head kidneys in hatchlings of scoloplos armiger (annelida: orbiniida): implications for the...
TRANSCRIPT

J. Mar. BioI. Ass. U.K. (1998), 78, 183--192 Printed in Great Britain
183
HEAD KIDNEYS IN HATCHLINGS OF SCOLOPLOS ARMIGER
(ANNELIDA: ORBINIIDA): IMPLICATIONS FOR THE OCCURRENCE
OF PROTONEPHRIDIA IN LECITHOTROPHIC LARVAE
THOMAS BARTOLOMAEUS
n. Zoologisches Institut und Musewn der Universitat Gottingen, Berliner StraBe 28, D-7073 Gottingen, Germany
It is generally believed that lecithotrophic larvae of annelids do not possess functional excretory organs. However, as in certain annelids the planktotrophic trochophora larva has been secondarily modified into a lecithotrophic developmental stage and because protonephridia are characteristic for the trochophora, lecithotrophic developmental stages should also possess such organs. To test this assumption hatchlings of the orbiniidan Scoloplos armiger, which develops directly without a free-living larval stage, were investigated ultrastructurally. Each hatchling possesses a pair of protonephridia which lie caudal to the eyes and almost level with the frontal margin of the foregut. Each organ consists of three multiciliated cells, a terminal cell, a duct cell and a nephropore cell. The terminal cell bears a distally oriented hollow cytoplasmic cylinder, which surrounds the cilia. Adherens junctions connect this structure to the duct cell. Several clefts and pores perforate the wall of the hollow cylinder. Extracellular material covers the pores and clefts and thus may function as a molecular sieve during filtration. A comparison with the protonephridia of other annelid larvae reveals: (1) that one pair of protonephridial head kidneys consisting of a terminal cell, a duct cell and a nephropore cell must be assumed for the trochophore in the ground pattern of annelids and (2) that these organs are preserved when lecithotrophic larval stages evolved within the Annelida.
INTRODUCTION
In the annelid trochophore, the first pair of nephridia formed during development is located almost at the level of the larval mouth, more or less closely behind the eyes. Thus, atthis stage, the nephridia are located in that region of the larva which becomes highly modified during the formation of the head and the peristomial region. These nephridia were termed head kidneys (Kopfnieren) by Hatschek (1886). Hatschek himself believed that these organs open freely into the blastocoel, whereas subsequent classical investigations demonstrated that these organs are blindly closed. To date the conception that one pair of head kidneys are characteristic for the annelid trochophore and that these organs are protonephridia is widely accepted (Bartolomaeus & Ax, 1992; Korn, 1982 pp. 133, 159-161; Salvini-Plawen, 1980, 1988; Smith, 1992). This leads to the assumption that the trochophore of the annelid stem species already had such excretory organs.
On the other hand, it is generally believed that head kidneys are absent or at least non-functional in lecithotrophic annelid larvae (Anderson, 1973 p. 29; Heimler, 1988; Schroeder & Hermans, 1975; Smith, 1992). But, if protonephridia are characteristic for
author's copy

184 T. BARTOLOMAEUS
the trochophore, which is assumed to belong to the ground pattern of annelids, then these organs should also exist in lecithotrophic larvae. This assumption has recently been tested successfully for the modified, lecithotrophic trochophore of Pectinaria auricoma (O.F. Miiller, 1776) (Terebelllida): protonephridia were found in these larvae (Bartolomaeus, 1995). Scoloplos armiger (O.F. Miiller, 1776) develops directly and has no free swimming larval stage (Anderson, 1959). When the juvenile hatches from the cocoon, it has already 11 segments. Head kidneys have not been described in any of its developmental stages thus far. In order to test the above mentioned assumption on the existence of head kidneys in lecithotrophic larvae, they were investigated ultrastructurally. The structure of their protonephridia confirms the existing assumptions on the organization of protonephridial head kidneys in annelid larvae, which have been hypothezised to originally consist of three cells, Le. a terminal cell with a filter, a duct cell and a nephropore cell (Bartolomaeus & Ax, 1992).
MATERIALS AND METHODS
Cocoons of Scoloplos armiger were collected at low tide in the Wadden Sea of the Isle of Sylt. They were kept at 15°C in the laboratory until hatching of the juveniles. The animals were fixed in 2·5% glutaraldehyde buffered in 0·1 M sodium-cacodylate (pH 7·4) and containing ruthenium red at 4°C for 30 min, 12 h and 3 d after hatching. The animals were washed twice in the same buffer and postfixed in 1 % osmium tetroxide buffered in the same manner, then dehydrated in an acetone series and embedded in Araldite. Silver interference coloured sections (60-70nm) cut on a Reichert Ultracut E microtome with a diamond knife were kept on formvar-covered, single slot copper grids and stained with uranyl acetate and lead citrate in an LKB Ultrostainer. The anterior end of two animals of each stage were cross-sectioned. The protonephridia were reconstructed from a complete series of sections and examined using Zeiss EM 900 and EM 10B electron microscopes. When fixed for electron microscopy, the younger animal (12 h after hatching) had a body length of 1·1 mm and the older one (3 d after hatching) measured 1 ·3 flm in length.
OBSERVATIONS
The youngest animal investigated was fixed at a developmental stage which corresponds to that of the day 8 stage according to Anderson (1959), (Figure lA). The hatchling has one pair of head kidneys which lie in a ventrolateral position � 17 flm caudal to the eyes. Here, the protonephridia are situated among the cells of the body wall muscles and the epidermis. An extracellular matrix (ECM) partly surrounds the organs and separates the filtration area of the terminal cell from gut cells and epidermal cells (Figure 2A). Each protonephridium consists of three multiciliated cells: one terminal cell, one duct cell and one nephropore cell. The entire organ has a
length of almost 65 flm (Figure lB). The orientation of the longitudinal axis differs in the various sections of the organ: the terminal cell is orientated almost parallel to the
author's copy

HEAD KIDNEYS OF SCOLOPLOS HATCHLINGS 185
Figure 1. Scoloplos armiger, hatchling: (A) position of the head kidneys (arrow) in the animal; (B) schematical reconstruction of the protonephridiurn; (C) cross section of the filter; (D) sagittal section of the filter as redrawn from EM micrographs. DC, duct cell; NC, nephropore cell; TC, terminal celL
dorso-ventral axis of the animal, the course of the duct follows the fronto-caudal axis, and the nephropore cell finally pierces the epidermis parallel to the medio-Iateral axis of the animal.
The terminal cell rests partly on the ECM which surrounds the frontal edge of the foregut. Muscle cells surround the terminal cell, and the two types of cells are not separated by an ECM, except in the filtration area. The nucleus lies laterally to the longitudinal axis of the cell. Mitochondria are located in the proximity of the ciliary basal structures. In addition to these organells, the perikaryon contains vesicles of different size and large electron-dense stained lipid vesicles which are restricted to the vicinity of the nucleus. The terminal cell bears up to 25 cilia. The ciliary axoneme emanates from the basal body, which bears a small horizontal basal foot and a 1 ·5 ,urn long vertical and striated rootlet. Short microvilli are interspersed between the bases of the cilia. Additional microvilli surround the central bundle of cilia (Figure 1C,D). These microvilli emanate from the inner wall of a distally oriented hollow cytoplasmic cylinder, which originates from the the terminal. This cylinder is almost 7,um long and tightly surrounds the central bundle of cilia; its diameter of � 2,um diminishes
author's copy

186 T. BARTOLOMAEUS
towards the duct (Figure 2A). The diameter of the wall of the cylinder varies between 77 and 500 nm and is perforated by several clefts and pores. The arrangement of the clefts does not form any particular pattern. Nevertheless, they seem to form two distinct groups, which are separated by a poreless part of the cylinder. The clefts and pores are 20nm wide (N=25, SD=3·2nm). They are partly bridged by an electrondense material, which is presumed to represent the actual site of filtration in the protonephridium (Figure 2B,C). Thus, the perforated hollow cylinder represents the supporting structure for the filtration barrier, which consists of extracellular material. This structure will be referred to as the filter in the following. In the solid part of the filter filament and microtubules can be seen to extend towards the perikaryon (Figure 2A).
The distal edge of the cylinder is connected to the duct cell by adherens junctions. The duct is percellular, and the duct cell completely surrounds the ciliary bundle of the terminal cell. No microvilli can be found inside the duct, except in the most distal part the duct cell (Figure 3B). Here, eight short microvilli generally surround each cilium of the duct cell, and additional microvilli extend into the duct lumen. The cilia are attached to the cytoplasm of the duct cell in the same manner as in the terminal cell. In addition to the nucleus, the cytoplasm contains several mitochondria, numerous small vesicles and a few larger electron-dense stained lipid vesicles. Coated vesicles, which seem to indicate the existence of receptor mediated endocytosis, were only found in the most distal section of the duct. During its course, the duct cell penetrates the subepidermal ECM and runs between the bases of the epidermal cells for a distance of 17 f.1m.
The nephropore cell is bottle-shaped and lies completely intraepidermally. In addition to the nucleus, its cytoplasm contains vesicles, mitochondria and a very few large lipid vesicles. The nephropore cell rests on the subepidermal ECM. Therefore the distal section of the duct cell runs alongside the nephropore cell for a few micrometres (Figure 3B). The two cells are connected by desmosomes and adherens junctions. The adluminal cell surface bears several microvilli. The slender neck of this cell is 12 f.1m long, but only 1·5 f.1m thick. It extends laterally and its distal margin is connected to the adjacent epidermal cells by adherens and septate junctions. The pore lies 1 f.1m beneath the cuticle. The cilia are attached to the cytoplasm in the same manner as in the terminal cell. Most of the cilia originate from a lateral pouch. A few micrometres further, they merge with those of the duct cell. A few of them pass through the bottle-neck and extend to the exterior on the left side of the animal. On the right side, the cilia extend into a small subcuticular compartment, which does not directly communicate with the external medium.
No head kidneys were found in those stages which were fixed 3 d after hatching (corresponding a 10-d embryo according to Anderson (1959)). In those regions where these organs were to be expected several degenerating prototrochal cells sunk below the epidermal cell layer were found, but no organs or structures resembling functioning or degenerating protonephridia were observed.
author's copy

HEAD KIDNEYS OF SCOLOPLOS HATCHLINGS 187
Figure 2. Scalaplas armiger, hatchling: (A) sagittal section of the filter of the terminal cell (small arrows mark the extracellular matrix). Inset: solid part of the filter wall of the terminal cell with fibrils and microtubules (arows). (B) Pore in the wall of the filter, bridged by an electron-dense diaphragm (arrow). (C) Cross-section of the terminal cell (arrows mark filter clefts). EC, epidermal cell; DC, duct cell; Cc, gut cell; TC, terminal cell.
author's copy
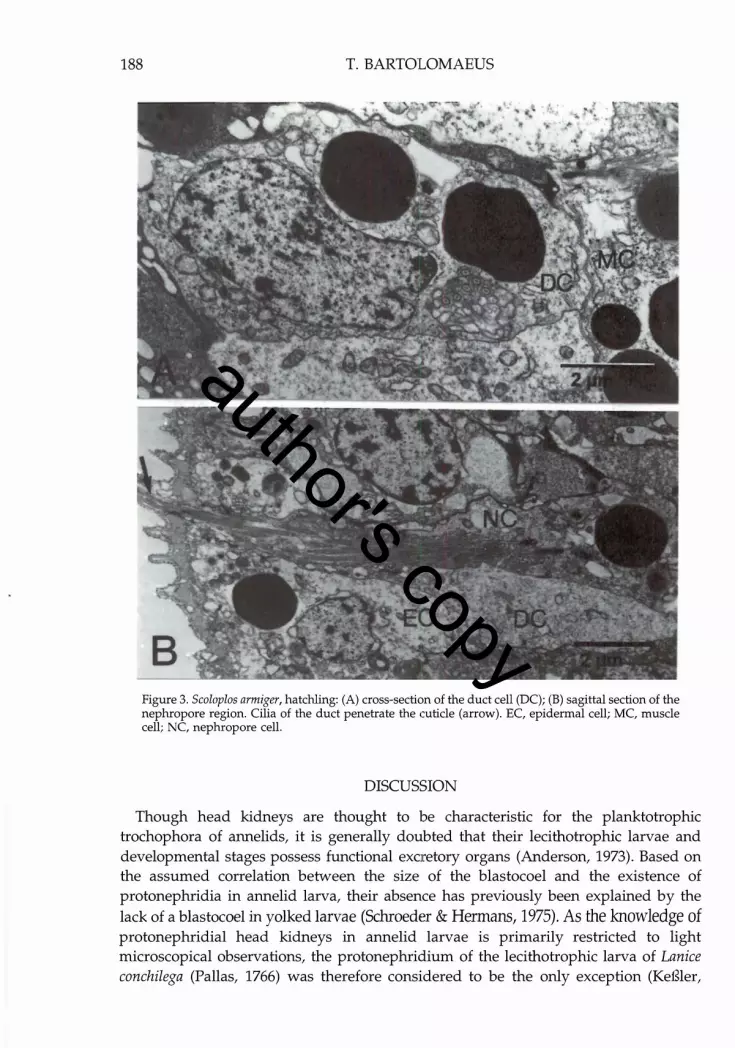
188 T. BARTOLOMAEUS
Figure 3. SeaIapIas armiger, hatchling: (A) cross-section of the duct cell (DC); (B) sagittal section of the nephropore region. Cilia of the duct penetrate the cuticle (arrow). EC, epidermal cell; MC, muscle cell; NC, nephropore cell.
DISCUSSION
Though head kidneys are thought to be characteristic for the planktotrophic trochophora of annelids, it is generally doubted that their lecithotrophic larvae and developmental stages possess functional excretory organs (Anderson, 1973). Based on the assumed correlation between the size of the blastocoel and the existence of protonephridia in annelid larva, their absence has previously been explained by the lack of a blastocoel in yolked larvae (Schroeder & Hermans, 1975). As the knowledge of protonephridial head kidneys in annelid larvae is primarily restricted to light microscopical observations, the protonephridium of the lecithotrophic larva of Lanice conchilega (pall as, 1766) was therefore considered to be the only exception (KeBler,
author's copy

HEAD KIDNEYS OF SCOLOPLOS HATCHLINGS 189
1963). Ultrastructurally, however, no filtration site could be detected in these protonephridia, so that the organs were regarded as non-functional (Heimler, 1981;
Smith, 1992). Nevertheless, such head kidneys have to be expected in those lecithotrophic larvae which have evolved from planktotrophic larvae and have been found in such derived larval forms (Bartolomaeus, 1995). According to the data in this paper such a retention also seems to be true for lecithotrophic developmental stages. This leads one to expect that protonephridial head kidneys should normally be present during annelid development, irrespective of the mode of development. Several records of larval protonephridia from light microscopy investigations of capsule larvae in certain oligochaetes and leeches support this assumption (Goodrich, 1945; Jamieson, 1992 p. 273; Fernandez et al., 1992 p. 356). The occurrence of these protonephridia can be interpreted as having been adopted from an ancestor which had a trochophore larva.
Compared to the large number of different annelid larvae, however, investigations into the ultrastructure of head kidneys are restricted to 14 annelid species (Bartolomaeus, 1995). With the exception of larval spionidans Pygospio elegans Claparede, 1863 and Polydora ciliata Gohnston, 1838), which both have metanephridial head kidneys (Schlotzer-Schrehardt, 1992; Bartolomaeus, in preparation), these organs are always protonephridia.
The protonephridial cells are multiciliated in Scoloplos armiger hatchlings and the ciliary flame is surrounded by several short microvilli. A comparable situation is known from sabellidan larvae, whereas other larvae have multiciliated terminal cells with rings of microvilli surrounding each cilium (Pemerl, 1965; Wessing & Polenz, 1974; Smith & Ruppert, 1988). The organization of the filter resembles that of the intersitital annelid Apodotrocha progenerans Westheide & Riser, 1983 (Westheide, 1985).
Based on the structural data from the known head kidneys, however, it seems impossible to obtain an indication of the phylogenetic position of orbiniidans. Nevertheless, the detection of a head kidney in the S. armiger leads to some considerations of the fate of protonephridia during evolution.
During the last decade, there has been a substantial accumulation of data from transmission electron microscopy of protonephridia. When these data were evaluated comparatively, a concept of the head kidney of the ancestral polychaete larva emerged (Bartolomaeus, 1993; Bartolomaeus & Ax, 1992; Smith, 1992). According to this concept, the head kidneys were protonephridia and composed of three monociliated cells (i.e. a terminal cell, a duct cell and a nephropore cell) in the ancestral trochophore. Such a protonephridium has been retained in the larva of Magelona mirabilis Johnston, 1865 (Bartolomaeus, 1995). The nephropore cell of this protonephridium lies intraepidermally and conducts the fluid to the exterior. The duct cells contain coated vesicles and lysosomes which both seem to indicate a modification of the ultrafiltrate flowing inside the duct. The terminal cell bears a hollow cytoplasmic cylinder which is attached to the duct cell by adhaerens junctions. The wall of this cylinder is perforated by pores which are bridged by extracellular material which is presumed to serve as a molecular sieve. This cylinder, which serves as supporting structure for the actual filtration barrier, has been called the filter. It surrounds the central cilium and its ring of elongated, strengthened microvilli. Except in certain phyllodocids, such a filter has been retained in all protonephridial terminal
author's copy

190 T. BARTOLOMAEUS
cells of annelid larvae, thus far investigated (Bartolomaeus, 1995). Within some phyllodocidan protonephridia the function of the filter has been adopted by the ring of microvilli which surround the cilium (Bartolomaeus, 1989; Smith & Ruppert, 1988).
A protonephridium consisting of these three cells has been found in larval Magelona mirabilis (Bartolomaeus, 1995), Chaetopterus variopedatus (Reinier, 1804) (BonchBruevich & Malakhov, 1986), Spirorbis spirorbis (Linne, 1758) (Bartolomaeus, in preparation) and now in the hatchling of the orbiniidan Scoloplos armiger. In contrast to M. mirabilis , these kidneys are entirely multiciliated, which is regarded as apomorphic. The plesiomorphic state of a monociliated and tricellular protonephridium is confirmed by the fact that such protonephridia have also been described in adult Gnathostomulida and Gastrotricha (Fischer, 1994; Lammert, 1985; Ruppert, 1991). Two competing hypotheses, i.e. the one presuming a small, acoelomate bilaterian ancestor (Ax, 1989, 1996, pp. 110-126) and the one which presumes a largebody bilaterian stem species with a larva (Rieger et al., 1989), explain these findings. If one accepts the hypothesis that the stem species of the Bilateria has been a small and compactly organized animal, it can be assumed that such a protonephridium must already have evolved in the stem lineage of the Bilateria, because protonephridia are the only functional excretory organs in a compact (acoelomate) organism (Ruppert & Smith, 1988). During the evolution of a biphasic life cycle the information for this excretory organ has been preserved in the early compact developmental stage, the larva (Bartolomaeus & Ax, 1992). Therefore, the primary monociliated and tricellular organization of the protonephridia can still be found in certain trochophorae or developmental stages of the annelids. Following the hypothesis of a large body stem species with a larva, it can be assumed that for functional reasons its larva must have had protonephridia. These organs would have been preserved in any larva of bilaterian with a large body and consequently also in the annelid larva. The data presented here, however, do not allow the rejection of one of the two hypotheses.
Both hypotheses, however, lead one to expect that there must be more annelid larvae or developmental stages which have retained the mentioned plesiomorphic design of protonephridia. Such protonephridia would, on the other hand, be unremarkable and hardly detectable under a light microscope, exspecially in lecithotrophic stages because of the high content of yolk granules in their cells: this could be another reason why there is only so little information about head kidneys in lecithotrophic larvae.
I would like to thank Professor Dr Peter Ax for some helpful comments on the manuscript and Miss Christina F6rster who kindly sectioned the animals. My thanks are also due to three unknown referees for their sincere and helpful comments. D.F. Whybrew kindly checked the English manuscript. This investigation was supported by the Akademie der Wissenschaften und der Literatur.
REFERENCES
Ax, P., 1989. Basis phylogenetic systematization of the Metazoa. In The hierarchy of life (ed. B. Fernholm et al.), pp. 229-245. Amsterdam: Elsevier.
Ax, P., 1996. Multicellular animals: a new approach to the phylogenetic order in nature, vol. 1. Berlin: Springer Verlag.
author's copy

HEAD KIDNEYS OF SCOLOPLOS HATCHLINGS 191
Anderson, D.T., 1959. The embryology of the polychaete Scoloplos armiger. Quarterly Journal of microscopical Sciences, 100, 89-166.
Anderson, D.T., 1973. Embryology and phylogeny in annelids and arthropods. Oxford: Pergamon Press.
Bartolomaeus, T., 1989. Ultrastructure and development of the nephridia in Anaitides mucosa (Annelida, Polychaeta). Zoomorphology, 109, 15-32.
Bartolomaeus, T., 1993. Ultrastructure of the protonephridia in the larva of Autolytus prolifer (Annelida, Syllidae): implications for annelid phylogeny. Microfauna Marina, 8, 55-64.
Bartolomaeus, T., 1995. Ultrastructure of the protonephridia in larval Magelona mirabilis (Spionida) and Pectinaria auricoma (Terebellida): head kidneys in the ground pattern of the Annelida. Microfauna Marina, 10, 117-141.
Bartolomaeus, T. & Ax, P., 1992. Protonephridia and Metanephridia - their relation within the Bilateria. Zeitschrift Jiir zoologische Systematik und Evolutionsforschung, 30, 21-45.
Bonch-Bruevich, E.Y. & Malakhov, V.Y., 1986. Organization of the early larva of the polychaete Chaetopterus variopedatus (Spiomorpha, Chaetopteridae). Doklady Akademii Nauk, SSSR, 292, 1013-1015.
Fernandez, J., TeIlez, V. & Olea, N., 1992. Hirudinea. In Microscopic anatomy of invertebrates Vol. 7. Annelida (ed. F.W. Harrison and S.L. Gardiner), pp. 323-394. New York: Wiley-Liss, Ine.
Fischer, u., 1994. Additional aspects to the protonephridial system of Dactylopodola baltica (Gastrotricha, Macrodasyida). Microfauna Marina, 9, 285-289.
Goodrich, E.5., 1945. The study of genital ducts and nephridia since 1895. Quarterly Journal of Microscopical Sciences, 86, 13-329.
Hatschek, B., 1886. Zur Entwicklung des Kopfes von Polygordius. Arbeiten aus dem Zoologischen Institut der Universitct"t Wien und der Zoologischen Station Triest, 6, 109-210.
Heimler, W., 1981. Untersuchungen zur Larvalentwicklung von Lanice conchilega (pallas, 1766) (Polychaeta, Terebellomorpha). H. Bau und Ultrastruktur der Trochophora-Larve. Zoologische Jahrbiicher, Abteilung Anatomie und Ontogenie, 106, 236-277.
Heimler, W., 1988. Larvae. In The ultrastructure of the Polychaeta (ed. W. Westheide and e.o. Hermans), pp. 353-371. Stuttgart: Gustav Fischer. [Microfauna Marina, 4.]
Jamieson, B.G.M., 1992. Oligochaeta. In Microscopic anatomy of invertebrates, vol. 7 (ed. F.W. Harrison and S.L. Gardiner), pp. 217-322. New York: Wiley-Liss, Inc.
KeBler, M., 1963. Die Entwicklung von Lanice conchilega (pallas) mit besonderer Beriicksichtigung der Lebensweise. Helgolil"nder wissenschaftliche Meeresuntersuchungen, 8, 425-476.
Korn, H., 1982. Annelida. In Morphogenese der Tiere, Lief. 5 H-I (series ed. F. Seidel), pp. 1-599. Jena: G. Fischer.
Lammert, V., 1985. The fine structure of the protonephridia in Gnathostomulida and their comparison within the Bilateria. Zoomorphology, 105, 308-316.
Pemerl, J., 1965. Ultrastructure of the protonephridium of the trochophore larva of Serpula vermicularis (Annelida, Polychaeta). American Zoologist, S, 666-667.
Rieger, R.M., Haszprunar, G. & Schuchert, P. 1989. On the origin of the Bilateria: traditional views and recent alternative concepts. In The early evolution of Metazoa and the significance of problematic taxa (ed. A.M. Simonetta and s.c. Morris), pp. 107-112. Cambridge University Press.
Ruppert, E.E. & Smith, P.R., 1988. The functional organization of filtration nephridia. Biological Reviews, 171, 231-258.
Ruppert, E.E., 1991. Gastrotricha. In Microscopic anatomy of invertebrates. Vol. 4. Aschelminthes (ed. F.W. Harrsion and E.E. Ruppert), pp. 41-100. New York: Wiley-Liss, Ine.
Salvini-Plawen, L. von, 1980. Was ist eine Trochophora? Eine Analyse der Larventypen mariner Protostomier. Zoologische Jahrbiicher, Abteilung Anatomie und Ontogenie, 103, 389-423.
Salvini-Plawen, L. von, 1988. Annelida and Mollusca - a prospectus. In The ultrastructure of the Polychaeta (ed. W. Westheide and e.O. Hermans), pp. 383-396. Stuttgart: Gustav Fischer. [Micro fauna Marina, 4.]
Schlotzer-Schrehardt, u., 1992. Ultrastrukutrelle Untersuchungen zur Reproduktion und Postembryonalentwicklung einschliefllich Adultorganisation von Pygospio elegans Claparede, 1863. PhD thesis, University Erlangen-Niirnberg, Germany.
author's copy

192 T. BARTOLOMAEUS
Schroeder, P.C & Hermans, CO., 1975. Annelida: Polychaeta. In Reproduction of marine invertebrates. Vol. 3. Annelids and echiurans (ed. A.C Giese and J.S. Pearse), pp. 1-213. New York: Academic Press.
Smith, P.R., 1992. Polychaeta: excretory system. In: Microscopic anatomy of invertebrates. Vol. 7. Annelida (ed. F.W. Harrison and S.L. Gardiner), pp. 71-108. New York: Wiley-Liss, Inc.
Smith, P.R. & Ruppert, E.E., 1988. Nephridia. In The ultrastructure of the Polychaeta (ed. W. Westheide and CO. Hermans), pp. 231-262. Stuttgart: Gustav Fischer. [Microfauna Marina, 4.]
Wessing, A. & Polenz, A., 1974. Structure, development and function of the protonephridium in the trochophore of Pomatoceros triqueter (Annelida, Polychaeta, Sedentaria). Cell and Tissue Research, 156, 21-33.
Westheide, W., 1985. Ultrastructure of the protonephridia in the dorvilleid polychaete Apodotrocha progenerans (Annelida). Zoologica Scripta, 14, 273-278.
Submitted 13 February 1996. Accepted 12 September 1996.
author's copy