heterologous stacking of prion protein peptides reveals structural details of fibrils and...
TRANSCRIPT

Heterologous Stacking of Prion Protein Peptides RevealsStructural Details of Fibrils and Facilitates CompleteInhibition of Fibril Growth□S
Received for publication, December 4, 2008, and in revised form, March 13, 2009 Published, JBC Papers in Press, March 19, 2009, DOI 10.1074/jbc.M809151200
Ronald S. Boshuizen‡1, Veronica Schulz‡, Michela Morbin§, Giulia Mazzoleni§, Rob H. Meloen‡¶,and Johannes P. M. Langedijk‡
From ‡Pepscan Therapeutics B.V., Zuidersluisweg 2, 8243 RC AB Lelystad, The Netherlands, §Fondazione Istituto Di Ricovero e Curaa Carattere Scientifico-Instituto Neurologico “Carlo Besta,” via Celoria 11, 20133 Milan, Italy, and ¶Academic Biomedical Centre,University of Utrecht, Yalelaan 1, Utrecht, The Netherlands
Fibrils play an important role in the pathogenesis of amyloi-dosis; however, the underlying mechanisms of the growth proc-ess and the structural details of fibrils are poorly understood.Crucial in the fibril formation of prion proteins is the stacking ofPrP monomers. We previously proposed that the structure ofthe prion protein fibril may be similar as a parallel left-handed�-helix. The �-helix is composed of spiraling rungs of parallel�-strands, and in the PrP model residues 105–143 of each PrPmonomer can contribute two �-helical rungs to the growingfibril. Here we report data to support this model. We show thattwo cyclized human PrP peptides corresponding to residues105–124 and 125–143, based on two single rungs of the left-handed �-helical core of the human PrPSc fibril, show sponta-neous cooperative fibril growth in vitro by heterologous stack-ing. Because the structural model must have predictive value,peptides were designed based on the structure rules of the left-handed �-helical fold that could stack with prion protein pep-tides to stimulate or to block fibril growth. The stimulator pep-tide was designed as an optimal left-handed �-helical fold thatcan serve as a template for fibril growth initiation. The inhibit-ing peptide was designed to bind to the exposed rung but frus-trate the propagation of the fibril growth. The single inhibitorypeptide hardly shows inhibition, but the combination of theinhibitory with the stimulatory peptide showed complete inhi-bition of the fibril growth of peptide huPrP-(106–126). More-over, the unique strategy based on stimulatory and inhibitorypeptides seems a powerful new approach to study amyloido-genic fibril structures in general and could prove useful for thedevelopment of therapeutics.
Transmissible spongiform encephalopathies are neurode-generative disorders in a wide range of mammalian species,including Creutzfeldt-Jacob disease in man, scrapie in sheep,and bovine spongiform encephalopathy in cattle. The deposi-tion of aggregated prion protein fibrils on and in neurons isregarded to be the source of these neurodegenerative diseasesand is frequently associated with occurrence of Congo red pos-itivity (1–3). The fibrils are formed by the conformational
change of the prion protein (PrPc)2 into the scrapie form(PrPSc). Themisfolded conformer of the prion protein (PrPSc) isconsidered as the causative agent in these diseases according tothe protein-only hypothesis (4). Studies have shown the toxicityof fibrils of the full-length recombinant mammalian prion pro-tein as well as soluble �-rich oligomers to cultured cells andprimary neurons (5).It is still unknown how much of the whole PrPSc molecule is
involved in the fibril growth. It is shown that the N-terminalpart of PrP, specifically residues 112–141, can go through con-formational changes involving �-strand formation, which sub-sequently triggers fibril growth (6–8), and solid state NMRstudies showed that residues 112–141 are part of the highlyordered core of huPrP-(23–144) (9). It was previously shownthat peptides based on the 89–143 region of the human PrPprotein can form fibrils rich in �-sheet structure which are bio-logically active in transgenic mice (10). Within this region it isthe huPrP-(106–126) peptide that is the smallest known regionof PrP that forms fibrils that are toxic and resemble the physi-ological properties of PrPSc (11–16). The formation of PrPSc isconsidered to be a two-step event; first, there is the bindingbetween PrPc and PrPSc and subsequently the conformationalconversion from PrPc into PrPSc occurs. Mutation studies in aprion-infected neuroblastoma cell line showed that in mousePrP the regions 101–110 and 136–158 are crucial for the bind-ing and conversion events, respectively (17). Because preven-tion of fibril growth is the prime therapeutic target, detailedstructural knowledge of the fibril is essential for understandingthemechanism of fibril growth. However, structural analysis ofamyloid fibrils is hampered by insolubility, isomorphism, andaggregation. X-ray diffraction of several amyloid fibrils revealeda so-called cross-� diffraction pattern which indicates that thefibrils contain �-strands perpendicular to the fibril axis andhydrogen bonds in parallel (18, 19). Thus, for fibril growth the�-strands have to stack on top of each other. Several structureshave been suggested to explain the structure of the stacked�-strands; e.g. a parallel in register organization of stacked �hairpins (24) or the comparable dry steric zipper structure (25).
□S The on-line version of this article (available at http://www.jbc.org) containsa supplemental figure.
1 To whom correspondence should be addressed. Tel.: 31-320-225331; Fax:31-320-225301; E-mail: [email protected].
2 The abbreviations used are: PrPc, cellular isoform of PrP; PrPSc, scrapie iso-form of PrP; huPrP, human prion protein; Fmoc, fluorenylmethoxycar-bonyl; LpxA, UDP-N-acetylglucosamine acyltransferase; AU, arbitrary units;HPLC, high performance liquid chromatography; MS, mass spectroscopy;PBS, phosphate-buffered saline.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 284, NO. 19, pp. 12809 –12820, May 8, 2009© 2009 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
MAY 8, 2009 • VOLUME 284 • NUMBER 19 JOURNAL OF BIOLOGICAL CHEMISTRY 12809
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from
by guest on February 9, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from

Previously, we and other groups suggested that the �-sheetstructures in the PrPSc fibril may be similar to the topologicallymost simple class of �-sheets; that is, the parallel left-handed�-helix (Fig. 1A) (6, 20, 21). The left-handed�helix is formedbytriangular progressive coils (rungs) of 18–20 residues. Eachrung is formed by three hexapeptide motifs, which results in anapproximate 3-fold symmetry. Backbone-backbone hydrogenbonding and stacking of the side chains in adjacent rungs con-tribute to the folding of�-helical rungs.We suggested that eachPrPSc monomer contributes two left-handed �-helical rungs tothe fibril, comprising residues 105–124 and 125–143 (Fig. 1A).This two-rung structural model was recently confirmed foramyloid fibrils of the HET-s prion by NMR analysis (22). Incontrast to fibrils which are composed of homologous stacks ofidentical peptides, e.g. the A� peptide (23), the PrPSc fibril ismore complex because it is composed of heterologous stacks ofat least two peptides. For homologous stacking of two identicalpeptides, the complementarity issue is relatively simple becausethe identical side chains are in register (e.g. Ile-Ile, Val-Valstacking, and Asn ladders). However, in the case of heterolo-gous stacking, the side chains of the additional heterologouspeptide needs to be complementary with the other peptide toallow fibril growth.To investigate whether the suggested rungs 105–123 and
125–143 from human PrP could be complementary (20), westudied the homologous stacking and the heterologous stacking
of linear and cyclized prion protein peptides comprising thehuPrP-(105–143) region (KTNMKHMAGAAAAGAVVG-GLGGYMLGSAMSRPIIHFGS). Qualitative and semiquantita-tive analysis were done by electron microscopy and Congo redstaining. The quantification of the fibril formationwas assessedby thioflavin S staining, inwhich the addition of polyanions (e.g.heparin) enhance the�-sheet formation of peptides comprisingthe 82–143 region of PrP and improve the reproducibility of thefibril growth (24). This study provides first evidence of heterol-ogous stacking by two isolated putative �-strand layers (orrungs) of the human prion protein with fibril formation as aresult. The left-handed �-helix structure provided insight forthe “stack-and-stop” approach. With this approach a mix of astimulatory peptide and an inhibitory peptide could completelyblock fibril formation. The stimulatory peptide was based onthe 125–143 region that was optimized to serve as a foldingtemplate for the consecutive stacking of the 106–126 peptide.This cooperative fibril growth was completely inhibited by theinhibitory peptide based on peptides 106–126 with strategicD-amino acid and/or proline substitutions. The findings in thisstudy support models in which the sequential strands in a fibrilmust somehow spiral up- or downward along the fibril axis, e.g.like the hypothetical left-handed �-helical structure of PrPScfibrils (20). Furthermore, it allows the development of welldefined small protein modules which can be used for structurestudies of the 82–143 domain of PrPSc and the development oftherapeutics.
EXPERIMENTAL PROCEDURES
Peptides—Peptides were synthesized by Fmoc chemistry andpurified by reversed phase HPLC. For the peptide-peptideinteraction studies some peptides were N-terminal-biotiny-lated by Fmoc chemistry via an aminohexanoic acid spacer. Thepeptides were analyzed for identity by electron spray desorp-tion mass spectrometry according to previously describedmethods (25, 26). Peptides were cyclized by oxidation of the C-andN-terminal cysteines. For the disulfide oxidation 0.1mg/mlpeptide was incubated overnight at 4 °C in an ammonium car-bonate solution (0.1%). The oxidation reaction was monitoredby HPLC/MS analysis. When the cyclization was complete theproduct was purified by reversed phase HPLC and analyzed byHPLC/MS.All cyclized peptides yielded a purity of�95%basedon UV detection at 215 nm. After completion of the cyclizationreaction, trifluoroacetic acid (10% in H2O) was added until thepH was �4. The peptides were lyophilized 3 times from aceto-nitrile (50% (v/v) in water) and stored at �20 °C in 1.4-mlpolypropylene tubes (Micronics, Lelystad, the Netherlands) indry aliquots corresponding with amounts that make up a pep-tide concentration of 200 �M when dissolved in a 120 �l vol-ume. All cooperativity experiments were performed in 50 mMphosphate buffer, pH 5.0, and 10 mM borate buffer, pH 8.5. Ingeneral, similar results were found for experiments performedin pH 5.0, but cooperativity effects were more pronounced inthe pH 8.5 buffer.Detection of Fibril Formation by Thioflavin S Staining and
Cooperativity Calculations—The peptides were dissolved justbefore the start of each experiment at a concentration of 200�Min phosphate buffer (50 mM, pH 5.0) or borate buffer (10 mM,
FIGURE 1. A, theoretical model of the fibrillogenic core of PrPSc. In the PrPSc
model based on the left-handed �-helix structure, each PrPSc monomer con-tributes two stacked rungs to the fibril (different color for each monomer).The protofibril is formed by consecutive stacking of the two windings. Thestack of two rungs provides enough elevation to accommodate the remain-ing part (residues � 146 –253) of the PrPSc molecule (20). B, the left-handed�-helix structure of LpxA-based on x-ray crystallography. In the left-handed�-helix structure of LpxA (PDB code 1LXA) rungs 6 and 7 are indicated (red)that were used for the heterologous stacking studies. Linear and cyclizedpeptides based on rung 6 and rung 7 were modified to satisfy the ideal left-handed �-helix motif (see “LpxA Peptides” under “Results”) and tested fortheir intrinsic and cooperative fibrillogenicity. C, left-handed �-helical rungbased on rung 6 of LpxA. The rung is formed by three hexapeptide motifs,which results in an approximate 3-fold symmetry. A left-handed �-helicalrung can be cyclized by a disulfide bridge after the introduction of a cysteineat position 2 of the first hexapeptide and position 1 of the fourth hexapeptide(according to the numbering used for the hexapeptide repeats in the left-handed �-helix).
Heterologous Stacking by Prion Protein Peptides
12810 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 19 • MAY 8, 2009
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from

pH 8.5) and kept on ice. For the cooperativity studies equal vol-umes of the peptide solutions were mixed. Subsequently the pep-tide solutions were incubated for 1 h at 37 °C. For thioflavin Sstaining, 50-�l aliquots were transferred into microtiter wells(FluoroNunc LumiNunc polystyrene plates, NUNC, Roskilde,Denmark) in which 5 �l of a thioflavin S solution (1.1 mg/ml inH2O; Sigma) was already present, and the microtiter plate wasagitated briefly. Plates were kept covered with microtiter cover-tape (Costar plate sealers, Corning Inc., Corning,NY) tominimizeevaporation. After 90 min, the fluorescence signal was measuredwith a Wallac Victor 1420 Multilabel counter (PerkinElmer LifeSciences) using the following instrumental conditions: 0.1-smeas-urement time, normal emission aperture, excitation and emissionwavelengths 450 and 535 nm, respectively, lamp energy 10,000(arbitrary units). Before the cooperativity calculation, the back-ground signals of the corresponding buffers were subtracted fromthe peptide samples. Background valueswere determined by thio-flavin S staining of the buffers. The cooperativity factor was calcu-latedby the following formula:measuredcooperativity/theoreticalcooperativity.Measured cooperativity is the fluorescence signal ofthemixture (peptide 1� peptide 2), and the theoretical cooperat-ivity is the average fluorescence signal of the individual peptides:(peptide 1�peptide 2)/2. Cooperativity is considered to be signif-icant if the measured cooperativity was significantly higher thanthe theoretical cooperativity and the fluorescence signal of thepeptides individually.HPLC Analyses of Dimerization Studies—Equal molar
amounts of peptide huPrP-(125–143)-E4C and huPrP-(105–126)-RxC or huPrP-(106–126)-RxC (C � Cys, E � Glu, R �Arg, x � 2, 3, or 4) were mixed and incubated for 1, 6, and 24 hat 37 °C in borate buffer, pH 8.5. Subsequently samples forHPLC/MS analysis were taken, and the dimerization reactionwas stopped by the addition of 3 �l of trifluoroacetic acid (10%v/v in water) to a 50-�l sample. The formation of dimers wasstudied by HPLC analysis (Alliance, Waters Corp., Milford,MA) with a C18 reversed phase column and detected simulta-neously with a photodiode array detector and mass spectrometrydetector; eluent was H2O � 0.05% trifluoroacetic acid and aceto-nitrile � 0.05% trifluoroacetic acid with a gradient of 2%/min5–65% acetonitrile. All reagents were at least HPLC grade.Electron Microscopy and Congo Red Staining—Peptide PrP-
(107–123), PrP-(127–142) and cyclized peptide rung 6 andrung 7 alone or in equimolar mixture were dissolved in boratebuffer (200 �M final concentration). Peptide suspensions wereincubated at 37 °C for 1, 24, and 48 h. At each time point, sam-ple aliquots were analyzed by light and electron microscopy.For light microscopy, 10 �l of suspension were air-dried onpoly-L-lysine-coated slides (Bio-Optica, Milan, Italy), stainedwith Congo red, and viewed under polarized light (NikonEclipse E-800, Japan). For ultrastructural examination, 5 �l ofsuspension were applied to Formvar-carbon 200-mesh nickelgrids for 6 min, negatively stained with uranyl acetate, andobserved with an electron microscope (EM109 Zeiss, Oberko-ken, Germany) operated at 80 kV at a standard magnification(�30,000), calibrated with an appropriate grid. The sampleswere evaluated for the presence and relative amount of oli-gomers, amorphous aggregates, filamentous structures, andfibrils by two independent operators.
Peptide-Peptide Interactions—Peptide-peptide interactionswere measured using pepscan analysis (27). Biotinylated pep-tideswere incubatedwith peptideswhichwere synthesized on asolid phase polypropylene support (minicard). The credit card-sized minicard contains 455 wells in which peptide arrays ofoverlapping linear and cyclized 18-mer and linear 22-mer pep-tides based on the PrP 97–150 region were synthesized as pre-viously described (28). Before use, the wells with the solid phasepeptides were washed with PBS-Tween (7.5 mM phosphate,0,14MNaCl, 0,5%Tween 80, pH7.4)) for 30min.Next, thewellswere blocked with 5% bovine serum albumin, 5% horse serum,1%Tween 80 in PBS for 1 h at 25 °C to reduce aspecific binding.Subsequently, the wells were washed 3 times for 5 min with 1%Tween 80 in PBS. The biotinylated PrP peptide 106–126 wasdissolved in water, heated for 20 min at 90 °C, and immediatelykept on ice to minimize aggregation before the incubation withthe solid phase peptide arrays. After cooling of the biotinylated106–126 peptide and shortly before use, a bovine serum albu-min solution (10% bovine serum albumin, 1%Tween 80 in PBS)was addeduntil a final concentrationof 1%bovine serumalbumin.The solid phase peptide arrays on the wells were incubated withthe biotinylated peptide 106–126 solution (3�l perwell) for 1 h at25 °C. After the wells were washed they were incubated for 1 h at25 °Cwith streptavidin-horseradishperoxidase (1/1000)diluted in5% bovine serum albumin plus horse serum for the detection ofboundbiotinylated106–126peptide.Next, thewellswerewashed,and the background signal of the wells wasmeasured with a char-ge-coupled device (CCD) camera. Subsequently the wells werewashedonce (0.1%Tween80 inPBS) and then incubated for 1h atroom temperature with substrate (0.5 g/liter 2,2�-azino-di[3-ethyl-benzthiazolinesulfonate(6)]diammonium salt (ABTS),0.006%H2O2 in 0.18 MNa2HPO4, 0.22 M citric acid, added untilpH 4). The color development was detected at 405 nmwith thecharge-coupled device camera.Inhibition Studies—The inhibition studies were performed
as previously described (24). In short, fibril formation of thelinear peptide huPrP-(106–126)wasmeasured individually andin combinationwith a stimulating or inhibiting peptide or amixof the stimulating and inhibiting peptides. The fibril formationof the stimulator and inhibiting peptides were also measuredindividually. Linear human prion protein peptides were dis-solved in 30 �l of phosphate buffer (50 mM, pH 5) at a concen-tration of 200 �M. Also, the stimulator peptide and each in-hibiting peptide were dissolved in phosphate buffer at aconcentration of 200 or 400 �M. For the combination of thehuPrP-(106–126) peptide with one stimulator or inhibitingpeptide, the linear huPrP-(106–126) peptide was dissolved in30 �l of 200 �M stimulator or inhibiting peptide solution,resulting in a final concentration of 200 �M concentration ofeach peptide. For the combination of huPrP-(106–126) peptidewith a mix of the stimulator peptide and an inhibiting peptide,peptide huPrP-(106–126) was dissolved in 17.5 �l (400 �M) ofstimulator peptide solution plus 17.5 �l (400 �M) inhibitingpeptide solution. This resulted in a final concentration of 200�M concentration of each peptide. Subsequently, the peptideswith or without inhibiting peptide were incubated for 1 h at37 °C.Next, 30�l of heparinwas added to the huPrP-(106–126)peptide until a final concentration of 50 �g/ml to optimize the
Heterologous Stacking by Prion Protein Peptides
MAY 8, 2009 • VOLUME 284 • NUMBER 19 JOURNAL OF BIOLOGICAL CHEMISTRY 12811
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from

fibril growth. This resulted in a final concentration of 100 �Mfor each individual peptide. Peptides were put in the incubatorat 37 °C for another hour. Next, a thioflavin S staining was per-formed. All measurements were performed in triplicate andcorrected for background. Negative values after backgroundcorrection were set at zero, as fluorescencemeasurements can-not be below background. The effect of the fibrillogenic peptideon the fibril growth was expressed as inhibition percentagesand was calculated as follows; for fibrillogenic peptide withinhibiting peptide, inhibition (%) � (p � i � c)/(p � i)100, inwhich the fluorescence values (AU) are given for fibrillogenicpeptide (p), inhibiting peptide (i), and the combination of fibril-logenic peptide with the inhibiting peptide (c); for the fibrillo-genic peptide with stimulator peptide and inhibiting peptide,inhibition (%) � (p � s � i � c)/(p � s � i)100, in which thefluorescence values (AU) are given for fibrillogenic peptide (p),stimulator peptide (s), inhibiting peptide (i), and the combina-tion of fibrillogenic peptide with the stimulator and inhibitingpeptide (c).
RESULTS
To determine how much of the 82–143 region of the humanprion protein is involved in fibril growth, peptides comprising res-idues106–126 (21-mer), 106–143 (38-mer), and82–143 (62-mer)were compared for the amount of fibril growth at equal molarconcentrations. Compared with the 21-mer peptide, the fibrilgrowth of the 62-mer and the 38-mer were �2.4- and 1.5-foldhigher, respectively (Fig. 2). The higher signalwith the longer pep-tides suggests that residues adjacent to the 106–126 region alsoparticipate in the fibril growth and contribute at least one extrarung to the growing fibril. In our proposed left-handed �-helixmodel (20) this would suggest that the minimal fibrillogenic coreof the PrPSc molecule is formed by two stacked rungs (Fig. 1A).N-Acetylglucoseamine-1-phosphate Uridyltransferase (LpxA)
Peptides—Because it was not known whether cooperativestacking of isolated peptides from a �-helical structure was fea-
sible, we used a known natural left-handed �-helix structure of theenzyme LpxA as a model system.Based on regular rungs of LpxA (Fig.1B) (29), we optimized two peptides(rung 6 and rung 7) to increase theprobability of heterologous stack-ing. Because the rungs are taken outof the structural context and to sat-isfy the ideal left-handed �-helixmotif,mutationswere introduced inrung 6. The Met on position 6 wasreplaced by a Gly, being the mostprevalent residue at that position inideal left-handed�-helices (30). TheAsn at position 3 of rung 6 forms ahydrogen bond with a residue fromrung 5, which is not a part of thestructure anymore; therefore, it wasreplaced by Thr an ideal alternativeat this position. The Cys at position15 of rung 6 was replaced by Thr,
and in rung 7 the Cys at position 3 was replaced by Ala. In rung7 the Leu at position 11 was replaced by Ile to improve thestacking with rung 6. The Asp at position 18 of rung 7 wasreplaced by an Ala to avoid disturbance with the polyionicextensions. For further reference in this paper the two peptidesthat are used for the cooperativity studies are coded rung 6 andrung 7. It was shown that peptides rung 6 and rung 7 can indeedbind thioflavin S and form fibrils (Fig. 3).The homologous stacking of rung 7 (intrinsic fluorescence)
was �3.8 times higher than the intrinsic fluorescence of rung 6(Fig. 3A). To enhance the probability of stacking, polyanionic(five glutamic acid residues) and polycationic (five arginine res-idues) extensions were added at the C terminus of the peptides;for further reference the polyionic extensions are denoted asneg and pos, respectively. Theoretically the probability of apotential stack of two peptides will be increased by the attrac-tion of the opposite charged extensions. Indeed the addition ofthe polyionic extensions increased the homologous stacking ofrung 6 by a factor of 2.5 (rung 6-pos� rung 6-neg) (Fig. 3A) andof rung 7 by a factor of 5 (rung 7-pos � rung 7-neg) (Fig. 3B).To facilitate the folding of rungs 6 and 7, cyclized versions of
the peptides were synthesized (Fig. 1C). The cyclizations limitthe flexibility of the peptides and allow the type of stackingsimilar to the typical rungs of a left-handed �-helix. Peptidec-rung 6 showed no fluorescence at all (Fig. 3A), but the addi-tion of polyionic extensions did increase the homologous stack-ing of c-rung 6 spectacularly by a factor of 12 comparedwith thelinear rung 6without polyionic extensions (Fig. 3A). In contrastto c-rung 6, the cyclized version of rung 7 (without polyionicextensions) was found to be much more fibrillogenic than itslinear form aswas shown by the 4-fold higher fluorescence level(Fig. 3B). In contrast to rung 6, the addition of polyionic exten-sions reduced the fluorescence level of the individual c-rung 7,but when the positive and negative charged c-rung 7 peptidesweremixed, the homologous stacking was a factor of 3.5 higherthan expected (Fig. 3B).
FIGURE 2. Fibril formation of three linear prion protein peptides at equimolar concentrations. Levels of thefibril growth of 12.5, 25, and 50 �M concentration of linear peptides huPrP-(106–126) (white bars), huPrP-(106–143)(light gray bars), huPrP-(82–143) (dark gray bars) and 25 �M negative control peptide huPrP-(106–126)-scrambled(NGAKALMGGHGATKVMVGAAA) (black bar) were determined by thioflavin S staining and subsequent fluorescencemeasurements. Data points are average values S.D. of triplicate measurements.
Heterologous Stacking by Prion Protein Peptides
12812 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 19 • MAY 8, 2009
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from
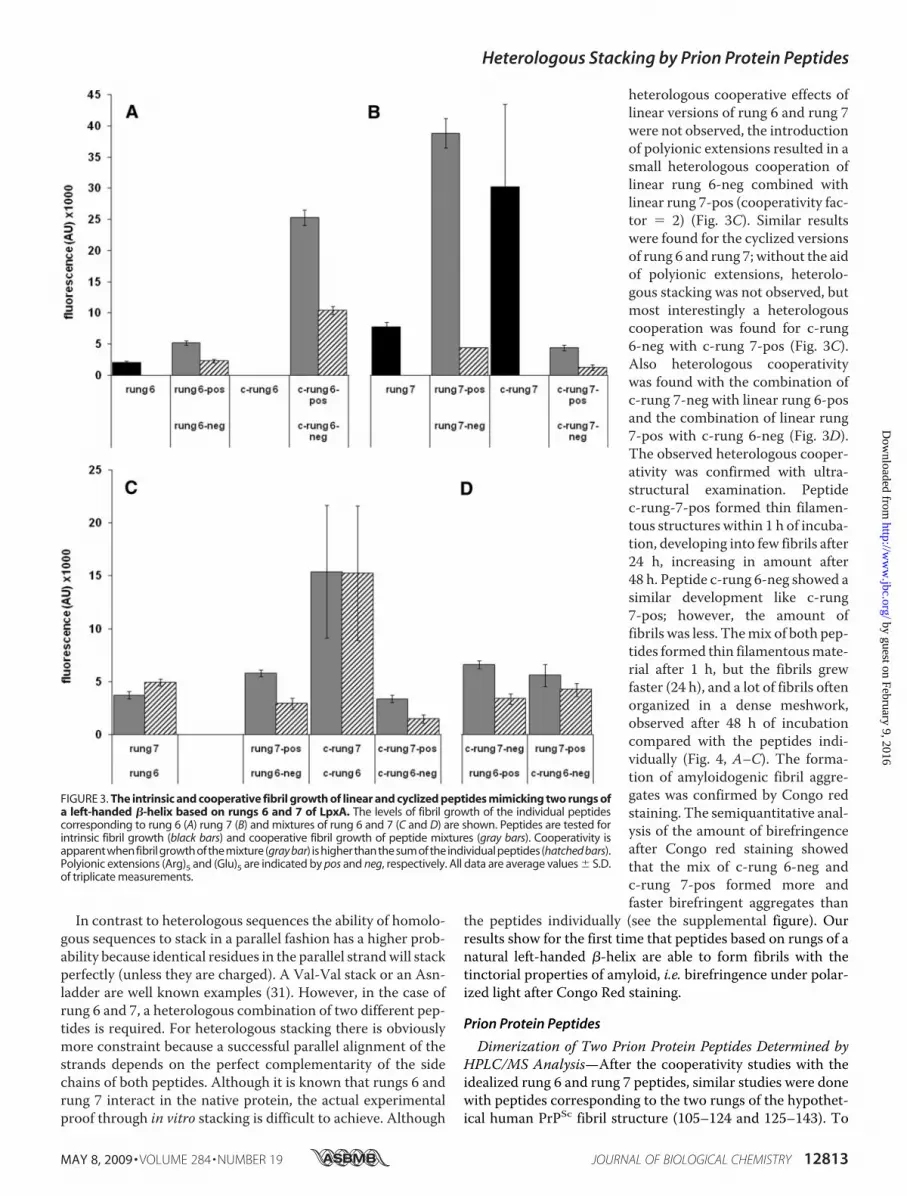
In contrast to heterologous sequences the ability of homolo-gous sequences to stack in a parallel fashion has a higher prob-ability because identical residues in the parallel strandwill stackperfectly (unless they are charged). A Val-Val stack or an Asn-ladder are well known examples (31). However, in the case ofrung 6 and 7, a heterologous combination of two different pep-tides is required. For heterologous stacking there is obviouslymore constraint because a successful parallel alignment of thestrands depends on the perfect complementarity of the sidechains of both peptides. Although it is known that rungs 6 andrung 7 interact in the native protein, the actual experimentalproof through in vitro stacking is difficult to achieve. Although
heterologous cooperative effects oflinear versions of rung 6 and rung 7were not observed, the introductionof polyionic extensions resulted in asmall heterologous cooperation oflinear rung 6-neg combined withlinear rung 7-pos (cooperativity fac-tor � 2) (Fig. 3C). Similar resultswere found for the cyclized versionsof rung 6 and rung 7; without the aidof polyionic extensions, heterolo-gous stacking was not observed, butmost interestingly a heterologouscooperation was found for c-rung6-neg with c-rung 7-pos (Fig. 3C).Also heterologous cooperativitywas found with the combination ofc-rung 7-neg with linear rung 6-posand the combination of linear rung7-pos with c-rung 6-neg (Fig. 3D).The observed heterologous cooper-ativity was confirmed with ultra-structural examination. Peptidec-rung-7-pos formed thin filamen-tous structures within 1 h of incuba-tion, developing into few fibrils after24 h, increasing in amount after48 h. Peptide c-rung 6-neg showed asimilar development like c-rung7-pos; however, the amount offibrils was less. Themix of both pep-tides formed thin filamentousmate-rial after 1 h, but the fibrils grewfaster (24 h), and a lot of fibrils oftenorganized in a dense meshwork,observed after 48 h of incubationcompared with the peptides indi-vidually (Fig. 4, A–C). The forma-tion of amyloidogenic fibril aggre-gates was confirmed by Congo redstaining. The semiquantitative anal-ysis of the amount of birefringenceafter Congo red staining showedthat the mix of c-rung 6-neg andc-rung 7-pos formed more andfaster birefringent aggregates than
the peptides individually (see the supplemental figure). Ourresults show for the first time that peptides based on rungs of anatural left-handed �-helix are able to form fibrils with thetinctorial properties of amyloid, i.e. birefringence under polar-ized light after Congo Red staining.
Prion Protein Peptides
Dimerization of Two Prion Protein Peptides Determined byHPLC/MS Analysis—After the cooperativity studies with theidealized rung 6 and rung 7 peptides, similar studies were donewith peptides corresponding to the two rungs of the hypothet-ical human PrPSc fibril structure (105–124 and 125–143). To
FIGURE 3. The intrinsic and cooperative fibril growth of linear and cyclized peptides mimicking two rungs ofa left-handed �-helix based on rungs 6 and 7 of LpxA. The levels of fibril growth of the individual peptidescorresponding to rung 6 (A) rung 7 (B) and mixtures of rung 6 and 7 (C and D) are shown. Peptides are tested forintrinsic fibril growth (black bars) and cooperative fibril growth of peptide mixtures (gray bars). Cooperativity isapparentwhenfibrilgrowthofthemixture(gray bar) ishigherthanthesumoftheindividualpeptides(hatched bars).Polyionic extensions (Arg)5 and (Glu)5 are indicated by pos and neg, respectively. All data are average values S.D.of triplicate measurements.
Heterologous Stacking by Prion Protein Peptides
MAY 8, 2009 • VOLUME 284 • NUMBER 19 JOURNAL OF BIOLOGICAL CHEMISTRY 12813
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from

investigate the interpeptide disulfide bridge formation in het-erodimers of stacked PrP peptides, we analyzed the dimeriza-tion reaction usingHPLC/MS. The different peptideswere syn-thesized with a polyionic extension and a free cysteine that
would allow interpeptide disulfideformation when the peptides wouldform a correct heterologous parallelalignment. The tendency to het-erodimerize was studied with linearpeptides (125–143)-E4C, (105–126)-RxC, and (106–126)-RxC (C � Cys,E � Glu, R � Arg, x � 2, 3, or 4).HPLC/MS analysis showed het-erodimerization of peptide (125–143)-E4C and (105–126)-R4C with-out the formation of homodimers(Fig. 5C). Peptide (105–126)-R4 canslowly homodimerize with itself (Fig.5B), but the heterodimerization with(125–143)-E4 was preferred (Fig. 5C).The rate of heterodimerization of(125–143)-E4C with (106–126)-RxCand (105–126)-RxC (x � 2–4)depended on the number of arginineresidues in the C-terminal extension(data not shown).Theheterodimerizationwastime-
dependent, ranging from 3 to 14%after 1 h and increased after 6 h ofincubation (21–73%) and 24 h (61–88%) (data not shown). The dimer-ization experiments indicated that,with the appropriate C-terminalextensions, linear prion proteinpeptides comprising regions 105–126 and 125–143 are able to het-erodimerize within 1 h without theformation of homodimers.Cooperativity of Two Prion Pro-
tein Peptides—To investigate thepossibility of the heterologousstacking by prion protein peptidesfrom the regions 105–123 and 124–143, based on the two rung model(20), these peptides were tested forthioflavin S staining, which is ameasure for the amount of fibrilgrowth. Peptides based on the 106–124 and 125–143 regions werecyclized by a disulfide bond tomimic the typical rungs of a left-handed �-helix (Fig. 1C) but alsoother models in which some kind ofspiraling of the parallel �-strand isessential. Interestingly, the fluores-cence level of the c-106–123 wasmuch higher (3-fold) than its lin-ear version (Fig. 6A). Also a
cyclized version of the 127–143 region showed someincrease in the fluorescence level compared with its linearform (Fig. 6B). Similar to the experiments with rung 6 and 7of LpxA, also the linear and cyclized versions of the PrP
FIGURE 4. Ultrastructural analysis of cyclized LpxA and cyclized prion protein peptides. Electron micro-graphs are shown of protofibrils formed in borate buffer pH 8.5 after 48 h of incubation at 37 °C. The LpxApeptide c-rung 6 neg (A) formed filamentous material, protofibrils, and some short fibrils, c-rung 7-pos (B)formed long unbranched and sometimes paired fibrils mixed with protofibrils, and the combination of both (C)formed a dense meshwork of long unbranched, often paired fibrils. Individual prion protein peptide c-127–142-neg (D) formed long unbranched and sometimes twisted fibrils, c-107–123-pos (E) formed very short andsometimes paired fibrils, and the combination of both (F) formed a meshwork of long, unbranched and some-times paired fibrils. The bar in C is applicable to all figures.
FIGURE 5. Heterodimerization of peptides huPrP-(125–143)-E4C and huPrP-(105–126)-R4C. Shown isHPLC analysis at 215 nm of the individual linear peptides huPrP-(125–143)E4C (top panel), huPrP-(105–126)R4C(middle panel), and the mix of both peptides (bottom panel) after six h incubation in borate buffer pH 8.5 at37 °C. The peaks were identified by mass spectrometry.
Heterologous Stacking by Prion Protein Peptides
12814 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 19 • MAY 8, 2009
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from

peptides did not show cooperativity without the help of poly-ionic extensions.The addition of polyionic extensions to peptides comprising
the PrP regions 105–123 and 124–143 reduced the fluores-cence levels 2–6-fold compared with the peptides without anextension (data not shown). However, the addition of polyionicextensions resulted in a significant heterologous cooperativityas was observed between linear peptides 105–124-pos and124–143-neg (cooperativity factor � 1.6) (Fig. 6C). The heter-ologous stacking improved spectacularly (cooperativity fac-tor � 6) when a combination of polyionic extensions andcyclizationwas used (Fig. 6D). A significant heterologous coop-erativity (cooperativity factor � 2) was also found between lin-ear 105–123-pos and c-127–142-neg, whereas c-107–123-poscombinedwith linear 124–142-neg showed some cooperativity(cooperativity factor 1.5) (Fig. 6E). Control experiments withpeptides unrelated to prion protein sequences (acetyl-CDGAVQPDGGQPAVRNER-amide and fluorescein iso-thiocyanate-aminohexanoic acid-LEDKIEELLSKIYHLENE-IARLAAAIRRR-amide) showed no significant cooperative
fluorescence levels with any of the linear or cyclized versions ofprion protein peptides with polyionic extensions based onregions 105–124 and 124–143 (data not shown). The ability toform amyloid fibrils was studied by electron microscopy, andCongo red staining for the peptides that displayed the highestcooperativity (Fig. 6D) were studied. Peptide c-127–142-negformed round-shaped structures (oligomers) and filamentousstructures within 1 h of incubation, after 24 h filamentousmaterial and protofibrils were found, whereas fibril aggre-gates were found after 48 h of incubation. Peptide c-107–123-pos showed dense amorphous structures, whereas after 24 hfilamentous structures were found, and after 48 h of incubationsome fibrils were also evident. Themix of both peptides formedprotofibrils among round-shaped structures (oligomers) andthin filamentous structures after 1 h of incubation. After 24 hthe mix formed few short amyloid fibrils, whereas after 48 haggregates of fibrils (no oligomers) were found in the mix in amuch higher density than the individual peptides (Fig. 4,D–F).Furthermore, the presence of amyloid fibril was confirmed byCongo red staining. The semiquantitative analysis of theamount of birefringence after Congo red staining showed thatthe mix of c-107–123-pos and c-127–142-neg not only formedmore amyloidogenic fibril aggregates than the peptides individ-ually, but also faster (supplemental figure). In contrast to theelectron microscopy and Congo red analysis, the thioflavinstaining is maximal within 2.5 h of incubation (including 1.5 hthioflavin staining). This indicates that probably the number ofthioflavin staining sites remain equal when oligomers aggregateinto protofibrils and mature fibrils (data not shown).Because the left-handed �-helix model predicted a comple-
mentary stack of two peptide rungs, it was investigatedwhetherthe influence of specific mutations in rung 1 (c-107–123-pos)and rung 2 (c-127–142-neg) could change the observed coop-erativity. As a positive control, a variant of rung 1 was synthe-sized in which Ala115 was substituted to valine. A better coop-erative stacking was expected from a valine mutation of Ala115,because the inside of the rung is better filled by a valine residueat this position according to the model. Indeed the heterolo-gous cooperativity of rung 2 with rung 1 in which Ala115 wassubstituted for valine showed an increase in fluorescence (Fig.7). Two variants of rung 2 were synthesized as negative con-trols; one variant comprising a scrambled 127–142 sequence(rung 2 scrambled) and one variant inwhich the size of the rung2 was reduced by six residues (rung 2-small: c-130–139-neg).As expected, the heterologous cooperativity between rung 1and rung 2 dropped from a cooperativity factor 3 to 1.5 whenthe size of rung 2 was reduced by six residues (rung 2-small) orwhen the sequence of rung 2 was scrambled (Fig. 7).Peptide-Peptide Interaction Studies—To investigate which
residues are involved in the heterologous stacking of prionprotein peptides, binding studies were performed with pep-tides comprising the 97–150 region of the human prion pro-tein. Biotinylated peptides huPrP-(106–126) (5 �g/ml) andhuPrP-(106–126)-pos (0.5 �g/ml) were tested for reactivi-ties with all overlapping linear (18- and 22-mer) and cyclized(18-mer) peptides of the PrP central domain (97–150) andas negative control peptides based on the LpxA pro-tein (biotinylated rung 7-pos), peptide KIAALKE-
FIGURE 6. Intrinsic and cooperative fibril growth of linear and cyclizedprion protein peptides comprising residues 106 –123 and 127–142. Thefibril growth of linear and cyclized peptides comprising the 106 –126 (A) and126 –143 regions (B) of PrP and the heterologous cooperation of linear (C),cyclized (D), and linear with cyclized (E) peptides based on PrP regions 106 –126 and 126 –143 are shown. The measured fluorescence levels are indicatedby gray bars, and the theoretical fluorescence levels of the heterologouscooperation are indicated by hatched bars. Polyionic extensions (Arg)5 and(Glu)5 are indicated by pos and neg, respectively. All data are average val-ues S.D. of triplicate measurements.
Heterologous Stacking by Prion Protein Peptides
MAY 8, 2009 • VOLUME 284 • NUMBER 19 JOURNAL OF BIOLOGICAL CHEMISTRY 12815
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from
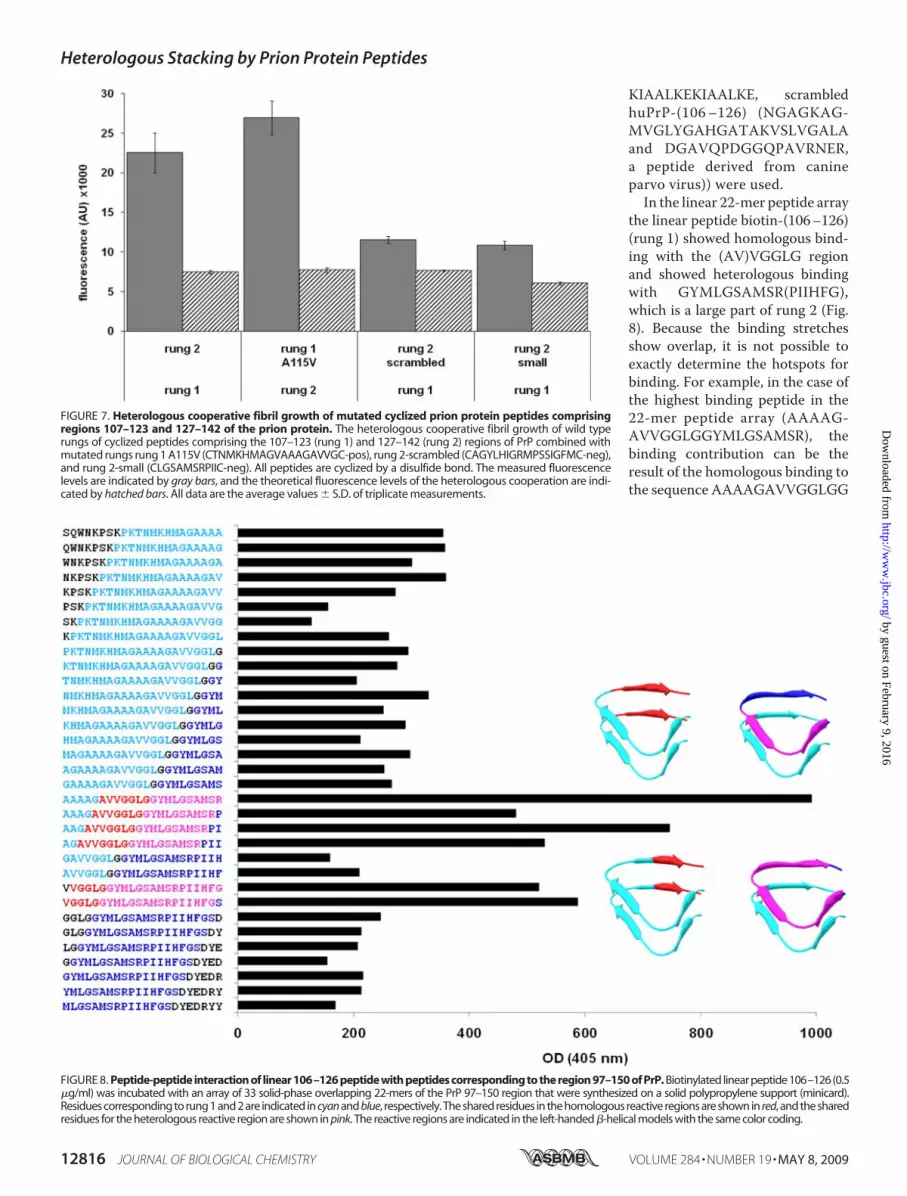
KIAALKEKIAALKE, scrambledhuPrP-(106–126) (NGAGKAG-MVGLYGAHGATAKVSLVGALAand DGAVQPDGGQPAVRNER,a peptide derived from canineparvo virus)) were used.In the linear 22-mer peptide array
the linear peptide biotin-(106–126)(rung 1) showed homologous bind-ing with the (AV)VGGLG regionand showed heterologous bindingwith GYMLGSAMSR(PIIHFG),which is a large part of rung 2 (Fig.8). Because the binding stretchesshow overlap, it is not possible toexactly determine the hotspots forbinding. For example, in the case ofthe highest binding peptide in the22-mer peptide array (AAAAG-AVVGGLGGYMLGSAMSR), thebinding contribution can be theresult of the homologous binding tothe sequence AAAAGAVVGGLGG
FIGURE 7. Heterologous cooperative fibril growth of mutated cyclized prion protein peptides comprisingregions 107–123 and 127–142 of the prion protein. The heterologous cooperative fibril growth of wild typerungs of cyclized peptides comprising the 107–123 (rung 1) and 127–142 (rung 2) regions of PrP combined withmutated rungs rung 1 A115V (CTNMKHMAGVAAAGAVVGC-pos), rung 2-scrambled (CAGYLHIGRMPSSIGFMC-neg),and rung 2-small (CLGSAMSRPIIC-neg). All peptides are cyclized by a disulfide bond. The measured fluorescencelevels are indicated by gray bars, and the theoretical fluorescence levels of the heterologous cooperation are indi-cated by hatched bars. All data are the average values S.D. of triplicate measurements.
FIGURE 8. Peptide-peptide interaction of linear 106–126 peptide with peptides corresponding to the region 97–150 of PrP. Biotinylated linear peptide 106–126 (0.5�g/ml) was incubated with an array of 33 solid-phase overlapping 22-mers of the PrP 97–150 region that were synthesized on a solid polypropylene support (minicard).Residues corresponding to rung 1 and 2 are indicated in cyan and blue, respectively. The shared residues in the homologous reactive regions are shown in red, and the sharedresidues for the heterologous reactive region are shown in pink. The reactive regions are indicated in the left-handed �-helical models with the same color coding.
Heterologous Stacking by Prion Protein Peptides
12816 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 19 • MAY 8, 2009
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from

or of the heterologous binding to YMLGSAMSR and mostlikely both. Similar binding interactions were found withbiotin-(106–126) and with biotin-(106–126)-pos in the linear18-mer and cyclized 18-mer peptide arrays. However, thesedata were even more difficult to interpret because the bindingstretches showed even more overlap (data not shown). No sig-nificant binding was observed with the negative control pep-tides (data not shown).Inhibition of Fibril Growth of Linear Prion Protein Peptide
huPrP-(106–126)—To find additional support for the hypo-thetical left-handed �-helical structure of human PrPSc, pep-tides were designed based on the structure of the left-handed�-helix that could stack with prion protein peptides to stimu-late or to block fibril growth. A stimulator peptide should havean optimal left-handed �-helical fold that can serve as a tem-plate for fibril growth initiation. The inhibitor peptide shouldbe able to bind to the exposed rung but frustrate the propaga-tion of the fibril growth. A proline is hardly ever present in theleft-handed �-helical rung except for position 1 at the corner ofthe triangle where the occurrence is very high. Especially at thetop or bottom rung the occurrence of the proline at position 1 ishigh because it decreases the conformational entropy and facil-itates folding of the �-helical structure (32). Therefore, a �-he-lix stimulator peptide (peptide 1 Fig. 9) was designed with a
proline at the first position of the middle hexapeptide motif ofrung 2 (G131P). Because optimized rung 2 (stimulator) has aproline at both corners of the triangle (positions 131 and 137),this peptide has a higher chance of the triangular arrangement(see Fig. 11B). Indeed, a strong cooperative fibril growth wasobserved (47%) when rung 1 (106–126) was mixed with theoptimized rung 2 (peptide 1 with the G131P mutation; Fig. 9).This is the first example of heterologous cooperative stackingwithout the aid of cyclization or polyionic extensions. Subse-quently, several attempts weremade for the design of inhibitingpeptides based on rung 1 with D-amino acid substitutions atpositions His111, Met112, Ala117, Ala118, and proline substitu-tions of Ala115 and Val122. The proline substitution of the resi-dues at the fifth position of the middle and last hexapeptide ofrung 1 frustrates the ideal fold of a�-helical rung, and the intro-duction of D-amino acids in rung 1 would clash with the sidechains of rung 2, and therefore, a reduced fibril growth cooper-ativity of rung 1 and rung 2was expected. Indeed, the inhibitingpeptides based on rung 1 (peptides 2–4 in Fig. 9) showed no ora low inhibition of fibril growth of rung 1, but when the stimu-lator peptide based on optimized rung 2was combinedwith theinhibiting peptides based on rung 1, a complete inhibition offibril growth was observed (Fig. 9). Furthermore the inhibitionlevel was inhibitor concentration-dependent (Fig. 10). These
FIGURE 9. Stimulation and inhibition of the fibril growth of linear prion protein peptide 106 –126. Peptide huPrP-(106 –126) was incubated with thestimulatory peptide (peptide 1) based on rung 2 of PrP that can serve as a template for fibril growth of huPrP-(106 –126) or inhibitory peptides (peptides 2–5)based on PrP region 106 –126 containing Pro (bold capitals) and/or D-amino acid residue mutations (bold lowercase letters) at strategic positions. Next peptidehuPrP-(106 –126) was incubated with a mix of the stimulatory (peptide 1) and inhibitory (peptides 2–5) peptides. The final concentration of each peptide was100 �M. The fibril growth was determined by thioflavin S staining and the stimulation and inhibition percentages were calculated as described under“Experimental Procedures.” All data are the average values S.D. of triplicate measurements.
Heterologous Stacking by Prion Protein Peptides
MAY 8, 2009 • VOLUME 284 • NUMBER 19 JOURNAL OF BIOLOGICAL CHEMISTRY 12817
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from

results confirm the role of cooperativity of the heterologouschains.
DISCUSSION
PrPc consists predominantly of �-helix structures, whereasthe conversion into PrPSc involves an increase of �-sheet con-tent. This conformational change promotes the fibril forma-tion. Although it is still unknown howmuch of the whole PrPScmolecule is involved in the fibril growth, solid-state NMR stud-ies showed that residues 112–141 are part of the highly orderedcore of huPrP-(23–144) (9). It is well accepted that the regioncomprising residues 89–143 can form fibrils rich in �-struc-tures (12, 14, 15). X-ray diffraction studies of amyloid fibrils ingeneral suggest a parallel �-strand organization (18, 19). Previ-ously, we and others proposed that the PrPSc fibril may fold as aparallel left-handed�-helix (6, 20, 21) and that each PrPScmon-omer contributes two left-handed �-helical rungs to the fibril,comprising residues 105–124 and 125–143 (Fig. 1A). This two-rung structural model was recently confirmed for amyloidfibrils of the HET-s prion by NMR analysis (22).The smallest known region of PrP that forms fibrils is peptide
huPrP-(106–126). These fibrils are toxic and resemble thephysiological properties of PrPSc (11, 16, 26, 33). The left-handed �-helix model predicts, however, that the elevation ofone PrPSc monomer requires at least two �-helical rungs ofheterologous sequences (20). Because in this model the 106–126 region can only make up one rung, the next rung needs tobe formed by residues C-terminal to the first rung. Support forthis was obtained with the thioflavin S staining studies with thelonger peptides that showed that flanking regions are alsoinvolved in the parallel �-sheet structure of the fibrils (Fig. 1).Subsequently we studied whether continuous heterologousstacking of the two regions would result into fibril growth.Therefore, peptides based on the 105–126 and 127–143 regionswere assayed for their individual and heterologous fibrilgrowth. Here we report experimental results supporting themodel by the heterologous stacking in vitro of two peptides
comprising the 105–126 and 127–143 regions of the humanprion protein.Heterologous Stacked Prion Protein Peptides Are Realistic
Mimics of the Theoretical Left-handed�-Helix—Using a similarapproach as for LpxA, we studied the heterologous stacking oflinear and cyclized prion protein peptides comprising the PrPregion 105–143. Because peptides comprising residues 105–124 and 125–143 have been suggested to form a heterologousstack in the left-handed �-helix model of PrP (20), the het-erodimerization of both regions was studied with HPLC/MSanalysis (Fig. 5). As expected, the electrostatic attraction of thecharged polyionic extensions at the C termini favored the het-erodimerization and disulfide bridge formation, favoring a par-allel organization of the peptides that may facilitate parallelstacking. Also, thioflavin S staining showed that the polyionicextensions increased the heterologous cooperative fibrilgrowth of linear peptides comprising regions 105–123 and124–143 (Fig. 6C). Irrespective of the hypothetical nature of theleft-handed �-helical model, the sequential strands in a fibrilmust somehow spiral up- or downward along the fibril axis,reminiscent to e.g. a left-handed�-helix. Therefore, also for PrPthe cyclized peptides are more realistic mimics of the putativespiraling rungs in a fibril than the linear ones. It was found thatc-106–123 was 3.2-fold more fibrillogenic than its linear ver-sion 106–126 (Fig. 6A). Furthermore, a striking 6-fold heterol-ogous cooperativity was observedwith peptides c-107–123-posand c-127–142-neg (Fig. 6D), which is convincingly supportedby the electronmicroscopy analysis (Fig. 4,D–F) andCongo redstaining (see the supplemental figure), both indicating a fasterand more intense aggregation of amyloidogenic fibrils com-pared with the individual peptides. Also, linear peptides canstack better with the cyclized heterologous peptide than withthe linear version (Fig. 6, E and C, respectively), suggesting thata conformationally stabilized peptide increases the chance ofheterologous stacking by serving as a template that can initiatefibril formation. The prion protein peptides showed evenhigher heterologous stacking then the positive control peptides(rung 6 and 7) derived from the known left-handed �-helixLpxA (Figs. 6D and 3D, respectively). A positive control forheterologous stacking of rung 1 and rung 2 was themutation ofAla115 into valine, which is located at the fifth position of thehexapeptide motif and is expected to fill the inside of the rungbetter then alanine and, therefore, would increase the stackingand fibril growth. As predicted, the A115V mutation showedhigh thioflavin S staining with the wild type rung 1 (Fig. 7). Anegative control was the scrambled version of rung 2 includinga proline at the unfavorable fifth position of the hexapeptidemotif. Another negative control was a rung 2 variant, whichwasreduced in size by six residues (c-130–139-neg), by which thecomplementary stacking with rung 1 is hampered. As pre-dicted, compared with the wild type rung 2, both negative con-trols showed less heterologous cooperative fibril growth withrung 1 (Fig. 7). The peptide-peptide interaction studies (Fig. 8)show that peptide 106–126 interacts homologously withsequences comprising the 106–126 region of PrP, which agreeswith the known fibril growth of huPrP-(106–126). Moreimportantly it was shown that the peptide 106–126 also inter-acts heterologously with the 125–143 region of PrP (Fig. 8).
FIGURE 10. The efficiency of fibril growth inhibitor peptide huPrP-(106 –126) with D-amino acid substitutions at positions His111, Met112, andAla116. 100 �M peptide huPrP-(106 –126) was incubated with 100 �M stimu-latory peptide based on rung 2 of PrP (peptide 1) and the inhibitor peptide(peptide 4), which was titrated from 100 to 5 �M. The fibril growth was deter-mined by thioflavin S staining, and the inhibition percentages were calcu-lated as described under “Experimental Procedures.” Subsequently the inhi-bition levels were standardized relative to the maximum inhibition level. Alldata are the average values S.D. of duplicate measurements.
Heterologous Stacking by Prion Protein Peptides
12818 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 19 • MAY 8, 2009
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from
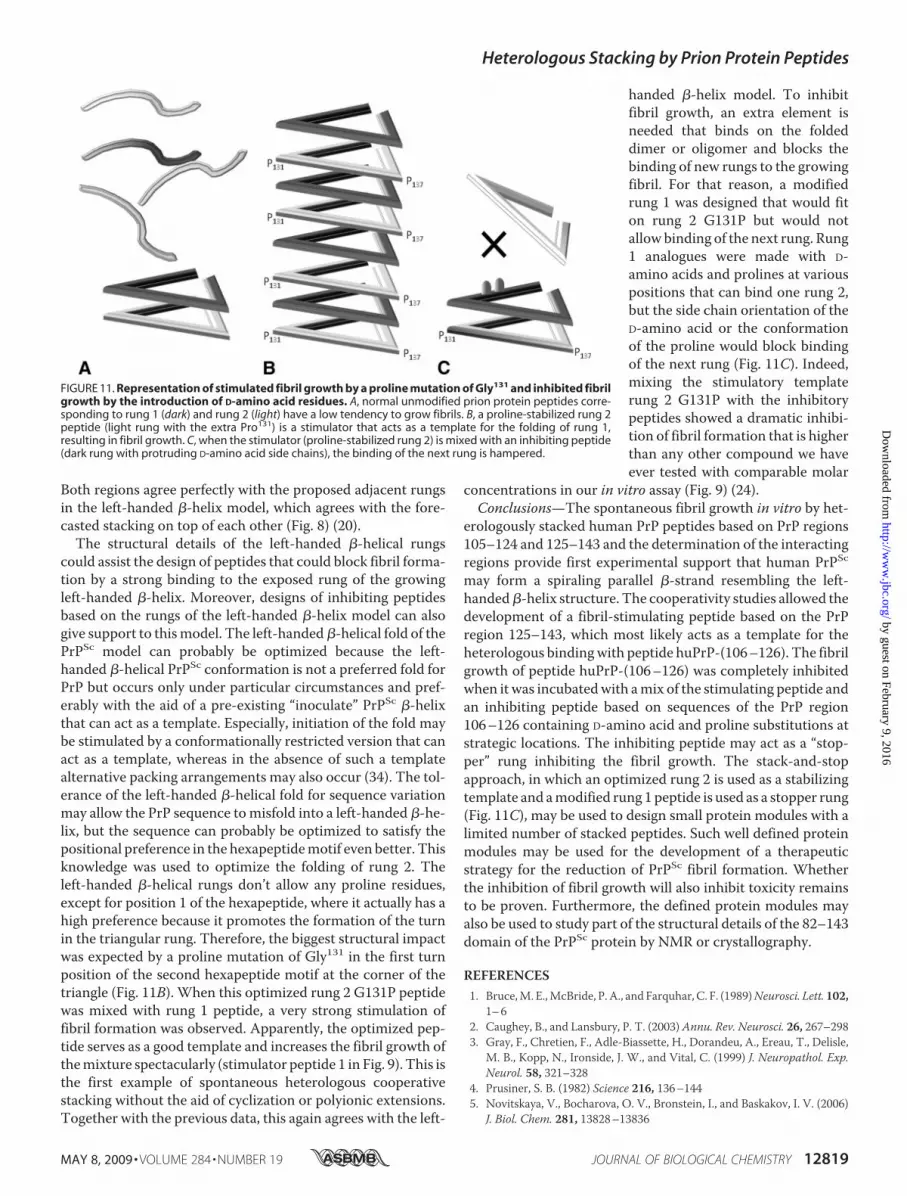
Both regions agree perfectly with the proposed adjacent rungsin the left-handed �-helix model, which agrees with the fore-casted stacking on top of each other (Fig. 8) (20).The structural details of the left-handed �-helical rungs
could assist the design of peptides that could block fibril forma-tion by a strong binding to the exposed rung of the growingleft-handed �-helix. Moreover, designs of inhibiting peptidesbased on the rungs of the left-handed �-helix model can alsogive support to thismodel. The left-handed�-helical fold of thePrPSc model can probably be optimized because the left-handed �-helical PrPSc conformation is not a preferred fold forPrP but occurs only under particular circumstances and pref-erably with the aid of a pre-existing “inoculate” PrPSc �-helixthat can act as a template. Especially, initiation of the fold maybe stimulated by a conformationally restricted version that canact as a template, whereas in the absence of such a templatealternative packing arrangements may also occur (34). The tol-erance of the left-handed �-helical fold for sequence variationmay allow the PrP sequence tomisfold into a left-handed �-he-lix, but the sequence can probably be optimized to satisfy thepositional preference in the hexapeptidemotif even better. Thisknowledge was used to optimize the folding of rung 2. Theleft-handed �-helical rungs don’t allow any proline residues,except for position 1 of the hexapeptide, where it actually has ahigh preference because it promotes the formation of the turnin the triangular rung. Therefore, the biggest structural impactwas expected by a proline mutation of Gly131 in the first turnposition of the second hexapeptide motif at the corner of thetriangle (Fig. 11B). When this optimized rung 2 G131P peptidewas mixed with rung 1 peptide, a very strong stimulation offibril formation was observed. Apparently, the optimized pep-tide serves as a good template and increases the fibril growth ofthemixture spectacularly (stimulator peptide 1 in Fig. 9). This isthe first example of spontaneous heterologous cooperativestacking without the aid of cyclization or polyionic extensions.Together with the previous data, this again agrees with the left-
handed �-helix model. To inhibitfibril growth, an extra element isneeded that binds on the foldeddimer or oligomer and blocks thebinding of new rungs to the growingfibril. For that reason, a modifiedrung 1 was designed that would fiton rung 2 G131P but would notallow binding of the next rung. Rung1 analogues were made with D-amino acids and prolines at variouspositions that can bind one rung 2,but the side chain orientation of theD-amino acid or the conformationof the proline would block bindingof the next rung (Fig. 11C). Indeed,mixing the stimulatory templaterung 2 G131P with the inhibitorypeptides showed a dramatic inhibi-tion of fibril formation that is higherthan any other compound we haveever tested with comparable molar
concentrations in our in vitro assay (Fig. 9) (24).Conclusions—The spontaneous fibril growth in vitro by het-
erologously stacked human PrP peptides based on PrP regions105–124 and 125–143 and the determination of the interactingregions provide first experimental support that human PrPScmay form a spiraling parallel �-strand resembling the left-handed�-helix structure. The cooperativity studies allowed thedevelopment of a fibril-stimulating peptide based on the PrPregion 125–143, which most likely acts as a template for theheterologous bindingwith peptide huPrP-(106–126). The fibrilgrowth of peptide huPrP-(106–126) was completely inhibitedwhen it was incubatedwith amix of the stimulating peptide andan inhibiting peptide based on sequences of the PrP region106–126 containing D-amino acid and proline substitutions atstrategic locations. The inhibiting peptide may act as a “stop-per” rung inhibiting the fibril growth. The stack-and-stopapproach, in which an optimized rung 2 is used as a stabilizingtemplate and amodified rung 1peptide is used as a stopper rung(Fig. 11C), may be used to design small protein modules with alimited number of stacked peptides. Such well defined proteinmodules may be used for the development of a therapeuticstrategy for the reduction of PrPSc fibril formation. Whetherthe inhibition of fibril growth will also inhibit toxicity remainsto be proven. Furthermore, the defined protein modules mayalso be used to study part of the structural details of the 82–143domain of the PrPSc protein by NMR or crystallography.
REFERENCES1. Bruce,M. E.,McBride, P. A., and Farquhar, C. F. (1989)Neurosci. Lett.102,
1–62. Caughey, B., and Lansbury, P. T. (2003) Annu. Rev. Neurosci. 26, 267–2983. Gray, F., Chretien, F., Adle-Biassette, H., Dorandeu, A., Ereau, T., Delisle,
M. B., Kopp, N., Ironside, J. W., and Vital, C. (1999) J. Neuropathol. Exp.Neurol. 58, 321–328
4. Prusiner, S. B. (1982) Science 216, 136–1445. Novitskaya, V., Bocharova, O. V., Bronstein, I., and Baskakov, I. V. (2006)
J. Biol. Chem. 281, 13828–13836
FIGURE 11. Representation of stimulated fibril growth by a proline mutation of Gly131 and inhibited fibrilgrowth by the introduction of D-amino acid residues. A, normal unmodified prion protein peptides corre-sponding to rung 1 (dark) and rung 2 (light) have a low tendency to grow fibrils. B, a proline-stabilized rung 2peptide (light rung with the extra Pro131) is a stimulator that acts as a template for the folding of rung 1,resulting in fibril growth. C, when the stimulator (proline-stabilized rung 2) is mixed with an inhibiting peptide(dark rung with protruding D-amino acid side chains), the binding of the next rung is hampered.
Heterologous Stacking by Prion Protein Peptides
MAY 8, 2009 • VOLUME 284 • NUMBER 19 JOURNAL OF BIOLOGICAL CHEMISTRY 12819
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from

6. Govaerts, C., Wille, H., Prusiner, S. B., and Cohen, F. E. (2004) Proc. Natl.Acad. Sci. U. S. A. 101, 8342–8347
7. Luhrs, T., Zahn, R., and Wuthrich, K. (2006) J. Mol. Biol. 357, 833–8418. Peretz, D., Williamson, R. A., Matsunaga, Y., Serban, H., Pinilla, C., Basti-
das, R. B., Rozenshteyn, R., James, T. L., Houghten, R. A., Cohen, F. E.,Prusiner, S. B., and Burton, D. R. (1997) J. Mol. Biol. 273, 614–622
9. Helmus, J. J., Surewicz, K., Nadaud, P. S., Surewicz, W. K., and Jaroniec,C. P. (2008) Proc. Natl. Acad. Sci. U. S. A. 105, 6284–6289
10. Kaneko, K., Ball, H. L.,Wille, H., Zhang, H., Groth, D., Torchia,M., Trem-blay, P., Safar, J., Prusiner, S. B., DeArmond, S. J., Baldwin, M. A., andCohen, F. E. (2000) J. Mol. Biol. 295, 997–1007
11. Forloni, G., Angeretti, N., Chiesa, R., Monzani, E., Salmona, M., Bugiani,O., and Tagliavini, F. (1993) Nature 362, 543–546
12. Laws, D. D., Bitter, H.M., Liu, K., Ball, H. L., Kaneko, K.,Wille, H., Cohen,F. E., Prusiner, S. B., Pines, A., andWemmer, D. E. (2001) Proc. Natl. Acad.Sci. U. S. A. 98, 11686–11690
13. Lim, K. H., Nguyen, T. N., Damo, S. M., Mazur, T., Ball, H. L., Prusiner,S. B., Pines, A., andWemmer, D. E. (2006) Solid State Nucl. Magn. Reson.29, 183–190
14. Salmona, M., Morbin, M., Massignan, T., Colombo, L., Mazzoleni, G.,Capobianco, R., Diomede, L., Thaler, F., Mollica, L., Musco, G., Kourie,J. J., Bugiani, O., Sharma, D., Inouye, H., Kirschner, D. A., Forloni, G., andTagliavini, F. (2003) J. Biol. Chem. 278, 48146–48153
15. Tagliavini, F., Prelli, F., Porro, M., Rossi, G., Giaccone, G., Farlow, M. R.,Dlouhy, S. R., Ghetti, B., Bugiani, O., and Frangione, B. (1994) Cell 79,695–703
16. Tagliavini, F., Prelli, F., Verga, L., Giaccone, G., Sarma, R., Gorevic, P.,Ghetti, B., Passerini, F., Ghibaudi, E., Forloni, G., et al. (1993) Proc. Natl.Acad. Sci. U. S. A. 90, 9678–9682
17. Abalos, G. C., Cruite, J. T., Bellon, A., Hemmers, S., Akagi, J., Mastrianni,J. A., Williamson, R. A., and Solforosi, L. (2008) J. Biol. Chem. 283,34021–34028
18. Pauling, L., and Corey, R. B. (1951) Proc. Natl. Acad. Sci. U. S. A. 37,729–740
19. Sunde,M., Serpell, L. C., Bartlam,M., Fraser, P. E., Pepys,M. B., and Blake,C. C. (1997) J. Mol. Biol. 273, 729–739
20. Langedijk, J. P., Fuentes, G., Boshuizen, R., andBonvin, A.M. (2006) J.Mol.Biol. 360, 907–920
21. Stork, M., Giese, A., Kretzschmar, H. A., and Tavan, P. (2005) Biophys. J.88, 2442–2451
22. Wasmer, C., Lange, A., Van Melckebeke, H., Siemer, A. B., Riek, R., andMeier, B. H. (2008) Science 319, 1523–1526
23. Luhrs, T., Ritter, C., Adrian,M., Riek-Loher, D., Bohrmann, B., Dobeli, H.,Schubert, D., and Riek, R. (2005) Proc. Natl. Acad. Sci. U. S. A. 102,17342–17347
24. Boshuizen, R. S., Morbin, M., Mazzoleni, G., Tagliavini, F., Meloen, R. H.,and Langedijk, J. P. (2007) Amyloid 14, 205–219
25. Fields, C. G., Lloyd, D. H., Macdonald, R. L., Otteson, K. M., and Noble,R. L. (1991) Pept. Res. 4, 95–101
26. Haik, S., Peyrin, J. M., Lins, L., Rosseneu, M. Y., Brasseur, R., Langeveld,J. P., Tagliavini, F., Deslys, J. P., Lasmezas, C., and Dormont, D. (2000)Neurobiol. Dis. 7, 644–656
27. Geysen, H. M., Meloen, R. H., and Barteling, S. J. (1984) Proc. Natl. Acad.Sci. U. S. A. 81, 3998–4002
28. Timmerman, P., Beld, J., Puijk, W. C., andMeloen, R. H. (2005) Chembio-chem. 6, 821–824
29. Raetz, C. R., and Roderick, S. L. (1995) Science 270, 997–100030. Iengar, P., Joshi, N. V., and Balaram, P. (2006) Structure 14, 529–54231. Tsai, H. H., Gunasekaran, K., and Nussinov, R. (2006) Structure 14,
1059–107232. Choi, J. H., Govaerts, C., May, B. C., and Cohen, F. E. (2008) Proteins 73,
150–16033. Salmona, M., Malesani, P., De Gioia, L., Gorla, S., Bruschi, M., Molinari,
A., Della Vedova, F., Pedrotti, B., Marrari, M. A., Awan, T., Bugiani, O.,Forloni, G., and Tagliavini, F. (1999) Biochem. J. 342, 207–214
34. Paravastu, A. K., Leapman, R. D., Yau, W. M., and Tycko, R. (2008) Proc.Natl. Acad. Sci. U. S. A. 105, 18349–18354
Heterologous Stacking by Prion Protein Peptides
12820 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 19 • MAY 8, 2009
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from
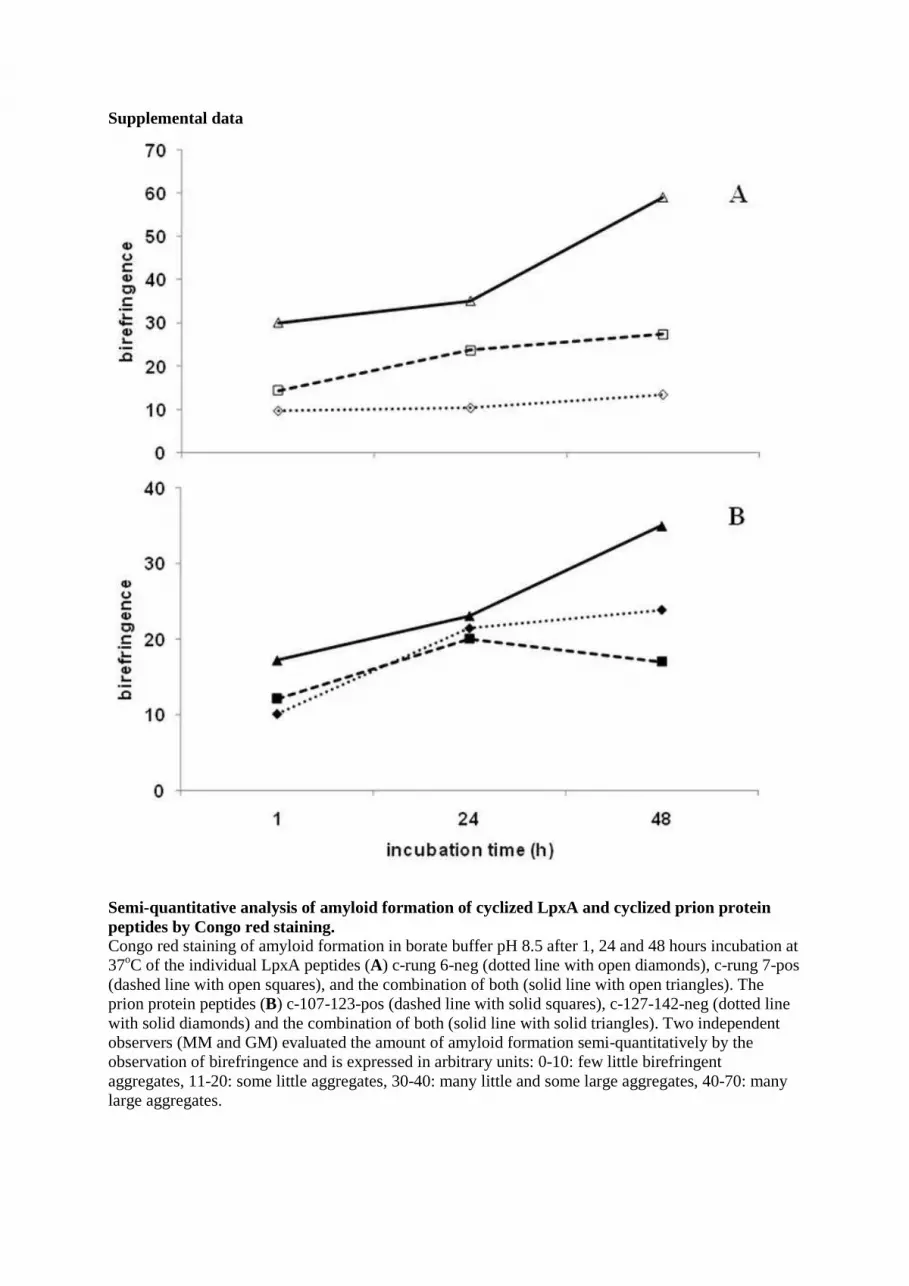
Supplemental data
Semi-quantitative analysis of amyloid formation of cyclized LpxA and cyclized prion protein peptides by Congo red staining. Congo red staining of amyloid formation in borate buffer pH 8.5 after 1, 24 and 48 hours incubation at 37oC of the individual LpxA peptides (A) c-rung 6-neg (dotted line with open diamonds), c-rung 7-pos (dashed line with open squares), and the combination of both (solid line with open triangles). The prion protein peptides (B) c-107-123-pos (dashed line with solid squares), c-127-142-neg (dotted line with solid diamonds) and the combination of both (solid line with solid triangles). Two independent observers (MM and GM) evaluated the amount of amyloid formation semi-quantitatively by the observation of birefringence and is expressed in arbitrary units: 0-10: few little birefringent aggregates, 11-20: some little aggregates, 30-40: many little and some large aggregates, 40-70: many large aggregates.

Meloen and Johannes P. M. LangedijkRonald S. Boshuizen, Veronica Schulz, Michela Morbin, Giulia Mazzoleni, Rob H.
Fibrils and Facilitates Complete Inhibition of Fibril GrowthHeterologous Stacking of Prion Protein Peptides Reveals Structural Details of
doi: 10.1074/jbc.M809151200 originally published online March 19, 20092009, 284:12809-12820.J. Biol. Chem.
10.1074/jbc.M809151200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2009/03/20/M809151200.DC1.html
http://www.jbc.org/content/284/19/12809.full.html#ref-list-1
This article cites 34 references, 16 of which can be accessed free at
by guest on February 9, 2016http://w
ww
.jbc.org/D
ownloaded from