immunological studies on peroxynitrite modified human dna
TRANSCRIPT

Life Sciences 77 (2005) 2626–2642
www.elsevier.com/locate/lifescie
Immunological studies on peroxynitrite modified human DNA
Kiran Dixit, MoinuddinT, Asif Ali1
Department of Biochemistry, Faculty of Medicine, A.M.U., Aligarh 202002, India
Received 30 August 2004; accepted 9 February 2005
Abstract
Peroxynitrite (ONOO�) is a strong and potent oxidizing and nitrating agent, formed by rapid reaction of two
highly reactive, nitric oxide and superoxide anion. The action of peroxynitrite generated by synergistic action of
diethylamine NONOate (a nitric oxide donor) and 1,4-hydroquinone (a superoxide donor), on human placental
DNA was monitored by ultraviolet and fluorescence spectroscopy, melting temperature studies, S1 nuclease
digestibility and alkaline agarose electrophoresis. The peroxynitrite modified human DNA (ONOO�-DNA) was
found to be highly immunogenic in rabbits inducing high titre immunogen specific antibodies. However, the
induced antibodies exhibited appreciable cross-reactivity with various polynucleotides and nucleic acids. The data
demonstrate that the antibodies, though cross-reactive, preferentially bind ONOO�-modified epitopes on DNA.
Visual detection of immune complex formation with native and ONOO�-DNA reiterated preferential binding with
modified human DNA. DNA modified by ONOO� presents unique epitopes which may be one of the factors for
the induction of autoantibodies in cancer patients.
D 2005 Published by Elsevier Inc.
Keywords: Peroxynitrite; DNA damage; Cancer; Autoantibodies
Introduction
Peroxynitrite (ONOO�) is formed in inflamed tissue by a diffusion limited reaction of nitric oxide
(NO) and superoxide anion (O2!�) (Huie and Padmaja, 1993). The formation of excess ONOO� has been
0024-3205/$ -
doi:10.1016/j.l
T Correspond
3091268.
E-mail add1 Present Ad
P.O. Box 641,
see front matter D 2005 Published by Elsevier Inc.
fs.2005.02.026
ing author. Department of Biochemistry, J.N. Medical College, A.M.U., Aligarh-202002, India. Tel.: +91 571
ress: [email protected] (Moinuddin).
dress: Department of Clinical Biochemistry, College of Medicine and Medical Sciences, King Khalid University,
Abha, Kingdom of Saudi Arabia.

K. Dixit et al. / Life Sciences 77 (2005) 2626–2642 2627
implicated in many pathological conditions including reperfusion injury, chronic inflammation,
atherosclerosis, neurodegenerative diseases and an increased cancer incidence in tissues under chronic
inflammatory conditions (Rachmilewitz et al., 1993; Salgo et al., 1995; Ohshima, 2003). Peroxynitrite is
a powerful oxidizing and nitrating agent that exhibits high reactivity at physiological pH. (Richeson et
al., 1998). Cellular DNA is an important molecular target for ONOO� and the radical is capable of
reacting with both the deoxyribose and nucleobases. Prominent DNA modifications induced by exposure
to ONOO� include formation of 8-nitroguanine and 8-oxyguanine as well as the induction of DNA
single-strand breaks (Szabo and Ohshima, 1997). Peroxynitrite is mutagenic in the sup F gene, inducing
primarily G to T transversions and deletions clustered at the 5Vend of the gene (Tretyakova et al., 2000;
Juedes and Wogan, 1996). The mutagenicity of ONOO� is believed to result from chemical
modifications at guanine leading to miscoding. DNA single strands generated by ONOO� leads to
activation of the nuclear enzyme poly(ADP-ribose) synthetase (PARS), its activation and subsequent
cellular processes play an important role in ONOO�-induced alteration in a variety of pathophysio-
logical conditions (Szabo and Dawson, 1998).
Tissue inflammation and chronic infection lead to the overproduction of nitric oxide and superoxide
which rapidly combine to yield peroxynitrite, which is thought to be involved in both cell death and an
increased cancer risk observed for inflamed tissues (Ohshima and Bartsch, 1994). Increased levels of
circulating antibodies and autoantibodies have been reported in the serum of patients with malignancies
(Chagnand et al., 1992; Faiderbe et al., 1992; Becker et al., 1994). Elevated levels of antinuclear
antibodies have been reported in the serum of cancer patients of hepatocellular carcinoma. It has been well
established that native DNA in B-conformation is not immunogenic where as various modified forms of
DNA are immunogenic and induce cross-reactive antibodies (Jansson, 1985; Hasan and Ali, 1990; Ali et
al., 1985). Recent studies from our laboratory have reported ROS-modified DNA and NO-modified DNA
to be an effective immunogenic stimulus and the induced antibodies showed polyspecificity (Alam et al.,
1993; Ara and Ali, 1993; Ahmad et al., 1997; Ashok and Ali, 1999; Dixit and Ali, 2001).
In the present study, the immunogenicity of ONOO�-modified human DNA has been investigated.
The induced antibodies have been characterized for their fine antigenic binding specificity towards a
variety of ONOO�-modified and unmodified nucleic acid polymers and synthetic polynucleotides so as
to gain insight into the mechanism of autoantibody production in cancer. The study also involved the
comparative immunobinding of antibodies present in sera of various cancer patients, with native and
ONOO�-modified human DNA.
Methods
Human placental DNA, diethylamine NONOate (DEA-NO), 1,4-hydroquinone, nuclease S1,
methylated bovine serum albumin (MBSA), diethylenetriaminepentaacetic acid (DTPA), 1H-imidazol-
1-oxy,2-(14-carboxyphenyl)-4, 5-dihydro-4,4,5,5-tetra methyl-3-oxide (carboxy-PTIO), mannitol, su-
peroxide dismutase (SOD), desferrioxamine, anti-human and anti-rabbit IgG alkaline phosphatase
conjugates, p-nitrophenyl phosphate, ethidium bromide, coomassie brilliant blue G250 and R250,
Freund’s complete and incomplete adjuvants, synthetic polynucleotides, Tween-20, and DNA bases
were obtained from Sigma Chemical Company (U.S.A.). Protein-A Sepharose CL-4B was obtained from
Genei, India. Polystyrene flat bottom ELISA plates were from Nunc, Denmark. All other chemicals used
were of highest grade available.

K. Dixit et al. / Life Sciences 77 (2005) 2626–26422628
Plasmid DNA (pUC18) was isolated in a highly purified form through gentle lysis of bacterial cells
followed by centrifugation to remove the bulk of chromosomal DNA using plasmid isolation Mega Kit
from Qiagen, U.S.A.
DNA purification
Commercially available human placental DNA (Sigma, U.S.A.) was purified free of proteins, RNA
and single stranded regions as described earlier (Ali et al., 1985). Purity of DNAwas confirmed by A260/
A280 ratio.
Modification of human placental DNA by peroxynitrite
The experiment was carried out by incubating human DNA (50 Ag) in Na-Pi buffer, pH 7.4 containing
0.1 mM DTPA, 0.1 mM DEA-NO and 0.1 mM 1,4-hydroquinone at 37 8C for 3 h. DEA-NO and 1,4-
hydroquinone were removed by extensive dialysis against PBS, pH 7.4. All the other nucleic acid
samples and DNA bases were similarly modified.
The UV absorption characteristics of native and ONOO�-DNA were recorded on Shimadzu UV-240
spectrophotometer. Fluorescence emission spectroscopical analysis on native and ONOO�-DNAsamples (5 Ag/ml each) was undertaken using the intercalating dye, ethidium bromide, as the external
chromophore (2.5 Ag/ml). The readings for EtBr-DNA and EtBr-ONOO�-DNA were recorded on a
Shimadzu RF-5301 Spectrofluorometer at an excitation wavelength of 325 nm.
Agarose gel electrophoresis
Native and ONOO�-DNA were mixed with a 1/10 volume of sample buffer (0.125% bromophenol
blue, 30% Ficoll-400, 5 mM EDTA in 10� electrophoresis buffer). The samples were electrophoresed
for 2 h at 30 mA on 1% agarose gels. The gels were stained with ethidium bromide (0.5 Ag/ml), viewed
by illumination under UV light and photographed.
Nuclease S1 digestion
ONOO�-DNA was characterized by nuclease S1 treatment followed by agarose gel electrophoresis
(Matsuo and Ross, 1987). The digesting capability of nuclease S1 under our experimental conditions
was checked using heat denatured calf thymus DNA. It was almost completely digested, whereas no
digestion was observed in the case of purified native DNA. One microgram each of native and ONOO�-DNAwas incubated with nuclease S1 (20 units/Ag DNA in acetate buffer, pH 5.0, at 37 8C for 30 min).
The reaction was stopped by adding a 1/10 volume of 0.2 M EDTA, pH 8.0 and the sample
electrophoresed on 1% agarose gel for 2 h at 30 mA. The nucleic acid bands were visualized under UV
light after staining with ethidium bromide.
Reaction of plasmid DNA with peroxynitrite
The conversion of covalently closed circular-double stranded supercoiled DNA (form I) to a
relaxed open circle (form II) was used to investigate DNA single strand breaks induced by the

K. Dixit et al. / Life Sciences 77 (2005) 2626–2642 2629
combined action of DEA-NO and 1,4-hydroquinone. The experiment was carried out by
incubating plasmid PUC 18 DNA (100 Ag) in 100 mM PBS, pH 7.4 containing 0.1 mM
DTPA and either 0.1 mM DEA-NO alone or 0.1 mM 1,4-hydroquinone alone, or these two
compounds in combination at 37 8C for 1 h (final volume 10 ml). The reaction was stopped
by the addition of 2 Al sample buffer and the samples electrophoresed on 1% agarose gel for
2 h at 30 mA. The nucleic acid bands were visualized under UV light after ethidium bromide
staining.
Thermal denaturation studies
Thermal denaturation of native and ONOO�-DNA under identical conditions was evaluated by a
temperature scan from 30 8C to 95 8C at an increment of 1 8C/min on a Shimadzu UV-240
spectrophotometer equipped with a temperature programmer and controller assembly. The change in
absorbance at 260 nm was recorded and melting temperature (Tm) of the samples calculated (Ara and
Ali, 1992).
Quenching studies
The effects of various quenchers like superoxide dismutase (SOD) nitric oxide-trapping agent
(carboxy-PTIO at 10 mM), hydroxyl radical scavenger (d-mannitol at 10 mM), antioxidants (ascorbic
acid at 10 mM and uric acid at 0.2 mM) and a metal ion chelator (desferrioxamine at 10 mM) on DNA
modification were studied by incubating DNA (50 Ag) in 100 mM PBS pH 7.4 containing 0.1 mM
DTPA with 0.1 mM DEA-NO plus 0.1 mM 1,4-hydroquinone at 37 8C for 3 h. Percent quenching was
calculated from absorbance at A260 nm.
Immunization schedule
The immunization of random bred, female, New Zealand white rabbits was performed as described
previously (Hasan et al., 1991). Briefly, rabbits (n=4; two each for native and ONOO�-DNAantigens) were immunized intramuscularly at multiple sites with 50 Ag of antigen complexed with
methylated BSA in the ratio of 1:1 (w/w) and emulsified with an equal volume of Freund’s complete
adjuvant. The animals were boosted in Freund’s incomplete adjuvant at weekly intervals for 7 weeks
with the same amount of antigen. Test bleeds were performed 7 days post boost, which gave,
appropriate titre of the antibody. The animals were bled and the serum separated from the blood
(preimmune and immune) was heated at 56 8C for 30 min to inactivate complement proteins and
stored at �20 8C.
Purification of IgG
Immunoglobulin G was isolated from immune sera on Protein A-Sepharose CL-4B column. Sera
diluted (1:1) with PBS (pH 7.4) were passed through the column. After washing the unbound proteins
with PBS (pH 7.4) the bound IgG was eluted with 0.58% acetic acid and 0.85% sodium chloride (Alam
et al., 1993). To prevent the effect of acidic pH of eluting buffer on IgG, the eluate was collected in 1.0
ml of 1 M Tris–HCl buffer (pH 8.5).

K. Dixit et al. / Life Sciences 77 (2005) 2626–26422630
Enzyme linked immunosorbent assay (ELISA)
Antibodies against native and ONOO�-DNA were detected and quantitated by ELISA performed
on flat bottom 96-wells, polystyrene immunoplates (Nunc, maxisorp) as described previously (Ali
and Alam, 2002). Briefly the plates were coated with 100 Al of antigen (2.5 Ag/ml) for 2 h at room
temperature and overnight at 4 8C. After washing three times with TBS-T (20 mM Tris, 2.68 mM
KCl, 150 mM NaCl, pH 7.4 containing 0.05% Tween-20), unoccupied sites were blocked with 150
Al of 1.5% BSA in TBS (10 mM Tris, 150 mM NaCl, pH 7.4) for 4–6 h at room temperature. The
test serum serially diluted in TBS-T (100 Al/well) was absorbed for 2 h at room temperature and
overnight at 4 8C. Bound antibodies were assayed with anti-rabbit IgG alkaline phosphatase
conjugate using p-nitrophenyl phosphate as conjugate. The absorbance of each well was monitored
at 410 nm on an automatic microplate reader. Proper controls were included in each set of
experiments.
Competition ELISA
Antibody specificity was ascertained by competitive binding assay (Dixit and Ali, 2001). Varying
concentrations of inhibitors (0–20 Ag/ml) were allowed to interact with a constant quantity of antibody
(50 Ag/ml IgG or 1/10 diluted serum) for 2 h at room temperature and overnight at 4 8C. The mixture
was added to antigen coated plates and the bound antibody was diluted as described in the direct binding
ELISA. Inhibition was expressed as the amount of inhibitor required for 50% elimination in the antibody
binding to the solid phase antigen.
Band shift assay
For the visual detection of antigen–antibody interaction and the formation of immune complexes,
band shift assay was performed on 1% agarose (Dixit and Ali, 2004). Varying amounts (0–60 Ag) of IgGwere incubated with a fixed amount (0.5 Ag) of antigen for 2 h at 37 8C and overnight at 4 8C. Theresulting immune complex was electrophoresed on 1% agarose for 2 h at 30 mA in 40 mM Tris-acetate-9
mM EDTA buffer (TAE), pH 8.0. The gel was stained with ethidium bromide and photographed under
UV illumination.
Results
Human placental DNA purified free of proteins, RNA and single stranded regions (Ali et al., 1985)
was exposed to peroxynitrite generated by DEA-NO plus 1,4-hydroquinone. In order to optimize the
dose of DEA-NO and 1,4-hydroquinone for peroxynitrite generation, the reaction was carried out with
varying concentrations (0.1 mM–0.5 mM) of both the chemicals. The results were analyzed by UV-
absorption spectra (data not shown). DNA subjected to 0.5 mM of DEA-NO and 0.5 mM of 1,4-
hydroquinone showed moderate modification, whereas at lower concentrations of 0.4 to 0.1 mM the
modification was found increased. DNA subjected to 0.1 mM DEA-NO and 0.1 mM 1,4 hydroquinone
showed maximum modification. However, when the concentrations were reduced below 0.1 mM, the
results were not consistent vis-a-vis DNA damage. All the experiments were run in duplicates. Based on
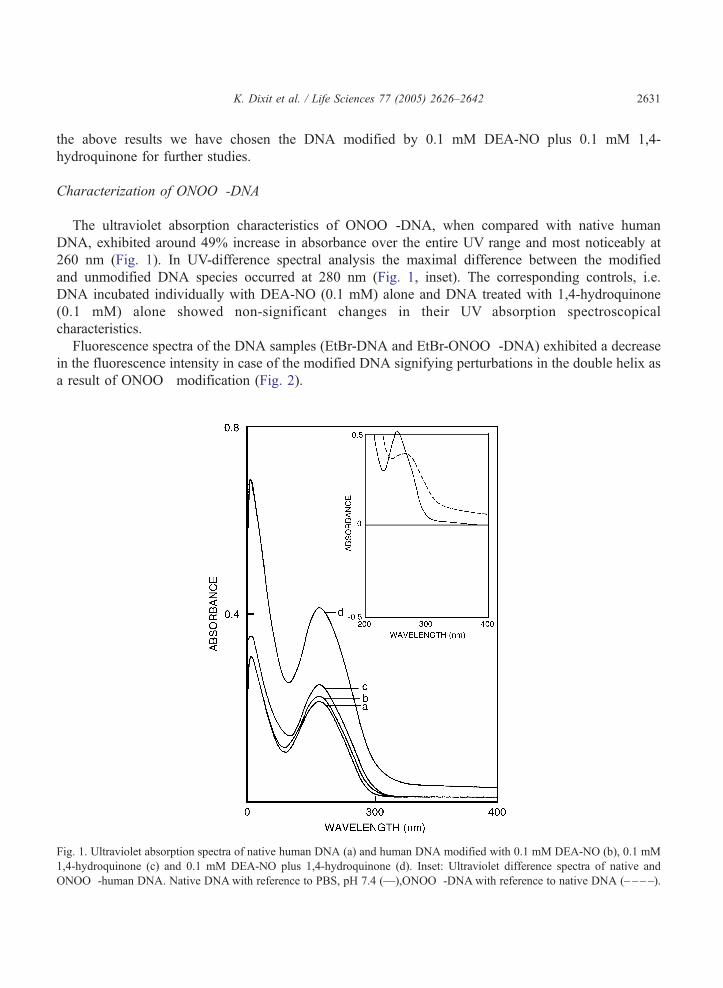
K. Dixit et al. / Life Sciences 77 (2005) 2626–2642 2631
the above results we have chosen the DNA modified by 0.1 mM DEA-NO plus 0.1 mM 1,4-
hydroquinone for further studies.
Characterization of ONOO�-DNA
The ultraviolet absorption characteristics of ONOO�-DNA, when compared with native human
DNA, exhibited around 49% increase in absorbance over the entire UV range and most noticeably at
260 nm (Fig. 1). In UV-difference spectral analysis the maximal difference between the modified
and unmodified DNA species occurred at 280 nm (Fig. 1, inset). The corresponding controls, i.e.
DNA incubated individually with DEA-NO (0.1 mM) alone and DNA treated with 1,4-hydroquinone
(0.1 mM) alone showed non-significant changes in their UV absorption spectroscopical
characteristics.
Fluorescence spectra of the DNA samples (EtBr-DNA and EtBr-ONOO�-DNA) exhibited a decrease
in the fluorescence intensity in case of the modified DNA signifying perturbations in the double helix as
a result of ONOO� modification (Fig. 2).
Fig. 1. Ultraviolet absorption spectra of native human DNA (a) and human DNA modified with 0.1 mM DEA-NO (b), 0.1 mM
1,4-hydroquinone (c) and 0.1 mM DEA-NO plus 1,4-hydroquinone (d). Inset: Ultraviolet difference spectra of native and
ONOO�-human DNA. Native DNAwith reference to PBS, pH 7.4 (—),ONOO�-DNAwith reference to native DNA (– – – –).
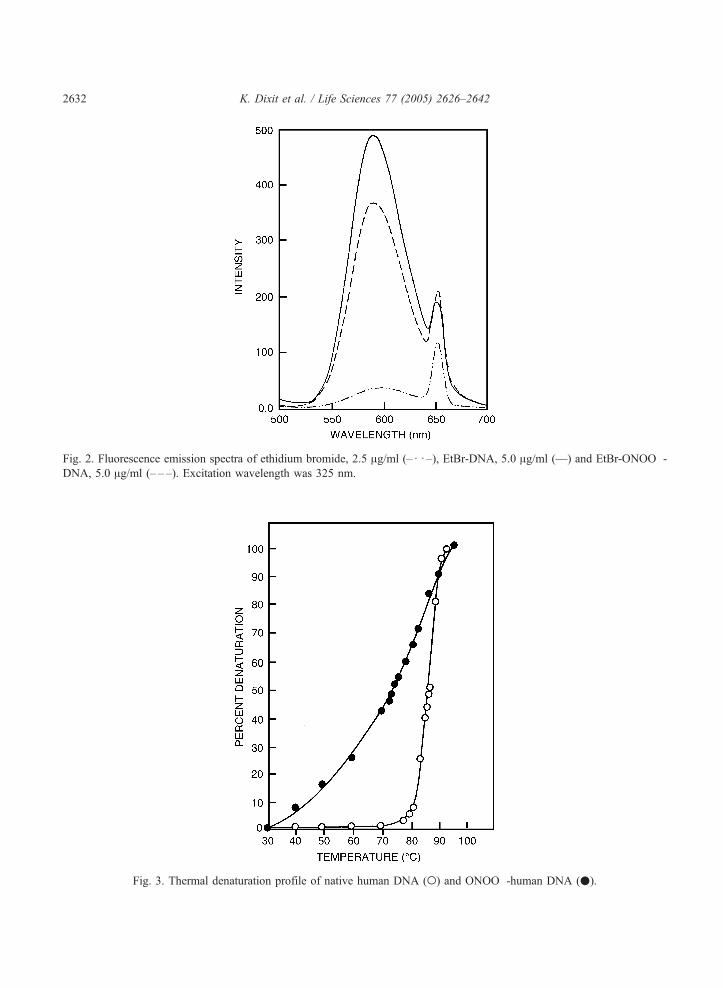
Fig. 2. Fluorescence emission spectra of ethidium bromide, 2.5 Ag/ml (–d d –), EtBr-DNA, 5.0 Ag/ml (—) and EtBr-ONOO�-
DNA, 5.0 Ag/ml (– – –). Excitation wavelength was 325 nm.
Fig. 3. Thermal denaturation profile of native human DNA (o) and ONOO�-human DNA (.).
K. Dixit et al. / Life Sciences 77 (2005) 2626–26422632

Table 1
Ultraviolet and thermal denaturation characteristics of native and ONOO�-modified human DNA
Parameter Native DNA Modified DNA
Absorbance ratio (A260/A280) 1.76 1.4
Hyperchromicity (%) at 95 8C 36.5 24
Melting temperature (Tm), 8C 86.0 71
Onset of duplex melting, 8C 78 31.5
K. Dixit et al. / Life Sciences 77 (2005) 2626–2642 2633
The melting profile of native and ONOO�-DNA (Fig. 3) was analyzed between the temperature range
30 8C and 95 8C. Increase in absorbance at 260 nm was taken as a measure of helix denaturation. The
process was characterized by determining the percent DNA in denatured state as a function of increasing
temperature and then computing the melting temperature (Tm). The Tm for ONOO�-DNAwas found to
be 71 8C and that of its unmodified form 86 8C indicating a net decrease of 15 8C in the Tm for modified
DNA. Moreover, the melting curve for modified DNA indicated a progressive and heterogeneous local
melting with the increase in temperature as compared to more homogeneous global melting in the
control. Physicochemical characteristics of native and ONOO�-DNA are summarized in Table 1.
To further evaluate the effect of peroxynitrite on human DNA, agarose gel electrophoresis was
performed. The electrophoretic pattern of DNA modified individually by DEA-NO and 1,4-
hydroquinone as well as by their combined action is shown in Fig. 4. DNA modified by ONOO�
migrated as a discrete band minus the characteristic stretch exhibited by the native DNA. Whereas DNA
treated individually to DEA-NO and 1,4-hydroquinone showed similar migration as native DNA.
Fig. 4. Agarose gel-electrophoresis of native and ONOO�-human DNA. Lanes: (1) Native DNA; (2) DNA modified with 0.1
mM DEA-NO; (3) DNA modified with 0.1 mM 1,4-hydroquinine; (4) DNA modified with 0.1 mM DEA-NO plus 1,4-
hydroquinone. DNA concentration was 0.5 Ag/ml in all the lanes.
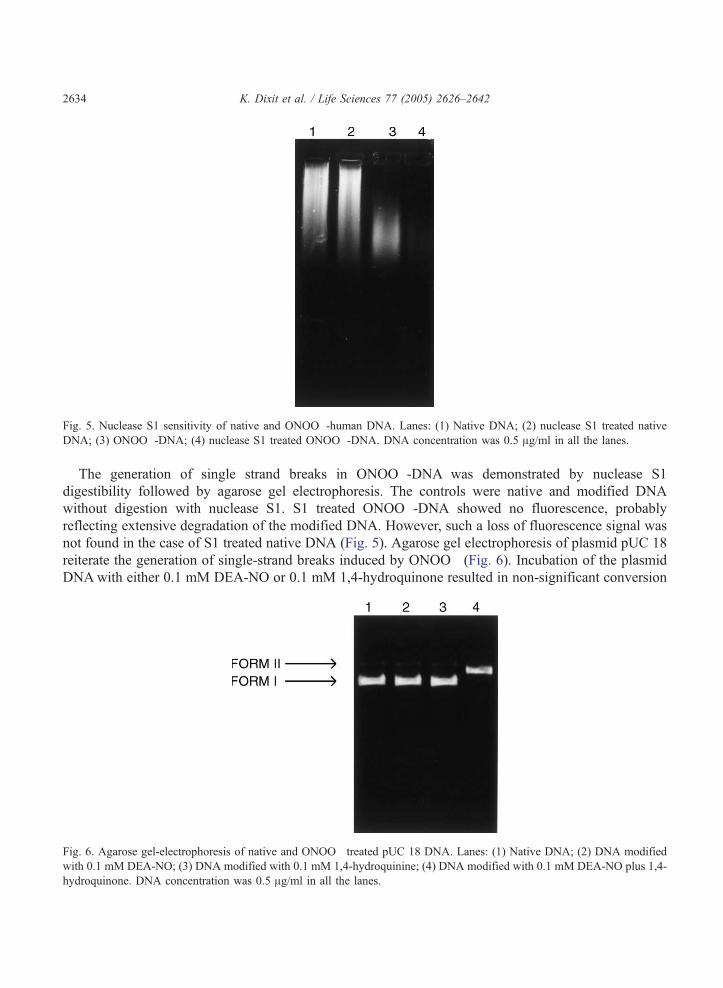
Fig. 5. Nuclease S1 sensitivity of native and ONOO�-human DNA. Lanes: (1) Native DNA; (2) nuclease S1 treated native
DNA; (3) ONOO�-DNA; (4) nuclease S1 treated ONOO�-DNA. DNA concentration was 0.5 Ag/ml in all the lanes.
K. Dixit et al. / Life Sciences 77 (2005) 2626–26422634
The generation of single strand breaks in ONOO�-DNA was demonstrated by nuclease S1
digestibility followed by agarose gel electrophoresis. The controls were native and modified DNA
without digestion with nuclease S1. S1 treated ONOO�-DNA showed no fluorescence, probably
reflecting extensive degradation of the modified DNA. However, such a loss of fluorescence signal was
not found in the case of S1 treated native DNA (Fig. 5). Agarose gel electrophoresis of plasmid pUC 18
reiterate the generation of single-strand breaks induced by ONOO� (Fig. 6). Incubation of the plasmid
DNAwith either 0.1 mM DEA-NO or 0.1 mM 1,4-hydroquinone resulted in non-significant conversion
Fig. 6. Agarose gel-electrophoresis of native and ONOO� treated pUC 18 DNA. Lanes: (1) Native DNA; (2) DNA modified
with 0.1 mM DEA-NO; (3) DNA modified with 0.1 mM 1,4-hydroquinine; (4) DNA modified with 0.1 mM DEA-NO plus 1,4-
hydroquinone. DNA concentration was 0.5 Ag/ml in all the lanes.
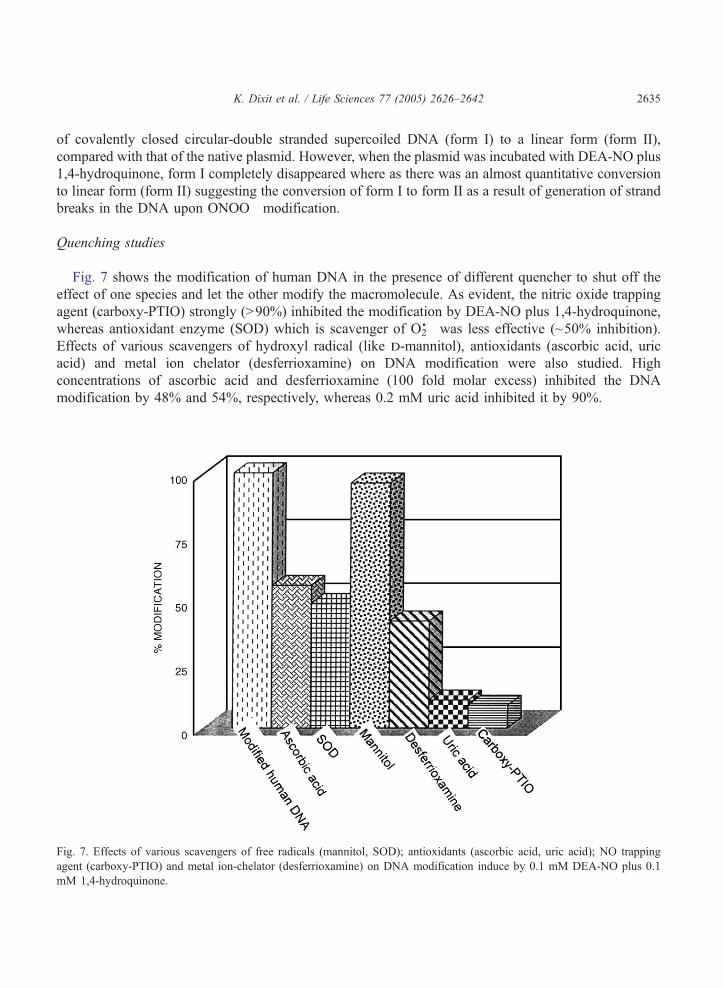
K. Dixit et al. / Life Sciences 77 (2005) 2626–2642 2635
of covalently closed circular-double stranded supercoiled DNA (form I) to a linear form (form II),
compared with that of the native plasmid. However, when the plasmid was incubated with DEA-NO plus
1,4-hydroquinone, form I completely disappeared where as there was an almost quantitative conversion
to linear form (form II) suggesting the conversion of form I to form II as a result of generation of strand
breaks in the DNA upon ONOO� modification.
Quenching studies
Fig. 7 shows the modification of human DNA in the presence of different quencher to shut off the
effect of one species and let the other modify the macromolecule. As evident, the nitric oxide trapping
agent (carboxy-PTIO) strongly (N90%) inhibited the modification by DEA-NO plus 1,4-hydroquinone,
whereas antioxidant enzyme (SOD) which is scavenger of O2!� was less effective (~50% inhibition).
Effects of various scavengers of hydroxyl radical (like d-mannitol), antioxidants (ascorbic acid, uric
acid) and metal ion chelator (desferrioxamine) on DNA modification were also studied. High
concentrations of ascorbic acid and desferrioxamine (100 fold molar excess) inhibited the DNA
modification by 48% and 54%, respectively, whereas 0.2 mM uric acid inhibited it by 90%.
Fig. 7. Effects of various scavengers of free radicals (mannitol, SOD); antioxidants (ascorbic acid, uric acid); NO trapping
agent (carboxy-PTIO) and metal ion-chelator (desferrioxamine) on DNA modification induce by 0.1 mM DEA-NO plus 0.1
mM 1,4-hydroquinone.
K. Dixit et al. / Life Sciences 77 (2005) 2626–26422636
Antigenicity of ONOO�-DNA
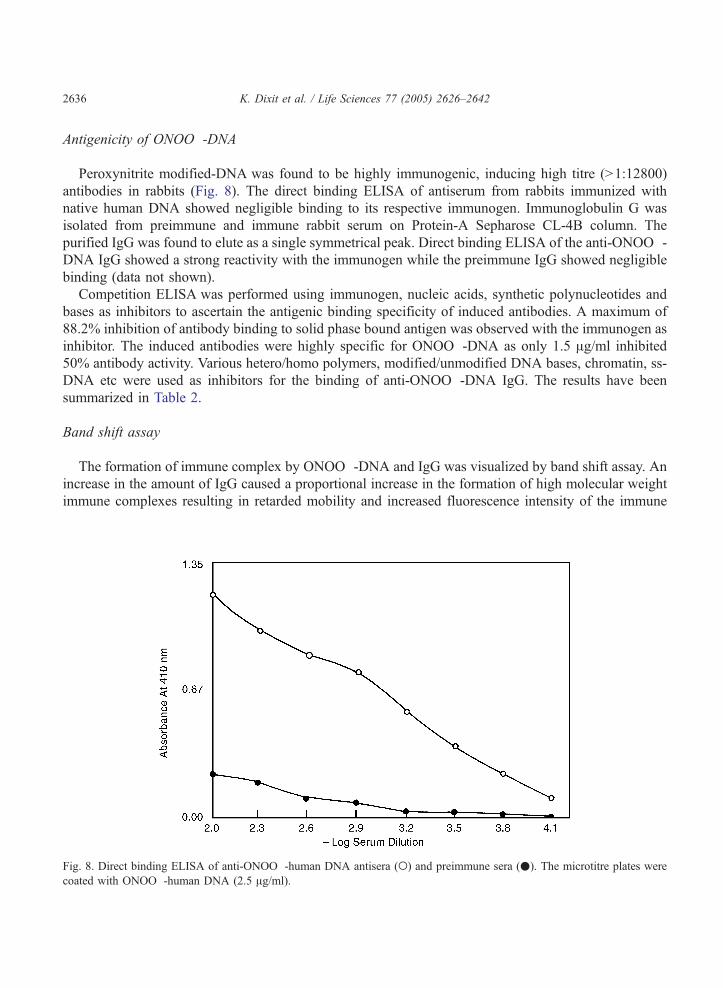
Peroxynitrite modified-DNA was found to be highly immunogenic, inducing high titre (N1:12800)
antibodies in rabbits (Fig. 8). The direct binding ELISA of antiserum from rabbits immunized with
native human DNA showed negligible binding to its respective immunogen. Immunoglobulin G was
isolated from preimmune and immune rabbit serum on Protein-A Sepharose CL-4B column. The
purified IgG was found to elute as a single symmetrical peak. Direct binding ELISA of the anti-ONOO�-DNA IgG showed a strong reactivity with the immunogen while the preimmune IgG showed negligible
binding (data not shown).
Competition ELISA was performed using immunogen, nucleic acids, synthetic polynucleotides and
bases as inhibitors to ascertain the antigenic binding specificity of induced antibodies. A maximum of
88.2% inhibition of antibody binding to solid phase bound antigen was observed with the immunogen as
inhibitor. The induced antibodies were highly specific for ONOO�-DNA as only 1.5 Ag/ml inhibited
50% antibody activity. Various hetero/homo polymers, modified/unmodified DNA bases, chromatin, ss-
DNA etc were used as inhibitors for the binding of anti-ONOO�-DNA IgG. The results have been
summarized in Table 2.
Band shift assay
The formation of immune complex by ONOO�-DNA and IgG was visualized by band shift assay. An
increase in the amount of IgG caused a proportional increase in the formation of high molecular weight
immune complexes resulting in retarded mobility and increased fluorescence intensity of the immune
Fig. 8. Direct binding ELISA of anti-ONOO�-human DNA antisera (o) and preimmune sera (.). The microtitre plates were
coated with ONOO�-human DNA (2.5 Ag/ml).
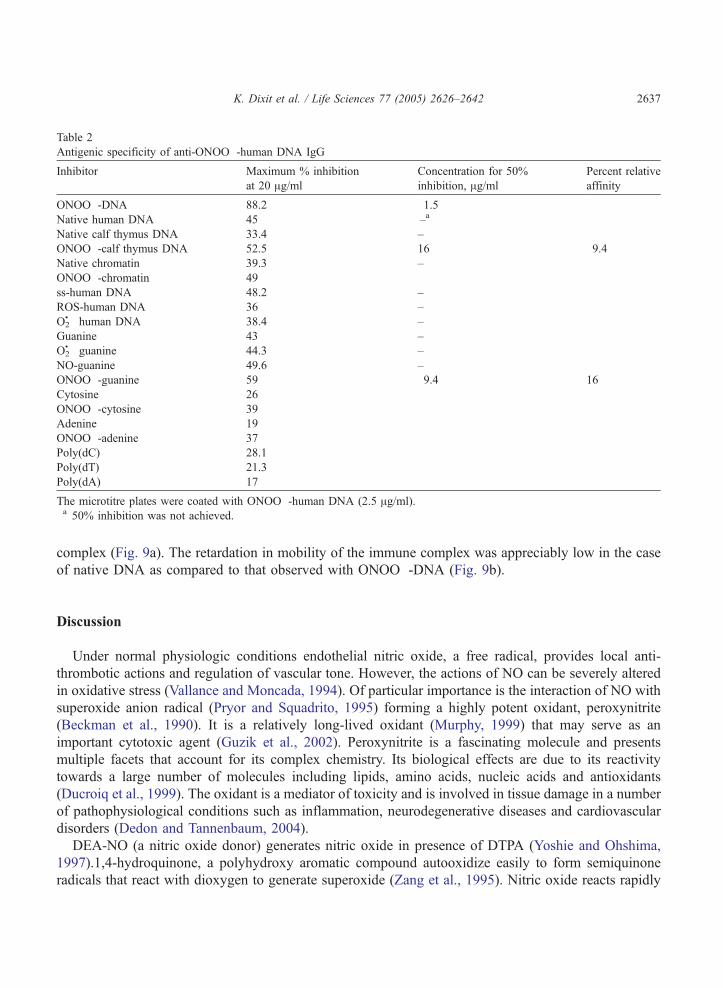
Table 2
Antigenic specificity of anti-ONOO�-human DNA IgG
Inhibitor Maximum % inhibition
at 20 Ag/ml
Concentration for 50%
inhibition, Ag/ml
Percent relative
affinity
ONOO�-DNA 88.2 1.5
Native human DNA 45 –a
Native calf thymus DNA 33.4 –
ONOO�-calf thymus DNA 52.5 16 9.4
Native chromatin 39.3 –
ONOO�-chromatin 49
ss-human DNA 48.2 –
ROS-human DNA 36 –
O2!� human DNA 38.4 –
Guanine 43 –
O2!� guanine 44.3 –
NO-guanine 49.6 –
ONOO�-guanine 59 9.4 16
Cytosine 26
ONOO�-cytosine 39
Adenine 19
ONOO�-adenine 37
Poly(dC) 28.1
Poly(dT) 21.3
Poly(dA) 17
The microtitre plates were coated with ONOO�-human DNA (2.5 Ag/ml).a 50% inhibition was not achieved.
K. Dixit et al. / Life Sciences 77 (2005) 2626–2642 2637
complex (Fig. 9a). The retardation in mobility of the immune complex was appreciably low in the case
of native DNA as compared to that observed with ONOO�-DNA (Fig. 9b).
Discussion
Under normal physiologic conditions endothelial nitric oxide, a free radical, provides local anti-
thrombotic actions and regulation of vascular tone. However, the actions of NO can be severely altered
in oxidative stress (Vallance and Moncada, 1994). Of particular importance is the interaction of NO with
superoxide anion radical (Pryor and Squadrito, 1995) forming a highly potent oxidant, peroxynitrite
(Beckman et al., 1990). It is a relatively long-lived oxidant (Murphy, 1999) that may serve as an
important cytotoxic agent (Guzik et al., 2002). Peroxynitrite is a fascinating molecule and presents
multiple facets that account for its complex chemistry. Its biological effects are due to its reactivity
towards a large number of molecules including lipids, amino acids, nucleic acids and antioxidants
(Ducroiq et al., 1999). The oxidant is a mediator of toxicity and is involved in tissue damage in a number
of pathophysiological conditions such as inflammation, neurodegenerative diseases and cardiovascular
disorders (Dedon and Tannenbaum, 2004).
DEA-NO (a nitric oxide donor) generates nitric oxide in presence of DTPA (Yoshie and Ohshima,
1997).1,4-hydroquinone, a polyhydroxy aromatic compound autooxidize easily to form semiquinone
radicals that react with dioxygen to generate superoxide (Zang et al., 1995). Nitric oxide reacts rapidly
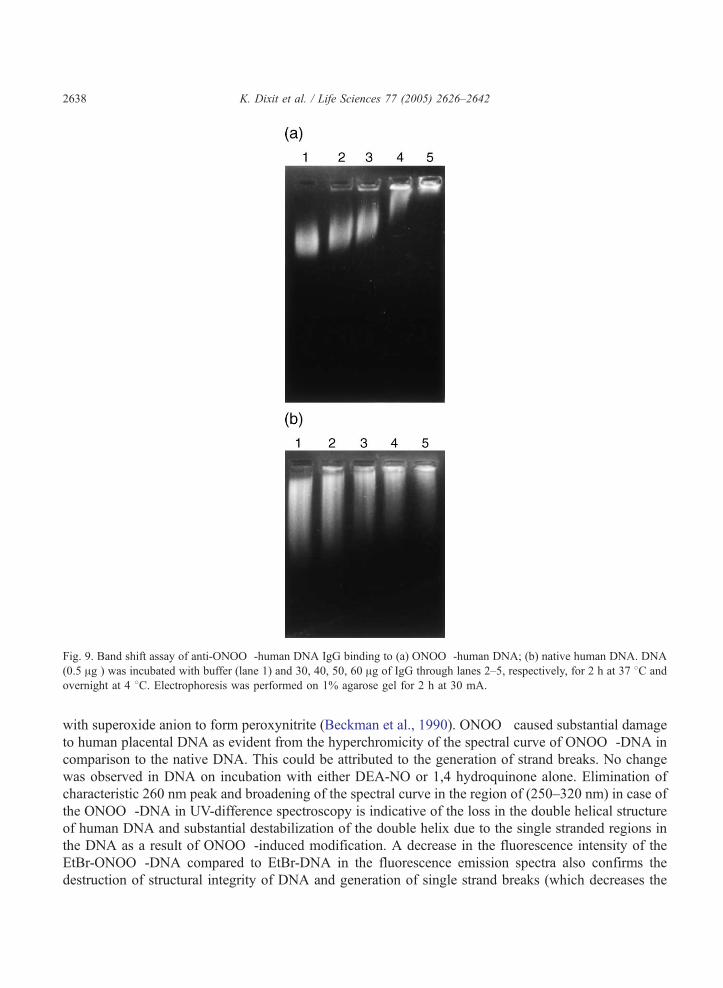
Fig. 9. Band shift assay of anti-ONOO�-human DNA IgG binding to (a) ONOO�-human DNA; (b) native human DNA. DNA
(0.5 Ag ) was incubated with buffer (lane 1) and 30, 40, 50, 60 Ag of IgG through lanes 2–5, respectively, for 2 h at 37 8C and
overnight at 4 8C. Electrophoresis was performed on 1% agarose gel for 2 h at 30 mA.
K. Dixit et al. / Life Sciences 77 (2005) 2626–26422638
with superoxide anion to form peroxynitrite (Beckman et al., 1990). ONOO� caused substantial damage
to human placental DNA as evident from the hyperchromicity of the spectral curve of ONOO�-DNA in
comparison to the native DNA. This could be attributed to the generation of strand breaks. No change
was observed in DNA on incubation with either DEA-NO or 1,4 hydroquinone alone. Elimination of
characteristic 260 nm peak and broadening of the spectral curve in the region of (250–320 nm) in case of
the ONOO�-DNA in UV-difference spectroscopy is indicative of the loss in the double helical structure
of human DNA and substantial destabilization of the double helix due to the single stranded regions in
the DNA as a result of ONOO�-induced modification. A decrease in the fluorescence intensity of the
EtBr-ONOO�-DNA compared to EtBr-DNA in the fluorescence emission spectra also confirms the
destruction of structural integrity of DNA and generation of single strand breaks (which decreases the

K. Dixit et al. / Life Sciences 77 (2005) 2626–2642 2639
efficiency of ethidium bromide intercalation). The alterations in the human DNA are also evident from
the decrease in melting temperature primarily due to generation of single strand breaks and due to altered
hydrogen bonding between base pairs.
Single strand breaks generated by ONOO� can arise from two processes—i.e. either from direct sugar
damage, which involves abstraction of hydrogen leading to formation of sugar radical (Szabo and
Ohshima, 1997) or indirectly via base damage which rapidly depurinates to generate abasic sites finally
culminating in single strand breaks. Thus, it can be concluded that the above modifications (sugar damage
and base modifications) are responsible for generation of single strand breaks in human DNA.
Appreciable evidence for the generation of strand breaks in the human DNA as a consequence of
ONOO� modification was gathered by nuclease S1 digestibility and plasmid nicking approach. DNA
strand breaks are induced synergistically when plasmid DNA is incubated in presence of both DEA-NO
and 1,4 hydroquinone. DEA-NO or 1,4 hydroquinone alone induced no appreciable change. Our results
are consistent with the observations that peroxynitrite, rather than NO is responsible for DNA strand
breakage (Yoshie and Ohshima, 1997).
Studies by various radical scavenging species indicate that single strand breaks and modification in
DNA induced by NO plus 1,4 hydroquinone can be inhibited by NO trapping agent (carboxy-PTIO) and
O2!� scavenger (SOD), suggesting that concurrent generation of NO and O2
!� is necessary for DNA
modification. Native DNA is a poor immunogen (Isenberg et al., 1994); however, conformational
isomers of the B-form have proved to be highly immunogenic (Jansson, 1985; Hasan and Ali, 1990; Ali
et al., 1985). The ONOO�-DNA was a potent immunizing stimulus inducing high titre antibodies in
rabbits which were highly immunogen specific but showing cross-reactivity with the native DNA.
Modification of DNA by ONOO� might have generated potential epitopes against which antibodies are
raised. The cross-reactivity with native human DNA, native calf thymus DNA may be due to recognition
of the common antigenic determinant, possibly the sugar phosphate backbone. Gel retardation data
further reiterated the preferential recognition of ONOO�-DNA over native DNA. Native calf thymus
DNA and native chromatin on peroxynitrite modification were more inhibitory than their unmodified
counterparts. The immune IgG was relatively more specific to peroxynitrite modified guanine, cytosine
and adenine. Peroxynitrite reacts significantly only with guanine which upon oxidation and nitration
leads to mutagenicity and strand breaks, respectively (Vermilov et al., 1995; Kennedy et al., 1997). In
addition, the induced antibodies showed recognition of various synthetic polynucleotides. Analysis of
the data indicates that anti-ONOO�-DNA IgG is immunogen specific and the varying degree of cross-
reactivity is due to sharing of common antigenic determinants which might be the modified bases or
peroxynitrite-induced lesions.
It has been proposed that tissue inflammation, immunostimulation (due to chronic infection) and
oxidative stress lead to the overproduction of nitric oxide and superoxide (Tretyakova et al., 2000).
These two species rapidly combine to yield peroxynitrite which in turn reacts with proteins (Beckman,
1996) resulting in their denaturation/modification or it could react with exposed nucleic acids leading to
single strand break generation which are potent signals for apoptosis (Levine, 1997). The subsequent
release of the altered DNA during apoptosis may enable it to act as an antigen inducing antibodies cross-
reactive with native DNA (Ahsan et al., 2003). The ONOO�-DNA thus acts as an autoantigen against
which anti-DNA antibodies could be generated.
Carcinogenesis is induced by altered DNA or tissue damage, mutations and chromosomal aberrations
(Gordon and Weitzman, 1990; Ames et al., 1993) Peroxynitrite levels are elevated in inflammation and
infection, and play an important role in carcinogenesis (Vamvakas and Schmidt, 1997; Ohshima, 2003).

K. Dixit et al. / Life Sciences 77 (2005) 2626–26422640
Peroxynitrite induced DNA damage leading to mutations has been strongly implicated in carcinogenesis
(Tretyakova et al., 2000; Dedon and Tannenbaum, 2004). If these mutations relate to critical genes,
initiation, and/or progression of cancer can occur. Our preliminary studies using antibodies from cancer
sera showed higher inhibition by ONOO�-DNA in comparison to native DNA when the immobilized
antigen was ONOO�-DNA. Studies on the binding specificity of cancer autoantibodies clearly
demonstrated that ONOO�-modified DNA is a better inhibitor of naturally occurring antibodies in most
of the cancer sera studied. Peroxynitrite appears to enhance the antigenicity of native DNA, thus
suggesting possible roles of ONOO�-induced neo-epitopes in the damaged DNA in production of
autoantibodies in cancer patients.
Acknowledgements
The authors are grateful to the University Grants Commission, Government of India for providing
financial assistance for this study vide grant no. F.3-16/2001(Sr-II). The authors are also grateful to Prof.
Rashid Ali for providing research facilities and for the critical analysis of the manuscript.
References
Ahmad, J., Ashok, B.T., Ali, R., 1997. Reactive oxygen species modified thymine and poly (dT) represent unique epitopes for
human anti-DNA autoantibodies. Immunology Letters 58, 69–74.
Ahsan, A., Ali, A., Ali, R., 2003. Oxygen free radicals and systemic autoimmunity. Clinical and Experimental Immunology
131, 398–404.
Alam, K., Ali, A., Ali, R., 1993. The effect of hydroxyl radical on the antigenicity of native DNA. Federation of European
Biochemical Societies Letters 319, 66–70.
Ali, R., Alam, K., 2002. Evaluation of antibodies against oxygen free radical modified DNA by ELISA. In: Armstrong, D.
(Ed.), Methods in Molecular Biology: Oxidative Stress Biomarkers and Antioxidant Protocols, vol. 186 (Ist edition).
Humana Press, New Jersey, pp. 171–181.
Ali, R., DerSimonian, H., Stollar, B.D., 1985. Binding of monoclonal anti-native DNA autoantibodies to DNA of varying size
and conformation. Molecular Immunology 22, 1415–1422.
Ames, B.N., Shigenaga, M.K., Hagen, T.M., 1993. Oxidants, antioxidants, and the degenerative diseases of aging. Proceedings
of the National Academy of Sciences 90, 7915–7922.
Ara, J., Ali, R., 1992. Reactive oxygen species modified DNA fragments of varying size are the preferred antigen for human
anti-DNA autoantibodies. Immunology Letters 34, 195–200.
Ara, J., Ali, R., 1993. Polynucleotide specificity of anti-ROS DNA antibodies. Clinical and Experimental Immunology 94,
134–139.
Ashok, B.T., Ali, R., 1999. Antigen binding characteristics of experimentally induced antibodies against hydroxyl radical
modified native DNA. Autoimmunity 29, 11–19.
Becker, J.C., Winkler, B., Klingert, S., Brocker, E.B., 1994. Anti-phospholipid syndrome associated with immunotherapy for
patients with melanoma. Cancer 73, 1621–1624.
Beckman, J.S., 1996. Oxidative damage and tyrosine nitration by peroxynitrite. Chemical Research in Toxicology 9,
836–844.
Beckman, J.S., Beckman, T.W., Chen, J., Marshall, B.A., Frecman, B.A., 1990. Apparent hydroxyl radical production by
peroxynitrite: implications for endothelial injury from nitric oxide and superoxide. Proceedings of the National Academy of
Sciences of the United States of America 87, 1620–1624.
Chagnand, J.L., Faiderbe, S., Geffard, M., 1992. Identification and immunochemical characterisation of IgA in sera of patients
with mammary tumour. International Journal of Cancer 50, 395–401.

K. Dixit et al. / Life Sciences 77 (2005) 2626–2642 2641
Dedon, P.C., Tannenbaum, S.R., 2004. Reactive nitrogen species in the chemical biology of inflammation. Archives of
Biochemistry and Biophysics 423, 12–22.
Dixit, K., Ali, R., 2001. Antigen binding characteristics of antibodies induced against nitric oxide modified plasmid DNA.
Biochimica et Biophysica Acta 1528, 1–8.
Dixit, K., Ali, R., 2004. Role of nitric oxide modified DNA in the etiopathogenesis of systemic lupus erythematosus. Lupus 13,
95–100.
Ducroiq, C., Blanchard, B., Pignatelli, B., Ohshima, H., 1999. Peroxynitrite: an endogenous oxidizing and nitrating agent.
Cellular and Molecular Life Sciences 55, 1068–1077.
Faiderbe, S., Chagnand, J.L., Geffard, M., 1992. Anti-phosphonisitide autoantibodies in sera of cancer patients: isotypic and
immunochemical characterisation. Cancer Letters 66, 35–41.
Gordon, L.L., Weitzman, S.A., 1990. Inflammation and cancer: role of phagocyte-generated oxidants in carcinogenesis. Blood
76, 655–663.
Guzik, T.J., West, N.E.J., Pillai, R., Taggart, D.P., Channon, K.M., 2002. Nitric oxide modulates superoxide release and
peroxynitrite formation in human blood vessels. Hypertension 39, 1088–1094.
Hasan, R., Ali, R., 1990. Antibody recognition of common epitopes on Z-DNA and native DNA brominated under high salt.
Biochemistry International 20, 1077–1188.
Hasan, R., Ali, A., Ali, R., 1991. Antibodies against DNA-psoralen crosslink recognizes unique conformation. Biochimica et
Biophysica Acta 1073, 507–513.
Huie, R.E., Padmaja, S., 1993. The reaction of NO with superoxide. Free Radical Research Communications 18, 195–199.
Isenberg, D.A., Ehrenstein, M.R., Longhurst, C., Kalsi, J., 1994. The origin, sequence, structure and sequence of developing
anti-DNA antibodies—a human perspective. Arthritis and Rheumatism 37, 169–180.
Jansson, G., 1985. Formation of antibodies to native DNA in rats after administration of native DNA treated with xanthine–
xanthine oxidase system. Free Radical Research Communications 1, 119–122.
Juedes, M.J., Wogan, G.N., 1996. Peroxynitrite-induced mutational spectra of pSP189 following replication in bacteria and
human cells. Mutation Research 349, 51–61.
Kennedy, L.J., Moore, K., Caulfield, J.L., Tannenbaum, S.R., Dedon, P.C., 1997. Quantitation of 8-oxoguanine and strand
breaks produced by four oxidizing agents. Chemical Research in Toxicology 10, 386–392.
Levine, A.J., 1997. p53, the cellular gatekeeper for growth and division. Cell 88, 323–331.
Matsuo, N., Ross, P.M., 1987. Measurement of interstrand cross-link frequency and distance between interruptions in DNA
exposed to 4,5V, 8-trimethylpsoralen and near-ultraviolet light. Biochemistry 26, 2001–2009.
Murphy, M.P., 1999. Nitric oxide and cell death. Biochimica et Biophysica Acta 1411, 401–414.
Ohshima, H., 2003. Genetic and epigenetic damage induced by reactive nitrogen species: implications in carcinogenesis.
Toxicology Letters 140, 99–104.
Ohshima, H., Bartsch, H., 1994. Chronic infections and inflammatory processes as cancer risk factors: possible role of nitric
oxide in carcinogenesis. Mutation Research 305, 253–264.
Pryor, W.A., Squadrito, G., 1995. Chemistry of peroxynitrite: a product from the reaction of nitric oxide with superoxide.
American Journal of Physiology 268, L699–L722.
Rachmilewitz, D., Stamler, D., Karmeli, F., Mullins, M.E., Singel, D.J., Loscalzo, J., Xavier, R.J., Podelsky, D.K., 1993.
Peroxynitrite induced rat colitis—a new model of colonic inflammation. Gastroenterology 105, 1681–1688.
Richeson, C.E., Mulder, P., Bowry, V.W., Ingold, K.U., 1998. The complex chemistry of peroxynitrite decomposition: new
insights. Journal of the American Chemical Society 120, 7211–7219.
Salgo, M.G., Bermudez, E., Squadrito, G.L., Pryor, W.A., 1995. Peroxynitrite causes DNA damage and oxidation of thiols in rat
thymocytes. Archives of Biochemistry and Biophysics 322, 500–505.
Szabo, C., Dawson, V.L., 1998. Role of poly (ADP-ribose) synthetase in inflammation and ischaemia reperfusion. Trends in
Pharmacological Sciences 19, 287–298.
Szabo, C., Ohshima, H., 1997. DNA damage induced by peroxynitrite: subsequent biological effects. Nitric Oxide 1, 373–385.
Tretyakova, N.Y., Burney, S., Tamir, B., Wishnok, J.S., Dedon, P.C., Wogan, G.N., Tannenbaum, S.R., 2000.
Peroxynitrite-induced DNA damage in the sup F gene: correlation with the mutational spectrum. Mutation Research
447, 287–303.
Vallance, P., Moncada, S., 1994. Nitric oxide: from mediator to medicines. Journal of Royal College of Physicians (London) 28,
209–219.
Vamvakas, S., Schmidt, H.H.H.W., 1997. Just say NO to cancer. Journal of the National Cancer Institute 89, 406–407.

K. Dixit et al. / Life Sciences 77 (2005) 2626–26422642
Vermilov, V., Rubio, J., Ohshima, H., 1995. 8-nitroguanine in DNA treated with peroxynitrite in vitro and its rapid removal
from DNA by depurination. Federation of European Biochemical Societies Letters 376, 207–210.
Yoshie, Y., Ohshima, H., 1997. Nitric oxide synergistically enhance DNA strand breakage induced by polyhydroxy
aromatic compounds, but inhibits that induced by the Fenton reaction. Archives of Biochemistry and Biophysics 342,
13–21.
Zang, L.Y., Stone, K., Pryor, W., 1995. Detection of free radicals in aqueous extracts of cigarette tar by electron spin resonance.
Free Radical Biology & Medicine 19, 161–167.