generalized tetracycline induced cre recombinase expression through the rosa26 locus of recombinant...
TRANSCRIPT

GR
CBa
b
a
ARRA
KCRCTK
1
iHarsNacft
oT
0d
Journal of Neuroscience Methods 176 (2009) 16–23
Contents lists available at ScienceDirect
Journal of Neuroscience Methods
journa l homepage: www.e lsev ier .com/ locate / jneumeth
eneralized tetracycline induced Cre recombinase expression through theOSA26 locus of recombinant mice
ristina M. Bäckmana,∗, YaJun Zhanga, Nasir Malika, Lufei Shana,arry J. Hoffera, Heiner Westphalb, Andreas C. Tomaca
Cellular Neurobiology Branch, National Institute on Drug Abuse, National Institutes of Health, Baltimore, MD 21224, USALaboratory of Mammalian Genes and Development, National Institute of Child Health and Human Development, Bethesda, MD 20892, USA
r t i c l e i n f o
rticle history:eceived 9 May 2008eceived in revised form 14 August 2008ccepted 16 August 2008
eywords:re–loxPOSA26onditionaletracyclinenockin
a b s t r a c t
Inducible Cre recombinase systems have been developed to bypass initial lethal phenotypes and to pro-vide access to later embryonic or adult phenotypes. Here we describe the generation of a recombinantmouse that combines a tetracycline dependent switch with generalized Cre recombinase expression bytargeting the ubiquitously expressed ROSA26 locus. This transgenic strain was developed using a simpli-fied gene delivery system integrating both elements, the reverse tetracycline controlled trans-activator(rtTA) and rtTA inducible promoter into a single vector. In this transgenic strain, the endogenous ROSA26promoter drives rtTA expression through a splice acceptor site. The tetracycline inducible promoter, clonedin opposite orientation to the ROSA26 locus and separated from the rtTA element by a 5 kb human p53intron, drives Cre recombinase expression. Crossing these mice with a Cre reporter strain showed that CreDNA-mediated recombination was ubiquitously and effectively induced during various prenatal devel-opmental windows. Background Cre recombinase expression levels were observed in some tissues in theabsence of the inducer, mostly during late embryonic developmental stages and in adult animals. Back-ground recombination levels were low during development and most prominent in nervous tissue. Crerecombinase expression could not be effectively induced in adult animals. While rtTA mRNA levels werehigh in developmental and adult tissues, Cre recombinase mRNA levels remained low after doxycycline
treatment. The mouse strain described here provides a valuable tool to further analyze the function ofgenes during specific developmental windows, by allowing the effective inactivation of their functionof em
wit
gbiri
throughout defined stages
. Introduction
Exploration of gene function in vivo has been markedly facil-tated by the use of transgenic and gene targeting technology.owever, there are clear limitations that restrict the utility of thispproach. In many cases, transgene expression or gene inactivationesults in serious developmental defects or embryonic lethality,o that only the earliest phenotype can be explored (Lobe andagy, 1998; Marth, 1996; Rajewsky et al., 1996). In addition, the
bsence of a particular gene during development often leads toompensatory mechanisms thereby obscuring interpretation of itsunction. Therefore, the ability to effectively induce genetic muta-ions during specific developmental windows and in adult animals∗ Corresponding author at: National Institute on Drug Abuse, Cellular Neurobiol-gy Section, 5500 Nathan Shock Drive, Baltimore, MD 21224, USA.el.: +1 443 740 2744; fax: +1 443 740 2123.
E-mail address: [email protected] (C.M. Bäckman).
itroiamtrr
165-0270/$ – see front matter. Published by Elsevier B.V.oi:10.1016/j.jneumeth.2008.08.024
bryonic development.Published by Elsevier B.V.
ould be advantageous, as this approach eliminates many of thenherent problems associated with the generation of conventionalransgenic mutant mice.
Spatially and temporally regulated genetic mutations can beenerated in mice by using an inducible recombination systemased on Cre recombinase, a site-specific recombinase in the
ntegrase family. The Cre–loxP system uses P1 bacteriophage Creecombinase to catalyze the excision of DNA located between flank-ng loxP sites, and it has been widely used since its first applicationn animal models (Nagy, 2000). No recombination occurs betweenhe loxP sites until Cre enzyme is introduced, thus the alterations areeferred to as conditional alterations. For conditional gene knock-uts, loxP sites are placed on either side of a critical exon(s); thusntroduction of Cre recombinase leads to the excision of the exon(s)
nd effectively a gene knockout. This strategy allows the develop-ent of mouse lines with silent genetic mutations (loxP flanked)hat can be activated by Cre mediated excision. Cre mediatedecombination can be introduced by crossing the mouse line car-ying the silent mutation with a Cre transgenic mouse. Depending

eurosc
oaabnt
cetsesmtmhitgoTrAfleoHiamlewa
slotkRbttalismnw
2
2
pXC12tT
ahbictFGResiatspTC3Tst
2
ibHsRnRsaCf(Affagapb(R
2
Rd(tRtRp
C.M. Bäckman et al. / Journal of N
n the expression pattern of the Cre transgene, the genomic alter-tions can be tailored in a general or tissue specific manner. Thispproach allows the establishment of lines that would otherwisee embryonic lethal. Unfortunately, introduction of Cre recombi-ase in this manner does not allow temporal control apart fromhe onset of Cre expression.
To create temporally controlled somatic mutations in the mouse,hemically inducible forms of Cre have been developed (Albaneset al., 2002; Garcia and Mills, 2002; Mills, 2001). At present,he reversed tetracycline-controlled transcriptional activator (rtTA)ystem, which activates the responsive elements only in the pres-nce of tetracycline (Gossen and Bujard, 1992), has proven to beuccessful for the generation of temporally controlled transgenicodels (Mansuy and Bujard, 2000; Morozov et al., 2003). Since
he first tetracycline-inducible gene expression system was imple-ented nearly a decade ago, several modifications of this approach
ave been reported to improve the control of gene expressionn vivo. However, currently available Cre recombinase linked toetracycline regulated transgenic methods require most often theeneration of two transgenic strains, one carrying the transgenef interest, in this case Cre recombinase, under the control of theet Operator (a tetracycline sensitive promoter), and the other theeverse transactivator TA (rtTA) transgene (Ryding et al., 2001).
third line containing the silent mutation or gene of interestanked by loxP sites is necessary. Crossing these three lines gen-rates progeny with the necessary transgenes to allow regulationf Cre recombinase upon administration of the inducer tetracycline.owever, crossing and analysis of animals transgenic for the three
ndividual components of the system is costly and time consumingnd genetic segregation of the control elements during breedingay also be an issue. Therefore, the generation of a single transgenic
ine containing all the necessary elements to allow the induciblexpression of Cre recombinase would provide a simplified frame-ork for the generation of conditional transgenic and knockout
nimal models.To improve the usefulness of the tetracycline regulated Cre–loxP
ystem we have generated a transgenic system that combines twoayers of regulation in the same transgenic line provided by the usef an endogenous cell promoter and treatment with doxycyclineo induce Cre recombinase expression. We have developed a novelnockin mouse strain that utilizes the endogenous and ubiquitousOSA26 promoter to drive the ubiquitous expression of Cre recom-inase in an inducible manner. We targeted the ROSA26 locus intohe first intron with a targeting vector containing (1) a splice accep-or (SA) followed by the newly developed transactivator rtTA-M2,nd (2) a tetracycline responsive minimal promoter (TRE-tight) fol-owed by the Cre recombinase sequence. To minimize any potentialnterference between both elements, they were placed in oppo-ite directions and spaced by a human p53 intron (5 kb). This novelouse strain was developed to allow regulation of gene recombi-
ation in a generalized and inducible manner after a single crossith mice containing a silent loxP mutation.
. Material and methods
.1. Generation of R26rtTA-TRECre knockin mice
A slightly modified backbone targeting vector (derived fromROSA26-1) was generated by insertion of a PacI linker into thehoI creating plasmid pROSA26-1.PacI. The tetracycline controlled
re cassette was released from plasmid pSA/M2alt/P53/CMV*-/nlsCRE/FRT/PGK-neo/FRT (for cloning details see Bäckman et al.,004) by PacI digestion and cloned into the unique XbaI site ofhe pROSA26-1.PacI vector, creating targeting plasmid pR26Cki.he vector was linearized and electroporated into R1 ES cellsRAtFf
ience Methods 176 (2009) 16–23 17
s previously described (Fig. 1A, Tomac et al., 2000). Desiredomologous recombination events were confirmed by Southernlot hybridization (Fig. 1B) with a probe external to the target-
ng vector. Targeted ES cells were injected into blastocysts of57bl/6 mice to generate chimeric mice. To eliminate the nega-ive selection cassette (Fig. 1A), germline mice were crossed withlpe deleter mice generated in 129 background (129S4/SvJaeSor-t(ROSA)26Sortm1(FLP1)Dym/J; Jackson Laboratories, Bar Harbor, ME).emoval of the neomycin cassette was confirmed in several gen-rations by PCR amplification with previously tested primerspanning from the p53 intron into the neomycin cassette. Genotyp-ng of mice carrying the mutation was performed by PCR analysiss follows below. Primers and fluorogenic Taqman probes forhe 5′ nuclease assay were designed with the Beacon Designeroftware (Premier Biosoft, Palo Alto, CA) for amplification androbe binding in the wild-type (5′-CCCTCTTCCCTCGTGATCTG-3′ 5′-CCCTCCAATTTTACACCTGTTC -3′ Taqman-CAGGACAACGCCCACA-ACCAGG) and the targeted alleles (5′-CTGTGTCAGCAAGGCTTCTC-′ 5′-TGTCTCTCTTTCCTCTTTTGCTAC-3′ Taqman-CCTGTTCCTCCAA-ACGCAGCCCA) in a multiplex assay. The resulting FAM and VICignals corresponded to the wild type and targeted alleles, respec-ively.
.2. Histology and Xgal staining
We tested the activity of the R26rtTA-TRECre transgene by cross-ng the mice to the ROSA26XneorXlacZ reporter mouse developedy Soriano, 1999 (129S-Gt(ROSA)26Sor/J; Jackson Laboratories, Bararbor, ME), which carry a sequence encoding a loxP flanked neor
equence blocking the transcription of the downstream lacZ gene.emoval of the neor gene sequence by Cre mediated recombi-ation leads to expression of b-galactosidase. Single transgenic26rtTA-TRECre mice were crossed to ROSA26XneorXlacZ reportertrain to define efficiency of recombination, as well as backgroundctivity. To determine the time course and efficiency of induciblere excision during different developmental windows, pregnant
emales were provided with food pellets containing doxycycline12 g/kg) during the following periods: E9–E11, E13–E15, E16–PD1.dult animals were treated with the same dose of doxycycline
or 7 consecutive days before analysis. Control animals receivedood ad libitum. Adult and pregnant mice were deeply anesthetizednd embryos and tissues were thereafter collected following NIHuidelines, and fresh frozen. Cryostat sections from embryos anddult tissues (30 mm) were mounted on slides and fixed with 4%araformaldehyde for 10 min. Xgal staining of tissue sections for-galactosidase detection was carried out as previously describedLobe et al., 1999). Sections were counterstained with Nuclear Fasted for better visualization of cellular elements.
.3. RNA isolation and real time quantitative PCR
Combination of Taqman probes (Universal Probe Library,oche, Indianapolis, IN) and primer pairs (IDT, CoralVille, IA) wereesigned using the Universal Probe Libray Design Center softwarehttps://www.roche-applied-science.com/sis/rtpcr/upl/index.jsp)o quantify levels of (1) Cre recombinase and rtTA mRNA. TotalNA was extracted as follows. Whole embryos (E11) or specificissues (PD1, adult) were dissected and immediately used forNA isolation. Tissue samples were homogenized (Ultrasonicrocessor, Cole-Parmer, Vermon Hills, IL) in lysis buffer. Total
NA was extracted using the RNAqueous Ambion kit (Ambion,ustin, TX), following manufacturers instructions. Total RNA wasreated with DNAse I to eliminate genomic DNA contamination.or cDNA synthesis, 2 �g total RNA was mixed following manu-acturers instructions for the Superscript III reverse transcriptase
18 C.M. Bäckman et al. / Journal of Neuroscience Methods 176 (2009) 16–23
Fig. 1. Generation of R26rtTA-TRECre-Neo knockin mice by homologous recombination. (A) Maps showing the endogenous ROSA26 locus (ROSA26wt) and targeted variants(R26rtTA-TRECre-Neo and R26rtTA-TRECre). The transgene was targeted into the first intron of the ROSA26 locus, at a unique XbaI site (X). Our targeting vector contained a splicea r TREF sses wa ith Ect
kcRFTda6mvepet
2
ecAob
3
3a
fcseueaberbE
e(awIt
Xdrahetitcwl
RtdswtglfambpdweinXd
3
ltaraen
cceptor (SA) followed by the rtTA transgene. The tetracycline sensitive promoter oRT-flanked neomycin selection cassette (Neo) was deleted in germline mice by crore shown (EV; EcoRV). (B) Southern blot hybridization of ES-cell DNA digested wargeted allele gives a 3.0-kb fragment.
it (Invitrogen, Carlsbad, CA). Distilled water was added to theDNA mixture to a final volume of 80 �l. Real time quantitativeT-PCR (qRT-PCR) analyses were conducted on the ABI HT7900ast Real time PCR system. All PCR samples were run as duplicates.he reproducibility of results was determined by inspection ofuplicate samples. After an initial incubation step for 10 mint 95 C, qRT-PCR was carried out using 40 cycles (95 C for 15 s,0 C for 60 s). The standard curve method was used to compareRNA expression levels between groups. To account for sample
ariability, the mRNA expression levels were normalized to thexpression levels of the housekeeping genes: (1) b-actin, and (2)hosphoglycerate kinase 1 (PGK). Since normalization to bothndogenous control genes led to similar results, we present onlyhe data normalized to PGK expression.
.4. Statistical analysis
The relative amounts of Cre and rtTA mRNA were calculated forach sample by expressing the corresponding mRNA levels as a per-entage of PGK and b-actin contained in each analyzed sample.ll values are presented as mean ± S.E.M. Statistical comparisonsf data among the groups were analyzed by ANOVA; additionaletween-group comparisons were made using Fisher post hoc tests.
. Results
.1. Doxycycline-dependent Cre excision in embryos and adultnimals
To study Cre mediated recombination, embryos heterozygousor both the R26rtTA-TRECre and ROSA26XneorXlacZ transgenes wereollected at various stages between E11 and postnatal day 1 andtained with Xgal to visualize LacZ activity. As seen in Fig. 2,mbryos treated with doxycycline from E9 to E11 displayed ubiq-itous and very effective DNA recombination. The bitransgenicmbryos were dark blue and Xgal staining was evenly detectedlong the entire embryonic axis. In the absence of the inducer,-galactosidase was present in a few scattered cells along thembryonic axis, indicating tight regulation of Cre induction. Theseesults indicate inducible and effective Cre mediated DNA recom-ination during this early developmental window (Fig. 2B, C, D and).
To analyze the degree of recombination during later stages,mbryos were exposed to doxycycline from E13 to E15 and E16–PD1
Fig. 3). E15 embryos and postnatal day 1 mice displayed gener-lized DNA recombination. However, during these developmentalindows we observed a shift towards partial DNA recombination.ncomplete or partial recombination was most marked in the cen-ral nervous system (CNS). While ubiquitous and quite extensive
(irdp
(C*) was cloned in opposite orientation and separated by a human p53 intron. Theith an Flp-deleter line, thereby generating R26rtTA-TRECre . Relevant restriction sites
oRV and using a 5′ probe. The wild-type allele gives a 11.0-kb fragment while the
gal staining could be detected in peripheral organs during bothevelopmental stages, the brain and spinal cord displayed DNAecombination in a segregated fashion (see Fig. 3). Also, in thebsence of the inducer, the number of Xgal positive cells was clearlyigher at E16 and PD1, as compared to E11 (data not shown). How-ver, the presence of leaking cells was absent in most embryonicissues and positive cells were still few and scattered in the brain,ndicating a controlled regulation of Cre induction. These qualita-ive results indicate that the R26rtTA-TRECre deleter line promotesonditional recombination during these developmental windows,ith maximal recombination efficiency outside the CNS and with
ow levels of leaking activity.Adult animals heterozygous for both, the R26rtTA-TRECre and
OSA26XneorXlacZ transgenes were treated with food pellets con-aining doxycycline (12 gr/kg) for 2 weeks. As a negative control,ouble transgenic animals were treated with food ad libitum. Tis-ue samples corresponding to brain, heart, liver, lung and kidneyere taken 1 week after treatment and stained to detect beta galac-
osidase activity. Treatment with the inducer did not induce betaalactosidase expression in bitransgenic animals. Xgal expressionevels remained low in both experimental and control animals. Aew scattered Xgal positive cells could be observed in kidney, lungnd liver, indicating low leaking activity. Qualitative analyses deter-ined that leaking of Cre recombinant expression was highest in
rain tissue, as compared to other organs analyzed (Fig. 4). Xgalositive cells could be observed in several brain regions, in bothoxycycline treated and untreated brains. These data correlate wellith Cre recombinase leaking activity observed during different
mbryonic stages, with the highest background levels also observedn brain and spinal cord. The following data suggest Cre recombi-ase expression is not inducible in adult animals, as generalizedgal activity cannot be induced and detected after treatment withoxycycline.
.2. rtTA and Cre mRNA expression in embryos and adult animals
In order to investigate the effectiveness of the dual cassette, uti-ized in this study, to induce Cre recombinase expression throughhe ROSA26 locus, E11 embryos and selected organs from PD1 anddult animals were collected to quantify rtTA and Cre mRNA levels.tTA mRNA was highly expressed in E11 embryos, and heart, brainnd liver from PD1 and adult animals (Fig. 5). rtTA levels were high-st in E11 embryos and PD1 brain tissue. rtTA mRNA levels wereormalized against the housekeeping gene phosphoglycerokinase
PGK), a medium copy-expressed gene encoding for an enzymenvolved in glycolysis. These data suggest that the R26rtTA-TRECreecombinant mice can effectively drive rtTA expression duringevelopment and in adult animals through the endogenous ROSA26romoter.
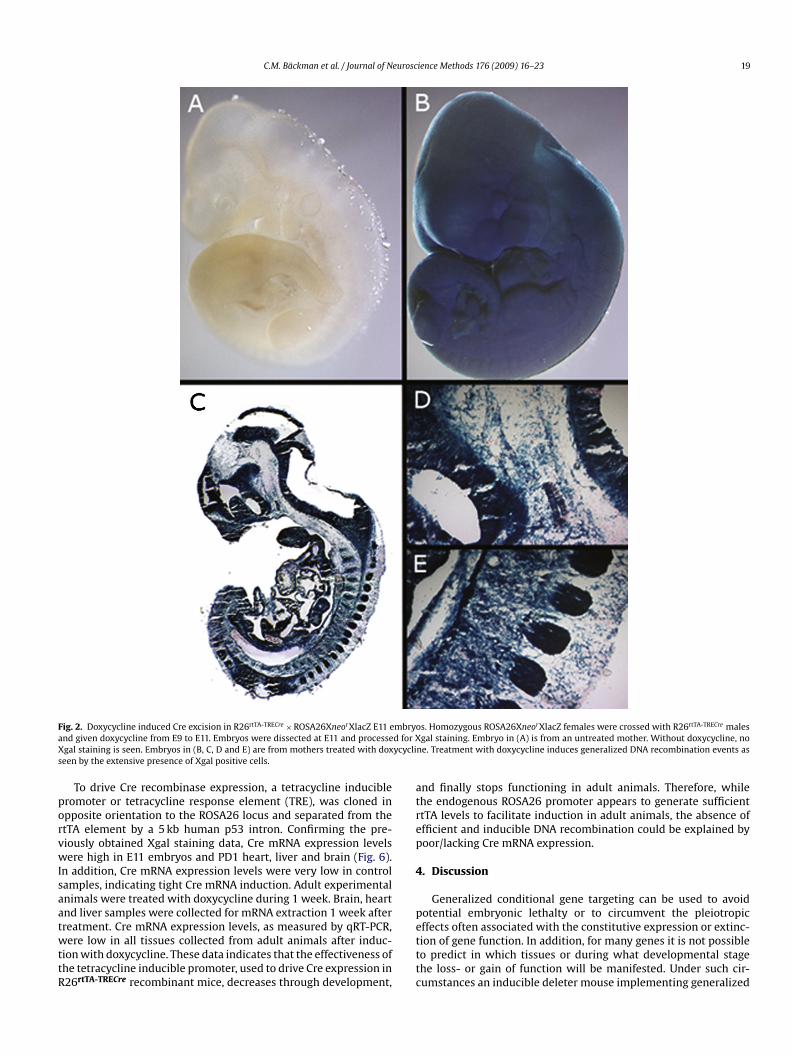
C.M. Bäckman et al. / Journal of Neuroscience Methods 176 (2009) 16–23 19
F mbryoa d for XX ycyclis
porvwIsaatwttR
atrep
4
p
ig. 2. Doxycycline induced Cre excision in R26rtTA-TRECre × ROSA26XneorXlacZ E11 end given doxycycline from E9 to E11. Embryos were dissected at E11 and processegal staining is seen. Embryos in (B, C, D and E) are from mothers treated with doxeen by the extensive presence of Xgal positive cells.
To drive Cre recombinase expression, a tetracycline inducibleromoter or tetracycline response element (TRE), was cloned inpposite orientation to the ROSA26 locus and separated from thetTA element by a 5 kb human p53 intron. Confirming the pre-iously obtained Xgal staining data, Cre mRNA expression levelsere high in E11 embryos and PD1 heart, liver and brain (Fig. 6).
n addition, Cre mRNA expression levels were very low in controlamples, indicating tight Cre mRNA induction. Adult experimentalnimals were treated with doxycycline during 1 week. Brain, heartnd liver samples were collected for mRNA extraction 1 week after
reatment. Cre mRNA expression levels, as measured by qRT-PCR,ere low in all tissues collected from adult animals after induc-ion with doxycycline. These data indicates that the effectiveness ofhe tetracycline inducible promoter, used to drive Cre expression in26rtTA-TRECre recombinant mice, decreases through development,
etttc
s. Homozygous ROSA26XneorXlacZ females were crossed with R26rtTA-TRECre malesgal staining. Embryo in (A) is from an untreated mother. Without doxycycline, no
ne. Treatment with doxycycline induces generalized DNA recombination events as
nd finally stops functioning in adult animals. Therefore, whilehe endogenous ROSA26 promoter appears to generate sufficienttTA levels to facilitate induction in adult animals, the absence offficient and inducible DNA recombination could be explained byoor/lacking Cre mRNA expression.
. Discussion
Generalized conditional gene targeting can be used to avoidotential embryonic lethalty or to circumvent the pleiotropic
ffects often associated with the constitutive expression or extinc-ion of gene function. In addition, for many genes it is not possibleo predict in which tissues or during what developmental stagehe loss- or gain of function will be manifested. Under such cir-umstances an inducible deleter mouse implementing generalized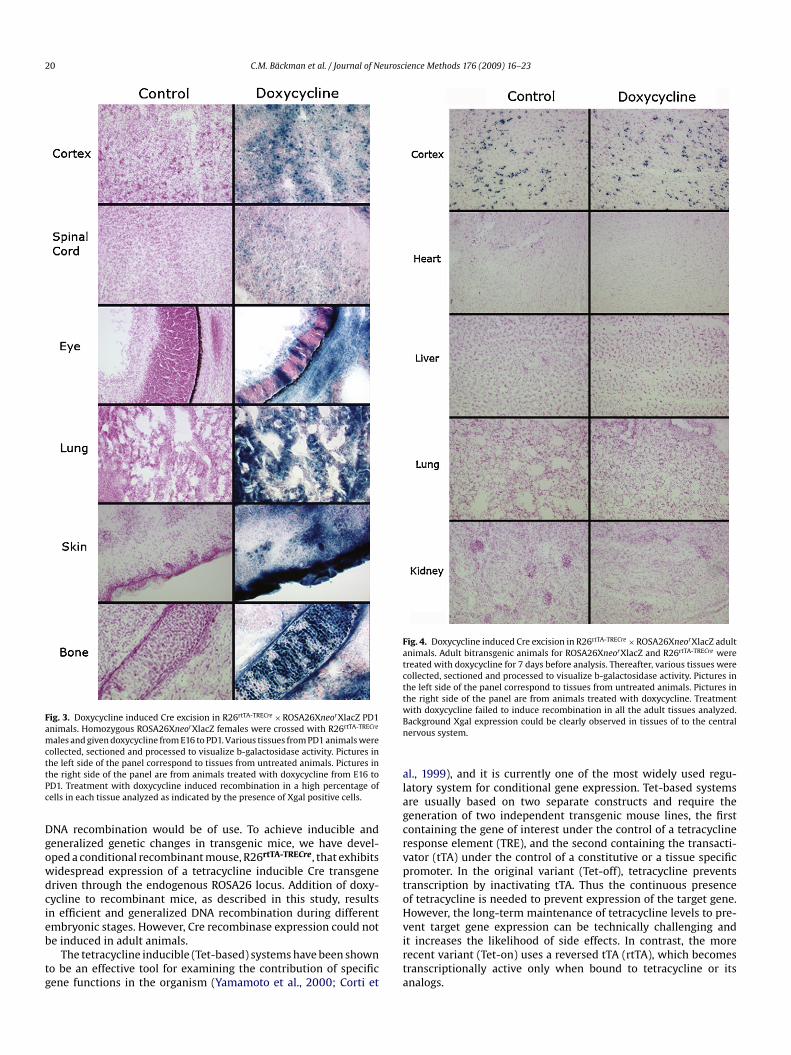
20 C.M. Bäckman et al. / Journal of Neuroscience Methods 176 (2009) 16–23
Fig. 3. Doxycycline induced Cre excision in R26rtTA-TRECre × ROSA26XneorXlacZ PD1animals. Homozygous ROSA26XneorXlacZ females were crossed with R26rtTA-TRECre
males and given doxycycline from E16 to PD1. Various tissues from PD1 animals werecollected, sectioned and processed to visualize b-galactosidase activity. Pictures inttPc
Dgowdcieb
tg
Fig. 4. Doxycycline induced Cre excision in R26rtTA-TRECre × ROSA26XneorXlacZ adultanimals. Adult bitransgenic animals for ROSA26XneorXlacZ and R26rtTA-TRECre weretreated with doxycycline for 7 days before analysis. Thereafter, various tissues werecollected, sectioned and processed to visualize b-galactosidase activity. Pictures inthe left side of the panel correspond to tissues from untreated animals. Pictures inthe right side of the panel are from animals treated with doxycycline. TreatmentwBn
alagcrvptoH
he left side of the panel correspond to tissues from untreated animals. Pictures inhe right side of the panel are from animals treated with doxycycline from E16 toD1. Treatment with doxycycline induced recombination in a high percentage ofells in each tissue analyzed as indicated by the presence of Xgal positive cells.
NA recombination would be of use. To achieve inducible andeneralized genetic changes in transgenic mice, we have devel-ped a conditional recombinant mouse, R26rtTA-TRECre, that exhibitsidespread expression of a tetracycline inducible Cre transgeneriven through the endogenous ROSA26 locus. Addition of doxy-ycline to recombinant mice, as described in this study, resultsn efficient and generalized DNA recombination during different
mbryonic stages. However, Cre recombinase expression could note induced in adult animals.The tetracycline inducible (Tet-based) systems have been showno be an effective tool for examining the contribution of specificene functions in the organism (Yamamoto et al., 2000; Corti et
virta
ith doxycycline failed to induce recombination in all the adult tissues analyzed.ackground Xgal expression could be clearly observed in tissues of to the centralervous system.
l., 1999), and it is currently one of the most widely used regu-atory system for conditional gene expression. Tet-based systemsre usually based on two separate constructs and require theeneration of two independent transgenic mouse lines, the firstontaining the gene of interest under the control of a tetracyclineesponse element (TRE), and the second containing the transacti-ator (tTA) under the control of a constitutive or a tissue specificromoter. In the original variant (Tet-off), tetracycline preventsranscription by inactivating tTA. Thus the continuous presencef tetracycline is needed to prevent expression of the target gene.owever, the long-term maintenance of tetracycline levels to pre-ent target gene expression can be technically challenging and
t increases the likelihood of side effects. In contrast, the moreecent variant (Tet-on) uses a reversed tTA (rtTA), which becomesranscriptionally active only when bound to tetracycline or itsnalogs.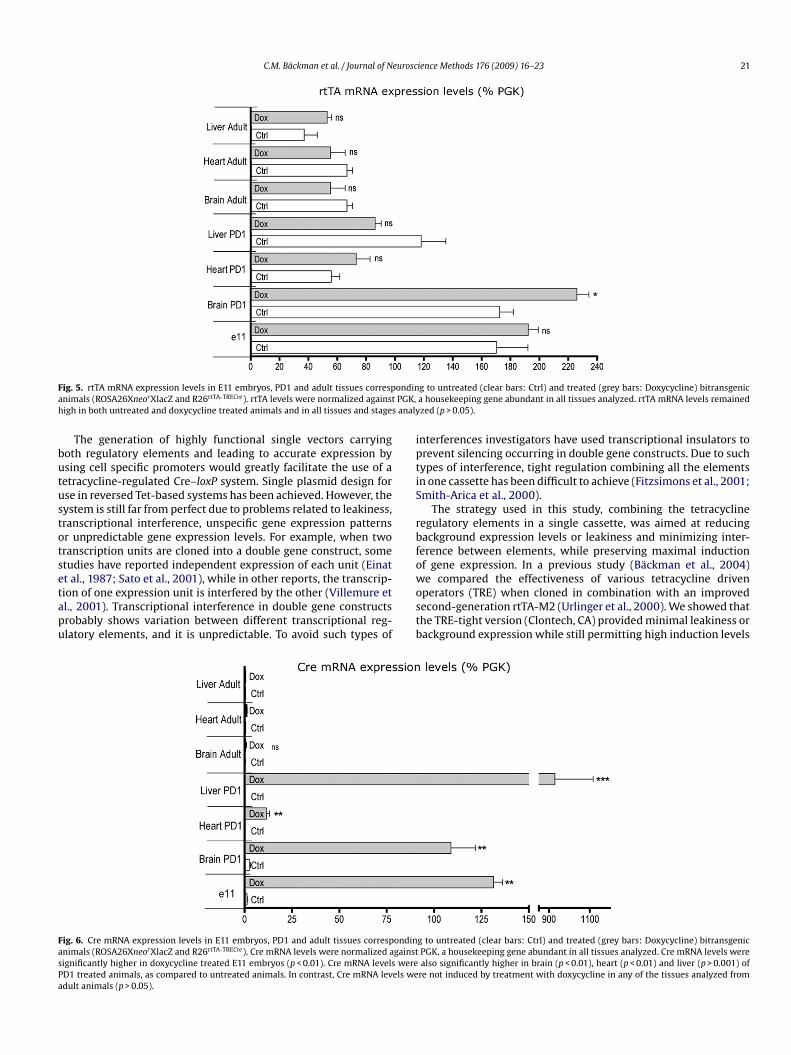
C.M. Bäckman et al. / Journal of Neuroscience Methods 176 (2009) 16–23 21
F ondina PGK,h analy
butustotsetapu
iptiS
rbfo
FasPa
ig. 5. rtTA mRNA expression levels in E11 embryos, PD1 and adult tissues correspnimals (ROSA26XneorXlacZ and R26rtTA-TRECre). rtTA levels were normalized againstigh in both untreated and doxycycline treated animals and in all tissues and stages
The generation of highly functional single vectors carryingoth regulatory elements and leading to accurate expression bysing cell specific promoters would greatly facilitate the use of aetracycline-regulated Cre–loxP system. Single plasmid design forse in reversed Tet-based systems has been achieved. However, theystem is still far from perfect due to problems related to leakiness,ranscriptional interference, unspecific gene expression patternsr unpredictable gene expression levels. For example, when tworanscription units are cloned into a double gene construct, sometudies have reported independent expression of each unit (Einat
t al., 1987; Sato et al., 2001), while in other reports, the transcrip-ion of one expression unit is interfered by the other (Villemure etl., 2001). Transcriptional interference in double gene constructsrobably shows variation between different transcriptional reg-latory elements, and it is unpredictable. To avoid such types ofwostb
ig. 6. Cre mRNA expression levels in E11 embryos, PD1 and adult tissues correspondinnimals (ROSA26XneorXlacZ and R26rtTA-TRECre). Cre mRNA levels were normalized againstignificantly higher in doxycycline treated E11 embryos (p < 0.01). Cre mRNA levels wereD1 treated animals, as compared to untreated animals. In contrast, Cre mRNA levels wedult animals (p > 0.05).
g to untreated (clear bars: Ctrl) and treated (grey bars: Doxycycline) bitransgenica housekeeping gene abundant in all tissues analyzed. rtTA mRNA levels remainedzed (p > 0.05).
nterferences investigators have used transcriptional insulators torevent silencing occurring in double gene constructs. Due to suchypes of interference, tight regulation combining all the elementsn one cassette has been difficult to achieve (Fitzsimons et al., 2001;mith-Arica et al., 2000).
The strategy used in this study, combining the tetracyclineegulatory elements in a single cassette, was aimed at reducingackground expression levels or leakiness and minimizing inter-erence between elements, while preserving maximal inductionf gene expression. In a previous study (Bäckman et al., 2004)
e compared the effectiveness of various tetracycline drivenperators (TRE) when cloned in combination with an improvedecond-generation rtTA-M2 (Urlinger et al., 2000). We showed thathe TRE-tight version (Clontech, CA) provided minimal leakiness orackground expression while still permitting high induction levels
g to untreated (clear bars: Ctrl) and treated (grey bars: Doxycycline) bitransgenicPGK, a housekeeping gene abundant in all tissues analyzed. Cre mRNA levels werealso significantly higher in brain (p < 0.01), heart (p < 0.01) and liver (p > 0.001) ofre not induced by treatment with doxycycline in any of the tissues analyzed from

2 eurosc
u(tepatecgttepilctgtsSswfiStTcwlos(niaaITeiRpRAncatIdlrrtd2waatbap
rio
dstmeiolatwtretso
A
o
R
A
B
B
C
C
E
F
G
G
H
H
L
L
M
M
2 C.M. Bäckman et al. / Journal of N
pon exposure to the inducer. Therefore, both these elementsrtTA-M2 and TRE-tight) were used in this study to generate theargeting vector. In addition, to further minimize potential interfer-nce between both elements when carried in a single cassette, welaced the rtTA-M2 and TRE-tight elements in opposite directions,nd separate them with a p53 intron spacer to reduce potentialranscriptional interference (Utomo et al., 1999). When bothlements were cloned in this manner and transfected into HK293ells, we were able to induce high expression levels of a reporterene after exposure to doxycycline (Bäckman et al., 2004). In addi-ion, to achieve generalized gene expression we opted to targethe endogenous ROSA26 locus by homologous recombination. Thendogenous ROSA26 locus remains active through the mouse lifes-an, and has been shown to provide generalized gene expression
n developing and adult animals. In addition, the use of a genomicocus to drive gene expression bypasses many of the problems asso-iated with random DNA integration. Previous studies have utilizedhe ROSA26 locus to achieve generalized gene expression in trans-enic animals. For example, Nagy and collaborators (2005) targetedhe rtTA transgene into the ROSA26 locus. In this study, rtTA expres-ion was conditional to a Cre recombinase-mediated excision of aTOP region from the ROSA26 locus. This recombinant mouse washown to express the rtTA transgene in tissues corresponding toell-characterized Cre lines, and rtTA expression levels were suf-cient to induce recombination in developing and adult animals.imilarly, the recombinant mice generated in this study utilizedhe endogenous ROSA26 promoter to drive rtTA-M2 expression.o investigate the ability of the R26rtTA-TRECre line to implementonditional DNA recombination in the embryo and adult animals,e crossed our recombinant line with a Cre dependent reporter
ine ROSA26XneorXlacZ. Nearly complete DNA recombination wasbserved during early developmental stages (E9–E15) after expo-ure to the inducer. However, during later developmental stagesE15–PD1) the efficiency of inducible Cre mediated DNA recombi-ation was clearly reduced, as different organs revealed differences
n the percentage of Xgal positive cells. An interesting finding in ournalysis was that, recombination efficiency during developmentppeared to be most problematic in the central nervous system.n addition, recombination could not be induced in adult animals.hese results have some similarities to those observed by Hameyert al. (2007), as they reported a lack of Cre mediated recombinationn the adult brain of a tamoxifen deleter strain targeted in theOSA26 locus. In this report the authors failed to detect CreERT2rotein signal in brain extracts, suggesting the inability of theOSA26 locus to drive expression of the transgene in this tissue.lso, in their study, Cre mediated DNA recombination was of mosaicature in adult peripheral tissues. Interestingly, in our study weould detect high levels of rtTA-M2 mRNA expression in all tissuesnalyzed during development and in adult animals, suggesting thathe endogenous ROSA26 promoter remains constitutively active.n contrast, while Cre recombinase mRNA levels were high duringevelopment after exposure to the inducer, Cre mRNA expression
evels were low in adult animals. Therefore, the absence of DNAecombination in adult animals is best explained by deficient Creecombinase expression. Previous studies have suggested thatransgenes integrated into animals in some cases may be silencedue to changes in chromatin structure (Mutskov and Felsenfeld,004). In addition, transcriptional interference has been reportedhen using double gene constructs in vitro and in vivo (Hasegawa
nd Nakatsuji, 2002; Villemure et al., 2001). It is possible that
lthough transcriptional interference may not completely silenceransgene expression in stem or progenitor cells, expression maye downregulated during development (Strathdee et al., 2006). Inddition, Bagchi et al. (2006) reported that the activity of the CMVromoter was downregulated in differentiated neurons, whileM
M
M
ience Methods 176 (2009) 16–23
emaining active in precursor and stem cells. These previous stud-es and the findings in this study clearly show the unpredictabilityf manipulating the genome to produce a desired phenotype.
In summary we have presented a qualitative analysis of a newlyeveloped R26rtTA-TRECre deleter line. The tetracycline regulatoryystem presented here provides investigators with a tool for gainingemporal control of transgene expression and gene ablation during
ouse development. Knockout mouse models have revealed sev-ral genes that interfere with embryo formation. A clear examples sonic hedgehog (SHH), a gene expressed through early devel-pment and essential for neural tube formation. Knockout miceacking SHH display abnormalities as early as embryonic day 7,nd lack the spinal cord and all the ventral cell types withinhe neural tube (Chiang et al., 1996). By delaying SHH deletion,e may investigate the function of this gene as it interacts with
he organism during later developmental windows. Therefore, theecombinant animals generated in this study should be useful forxperiments designed to address certain biological questions per-aining to developmental processes. The ability to delete genes atpecific times during development could provide further definitionf their roles during maturation.
cknowledgement
This study was supported by the Intramural Research Programf the NIH.
eferences
lbanese C, Hulit J, Sakamaki T, Pestell RG. Recent advances in inducible expressionin transgenic mice. Semin Cell Dev Biol 2002;13(2):129–41.
äckman CM, Zhang Y, Hoffer BJ, Tomac AC. Tetracycline-inducible expression sys-tems for the generation of transgenic animals: a comparison of various induciblesystems carried in a single vector. J Neurosci Methods 2004;139(2):257–62.
agchi B, Kumar M, Mani S. CMV promotor activity during ES cell differentia-tion: potential insight into embryonic stem cell differentiation. Cell Biol Int2006;30:505–13.
hiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H, Beachy PA. Cyclopiaand defective axial patterning in mice lacking Sonic hedgehog gene function.Nature 1996;383:407–13.
orti O, Sabaté O, Horellou P, Colin P, Dumas S, Buchet D, et al. A single adenovirusvector mediates doxycycline-controlled expression of tyrosine hydroxylase inbrain grafts of human neural progenitors. Nat Biotechnol 1999;17(4):349–54.
inat P, Bergman Y, Yaffe D, Shani M. Expression in transgenic mice of two genes ofdifferent tissue specificity integrated into a single chromosomal site. Genes Dev1987;1:1075–84.
itzsimons HL, Mckenzie JM, During MJ. Insulators coupled to a minimal bidirec-tional tet cassette for tight regulation of rAAV-mediated gene transfer in themammalian brain. Gene Ther 2001;8(22):1675–81.
arcia EL, Mills AA. Getting around lethality with inducible Cre-mediated excision.Semin Cell Dev Biol 2002;13(2):151–8.
ossen M, Bujard H. Tight control of gene expression in mammaliancells by tetracycline-responsive promoters. Proc Natl Acad Sci USA1992;89(12):5547–51.
ameyer D, Loonstra A, Eshkind L, Schmitt S, Antunes C, Groen A, et al. Toxicity ofligand-dependent Cre recombinases and generation of a conditional Cre deletermouse allowing mosaic recombination in peripheral tissues. Physiol Genomics2007;31(1):32–41.
asegawa K, Nakatsuji N. Insulators prevent transcriptional interference betweentwo promoters in a double gene construct for transgenesis. FEBS Lett2002;520:47–52.
obe CG, Nagy A. Conditional genome alteration in mice. Bioessays1998;20(3):200–8.
obe CG, Koop KE, Kreppner W, Lomeli H, Gertsenstein M, Nagy A. Z/AP, a doublereporter for Cre-mediated recombination. Dev Biol 1999;208:281–92.
ansuy IM, Bujard H. Tetracycline-regulated gene expression in the brain. Curr OpinNeurobiol 2000;10(5):593–6.
arth JD. Recent advances in gene mutagenesis by site-directed recombination. JClin Invest 1996;97(9):1999–2002.
ills AA. Changing colors in mice: an inducible system that delivers. Genes Dev2001;15:1461–7.
orozov A, Kellendonk C, Simpson E, Tronche F. Using conditional mutagenesis tostudy the brain. Biol Psychiatry 2003;54(11):1125–33.
utskov V, Felsenfeld G. Silencing of transgene transcription precedes methylationof promoter DNA and histone H3 lysine 9. EMBO J 2004;23(1):138–49.

eurosc
N
R
R
S
S
S
S
T
U
U
C.M. Bäckman et al. / Journal of N
agy A. Cre recombinase: the universal reagent for genome tailoring. Genesis2000;26:99–109.
ajewsky K, Gu H, Kühn R, Betz UA, Müller W, Roes J, et al. Conditional gene targeting.J Clin Invest 1996;98(3):600–3.
yding AD, Sharp MG, Mullins JJ. Conditional transgenic technologies. J Endocrinol2001;171(1):1–14.
ato M, Watanabe T, Oshida A, Nagashima A, Miyazaki J-I, Kimura M. Usefulnessof double gene construct for rapid identification of transgenic mice exhibitingtissue-specific gene expression. Mol Reproduction and Dev 2001;60:446–56.
mith-Arica JR, Morelli AE, Larregina AT, Smith J, Lowenstein PR, CastroMG. Cell-type-specific and regulatable transgenesis in the adult brain:adenovirus-encoded combined transcriptional targeting and inducible trans-
gene expression. Mol Ther 2000;2(6):579–87.oriano P. Generalized lacZ expression with the ROSA26 Cre reporter strain. NatGenet 1999;21(1):70–1.
trathdee D, Ibbotson H, Grant SGN. Expression of transgenes targeted to theGt(ROSA)26Sor locus is orientation dependent. PLoS ONE 2006;1(December20):e4.
V
Y
ience Methods 176 (2009) 16–23 23
omac AC, Grinberg A, Huang SP, Nosrat C, Wang Y, Borlongan C, et al.Glial cell-line derived neurotrophic factor receptor alpha1 availability reg-ulates glial cell-line derived neurotrophic factor signaling: evidence frommice carrying one or two mutated alleles. Neuroscience 2000;95(4):1011–23.
rlinger S, Baron U, Thellmann M, Hasan MT, Bujard H, Hillen W. Exploring thesequence space for tetracycline-dependent transcriptional activators: novelmutations yield expanded range and sensitivity. Proc Natl Acad Sci USA2000;97(14):7963–8.
tomo ARH, Nikitin AY, Lee W-H. Temporal, spatial, and cell typr-specific con-trol of Cre-mediated DNA recombination in transgenic mice. Nat Biotechnol1999;17:1091–6.
illemure JF, Savard N, Belmaaza A. Promoter suppression in cultures mammaliancells can be blocked by the chicken beta-globin chromatin insulator 5′HS4 andmatrix/scaffold attachment regions. J Mol Biol 2001;312:963–74.
amamoto A, Lucas JJ, Hen R. Reversal of neuropathology and motor dys-function in a conditional model of Huntington’s disease. Cell 2000;101(1):57–66.