the native-like conformation of ure2p in fibrils assembled under physiologically relevant conditions...
TRANSCRIPT

The native-like conformation of Ure2p in fibrils assembledunder physiologically relevant conditions switches to an
amyloid-like conformation upon heat-treatment of the fibrils
Luc Bousset,a Fatma Briki,b Jean Doucet,b,c and Ronald Melkia,*
a Laboratoire d�Enzymologie et Biochimie Structurales, CNRS, Baatiment 34 Avenue de la Terrasse, Gif-sur-Yvette Cedex FR-91198, Franceb Laboratoire d�Utilisation du Rayonnement Electromagn�eetique, Universit�ee Paris Sud, BP 34, Orsay 91898, France
c European Synchrotron Radiation Facility, Boııte Postale 220, Grenoble Cedex 38043, France
Received 11 September 2002, and in revised form 6 November 2002
Abstract
The [URE3] phenotype in the yeast Saccharomyces cerevisiae is inherited by a prion mechanism involving self-propagating Ure2p
aggregates. It is believed that assembly of intact Ure2p into fibrillar polymers that bind Congo Red and show yellow-green bire-
fringence upon staining and are resistant to proteolysis is the consequence of a major change in the conformation of the protein. We
recently dissected the assembly process of Ure2p and showed the protein to retain its native a-helical structure upon assembly into
protein fibrils that are similar to amyloids in that they are straight, bind Congo red and show green-yellow birefringence and have an
increased resistance to proteolysis (Bousset et al., 2002). Here we further show using specific ligand binding, FTIR spectroscopy and
X-ray fiber diffraction that Ure2p fibrils assembled under physiologically relevant conditions are devoid of a cross-b core. The X-ray
fiber diffraction pattern of these fibrils reveals their well-defined axial supramolecular order. By analyzing the effect of heat-treat-
ment on Ure2p fibrils we bring evidences for a large conformational change that occurs within the fibrils with the loss of the ligand
binding capacity, decrease of the a helicity, the formation of a cross-b core and the disappearance of the axial supramolecular order.
The extent of the conformational change suggests that it is not limited to the N-terminal part of Ure2p polypeptide chain. We show
that the heat-treated fibrils that possess a cross-b core are unable to propagate their structural characteristic while native-like fibrils
are. Finally, the potential evolution of native-like fibrils into amyloid fibrils is discussed.
� 2003 Elsevier Science (USA). All rights reserved.
Keywords: Prion; Saccharomyces cerevisiae; Ure2p; Assembly; Amyloid fibrils; Conformational transition; X-ray fiber diffraction
1. Introduction
An intriguing group of neurodegenerative diseases,
among which transmissible spongiform encephalopa-
thies (TSEs) in mammals are believed to be due to the
infectious protein (prion) PrP (Bolton et al., 1982). In
the yeast Saccharomyces cerevisiae, at least four proteinscause heritable, self-perpetuating, changes in phenotype.
These proteins are Ure2p, Sup35p, and Rnq1p, they are
responsible of the phenotypes [URE3] (Lacroute, 1971),
½PSIþ� (Cox, 1965), and ½PINþ� (induction of PSIþ)
(Derkatch et al., 2001; Sondheimer and Lindquist,
2000), respectively. A fourth protein, New1p (Santoso
et al., 2000), is believed to have prion properties but no
specific phenotype.
Ure2p is involved in a signal transduction pathway
that regulates nitrogen catabolism (Magasanik, 1992).
[URE3] is due to the self-propagation of altered formsof Ure2p (Wickner, 1994). In wild type cells, Ure2p is
dispersed throughout the cytoplasm while it forms large
globular or filiform aggregates in cells carrying the
[URE3] phenotype (Edskes et al., 1999; Fernandez-
Bellot et al., 2000). Overexpression of Ure2p 65 N-ter-
minal amino acid residues in wild type cells is sufficient
to induce de novo appearance of [URE3] while the ex-
pression of the complementary C-terminal region in
Journal of Structural Biology 141 (2003) 132–142
www.elsevier.com/locate/yjsbi
Journal of
StructuralBiology
* Corresponding author. Fax: +33-1-69-82-31-29.
E-mail addresses: [email protected] (J. Doucet), melki@-
lebs.cnrs-gif.fr (R. Melki).
1047-8477/03/$ - see front matter � 2003 Elsevier Science (USA). All rights reserved.
doi:10.1016/S1047-8477(02)00606-8

yeast cells lacking the URE2 gene restores the functionof the gene (Masison and Wickner, 1995). This is at the
origin of the definition of a prion-forming (residues 1–
65) and a functional (residues 65–354) part. The physical
boundary between the two domains was redefined and
their respective properties accessed using purified solu-
ble native Ure2p (Thual et al., 1999). The N-terminal
region extending from residues 1–93 is poorly structured
while the C-terminal region extending from residues 94–354 is compactly folded (Thual et al., 1999). The crystal
structure of Ure2p 95–354 was solved (Bousset et al.,
2001a; Umland et al., 2001). Ure2p 95–354 is highly a-helical. It is formed by two sub-domains and has a fold
similar to that of glutathione-S-transferases (GSTs)
(Board et al., 2000). It binds glutathione (GSH) and
related compounds with high affinity in a cleft running
along the domain interface that resembles the active siteof GSTs (Bousset et al., 2001b).
We have shown previously that authentic Ure2p can
assemble in vitro into fibrils under physiologically rele-
vant conditions (Thual et al., 2001, 1999). Assembly is
accompanied by an increase in the resistance to prote-
olysis (Thual et al., 1999). Interestingly however, the
degradation patterns of soluble and assembled Ure2p
are very similar suggesting that the protein remains in anative-like conformation in the fibrils. Ure2p fibrils bind
the dye Congo red (Thual et al., 1999). This finding by
itself does not mean that the fibrils are amyloids since
binding of Congo red is not specific of amyloids
(Khurana et al., 2001). Binding is however accompanied
by the amyloid characteristic yellow-green birefringence
in cross-polarized light (Taylor et al., 1999).
A large number of proteins that possess unrelatedstructures form amyloid-like fibrils under given experi-
mental conditions (Carrell and Gooptu, 1998; Fandrich
et al., 2001; Krebs et al., 2000; Lim et al., 2000;
Morozova-Roche et al., 2000). This suggests that a
common factor drive the formation of fibrils despite
their highly differing primary structures. Up to now, this
factor was thought to be a partial unfolding–refolding
process (Dobson, 1999), leading to the formation ofintermolecular bonding through b-sheets. These b-sheetsform a cross-b core running all along the fibril with the
hydrogen-bonded polypeptide chains running normal to
the fibril direction and show a characteristic cross-bmeridional 4.7–4.8�AA reflection in X-ray fiber diffraction
patterns (Sunde et al., 1997).
We recently dissected the assembly process of Ure2p
and showed the protein to retain its native a-helicalstructure upon assembly into protein fibrils (Bousset
et al., 2002). The absence of a major conformational
change upon Ure2p assembly into fibrils that exhibit
several characteristics of amyloids can be readily ex-
plained if one assumes that the conformational change
that drives Ure2p assembly only affects the poorly
structured N-terminal domain of the protein. This
region extends over 65–94 amino acid residues out of thetotal 354 amino acids of Ure2p.
Here we further show using X-ray fiber diffraction
that Ure2p fibrils assembled under physiologically rele-
vant conditions are devoid of a cross-b structure. While
trying to improve Ure2p fibrils alignment to elucidate
the structure of the fibrillar form of Ure2p, we observed
a change in their architectures. While the a-helicalcontent of Ure2p in fibrils assembled at 20 �C was verysimilar to that of the native protein, fibrils exposed
to 60 �C were essentially made of b-sheets. The two
kinds of fibrils are indistinguishable in the electron
microscope. The observation of protein fibrils with dif-
ferent structures and morphologies, either within the
same preparation, or under different physico-chemical
conditions, has already been reported (Kad et al., 2001),
but the absence of a cross-b core signal is withoutprecedent for amyloid fibrils. The absence of a cross
b-sheet core in Ure2p fibrils obtained under physiolog-
ical conditions raises the question of the necessity of
such a core in fibril formation and of its role in their
stabilization.
Upon heating the native-like fibrils in solution, a
change in their architecture occurs with the formation of
a cross b-sheet core. The proteinase K degradationpatterns of Ure2p in native-like and heated fibrils are
unrelated. Moreover, while Ure2p in native-like fibrils
binds GSH in a manner indistinguishable from that of
the native unassembled form of the protein (Bousset et
al., 2001a), the heat-treated fibrils have lost their binding
capacity. All together, our data indicate that a major
change in the conformation of Ure2p within the fibrils
occurs upon heating. This change corresponds to thetransition from a mainly a-helical to a mainly b-sheetstructure, i.e., a transition from a helical polymer to an
amyloid fibril. Interestingly, while native-like fibrils
propagate in a catalytic manner upon dilution in the
presence of soluble Ure2p, heat-treated fibrils are inert
end products. This indicates that the cross-b rich Ure2p
fibrils are irrelevant to the prion concept while native-
like fibrils are.
2. Materials and methods
2.1. Purification and crystallization
Full length Ure2p and its C-terminal region extending
from residues 95–354 were overexpressed in Escherichiacoli and purified as previously described (Thual et al.,
2001, 1999). Protein concentrations were determined
spectrophotometrically (HP 8453 diode array spectro-
photometer, Hewlett-Packard) using an extinction co-
efficient of 0.67mg cm2 at 280 nm and a molecular
weight of 40 200 or by the Bradford methods (Bradford,
1976). Crystals of full length Ure2p and Ure2p 95–354
L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142 133

were obtained using the crystallization conditions de-scribed in (Bousset et al., 2001a).
2.2. Assembly of Ure2p into fibrils
Assembly of Ure2p into fibrils was monitored using a
thioflavin-T binding assay (McParland et al., 2000).
Ure2p (60 lM) in 20mM Tris, pH 7.5, 100mM KCl was
incubated at 7 �C. At regular time intervals, 8 ll aliquotswere removed from the solution, mixed with 300 ll of
thioflavin-T (10 lM) from Sigma and incubated at 20 �Cfor 10min. Thioflavin-T binding was measured by av-
eraging the emission signal over 30 s using an AM-
INCO-Bowman series2 spectrofluorometer set at 440 nm
(excitation) and 480 nm (emission). Ure2p fibrils were
negatively stained on carbon-coated grids (200 mesh)
with 1% uranyl acetate and examined in a Philips EM410 electron microscope.
2.3. Binding of acetyl-2-dimethylaminonaphtalene gluta-
thione to Ure2p fibrils
Glutathione (Roche), 50mM, in HEPES, pH 7.5,
was incubated with 20mM 6-bromoacetyl-2-dim-
ethylaminonaphtalene (Molecular Probes, Eugene, OR)for 2 h at room temperature in order to allow the for-
mation of a thioether bond between the SH group of the
molecule and the probe. Binding of acetyl-2-dim-
ethylaminonaphtalene-glutathione (ADAN-glutathione)
to untreated or heat-treated Ure2p fibrils in 20mM Tris,
pH 7.5, 100mM KCl was monitored in a 10� 2mm
quartz cuvette (Hellma) thermostated at 20 �C in a
AMINCO-Bowman series2 spectrofluorometer. Theexcitation and emission monochromators were set at 380
and 500 nm, respectively.
2.4. Proteolytic digestions
Assembly of Ure2p (2.0mg/ml) in 20mM Tris, pH
7.5, 100mM KCl into fibrils was allowed to completion
by incubating soluble Ure2p for 80 h at 7 �C. The samesample was then split into two aliquots. The first was
incubated without any additional treatment at 37 �Cwith Proteinase K from Roche (2.4 lg/ml), while the
second was incubated for 60min at 60 �C prior to the
proteolytic treatment. Aliquots from each sample were
removed at different time intervals following addition of
the protease and transferred into Eppendorf tubes
maintained at 95 �C containing sample denaturing buffer(50mM Tris–HCl, pH 6.8, 4% SDS, 2% b-mercap-
toethanol, 12% glycerol 0.01% bromophenol blue) in
order to arrest immediately the cleavage reaction. After
incubation of each tube for 10min at 95 �C, the samples
were processed to monitor the time course of Ure2p
cleavage by SDS–PAGE (Laemmli, 1970).
2.5. X-ray diffraction (XRD)
Assembly of Ure2p (4.0mg/ml) in 20mM Tris, pH
7.5, 100mM KCl into fibrils was allowed to completion
by incubating soluble Ure2p for 48 h at 20 �C. The
sample was then dialyzed against water for 4 h and split
into two aliquots. One of the aliquots was heated for 1 h
at 60 �C the other was untreated. About 25 ll of each
aliquot were introduced into glass capillaries 1.5mmdiameter. The fibrils were aligned either in a magnetic
field (1 T) at 20 �C for one week or evaporated at room
temperature. The material was concentrated following
these treatments into a thin disk of about 0.3mm
thickness, delimited by the capillary cross-section.
X-ray data collection was carried out on two syn-
chrotron beamlines, D43 at LURE (Orsay, France) and
ID14-EH1 at the European Synchrotron RadiationFacility (Grenoble, France). D43 beamline uses a colli-
mated and monochromatic incident beam (0.5mm di-
ameter cross-section, wavelength 1.45�AA) selected by a
Ge (1 1 1) bent monochromator. A transmission geom-
etry setting was chosen with a sample-detector distance
of 100mm, giving access to periodicities 30 and 2.5�AA.
The two-dimensional scattering patterns were recorded
on FUJI PhosphorImage plates located perpendicular tothe incident beam and read on a AMERSHAM scanner
(STORM) with a 200 lm pixel size and 0.08� angular
resolution. The X-ray beam (wavelength 0.934�AA) had a
diameter of 50 lm on ID14-EH1 and allowed the
analysis of sample sections 100 times smaller than at
LURE, thus increasing the probability to focus
the beam on well-oriented regions. The combination of
the 80 lm pixel size of the MarResearch CCD detectorwith the 150mm sample-to-detector distance gave access
to periodicities between 80 and 2�AA, with an angu-
lar resolution of 0.02�. Data processing involved sub-
traction of the scattering pattern due to the glass
capillary.
2.6. Fourier transform infrared spectrometry (FT-IR)
Infrared spectra were collected using a Spectra Tech
infrared microscope equipped with a small aperture
ð5� 5lm2Þ at the synchrotron radiation facility,
Beamline MIRAGE (Polack et al., 1999) at LURE,
Orsay, France. Spectra of untreated and heat-treated
Ure2p fibrils recovered by sedimentation at 100 000g for
15min and that of crystals of Ure2p 95–354 were re-
corded at 8 cm�1 resolution. Five hundred and twelvespectra were collected before performing Fourier
transform. A linear baseline subtraction was performed
to account for the gradual decay in the beam intensity
due to synchrotron source decay. The amide I band
(1600–1700 cm�1) of the spectra was subjected to a fit-
ting procedure using six gaussian distributions centered
134 L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142

at the frequencies of well characterized secondarystructures (Stuart, 1997). Each secondary structure is
characterized by its frequency and by an interval of a
given width. During the fitting procedure the peak po-
sitions were free to vary inside that interval. Each peak
width was limited to 25 cm�1 while peak height was free.
The fitting was achieved using the fitting �module� of theOrigin package. It is worthwhile to note that all the
frequencies were present on the second derivative of thespectra. The contribution of each curve to the amide I
band was assessed by integrating the area under the
curve and then normalizing to the total area under the
amide I band.
3. Results
3.1. Untreated and heat-treated Ure2p fibrils differ in their
ligand binding capacities
Native unassembled Ure2p binds with high affinity
ADAN-GSH, a substrate analogue of GSH (Bousset et
al., 2001b). The GSH binding site is located in the
functional domain of the protein, which is made of twosub-domains connected by a short linker region (Bousset
et al., 2001a), each of which is involved in substrate
binding site. ADAN-GSH is thus an excellent tool for
probing whether the fibrils contain native-like subunits.
Fig. 1. Native-like and heat-treated Ure2p fibrils differ by their binding capacity of GSH but are indistinguishable on electron micrographs. (A)
Saturation of native-like (�) and heat-treated (N) Ure2p fibrils (1mg/ml) by ADAN-GSH. (B and C) Negative stained electron micrographs of
native-like and heat-treated Ure2p fibrils, respectively. Bar, 0.2lm.
L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142 135

Fig. 1A shows the saturation by ADAN-GSH of Ure2pfibrils generated under physiologically relevant condi-
tions. Comparison of the data with that obtained for
soluble dimeric Ure2p (Bousset et al., 2001b) strongly
suggests that the C-terminal domain of the protein re-
tains a native conformation within the fibrils. The
ADAN-GSH binding capacity of Ure2p is lost upon
incubation of native-like fibrils in solution at 60 �C for
1 h (Fig. 1A). We conclude from this observation that aconformational rearrangement occurs during heat
treatment that reveals by the loss of GSH binding ca-
pacity. To determine whether this change shows in the
overall shape of the fibrils, untreated and heat-treated
Ure2p fibrils were observed in the electron microscope
following negative staining. The two kinds of fibrils were
found indistinguishable (Figs. 1B and C). Both kinds of
fibrils are 20 nm wide and over 1 lm long. We con-clude from this observation that the conformational
rearrangement that leads to the loss of ligand binding
capacity of Ure2p does not perturb the overall mor-
phology of the fibrils. We further conclude that heat
treatment of the native-like assembled form of Ure2p
in solution is not sufficient to disrupt their fibrillar
structure.
3.2. The proteolytic patterns of untreated and heat-treated
Ure2p fibrils differ significantly
Using proteinase K treatment, we previously showed
that the time courses of degradation of the soluble and
assembled forms of Ure2p differ significantly. Interest-
ingly however the patterns show no significant differ-
ences in the polypeptide species generated which suggestthat the overall conformation of Ure2p is not altered
upon assembly into fibrils (Thual et al., 1999). The
degradation pattern of soluble and assembled Ure2p
was further shown to be indistinguishable from that of
Ure2p 95–354 indicating that the polypeptide species
generated upon proteinase K treatment are derived from
the C-terminal domain of the protein (Thual et al.,
2001). Proteinase K treatment is thus an excellent toolfor probing conformational changes that occurs in the
compactly folded, mainly a-helical C-terminal domain
of Ure2p. Fig. 2 shows the degradation products of
untreated and heat-treated Ure2p fibrils. In native-like
fibrils, full-length Ure2p is degraded within 2min into a
polypeptide with a molecular mass of 30 kDa that cor-
responds to Ure2p 95–354. Six other polypeptides with
apparent molecular masses 29, 24, 22, 20, 19, and14 kDa are generated. Three polypeptides resist the
treatment for over 30min (29, 20, and 14 kDa). They are
degraded 60min after the onset of the cleavage reaction.
Full-length Ure2p resists proteinase K treatment for
over 5min in heat-treated fibrils. Its degradation at early
times following the onset of proteinase K treatment
generate a cluster of polypeptides that have apparent
molecular masses in the range 34–27 kDa. These trun-
cated Ure2p polypeptides represent over 90% of the
degradation products. Minor polypeptides with appar-
ent molecular masses between 26 and 20 kDa are also
generated. At later stages of the degradation reaction
three polypeptides with apparent molecular masses 30,29, and 23 kDa that differ from those generated from
untreated fibrils are observed. We conclude from these
observations that a major change in the conformation of
Ure2p in the fibrils occurs during heating with the loss
or protection of several proteinase K cleavage sites
probably due to increased packing of Ure2p molecules
within the fibrils.
3.3. XRD reveals different molecular architectures for
native-like and heat-treated Ure2p fibrils
Examination of the X-ray scattering patterns from
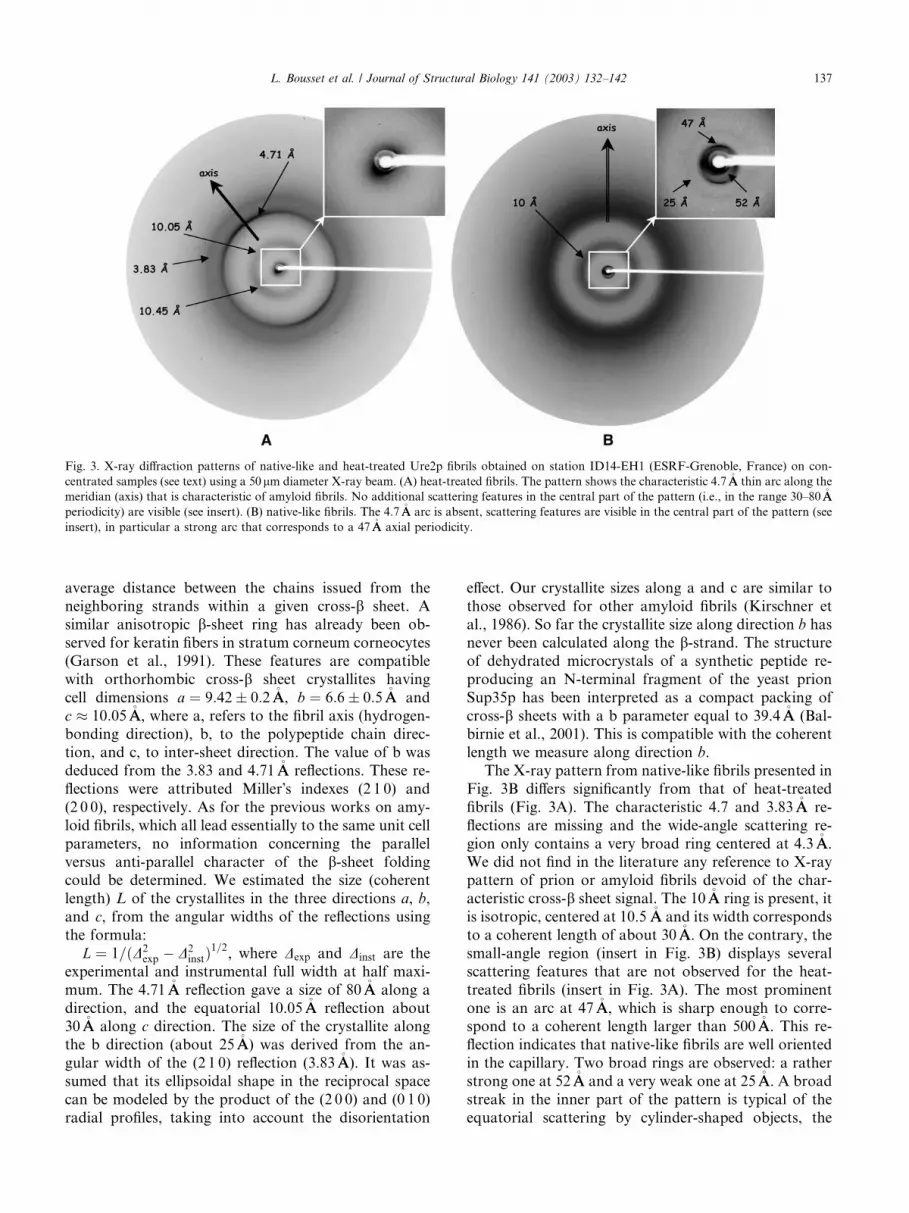
heat-treated, Fig. 3A, and native-like fibrils Fig. 3B,
reveals changes in the molecular organization of Ure2p
within the fibrils.
The pattern in Fig. 3A resembles the classical patternof others amyloid fibrils, with the characteristic cross-bsheet reflection at 4:71 0:01�AA spacing, the rather
broad ring around 10�AA, and another reflection at
3:83 0:02�AA. The pattern is anisotropic, the 4.71 and
3.83�AA rings are slightly reinforced along a direction
which can be assumed to be the fibrils direction (me-
ridian). The intensity profile of the 4.71�AA reflection is
regular and unlikely to be due to the overlap of the re-flections of several structural features. More interesting,
the 10�AA ring is also anisotropic, its maximum along the
meridian and the equator are respectively located at
10:05 0:05 and 10:45 0:05�AA. This clearly indicates
that the ‘‘10 �AA’’ ring originates from two different
structural features, the cross b intersheet distance gives
rise to the equatorial reflection, whilst the origin of the
meridional reflection is unclear. It could come from an
Fig. 2. The degradation patterns of native-like and heat-treated Ure2p
fibrils are unrelated. Native-like (A) and heat-treated (B) Ure2p fibrils
were subjected to proteinase K treatment, and the time courses of the
digestions monitored by SDS–PAGE (15%) followed by immunode-
tection using an anti-Ure2p polyclonal antibody. Time points (in
minutes) are shown at the top of each gel. Molecular mass markers (in
kDa) and the migration front (MF) of the gel are shown on the left.
136 L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142
average distance between the chains issued from the
neighboring strands within a given cross-b sheet. A
similar anisotropic b-sheet ring has already been ob-
served for keratin fibers in stratum corneum corneocytes
(Garson et al., 1991). These features are compatible
with orthorhombic cross-b sheet crystallites having
cell dimensions a ¼ 9:42 0:2�AA, b ¼ 6:6 0:5�AA and
c � 10:05�AA, where a, refers to the fibril axis (hydrogen-bonding direction), b, to the polypeptide chain direc-
tion, and c, to inter-sheet direction. The value of b was
deduced from the 3.83 and 4.71�AA reflections. These re-
flections were attributed Miller�s indexes (2 1 0) and
(2 0 0), respectively. As for the previous works on amy-
loid fibrils, which all lead essentially to the same unit cell
parameters, no information concerning the parallel
versus anti-parallel character of the b-sheet foldingcould be determined. We estimated the size (coherent
length) L of the crystallites in the three directions a, b,
and c, from the angular widths of the reflections using
the formula:
L ¼ 1=ðD2exp � D2
instÞ1=2, where Dexp and Dinst are the
experimental and instrumental full width at half maxi-
mum. The 4.71�AA reflection gave a size of 80�AA along a
direction, and the equatorial 10.05�AA reflection about30�AA along c direction. The size of the crystallite along
the b direction (about 25�AA) was derived from the an-
gular width of the (2 1 0) reflection (3.83�AA). It was as-
sumed that its ellipsoidal shape in the reciprocal space
can be modeled by the product of the (2 0 0) and (0 1 0)
radial profiles, taking into account the disorientation
effect. Our crystallite sizes along a and c are similar to
those observed for other amyloid fibrils (Kirschner et
al., 1986). So far the crystallite size along direction b has
never been calculated along the b-strand. The structure
of dehydrated microcrystals of a synthetic peptide re-
producing an N-terminal fragment of the yeast prion
Sup35p has been interpreted as a compact packing of
cross-b sheets with a b parameter equal to 39.4�AA (Bal-birnie et al., 2001). This is compatible with the coherent
length we measure along direction b.
The X-ray pattern from native-like fibrils presented in
Fig. 3B differs significantly from that of heat-treated
fibrils (Fig. 3A). The characteristic 4.7 and 3.83�AA re-
flections are missing and the wide-angle scattering re-
gion only contains a very broad ring centered at 4.3�AA.
We did not find in the literature any reference to X-raypattern of prion or amyloid fibrils devoid of the char-
acteristic cross-b sheet signal. The 10�AA ring is present, it
is isotropic, centered at 10.5 �AA and its width corresponds
to a coherent length of about 30�AA. On the contrary, the
small-angle region (insert in Fig. 3B) displays several
scattering features that are not observed for the heat-
treated fibrils (insert in Fig. 3A). The most prominent
one is an arc at 47�AA, which is sharp enough to corre-spond to a coherent length larger than 500�AA. This re-
flection indicates that native-like fibrils are well oriented
in the capillary. Two broad rings are observed: a rather
strong one at 52�AA and a very weak one at 25�AA. A broad
streak in the inner part of the pattern is typical of the
equatorial scattering by cylinder-shaped objects, the
Fig. 3. X-ray diffraction patterns of native-like and heat-treated Ure2p fibrils obtained on station ID14-EH1 (ESRF-Grenoble, France) on con-
centrated samples (see text) using a 50lm diameter X-ray beam. (A) heat-treated fibrils. The pattern shows the characteristic 4.7�AA thin arc along the
meridian (axis) that is characteristic of amyloid fibrils. No additional scattering features in the central part of the pattern (i.e., in the range 30–80�AA
periodicity) are visible (see insert). (B) native-like fibrils. The 4.7�AA arc is absent, scattering features are visible in the central part of the pattern (see
insert), in particular a strong arc that corresponds to a 47�AA axial periodicity.
L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142 137
fibrils in this case. Therefore, the 47�AA sharp arc is me-
ridional and reveals the existence of a long-range peri-
odicity along the fibrils. The broadness of the 52 and25�AA rings indicate that these features probably arise
from lateral intra-fibril interferences between protofibril
subunits. We conclude from these observations that
native-like and heat-treated fibrils possess each a char-
acteristic architecture. The two architectures are dis-
tinct. This reflects two different spatial organizations ofUre2p molecules within the two kinds of fibrils. We
further conclude that the characteristics of native-like
fibrils differ significantly from that of authentic amy-
loids.
3.4. Native-like and heat-treated Ure2p fibrils differ by
their secondary structural content
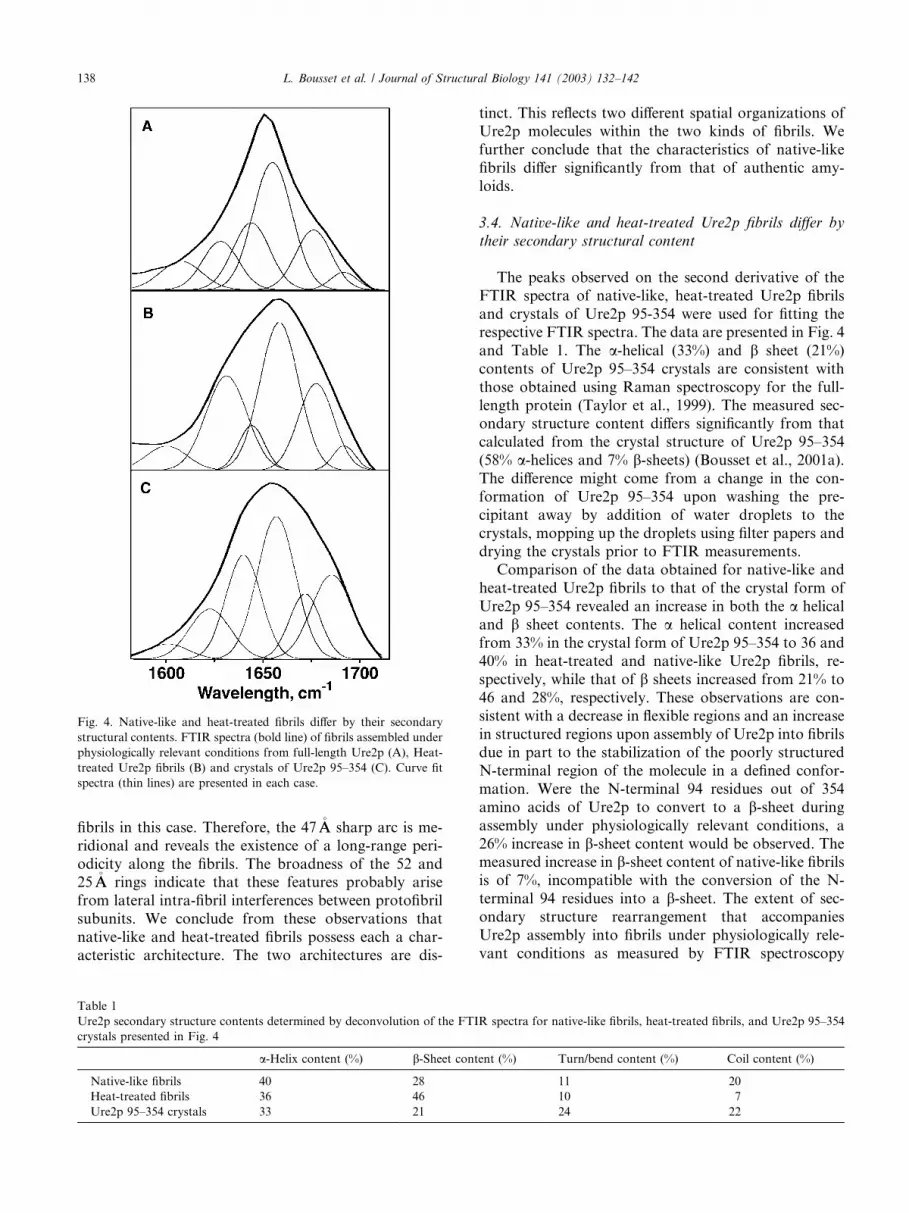
The peaks observed on the second derivative of the
FTIR spectra of native-like, heat-treated Ure2p fibrils
and crystals of Ure2p 95-354 were used for fitting the
respective FTIR spectra. The data are presented in Fig. 4
and Table 1. The a-helical (33%) and b sheet (21%)
contents of Ure2p 95–354 crystals are consistent with
those obtained using Raman spectroscopy for the full-
length protein (Taylor et al., 1999). The measured sec-ondary structure content differs significantly from that
calculated from the crystal structure of Ure2p 95–354
(58% a-helices and 7% b-sheets) (Bousset et al., 2001a).
The difference might come from a change in the con-
formation of Ure2p 95–354 upon washing the pre-
cipitant away by addition of water droplets to the
crystals, mopping up the droplets using filter papers and
drying the crystals prior to FTIR measurements.Comparison of the data obtained for native-like and
heat-treated Ure2p fibrils to that of the crystal form of
Ure2p 95–354 revealed an increase in both the a helical
and b sheet contents. The a helical content increased
from 33% in the crystal form of Ure2p 95–354 to 36 and
40% in heat-treated and native-like Ure2p fibrils, re-
spectively, while that of b sheets increased from 21% to
46 and 28%, respectively. These observations are con-sistent with a decrease in flexible regions and an increase
in structured regions upon assembly of Ure2p into fibrils
due in part to the stabilization of the poorly structured
N-terminal region of the molecule in a defined confor-
mation. Were the N-terminal 94 residues out of 354
amino acids of Ure2p to convert to a b-sheet during
assembly under physiologically relevant conditions, a
26% increase in b-sheet content would be observed. Themeasured increase in b-sheet content of native-like fibrils
is of 7%, incompatible with the conversion of the N-
terminal 94 residues into a b-sheet. The extent of sec-
ondary structure rearrangement that accompanies
Ure2p assembly into fibrils under physiologically rele-
vant conditions as measured by FTIR spectroscopy
Fig. 4. Native-like and heat-treated fibrils differ by their secondary
structural contents. FTIR spectra (bold line) of fibrils assembled under
physiologically relevant conditions from full-length Ure2p (A), Heat-
treated Ure2p fibrils (B) and crystals of Ure2p 95–354 (C). Curve fit
spectra (thin lines) are presented in each case.
Table 1
Ure2p secondary structure contents determined by deconvolution of the FTIR spectra for native-like fibrils, heat-treated fibrils, and Ure2p 95–354
crystals presented in Fig. 4
a-Helix content (%) b-Sheet content (%) Turn/bend content (%) Coil content (%)
Native-like fibrils 40 28 11 20
Heat-treated fibrils 36 46 10 7
Ure2p 95–354 crystals 33 21 24 22
138 L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142

further confirm the lack of cross b-core in native-likefibrils as observed in X-ray fiber diffraction images. We
conclude, therefore that the fibrils do not comprise a
cross b-core that is typical of amyloids. In contrast, heat
treatment of Ure2p fibrils increases significantly (65%)
their b sheet content with the concomitant appearance
of the 4.7�AA band in X-ray fiber diffraction images typ-
ical of amyloids.
4. Discussion
The data presented above further support our recent
findings that the fibrils assembled from full-length
Ure2p under physiologically relevant conditions despite
having many properties akin to amyloid such as an in-
creased resistance to proteolysis and yellow-green bire-fringence upon Congo Red binding are devoid of the
amyloid characteristic cross b-sheet core (Bousset et al.,
2002). When native-like fibrils are heated, a major
conformational change occurs with the constitution of a
cross b-sheet core typical of amyloid fibrils. This rear-
rangement is accompanied by a major conformational
change in Ure2p with the loss of the ability to bind
GSH, a change in the proteolytic pattern and an in-crease in b-sheet content in the fibrils. The transition
from a helical polymer made of native-like polypeptide
chains to a cross-b structure made of non-native mole-
cules does not reveal in a change in fibril morphology.
Indeed, electron micrographs of untreated and heat
treated fibrils are indistinguishable.1 This transition
shows however in X-ray diffraction patterns and FTIR
measurements.A plausible low-resolution structural model for na-
tive-like and heat-treated Ure2p fibrils needs to account
for EM, FT-IR, and XRD observations. Native-like fi-
brils contain no cross-b core. Their longitudinal orga-
nization is dominated by a 47�AA long-range axial repeat
that does not arise from a helical coil but probably from
the constituting Ure2p molecules since it corresponds
with one of the dimensions of Ure2p. Indeed, the vol-ume of hydrated Ure2p monomer derived from the
crystal structure of Ure2p 95–354 (Bousset et al., 2001a)
is 69:2� 27:8� 52:1�AA3
while that of the dimer is
83:1� 50:4� 53:6�AA3. Native-like fibrils are built either
of monomeric or dimeric molecules associated in a non-
native manner (Bousset et al., 2002). The 47�AA repeat is
compatible with either one of the dimensions of the
monomer or with that of the dimer. Furthermore, thepresence of equatorial broad reflections at 52 and 25 �AAsuggest that Ure2p fibrils could be constituted by the
lateral association of protofibrils that are about 50 �AA in
diameter. The scattered intensity IðSÞ by one fibril could
be modeled as: IðSÞ ¼ ZðSÞ:jF ðSÞj2, where ZðSÞ is the
interference function, i.e. the Fourier transform of the
distribution function of the protofibrils within the fibril,
and F ðSÞ is the Fourier transform of the electron densityof the protofibril section. If we assume that the pro-
tofibrils are cylindrical in shape (with a radius r),
F ðSÞ / r2J1ðuÞ=u, with u ¼ 2prS. The scattering inten-
sity assuming a square lattice with unit-cell length equal
to 50�AA leads to scattering peaks close to 50 and 25 �AA.
Heat-treated Ure2p fibrils although indistinguishable
from native-like fibrils in the electron microscope have
lost the axial 47�AA repeat that characterizes native-likefibrils. Thus heat treatment yields fibrils devoid of the
native-like fibril characteristic axial repeat. These fibrils
show instead the typical cross b-sheet core of amyloids.
The radial size of the b-sheet core is 30�AA. Strikingly
however and in contrast to models of amyloid fibrils
made of transthyretin (Blake and Serpell, 1996) and the
SH3 domain of bovine phosphatidyl-inositol kinase
p85a subunit (Jimenez et al., 1999), Ure2p fibrils exhibita rather small coherence length (80�AA) which indicates
that their b-core is short ranged.
Although the coherence length is small, it is reason-
able to consider that the fibrils are made of a continuous
cross-b core running along their axis. The limited
number of scattering features in the XRD pattern indi-
cate that heat-treated Ure2p fibrils differ from other
amyloid fibrils (Blake and Serpell, 1996; Jimenez et al.,1999) in that they show no helical axial repeat.
The 30�AA b-sheet core is due to a conformational
change either within a protofibril or at the interface be-
tween two adjacent protofibrils. Given that the overall b-sheet content of the fibrils is 50%, the surface area that is
not involved in the b core must represent the remaining
50% of the overall surface in a fibril cross-section. A
scheme summarizing these parameters is shown in Fig. 5.Interestingly, the diameter of these protofibrils remains
similar to that made of native-like polypeptide chains,
thus accounting for the absence of measurable change in
fibril width as determined by electron microscopy.
4.1. Biological significance
Yeast prions assemble in vitro in a catalytic mannerinto fibrils. The limiting step in the assembly reaction is
the nucleation step (Glover et al., 1997; Thual et al.,
1999). This is strongly in favor of a polymerization
model in which prion molecules assemble in a manner
similar to the crystallization of a solute molecule in so-
lution. In such a process the energetically unfavorable
nucleation step is followed by a more favorable elon-
1 The coherence length calculated from the X-ray diffraction
patterns is not readily comparable to the persistence length of the
fibrils that can be derived from electron micrographs. Indeed, the loss
of regular periodicity does not necessarily show in a change in the
rigidity of a fibril. It is therefore not unusual to observe similar
morphologies for native-like and heat-treated fibrils at the electron
microscope resolution used in this study.
L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142 139

gation reaction that proceeds in a closed system until
equilibrium is reached between unassembled and poly-
meric species. It is reasonable to envisage that Ure2p has
the ability to assemble into different polymers in a
manner similar to the crystallization of a large number
of proteins in different crystal lattices. Although thepresent work does not bring evidence for such a process,
we clearly show that a major conformational change
occurs upon heating the fibrils. Such a change is similar
to that observed upon heating protein polymers (Ast-
bury and Woods, 1933).
The finding that native-like and heat-treated Ure2p
fibrils are indistinguishable in the electron microscope
although they differ both in their overall secondarystructure content and their intrinsic supramolecular or-
ganization suggests that the fibrils could be made of the
arrangement of Ure2p polypeptide chains in at least two
manners. Such different arrangements should reflect in
the elongation properties of the fibrils since the geome-
try of polymers ends regulates their growth rate by
favoring or unfavoring intermolecular interactions be-
tween soluble Ure2p polypeptide chains and fibrils ends.Alternatively, the growth rate of the two kinds of fibrils
may be regulated by the higher or lower propensity of
defined Ure2p unfolding intermediates to interact with
fibril ends. Finally, such different arrangements are
expected to imprint a specific orientation to Ure2pmolecules upon incorporation at fibrils ends leading to
the propagation of a defined structural motif that could
account for the experimental differences in conver-
sion efficiencies at the origin of the strain concept in
prion propagation (Schlumpberger et al., 2001; Uptain
et al., 2001).
To determine whether the two types of Ure2p fibrils
can propagate their respective structural characteristicsduring the construction of the fibrils by either selecting
Ure2p molecules in different conformations or imprint-
ing a specific orientation or conformation to newly in-
corporated Ure2p molecules, native-like seeds and heat
treated seeds were diluted in the presence of soluble
Ure2p and the structure of newly assembled fibrils as-
sessed by proteinase K treatment. Ure2p was found to
assemble into the native-like type of fibrils when seededeither by native-like or heat-treated fibrils (not shown).
This result either indicates that heat-treated fibrils ends
are indistinguishable structurally from that of native-
like fibrils or that they are unable to elongate. To dis-
tinguish between the two possibilities, Ure2p assembly
kinetics were measured following seeding by the two
kinds of fibrils. The data are shown in Fig. 6. The as-
sembly of Ure2p in the presence of native-like fibrils wasfound to proceed without a lag phase in accord with the
limiting nucleation phase being bypassed. In contrast, a
lag phase precedes the elongation phase in Ure2p as-
sembly reactions in the presence of heat-treated fibrils
indicating de novo nucleation of native-like Ure2p fibrils.
Thus, Ure2p does neither interact with heat-treated fibril
ends nor incorporate into such preformed fibrils.
We recently proposed by analogy to the mechanismof assembly of serpins into protein fibrils (Carrell and
Gooptu, 1998) two scenarios that account for the lim-
ited structural rearrangement at the origin of Ure2p
assembly under physiologically relevant conditions
Fig. 6. Native like fibrils seed Ure2p assembly into protein fibrils, while
heat-treated fibrils do not. Time courses of Ure2p (170lM) assembly
into protein fibrils in 50mM Tris, pH 7.5, 100mM KCl, in the absence
of added nuclei (d, dotted line), upon addition of 5.5lM pre-assem-
bled native-like (j) or 5.5 lM heat-treated (m, plain line) Ure2p fibrils.
The assembly of Ure2p into protein fibrils was monitored by Thio-
flavin-T binding.
Fig. 5. Schematic representation of a cross b-sheet core indicating the
dimensions of the unit cell and the domain size of heat-treated Ure2p
fibres. The folding elements that spread from the core are purely hy-
pothetical. They are represented to underline the existence of non b-sheet parts representing about 50% of the molecules that are located
outside the core.
140 L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142

(Bousset et al., 2002). Other scenarios where b-sheetsorganized in helices running along the core of the fibrils
have been recently proposed for the assembly of syn-
thetic polypeptides into amyloid fibrils (Perutz et al.,
2002; Wille et al., 2002). In these models, the b-sheetcore of the fibrils is therefore expected to resist prote-
olysis. These scenarios do not apply for the assembly of
full-length Ure2p into fibrils under physiologically rel-
evant conditions as the degradation patterns of solubleand assembled Ure2p generate identical polypeptides
(Thual et al., 1999) and the fibrils lack the 4.7�AA reflec-
tion that characterizes amyloids. The finding that heat-
treatment of native like Ure2p fibrils generates b-sheetrich fibrils indicates that parts of Ure2p molecules in the
fibrils are flexible and that the heat-induced structural
rearrangement leads to the reorganization within the
fibrils of Ure2p flexible parts into anti-parallel b-sheetthat may be organized in helices. Ure2p N-terminal
domain and Ure2p 94–200 fragment that contains a b-sheet (Bousset et al., 2001a) have the highest probability
to be involved in such a transition. Thus, future efforts
to identify Ure2p regions that are at the origin of the a-helical to b-sheet transition would allow a better com-
prehension of the flexibility of Ure2p within the fibrils.
Amyloid formation is believed to be a generic prop-erty of polypeptide chains. It is also believed that the
form of a given protein that assembles into amyloid fi-
bers is a partially unfolded polypeptide chain. The fact
that only a subset of proteins possess the capacity to
form such polymers under non-denaturing conditions
has been attributed to an insufficient amount of folding
intermediate that plays the role of amyloid precursor
(Dobson, 1999). To populate the folding intermediatesthat are precursors of amyloid fibrils it is common to
incubate various polypeptide chains at high tempera-
tures and/or extreme pHs (Jimenez et al., 2002). The
transition we report here differs significantly from the
partial unfolding of a polypeptide chain in solution and
its consequent assembly into amyloid fibrils as the a-helical to b-sheet transition occurs within the fibrils.
Finally, the ability of Ure2p fibrils obtained underphysiological conditions to evolve in vivo into the b-sheet rich fibrils remains to be determined and the
function of such an evolution elucidated. It is important
to bear in mind though that while native-like fibrils as-
semble in a nucleation-dependent manner, the nucle-
ation phase being abolished upon addition of minute
amounts of preformed fibrils to soluble Ure2p (Thual et
al., 1999), a property central to prion propagation(Bousset and Melki, 2002), heat-treated fibrils do not
meet with this property. This strongly suggests that b-sheet rich fibrils if formed in vivo are inert end products.
In the yeast cytosol, Ure2p fibrils appear to be packed
into very dense, high molecular weight particles (Fer-
nandez-Bellot et al., 2000) that may correspond to su-
pramolecular assemblies. This packing is either the
consequence of fibrils ageing in a particular physico-chemical microenvironment or due to the effect of pro-
teins associated to Ure2p fibrils. It is reasonable to
envisage that such a packing could be the consequence
of a conformational change within the fibrils. In prin-
ciple, Ure2p fibrils in such bundles must be well oriented
and therefore suitable for X-ray fiber diffraction mea-
surements. Partner proteins associated to the fibrils may
affect the intrinsic structure of the fibrils. Alternatively,they may produce an additional signal that superim-
poses to that of the fibrils X-ray diffraction patterns.
Thus, future efforts to isolate fibril bundles free of as-
sociated proteins for X-ray diffraction studies must be
attempted. It would also be very interesting to carry out
such studies on fibrils with associated proteins.
Acknowledgments
We gratefully acknowledge the expert help of Hassan
Belrhali on beamline ID14-EH1 in data collection and
Paul Dumas for his help in FTIR measurements. Wethank Dr. Solange Morera for allowing us to meet and
initiate our collaboration and Dr. Virginie Redeker and
Sheena Radford for helpful discussions. This work was
supported by the French Ministry of Research and
Technology, the Centre National de la Recherche Sci-
entifique and the Association pour la Recherche sur le
Cancer.
References
Astbury, W.T., Woods, H.J., 1933. Philos. Trans. A 232, 333–394.
Balbirnie, M., Grothe, R., Eisenberg, D.S., 2001. An amyloid-forming
peptide from the yeast prion Sup35 reveals a dehydrated b-sheetstructure for amyloid. Proc. Natl. Acad. Sci. USA 98, 2375–2380.
Blake, C., Serpell, L., 1996. Synchrotron X-ray studies suggest that the
core of the transthyretin amyloid fibril is a continuous b-sheethelix. Structure 4, 989–998.
Board, P.G., Coggan, M., Chelvanayagam, G., Easteal, S., Jermiin,
L.S., Schulte, G.K., Danley, D.E., Hoth, L.R., Griffor, M.C.,
Kamath, A.V., Rosner, M.H., Chrunyk, B.A., Perregaux, D.E.,
Gabel, C.A., Geoghegan, K.F., Pandit, J., 2000. Identification,
characterization, and crystal structure of the X class glutathione
transferases. J. Biol. Chem. 275, 24798–24806.
Bolton, D.C., McKinley, M.P., Prusiner, S.B., 1982. Identification of a
protein that purifies with the scrapie prion. Science 218, 1309–1311.
Bousset, L., Belrhali, H., Janin, J., Melki, R., Morera, S., 2001a.
Structure of the globular region of the prion protein Ure2 from the
yeast Saccharomyces cerevisiae. Structure (Camb) 9, 39–46.
Bousset, L., Belrhali, H., Melki, R., Morera, S., 2001b. Crystal
structures of the yeast prion Ure2p functional region in complex
with glutathione and related compounds. Biochemistry 40, 13564–
13573.
Bousset, L., Melki, R., 2002. Similar and divergent features in
mammalian and yeast prions. Microbes Infect. 4, 461–469.
Bousset, L., Thomson, N.H., Radford, S.E., Melki, R., 2002. The yeast
prion Ure2p retains its native alpha-helical conformation upon
assembly into protein fibrils in vitro. EMBO J. 21, 2903–2911.
L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142 141

Bradford, M.M., 1976. A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal. Biochem. 72, 248–254.
Carrell, R.W., Gooptu, B., 1998. Conformational changes and disease—
serpins, prions, andAlzheimer�s.Curr.Opin. Struct.Biol. 8, 799–809.
Cox, B.S., 1965. PSI, a cytoplasmic suppressor of super-suppressor in
yeast. Heredity 20, 505–521.
Derkatch, I.L., Bradley, M.E., Hong, J.Y., Liebman, S.W., 2001.
Prions affect the appearance of other prions: the story of [PIN(+)].
Cell 106, 171–182.
Dobson, C.M., 1999. Protein misfolding, evolution and disease.
Trends Biochem. Sci. 24, 329–332.
Edskes, H.K., Gray, V.T., Wickner, R.B., 1999. The [URE3] prion is
an aggregated form of Ure2p that can be cured by overexpression
of Ure2p fragments. Proc. Natl. Acad. Sci. USA 96, 1498–1503.
Fandrich, M., Fletcher, M.A., Dobson, C.M., 2001. Amyloid fibrils
from muscle myoglobin. Nature 410, 165–166.
Fernandez-Bellot, E., Guillemet, E., Cullin, C., 2000. The yeast prion
[URE3] can be greatly induced by a functional mutated URE2
allele. EMBO J. 19, 3215–3222.
Garson, J.C., Doucet, J., Leveque, J.L., Tsoucaris, G., 1991. Oriented
structure in human stratum corneum revealed by X-ray diffraction.
J. Invest. Dermatol. 96, 43–49.
Glover, J.R., Kowal, A.S., Schirmer, E.C., Patino, M.M., Liu, J.J.,
Lindquist, S., 1997. Self-seeded fibers formed by Sup35, the protein
determinant of [PSI+], a heritable prion-like factor of S. cerevisiae.
Cell 89, 811–819.
Jimenez, J.L., Guijarro, J.I., Orlova, E., Zurdo, J., Dobson, C.M.,
Sunde, M., Saibil, H.R., 1999. Cryo-electron microscopy structure
of an SH3 amyloid fibril and model of the molecular packing.
EMBO J. 18, 815–821.
Jimenez, J.L., Nettleton, E.J., Bouchard, M., Robinson, C.V., Dob-
son, C.M., Saibil, H.R., 2002. The protofilament structure of
insulin amyloid fibrils. Proc. Natl. Acad. Sci. USA 99, 9196–9201.
Kad, N.M., Thomson, N.H., Smith, D.P., Smith, D.A., Radford, S.E.,
2001. b(2)-microglobulin and its deamidated variant, N17D form
amyloid fibrils with a range of morphologies in vitro. J. Mol. Biol.
313, 559–571.
Khurana, R., Uversky, V.N., Neilsen, L., Fink, A.L., 2001. Is Congo
red an amyloid-specific dye. J. Biol. Chem. 276, 22715–22721.
Kirschner, D.A., Abraham, C., Selkoe, D.J., 1986. X-ray diffraction
from intraneuronal paired helical filaments and extraneuronal
amyloid fibers in Alzheimer disease indicates cross-b conformation.
Proc. Natl. Acad. Sci. USA 83, 503–507.
Krebs, M.R., Wilkins, D.K., Chung, E.W., Pitkeathly, M.C., Cham-
berlain, A.K., Zurdo, J., Robinson, C.V., Dobson, C.M., 2000.
Formation and seeding of amyloid fibrils from wild-type hen
lysozyme and a peptide fragment from the b-domain. J. Mol. Biol.
300, 541–549.
Lacroute, F., 1971. Non-Mendelian mutation allowing ureidosuccinic
acid uptake in yeast. J. Bacteriol. 106, 519–522.
Laemmli, U.K., 1970. Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227, 680–685.
Lim, A., Makhov, A.M., Bond, J., Inouye, H., Connors, L.H., Griffith,
J.D., Erickson, B.W., Kirschner, D.A., Costello, C.E., 2000.
Betabellins 15D and 16D, de novo designed b-sandwich proteins
that have amyloidogenic properties. J. Struct. Biol. 130, 363–370.
Magasanik, B., 1992. The Molecular and Cellular Biology of the Yeast
Saccharomyces cerevisiae. Cold Spring Harbor Laboratory Press,
Plainview, NY.
Masison, D.C., Wickner, R.B., 1995. Prion-inducing domain of yeast
Ure2p and protease resistance of Ure2p in prion-containing cells.
Science 270, 93–95.
McParland, V.J., Kad, N.M., Kalverda, A.P., Brown, A., Kirwin-
Jones, P., Hunter, M.G., Sunde, M., Radford, S.E., 2000. Partially
unfolded states of b(2)-microglobulin and amyloid formation in
vitro. Biochemistry 39, 8735–8746.
Morozova-Roche, L.A., Zurdo, J., Spencer, A., Noppe, W., Receveur,
V., Archer, D.B., Joniau, M., Dobson, C.M., 2000. Amyloid fibril
formation and seeding by wild-type human lysozyme and its
disease-related mutational variants. J. Struct. Biol. 130, 339–351.
Perutz, M.F., Pope, B.J., Owen, D., Wanker, E.E., Scherzinger, E.,
2002. Aggregation of proteins with expanded glutamine and
alanine repeats of the glutamine-rich and asparagine-rich domains
of Sup35 and of the amyloid beta-peptide of amyloid plaques. Proc.
Natl. Acad. Sci. USA 99, 5596–5600.
Polack, F., Mercier, R., Nahon, L., Armellin, C., Marx, J.P., Tanguy,
M., Couprie, M.E., P.D, 1999. Optical design and performances of
the IR microscope beamline at SuperACO. In: Carr, G.L., Dumas,
P. (Eds.), SPIE Proceedings (1999), Paris, France.
Santoso, A., Chien, P., Osherovich, L.Z., Weissman, J.S., 2000.
Molecular basis of a yeast prion species barrier. Cell 100, 277–288.
Schlumpberger, M., Prusiner, S.B., Herskowitz, I., 2001. Induction of
distinct [URE3] yeast prion strains. Mol. Cell. Biol. 21, 7035–7046.
Sondheimer, N., Lindquist, S., 2000. Rnq1: an epigenetic modifier of
protein function in yeast. Mol. Cell. 5, 163–172.
Stuart, B., 1997. Biological application of Infrared Spectroscopy.
Wiley, Sydney, Australia.
Sunde, M., Serpell, L.C., Bartlam, M., Fraser, P.E., Pepys, M.B.,
Blake, C.C., 1997. Common core structure of amyloid fibrils by
synchrotron X-ray diffraction. J. Mol. Biol. 273, 729–739.
Taylor, K.L., Cheng, N., Williams, R.W., Steven, A.C., Wickner,
R.B., 1999. Prion domain initiation of amyloid formation in vitro
from native Ure2p. Science 283, 1339–1343.
Thual, C., Bousset, L., Komar, A.A., Walter, S., Buchner, J., Cullin,
C., Melki, R., 2001. Stability, folding, dimerization, and assembly
properties of the yeast prion Ure2p. Biochemistry 40, 1764–1773.
Thual, C., Komar, A.A., Bousset, L., Fernandez-Bellot, E., Cullin, C.,
Melki, R., 1999. Structural characterization of Saccharomyces
cerevisiae prion-like protein Ure2. J. Biol. Chem. 274, 13666–
13674.
Umland, T.C., Taylor, K.L., Rhee, S., Wickner, R.B., Davies, D.R.,
2001. The crystal structure of the nitrogen regulation fragment of
the yeast prion protein Ure2p. Proc. Natl. Acad. Sci. USA 98,
1459–1464.
Uptain, S.M., Sawicki, G.J., Caughey, B., Lindquist, S., 2001. Strains
of [PSI(+)] are distinguished by their efficiencies of prion-mediated
conformational conversion. EMBO J. 20, 6236–6245.
Wickner, R.B., 1994. [URE3] as an altered URE2 protein: evidence for
a prion analog in Saccharomyces cerevisiae. Science 264, 566–569.
Wille, H., Michelitsch, M.D., Guenebaut, V., Supattapone, S., Serban,
A., Cohen, F.E., Agard, D.A., Prusiner, S.B., 2002. Structural
studies of the scrapie prion protein by electron crystallography.
Proc. Natl. Acad. Sci. USA 99, 3563–3568.
142 L. Bousset et al. / Journal of Structural Biology 141 (2003) 132–142