fight or flight: an investigation of aggressive behavior and predator avoidance in two populations...
TRANSCRIPT

PRIMARY RESEARCH PAPER
Fight or flight: an investigation of aggressive behaviorand predator avoidance in two populations of blue crabs(Callinectes sapidus Rathbun) in New Jersey
Jessica M. Reichmuth • James MacDonald •
Jonathan Ramirez • Judith S. Weis
Received: 14 April 2010 / Revised: 13 July 2010 / Accepted: 30 August 2010
� Springer Science+Business Media B.V. 2010
Abstract Recent literature has suggested aggres-
sion may be context dependent. The purpose of this
investigation was to examine aggressive and predator
avoidance behaviors in juvenile blue crabs of two
populations. Furthermore, we wanted to determine
whether aggression persisted into the adult stages.
Juvenile blue crabs collected from an impacted
estuary, the Hackensack Meadowlands (HM), were
found to attack a threatening stimulus significantly
more often (70%) than conspecifics from a less
impacted estuary (Tuckerton—TK). TK juveniles
responded significantly more often with a flight
(*35%) or mixed response (*30%). Additionally,
HM juveniles were significantly more successful than
TK juveniles at avoiding an adult blue crab predator
when sandy substrate was present in laboratory
experiments. However, the video clarity made it
impossible to determine which interactions were
allowing survival. To determine if ‘‘aggression’’
exhibited by the HM juveniles was the reason for
their enhanced survival, follow-up predator avoid-
ance experiments were conducted without substrate
and videotaped. The results of these experiments
suggest that aggression per se is not the reason since
aggressive juveniles were no more successful than
non-aggressive individuals. The aggressive behavior
exhibited by HM juveniles continues into the adult
stages. This behavior may be important to recognize
when estimating population size as well as local
fishery efforts.
Keywords Aggression � Behavior � Blue crab �Callinectes sapidus � Predator avoidance
Introduction
An organism’s propensity for aggression may affect
interactions between individuals. This propensity for
aggression may also help determine its foraging
success (e.g., subduing resisting prey and pressing
attacks) or defending other items such as mates or
refuge, from other individuals (Kaiser et al., 1990).
Aggression has been widely observed in many types
of invertebrates. For example, larger and more
aggressive wild octopuses (Abdopsus aculeatus
d’Orbigny) were more successful at gaining time to
successfully mate with females (Huffard et al., 2010).
Handling editor: L.B. Kats
J. M. Reichmuth � J. Ramirez � J. S. Weis
Department of Biological Sciences, Rutgers, The State
University of New Jersey, 195 University Avenue,
Newark, NJ 07102, USA
J. MacDonald
New York Sea Grant, c/o NYS DEC, 47-40 21st Street,
Long Island City, NY 11101, USA
J. M. Reichmuth (&)
Department of Biology, Augusta State University,
2500 Walton Way, Augusta, GA 30904, USA
e-mail: [email protected]
123
Hydrobiologia
DOI 10.1007/s10750-010-0460-z

Other examples of aggression have been observed
with reef-building corals (Romano, 1990), ants
(Vasquez & Silverman, 2008), sea urchins (Shulman,
1990), crayfish (Graham & Heberholz, 2009), and
crabs (Huntingford et al., 1995).
Crustaceans are good models for observing aggres-
sive and/or agonistic behavior for a number of
reasons: ease of accessibility, known physiology,
size, and social behavior (Kravitz & Huber, 2003).
Both adult and juvenile blue crabs are known to be
aggressive, especially toward conspecifics (Moksnes
et al., 1997; Clark et al., 1999a). Such encounters can
leave a crab injured or missing appendages. Because
of such an alteration, an individual can have a
presumed competitive and/or energetic disadvantage
as well as increased predation risk (Juanes & Smith,
1995). The blue crab’s most formidable defensive (or
offensive) weapons are its sharp, strong chelae. How
an individual uses these weapons may affect its
prospects for survival in an encounter with a predator
or conspecific competitor. Jachowski (1974) offered
the first experimental study of agonistic behavior by
C. sapidus. He suggested that agonistic encounters
with other individuals were highly dependent on the
presence of food or a potential mate. More recent
literature (e.g., Nye, 1989; Wolcott & Hines, 1989,
1990; Clark et al., 1999b) has focused on aggressive
behavior associated with foraging activities and has
shown agonistic activity occurring simultaneously
with periods of feeding. Not only did the presence of
conspecifics interfere with feeding and foraging, but
some adult crabs spent up to 40% of their time
engaged in agonistic activities (Clark et al., 1999a, b).
Studies using telemetry supported the idea that the
frequency and nature of interactions occurring
between C. sapidus conspecifics are dependent on
the density of crabs in a specific area at one time and,
as a result, have important implications for fisheries
management. A field study found an increase of
agonistic behavior in crabs when using certain kinds
of commercial crab pots (Vasquez-Archdale et al.,
2003). Since crab pots are sometimes used in
population estimates, it is possible that agonistic
behaviors may affect the likelihood of crabs entering
the pot and thus impact the count.
Aggressive behavior has been correlated with
boldness and reactive behavior, which has implica-
tions for predator avoidance behavior. Individuals
that are more reactive in the presence of a predator
potentially increase the chance of being eaten (Sih
et al., 2004a, b). However, other studies have
suggested that the increase of aggression/agonistic
behavior toward a predator can increase the chance of
survival (Whitehouse, 1997; Reany & Backwell,
2007). Juvenile C. sapidus normally avoid predation
by fleeing and/or by burying themselves in the
sediment. Some juveniles will even stand their
ground and use their chelae for defense, an aggressive
act that is more characteristic of larger adults.
Recent animal behavior research has focused on
differences among individuals in a species and
classifying the ‘‘behavioral syndrome’’ of a popula-
tion (Sih et al., 2004a, b). The most common of
these behavioral types are aggression and the shy-
bold regime, which have been investigated mostly in
vertebrates, such as fish (e.g., Coleman & Wilson,
1998; Ward et al., 2004). The behavior of brachyu-
ran crabs has also been widely studied (e.g., Hazlett,
1971, 1972; Jachowski, 1974; Crane, 1975), but not
necessarily in this context. Just as in other animal
populations, aggressive and defensive responses can
vary across a population as well as between
populations, and could impact predator avoidance
abilities. Recently, aggression and bold behavior
have been correlated with the success of an invasive
crayfish, Pacifcastacus leniusculus Dana (Pintor
et al., 2008). In another study, risk-taking behavior
predicted aggression and mating success in the
fiddler crab, Uca mjoebergi Rathbun (Reany &
Backwell, 2007).
The blue crab is a swimming decapod that is
estuarine dependent and widely distributed from
Nova Scotia to northern Argentina. They inhabit
estuaries and near shore coastal waters to depths of at
least 36 m and are a year-long resident of New Jersey
estuaries (Norse, 1977). This species is not only
important ecologically, but economically as well. The
Chesapeake Bay produces approximately 13,000 MT
of crabs a year, but in the northern Atlantic states the
crab provides access to a localized fishery (Jop et al.,
1997). Blue crabs are part of a predatory guild that
structures the dynamics of estuarine soft bottom
communities, which makes them important in the
estuarine food web. They are not only predators, but
also scavengers, and are themselves sometimes prey
for fishes and carnivorous crabs, especially as juve-
niles (Virnstein, 1979; Nelson, 1981; Edwards et al.,
1982; Hines et al., 1990).
Hydrobiologia
123
In this investigation of aggression, we compared
juveniles of two populations for their response to a
threatening stimulus and their predator avoidance
abilities. One population was from the Hackensack
Meadowlands (HM), an estuarine system with a long
history of impact, and the other from a less impacted
site in southeastern NJ, Tuckerton (TK). We hypoth-
esized that if one population of juvenile blue crabs
was more aggressive/reactive, this would give an
advantage with a predator. We also examined the
tendency of adult crabs from the same two systems to
enter crab pots. We expected that aggressive indi-
viduals, once inside a pot, would keep other crabs out
of the baited pot.
Materials and methods
Study sites
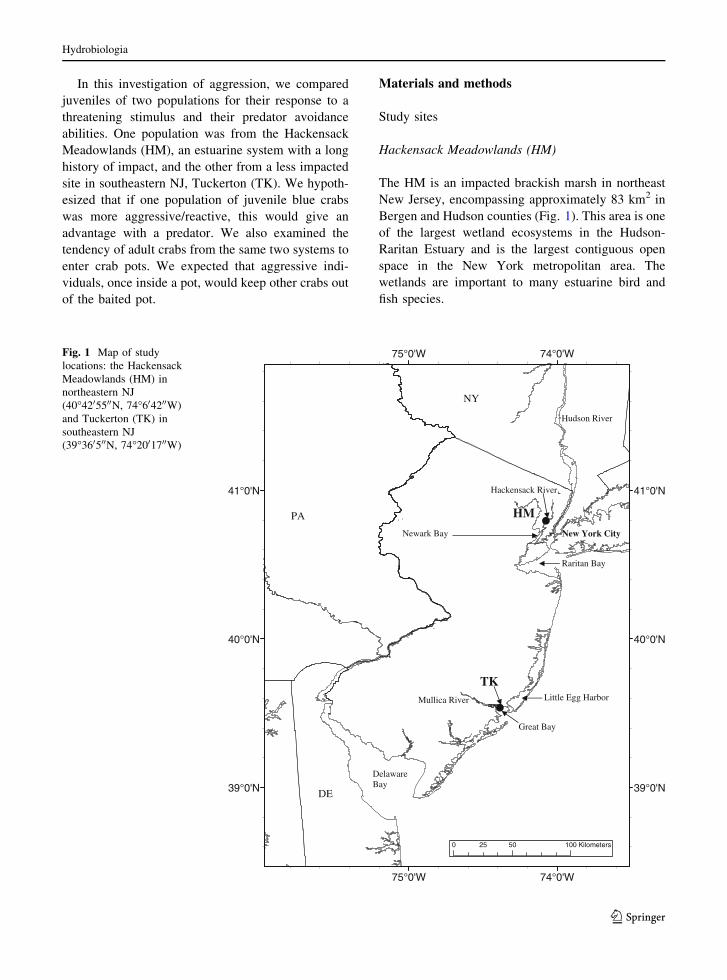
Hackensack Meadowlands (HM)
The HM is an impacted brackish marsh in northeast
New Jersey, encompassing approximately 83 km2 in
Bergen and Hudson counties (Fig. 1). This area is one
of the largest wetland ecosystems in the Hudson-
Raritan Estuary and is the largest contiguous open
space in the New York metropolitan area. The
wetlands are important to many estuarine bird and
fish species.
74°0'W
74°0'W
75°0'W
75°0'W
41°0'N 41°0'N
40°0'N 40°0'N
39°0'N 39°0'N
0 50 100 sretemoliK52
Newark Bay New York City
Hudson River
Great Bay
TKMullica River Little Egg Harbor
Raritan Bay
Hackensack River
HM
Delaware Bay
PA
DE
NY
Fig. 1 Map of study
locations: the Hackensack
Meadowlands (HM) in
northeastern NJ
(40�4205500N, 74�604200W)
and Tuckerton (TK) in
southeastern NJ
(39�360500N, 74�2001700W)
Hydrobiologia
123

Tuckerton (TK)
The Mullica River-Great Bay estuary comprised
225 km2 of salt marsh and 145 km2 of shallow
estuarine waters. The surrounding area is protected
by the Edwin B. Forsythe National Wildlife Refuge
and the Great Bay Wildlife Management Area
(Fig. 1). The system is less impacted and has been
designated as the Jacques Cousteau National Estua-
rine Research Reserve (NERR). TK is about 160 km
south of HM.
Experimental design
Crabs were collected from various sites within HM
and TK using a seine net and otter trawl and brought
back to the laboratory. They were kept in aerated
tanks with a sand depth of 1 cm and artificial
seawater (Instant Ocean�) at their native salinity
(HM: 15 psu; TK: 30 psu). A 14/10 light cycle was
kept throughout the field season. TK crabs were fed
with a diet of ribbed mussels (Geukensia demissa
Dillwyn) and Atlantic menhaden (Brevoortia tyran-
nus Latrobe) collected from TK, while HM crabs
were fed with a diet of menhaden and mummichogs
collected from HM; all crabs were fed three times a
week after which the water was changed. Intermolt
crabs were acclimated to laboratory conditions for
48 h prior to the beginning of experimentation.
Response to a threatening stimulus
Crabs [HM: mean carapace width, CW = 52 ±
4.8 mm; N = 41; TK: mean CW = 48 ± 5.3 mm
(SE), N = 58] were constrained inside a small,
inverted opaque container positioned. This container
was positioned at one end inside a 38-l aquarium with
a depth of 5-cm artificial sea water (salinity of
22 psu; no substrate). Each crab was allowed to
acclimate for 10 min. Substrate was not used so that
specific behaviors could be isolated. The aquarium
was covered on three sides with opaque, non-
reflective paper to restrict the crab’s peripheral vision
and prevent it from reacting to its own reflection or
movement outside the aquarium. The top two-thirds
of the remaining side were covered as well for the
same reason; the bottom third was left clear for
observation.
An individual crab, from either HM or TK, was
isolated in the opaque container. Once isolated, a
stimulus consisting of a black rubber stopper 44-mm
diameter attached to a dowel 54 cm in length was
slowly lowered into the other end of the aquarium.
After 10 min, the opaque container was slowly
removed. The stopper was slowly pushed toward
the crab by an investigator who stood beyond the
opaque side of the aquarium. No part of this
researcher ever appeared directly over the aquarium.
A second observer, stationed several feet back from
the aquarium to avoid provoking a reaction from the
crab, recorded the nature of the crab’s reaction.
Three response types were recorded. A ‘‘flee’’
reaction was defined as the crab quickly moving away
from the stopper, and ‘‘attack’’ was a lunge toward it.
A ‘‘mixed’’ response was a lunge toward the stopper
immediately followed by a quick retreat to the
opposite end of the tank. If the crab was facing away
from the stimulus, especially active, or moved
immediately across the aquarium before the stimulus
was activated, then the trial was discontinued. The
observer recording the reaction was blind to the
population of crab being tested. Statistical analyses
were conducted using Statistix 7.0 and GraphPad
Prism 4.0 software. Differences in the responses to
the stimulus were analyzed using v2 tests.
Survival with an adult blue crab predator
with substrate
Adult blue crab predators from these two sites were
highly variable in their time to capture juvenile blue
crabs (Reichmuth et al., 2009). To best determine the
exact vulnerability of the juveniles, we set up an
experiment to determine which juveniles were more
likely to be captured.
In this experimental set-up, one adult predator and
two juveniles from each population were used. The
experiment was conducted in a 76-l aquarium with
*35 mm sand covering the bottom and salinity of
22 psu, which is similar to field conditions. The
aquarium was covered with opaque paper on all four
sides so that the crabs did not get distracted by
outside movement or reflections. The four juveniles
(mean CW: HM = 32 ± 2.8 mm, N = 110; TK =
35 ± 2.6 mm (SE), N = 110) were added to the
aquarium and allowed to acclimate for 1 h; sizes and
sexes were noted and recorded. Swimming paddles
Hydrobiologia
123

on juvenile crabs from one population were marked
with a black Sharpie� in order to determine the
survivors; each population was marked in alternate
trials.
After the juvenile’s acclimation period, one adult
crab that had been food deprived for 48 h (male or
female, mean CW: HM = 101 ± 8.2 mm, TK = 94 ±
7.4 mm) was added. Adults were usually 80–100 mm
larger than the juveniles. Preliminary studies deter-
mined sex of the predator was not a factor in the
observed differences of survivorship. All trials were
recorded using a closed circuit digital camera
(Ikegami Tsushinki Co. Ltd) with a 7.5–75-mm lens
(Canon) and digital disk recorder (Panasonic, Model
WJ-HD 309). The experiment ended once two
juveniles were consumed. The number and identity
(which population) of juveniles consumed was also
recorded, and data were analyzed using v2 tests.
Survival with a predator without substrate
Baseline threatening stimulus test In the video from
the previous experiment, it was often impossible to
view the interactions between the juvenile(s) and
adults clearly. In order to closely observe the
interactions, we designed a follow-up experiment
without substrate.
Juvenile crabs were pre-screened for aggressive
tendencies. One juvenile crab [HM, mean CW =
28 ± 2.9 mm; TK, mean CW = 31 mm ± 2.3 mm
(SE)] from either population was placed in an
opaque rectangular Nalgene� container (dimensions:
L = 23 cm, W = 13.5 cm, H = 13.5 cm). The con-
tainer did not have substrate, but had a water depth of
4 cm. Individual crabs were allowed to acclimate for
30 min before testing. Each crab was tested three
times using a similar threatening stimulus: a #2
stopper (diameter: 2 cm) attached to a wooden dowel
(length: 52.5 cm) with a 10-min rest period between
tests. Responses were recorded as in the previous
section. The crab was classified as either ‘‘aggres-
sive,’’ ‘‘non-aggressive,’’ or ‘‘mixed’’ as a result of
the responses from this baseline test. Only TK
juveniles were used for this experiment, and by
doing this, we were able to isolate aggressive
behavior as a factor in avoiding predators. After this
classification, they were used in the following
experiment.
Predator avoidance
This predator avoidance experimental set-up is sim-
ilar to the experiment described earlier. The set-up
used a 76-l aquarium, but without sand to allow better
visual observations of behavioral interactions. Addi-
tionally, white paper was placed underneath the
aquarium to further enhance visibility when using the
low-light digital recorder. This set-up does not
simulate field conditions, but was necessary in order
to visualize the interactions between the predator and
the prey.
Two juveniles were used in each experiment in
one of the following combinations: one aggres-
sive ? one non-aggressive, one aggressive ? one
mixed, or one non-aggressive ? one mixed. All trials
were recorded using a closed circuit digital camera
(Ikegami Tsushinki Co. Ltd) with a 7.5–75-mm lens
(Canon) and digital disk recorder (Panasonic, Model
WJ-HD 309). The juveniles were marked with a
Sharpie� in order to distinguish them on the video
and allowed to acclimate to the tank for 1 h. After the
acclimation period, an adult male blue crab that had
been food deprived for 48 h was introduced to the
tank. The experiment ended after 48 h. The first
juvenile to be consumed and any other interactions
with the predator were recorded and analyzed using
v2 tests.
Adult crab pot experiment
To test if aggression continued into the adult stages,
which could ultimately affect entry into crab traps,
adults from both populations were pre-screened for
aggressive behavior. Adult blue crabs [HM: mean
CW = 118 mm ± 9.2 mm; TK: mean CW = 96 ±
8.7 mm (SE)] from either population were isolated in
a Nalgene� container (dimensions: L = 20 cm,
W = 21.5 cm, H = 6.5 cm) for 15 min. After the
acclimation period, a hand was passed over the
container. If the crab lunged, the individual was
classified as aggressive; if it did not, it was classified
as non-aggressive.
Crab pot experiments were conducted outdoors in a
large circular tank (diameter: 1.7 m; height: 1.5 m).
The tank had a sand depth of 1 cm and water depth of
0.5 m. Five adult crabs that had food deprived for
48 h were placed in the tank simultaneously and
allowed to acclimate for 30 min. Only crabs from one
Hydrobiologia
123
population were used in each trial (HM: N = 11 trials;
TK: N = 9 trials). After acclimation, one commercial
crab pot (dimensions: 1.0 m 9 1.3 m 9 0.75 m) with
bait (Atlantic menhaden, B. tyrannus) was placed on
the bottom of the tank. Crabs were checked after 24
and 48 h. After 48 h, the experiment ended and crab
positions within the tank and pot were recorded. The
number of crabs that had entered the pot at 24 and
48 h were analyzed using Student’s t tests.
Results
Response to a threatening stimulus
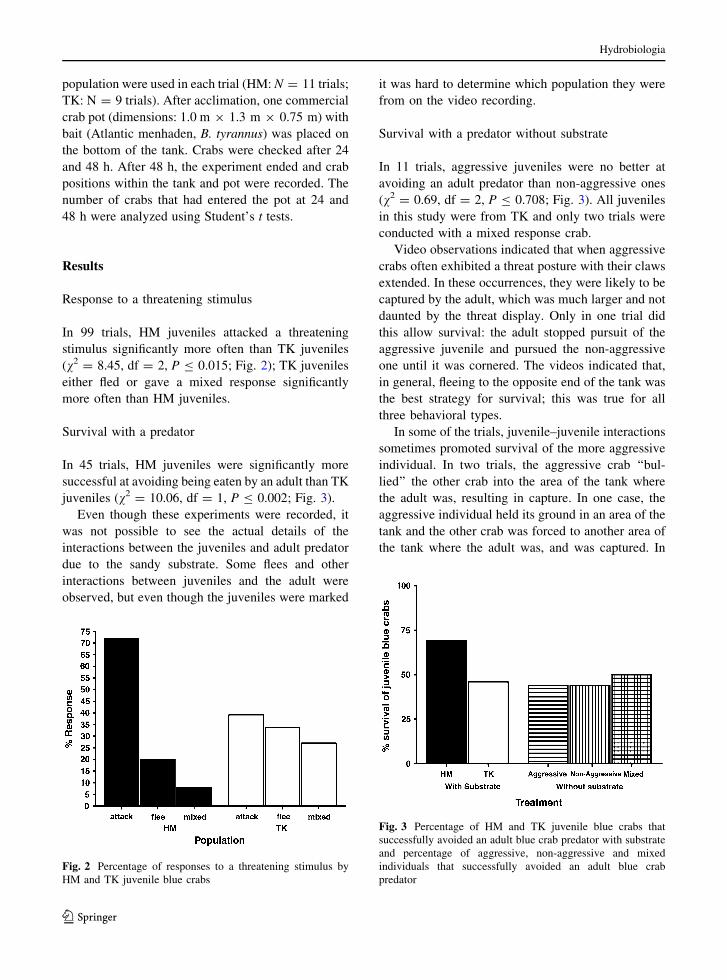
In 99 trials, HM juveniles attacked a threatening
stimulus significantly more often than TK juveniles
(v2 = 8.45, df = 2, P B 0.015; Fig. 2); TK juveniles
either fled or gave a mixed response significantly
more often than HM juveniles.
Survival with a predator
In 45 trials, HM juveniles were significantly more
successful at avoiding being eaten by an adult than TK
juveniles (v2 = 10.06, df = 1, P B 0.002; Fig. 3).
Even though these experiments were recorded, it
was not possible to see the actual details of the
interactions between the juveniles and adult predator
due to the sandy substrate. Some flees and other
interactions between juveniles and the adult were
observed, but even though the juveniles were marked
it was hard to determine which population they were
from on the video recording.
Survival with a predator without substrate
In 11 trials, aggressive juveniles were no better at
avoiding an adult predator than non-aggressive ones
(v2 = 0.69, df = 2, P B 0.708; Fig. 3). All juveniles
in this study were from TK and only two trials were
conducted with a mixed response crab.
Video observations indicated that when aggressive
crabs often exhibited a threat posture with their claws
extended. In these occurrences, they were likely to be
captured by the adult, which was much larger and not
daunted by the threat display. Only in one trial did
this allow survival: the adult stopped pursuit of the
aggressive juvenile and pursued the non-aggressive
one until it was cornered. The videos indicated that,
in general, fleeing to the opposite end of the tank was
the best strategy for survival; this was true for all
three behavioral types.
In some of the trials, juvenile–juvenile interactions
sometimes promoted survival of the more aggressive
individual. In two trials, the aggressive crab ‘‘bul-
lied’’ the other crab into the area of the tank where
the adult was, resulting in capture. In one case, the
aggressive individual held its ground in an area of the
tank and the other crab was forced to another area of
the tank where the adult was, and was captured. In
Fig. 2 Percentage of responses to a threatening stimulus by
HM and TK juvenile blue crabs
Fig. 3 Percentage of HM and TK juvenile blue crabs that
successfully avoided an adult blue crab predator with substrate
and percentage of aggressive, non-aggressive and mixed
individuals that successfully avoided an adult blue crab
predator
Hydrobiologia
123
another case, however, a scuffle between juveniles
alerted the adult and the aggressive individual was
captured.
Adult crab pot experiment
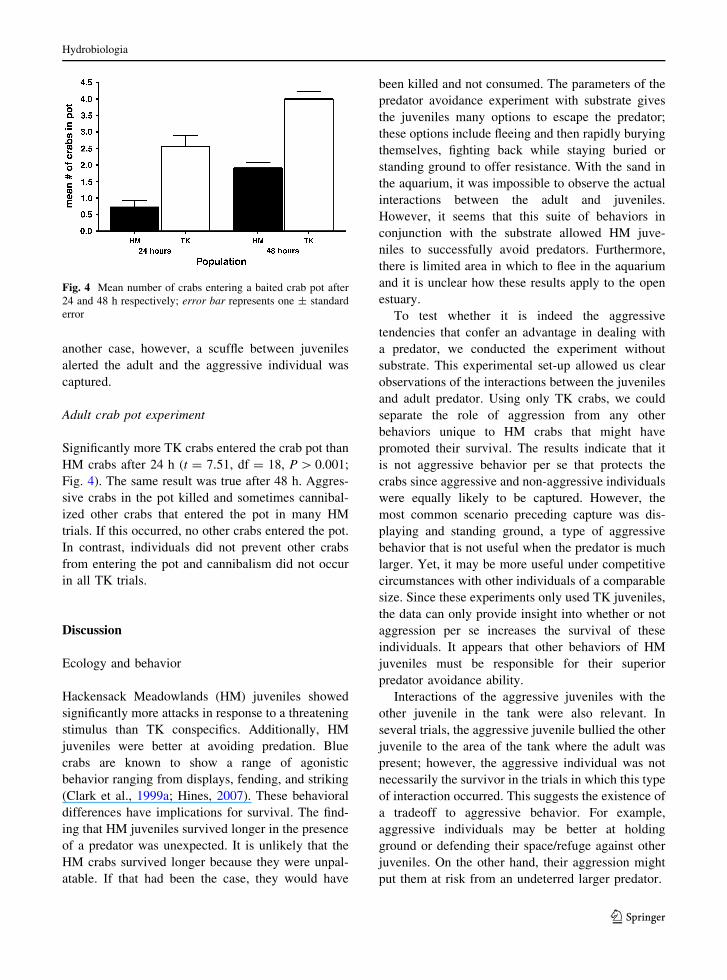
Significantly more TK crabs entered the crab pot than
HM crabs after 24 h (t = 7.51, df = 18, P [ 0.001;
Fig. 4). The same result was true after 48 h. Aggres-
sive crabs in the pot killed and sometimes cannibal-
ized other crabs that entered the pot in many HM
trials. If this occurred, no other crabs entered the pot.
In contrast, individuals did not prevent other crabs
from entering the pot and cannibalism did not occur
in all TK trials.
Discussion
Ecology and behavior
Hackensack Meadowlands (HM) juveniles showed
significantly more attacks in response to a threatening
stimulus than TK conspecifics. Additionally, HM
juveniles were better at avoiding predation. Blue
crabs are known to show a range of agonistic
behavior ranging from displays, fending, and striking
(Clark et al., 1999a; Hines, 2007). These behavioral
differences have implications for survival. The find-
ing that HM juveniles survived longer in the presence
of a predator was unexpected. It is unlikely that the
HM crabs survived longer because they were unpal-
atable. If that had been the case, they would have
been killed and not consumed. The parameters of the
predator avoidance experiment with substrate gives
the juveniles many options to escape the predator;
these options include fleeing and then rapidly burying
themselves, fighting back while staying buried or
standing ground to offer resistance. With the sand in
the aquarium, it was impossible to observe the actual
interactions between the adult and juveniles.
However, it seems that this suite of behaviors in
conjunction with the substrate allowed HM juve-
niles to successfully avoid predators. Furthermore,
there is limited area in which to flee in the aquarium
and it is unclear how these results apply to the open
estuary.
To test whether it is indeed the aggressive
tendencies that confer an advantage in dealing with
a predator, we conducted the experiment without
substrate. This experimental set-up allowed us clear
observations of the interactions between the juveniles
and adult predator. Using only TK crabs, we could
separate the role of aggression from any other
behaviors unique to HM crabs that might have
promoted their survival. The results indicate that it
is not aggressive behavior per se that protects the
crabs since aggressive and non-aggressive individuals
were equally likely to be captured. However, the
most common scenario preceding capture was dis-
playing and standing ground, a type of aggressive
behavior that is not useful when the predator is much
larger. Yet, it may be more useful under competitive
circumstances with other individuals of a comparable
size. Since these experiments only used TK juveniles,
the data can only provide insight into whether or not
aggression per se increases the survival of these
individuals. It appears that other behaviors of HM
juveniles must be responsible for their superior
predator avoidance ability.
Interactions of the aggressive juveniles with the
other juvenile in the tank were also relevant. In
several trials, the aggressive juvenile bullied the other
juvenile to the area of the tank where the adult was
present; however, the aggressive individual was not
necessarily the survivor in the trials in which this type
of interaction occurred. This suggests the existence of
a tradeoff to aggressive behavior. For example,
aggressive individuals may be better at holding
ground or defending their space/refuge against other
juveniles. On the other hand, their aggression might
put them at risk from an undeterred larger predator.
Fig. 4 Mean number of crabs entering a baited crab pot after
24 and 48 h respectively; error bar represents one ± standard
error
Hydrobiologia
123

At the same time, the juvenile crabs that were most
aggressive toward the stopper did not always show an
aggressive posture (i.e., bold behavior) toward the
predator suggesting that this behavior may be context
dependent. Similar results were found in sunfish.
Individuals were classified as bold when approaching
a meter stick, but the same bold individual did not
inspect a novel food source (Coleman & Wilson,
1998). The behavioral plasticity observed in the
predator avoidance experiments is adaptive to the
circumstances. Additional studies will be needed to
ascertain exactly what aspects of behavior are most
closely associated with the survival of HM juveniles,
since increased propensity for aggression per se is not
the critical factor.
Adult blue crabs are responsible for 75–97% of
mortality in juvenile blue crabs within Chesapeake
Bay (Hines & Ruiz, 1995). In the Gulf of Mexico,
this value ranges from 85 to 91% (Heck & Coen,
1995); in New Jersey, mortality ranges from 10 to
45% (Wilson et al., 1990a, b). However, in a previous
study, HM adults were found to be poor predators on
juvenile blue crabs under laboratory conditions, and
few crab parts were found in stomach contents of
field-collected individuals (Reichmuth et al., 2009).
Poor prey capture by adults, combined with effective
predator avoidance by juveniles, suggests that less
cannibalism may be occurring within the Meadow-
lands, which should allow larger numbers of juve-
niles to survive. If this is the case, juvenile blue crabs
may be overcrowded in a degraded, patchy habitat.
Studies have shown that increased agonistic encoun-
ters occur in populations of blue crabs in crowded
conditions (Mansour & Lipcius, 1991; Clark et al.,
2000, 1999a, b), and this may be one reason why HM
juveniles showed increased aggression. Funnel-web
spiders (Agelenopsis aperta) were highly aggressive
under low food availability conditions (Maupin &
Riechert, 2001). Furthermore, adult blue crabs were
found to exhibit increased agonism under varying
food availability (Mansour & Lipcius, 1991). In these
situations, resources are limited and competition is
increased, which favors aggressive individuals (Sih
et al., 2004a, b). Increased aggression may also be a
response to greater predation risk in the environment,
as has been seen in populations of various species
(spiders: Whitehouse, 1997; fiddler crabs: Reany &
Backwell, 2007). However, this does not appear
likely within the Meadowlands since a major predator
of juvenile blue crabs—adult blue crabs—are poor
predators and eat few juveniles (Reichmuth et al.,
2009). Whether predation by other species is greater
at HM than TK is not known.
However, another possible reason for altered
aggressive behavior is effects of contaminants. The
altered feeding behavior of adult HM blue crabs
(Reichmuth et al., 2009) is related to the environment
(Windham et al., 2004). When exposed to ‘‘clean
food’’ or a less impacted environment for 8 weeks,
HM adults predatory behavior improved and became
similar in behavior to crabs from TK, suggesting that
contaminants are the cause for the decreased preda-
tory behavior observed in HM adults (Reichmuth,
2009; Reichmuth et al., 2010).
Ecology of crab pots
The results of the crab pot experiment suggest that
HM crabs remain aggressive into the adult life stages.
Aggressive behavior can have serious implications on
the ecology of an organism if the individuals are
aggressive in novel or inappropriate situations (Sih
et al., 2004a, b). In our initial collections of juveniles
from HM, we put them in containers together, and by
the time we returned to the laboratory in under an
hour many had been killed by others, unlike TK
crabs. This experience prompted the current behav-
ioral studies.
A few of the crab pot trials with HM crabs resulted
in a crab being killed and eaten. The other crabs may
not have entered the trap due to the scent of the
injured conspecifics. Field experiments using crab
pots baited with an injured blue crab caught fewer
crabs than traps baited with menhaden (Ferner et al.,
2005). Another study using odor plumes containing
metabolites of injured crabs found crabs reduced their
foraging behavior and movement (Moir & Weiss-
burg, 2008). It is possible that metabolites released
from the injured crab in our mesocosm acted as a
deterrent.
Our results have important implication for popu-
lation surveys, suggesting caution when using crab
pots in population estimates. These results suggest an
apparent difference in the propensity for individual
crabs to enter the pot, which could also affect local
fisheries. When we used crab pots in the Meadow-
lands, few crabs were caught, and on occasion, an
inhabitant would be dead or severely damaged. Using
Hydrobiologia
123

the same effort in TK, many more crabs were caught
(personal observation) without damage to the inhab-
itants. The poor field catch with baited crab traps is
not an accurate representation of the blue crab
abundance within the Meadowlands since other
fishing techniques (seines and trawls) were far more
successful. A survey using only trapping as a
technique might result in severe undercounting if
behavior is not taken into account.
Conclusion
Hackensack Meadowlands juveniles were more
aggressive when threatened with a stimulus and were
also significantly better than crabs from the less
impacted site (Tuckerton) at avoiding predators.
However, it appears that aggressive behavior itself
does not confer an advantage with a larger predator,
but rather it may be more important in interactions
with similar-sized conspecifics in gaining refuge or
protecting a prime habitat patch. Aggressive behavior
also reduced the rate in which crabs entered baited
traps, which may have implications in using this
method for population counts or a localized fishery.
Acknowledgments This research was funded in part by
grants from the Rutgers University Marine Field Station
Graduate Student Research Fund and the Meadowlands
Environmental Research Institute (MERI). The authors would
like to thank B. Bragin (New Jersey Meadowlands
Commission) and J. Grzyb (MERI) for their help with
collection and boat time on the Hackensack River as well as
Dr. T. Glover for statistical assistance. We would also like to
thank the many undergraduate and high school students who
helped with collecting and maintaining specimens,
experimental set-up, and data collection in the lab.
References
Clark, M. C., T. G. Wolcott, D. L. Wolcott & A. H. Hines,
1999a. Intraspecific interference among foraging blue
crabs Callinectes sapidus: interactive effects of predator
density and prey patch distribution. Marine Ecology
Progress Series 178: 69–78.
Clark, M. C., T. G. Wolcott, D. L. Wolcott & A. H. Hines,
1999b. Foraging and agonistic activity co-occur in free-
ranging blue crabs (Callinectes sapidus): observations of
animals by ultrasonic telemetry. Journal of Experimental
Marine Biology and Ecology 233: 143–160.
Clark, M. C., T. G. Wolcott, D. L. Wolcott & A. H. Hines,
2000. Foraging behavior of an estuarine predator, the blue
crab Callinectes sapidus in a patchy environment.
Ecography 23: 21–31.
Coleman, K. & D. S. Wilson, 1998. Shyness and boldness in
pumpkinseed sunfish: individual differences are context-
specific. Animal Behavior 56: 927–936.
Crane, J., 1975. Fiddler Crabs of the World. Princeton
University Press, Princeton.
Edwards, D. C., D. O. Conover & F. Sutter III, 1982. Mobile
predators and the structure of marine intertidal commu-
nities. Ecology 63: 1175–1180.
Ferner, M. C., D. L. Smee & Y. P. Chang, 2005. Cannibalistic
crabs respond to the scent of injured conspecifics: danger
or dinner? Marine Ecology Progress Series 300: 193–200.
Graham, M. E. & J. Heberholz, 2009. Stability of dominance
relationships in crayfish depends on social context.
Animal Behavior 77: 195–199.
Hazlett, B. A., 1971. Interspecific fighting in three species of
brachyuran crabs from Hawaii. Crustaceana 20: 308–314.
Hazlett, B. A., 1972. Response to agonistic postures by the
spider crab Microphyrys bicornutus. Marine Behavioral
Physiology 1: 85–92.
Heck, J. L. Jr., & L. D. Coen, 1995. Predation and the abun-
dance of juvenile blue crabs: a comparison of selected east
and Gulf coast (USA) studies. Bulletin of Marine Science
57: 877–883.
Hines, A. H., 2007. Ecology of juvenile and adult blue crabs. In
Kennedy, V. S. & L. E. Cronin (eds), The Blue Crab,
Callinectes sapidus. Maryland Sea Grant College, College
Park, MD: 565–654.
Hines, A. H., A. M. Haddon & L. A. Wiechert, 1990. Guild
structure and foraging impact of blue crabs and epibenthic
fish in a subestuary of Chesapeake Bay. Marine Ecology
Progress Series 67: 105–126.
Hines, A. H. & G. M. Ruiz, 1995. Temporal variation in
juvenile blue crab mortality: nearshore shallows and
cannibalism in Chesapeake Bay. Bulletin of Marine
Science 57: 884–901.
Huffard, C. L., R. L. Caldwell & F. Boneka, 2010. Male–male
and male–female aggression may influence mating asso-
ciations in wild octopuses (Abdopus aculeatus). Journal of
Comparative Psychology 124: 38–46.
Huntingford, F. A., A. C. Taylor, I. P. Smith & K. E. Thorpe,
1995. Behavioural and physiological studies of aggression
in swimming crabs. Journal of Experimental Marine
Biology and Ecology 233: 143–160.
Jachowski, R. L., 1974. Agonistic behavior of the blue crab,
Callinectus sapidus Rathbun. Bulletin of Marine Science
6: 232–253.
Jop, K. M., R. C. Biever, J. R. Hoberg & S. P. Shepherd, 1997.
Analysis of metals in blue crabs, Callinectes sapidus,
from two Connecticut estuaries. Bulletin of Environmen-
tal Contamination and Toxicology 58: 311–317.
Juanes, F. & L. D. Smith, 1995. The ecological consequences
of limb damage and loss in decapod crustaceans: a review
and prospectus. Journal of Experimental Marine Biology
and Ecology 93: 197–223.
Kaiser, M. J., R. N. Hughes & D. G. Reid, 1990. Chelal
morphometry, prey-size selection and aggressive compe-
tition in green and red forms of Carcinus maenas (L.).
Journal of Experimental Marine Biology and Ecology
140: 121–134.
Hydrobiologia
123

Kravitz, E. A. & R. Huber, 2003. Aggression in invertebrates.
Current Opinion in Neurobiology 13: 736–743.
Mansour, R. A. & R. N. Lipcius, 1991. Density-dependent
foraging and mutual interference in blue crabs preying
upon infaunal clams. Marine Ecology Progress Series 72:
236–249.
Maupin, J. L. & S. E. Riechert, 2001. Superfluous killing in
spiders: a consequence of adaptation to food-limited
environments? Behavioral Ecology 12: 569–576.
Moir, F. & M. J. Weissburg, 2008. Cautious cannibals:
behavioral responses of juvenile and adult blue crabs to
the odor of injured conspecifics. Journal of Experimental
Marine Biology and Ecology. doi:10.1016/j.jembe.2008.
10.026.
Moksnes, P. O., R. N. Lipcius, L. Pihl & J. Montfrans, 1997.
Cannibal-prey dynamics in young juveniles and post lar-
vae of the blue crab. Journal of Experimental Marine
Biology and Ecology 215: 157–187.
Nelson, W. G., 1981. Experimental studies of decapod and fish
predation on seagrass macrobenthos. Marine Ecology
Progress Series 5: 141–149.
Norse, E. A., 1977. Aspects of the zoogeographic distribution
of Callinectes (Brachyuran: Portunidae). Bulletin of
Marine Science 27: 440–447.
Nye, L. A., 1989. Variation in feeding behavior of blue crabs
(Callinectes sapidus Rathbun) measured by ultrasonic
biotelemetry. M.Sc. thesis, North Carolina State Univer-
sity, Raleigh: 82 pp.
Pintor, L. M., A. Sih & M. L. Bauer, 2008. Differences in
aggression, activity and boldness between native an
introduced populations of an invasive crayfish. Oikos 117:
1629–1636.
Reany, L. T. & P. R. Y. Backwell, 2007. Risk-taking behavior
predicts aggression and mating success in a fiddler crab.
Behavioral Ecology 18: 521–525.
Reichmuth, J. M., 2009. Behavioral ecology and population
genetics in two populations of blue crabs Callinectes sa-pidus (Rathbun) in New Jersey. Ph.D. Thesis, Rutgers,
The State University of New Jersey: 125 pp.
Reichmuth, J. M., R. Roudez, T. Glover & J. S. Weis, 2009.
Differences in prey capture behavior in populations of
blue crab (Callinectes sapidus Rathbun) from contami-
nated and clean estuaries in New Jersey. Estuaries and
Coasts 32: 298–308.
Reichmuth, J. M., P. Weis & J. S. Weis, 2010. Bioaccumula-
tion and depuration of metals in blue crabs (Callinectessapidus Rathbun) from a contaminated and clean estuary.
Environmental Pollution 158: 361–368.
Romano, S. L., 1990. Long-term effects of interspecific
aggression on growth of the reef-building corals
Cyphastrea ocellina (Dana) and Pocillopora damicomis
(Linnaeus). Journal of Experimental Marine Biology and
Ecology 140: 135–146.
Shulman, M. J., 1990. Aggression among sea urchins on
Caribbean coral reefs. Journal of Experimental Marine
Biology and Ecology 140: 197–207.
Sih, A., A. Bell & C. J. Johnson, 2004a. Behavioral syndromes:
an ecological and evolutionary overview. Trends in
Ecology and Evolution 19: 372–378.
Sih, A., A. Bell, C. J. Johnson & R. E. Ziemba, 2004b.
Behavioral syndromes: an integrative overview. Quarterly
Review of Biology 79: 241–277.
Vasquez-Archdale, M., K. Anraku, T. Yamamoto & N. Hig-
ashitani, 2003. Behavior of the Japanese rock crab ‘Ishi-
gani’ Charybdis japonica towards two collapsible baited
pots: evaluation of capture effectiveness. Fisheries Sci-
ence 69: 785–791.
Vasquez, G. M. & J. Silverman, 2008. Intraspecific aggression
and colony fusion in the Argentine ant. Animal Behavior
75: 583–593.
Virnstein, R. W., 1979. Predation of estuarine infauna:
response patterns of component species. Estuaries 2:
69–86.
Ward, A. J. W., P. Thomas, P. J. B. Hart & J. Krause, 2004.
Correlates of boldness in the three-spined stickleback
(Gasterosteus aculeatus). Behavioral and Ecological
Sociobiology 55: 561–568.
Wilson, K. A., K. W. Able & K. L. Heck Jr., 1990a. Predation
rates on juvenile blue crabs in estuarine nursery habitats:
evidence for the importance of macroalgae (Ulva lactuca).
Marine Ecology Progress Series 48: 243–251.
Wilson, K. A., K. W. Able & K. L. Heck Jr., 1990b. Habitat
use by juvenile blue crabs: a comparison among habitats
in southern New Jersey. Bulletin of Marine Science 46:
105–114.
Whitehouse, M. E. A., 1997. The benefits of stealing from a
predator: foraging rates, predation risk, and intraspecific
aggression in the kleptoparasitic spider Argyrodes antip-odiana. Behavioral Ecology 8: 665–667.
Wolcott, T. G. & A. H. Hines, 1989. Ultrasonic biotelemetry of
muscle activity from free-ranging marine animals: a new
method for studying foraging by blue crabs (Callinectessapidus). Biological Bulletin 176: 50–56.
Wolcott, T. G. & A. H. Hines, 1990. Ultrasonic telemetry of
small-scale movements and microhabitat selection by
molting blue crabs (Callinectes sapidus). Bulletin of
Marine Science 46: 83–94.
Windham, L., J. S. Weis & P. Weis, 2004. Metal dynamics of
plant litter of Spartina alterniflora and Phragmites aus-tralis in metal-contaminated salt marshes. Part 1. Patterns
of decomposition and metal uptake. Environmental Tox-
icology and Chemistry 23: 1520–1528.
Hydrobiologia
123