups. Acute Mg2+ repletion significantly elevated serum
Mg2+ levels above those in the MgD, however, the level
remained lower than those in the sham control groups
(Table 1). Of interest is that 4 hours after IR, there was a
significant increase in serum Mg2+ that was also observed
even in the MgD group suggesting that mesenteric IR
mobilizes Mg2+ (Table 1). Serum levels of potassium (K+)
or calcium (Ca2+) were not altered significantly by either
diet or treatment (Table 1).
Effects of MgD on red blood cell (RBC) glutathione
To confirm previous results showing that endogenous
antioxidant defenses were impaired by MgD, we observed
that total glutathione levels of the RBC samples from the
MgD rats reduced gradually over time. Blood glutathione
levels were unchanged after 1 week (6.30 ± 0.38 vs.
5.28 ± 0.29 lM)), but were decreased significantly to 57%
of levels in MgS rats at the end of the second week
(6.22 ± 0.33 vs. 3.61 ± 0.21 lM, p < 0.01), and to 49% by
the end of third week of deficiency (6.11 ± 0.30 vs.
3.09 ± 0.22 lM, p < 0.01). In data not shown, the levels of
GSSG in either the Mg deficient or Mg control RBC
samples were less than 1% of the total glutathione.
MgD effects on intestinal morphology and function
Histology
When compared to the control diet (MgS), the MgD alone
did not alter mucosal injury (Fig. 1A, B). In contrast, MgD
alone induced significant PMN infiltration after 1 week
that was still evident at 3 weeks (Fig. 2A). PMN infiltra-
tion in the intestine involves the leukocyte b2 integrins,
especially CD11b/CD18 and is associated with increased
expression of ICAM. Immunohistochemistry of jejunal
tissue sections obtained from MgS rats stained with
anti-CD11b antibody showed the presence of PMNs and
confirmed the MgD-increased PMN infiltration (Fig. 2C
vs. 2D) that was observed in Giemsa-stained sections under
light microscopy (Fig. 2B). Staining with ICAM-1 anti-
body showed a specific, but low level of, staining in the
endothelium along the vascular wall in MgS rats (Fig. 3A),
demonstrating that ICAM-1 is constitutively and moder-
ately expressed. There was evidence of marked vasodila-
tion in the MgD group associated with increased ICAM-1
staining within the wall of submucosal vessels (Fig. 3B).
ICAM-1 is an adhesion molecule involved in PMN infil-
tration and therefore, the increased staining for ICAM-1 in
blood vessels in MgD rats (Fig. 3A vs. 3B) is consistent
with the increased numbers of PMN (Fig. 2).
mRNA expression of inflammatory mediators
To determine if this PMN infiltration was associated with
changes in the proinflammatory mediators, we determined
mRNA expression of NOS-2, COX-2, and NFjB in full
Table 1 Changes in Serum magnesium (Mg2+), potassium (K+) and calcium (Ca2+) levels in response to mesenteric IR in rats fed Mg2+
sufficient (MgS) or Mg2+ deficient (MgD) diets or in MgD rats after acute Mg2+ repletion (MgR)
MgS MgD MgR
SHAM IR-30 IR-30/4 SHAM IR-30 IR-30/4 SHAM IR-30 IR-30/4
Mg2+ 1.8 ± 0.1 2.3 ± 0.2* 2.9 ± 0.2* 0.7 ± 0.1* 0.8 ± 0.1 1.3 ± 0.1 ** 1.5 ± 0.1 1.5 ± 0.6 2.5 ± 0.3**
K+ 8.9 ± 0.4 10.3 ± 0.7 9.5 ± 0.7 7.4 ± 0.5 10.9 ± 0.9 12.2 ± 2.5 7.7 ± 0.6 8.6 ± 0.1 12.2 ± 1.6
Ca2+ 11.2 ± 0.5 10.7 ± 0.4 11.9± .6 11.2 ± 0.4 11.4 ± 0.2 11.5 ± 0.6 11.5 ± 0.2 11.9 ± 0.2 11.0 ± 0.4
n ‡ 5 for each group; *p < 0.05 vs MgS; **p < 0.05 vs. respective SHAM; K+ in mEq/l; Ca2+ in mg/dl
62 Mol Cell Biochem (2007) 306:59–69
123

thickness sections of mid-jejunum. MgD alone did not
significantly alter expression of NOS-2 (1.0 ± 1.5 vs.
1.0 ± 1.1 fold), COX-2 (1.0 ± 0.2 vs. 0.6 ± 0.2 fold), or
NFjB (1.0 ± 0.2 vs. 0.7 ± 0.1 suggesting that MgD in-
duces a subclinical inflammation.
Vascular macromolecular leak
PMN infiltration in the small intestine is associated with
reduced levels of NOS-3 activity and evidence of increased
vascular permeability to macromolecules [20]. When
compared to MgS, full thickness sections of small intestine
taken from MgD (3 week) showed decreased NOS-3
mRNA expression in (Fig. 4A) coincident with decreased
staining for NOS-3 in the submucosal nerves as well as in
the walls of blood vessels in the submucosa (Fig. 4C vs.
4D). The decrease in NOS-3 expression (Fig. 4A) was
associated with an increase in vascular leak in both the
small intestine and lung (Fig. 4B).
Mucosal function
Secretory responses to acetylcholine were unchanged after
1 week, but were significantly inhibited after 3 weeks
(Fig. 5A). Glucose absorption was unaltered by MgD
(Fig. 5B) and is consistent with the lack of effect of
3 weeks of MgD on mucosal injury and on body weight
(301.1 ± 19.7 vs. 295.6 ± 8.6 gms).
Effects of MgD and acute MgR on responses to
mesenteric ischemia/reperfusion
Mucosal injury
Since MgD alone induced a proinflammatory state in the
gut, we next determined if MgD altered the response of the
small intestine to mesenteric IR, a known inducer of
oxidative stress. Neither MgS nor 1 or 3 weeks of the MgD
diet resulted in development of mucosal injury after a
Fig. 1 Sections of fixed tissue
taken from mid-jejunum in MgS
(A) and MgD (B) rats were cut
and mounted onto slides and
stained with Giemsa. MgD did
not alter intestinal morphology.
One representative picture from
each group of at least 6–8/group
rats is shown (Magnification,
·20)
A
MgS 1WMgD 3WMgD0
10
20
30
** **)fph/#(
NM
PB
C D
Fig. 2 Rats were fed a Mg2+
sufficient diet (MgS) or a Mg2+
deficient diet for either 1 week
(1W MgD) or 3 weeks (3W
MgD). (A) Both 1W MgD and
3W MgD increased the number
of PMNs; (B) Giemsa-stained
section of 3WMgD rat showing
increased PMN infiltration
(arrowhead); (C–D) Frozen
tissue blocks of mid-jejunum
were prepared and the sections
were cut for immunofluoresence
staining for CD11b. The
intensity of the staining was
determined by establishing
settings for the samples from the
individual vehicle groups and
using the same conditions to
evaluate the samples from the
treated groups. MgD increased
neutrophil infiltration (D) when
compared to MgS (C)
Mol Cell Biochem (2007) 306:59–69 63
123

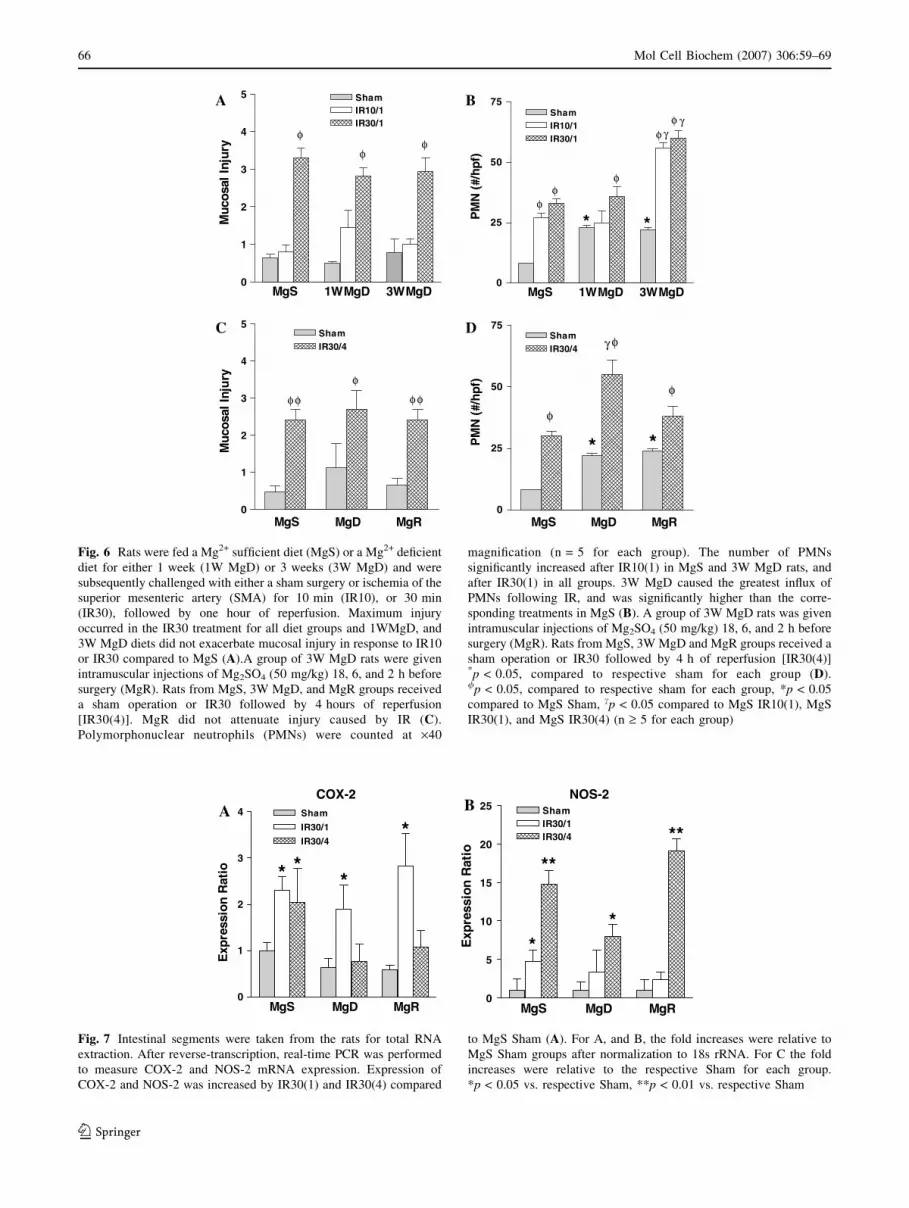
10 minute period of ischemia and 1 hour of perfusion (IR-
10/1), but all groups exhibited a comparable and significant
mucosal injury after IR-30/1 (Fig. 6A) or IR-30/4 (Fig. 6C)
of reperfusion. Acute Mg2+ repletion (MgR) did not
diminish mucosal injury induced by IR-30/4 (Fig. 6D).
Inflammation
In the MgS rats, IR-10/1 induced PMN influx in the
absence of mucosal damage (Fig. 6A, C). In contrast, there
was a significant increase in PMN infiltration following
IR-30/1 (Fig. 6B) and IR-30/4 (Fig. 6D) in the MgS group
that was associated with the mucosal damage (6A, C).
MgD exaggerated the inflammatory response following
IR30/1 (Fig. 6B) and IR30/4 (Fig. 6D). MgR blunted the
MgD-induced enhancement of PMN infiltration to levels
observed in the MgS group (Fig. 6D).
mRNA expression of inflammatory mediators
Since MgD enhanced PMN infiltration in response to
mesenteric IR, we determined if this was associated with
altered expression of inflammatory mediators. In MgS rats,
IR-30/1 significantly increased mRNA expression of COX-
2 and NOS-2 consistent with IR-induced injury and PMN
infiltration (Fig. 7A, B). Expression of COX-2 remained
elevated in IR-30/4 (Fig. 7A) while expression of NOS-2
increased further after IR-30/4 (Fig. 7B) in MgS rats. In the
MgD group, the expression of COX-2 (Fig. 7A) was sim-
ilar to that in the MgS rats after IR-30/1. In contrast, after
IR-30/4 COX-2 expression was significantly less in the
MgD, than in MgS, rats suggesting that MgD shortens the
duration of the upregulation of COX-2 in response to IR.
MgR did not alter the effects of MgD on COX-2 expression
(Fig. 7A). The expression of NOS-2 in response to IR was
both delayed and attenuated in the MgD group (Fig. 7B).
MgR did not alter the MgD-induced delay in NOS-2
expression (Fig. 7B), but the level of expression at IR30/4
was similar to that in the MgS group.
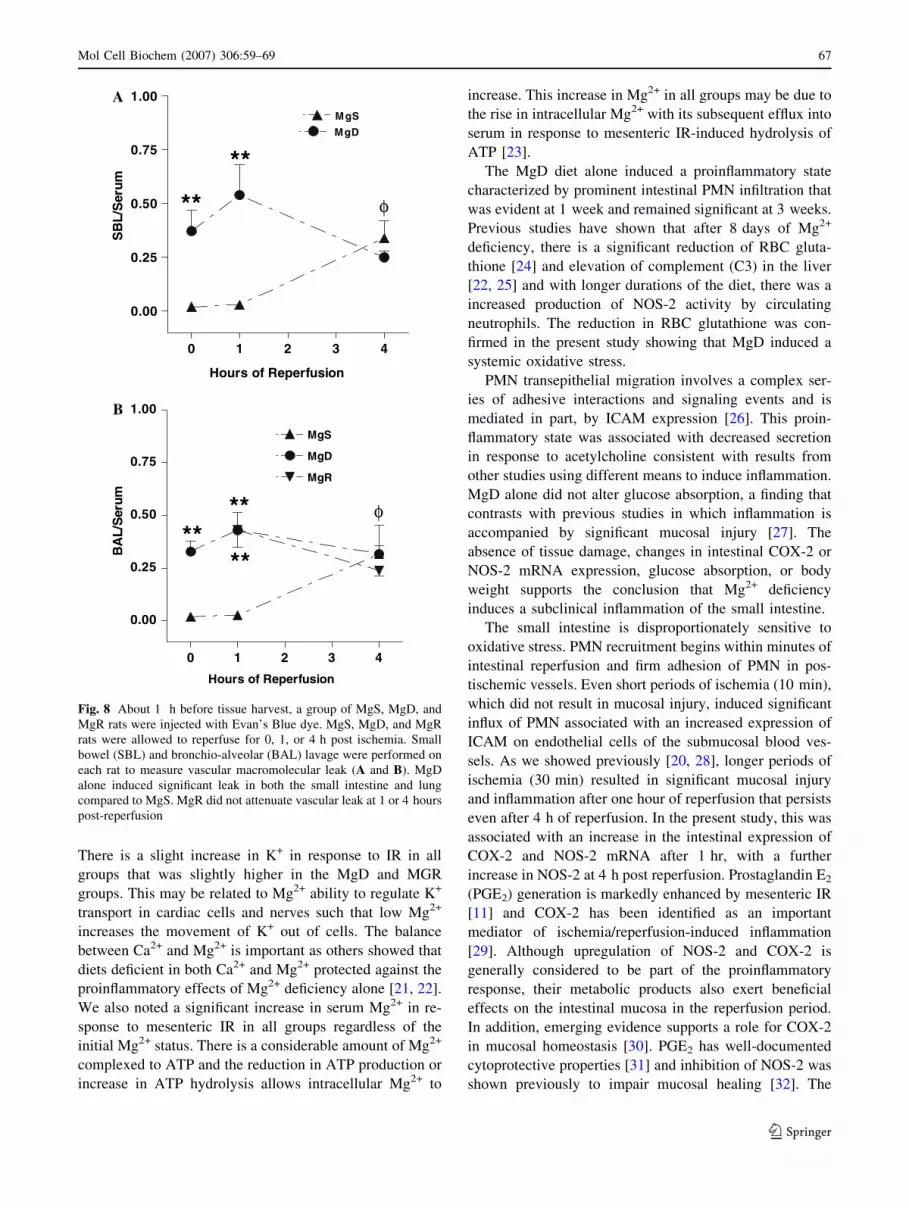
Vascular macromolecular leak
When compared to sham-operated MgS rats, IR increased
intestinal vascular leak in both the small intestine (SBL)
and lung (BAL), but only after 4 h (Fig. 8A, B). In con-
trast, MgD alone induced significant vascular leak in the
small intestine and lung that remained elevated, but was
not further enhanced, after 1 or 4 h of reperfusion. Acute
Mg2+ repletion of MgD rats did not improve basal or
IR-induced changes in vascular permeability in either
region (Fig. 8B).
Discussion
The intestine is the site of Mg2+ absorption, yet little is
known about the effects of hypomagnesmia on the small
intestine. In the present study, rats were fed a Mg2+ defi-
cient diet to induce a gradual hypomagnesmia over a
3 week period. Significant inflammation in the absence of
mucosal injury in the small intestine was evident after
1 week and remained significant after 3 weeks on the diet.
The intestinal inflammation after 3 weeks of MgD was
Fig. 3 Rats were fed a Mg2+ sufficient diet (MgS) or a Mg2+ deficient
diet for 3 weeks (3W MgD). Sections (5 l) of frozen tissue blocks of
mid-jejunum taken from rats after 3 weeks on the MgS and MgD diet
were prepared for immunofluoresent staining of ICAM-1 (arrows).
The intensity of the staining was determined by establishing settings
for the samples from the individual vehicle groups and using the same
conditions to evaluate the samples from the infected or treated groups.
ICAM-1 positive staining was very specific for the endothelium of the
vascular walls (arrows). There is a low-level, constitutive expression
of ICAM-1 in MgS, however, 3W MgD greatly enhanced the intensity
of ICAM-1 staining of rat endothelium
64 Mol Cell Biochem (2007) 306:59–69
123

associated with significant changes in intestinal and pul-
monary vascular permeability as well as intestinal function.
MgD also induced an increased sensitivity to mesenteric
ischemia reperfusion characterized by inflammation in re-
sponse to a normally non-injurious period of intestinal
ischemia (10 min) and amplification of PNM infiltration
after an injurious period (30 min) of ischemia. Acute Mg2+
repletion was only partially effective in reversing the MgD-
induced changes in intestinal morphology and function.
In the present study, we showed that rats fed Mg2+-
deficient diets for 3 weeks exhibited significant reductions
in serum Mg2+ without alterations in Ca2+ or K+. These
data indicate, however, that homeostasis of Ca2+ or K+ can
be maintained during a gradual onset of Mg2+ deficiency.
A NOS-3
MgS 3W MgD MgR0.00
0.25
0.50
0.75
1.00
1.25
* *
oitaR
noisserpxE
Vascular Leak
0.0
0.1
0.2
0.3
0.4
0.5 MgSMgD**
**
Intestine Lung
)mures/
LB
Sr
oL
AB (
k aeL
B
C D
Fig. 4 Rats were fed a Mg2+ sufficient diet (MgS) or a Mg2+ deficient
diet for 3 weeks (3W MgD). Separate groups of 3w MgD rats were
repleted with magnesium before study (MgR). Segments of intestine
were taken from each animal in each group to measure NOS-3 mRNA
expression using real-time PCR. Expression of NOS-3 was dimin-
ished by 3W MgD and MgR in rat small intestine (A). Acute Mg2+
repletion did not improve expression of NOS-3. *p < 0.05 vs. MgS.
Sections of frozen tissue blocks of mid-jejunum were prepared for
immunofluorescent staining for NOS-3. NOS-3 positive staining is
visible throughout the section and is similar in both groups in
myenteric plexus neurons (arrowheads). In contrast, staining for
NOS-3 in MgS (C) was greater in the submucosal plexus neurons and
blood vessels (arrows) compared with 3W MgD (D). The intensity of
the staining was determined by establishing settings for the samples
from the individual vehicle groups and using the same conditions to
evaluate the samples from the treated groups. About 1 h before tissue
harvest, a group of MgS, MgD, and MgR rats were injected with
Evan’s Blue dye. Small bowel and bronchio-alveolar lavage were
performed on each rat to measure vascular macromolecular leak (B).
MgD alone induced significant leak in both the small intestine and
lung compared to MgS. **p < 0.01 vs.MgS
Acetylcholine
-8 -7 -6 -5 -4 -3
0
20
40
60
80
100A B
MgS
3WMgD
*
1wMgD
ACETYLCHOLINE LOG [M]
(csI
NIE
GN
AH
Cµ
mc/A
2 )
Glucose
1 10 1000
100
200
300
MgS
3WMgD1WMgD
GLUCOSE [mM]
(csI
NIE
GN
AH
Cµ
mc /A
2 )
Fig. 5 Rats were fed a Mg2+
sufficient diet (MgS) or a Mg2+
deficient diet for either 1 week
(1W MgD) or 3 weeks (3W
MgD). Segments of muscle-free
jejuna were mounted in Ussing
chambers to measure secretion
in response to acetylcholine (A)
and absorption in response to
glucose (B)
Mol Cell Biochem (2007) 306:59–69 65
123
0
1
2
3
4
5
φ
φ φφ φ
ShamIR30/4
MgS MgRMgD
jury
nIlasocuM
A B
C D
0
25
50
75
φ
* *
φγ
γ
φ
φφ
ShamIR10/1IR30/1
MgS 1WMgD 3WMgD
)fph/#(N
MP
0
25
50
75
*
φγ
φ
*φ
ShamIR30/4
MgS MgD MgR
)fph/#(N
MP
0
1
2
3
4
5
φφ
φ
ShamIR10/1IR30/1
MgS 1WMgD 3WMgD
yrujnIlasocuM
Fig. 6 Rats were fed a Mg2+ sufficient diet (MgS) or a Mg2+ deficient
diet for either 1 week (1W MgD) or 3 weeks (3W MgD) and were
subsequently challenged with either a sham surgery or ischemia of the
superior mesenteric artery (SMA) for 10 min (IR10), or 30 min
(IR30), followed by one hour of reperfusion. Maximum injury
occurred in the IR30 treatment for all diet groups and 1WMgD, and
3W MgD diets did not exacerbate mucosal injury in response to IR10
or IR30 compared to MgS (A).A group of 3W MgD rats were given
intramuscular injections of Mg2SO4 (50 mg/kg) 18, 6, and 2 h before
surgery (MgR). Rats from MgS, 3W MgD, and MgR groups received
a sham operation or IR30 followed by 4 hours of reperfusion
[IR30(4)]. MgR did not attenuate injury caused by IR (C).
Polymorphonuclear neutrophils (PMNs) were counted at ·40
magnification (n = 5 for each group). The number of PMNs
significantly increased after IR10(1) in MgS and 3W MgD rats, and
after IR30(1) in all groups. 3W MgD caused the greatest influx of
PMNs following IR, and was significantly higher than the corre-
sponding treatments in MgS (B). A group of 3W MgD rats was given
intramuscular injections of Mg2SO4 (50 mg/kg) 18, 6, and 2 h before
surgery (MgR). Rats from MgS, 3W MgD and MgR groups received a
sham operation or IR30 followed by 4 h of reperfusion [IR30(4)]*p < 0.05, compared to respective sham for each group (D)./p < 0.05, compared to respective sham for each group, *p < 0.05
compared to MgS Sham, cp < 0.05 compared to MgS IR10(1), MgS
IR30(1), and MgS IR30(4) (n ‡ 5 for each group)
NOS-2
0
5
10
15
20
25
**
*
**
*
ShamIR30/1IR30/4
MgS MgD MgR
oitaR
noisserpxE
COX-2
0
1
2
3
4A B
* *
*
*
Sham
IR30/1
IR30/4
MgS MgD MgR
oitaR
noisserpxE
Fig. 7 Intestinal segments were taken from the rats for total RNA
extraction. After reverse-transcription, real-time PCR was performed
to measure COX-2 and NOS-2 mRNA expression. Expression of
COX-2 and NOS-2 was increased by IR30(1) and IR30(4) compared
to MgS Sham (A). For A, and B, the fold increases were relative to
MgS Sham groups after normalization to 18s rRNA. For C the fold
increases were relative to the respective Sham for each group.
*p < 0.05 vs. respective Sham, **p < 0.01 vs. respective Sham
66 Mol Cell Biochem (2007) 306:59–69
123
There is a slight increase in K+ in response to IR in all
groups that was slightly higher in the MgD and MGR
groups. This may be related to Mg2+ ability to regulate K+
transport in cardiac cells and nerves such that low Mg2+
increases the movement of K+ out of cells. The balance
between Ca2+ and Mg2+ is important as others showed that
diets deficient in both Ca2+ and Mg2+ protected against the
proinflammatory effects of Mg2+ deficiency alone [21, 22].
We also noted a significant increase in serum Mg2+ in re-
sponse to mesenteric IR in all groups regardless of the
initial Mg2+ status. There is a considerable amount of Mg2+
complexed to ATP and the reduction in ATP production or
increase in ATP hydrolysis allows intracellular Mg2+ to
increase. This increase in Mg2+ in all groups may be due to
the rise in intracellular Mg2+ with its subsequent efflux into
serum in response to mesenteric IR-induced hydrolysis of
ATP [23].
The MgD diet alone induced a proinflammatory state
characterized by prominent intestinal PMN infiltration that
was evident at 1 week and remained significant at 3 weeks.
Previous studies have shown that after 8 days of Mg2+
deficiency, there is a significant reduction of RBC gluta-
thione [24] and elevation of complement (C3) in the liver
[22, 25] and with longer durations of the diet, there was a
increased production of NOS-2 activity by circulating
neutrophils. The reduction in RBC glutathione was con-
firmed in the present study showing that MgD induced a
systemic oxidative stress.
PMN transepithelial migration involves a complex ser-
ies of adhesive interactions and signaling events and is
mediated in part, by ICAM expression [26]. This proin-
flammatory state was associated with decreased secretion
in response to acetylcholine consistent with results from
other studies using different means to induce inflammation.
MgD alone did not alter glucose absorption, a finding that
contrasts with previous studies in which inflammation is
accompanied by significant mucosal injury [27]. The
absence of tissue damage, changes in intestinal COX-2 or
NOS-2 mRNA expression, glucose absorption, or body
weight supports the conclusion that Mg2+ deficiency
induces a subclinical inflammation of the small intestine.
The small intestine is disproportionately sensitive to
oxidative stress. PMN recruitment begins within minutes of
intestinal reperfusion and firm adhesion of PMN in pos-
tischemic vessels. Even short periods of ischemia (10 min),
which did not result in mucosal injury, induced significant
influx of PMN associated with an increased expression of
ICAM on endothelial cells of the submucosal blood ves-
sels. As we showed previously [20, 28], longer periods of
ischemia (30 min) resulted in significant mucosal injury
and inflammation after one hour of reperfusion that persists
even after 4 h of reperfusion. In the present study, this was
associated with an increase in the intestinal expression of
COX-2 and NOS-2 mRNA after 1 hr, with a further
increase in NOS-2 at 4 h post reperfusion. Prostaglandin E2
(PGE2) generation is markedly enhanced by mesenteric IR
[11] and COX-2 has been identified as an important
mediator of ischemia/reperfusion-induced inflammation
[29]. Although upregulation of NOS-2 and COX-2 is
generally considered to be part of the proinflammatory
response, their metabolic products also exert beneficial
effects on the intestinal mucosa in the reperfusion period.
In addition, emerging evidence supports a role for COX-2
in mucosal homeostasis [30]. PGE2 has well-documented
cytoprotective properties [31] and inhibition of NOS-2 was
shown previously to impair mucosal healing [32]. The
A
B
0 1 2 3 4
0.00
0.25
0.50
0.75
1.00MgS
MgD
** φ
**
Hours of Reperfusion
mureS/L
BS
0 1 2 3 4
0.00
0.25
0.50
0.75
1.00
MgS
MgD
MgR
**φ
**
**
Hours of Reperfusion
mureS/L
AB
Fig. 8 About 1 h before tissue harvest, a group of MgS, MgD, and
MgR rats were injected with Evan’s Blue dye. MgS, MgD, and MgR
rats were allowed to reperfuse for 0, 1, or 4 h post ischemia. Small
bowel (SBL) and bronchio-alveolar (BAL) lavage were performed on
each rat to measure vascular macromolecular leak (A and B). MgD
alone induced significant leak in both the small intestine and lung
compared to MgS. MgR did not attenuate vascular leak at 1 or 4 hours
post-reperfusion
Mol Cell Biochem (2007) 306:59–69 67
123

maximal induction of NOS-2 occurs at 4 h into the reper-
fusion period, correlating with the observed initiation of
mucosal restitution [20, 33]. Taken together these data
suggest that the MgD attenuates IR-induced upregulation
of COX-2 and NOS-2, whose products may act to balance
the inflammation-induced injury and to promote sub-
sequent healing and restitution of the gut.
Mg2+ deficiency significantly enhanced the sensitivity of
the intestine to IR-induced inflammation. After 1 week of
the deficient diet there was an evident subclinical inflam-
mation with no change in mucosal morphology, when
compared to the MgS group. Rats on the MgD diet for
3 weeks exhibited a significant PMN infiltration after only
10 min of ischemia and an augmented response to 30 min
of ischemia at 1 and 4 h of reperfusion. An acute Mg2+
repletion paradigm, similar to that utilized for repletion in
the trauma patients, did not alter PMN infiltration, but
significantly blunted the exaggerated IR-induced PMN
influx observed in MgD rats; PMN infiltration is a hallmark
feature of mesenteric IR, but its role in the associated
mucosal injury may be limited. Recent studies have
implicated other factors including macrophages [34],
complement activation and deposition [28, 35] and/or
natural antibodies [36] as having a more direct effect.
These data indicate that Mg2+ has a specific anti-inflam-
matory effect in the intestinal response to oxidative stress
and that clinical repletion of Mg2+ may be beneficial in
preventing further inflammation in response to intestinal
trauma.
We showed previously that the post ischemic intestine
serves as a priming bed for circulating PMNs that
contribute to the development of a systemic inflammatory
response (SIRS) and pulmonary injury [12, 20, 33, 37]. In
the MgS group, there was an emergence of significant
endothelial permeability within the small intestine and lung
that developed after 4 h. The inflammation induced by
MgD alone was sufficient to induce increased endothelial
permeability that was, surprisingly, not worsened by
intestinal IR. Constitutive production of NO plays a key
role in the maintenance of endothelial as well as epithelial
permeability. MgD inhibited constitutive NOS-3 expres-
sion particular in the submucosal area, where the large
mucosal blood vessels are located. Neither NOS-3
expression nor the enhanced vascular permeability were
prevented by MgR.
In summary, this data shows that gradual development
of severe MgD over 3 weeks induces a subclinical
inflammation characterized by PMN infiltration and
enhanced vascular permeability without mucosal injury or
upregulation of expression of proinflammatory pathways.
MgD, however, enhanced the inflammatory responses to
mesenteric IR and altered the amplitude and the duration
of the expression of key mediators that contribute to
inflammation. Acute Mg2+ repletion of MgD rats was
ineffective in alleviating the proinflammatory effects
induced by MgD alone, but was able to blunt the MgD-
induced alteration in molecular and functional responses to
mesenteric IR. These data demonstrate the importance of
Mg2+ status and the limited benefits of therapeutic repletion
of Mg2+ serum levels in trauma patients.
Acknowledgements This work was supported by NIH grants
National Institutes of Health grants R01-HL-62282 and HL-65718
awarded to W.B.W. and AI 49316 to TSD, and USDA CRIS project
#1235-52000-055. The opinions and assertions in this article are those
of the authors and do not necessarily represent those of the U. S.
Department of Defense or the U. S. Department of Agriculture.
References
1. Dube L, Granry JC (2003) The therapeutic use of magnesium
in anesthesiology, intensive care and emergency medicine: a
review: [L’usage therapeutique du magnesium en anesthesiol-
ogie, reanimation et medecine d’urgence]. Can J Anesth
50:732–746
2. Noronha L, Matuschak G (2002) Magnesium in critical illness:
metabolism, assessment, and treatment. Intensive Care Medicine
28:667–679
3. Tosiello L (1996) Hypomagnesemia and diabetes mellitus. A
review of clinical implications. Arch Intern Med 156:1143–1148
4. Kahraman S, Ozgurtas T, Kayali H, Atabey C, Kutluay T,
Timurkaynak E (2003) Monitoring of serum ionized magnesium
in neurosurgical intensive care unit: preliminary results. Clinica
Chimica Acta 334:211–215
5. Kramer JH, Mak IT, Phillips TM, Weglicki WB (2003) Dietary
magnesium intake influences circulating pro-inflammatory neu-
ropeptide levels and loss of myocardial tolerance to postischemic
stress. Exp Biol Med (Maywood) 228:665–673
6. Weglicki WB, Phillips TM (1992) Pathobiology of magnesium
deficiency: a cytokine/neurogenic inflammation hypothesis. Am J
Physiol 263:R734–R737
7. Weglicki WB, Bloom S, Cassidy MM, Freedman AM, Atrakchi
AH, Dickens BF (1992) Antioxidants and the cardiomyopathy of
Mg-deficiency. Am J Cardiovasc Pathol 4:210–215
8. Wiles ME, Wagner TL, Weglicki WB (1997) Effect of acute
magnesium deficiency (MgD) on aortic endothelial cell (EC)
oxidant production. Life Sci 60:221–236
9. Moore FA, Haenel JB, Moore EE, Whitehill TA (1992) Incom-
mensurate oxygen consumption in response to maximal oxygen
availability predicts postinjury multiple organ failure. J Trauma
33:58–65
10. Rehrig S, Fleming SD, Anderson J, Guthridge JM, Rakstang J,
McQueen CE, Holers VM, Tsokos GC, Shea-Donohue T (2001)
Complement inhibitor, complement receptor 1-related gene/pro-
tein y-Ig attenuates intestinal damage after the onset of
mesenteric ischemia/reperfusion injury in mice. J Immunol
167:5921–5927
11. Stojadinovic A, Kiang J, Smallridge R, Galloway R, Shea-Don-
ohue T (1995) Induction of heat-shock protein 72 protects against
ischemia/reperfusion in rat small intestine. Gastroenterology
109:505–515
12. Ward DT, Lawson SA, Gallagher CM, Conner WC, Shea-Don-
ohue T (2000) Sustained nitric oxide production via l-arginine
administration ameliorates effects of intestinal ischemia-reper-
fusion. J Surg Res 89:13–19
68 Mol Cell Biochem (2007) 306:59–69
123

13. Hernandez LA, Grisham MB, Twohig B, Arfors KE, Harlan JM,
Granger DN (1987) Role of neutrophils in ischemia-reperfusion-
induced microvascular injury. Am J Physiol 253:H699–H703
14. Bienvenu K, Hernandez L, Granger DN (1992) Leukocyte
adhesion and emigration in inflammation. Ann NY Acad Sci
664:388–399
15. Grisham MB, Hernandez LA, Granger DN (1986) Xanthine
oxidase and neutrophil infiltration in intestinal ischemia. Am J
Physiol 251:G567–G574
16. Kubes P, Hunter J, Granger DN (1992) Ischemia/reperfusion-
induced feline intestinal dysfunction: importance of granulocyte
recruitment. Gastroenterology 103:807–812
17. Hayward R, Lefer AM (1998) Time course of endothelial-neu-
trophil interaction in splanchnic artery ischemia-reperfusion. Am
J Physiol Heart Circ Physiol 275:H2080–H2086
18. Mak IT, Komarov AM, Wagner TL, Stafford RE, Dickens BF,
Weglicki WB (1996) Enhanced NO production during Mg defi-
ciency and its role in mediating red blood cell glutathione loss.
Am J Physiol 271:C385–C390
19. Mak IT, Dickens BF, Komarov AM, Wagner TL, Phillips TM,
Weglicki WB (1997) Activation of the neutrophil and loss of
plasma glutathione during Mg-deficiency–modulation by nitric
oxide synthase inhibition. Mol Cell Biochem 176:35–39
20. Tavaf-Motamen H, Miner TJ, Starnes BW, Shea-Donohue T
(1998) Nitric oxide mediates acute lung injury by modulation of
inflammation. J Surg Res 78:137–142
21. Bussiere FI, Gueux E, Rock E, Mazur A, Rayssiguier Y (2002)
Protective effect of calcium deficiency on the inflammatory re-
sponse in magnesium-deficient rats. Eur J Nutr 41:197–202
22. Mak IT, Komarov AM, Kramer JH, Weglicki WB (2000) Pro-
tective mechanisms of Mg-gluconate against oxidative endothe-
lial cytotoxicity. Cell Mol Biol (Noisy -le-grand) 46:1337–1344
23. Murphy E, Steenbergen C, Levy LA, Raju B, London RE (1989)
Cytosolic free magnesium levels in ischemic rat heart. J Biol
Chem 264:5622–5627
24. Mak IT, Stafford R, Weglicki WB (1994) Loss of red blood cell
glutathione during Mg deficiency: prevention by vitamin E, D-
propranolol, and chloroquine. Am J Physiol 267:C1366–C1370
25. Bussiere FI, Tridon A, Zimowska W, Mazur A, Rayssiguier Y
(2003) Increase in complement component C3 is an early
response to experimental magnesium deficiency in rats. Life Sci
73:499–507
26. Kubes P, Ward PA (2000) Leukocyte recruitment and the acute
inflammatory response. Brain Pathol 10:127–135
27. Fleming SD, Starnes BW, Kiang JG, Stojadinovic A, Tsokos GC,
Shea-Donohue T (2002) Heat stress protection against mesenteric
I/R-induced alterations in intestinal mucosa in rats. J Appl
Physiol 92:2600–2607
28. Anderson J, Fleming SD, Rehrig S, Tsokos GC, Basta M, Shea-
Donohue T (2005) Intravenous immunoglobulin attenuates mes-
enteric ischemia-reperfusion injury. Clin Immunol 114:137–146
29. Sato N, Kozar RA, Zou L, Weatherall JM, Attuwaybi B, Moore-
Olufemi SD, Weisbrodt NW, Moore FA (2005) peroxisome
proliferator-activated receptor gamma mediates protection
against cyclooxygenase-2-induced gut dysfunction in a rodent
model of mesenteric ischemia/reperfusion. Shock 24:462–469
30. Wallace JL, Devchand PR (2005) Emerging roles for cyclooxy-
genase-2 in gastrointestinal mucosal defense. Br J Pharmacol
145:275–282
31. Blikslager AT, Roberts MC, Rhoads JM, Argenzio RA (1997)
Prostaglandins I2 and E2 have a synergistic role in rescuing
epithelial barrier function in porcine ileum. J Clin Invest
100:1928–1933
32. McCafferty DM, Mudgett JS, Swain MG, Kubes P (1997)
Inducible nitric oxide synthase plays a critical role in resolving
intestinal inflammation. Gastroenterology 112:1022–1027
33. Miner TJ, Tavaf-Motamen H, Stojadinovic A, Shea-Donohue T
(1999) Ischemia-reperfusion protects the rat small intestine
against subsequent injury. J Surg Res 82:1–10
34. Chen Y, Lui VCH, Rooijen NV, Tam PKH (2004) Depletion of
intestinal resident macrophages prevents ischaemia reperfusion
injury in gut. Gut 53:1772–1780
35. Fleming SD, Mastellos D, Karpel-Massler G, Shea-Donohue T,
Lambris JD, Tsokos GC (2003) C5a causes limited, polymor-
phonuclear cell-independent, mesenteric ischemia/reperfusion-
induced injury. Clin Immunol 108:263–273
36. Fleming SD, Shea-Donohue T, Guthridge JM, Kulik L, Wald-
schmidt TJ, Gipson MG, Tsokos GC, Holers VM (2002) Mice
deficient in complement receptors 1 and 2 lack a tissue injury-
inducing subset of the natural antibody repertoire. J Immunol
169:2126–2133
37. Lawson S, Ward DT, Conner C, Gallagher C, Tsokos G, Shea-
Donohue T (2002) Diabetic hyperglycemia: a facilitating factor
in systemic capillary leak. J Surg Res 105:95–101
Mol Cell Biochem (2007) 306:59–69 69
123