functional properties of ca 2+inhibitable type 5 and type 6 adenylyl cyclases and role of ca 2+...
TRANSCRIPT

Cell. Signal. Vol. 11, No. 9, pp. 651–663, 1999 ISSN 0898-6568/99 $ – see front matterCopyright 1999 Elsevier Science Inc. PII S0898-6568(99)00031-5
TOPICAL REVIEW
Functional Properties of Ca21-InhibitableType 5 and Type 6 Adenylyl Cyclases and Role of Ca21
Increase in the Inhibition of Intracellular cAMP ContentDanielle Chabardes,* Martine Imbert-Teboul and Jean-Marc Elalouf
URA 1859 CNRS, Service de Biologie Cellulaire, Departement deBiologie Cellulaire et Moleculaire, Bat 520, CEA Saclay, 91191 Gif sur Yvette, France
ABSTRACT. Among the different adenylyl cyclase (AC) isoforms, type 5 and type 6 constitute a subfamilywhich has the remarkable property of being inhibited by submicromolar Ca21 concentrations in addition to Gai-mediated processes. These independent and cumulative negative regulations are associated to a low basal enzy-matic activity which can be strongly activated by Gas-mediated interactions or forskolin. These properties ensurepossible wide changes of cAMP synthesis. Regulation of cAMP synthesis by Ca21was studied in cultured or nativecells which express naturally type 5 and/or type 6 AC, including well-defined renal epithelial cells. The resultsunderline two characteristics of the inhibition due to agonist-elicited increase of intracellular Ca21: i) Ca21 risesachieved through capacitive Ca21 entry or intracellular Ca21 release can inhibit AC to a similar extent; and ii)in a same cell type, different agonists inducing similar overall Ca21 rises elicit a variable inhibition of AC activity.The results suggest that a high efficiency of AC regulation by Ca21 is linked to a requisite close localization ofAC enzyme and Ca21 rises. cell signal 11;9:651–663, 1999. 1999 Elsevier Science Inc.
KEY WORDS. Ca21-inhibitable adenylyl cyclases, Intracellular cAMP content, Inhibition by Ca21 increase,AC5 or AC6-transfected cells, Intact cells, Isolated segments of the renal tubule
INTRODUCTION protein kinases A and C and/or the intracellular free concen-tration of Ca21 ([Ca21]i) (Table 1). The activity of each ade-Intracellular adenosine 39:59-cyclic monophosphate (cAMP)nylyl cyclase (AC) isoform described up to date thereforeplays a key role in the control of cellular functions. The in-can integrate signals emerging from the activation of othertracellular content of cAMP results from its synthesis, en-signalling pathways.sured by the activation of adenylyl cyclases, and from its hy-
This review focuses on the properties of the AC isoformsdrolysis, catalysed by phosphodiesterases. These two enzymetype 5 (AC5) and type 6 (AC6) and on the regulation offamilies comprise multiple isoforms, the regulatory proper-intracellular cAMP content resulting from their expressionties of which differ from one isoform to another [1].in different cell types. The properties of AC5 and AC6 en-Several excellent reviews have previously recalled thezymatic activity have been studied in transfected cells or ingeneral characteristics of adenylyl cyclases and some of thecells which naturally express these isoforms. Both approachesspecific properties exhibited by the nine isoforms cloned uphave allowed to define similar properties which, thus, charac-to date [1–6]. The regulatory factors include the bg subunitsterise Ca21-inhibitable AC. In membrane preparations, one ofof the heterotrimeric GTP binding proteins, the activity oftheir fundamental properties is to be inhibited by submicro-molar concentrations of Ca21 in addition to Gai-mediated
*Author to whom all correspondence should be addressed. Tel: 133-1- processes. In intact cells, the term “Ca21-inhibitable” AC is6908-9761; fax: 133-1-6908-3570; e-mail: [email protected] appropriate as evidenced by two lines of obser-Abbreviations: AC–adenylyl cyclase; AVP–arginine vasopressin; [Ca21]i–
intracellular free concentration of Ca21; [Ca21]e–extracellular free concen- vations that emerged from several recent studies discussedtration of Ca21; cAMP–adenosine 39:59-cyclic monophosphate; CCE–ca-
in this review. First, the magnitude of Ca21 inhibition ispacitive Ca21 entry; CT–calcitonin; CTAL–cortical portion of the thickascending limb; Gas, Gai and Gbg, stimulatory, inhibitory, and bg sub- highly variable from one cell type to another and the ex-units of guanine nucleotide-binding regulatory protein; IBMX–3-isobutyl- pression of one Ca21 inhibitable AC does not always confer1-methylxanthine; OMCD–outer medullary portion of the collecting duct;
a strong inhibition of AC activity by agonists elicitingPDE–phosphodiesterase; PGE2–prostaglandin E2; PKA–protein kinase A;PKC–protein kinase C; PTH–parathyroid hormone; RaKCaR–rat kidney [Ca21]i increase. Second, in a given cell type, any [Ca21]i in-extracellular Ca21-sensing receptor; Ro 20-1724–4-(3-butoxy-4-methoxy- crease is not able to decrease AC activity. These observa-benzyl)-2-imidazolidinone.
Received 5 December 1998; and accepted 27 January 1999. tions underline a dual specificity of the putative regulation

652 D. Chabardes et al.
TABLE 1. Differential regulation of mammalian adenylyl cyclase isoforms
AC isoforms Ca21 Gai Gbg PKA PKC
AC5 ↘ ↘ NC ↘ ↘ ↗AC6 ↘ ↘ NC ↘ ↘ NC
AC3 ↗ ↘a ↘ NC ? ↗ b
AC9 NC ↘c ? NC ? ?
AC1 ↗ d ↘e ↘ NC ↗ f
AC8 ↗ d NC ? ? NC
AC2 NC NC ↘g ↗ h ↘i ↗AC4 NC ↘j ↗ h ? ↘k
AC7 NC ? ↗ h ? ↗AC isoforms are listed as a function of their regulation by Ca21: direct inhibition by Ca21 in vitro and in vivo(AC5 and AC6), indirect inhibition by Ca21 in vivo (AC3 and AC9), stimulation by Ca21/calmodulin (CaM)in vitro and in vivo (AC1 and AC8), not regulated by Ca21 (AC2, AC4, and AC7). Detailed data obtainedwith AC5 and AC6 isoforms are given in the text. Activity: ↗ 5 increase in activity; ↘ 5 decrease in activity;? 5 effect unknown; NC 5 no change in activity. The following footnotes provide respective comments withthe corresponding references given in brackets.
a Ca21/CaM-dependent stimulation of Gas-stimulated activity observed with high Ca21 concentrations invitro [14] and CaM kinase II-mediated inhibition in vivo [15].
b Increase of all activities [50].c No effect in vitro [16] and calcineurin-mediated inhibition in vivo [17].d Stimulation mediated by Ca21/CaM in vivo and in vitro [1, 18, 19, 32, 33].e Modest inhibition of Ca21/CaM-stimulated activity in vitro [37] and pronounced inhibition in vivo [20].f Increase of forskolin- and Ca21/CaM-stimulated activity [22, 50].g No effect in vitro [38] and inhibition of forskolin-stimulated activity in vivo [21].h Activation of Gas-stimulated activity [24–28, 42].i Modest inhibition of Gas-stimulated activity [48].j Inhibition of forskolin-stimulated activity in vivo [21].k Inhibition of Gas-stimulated and bg-potentiated activity [23].
of AC activity by Ca21, i.e., the cell type involved and the and/or forskolin [7–11, 12, 29, 30, 34, 35]. Forskolin andGas-mediated processes induce a synergistic activation ofagonist eliciting [Ca21]i rise.
The first part of this review will recall the properties ob- AC6 [12, 30] and, although less pronounced, of AC5 [34,35]. The expression of similar amounts of AC2 and AC6tained in AC5- and AC6-transfected cells and their com-
parison to other AC isoforms. The second part will develop has allowed a careful comparison of their enzymatic activi-ties [29]. Whatever the concentration of Mg21 used in thethe properties of regulation observed in intact cells which
express preferentially, if not exclusively, AC5 and/or AC6 assay, basal activity of AC6 is at least 20-fold lower thanmRNAs and the role of [Ca21]i increase in the regulation of that of AC2. This difference does not appear linked to theintracellular cAMP content. Some examples will be taken catalytic capability of each isoform but rather to a differentfrom defined native cells of the renal tubule which appear stimulation by Mg21 [29]. A marked difference is observedparticularly suitable to emphasize several properties of Ca21- between AC6 and AC2 for (100 mM) forskolin-dependentinhibitable AC isoforms. stimulation (about 40- and 6-fold with AC6- and AC2-
transfected cells, respectively [29]).
REGULATORY PROPERTIES OFTYPE 5 AND TYPE 6 ADENYLYL CYCLASES
Regulatory FactorsMolecular cloning of type 5 [7, 8, 11] and type 6 [9–13] AC Several factors can decrease AC5 and AC6 enzymatic ac-has been performed from different cDNA libraries. The in- tivity. Concerning Ca21, free-Ca21 concentrations should betrinsic properties of these isoforms and the factors suscepti- accurately determined to detect a possible inhibition by lowble to regulate each enzymatic activity have been studied ei-
concentrations. In such experimental conditions, Cooperther in intact mammalian cells transfected with the isoformand collaborators have demonstrated that 0.3 mM free-Ca21
of interest or in membranes prepared from transfected cellsinduces a maximal inhibition of murine AC6 isoform [12]).or from insect cells infected with recombinant baculovirus.This result supported previous observations made on mem-In most experiments, the properties elicited by different ACbranes prepared from NCB-20 cells [31] from which AC6isoforms were studied in parallel, thus strengthening the dif-cDNA has been cloned [12]. In the same AC assay condi-ferential regulation observed from one AC type to anothertions, rabbit AC5 cloned from myocardium and expressed(Table 1).in COS-1 cells is inhibited with a higher sensitivity by sub-micromolar concentrations of Ca21 [8]. However, the con-
Basal and Stimulated AC Activity centration of free-Ca21 inducing half-maximal inhibition ofcanine AC5 expressed in Sf9 cells is only 7 mM [34]. TheType 5 and type 6 AC exhibit a low basal enzymatic activity
which is strongly stimulated by Gas-mediated activation discrepant observations noted above suggest that AC5 sen-

Cellular cAMP and Ca21-inhibitable Adenylyl Cyclase 653
sitivity to Ca21 may be linked to additional unknown factors analogues exhibit the same order of potency to decreaseAC5 and AC6 activity [7, 9].and/or may vary from one expression system to another.
The inhibition elicited by low concentrations of Ca21 doesnot depend on an interaction with calmodulin [12, 34]. A
Regulation by Protein Kinase Arecent report has established that Ca21 binds directly to thePremont et al. provided the first evidence of an inhibitory ef-C terminus of the first large cytosolic loop of AC5 [34]. Thisfect of protein kinase A (PKA) on AC5 and/or AC6 activ-region is conserved in AC6, but not in other AC isoformsity present in chick hepatocytes or S49 murine lymphoma[34]. However, the relatively high concentration of Ca21 ef-cells [13]. Regulation by PKA accounts, at least partly, forfective to inhibit AC in this study (see above) does notthe glucagon-induced heterologous desensitization of ade-allow one to conclude that this Ca21 binding site accountsnylyl cyclase previously observed in chick hepatocytes [46].for the inhibition of AC5 and AC6 by submicromolar con-
In membranes prepared from recombinant cells or with pu-centrations of Ca21. It is important to note that the activityrified enzymes, AC5 and AC6 are directly phosphorylatedof all AC isoforms can be inhibited by high concentrationsby protein kinase A catalytic unit [47, 48]. This phosphory-of Ca21: for example, the Ca21/calmodulin stimulable AC1lation decreases forskolin- or Gas-stimulated AC5 and AC6is activated with less than 1 mM free-Ca21; but, when free-activity by ,30–50%. AC5 basal activity is also decreasedCa21 is increased to about 10 mM, this activation is no[47] but not that of AC6 [48]. Kinetic analysis of forskolin-longer observed and the response slows down [32, 33, 36];activated AC5 shows that the Km for ATP is unaltered andsuch high Ca21 concentrations also induce a second phasethat inhibition results from a decrease in AC catalytic rateof inhibition in NCB-20 cells [31].[47]. Mutagenesis of AC6 cDNA has allowed the definitionPertussis toxin-dependent Gai subunits inhibit Gas-acti-of a single site of phosphorylation by PKA which appears es-vated type 5 and type 6 activity measured in membranessential in PKA-mediated inhibition of Gas-sensitive activ-prepared from recombinant Sf9 cells. Gai1, Gai2 and Gai3ity [48]. The phosphorylation by PKA does not affect AC1exhibit the same efficiency, leading to ,70–80% inhibitionand slightly decreases Gas-activated AC2 [48].[37, 38]. A similar inhibition is observed on forskolin-acti-
vated enzymes suggesting that Gai-mediated inhibition isindependent of an interaction with Gas. Ga0 has no effect, Regulation by Protein Kinase Cin contrast to the inhibition observed on forskolin- or Ca21/
In mammalian COS-7- or HEK-293-transfected cells, thecalmodulin-stimulated AC1 [38]. The strong inhibition of
activation of protein kinase C (PKC) isoforms by phorbolGai subunits on Ca21-inhibitable AC subtypes is also observed esters does not affect AC6 activity [39, 49, 50]. This resultin intact transfected COS-7 cells since Gai2 elicits ,80% in- is sustained by the stimulatory effect elicited by PKCs onhibition of type 6 AC activity [39]. Inhibition of cAMP syn- other AC isoforms studied in the same experiments andthesis can also be achieved through the pertussis toxin-insensi- most particularly on AC2. Also, a pretreatment with thetive Gaz subunit [40]. Tested on Gas-stimulated AC5, Gaz phorbol ester PMA does not impair Gai-mediated inhibi-is more potent than Gai1 in inhibiting AC activity [41]. tion of AC6 [39]. In contrast with these results, it has been
The activation of GTP-dependent heterotrimeric G pro- observed that a preparation of purified PKC isoenzymes de-teins liberates Gbg subunits which may regulate the activ- creases AC6 activity overexpressed in membranes preparedity of several AC isoforms as detailed in Table 1. A mixture from recombinant insect cells and induces a direct phos-of various bg subunits purified from bovine brain mem- phorylation of the enzyme [51]. The discrepant results ob-branes stimulates AC2 activity, but has no effect on AC5 tained in insect recombinant cells versus mammalian trans-or AC6 [11]. The cotransfection in HEK-293 cells of AC5 fected cells might be linked to the use of a purified preparationwith cloned b1 and g2 isoforms also shows that the enzy- of PKCs [51] which may contain different isoenzymes in ad-matic activity is not modified, in contrast to that observed dition to those regulated by phorbol esters. Thus, if differentwith AC 7 [42]. At variance with these results, recent data studies have clearly established that phorbol ester-sensitiveobtained in transfected COS-7 cells show that either b1g2 PKCs do not regulate AC6 activity [39, 49, 50], a possibleor b5g2 inhibits AC5 and AC6 activity [43]. Another AC effect of other PKC isoenzymes cannot be excluded.isoform was not tested in the same experiments, but the Using purified enzymes in an in vitro assay, Kawabe et al.same methodology has established previously that b1g2 have demonstrated that AC5 activity is stimulated by thestimulates and b5g2 inhibits AC2 activity, whereas both bg classical PKCa and the atypical diacylglycerol-insensitivecomplexes inhibit AC1 [44]. Results obtained in COS-7 PKCz [52]. These two PKC isoenzymes induce a cumulativecells, however, may be difficult to generalise because the increase in AC5 activity [52] and this dual stimulation hassensitivity of endogenous adenylyl cyclases to the regulation been confirmed in intact HEK-293 transfected cells [53].elicited by bg subunits appears particularly high [27]. PKCa and PKCz phosphorylate AC5 on distinct residues
All AC isoforms are inhibited by adenosine and some of [52] which are different from those phosphorylated by pro-its analogues through an interaction with an intracellular tein kinase A [47]. Interestingly, insulin increases forskolin-purine binding site (P-site). The magnitude of inhibition dependent cAMP accumulation in transfected cells and thisdepends on the analogue used and a differential sensitivity effect is potentiated by the coexpression of PKCz, but not
by that of PKCa [53]. This observation suggests stronglyis observed from one AC isoform to another [45]. Different

654 D. Chabardes et al.
that insulin-activated PKCz is able to stimulate AC5 activ- one cell type as suggested by morphological and functionality. It may be noted that the effect of insulin on protein syn- studies [61]. This conclusion is supported by the observationthesis is also mediated by the activation of PKCz [54]. The that the different agonists active on this cell type regulateactivation of AC5 by specific PKCs is to put together with the same pool of AC catalytic units or of intracellularprevious results obtained with epidermal growth factor cAMP [63, 64]. Other segments, such as the medullary col-(EGF) in HEK-293 cells transfected with different AC iso- lecting duct (OMCD), contain mainly two epithelial cellforms [55]. EGF stimulates adenylyl cyclase activity or types (principal and intercalated type A cells). In this het-cAMP accumulation in AC5-transfected cells but has no ef- erogeneous segment, however, functional and biochemicalfect in AC1-, AC2- or AC6- expressing cells. In contrast, data [61, 65, 66, 70, 73] allow the identification of a givenisoproterenol-stimulated AC activity is observed with all cell type by the specificity of the hormonal regulation of in-AC isoforms [55]. Different experimental conditions were tracellular cAMP content. Thus arginine vasopressintested in this study and the data support a specific require- (AVP) increases cAMP synthesis in the principal cellment of AC5 to observe a stimulation of cAMP synthesis whereas glucagon is active in the intercalated cell.by EGF. Similarly to that observed with insulin, the effect Northern blot analyses have demonstrated the presenceof EGF might be linked to the activation of specific PKCs of mRNAs encoding 4, 5, or 6 AC isoforms in the kidneyleading to the stimulation of AC5 activity. [9–11, 25]. The distribution of these mRNAs has been stud-
ied in different segments of the rat renal tubule [66, Fig. 1].The quantitative RT-PCR technique used allows determi-REGULATION OF INTRACELLULARnations of mRNAs from very small amounts of total RNAscAMP CONTENT AND PROPERTIES OF THEextracted from several microdissected samples of one de-INHIBITION ELICITED BY Ca21 IN INTACT CELLSfined segment [66, 67]. Type 6 mRNA is expressed at differ-
The expression of Ca21 inhibitable AC mRNAs does not al- ent levels all along the renal tubule whereas type 5 mRNAways correlate with the functional properties that are ob- is present only in the glomerulus and two portions of theserved [56, 57]. The properties of regulation linked to the collecting duct (Fig. 1). Type 4 mRNA, although highly ex-presence of one given mRNA isoform therefore may be dif- pressed in the rat kidney [25, 68], is expressed moderatelyficult to define in some tissues. Different factors can con- in the glomerulus but not in tubular segments. This obser-tribute to the difficulty of this analysis, such as the heteroge- vation suggests that type 4 mRNA is present in other renalneity of tissular or cellular preparations, the expression of an cell types such as vascular and/or interstitial cells. Since re-unknown AC isoform and/or of several isoforms in a same nal segments contain ,200–500 cells per millimetre tubu-cell as suggested from the distribution of AC mRNAs, see lar length, the data obtained with each mRNA correspondfor example [12, 21, 56–60]. However, different cell types to significant levels of expression, i.e., several mRNA cop-appear to be suitable examples to emphasise the characteris- ies per cell. Thus, if functional, type 5 and 6 adenylyl cy-tics of the regulation of cAMP content linked to the expres- clase proteins should be involved in cAMP synthesis alongsion of Ca21-inhibitable AC isoforms and to define the the renal tubule. In situ hybridization experiments on kid-properties of Ca21-induced inhibition. We will focus mainly ney sections have established that only type 6 mRNA is ex-on defined native cells of the renal tubule [61], different pressed in the principal cell and that both type 5 and 6preparations from cardiac tissue which have a strong expres-
mRNAs are present in the intercalated cell of the OMCDsion of AC5 and AC6 mRNAs [7–12, 62] and NCB-20 neu-[69]. Type 5 mRNA is also highly expressed in renal bloodroblastoma and C6-2B glioma cell lines which exhibit a pre-vessels (J. M. Verbavatz, personal communication).dominant expression of AC6 mRNA [12, 60]. Part I of the
The type 9 mRNA is also detected in the kidney [16, 68].next section summarises the distribution of Ca21-inhibitableThe level of expression is low and of comparable magnitudeAC mRNAs along the renal tubule and some functionalin the cortex and the medulla [68]. This isoform is not di-data relative to the regulation of AC activity observed in re-rectly inhibited by submicromolar concentrations of Ca21
nal segments. Part II emphasises the properties of the inhi-and is poorly stimulated by forskolin [16], in contrast to thatbition elicited by Ca21 in intact cell types and Part III re-observed on kidney membranes and isolated segments (seeports the data which support a regulation of adenylyl cyclasebelow). These results suggest that type 9 mRNA might haveactivity by local increases in [Ca21]i.a weak expression in the cells constitutive of the renal tu-bule and/or might be mainly expressed in other renal cells.I. Expression of Ca21-Inhibitable AC Isoforms
and Regulation of cAMP Synthesis in Renal Segments REGULATION OF ADENYLYL CYCLASE ACTIVITY BY Gas-,
Gai- AND PKC-MEDIATED INTERACTIONS IN RENAL SEG-DISTRIBUTION OF mRNAs ENCODING Ca21-INHIBITABLE ISO-MENTS. In different segments, low basal enzymatic activi-FORMS ALONG THE RENAL TUBULE. The monolayer of epi-ties contrast with the high stimulated values induced bythelial cells constitutive of the different segments of the re-hormones or neuromediators active on their respective tar-nal tubule exhibits unique properties. Each segment, free ofget sites [61, 63]. Table 2 shows examples of the strong stim-vascular and interstitial cells, can be obtained by microdis-ulation factors which are achieved in the CTAL and thesection (Fig. 1). It is very likely that some segments, such
as the cortical thick ascending limb (CTAL), contain only principal and intercalated cells of OMCD. Forskolin (1 or

Cellular cAMP and Ca21-inhibitable Adenylyl Cyclase 655
FIGURE 1. Distribution of AC4, AC5 and AC6 mRNAs along the rat renal tubule. Left: Quantitative RT-PCR analysis of AC5 andAC6 in different segments of the rat renal tubule. For each AC type, amplified cDNAs (28 cycles) corresponding to mutated (Mut)and wild type (WT) RNA have the same length and differ only by two base pairs to introduce a new restriction site in Mut cDNA.RT-PCR amplification of mutated and wild type RNAs is of a same efficiency and the coamplification of AC5 and AC6 templates ina same reaction tube does not modify the quantitation (66). AC6 mRNA is present all along the nephron whereas the expression ofAC5 is restricted to the glomerulus and the cortical and outer medullary portions of the collecting duct. Right: Schematic representationof a renal tubule giving some examples of quantitation of AC mRNAs in defined structures. The results are expressed as moleculesof mRNA per glomerulus or per millimetre of segment. Abbreviations: Glom–glomerulus; PCT–proximal convoluted tubule; CTAL–cortical portion of the thick ascending limb; CCD, OMCD, IMCD–cortical, outer medullary and inner medullary portion of the collect-ing duct, respectively. Data are reproduced from [66] with the permission of The American Society for Biochemistry and MolecularBiology.
5 mM) activates AC activity with a magnitude comparable 80% [79]. Different results establish that this observation isdue to a high homologous desensitization and to a small (ofto those elicited by hormonal concentrations inducing maxi-
mal effects [64, 76]. Also, forskolin has a synergistic action on ,20%) heterologous desensitization [79]. By comparison tothat observed in AC6-transfected cells (see Part 1), the het-AVP-induced cAMP production in the medullary portion of
the thick ascending limb [78] which expresses type 6 mRNA erologous process might result from the activation of PKA.The activation of PKC isoforms with phorbol esters does(Fig. 1). Pertussis toxin-sensitive pathways elicit high inhi-
bitions of AC activity (Table 2). not modify hormone-stimulated cAMP accumulation in thethick ascending limb and the vasopressin-sensitive cells ofThe preincubation (60 min) of thick ascending limbs with
AVP reduces AVP-dependent cAMP synthesis by about OMCD [80, 81] or pertussis toxin-sensitive inhibition in-
TABLE 2. Effect of different agonists on AC activity in CTAL and OMCD isolated from the rat kidney
Gas-interaction Gai-interactionCell type (stimulated over basal activity) (% inhibition of the hormonal response)
CTAL cell AVP-V2 (314); PTH (335) PGE2 (55%)b1-adrenergic agonists (38)CT-CT1a (327); Glucagon (333)
OMCDPrincipal cell AVP-V2 (320) a2-adrenergic agonists (85%)
A1-adenosine agonists (52%)
Intercalated cell Glucagon (310) PGE2 (62%)
The magnitude of stimulation of AC activity or the percentage of inhibition elicited by each agonist is given in parentheses. The stimulation of ACactivity was measured by using an AC microassay on permeabilized segments. The receptor subtype eliciting cAMP synthesis has been previously characterized[67, 71, 72] for some agonists. The inhibition of hormone-stimulated intracellular cAMP content has been determined from cAMP values measured onintact segments incubated in the presence of either IBMX, which blocks all PDEs types in the rat kidney [74], or Ro 20-1724, inhibitor of the low-Km,cAMP specific PDE 4 family. All inhibitions indicated here were sensitive to pertussis toxin. The data are from [61, 63, 64, 70, 73, 75–77].
Abbreviations: AVP 5 arginine vasopressin; PTH 5 parathyroid hormone; CT 5 calcitonin; PGE2 5 prostaglandin E2; CTAL 5 cortical thick ascendinglimb; OMCD 5 collecting duct microdissected from the outer medulla.

656 D. Chabardes et al.
duced by a2-adrenergic agonists [81]. To date, potential regula- with Gai-mediated processes. AC5 and AC6 isoformstherefore appear well-adapted to ensure, when necessary,tion of PKCs with the AC activity present in glucagon-sen-
sitive cells of the collecting duct has not been investigated. independent or simultaneous and combined inhibitions ofcAMP synthesis. Ca21-mediated inhibition is observed onforskolin- as well as on hormone-dependent cAMP synthe-
II. Regulation of cAMP Synthesis sis [31, 64, 82, 84, 85]. An additional observation can beby Ca21 and Properties of Ca21-Induced made if we consider not only the step of cAMP synthesis,Inhibition of Intracellular cAMP Content but the regulation of intracellular cAMP content, thus in-Adenylyl cyclase assays performed on membranes or perme- cluding the potential role of Ca21 to activate phosphodies-abilized cells have demonstrated that AC activity is inhib- tease activity. PDEs activation by Ca21 does not contributeited by submicromolar free-Ca21 concentrations in NCB-20 to the decrease of cAMP accumulation in cardiac myocytes[31] or C6-2B [82] cell lines, cultured pulmonary endothe- [84], NCB-20 cell line [90] or substance K receptor-trans-lial cells [83] or cardiac myocytes [84], and purified cardiac fected C6-2B cells [82]. However, in wild type C6-2B cellsarcolemmal vesicles [85]. AC sensitivity to free-Ca21 has line [91], cultured artery endothelial cells [83] and nativenot been studied in defined renal segments, but on cortical renal cells [64, 76, 88], Ca21-mediated inhibition of cAMPor medullary membranes prepared from rat kidney, for- content is linked to both a decrease in AC activity and anskolin-dependent AC activity is decreased by submicromo- increase in PDE activity (Table 5). As underlined by Coo-lar free-Ca21 concentrations [68]. This observation, the ex- per et al. [91], a modest inhibitory effect on AC activitypression of AC5 and AC6 mRNAs along the rat renal coupled to an increase in cAMP breakdown can lead to atubule (Fig. 1), and the similarity of the properties of regula- high inhibition of cAMP content. Thus, if Ca21-sensitivetion of AC activity observed in isolated segments (see PDEs are present in the cell type studied, the action of Ca21
above) and of those obtained in AC5- or AC6- transfected on cAMP hydrolysis amplifies its inhibitory effect on cAMPcells, strongly suggest the functional expression of Ca21in- synthesis. In the CTAL cell, it has been shown that Ca21-
induced cAMP breakdown is due to the activation of thehibitable AC types in these segments. In agreement withthis hypothesis, experiments performed on intact segments Ca21/calmodulin-dependent PDE family type I [64]. This
regulation combined with the Ca21-mediated inhibition ofhave established that cAMP synthesis is inhibited by ago-nist-induced [Ca21]i increase and/or by variations of extra- cAMP synthesis results in a strong decrease of intracellular
cAMP following stimulation of the Ca21-sensing receptorcellular Ca21 (Table 3).The results obtained on cell lines, cultured or native cells RaKCaR (Fig. 2).
In cultured cells, the inhibition of cAMP synthesis by ag-allow us to emphasise several properties that characteriseCa21-induced inhibition of AC activity (Table 4). Particu- onists inducing phospholipase C activation is often of low
magnitude [31, 60, 83, 90, 91]. A small effect is also ob-larly, the inhibition of hormone-dependent cAMP synthe-sis appears due to a direct interaction of Ca21 on catalytic served with bradykinin and angiotensin II in CTAL cells
(Table 5). In contrast, the activation of phospholipase C byunits, is insensitive to pertussis toxin, and is cumulative
TABLE 3. Inhibition of hormone-dependent cAMP synthesis by maneuvers inducing [Ca21]i increase
Cell type Experimental conditions % inhibition of Gas- stimulated AC activity
CTAL cell [Ca21]e: 1.5 mM vs. 0.5 35[Ca21]e: 2.5 mM vs. 0.5 65100 mM Neomycin 440.1 mM Angiotensin II 200.1 mM Bradykinin 182 mM Ionomycin No effect
OMCDPrincipal cell 0.3 mM PGE2 12a
(AVP-sensitive) 10 nM Ionomycin No effect1 mM Ionomycin 64[Ca21]e: 2.0 mM vs. 0 No effect
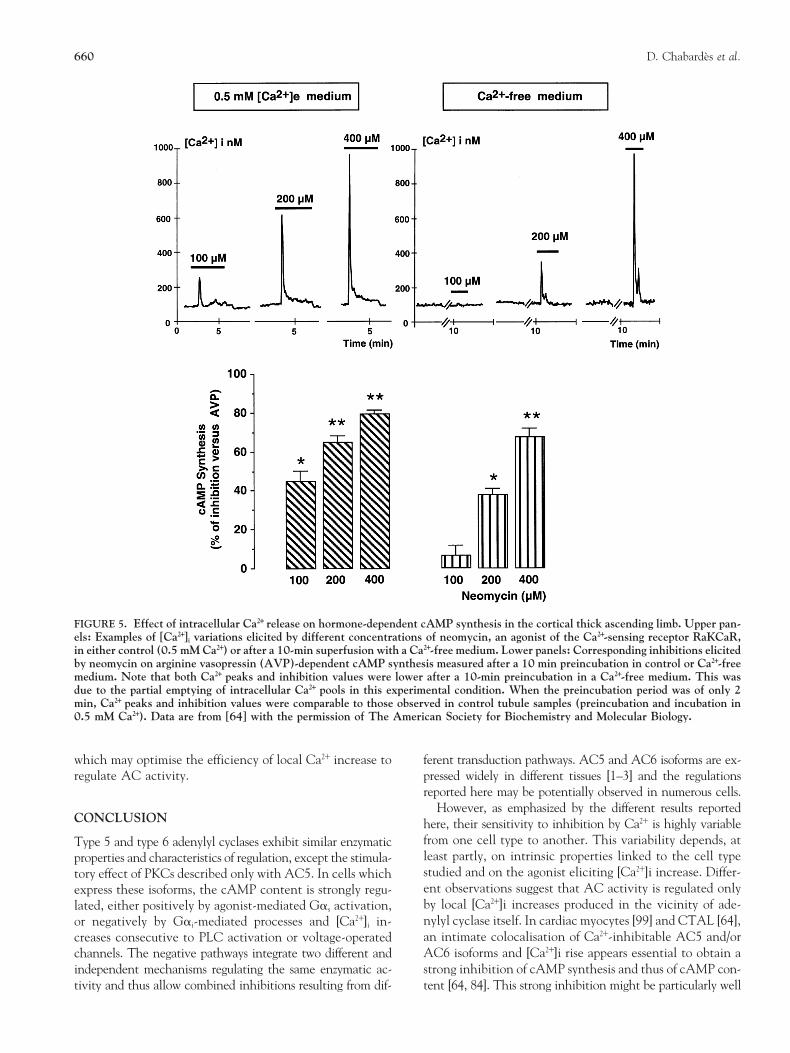
Intercalated cell Muscarinic agonists 51(glucagon-sensitive) [Ca21]e: 2.0 mM vs. 0 52
The inhibition of AC activity was measured on intact nephron segments in the presence of IBMX. CTAL: the inhibition elicited by variations of theconcentrations of extracellular Ca21 ([Ca21]e) is mediated by the Ca21-sensing receptor, RaKCaR [86, 87], coupled to PLC activation [64]; neomycin is anagonist of RaKCaR; pertussis toxin does not affect the inhibition due to [Ca21]e variations or angiotensin II [64]. OMCD: increases of [Ca21]e raise [Ca21]iand inhibit the response to glucagon [66] but there is no evidence demonstrating the presence of RaKCaR in this segment [86]; the inhibition elicitedby a high concentration of ionomycin in principal cell [66] has been also observed with A23187 [88]. CTAL cell and principal cell of the OMCD expresstype 6 AC; intercalated cell expresses type 5 and type 6 AC [66, 69]. The preferential localization of PGE2- and muscarinic agonists-mediated Ca21 increasesin either principal cells (sensitive to AVP) or intercalated cells (sensitive to glucagon) is based on their respective properties to inhibit hormone-stimulatedcAMP synthesis [66, 70, 76, 77] and on their additive effect on [Ca21]i increase in the OMCD (Aarab et al., unpublished data).
a The slight inhibition induced by PGE2 in the OMCD did not reach statistical significance [76] probably because of the biological scatter of the cAMPcontents from one microdissected segment to another.
Cellular cAMP and Ca21-inhibitable Adenylyl Cyclase 657
TABLE 4. Properties of Ca21-mediated inhibition of hormone-dependent cAMP synthesis in different cell types which exhibit apredominant expression of Ca21-inhibitable AC isoforms
Regulatory factor Ca21-mediated inhibition Cell type
Ca21/calmodulin No effect C6-2B cell lineNCB-20 cell lineCardiac preparations
[Ca21]i chelators Impaired C6-2B cell lineNCB-20 cell line
Bordetella Pertussis toxin No effect C6-2B cell lineNCB-20 cell lineCardiac preparationsRenal cell: CTAL
Gai Independent and cumulative effect with Gai-mediated process NCB-20 cell lineRenal cell: CTAL
Phorbol ester-sensitive PKCs No effect C6-2B cell lineNCB-20 cell lineRenal cell: CTAL
Results were obtained on C6-2B [82, 89] or NCB-20 [31, 90] cell lines, cultured cardiac myocytes [84], purified sarcolemmal vesicles [85] and corticalthick ascending limb (CTAL) microdissected from the rat kidney [64]. Antagonists of Ca21/calmodulin and Ca21 chelators have not been tested in CTALbut the usual inhibitions elicited by angiotensin II, bradykinin, and neomycin (Table 3) are no longer observed in Ca21-depleted segments (long pre-incubation time of tubule samples in Ca21-free medium).
the Ca21-sensing receptor RaKCaR elicits an inhibition as [64, 92] (Fig. 3). As discussed in the next section, this variabil-high as that induced by Gai-mediated processes [64]. ity is most likely to reflect a differential localisation of the
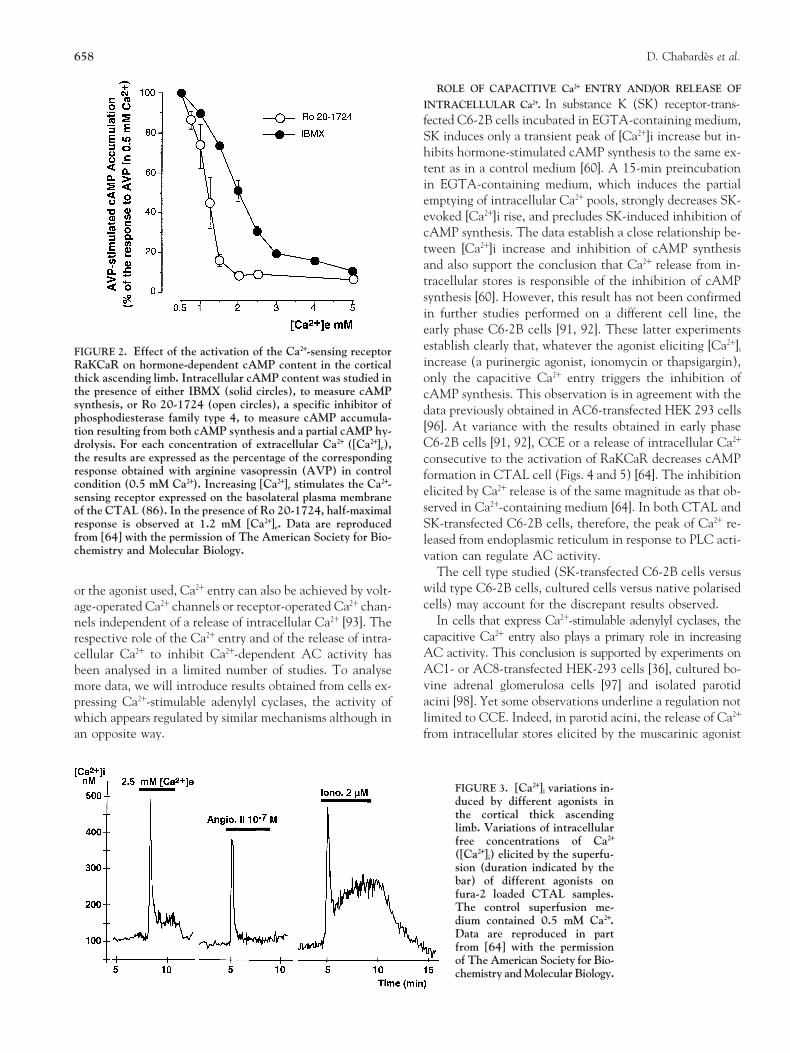
The same agonist induces variable inhibitions depending partners involved in the inhibition of cAMP synthesis, i.e.,on the cell type studied. For example, ionomycin (0.4 mM) adenylyl cyclase, release of intracellular Ca21 and/or influxis as potent as a purinergic agonist to increase [Ca21]i and to of extracellular Ca21 through Ca21 channels.inhibit slightly cAMP synthesis in C6-2B cell line [91]. Innative renal cells, 1 mM ionomycin inhibits strongly AVP-
III. Evidence Supporting thedependent cAMP synthesis in the OMCD (Table 3) [66].Colocalisation of Adenylyl Cyclase and of a Ca21
By contrast in the CTAL, a similar concentration has no ef-Entry Pathway and/or of a Release of Intracellular Ca21
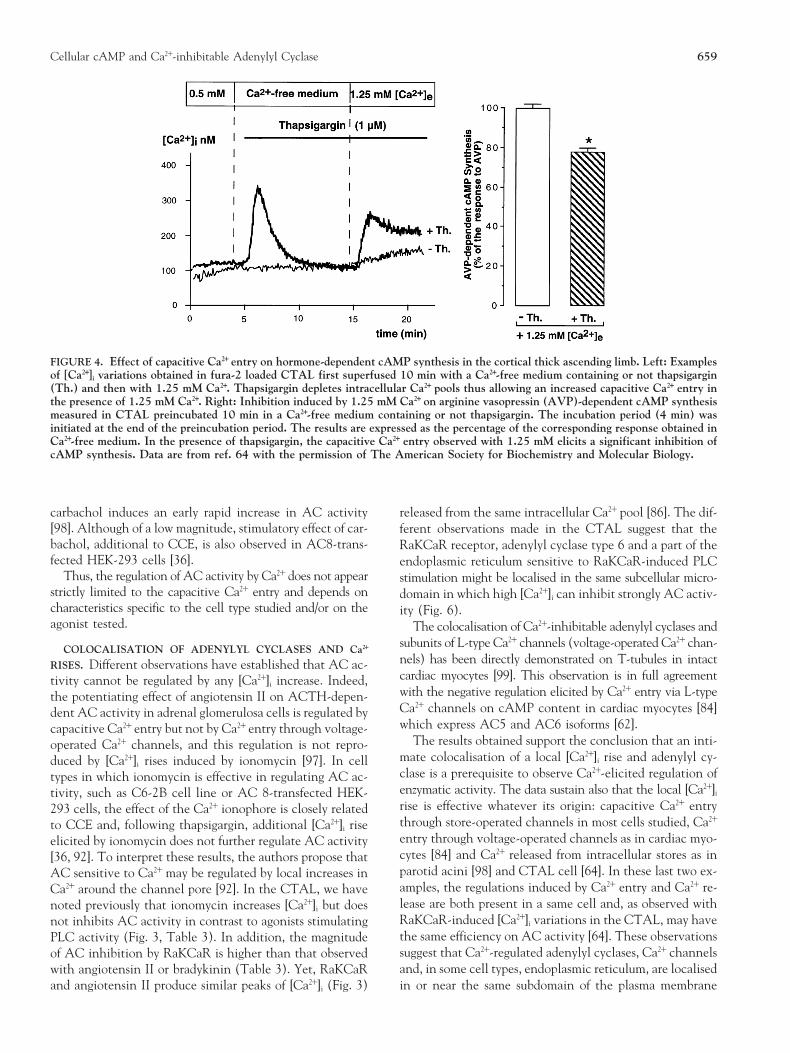
fect on AC activity (Table 3) although this agent induces aThe emptying of intracellular Ca21 stores by natural ago-high [Ca21]i rise (Fig. 3). In the same cell type, a variable effi-nists releasing Ca21 or by pharmacological agents (such asciency of inhibition of cAMP synthesis is also noted as a func-thapsigargin, an inhibitor of the endoplasmic reticular Ca21tion of the agonist tested (Table 5). This variability cannot bepump) switches on plasma membrane capacitive Ca21 entryexplained only by the pattern of [Ca21]i rise or by the magni-
tude of [Ca21]i increases which can be measured on whole cells (CCE) channels [93–95]. Depending on the cell type and/
TABLE 5. Role of PDEs in Ca21-mediated inhibition of cAMP accumulation
Agonists used Inhibition of cAMP synthesis Inhibition of cAMP accumulationto increase [Ca21]i (%) (%)
C6-2B cell line [91]Pre-treatment by thapsigargin 29 79Pre-treatment by purinergic agonist 16 33
Artery endothelial cell [83]Bradykinin 12 34
CTAL cell [64]Angiotensin II 20 46Bradykinin 18 471.5 mM [Ca21]e 35 732.5 mM [Ca21]e 65 91
OMCD, AVP-sensitive cell [76]Prostaglandin E2 12a 53
Inhibitions were obtained in intact cells studied in the presence (cAMP accumulation) or not (cAMP synthesis) of phosphodiesterase activities. Thevalues indicated are the percentages of inhibition calculated from the corresponding control. In C6-2B cells, the inhibitions were observed in experimentalprotocols studying the effect induced by a capacitive Ca21 entry. The results are from the references given in brackets.
a As commented in the legend of Table 3, this effect was not statistically significant; PGE2-mediated inhibition of AVP-dependent cAMP accumulationis mainly due to cAMP hydrolysis which represents more than 85% of the inhibition observed [76].
658 D. Chabardes et al.
ROLE OF CAPACITIVE Ca21 ENTRY AND/OR RELEASE OF
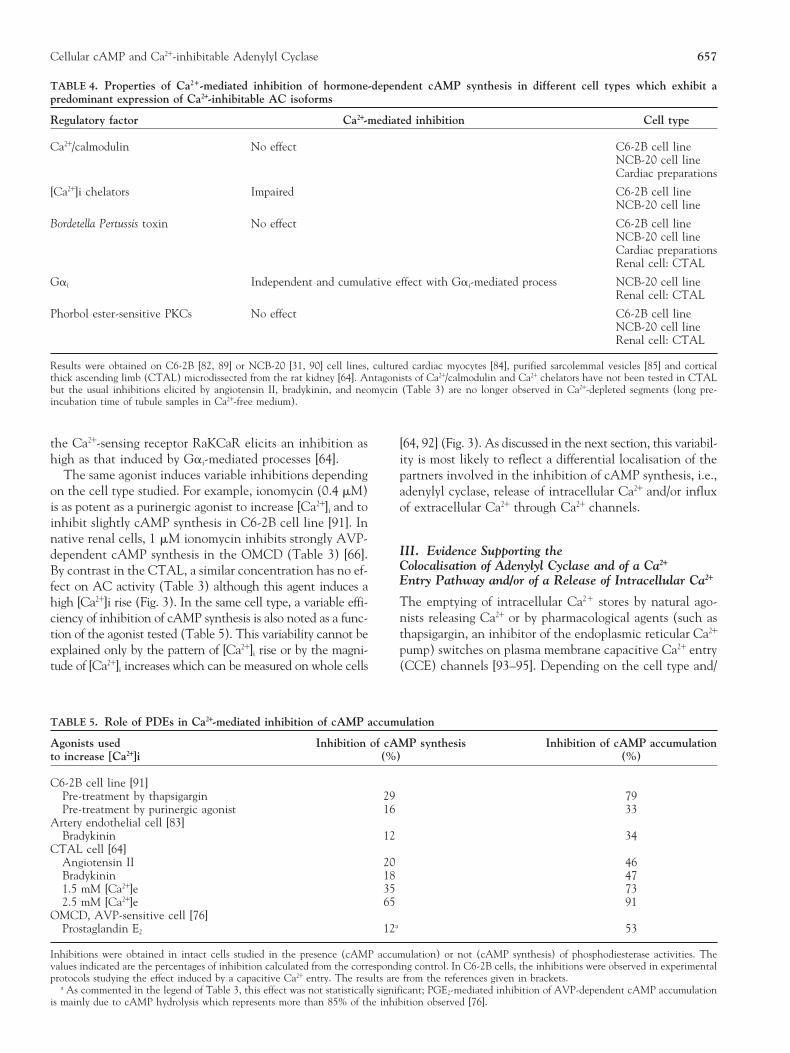
INTRACELLULAR Ca21. In substance K (SK) receptor-trans-fected C6-2B cells incubated in EGTA-containing medium,SK induces only a transient peak of [Ca21]i increase but in-hibits hormone-stimulated cAMP synthesis to the same ex-tent as in a control medium [60]. A 15-min preincubationin EGTA-containing medium, which induces the partialemptying of intracellular Ca21 pools, strongly decreases SK-evoked [Ca21]i rise, and precludes SK-induced inhibition ofcAMP synthesis. The data establish a close relationship be-tween [Ca21]i increase and inhibition of cAMP synthesisand also support the conclusion that Ca21 release from in-tracellular stores is responsible of the inhibition of cAMPsynthesis [60]. However, this result has not been confirmedin further studies performed on a different cell line, theearly phase C6-2B cells [91, 92]. These latter experimentsestablish clearly that, whatever the agonist eliciting [Ca21]iFIGURE 2. Effect of the activation of the Ca21-sensing receptorincrease (a purinergic agonist, ionomycin or thapsigargin),RaKCaR on hormone-dependent cAMP content in the cortical
thick ascending limb. Intracellular cAMP content was studied in only the capacitive Ca21 entry triggers the inhibition ofthe presence of either IBMX (solid circles), to measure cAMP cAMP synthesis. This observation is in agreement with thesynthesis, or Ro 20-1724 (open circles), a specific inhibitor of data previously obtained in AC6-transfected HEK 293 cellsphosphodiesterase family type 4, to measure cAMP accumula-
[96]. At variance with the results obtained in early phasetion resulting from both cAMP synthesis and a partial cAMP hy-C6-2B cells [91, 92], CCE or a release of intracellular Ca21drolysis. For each concentration of extracellular Ca21 ([Ca21]e),
the results are expressed as the percentage of the corresponding consecutive to the activation of RaKCaR decreases cAMPresponse obtained with arginine vasopressin (AVP) in control formation in CTAL cell (Figs. 4 and 5) [64]. The inhibitioncondition (0.5 mM Ca21). Increasing [Ca21]e stimulates the Ca21- elicited by Ca21 release is of the same magnitude as that ob-sensing receptor expressed on the basolateral plasma membrane
served in Ca21-containing medium [64]. In both CTAL andof the CTAL (86). In the presence of Ro 20-1724, half-maximalresponse is observed at 1.2 mM [Ca21]e. Data are reproduced SK-transfected C6-2B cells, therefore, the peak of Ca21 re-from [64] with the permission of The American Society for Bio- leased from endoplasmic reticulum in response to PLC acti-chemistry and Molecular Biology. vation can regulate AC activity.
The cell type studied (SK-transfected C6-2B cells versuswild type C6-2B cells, cultured cells versus native polarisedor the agonist used, Ca21 entry can also be achieved by volt-cells) may account for the discrepant results observed.age-operated Ca21 channels or receptor-operated Ca21 chan-
In cells that express Ca21-stimulable adenylyl cyclases, thenels independent of a release of intracellular Ca21 [93]. Thecapacitive Ca21 entry also plays a primary role in increasingrespective role of the Ca21 entry and of the release of intra-AC activity. This conclusion is supported by experiments oncellular Ca21 to inhibit Ca21-dependent AC activity hasAC1- or AC8-transfected HEK-293 cells [36], cultured bo-been analysed in a limited number of studies. To analysevine adrenal glomerulosa cells [97] and isolated parotidmore data, we will introduce results obtained from cells ex-acini [98]. Yet some observations underline a regulation notpressing Ca21-stimulable adenylyl cyclases, the activity oflimited to CCE. Indeed, in parotid acini, the release of Ca21which appears regulated by similar mechanisms although in
an opposite way. from intracellular stores elicited by the muscarinic agonist
FIGURE 3. [Ca21]i variations in-duced by different agonists inthe cortical thick ascendinglimb. Variations of intracellularfree concentrations of Ca21
([Ca21]i) elicited by the superfu-sion (duration indicated by thebar) of different agonists onfura-2 loaded CTAL samples.The control superfusion me-dium contained 0.5 mM Ca21.Data are reproduced in partfrom [64] with the permissionof The American Society for Bio-chemistry and Molecular Biology.
Cellular cAMP and Ca21-inhibitable Adenylyl Cyclase 659
FIGURE 4. Effect of capacitive Ca21 entry on hormone-dependent cAMP synthesis in the cortical thick ascending limb. Left: Examplesof [Ca21]i variations obtained in fura-2 loaded CTAL first superfused 10 min with a Ca21-free medium containing or not thapsigargin(Th.) and then with 1.25 mM Ca21. Thapsigargin depletes intracellular Ca21 pools thus allowing an increased capacitive Ca21 entry inthe presence of 1.25 mM Ca21. Right: Inhibition induced by 1.25 mM Ca21 on arginine vasopressin (AVP)-dependent cAMP synthesismeasured in CTAL preincubated 10 min in a Ca21-free medium containing or not thapsigargin. The incubation period (4 min) wasinitiated at the end of the preincubation period. The results are expressed as the percentage of the corresponding response obtained inCa21-free medium. In the presence of thapsigargin, the capacitive Ca21 entry observed with 1.25 mM elicits a significant inhibition ofcAMP synthesis. Data are from ref. 64 with the permission of The American Society for Biochemistry and Molecular Biology.
carbachol induces an early rapid increase in AC activity released from the same intracellular Ca21 pool [86]. The dif-[98]. Although of a low magnitude, stimulatory effect of car- ferent observations made in the CTAL suggest that thebachol, additional to CCE, is also observed in AC8-trans- RaKCaR receptor, adenylyl cyclase type 6 and a part of thefected HEK-293 cells [36]. endoplasmic reticulum sensitive to RaKCaR-induced PLC
Thus, the regulation of AC activity by Ca21 does not appear stimulation might be localised in the same subcellular micro-strictly limited to the capacitive Ca21 entry and depends on domain in which high [Ca21]i can inhibit strongly AC activ-characteristics specific to the cell type studied and/or on the ity (Fig. 6).agonist tested. The colocalisation of Ca21-inhibitable adenylyl cyclases and
subunits of L-type Ca21 channels (voltage-operated Ca21 chan-COLOCALISATION OF ADENYLYL CYCLASES AND Ca21
nels) has been directly demonstrated on T-tubules in intactRISES. Different observations have established that AC ac-cardiac myocytes [99]. This observation is in full agreementtivity cannot be regulated by any [Ca21]i increase. Indeed,with the negative regulation elicited by Ca21 entry via L-typethe potentiating effect of angiotensin II on ACTH-depen-Ca21 channels on cAMP content in cardiac myocytes [84]dent AC activity in adrenal glomerulosa cells is regulated bywhich express AC5 and AC6 isoforms [62].capacitive Ca21 entry but not by Ca21 entry through voltage-
The results obtained support the conclusion that an inti-operated Ca21 channels, and this regulation is not repro-mate colocalisation of a local [Ca21]i rise and adenylyl cy-duced by [Ca21]i rises induced by ionomycin [97]. In cellclase is a prerequisite to observe Ca21-elicited regulation oftypes in which ionomycin is effective in regulating AC ac-enzymatic activity. The data sustain also that the local [Ca21]itivity, such as C6-2B cell line or AC 8-transfected HEK-rise is effective whatever its origin: capacitive Ca21 entry293 cells, the effect of the Ca21 ionophore is closely relatedthrough store-operated channels in most cells studied, Ca21to CCE and, following thapsigargin, additional [Ca21]i riseentry through voltage-operated channels as in cardiac myo-elicited by ionomycin does not further regulate AC activitycytes [84] and Ca21 released from intracellular stores as in[36, 92]. To interpret these results, the authors propose thatparotid acini [98] and CTAL cell [64]. In these last two ex-AC sensitive to Ca21 may be regulated by local increases inamples, the regulations induced by Ca21 entry and Ca21 re-Ca21 around the channel pore [92]. In the CTAL, we havelease are both present in a same cell and, as observed withnoted previously that ionomycin increases [Ca21]i but doesRaKCaR-induced [Ca21]i variations in the CTAL, may havenot inhibits AC activity in contrast to agonists stimulatingthe same efficiency on AC activity [64]. These observationsPLC activity (Fig. 3, Table 3). In addition, the magnitudesuggest that Ca21-regulated adenylyl cyclases, Ca21 channelsof AC inhibition by RaKCaR is higher than that observedand, in some cell types, endoplasmic reticulum, are localisedwith angiotensin II or bradykinin (Table 3). Yet, RaKCaR
and angiotensin II produce similar peaks of [Ca21]i (Fig. 3) in or near the same subdomain of the plasma membrane
660 D. Chabardes et al.
FIGURE 5. Effect of intracellular Ca21 release on hormone-dependent cAMP synthesis in the cortical thick ascending limb. Upper pan-els: Examples of [Ca21]i variations elicited by different concentrations of neomycin, an agonist of the Ca21-sensing receptor RaKCaR,in either control (0.5 mM Ca21) or after a 10-min superfusion with a Ca21-free medium. Lower panels: Corresponding inhibitions elicitedby neomycin on arginine vasopressin (AVP)-dependent cAMP synthesis measured after a 10 min preincubation in control or Ca21-freemedium. Note that both Ca21 peaks and inhibition values were lower after a 10-min preincubation in a Ca21-free medium. This wasdue to the partial emptying of intracellular Ca21 pools in this experimental condition. When the preincubation period was of only 2min, Ca21 peaks and inhibition values were comparable to those observed in control tubule samples (preincubation and incubation in0.5 mM Ca21). Data are from [64] with the permission of The American Society for Biochemistry and Molecular Biology.
which may optimise the efficiency of local Ca21 increase to ferent transduction pathways. AC5 and AC6 isoforms are ex-regulate AC activity. pressed widely in different tissues [1–3] and the regulations
reported here may be potentially observed in numerous cells.However, as emphasized by the different results reported
CONCLUSION here, their sensitivity to inhibition by Ca21 is highly variablefrom one cell type to another. This variability depends, atType 5 and type 6 adenylyl cyclases exhibit similar enzymaticleast partly, on intrinsic properties linked to the cell typeproperties and characteristics of regulation, except the stimula-studied and on the agonist eliciting [Ca21]i increase. Differ-tory effect of PKCs described only with AC5. In cells whichent observations suggest that AC activity is regulated onlyexpress these isoforms, the cAMP content is strongly regu-by local [Ca21]i increases produced in the vicinity of ade-lated, either positively by agonist-mediated Gas activation,nylyl cyclase itself. In cardiac myocytes [99] and CTAL [64],or negatively by Gai-mediated processes and [Ca21]i in-an intimate colocalisation of Ca21-inhibitable AC5 and/orcreases consecutive to PLC activation or voltage-operatedAC6 isoforms and [Ca21]i rise appears essential to obtain achannels. The negative pathways integrate two different andstrong inhibition of cAMP synthesis and thus of cAMP con-independent mechanisms regulating the same enzymatic ac-
tivity and thus allow combined inhibitions resulting from dif- tent [64, 84]. This strong inhibition might be particularly well
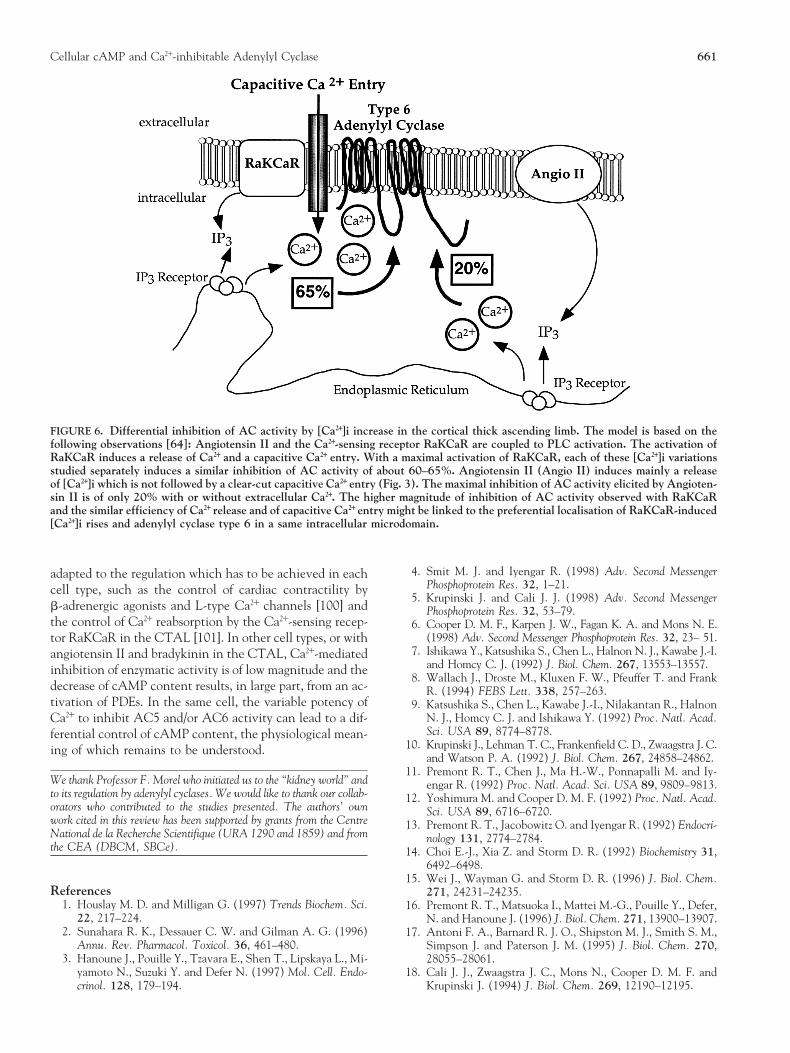
Cellular cAMP and Ca21-inhibitable Adenylyl Cyclase 661
FIGURE 6. Differential inhibition of AC activity by [Ca21]i increase in the cortical thick ascending limb. The model is based on thefollowing observations [64]: Angiotensin II and the Ca21-sensing receptor RaKCaR are coupled to PLC activation. The activation ofRaKCaR induces a release of Ca21 and a capacitive Ca21 entry. With a maximal activation of RaKCaR, each of these [Ca21]i variationsstudied separately induces a similar inhibition of AC activity of about 60–65%. Angiotensin II (Angio II) induces mainly a releaseof [Ca21]i which is not followed by a clear-cut capacitive Ca21 entry (Fig. 3). The maximal inhibition of AC activity elicited by Angioten-sin II is of only 20% with or without extracellular Ca21. The higher magnitude of inhibition of AC activity observed with RaKCaRand the similar efficiency of Ca21 release and of capacitive Ca21 entry might be linked to the preferential localisation of RaKCaR-induced[Ca21]i rises and adenylyl cyclase type 6 in a same intracellular microdomain.
4. Smit M. J. and Iyengar R. (1998) Adv. Second Messengeradapted to the regulation which has to be achieved in eachPhosphoprotein Res. 32, 1–21.cell type, such as the control of cardiac contractility by
5. Krupinski J. and Cali J. J. (1998) Adv. Second Messengerb-adrenergic agonists and L-type Ca21 channels [100] and Phosphoprotein Res. 32, 53–79.the control of Ca21 reabsorption by the Ca21-sensing recep- 6. Cooper D. M. F., Karpen J. W., Fagan K. A. and Mons N. E.
(1998) Adv. Second Messenger Phosphoprotein Res. 32, 23– 51.tor RaKCaR in the CTAL [101]. In other cell types, or with7. Ishikawa Y., Katsushika S., Chen L., Halnon N. J., Kawabe J.-I.angiotensin II and bradykinin in the CTAL, Ca21-mediated
and Homcy C. J. (1992) J. Biol. Chem. 267, 13553–13557.inhibition of enzymatic activity is of low magnitude and the8. Wallach J., Droste M., Kluxen F. W., Pfeuffer T. and Frank
decrease of cAMP content results, in large part, from an ac- R. (1994) FEBS Lett. 338, 257–263.tivation of PDEs. In the same cell, the variable potency of 9. Katsushika S., Chen L., Kawabe J.-I., Nilakantan R., Halnon
N. J., Homcy C. J. and Ishikawa Y. (1992) Proc. Natl. Acad.Ca21 to inhibit AC5 and/or AC6 activity can lead to a dif-Sci. USA 89, 8774–8778.ferential control of cAMP content, the physiological mean-
10. Krupinski J., Lehman T. C., Frankenfield C. D., Zwaagstra J. C.ing of which remains to be understood.and Watson P. A. (1992) J. Biol. Chem. 267, 24858–24862.
11. Premont R. T., Chen J., Ma H.-W., Ponnapalli M. and Iy-We thank Professor F. Morel who initiated us to the “kidney world” and engar R. (1992) Proc. Natl. Acad. Sci. USA 89, 9809–9813.to its regulation by adenylyl cyclases. We would like to thank our collab- 12. Yoshimura M. and Cooper D. M. F. (1992) Proc. Natl. Acad.orators who contributed to the studies presented. The authors’ own Sci. USA 89, 6716–6720.work cited in this review has been supported by grants from the Centre 13. Premont R. T., Jacobowitz O. and Iyengar R. (1992) Endocri-National de la Recherche Scientifique (URA 1290 and 1859) and from nology 131, 2774–2784.the CEA (DBCM, SBCe). 14. Choi E.-J., Xia Z. and Storm D. R. (1992) Biochemistry 31,
6492–6498.15. Wei J., Wayman G. and Storm D. R. (1996) J. Biol. Chem.
References 271, 24231–24235.1. Houslay M. D. and Milligan G. (1997) Trends Biochem. Sci. 16. Premont R. T., Matsuoka I., Mattei M.-G., Pouille Y., Defer,
22, 217–224. N. and Hanoune J. (1996) J. Biol. Chem. 271, 13900–13907.2. Sunahara R. K., Dessauer C. W. and Gilman A. G. (1996) 17. Antoni F. A., Barnard R. J. O., Shipston M. J., Smith S. M.,
Annu. Rev. Pharmacol. Toxicol. 36, 461–480. Simpson J. and Paterson J. M. (1995) J. Biol. Chem. 270,3. Hanoune J., Pouille Y., Tzavara E., Shen T., Lipskaya L., Mi- 28055–28061.
yamoto N., Suzuki Y. and Defer N. (1997) Mol. Cell. Endo- 18. Cali J. J., Zwaagstra J. C., Mons N., Cooper D. M. F. andKrupinski J. (1994) J. Biol. Chem. 269, 12190–12195.crinol. 128, 179–194.

662 D. Chabardes et al.
19. Choi E.-J., Wong S. T., Hinds T. R. and Storm D. R. (1992) 51. Lai H.-L., Yang T.-H., Messing R. O., Ching Y.-H., LinS.-C., and Chern Y. (1997) J. Biol. Chem. 272, 4970–4977.J. Biol. Chem. 267, 12440–12442.
52. Kawabe J.-I., Iwami G., Ebina T., Ohno S., Katada T., Ueda20. Nielsen M. D., Chan G. C. K., Poser S. W. and Storm D. R.Y., Homcy C. J. and Ishikawa Y. (1994) J. Biol. Chem. 269,(1996) J. Biol. Chem. 271, 33308–33316.16554–16558.21. Marjamaki A., Sato M., Bouet-Alard R., Yang Q., Limon-
53. Kawabe J.-I., Ebina T., Toya Y., Oka N., Schwencke C.,Boulez I., Legrand C. and Lanier S. M. (1997) J. Biol. Chem.Duzic E. and Ishikawa Y. (1996) FEBS Lett. 384, 273–276.272, 16466–16473.
54. Mendez R., Kollmorgen G., White M. F. and Rhoads R. E.22. Choi E.-J., Wong S. T., Dittman A. H. and Storm D. R.(1997) Mol. Cell. Biol. 17, 5184–5192.(1993) Biochemistry 32, 1891–1894.
55. Chen Z., Nield, H. S., Sun H., Barbier A. and Patel T. B.23. Zimmermann G. and Taussig R. (1996) J. Biol. Chem. 271,(1995) J. Biol. Chem. 270, 27525–27530.27161–27166.
56. Mons N., and Cooper D. M. F. (1994) J. Neuroendocrinol. 6,24. Tang W.-J. and Gilman A. G. (1991) Science 254, 1500–1503.665–671.25. Gao B. and Gilman A. G. (1991) Proc. Natl. Acad. Sci. USA
57. Hellevuo K., Hoffman P. L. and Tabakoff B. (1996) J. Neuro-88, 10178–10182.chem. 67, 177–185.26. Watson P. A., Krupinski J., Kempinski A. M. and Franken-
58. Paulssen R. H., Johansen P. W., Gordeladze J. O., Nymoenfield C. D. (1994) J. Biol. Chem. 269, 28893–28898.O., Paulssen E. J. and Gautvik K. M. (1994) Eur. J. Biochem.27. Federman A. D., Conklin B. R., Schrader K. A., Reed R. R.222, 97–103.and Bourne H. R. (1992) Nature 356, 159–161.
59. Manolopoulos V. G., Liu J., Unsworth B. R. and Lelkes P. I.28. Lustig K. D., Conklin B. R., Herzmark P., Taussig R. and(1995) Biochem. Biophys. Res. Commun. 208, 323–331.Bourne H. R. (1993) J. Biol. Chem. 268, 13900–13905.
60. DeBernardi M. A., Munshi R., Yoshimura M., Cooper D. M. F.29. Pieroni J. P., Harry A., Chen J., Jacobowitz O., Magnusson R. P.and Brooker G. (1993) Biochem. J. 293, 325–328.and Iyengar R. (1995) J. Biol. Chem. 270, 21368–21373.
61. Morel F. and Doucet A. (1986) Physiological Reviews 66,30. Harry A., Chen Y., Magnusson R., Iyengar R. and Weng G.377–468.(1997) J. Biol. Chem. 272, 19017–19021.
62. Yu H. J., Unnerstall J. R. and Green R. D. (1995) FEBS Let-31. Boyajian C. L., Garritsen A. and Cooper D. M. F. (1991) J.ters 374, 89–94.Biol. Chem. 266, 4995–5003.
63. Morel F., Imbert-Teboul M. and Chabardes D. (1987) Kidney32. Wu Z., Wong S. T. and Storm D. R. (1993) J. Biol. Chem.Inter. 31, 512–520.268, 23766–23768.
64. De Jesus Ferreira M. C., Helies-Toussaint C., Imbert-Teboul33. Feinstein P. G., Schrader K. A., Bakalyar H. A., Tang W.-J., M., Bailly C., Verbavatz J.-M., Bellanger A.-C. and Cha-Krupinski J., Gilman A. G. and Reed R. R. (1991) Proc. bardes D. (1998) J. Biol. Chem. 273, 15192–15202.Natl. Acad. Sci. USA 88, 10173–10177. 65. Knepper M. A., Nielsen S., Chou C.-L. and DiGiovanni S.34. Scholich K., Barbier A. J., Mullenix J. B. and Patel T. B. R. (1994) Seminars in Nephrology 14, 302–321.
(1997) Proc. Natl. Acad. Sci. USA 94, 2915–2920. 66. Chabardes D., Firsov D., Aarab L., Clabecq A., Bellanger A.-C.,35. Sutkowski E. M., Tang W.-J., Broome C. W., Robbins J. D. Siaume-Perez S. and Elalouf J.-M. (1996) J. Biol. Chem. 271,
and Seamon K. B. (1994) Biochemistry 33, 12852–12859. 19264–19271.36. Fagan K. A., Mahey R. and Cooper D. M. F. (1996) J. Biol. 67. Elalouf J.-M., Buhler J.-M., Tessiot C., Bellanger A.-C.,
Chem. 271, 12438–12444. Dublineau I. and Rouffignac C. de (1993) J. Clin. Invest. 91,37. Taussig R., Iniguez-Lluhi J. A. and Gilman A. G. (1993) Sci- 264–272.
ence 261, 218–221. 68. Shen T., Suzuki Y., Poyard M., Miyamoto N., Defer N. and38. Taussig R., Tang W.-J., Hepler J. R. and Gilman A. G. Hanoune J. (1997) Am. J. Physiol. 273, C323–C330.
(1994) J. Biol. Chem. 269, 6093–6100. 69. Verbavatz J.-M., Helies C., Aarab L., Gasc J.-M. and Cha-39. Chen J. and Iyengar R. (1993) J. Biol. Chem. 268, 12253–12256. bardes D. (1998) J. Amer. Soc. Nephrol. 9, 432A.40. Wong Y. H., Conklin B. R. and Bourne H. R. (1992) Science 70. Aarab L., Siaume-Perez S. and Chabardes D. (1999) Br. J.
255, 339–342. Pharmacol. 126, 1041–1049.41. Kozasa T. and Gilman A. G. (1995) J. Biol. Chem. 270, 71. Firsov D., Mandon B., Morel A., Merot J., Le Maout S., Bel-
1734–1741. langer A.-C., Rouffignac C. de, Elalouf J.-M. and Buhler J.-M.42. Yoshimura M., Ikeda H. and Tabakoff B. (1996) Mol. Phar- (1994) Pfluger’s Arch. Eur. J. Physiol. 429, 79–89.
macol. 50, 43–51. 72. Firsov D., Bellanger A.-C., Marsy S. and Elalouf J.-M. (1995)43. Bayewitch M. L., Avidor-Reiss T., Levy R., Pfeuffer T., Nevo I., Am. J. Physiol. 269, F702–F709.
Simonds W. F. and Vogel Z. (1998) FASEB J. 12, 1019–1025. 73. Chabardes D., Montegut M., Imbert-Teboul M. and Morel F.44. Bayewitch M. L., Avidor-Reiss T., Levy R., Pfeuffer T., Nevo (1984) Mol. Cell. Endocrinol. 37, 263–275.
I., Simonds W. F. and Vogel Z. (1998) J. Biol. Chem. 273, 74. Hoey M. and Houslay M. D. (1990) Biochem. Pharmacol. 40,2273–2276. 193–202.
45. Johnson R. A., Desaubry L., Bianchi G., Shoshani I., Lyons E., 75. Torikai S. and Kurokawa K. (1983) Am. J. Physiol. 245,Jr., Taussig R., Watson P. A., Cali J. J., Krupinski J., Pieroni J. F58–F66.P. and Iyengar R. (1997) J. Biol. Chem. 272, 8962–8966. 76. Chabardes D., Montegut M., Zhou Y. and Siaume-Perez S.
46. Premont R. T. and Iyengar R. (1988) J. Biol. Chem. 263, (1990) Mol. Cell. Endocrinol. 73, 111–121.16087–16095. 77. Aarab L., Montegut M., Siaume-Perez S., Imbert-Teboul M.
47. Iwami G., Kawabe J.-I., Ebina T., Cannon P. J., Homcy C. and Chabardes D. (1993) Pfluger’s Arch. 423, 397–405.J. and Ishikawa Y. (1995) J. Biol. Chem. 270, 12481–12484. 78. Murayama N., Werness J. L., Kusano E., Christensen S. and
48. Chen Y., Harry A., Li J., Smit M. J., Bai X., Magnusson R., Dousa T. P. (1983) J. Cyclic Nucleotide Protein Phosphor. Res.Pieroni J. P., Weng G. and Iyengar R. (1997) Proc. Natl. 9, 427–433.Acad. Sci. USA 94, 14100–14104. 79. Dublineau I., Pradelles P., Rouffignac C. de and Elalouf J.-M.
49. Yoshimura M. and Cooper D. M. F. (1993) J. Biol. Chem. (1990) Proc. Natl. Acad. Sci. USA 87, 7583–7587.268, 4604–4607. 80. Firsov D., Aarab L., Mandon B., Siaume-Perez S., Rouffignac
50. Jacobowitz O., Chen J., Premont R. T. and Iyengar R. (1993) C. de and Chabardes D. (1995) Pfluger’s Arch. Eur. J. Phys-iol. 429, 636–646.J. Biol. Chem. 268, 3829–3832.

Cellular cAMP and Ca21-inhibitable Adenylyl Cyclase 663
81. Aarab L., Siaume-Perez S. and Chabardes D. (1993) Pfluger’s 91. Chiono M., Mahey R., Tate G. and Cooper D. M. F. (1995)J. Biol. Chem. 270, 1149–1155.Arch. 425, 417–425.
92. Fagan K. A., Mons N. and Cooper D. M. F. (1998) J. Biol.82. DeBernardi M. A., Seki T. and Brooker G. (1991) Proc.Chem. 273, 9297–9305.Natl. Acad. Sci. USA 88, 9257–9261.
93. Fasolato C., Innocenti B. and Pozzan T. (1994) Trends Phar-83. Stevens T., Nakahashi Y., Cornfield D. N., McMurtry I. F.,macol. Sci. 15, 77–83.Cooper D. M. F. and Rodman D. M. (1995) Proc. Natl.
94. Berridge M. J. (1995) Biochem. J. 312, 1–11.Acad. Sci. USA 92, 2696–2700.95. Birnbaumer L., Zhu X., Jiang M., Boulay G., Peyton M.,84. Yu H.J., Ma H. and Green R. D. (1993) Mol. Pharmacol. 44,
Vannier B., Brown D., Platano D., Sadeghi H., Stefani E.689–693.and Birnbaumer M. (1996) Proc. Natl. Acad. Sci. USA 93,85. Colvin R. A., Oibo J. A. and Allen R. A. (1991) Cell Cal-15195–15202.cium 12, 19–27.
96. Cooper D. M. F., Yoshimura M., Zhang Y., Chiono M. and86. Champigneulle A., Siga E., Vassent G. and Imbert-Teboul M. Mahey R. (1994) Biochem. J. 297, 437–440.(1997) J. Membrane Biol. 156, 117–129. 97. Burnay M. M., Vallotton M. B., Capponi A. M. and Rossier
87. Riccardi D., Lee W.-S., Lee K., Segre G. V., Brown E. M. and M. F. (1998) Biochem. J. 330, 21–27.Hebert S. C. (1996) Am. J. Physiol. 271, F951–F956. 98. Watson E. L., Wu Z., Jacobson K. L., Storm D. R., Singh
88. Kusano E., Murayama N., Werness J. L., Christensen S., J. C. and Ott S. M. (1998) Am. J. Physiol. 274, C557–C565.Homma S., Yusufi A. N. K. and Dousa T. P. (1985) Am. J. 99. Gao T., Puri T. S., Gerhardstein B. L., Chien A. J., Green R. D.Physiol. 249, F956–F966. and Hosey M. M. (1997) J. Biol. Chem. 272, 19401–19407.
89. DeBernardi M. A., Munshi R. and Brooker G. (1993) Mol. 100. Cooper D. M. F. and Brooker G. (1993) Trends Pharmacol.Pharmacol. 43, 451–458. Sci. 14, 34–36.
90. Garritsen A., Zhang Y., Firestone J. A., Browning M. D. and 101. Brown E. M. and Hebert S. C. (1995) J. Am. Soc. Nephrol.6, 1530–1540.Cooper D. M. F. (1992) J. Neurochem. 59, 1630–1639.