how does iron deficiency disrupt the electron flow in photosystem i of lettuce leaves?
TRANSCRIPT

P
Hl
NMa
b
c
a
ARRAA
KACFFP
I
cinoecf(ctch
dffNPc
0h
Journal of Plant Physiology 170 (2013) 1400– 1406
Contents lists available at ScienceDirect
Journal of Plant Physiology
journa l h o me page: www.elsev ier .com/ locate / jp lph
hysiology
ow does iron deficiency disrupt the electron flow in photosystem I of lettuceeaves?
ajoua Msilinia,b, Jemâa Esseminea,c, Maha Zaghdoudia,b, Johanne Harnoisa,okhtar Lachaâlb, Zeineb Ouerghib, Robert Carpentiera,∗
Groupe de Recherche en Biologie Végétale (GRBV), Université du Québec à Trois-Rivières, Trois-Rivières, Québec G9A 5H7, CanadaUnité de Physiologie et de Biochimie de la Tolérance au Sel chez les Plantes, Faculté des Sciences de Tunis, Campus Universitaire, 1060, TunisiaPlant Biology and Biotechnology Laboratory, Science Faculty of Tunis, Tunis University of El Manar, 1060, Tunisia
r t i c l e i n f o
rticle history:eceived 6 February 2013eceived in revised form 8 May 2013ccepted 8 May 2013vailable online 6 June 2013
eywords:
a b s t r a c t
The changes observed photosystem I activity of lettuce plants exposed to iron deficiency were inves-tigated. Photooxidation/reduction kinetics of P700 monitored as �A820 in the presence and absence ofelectron transport inhibitors and acceptors demonstrated that deprivation in iron decreased the popu-lation of active photo-oxidizable P700. In the complete absence of iron, the addition of plant inhibitors(DCMU and MV) could not recover the full PSI activity owing to the abolition of a part of P700 centers. Inleaves with total iron deprivation (0 �M Fe), only 15% of photo-oxidizable P700 remained. In addition, iron
+
bsorbance changesyclic electron flowe deficiencyluorescence measurements700 oxidation/re-reduction kineticsdeficiency appeared to affect the pool size of NADP as shown by the decline in the magnitude of the firstphase of the photooxidation kinetics of P700 by FR-light. Concomitantly, chlorophyll content graduallydeclined with the iron concentration added to culture medium. In addition, pronounced changes werefound in chlorophyll fluorescence spectra. Also, the global fluorescence intensity was affected. The abovechanges led to an increased rate of cyclic electron transport around PSI mainly supported by stromal
reductants.ntroduction
In plant chloroplasts, a portion of light energy absorbed byhlorophylls (Chls) and carotenoids is eventually stored as chem-cal energy. This conversion of energy from one form to another,amed also photosynthesis, is a complex process that dependsn cooperation between many pigment molecules and a group oflectron transfer proteins. In green plants, the pigment–proteinomplexes serve as an antenna complex, collecting light and trans-erring the energy to the reaction center (RC). Photosystem IIPSII) is composed of two antenna systems: inner antenna locatedlose to the RC and peripheral antenna. The inner antenna has
wo Chl–proteins CP47 and CP43 and the peripheral antenna,alled light harvesting complex II (LHCII), is the major light-arvesting protein of PS II. Photosystem I (PSI) is composed ofAbbreviations: Chl, chlorophyll; Cyt, cytochrome; DCMU, 3 (3,4-ichlorophenyl)-1,1-dimethylurea; EDTA, ethylenediaminetetracetic acid; FNR,erredoxin:NAD(P)H reductase; FQR, ferredoxin:plastoquinone reductase; FR,ar-red; LHCI and LHCII, light harvesting complexes I and II; MV, methyl viologen;dh, NAD(P)H dehydrogenase complex; P700, reaction center of photosystem I;AM, pulse amplitude modulation; PSI and PSII, photosystems I and II; RC, reactionenter; Rubisco, ribulose-1,5-diphosphate carboxylase/oxygenase.∗ Corresponding author. Tel.: +1 819 376 5011; fax: +1 819 376 5057.
E-mail address: [email protected] (R. Carpentier).
176-1617/$ – see front matter © 2013 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.jplph.2013.05.004
© 2013 Elsevier GmbH. All rights reserved.
light harvesting complex I (LHCI) and a core component (CCI).LHCI serves as an accessory antenna with some LHCII subunits toharvest light and funnel its energy to the reaction center of PSI(P700).
Iron is known as an essential element for plant growth and nutri-tion but the effect of Fe deficiency at the photosynthetic level is notwell known. Iron being an essential element for Chl biosynthesis,pigment–protein complexes are the first target of iron deficiency.It is commonly known that iron limitation induces leaf chlorosisdue to the Chl content decrease. Consequently, a pronounced irondeficiency results in a reduction of the amount of light harvest-ing complexes in both PSI and PSII (Morales et al., 1994; Timperioet al., 2007). In sugar beet, Morales et al. (1994) have shown thatiron deficiency leads to major decrease in the absolute valuesof Fm from PSII and PSI measured at 77 K. Similar results werefound with cyanobacteria grown in an iron deficient medium. Inthese cells, iron deficiency induced an alteration in the compo-sition of pigment–protein complexes forming the IsiA antennaprotein supercomplex (Chauhan et al., 2011). These changes canbe evidenced by low temperature fluorescence emission spectra(77 K) and by green gel electrophoresis. In cyanobacteria, iron
stressed cells are locked in state 1, associated with a charac-teristic increase of PSII fluorescence at 685 nm and a slightlydecrease of the PSI fluorescence peak at 725 nm (Ivanov et al., 2000,2006).
t Phys
etqbtitfatccctwes2dsektS
dvtmawi
M
P
(taeswcfPt
I
iiKt(
I
a1mc
N. Msilini et al. / Journal of Plan
PSI appears to be the first target of iron deficiency (Moseleyt al., 2002), perhaps because of its high Fe content as the electronransfer involves 12 iron atoms in three Fe–S centers. As a conse-uence, PSI can be less tolerant than PSII in the case of iron stressecause any iron deficiency may have a site of action on the accep-or side of PSI where are localized the majority of Fe–S centers. Inron deficient Cyanobacteria the PSI:PSII ratio decreases from 4:1o 1:1 (Straus, 1994) and in Acaryochloris cells the level of PSI wasound to decrease compared with that of PSII (Chen et al., 2005). Inddition, Timperio et al. (2007) showed that PSI is the most sensi-ive complex, with reduction of both the light-harvesting and coreomplex. In cyanobacteria iron deficiency caused an increase in theyclic electron flow around PSI as a consequence of the anatomi-al alterations and reduction of linear electron transport activityhrough PSII (Michel and Pistorius, 2004). Two alternative path-ays driven by PSI are known: the ferredoxin-dependent cyclic
lectron transport pathway and electron donation from solubletromal reductants such as NADPH or/and NADH (Bukhov et al.,001). In intact leaves, cyclic electron transport activity has beenetected indirectly by measuring the P700 re-reduction rate afterwitching off the far-red (FR)-light (Schreiber et al., 1988; Asadat al., 1992; Klughammer and Schreiber, 1998). The re-reductioninetics of P700 in dark-adapted leaves accelerates when elec-rons coming from the stroma are donated to PSI (Munekage andhikanai, 2005).
In the present study we have examined the effect of differentegree of iron deficiency in leaf disks from lettuce plants. The inivo photo-oxidation/reduction kinetics of P700 and the Chl con-ent were greatly affected. Besides a strong decline in active P700,
easurements of the dark re-reduction of oxidized P700 (P700+)fter the termination of FR-light have shown that the different path-ays of cyclic electron flow around PSI were largely modified under
ron deficiency conditions.
aterial and methods
lant growth
After germination in Petri dishes with distilled water for 7 d25 ◦C in the dark), lettuce (Lactuca sativa var. longifolia) plants wereransferred to pots containing 1 L of Hoagland’s nutrient solutionnd were kept for 21 d in control solution containing 20 �M FeIII-thylenediaminetetracetic acid (EDTA). Then, lettuce plants wereeparated into different groups where each group was irrigatedith the same Hoagland’s nutrient solution but with various iron
oncentrations ranging from 20 �M (control) to 0, 5, 10 and 15 �Mor 5 or 10 supplementary days. All solutions were replaced weekly.lants were cultivated in a growth chamber at 22/18 ◦C of day/nightemperature regime under a photoperiod of 16/8 h light/darkness.
solation of thylakoid membranes
Thylakoid membranes were isolated from lettuce leaves accord-ng to Joly et al. (2005). The resulting pellet was finally suspendedn 20 mM MES–NaOH (7.6), 400 mM sucrose, 10 mM NaCl, 20 mMCl, and 2 mM MgCl2 and stored at −80 ◦C until use. Chl concentra-
ion was calculated following the procedure outlined in Porra et al.1989).
nhibitor treatments
Whole untreated or iron deficient lettuce leaves were floated in
shaker (120 rpm) on a solution containing 3 (3,4-dichlorophenyl)-,1-dimethylurea (DCMU) (200 �M) alone or DCMU together withethyl viologen (MV) (100 �M) 90 min in darkness. In the latterase, MV was added 1 h after the leaves were floated in DCMU
iology 170 (2013) 1400– 1406 1401
solution to avoid oxidative damage elicited by this agent. The stocksolutions of DCMU were prepared in ethanol and MV in water. Allinhibitor solutions were prepared fresh. We note that the absorp-tion of inhibitors in lettuce leaves takes much time if compared toother material such as Arabidopsis (Essemine et al., 2011a, 2011b)or Cucumis sativus L. (Govindachary et al., 2007) leaves. This maybe owing to the complex structure of bundle sheath cells of lettuceleaves, which plays the role of a barrier facing the easy assimilationof inhibitors into leaves.
Redox state of P700
Photo-oxidation/reduction of P700 was monitored in darkadapted leaves at 23 ◦C as the light-induced absorbance changeat 820 nm (�A820) using the ED-P700DW dual wavelength unitconnected via a PAM-101 fluorometer (Walz, Effeltrich, Germany).The ED-P700DW detects strictly the differential absorbancechanges (810 − 860 nm) peaking at a single wavelength band820 nm ascribed for the P700+ cation radical absorption andremoves the plastocyanin absorbance changes (Klughammer andSchreiber, 1998). The photo-oxidation reaction was induced bya beam of FR-light (peaking wavelength is 735 nm; intensity of∼75 �mol m−2 s−1) obtained from a light emitting diode (RoithnerLasertchnik Gmbh, Vienna, Austria) passed through a supplemen-tary RG-9 filter (Schott, Mainz, Germany). The sampling rate was3 ms/point in Wincontrol software, which was used to acquire data.
Fluorescence measurements
The room temperature (298 K) and low temperature (77 K) spec-tra of fluorescence emission were measured with a Perkin-ElmerLS55 spectrofluorimeter equipped with a red-sensitive photo-multiplier R928. Chl fluorescence was excited at 436 nm. Theexcitation and emission spectral widths were fixed at 5 and 2.5 nm,respectively. Emission spectra were corrected according to thephotomultiplier sensitivity using the correction factor spectrumprovided by Perkin-Elmer. The Chl content of the samples wasadjusted to 5 �g mL−1. Low temperature spectra were measuredas reported previously (Rajagopal et al., 2002) in the presence of60% glycerol.
Results
Photo-oxidation kinetics of P700 by FR-light
Fig. 1A shows the oxidation/reduction kinetics of P700 mea-sured in dark-adapted lettuce leaf disks as the absorbance changesat 820 nm with a saturating 20-s FR-light (see Material and meth-ods section). The photo-oxidation kinetics of P700 in control leaves(20 �M Fe) followed a biphasic pattern (Fig. 1A, trace 20 �M). Thefirst fast phase that represents a rapid increase in the level of oxi-dized P700 (P700+) was followed by a dip. Then, the second slowerphase led to a steady state level much before 20 s. Less P700 couldbe photo-oxidized in plants submitted to iron limitation for 10 d. Asshown in Fig. 1B, the amplitude of the detected P700+ in untreatedand iron deficient plants declined as a function of the degree ofiron deprivation. This inhibition reached 85% with the completedeprivation of iron (0 �M). The magnitude of the rapid initial phasestrongly declined and was fully abolished at 0 �M iron (Fig. 1A). For0, 5 and 10 �M Fe, the steady-state level of P700+ was reached ear-lier in comparison with that observed in untreated plants (20 �MFe) or with moderate iron limitation (15 �M Fe).
Re-reduction of P700 after the termination of FR-light
The decay of P700+ observed after switching off the FR-light fit-ted well with the sum of three first order exponential decay curves
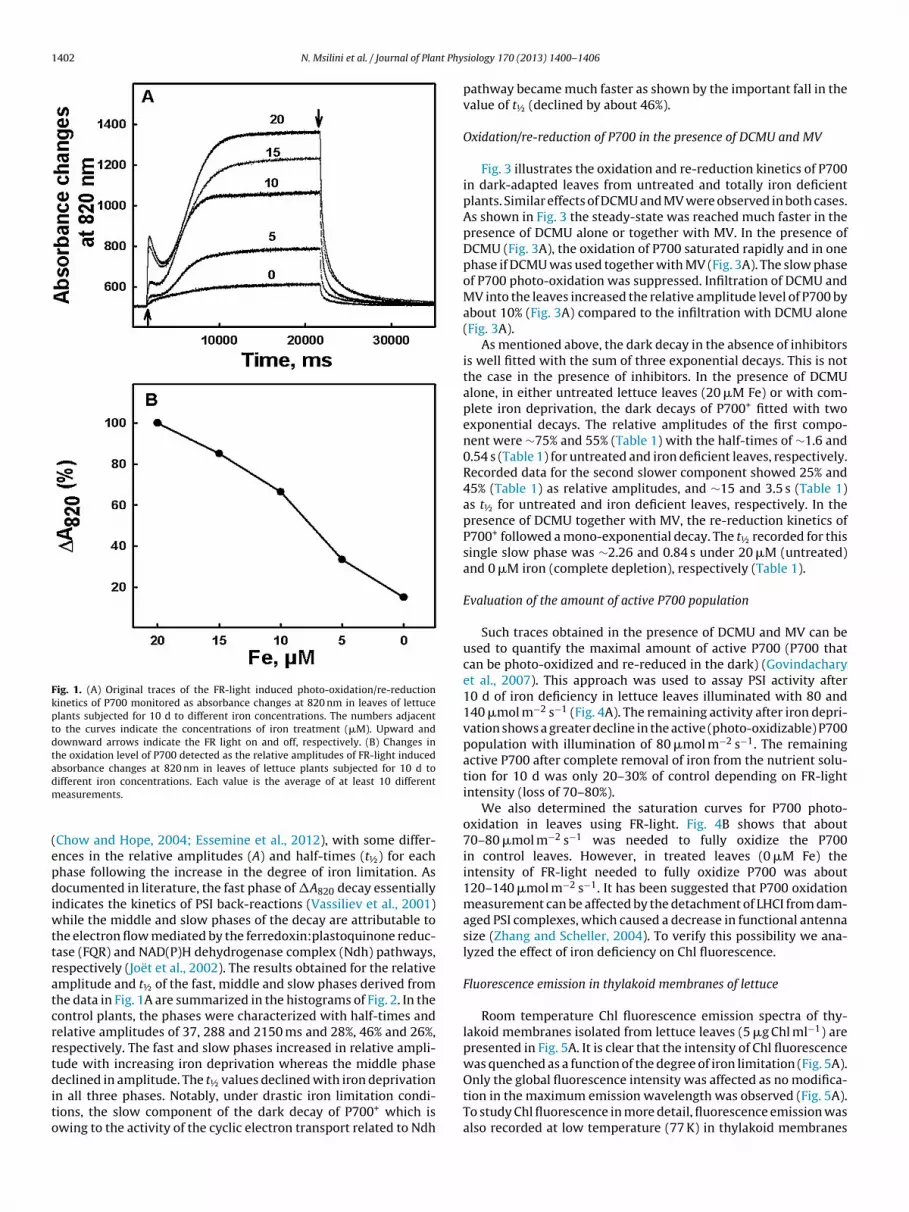
1402 N. Msilini et al. / Journal of Plant Phy
Fig. 1. (A) Original traces of the FR-light induced photo-oxidation/re-reductionkinetics of P700 monitored as absorbance changes at 820 nm in leaves of lettuceplants subjected for 10 d to different iron concentrations. The numbers adjacentto the curves indicate the concentrations of iron treatment (�M). Upward anddownward arrows indicate the FR light on and off, respectively. (B) Changes inthe oxidation level of P700 detected as the relative amplitudes of FR-light inducedabsorbance changes at 820 nm in leaves of lettuce plants subjected for 10 d todm
(epdiwttratcrrtdito
Only the global fluorescence intensity was affected as no modifica-
ifferent iron concentrations. Each value is the average of at least 10 differenteasurements.
Chow and Hope, 2004; Essemine et al., 2012), with some differ-nces in the relative amplitudes (A) and half-times (t½) for eachhase following the increase in the degree of iron limitation. Asocumented in literature, the fast phase of �A820 decay essentially
ndicates the kinetics of PSI back-reactions (Vassiliev et al., 2001)hile the middle and slow phases of the decay are attributable to
he electron flow mediated by the ferredoxin:plastoquinone reduc-ase (FQR) and NAD(P)H dehydrogenase complex (Ndh) pathways,espectively (Joët et al., 2002). The results obtained for the relativemplitude and t½ of the fast, middle and slow phases derived fromhe data in Fig. 1A are summarized in the histograms of Fig. 2. In theontrol plants, the phases were characterized with half-times andelative amplitudes of 37, 288 and 2150 ms and 28%, 46% and 26%,espectively. The fast and slow phases increased in relative ampli-ude with increasing iron deprivation whereas the middle phaseeclined in amplitude. The t½ values declined with iron deprivation
n all three phases. Notably, under drastic iron limitation condi-ions, the slow component of the dark decay of P700+ which iswing to the activity of the cyclic electron transport related to Ndh
siology 170 (2013) 1400– 1406
pathway became much faster as shown by the important fall in thevalue of t½ (declined by about 46%).
Oxidation/re-reduction of P700 in the presence of DCMU and MV
Fig. 3 illustrates the oxidation and re-reduction kinetics of P700in dark-adapted leaves from untreated and totally iron deficientplants. Similar effects of DCMU and MV were observed in both cases.As shown in Fig. 3 the steady-state was reached much faster in thepresence of DCMU alone or together with MV. In the presence ofDCMU (Fig. 3A), the oxidation of P700 saturated rapidly and in onephase if DCMU was used together with MV (Fig. 3A). The slow phaseof P700 photo-oxidation was suppressed. Infiltration of DCMU andMV into the leaves increased the relative amplitude level of P700 byabout 10% (Fig. 3A) compared to the infiltration with DCMU alone(Fig. 3A).
As mentioned above, the dark decay in the absence of inhibitorsis well fitted with the sum of three exponential decays. This is notthe case in the presence of inhibitors. In the presence of DCMUalone, in either untreated lettuce leaves (20 �M Fe) or with com-plete iron deprivation, the dark decays of P700+ fitted with twoexponential decays. The relative amplitudes of the first compo-nent were ∼75% and 55% (Table 1) with the half-times of ∼1.6 and0.54 s (Table 1) for untreated and iron deficient leaves, respectively.Recorded data for the second slower component showed 25% and45% (Table 1) as relative amplitudes, and ∼15 and 3.5 s (Table 1)as t½ for untreated and iron deficient leaves, respectively. In thepresence of DCMU together with MV, the re-reduction kinetics ofP700+ followed a mono-exponential decay. The t½ recorded for thissingle slow phase was ∼2.26 and 0.84 s under 20 �M (untreated)and 0 �M iron (complete depletion), respectively (Table 1).
Evaluation of the amount of active P700 population
Such traces obtained in the presence of DCMU and MV can beused to quantify the maximal amount of active P700 (P700 thatcan be photo-oxidized and re-reduced in the dark) (Govindacharyet al., 2007). This approach was used to assay PSI activity after10 d of iron deficiency in lettuce leaves illuminated with 80 and140 �mol m−2 s−1 (Fig. 4A). The remaining activity after iron depri-vation shows a greater decline in the active (photo-oxidizable) P700population with illumination of 80 �mol m−2 s−1. The remainingactive P700 after complete removal of iron from the nutrient solu-tion for 10 d was only 20–30% of control depending on FR-lightintensity (loss of 70–80%).
We also determined the saturation curves for P700 photo-oxidation in leaves using FR-light. Fig. 4B shows that about70–80 �mol m−2 s−1 was needed to fully oxidize the P700in control leaves. However, in treated leaves (0 �M Fe) theintensity of FR-light needed to fully oxidize P700 was about120–140 �mol m−2 s−1. It has been suggested that P700 oxidationmeasurement can be affected by the detachment of LHCI from dam-aged PSI complexes, which caused a decrease in functional antennasize (Zhang and Scheller, 2004). To verify this possibility we ana-lyzed the effect of iron deficiency on Chl fluorescence.
Fluorescence emission in thylakoid membranes of lettuce
Room temperature Chl fluorescence emission spectra of thy-lakoid membranes isolated from lettuce leaves (5 �g Chl ml−1) arepresented in Fig. 5A. It is clear that the intensity of Chl fluorescencewas quenched as a function of the degree of iron limitation (Fig. 5A).
tion in the maximum emission wavelength was observed (Fig. 5A).To study Chl fluorescence in more detail, fluorescence emission wasalso recorded at low temperature (77 K) in thylakoid membranes

N. Msilini et al. / Journal of Plant Physiology 170 (2013) 1400– 1406 1403
F s B, D
F ations
(bfnIg5
D
ttivi
THdb
ig. 2. Relative amplitudes, A in % (panels A, C and E) and half-times, t½ in ms (panelR light in dark adapted lettuce leaves submitted for 10 d to different iron concentr
Fig. 5B). At 77 K, the fluorescence emission spectrum is dominatedy two peaks at 685 and 731 nm which are ascribed to emissionrom PS II and PS I, respectively. After 10 d of treatment, the mag-itude of Chl fluorescence was decreased with iron deprivation.
n Fig. 5B, the spectra are normalized at 730 nm which shows thereater decline of the 685 nm band attributed to PSII and there is a
nm blue shift of the band at 730 nm with 0 �M iron.
iscussion
A number of mechanisms are known to control energy utiliza-ion in photosynthesis by modulating the anatomy and electron
ransfer in both photosystems. These mechanisms provide the flex-bility needed by the photosynthetic apparatus to adapt to a wideariety of environmental stress conditions. Iron availability is crit-cally important in determining the regulation and function ofable 1alf-times (t½ in s) and relative amplitudes (A in %) of each component of the re-reductiark adapted lettuce leaf discs floated in DCMU (200 �M) and MV (100 �M) as recorded
atches.
Fe (�M) Middle phase
20 0
A2 t½ A2 t½
DCMU 75 ± 4 1.60 ± 0.03 55 ± 4 0.54 ±DCMU + MV –/– –/– –/– –/–
and F) of each component of the re-reduction of P700+ following the termination of. Each value is the mean of 9–10 measurements with plants from different batches.
photosynthetic and diazotrophic growth (Shi et al., 2007). In thepresent work we studied the effect of iron deficiency on the PSIactivity of lettuce plants. Electron flow around PSI was also assessedusing the absorbance changes at 820 nm to follow the dark re-reduction of P700. The results indicate a strong influence of irondeficiency stress on the different cyclic electron transport pathwaysaround PSI and on the degree of PSI acceptor side limitation.
Iron depletion led to pronounced changes in the electron flowin PSI of lettuce leaves as determined by the oxidation–reductionkinetics of P700 in vivo. The magnitude of P700 photo-oxidation(Fig. 1A) in lettuce leaves irradiated with a 20 s of saturating FR-light strongly declined. After 10 d, about 80% of the PSI centers
were inactivated and may be destroyed in the plants cultivatedin a complete iron deprivation (Fig. 1A, trace 0 �M). The apparentlower oxidation level of P700 was also observed in iron deprivedcells of Synechococcus sp. (Ivanov et al., 2000). The initial fast phaseon of P700+ following the termination of FR-light in untreated and iron deficiencyin Fig. 3. Each value is the mean of 9–10 measurements with plants from different
Slow phase
20 0
A3 t½ A3 t½
0.03 25 ± 4 15.0 ± 0.1 45 ± 4 3.5 ± 0.3100 ± 10 2.3 ± 0.1 100 ± 10 0.8 ± 0.1

1404 N. Msilini et al. / Journal of Plant Physiology 170 (2013) 1400– 1406
Fig. 3. Original traces of the FR-light induced photo-oxidation and re-reductionkinetics of P700 monitored as absorbance changes at 820 nm in leaves of lettuceplants treated with 20 �M iron (A) or 0 �M iron (B). Leaf segments were incubatediDl
oiaiilpTri
slnibPaiireberPeat
Fig. 4. (A) Photo-oxidizable P700 in untreated and iron deficient lettuce leavesafter illumination with 80 and 140 �mol m−2 s−1 FR-light. The amplitude of �A820
obtained in leaves treated with DCMU and MV is used as the expression of PSI
n the dark in a solution with no inhibitor, with 200 �M DCMU, or with 200 �MCMU together with 100 �M MV. Upward and downward arrows indicate the FRight on and off, respectively.
f the FR-light induced photo-oxidation of P700 was abolishedn leaves submitted to strong iron deficiency. According to Joliotnd Joliot (2006), this phase is related to the pool size of NADP+
n vivo. Clearly, the size of the NADP+ pool was strongly diminishedn these leaves. A decrease of ribulose-1,5-diphosphate carboxy-ase/oxygenase (Rubisco) activity was previously demonstrated inlants treated with iron deficiency (Arulanantham et al., 1990).hus, the observed decrease of NADP+ pool is likely due to a lowatio of NADP+/NADPH associated with the NADPH accumulationn the absence of Benson–Calvin cycle activity.
The fast component of �A820 decay observed in the dark afterwitching off the FR-light was accelerated in iron-stressed lettuceeaves. Following the termination of 20 s FR-light, this compo-ent with relative amplitude of 28% and t½ of 37 ms was found in
ron-sufficient leaves to correspond to the kinetics of charge recom-ination, when there is no reducing pressure at the acceptor side ofSI (Vassiliev et al., 2001). Its relative amplitude was enhanced bybout 20% and the t½ declined to 24 ms in iron-deficient leaves. Thiss attributable to an increased yield of charge recombination as thentegrity of the PsaC subunit and other polypeptides of the stromalidge of PSI are strongly perturbed during iron deficiency (Yadavallit al., 2012a,b). Indeed, a three-fold decrease in the life-time of PSIack-reactions was observed after the extraction of FB (Vassilievt al., 2001). It has been demonstrated that the life-time of back-eactions sharply decreased with damage to the acceptor side of
SI centers (Golbeck and Bryant, 1991; Sonoike et al., 1995; Teichert al., 2000). The damaged acceptor side is probably responsible forn enhanced production of reactive oxygen species that affectedhe PSI complex and declined the amount of active P700 (Fig. 1).activity. Each data point is the average of 10 measurements. (B) FR-light satura-tion curve of P700 photo-oxidation in leaves of lettuce plants treated with differentiron concentrations.
Another aspect of the �A820 data modeled as three exponentialdecays is the marked changes in the t½ of the slow component thatcorresponds to the flux of electrons for P700 reduction through theNdh pathway. This component decayed in the dark with a t½ of2150 ms and 1166 ms in iron-sufficient and iron-deficient leaves,respectively. This reveals that the cyclic electrons flow through theNdh pathway was accelerated by about 1000 ms. In the presenceof saturating DCMU, two distinct phases of P700+ reduction wereobserved in lettuce leaves. Similar results were also reported byBukhov et al. (1999) for spinach leaves floated on a solution ofDCMU. According to Chow and Hope (2004), the first phase canbe attributed to electron donation from reduced ferredoxin to theintersystem pool. In the combined presence of DCMU and MV, asingle phase was observed. This slow phase which is insensitiveto MV has been associated to the electron donation from stromalNADPH to the PQ pool via Ndh (Corneille et al., 1998; Chow andHope, 2004). In such conditions, iron deficiency accelerated theflow of electron to P700+ through these two pathways (Table 1),indicating an increased capacity for cyclic electron transport in ironstressed leaves. The above trend was also observed in cyanobac-teria (Michel and Pistorius, 2004). It has been suggested that theincrease in cyclic electron flow around PSI under iron deficiencywas the consequence of the PSI-IsiA supercomplex in these cells(Michel et al., 2003) but this complex is absent in higher plants.
In the present case, the increased pool of NADPH as discussedabove provides stromal reductants in greater amount to sustain thecyclic pathway especially when the population of active PSI reac-tion centers is declined, which account for the increased relative
N. Msilini et al. / Journal of Plant Phys
F(o
aIpiTritcl2
acfpcdirewio(ide
ig. 5. Fluorescence emission spectra measured at room temperature (A) and at 77 KB) in isolated thylakoid membranes of lettuce plants submitted to different degreef iron limitation conditions. Each curve is the average of 12 different measurements.
mplitude and rate of the slow phase of P700 re-reduction (Fig. 2).t should be noted that the Cyt b6f complex, also needed in theathway of cyclic electron transport around PSI, hosts the Rieske
ron–sulfur center and is potentially damaged by iron depletion.he re-reduction of P700 in the population of PSI complexes thatemain active with low iron supply therefore involves the remain-ng undamaged Cyt b6f complexes. The enhanced electron flowhrough Ndh and FQR following iron deficiency as shown here isonsistent with the responses of plants subjected to other physio-ogical stress conditions (Govindachary et al., 2007; Bukhov et al.,004).
It has been shown that increased rates of cyclic electron flowround PSI leads to overestimation of the photo-oxidizable P700ontent (Kudoh and Sonoike, 2002; Bukhov et al., 2004). Thus, aaster P700+ re-reduction after iron depletion is likely to com-ete with re-oxidation of P700 by FR-light. In our experimentalonditions, the observed changes in light response for P700 oxi-ation showing an increased light requirement for saturation after
ron depletion (Fig. 4B) cannot be explained only by the increasedate of P700 reduction caused by this enhancement of the cycliclectron transfer. The Chl fluorescence quenching that increasedith iron depletion (Fig. 5A) and the decline in measurable P700
ndicates that strongly dissipative centers are formed with theccurrence of inactive PSI RC. Alternatively, Zhang and Scheller
2004) explained such increased light requirement by a decreasen functional antenna size caused by a detachment of LHCI fromamaged PSI complexes. This is also possible in our case. How-ver, as iron depletion inhibits Chl biosynthesis, iron deficiencyiology 170 (2013) 1400– 1406 1405
caused a direct depletion of antenna complexes as also reportedfor spinach (Timperio et al., 2007). A rapid loss of LHCI duringiron deficiency was also recently shown in Chlamydomonas rein-hardtii and rice (Yadavalli et al., 2012a,b). These results are ingood agreement with the observed general decline of Chl fluores-cence emission even though PSII antenna was more affected inthat respect. Depression of fluorescence emission of PSI was firstdescribed by Öquist (1974) and was ascribed to a depletion of PSIwithin the membrane. Guikema and Sherman (1983) also foundthat fluorescence emission at 716 was strongly influenced by irondeficiency. Antenna complexes harvest light energy and transferexcitation to the photosystem core. It is obvious that the primaryeffect of iron deficiency is the regression in the pigment synthesisand decreased content of LHCs that is likely the main source forthe increased light requirement reported here. During 10 d of irondeficiency treatment, the yellowing chlorosis increased as functionof the degree of iron limitation (not shown). The reduced Chl con-tent is the source of chlorosis (Chl deficiency) symptoms observedunder iron deficiency conditions (Spiller and Terry, 1980; Terry,1980; Straus, 1994).
The increased donation of electrons to PSI from a source alter-native to PSII in leaves submitted to iron deficiency may have somephysiological relevance. Accelerated donation of electrons fromstromal reductants to P700+ was also induced by different abioticstresses such as heat (Bukhov et al., 2000) or high NaCl concen-tration (Endo et al., 1995). Under these stress conditions as in thecase of iron stress, PSII activity is declined (Msilini et al., 2011) andtherefore a down-regulation of linear electron transport occurs.Even following moderate iron stress, the linear electron flow ismuch affected together with PSII activity (Msilini et al., 2011). Anincreased rate of cyclic electron flow around PSI appears as a pro-tective tool that may keep the energetic balance protecting ATPsynthesis (Heber and Walker, 1992). This protective role can besustained by PSI to some threshold only. Complete iron deficiencyfor longer time affects also PSI activity and subsequently leads tothe destruction of the whole photosynthetic machinery.
References
Arulanantham AR, Rao IM, Terry N. Limiting factors in photosynthesis. IV. Regener-ation of ribulose 1,5-bisphosphate limits photosynthesis at low photochemicalcapacity. Plant Physiol 1990;93:1465–75.
Asada K, Heber U, Schreiber U. Pool size of electrons that can be donated to P700+,as determined in intact leaves: donation to P700+ from stromal components viathe intersystem chain. Plant Cell Physiol 1992;33:927–32.
Bukhov NG, Wiese C, Neimanis S, Heber U. Heat sensitivity of chloroplasts andleaves: leakage of protons from thylakoids and reversible activation of cyclicelectron transport. Photosynth Res 1999;59:81–93.
Bukhov NG, Samson G, Carpentier R. Nonphotosynthetic reduction of the intersys-tem electron transport chain of chloroplast following heat stress. Steady-staterate. Photochem Photobiol 2000;72:351–7.
Bukhov NG, Samson G, Carpentier R. Non photosynthetic reduction of the intersys-tem electron transport chain of chloroplasts following heat stress. The pool sizeof stromal reductants. Photochem Photobiol 2001;74:438–43.
Bukhov NG, Govindachary S, Rajagopal S, Joly D, Carpentier R. Enhanced rates ofP700+ dark-reduction in leaves of Cucumis sativus L. photoinhibited at chillingtemperature. Planta 2004;218:852–61.
Chauhan D, Folea M, Jolley CC, Kouril R, Lubner CE, Lin S, et al. Novel photosyn-thetic strategy for adaptation to low-iron aquatic environments. Biochemistry2011;50:686–92.
Chen M, Bibby TS, Nield J, Larkuma A, Barber J. Iron deficiency induces a chlorophylld-binding Pcb antenna system around photosystem I in Acaryochloris marina.Biochim Biophys Acta 2005;1708:367–74.
Chow WS, Hope AB. Kinetics of reactions around the cytochrome bf complex studiedin intact leaf disks. Photosynth Res 2004;81:153–63.
Corneille S, Cournac L, Guedeney G, Havaux M, Peltier G. Reduction of the plas-toquinone pool by exogenous NADH and NADPH in higher plant chloroplasts.
Characterization of a NAD(P)H-plastoquinone oxidoreductase activity. BiochimBiophys Acta 1998;1363:59–69.Endo T, Schreiber U, Asada K. Suppression of quantum yield of photosystemII by hyperosmotic stress in Chlamydomonas reinhardtii. Plant Cell Physiol1995;36:1253–8.

1 t Phy
E
E
E
GG
G
H
I
I
J
J
J
K
K
M
M
M
M
406 N. Msilini et al. / Journal of Plan
ssemine J, Govindachary S, Ammar S, Bouzid S, Carpentier R. Abolition of photo-system I cyclic electron flow in Arabidopsis thaliana following thermal-stress.Plant Physiol Biochem 2011a;49:235–43.
ssemine J, Govindachary S, Ammar S, Bouzid S, Carpentier R. Functional aspectsof the photosynthetic light reactions in heat stressed Arabidopsis deficient indigalactosyl-diacylglycerol. J Plant Physiol 2011b;168:1526–33.
ssemine J, Govindachary S, Ammar S, Bouzid S, Carpentier R. Enhanced sensitiv-ity of the photosynthetic apparatus to heat stress in digalactosyl-diacylglyceroldeficient Arabidopsis. Environ Exp Bot 2012;80:16–26.
olbeck JH, Bryant DA. Photosystem I. Curr Topics Bioenerg 1991;16:83–177.ovindachary S, Bigras C, Harnois J, Joly D, Carpentier R. Changes in the mode of
electron flow to photosystem I following chilling-induced photoinhibition in aC3 plant, Cucumis sativus L. Photosynth Res 2007;94:333–45.
uikema JA, Sherman LA. Organization and function of chlorophyll in membranesof cyanobacteria during iron starvation. Plant Physiol 1983;73:250–6.
eber U, Walker D. Concerning a dual function of coupled cyclic electron transportin leaves. Plant Physiol 1992;100:1621–5.
vanov AG, Park YI, Miskiewicz E, Raven JA, Huner NPA, Öquist G. Iron stress restrictsphotosynthetic intersystem electron transport in Synechococcus sp. PCC 7942.FEBS Lett 2000;485:173–7.
vanov AG, Krol M, Sveshnikov D, Selstam E, Sandstrõm S, Koochek M, et al. Irondeficiency in cyanobacteria causes monomerization of photosystem I trimersand reduces the capacity for state transitions and the effective absorption crosssection of photosystem I in vivo. Plant Physiol 2006;141:1436–45.
oët T, Cournac L, Peltier G, Havaux M. Cyclic electron flow around photosystem I inC3 plants. In vivo control by the redox state of chloroplasts and involvement ofthe NADH-dehydrogenase complex. Plant Physiol 2002;128:760–9.
oliot P, Joliot A. Cyclic electron flow in C3 plants. Biochim Biophys Acta2006;1757:362–8.
oly D, Bigras C, Harnois J, Govindachary S, Carpentier R. Kinetic analyses of theOJIP chlorophyll fluorescence rise in thylakoid membranes. Photosynth Res2005;84:107–12.
lughammer C, Schreiber U. Measuring P700 absorbance changes in the near infra-red spectral region with a dual wavelength pulse modulation system. In: GarabG, editor. Photosynthesis: mechanisms and effects. Dordrecht: Kluwer AcademicPublishers; 1998. p. 4357–60.
udoh H, Sonoike K. Irreversible damage to photosystem I by chilling in the light:cause of the degradation of chlorophyll after returning to normal growth tem-perature. Planta 2002;215:541–8.
ichel KP, Pistorius EK. Adaptation of the photosynthetic electron transport chainin cyanobacteria to iron deficiency: the function of IdiA and IsiA. Physiol Plant2004;120:36–50.
ichel KP, Berry S, Hifney A, Pistorius EK. Adaptation to iron deficiency: a compar-ison between the cyanobacterium Synechococcus elongatus PCC 7942 wild-typeand a DpsA-free mutant. Photosynth Res 2003;75:71–84.
orales F, Abadía A, Belkhodja R, Abadía J. Iron deficiency-induced changes in the
photosynthetic pigment composition of field-grown Pyrus communis L. leaves.Plant Cell Environ 1994;17:1153–60.oseley JL, Allinger T, Herzog S, Hoerth P, Wehinger E, Merchant S, et al. Adaptationto Fe-deficiency requires remodelling of the photosynthetic apparatus. EMBO J2002;21:6709–20.
siology 170 (2013) 1400– 1406
Msilini N, Zaghdoudi M, Govindachary S, Lachaâl M, Ouerghi Z, Carpentier R.Inhibition of photosynthetic oxygen evolution and electron transfer from thequinone acceptor QA
− to QB by iron deficiency. Photosynth Res 2011;107:247–56.
Munekage Y, Shikanai T. Cyclic electron transport through photosystem I. PlantBiotechnol 2005;22:361–9.
Öquist G. Iron deficiency in the blue-green alga Anacystis nidulans: fluorescenceand absorption spectra recorded at 770 K. Physiol Plant 1974;31:55–8.
Porra RJ, Thompson WA, Kriedemann PE. Determination of accurate extinc-tion coefficients and simultaneous equations for assaying chlorophyll a andb extracted with four different solvents: verification of the concentration ofchlorophyll standards by atomic absorption spectroscopy. Biochim Biophys Acta1989;975:384–94.
Rajagopal S, Bukhov NG, Carpentier R. Changes in the structure ofchlorophyll–protein complexes and excitation energy transfer duringphotoinhibitory treatment of isolated photosystem I submembrane particles. JPhotochem Photobiol B 2002;67:194–200.
Schreiber U, Klughammer C, Neubauer C. Measuring P700 absorbance changesaround 830 nm with a new type of pulse modulation system. Z Naturforsch1988;43c:686–98.
Shi T, Sun Y, Falkowski PG. Effects of iron limitation on the expression of metabolicgenes in the marine cyanobacterium Trichodesmium erythraeum IMS101. Envi-ron Microbiol 2007;9:2945–56.
Sonoike K, Terashima I, Iwaki M, Itoh S. Destruction of photosystem I iron–sulfurcenters in leaves of Cucumis sativus L. by weak illumination at chilling temper-atures. FEBS Lett 1995;362:235–8.
Spiller S, Terry N. Limiting factors in photosynthesis. II. Iron stress diminishes photo-chemical capacity by reducing the number of photosynthetic units. Plant Physiol1980;65:121–5.
Straus NA. Iron deprivation: physiology and gene regulation. In: Bryant DA, editor.The molecular biology of cyanobacteria. Dordrecht, The Netherlands: Kluwer;1994. p. 731–50.
Teicher BH, Möller BL, Scheller HV. Photoinhibition of photosystem I in field-grownbarley (Hordeum vulgare L.): induction, recovery and acclimation. PhotosynthRes 2000;64:53–61.
Terry N. Limiting factors in photosynthesis. I. Use of iron stress to control photo-chemical capacity in vivo. Plant Physiol 1980;65:114–20.
Timperio AM, D’Amici GM, Barta C, Loreto F, Zolla L. Proteomics, pigment composi-tion, and organization of thylakoid membranes in iron-deficient spinach leaves.J Exp Bot 2007;58:3695–710.
Vassiliev IR, Antonkine ML, Golbeck JH. Iron–sulfur clusters in type I reaction centers.Biochim Biophys Acta 2001;1507:139–60.
Yadavalli V, Jolley CC, Malleda C, Thangaraj B, Fromme P, Subramanyam R. Alterationof proteins and pigments influence the function of photosystem I under irondeficiency from Chlamydomonas reinhardtii. PLoS ONE 2012a;7:e35084.
Yadavalli V, Neelam S, Rao ASVC, Reddy AR, Subramanyam R. Differential degra-
dation of photosystem I subunits under iron deficiency in rice. J Plant Physiol2012b;169:753–9.Zhang S, Scheller HV. Photoinhibition of photosystem I at chilling temper-ature and subsequent recovery in Arabidopsis thaliana. Plant Cell Physiol2004;45:1595–602.