f-81 skeleton from wadi mataha, jordan, and its bearing on human variability in the epipaleolithic...
TRANSCRIPT

F-81 Skeleton from Wadi Mataha, Jordan, and itsBearing on Human Variability in the Epipaleolithicof the Levant
J.T. Stock,1* S.K. Pfeiffer,2 M. Chazan,2 and J. Janetski3
1Leverhulme Centre for Human Evolutionary Studies, Department of Biological Anthropology,University of Cambridge, Cambridge CB2 3DZ, UK2Department of Anthropology, University of Toronto, Toronto, Ontario M5S 1A1, Canada3Department of Anthropology, Brigham Young University, Provo, Utah 84601
KEY WORDS Geometric Kebaran; Natufian; osteometrics; robusticity; cranial variation
ABSTRACT The discovery of a Middle Epipaleolithicadult skeleton (F-81) at the site of Wadi Mataha insouthern Jordan provides new insights into humanvariability in the Epipaleolithic of the Levant. Thispaper analyzes the skeletal morphology of Wadi MatahaF-81 in the context of other Epipaleolithic remainsfrom Jordan and Israel to assess the current evidencefor morphological variability throughout this period.The F-81 skeleton shares morphological features withearlier Epipaleolithic skeletons from Ohalo and NahalEin Gev, and later Natufian populations. Despite themorphological similarities, F-81 extends the range ofknown variability prior to the Natufian with its unu-
sually small stature and unique combination of morpho-logical characteristics. High levels of cranial andpostcranial robusticity suggest that the F-81 individualwas physically active and terrestrially mobile. Pro-nounced bilateral asymmetry in the upper limb sug-gests significant lateralization of habitual activity. Inthe context of Epipaleolithic remains, the F-81 skeletonprovides preliminary evidence for greater morphologicalvariability, terrestrial mobility, and lateralized habitualbehavior prior to the Natufian, and skeletal graciliza-tion between the Middle and Late Epipaleolithic in theLevant. Am J Phys Anthropol 128:453–465, 2005.' 2005 Wiley-Liss, Inc.
Human remains from the Upper Paleolithic (UP)and Early Epipaleolithic of the Levant are veryrare. The most complete skeletons include thewell-preserved Ohalo II male, dated to 19,000 BP(Hershkovitz et al., 1995), and the distorted NahalEin Gev I female, dated to ca. 24,000 years BP(Arensburg, 1977, 1981). There are two isolatedfrontal bones and a fragmentary mandible fromQafzeh Cave, and some human bone and toothfragments from Hayonim (Arensburg et al., 1990).
The Middle Epipaleolithic is also poorly repre-sented by human skeletal remains. There are iso-lated teeth, mandible, and postcranial fragmentsfrom El-Wad and Kebara, and a complete butdamaged skeleton from Ein Gev I, dated to13,000 BP (Arensburg and Bar-Yosef, 1973). Sev-eral skeletons were recovered from contexts asso-ciated with Geometric Kebaran lithics. Theseinclude two fragmentary skeletons from Neve-David, dated to between 12,610–13,400 BP (Kauf-man and Ronen, 1987), and two skeletons from thesite of Qasr Kharaneh in Jordan (Rolston, 1992).The remains from Neve-David have not yet beendescribed in anatomical detail, but a basic osteobio-graphical and osteometric description of the twomale skeletons from Qasr Kharaneh was published(Rolston, 1992).
The discovery of an adult skeleton (designatedF-81; Figs. 1, 2) at the site of Wadi Mataha in thePetra Basin of southern Jordan, in association
with Geometric Kebaran lithics, provides animportant addition to the Middle Epipaleolithicarchaeological record of the Levant. The earliestoccupation of Wadi Mataha dates to the GeometricKebaran, which corresponds to the Older Dryasand the onset of the Bølling-Allerød (Goring-Morris and Belfer-Cohen, 1998; Southon, 2002).An AMS radiocarbon date from burned animalbone places the Geometric Kebaran occupation at14,100 6 130 radiocarbon years BP, which cali-brates to 17,579–16,457 calibrated years. BP at2 sigma, using Calib 4.4 (Stuiver et al., 1998). TheF-81 skeleton was excavated from the GeometricKebaran level, and was found in association witha fragmentary juvenile skeleton. The F-81 burialwas positioned with the face down and hands and
*Correspondence to: J.T. Stock, Leverhulme Centre for HumanEvolutionary Studies, Department of Biological Anthropology, Uni-versity of Cambridge, Downing Street, Cambridge CB2 3DZ, UK.E-mail: [email protected]
Grant sponsor: Social Sciences and Humanities Research Councilof Canada.
Received 5 November 2003; accepted 10 June 2004.
DOI 10.1002/ajpa.20163Published online 28 April 2005 in Wiley InterScience
(www.interscience.wiley.com).
# 2005 WILEY-LISS, INC.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 128:453–465 (2005)
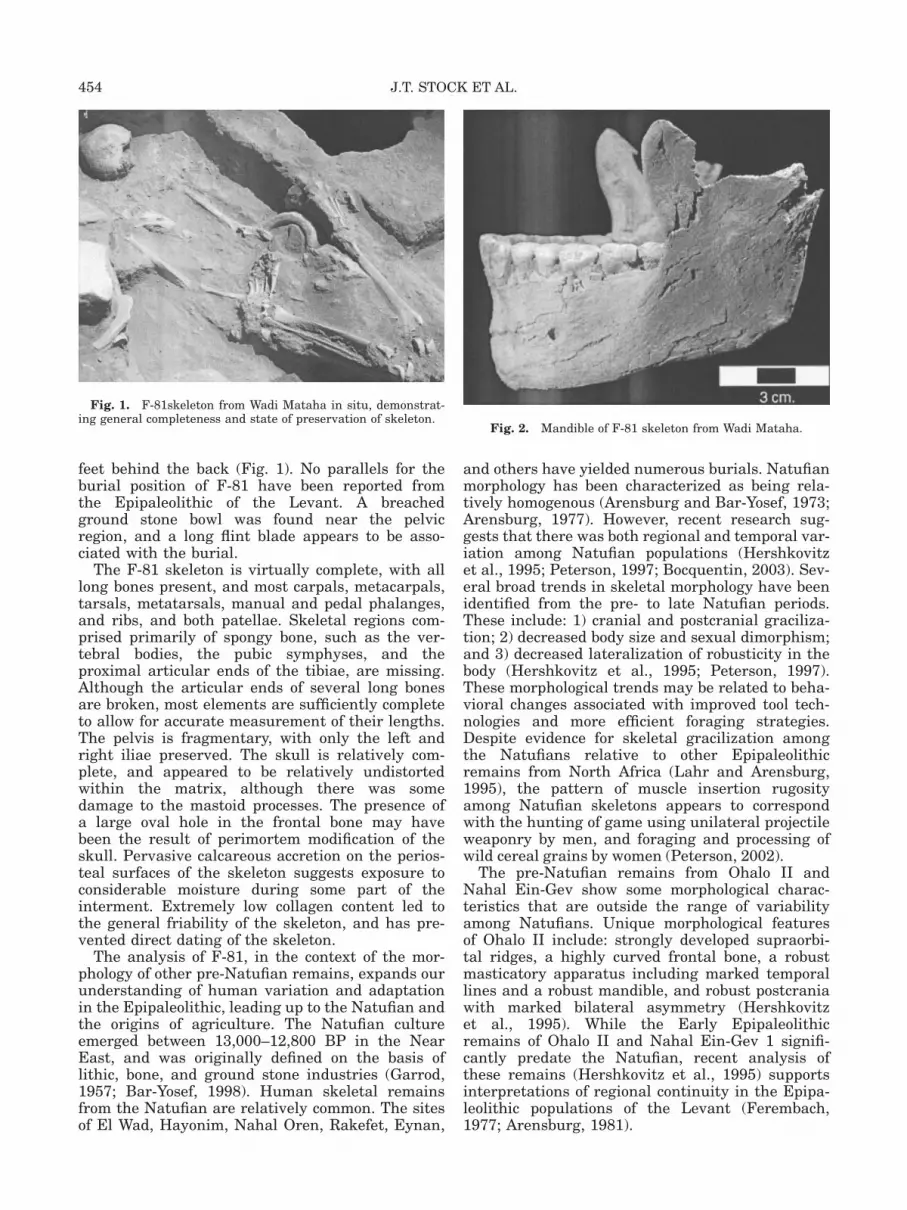
feet behind the back (Fig. 1). No parallels for theburial position of F-81 have been reported fromthe Epipaleolithic of the Levant. A breachedground stone bowl was found near the pelvicregion, and a long flint blade appears to be asso-ciated with the burial.
The F-81 skeleton is virtually complete, with alllong bones present, and most carpals, metacarpals,tarsals, metatarsals, manual and pedal phalanges,and ribs, and both patellae. Skeletal regions com-prised primarily of spongy bone, such as the ver-tebral bodies, the pubic symphyses, and theproximal articular ends of the tibiae, are missing.Although the articular ends of several long bonesare broken, most elements are sufficiently completeto allow for accurate measurement of their lengths.The pelvis is fragmentary, with only the left andright iliae preserved. The skull is relatively com-plete, and appeared to be relatively undistortedwithin the matrix, although there was somedamage to the mastoid processes. The presence ofa large oval hole in the frontal bone may havebeen the result of perimortem modification of theskull. Pervasive calcareous accretion on the perios-teal surfaces of the skeleton suggests exposure toconsiderable moisture during some part of theinterment. Extremely low collagen content led tothe general friability of the skeleton, and has pre-vented direct dating of the skeleton.
The analysis of F-81, in the context of the mor-phology of other pre-Natufian remains, expands ourunderstanding of human variation and adaptationin the Epipaleolithic, leading up to the Natufian andthe origins of agriculture. The Natufian cultureemerged between 13,000–12,800 BP in the NearEast, and was originally defined on the basis oflithic, bone, and ground stone industries (Garrod,1957; Bar-Yosef, 1998). Human skeletal remainsfrom the Natufian are relatively common. The sitesof El Wad, Hayonim, Nahal Oren, Rakefet, Eynan,
and others have yielded numerous burials. Natufianmorphology has been characterized as being rela-tively homogenous (Arensburg and Bar-Yosef, 1973;Arensburg, 1977). However, recent research sug-gests that there was both regional and temporal var-iation among Natufian populations (Hershkovitzet al., 1995; Peterson, 1997; Bocquentin, 2003). Sev-eral broad trends in skeletal morphology have beenidentified from the pre- to late Natufian periods.These include: 1) cranial and postcranial graciliza-tion; 2) decreased body size and sexual dimorphism;and 3) decreased lateralization of robusticity in thebody (Hershkovitz et al., 1995; Peterson, 1997).These morphological trends may be related to beha-vioral changes associated with improved tool tech-nologies and more efficient foraging strategies.Despite evidence for skeletal gracilization amongthe Natufians relative to other Epipaleolithicremains from North Africa (Lahr and Arensburg,1995), the pattern of muscle insertion rugosityamong Natufian skeletons appears to correspondwith the hunting of game using unilateral projectileweaponry by men, and foraging and processing ofwild cereal grains by women (Peterson, 2002).
The pre-Natufian remains from Ohalo II andNahal Ein-Gev show some morphological charac-teristics that are outside the range of variabilityamong Natufians. Unique morphological featuresof Ohalo II include: strongly developed supraorbi-tal ridges, a highly curved frontal bone, a robustmasticatory apparatus including marked temporallines and a robust mandible, and robust postcraniawith marked bilateral asymmetry (Hershkovitzet al., 1995). While the Early Epipaleolithicremains of Ohalo II and Nahal Ein-Gev 1 signifi-cantly predate the Natufian, recent analysis ofthese remains (Hershkovitz et al., 1995) supportsinterpretations of regional continuity in the Epipa-leolithic populations of the Levant (Ferembach,1977; Arensburg, 1981).
Fig. 1. F-81skeleton from Wadi Mataha in situ, demonstrat-ing general completeness and state of preservation of skeleton.
Fig. 2. Mandible of F-81 skeleton from Wadi Mataha.
454 J.T. STOCK ET AL.

Morphological variability within the LevantineEpipaleolithic may represent changes in skeletalanatomy associated with changes in habitual beha-vior. There is significant evidence that variability inhuman postcranial robusticity is related to differ-ences in habitual activity (see Ruff, 2000a andreferences therein). Further research suggests thatthis variability can be used to interpret behavioraldifferences between different populations of hunter-gatherers (Stock and Pfeiffer, 2001; Stock, 2002c).Human variability in cranial robusticity may berelated to tooth size and dental biomechanics, butthere is additional evidence for a correspondencebetween cranial robusticity and systemic patternsof robusticity resulting from general levels of physi-cal activity (Nawrocki, 1992; Lieberman, 1996).Given the evidence for relatively gracile skeletalmorphology in the Natufian and the in situ develop-ment of Epipaleolithic populations in the Levant(Ferembach, 1977; Arensburg, 1981; Lahr andArensburg, 1995; Hershkovitz et al., 1995), wewould expect to find evidence for skeletal graciliza-tion throughout the Early and Middle Epipaleo-lithic in the Near East.
When compared to later Natufian skeletons fromthe region, and to other Middle Epipaleolithic andearlier remains from Ohalo and Nahal Ein Gev,the study of the F-81 skeleton provides an opportu-nity to further evaluate morphological relation-ships among Epipaleolithic populations of theLevant, and to provide a preliminary quantifica-tion of skeletal robusticity in the Middle Epipaleo-lithic. Given the rarity of pre-Natufian burials,skeletal remains from these contexts provideessential evidence to determine the nature of adap-tive and behavioral changes among early humanforagers in this region. This paper compares themorphology of pre-Natufian skeletons and subse-quent remains from Natufian contexts in order toaddress the following questions: 1) Is the morphol-ogy of F-81 consistent with previously describeddifferences between the pre-Natufian and Natufianpopulations of the Levant? 2) Is there evidence formorphological change throughout the Epipaleo-lithic? 3) Is the skeletal robusticity of Wadi MatahaF-81, when compared to other hunter-gatherers,suggestive of particular patterns of habitual fora-ging behavior prior to the Natufian?
AGE AND SEX DETERMINATION
All aspects of the F-81 skeleton indicate a fullymature adult. Among cranial sutures, only the coro-nal suture appears to be partially obliterated ecto-cranially. The maxillary and mandibular thirdmolars are fully erupted and slightly worn. Thecusps of the first mandibular molars are worn tostages 6 and 7 of Scott (1979), while the secondmolar cusps are at stages 5 and 6 of wear. Each thirdmolar shows slight dentine exposure on one cusponly. There is relatively little difference in wear of
the first or second molars, indicating that toothwear was not rapid. Given the lack of evidence formarked osteoarthritis or alveolar resorption, anage range of approximately 35–55 years is likely.
Most extant sexually dimorphic features suggestthat F-81 is male. The bones of the pelvis arepoorly preserved, but one partial ilium shows astrongly developed pilaster. On the cranium, themastoid processes are damaged, but there are mod-erately developed superciliary arches, strong mus-cle markings in the nuchal region, and a high andwide palate. The root of the zygomatic archextends posteriorly from the external auditorymeatus. The mandible has moderate gonial ever-sion, a relatively vertical ascending ramus that isbroad antero-posteriorly, and a prominent mentaleminence. While the body size of the F-81 skeletonis small, joint sizes and skeletal robusticity arehigh relative to size.
MATERIALS AND METHODS
Standard cranial, dental, and postcranial metricmeasurements of the Wadi Mataha F-81 skeletonwere taken. These were analyzed within the con-text of skeletal variability in the Early Epipaleo-lithic, Geometric Kebaran, and Natufian, asderived from the literature and osteometric datacollected from human remains from Hayonim,Nahal Oren, Eynan, and Rakefet. These data arecontrasted with published osteometric data for pre-Natufian remains from Ohalo II, Nahal Ein Gev,Ein Gev, and Qasr Kharaneh, to place the mor-phology of the F-81 skeleton within a comparativecontext.
Robusticity of the F-81 crania and postcraniawere compared using a variety of approaches.Robusticity, in its most general definition, refers tothe strength of a skeletal element as reflected bythe size, shape, and organization of bone tissue.Using this definition, robusticity can refer tostrengthened elements of the cranium or postcra-nial skeleton. Since the major morphological differ-ences noted between pre-Natufian and Natufianremains involve a reduction in skeletal robusticity,an analysis of this aspect is essential to place theF-81 skeleton within the context of human varia-bility in the Levant. Previous research document-ing skeletal robusticity in Middle Easternpopulations used various methods of quantifica-tion, and so it is necessary to use severalapproaches to investigate skeletal gracilization indetail. External osteometric data can be used tocalculate indices of robusticity for individual longbones, while cranial vault thickness and thecortical thickness of long bone diaphyses provideadditional methods of quantifying robusticity.Comparisons of external osteometric data wereused as a basic method of investigating skeletalrobusticity. In addition, biomechanical propertiesof the F-81 long bones were quantified using cross-
455WADI MATAHA F-81 SKELETAL MORPHOLOGY

sectional geometric properties of the diaphyses,standardized to estimates of body size (Ruff,2000b). This method uses a combination of bipla-nar radiographs and molds of the periosteal sur-face (Trinkaus and Ruff, 1989; Trinkaus, 1997;Stock, 2002a) to quantify the distribution of corti-cal bone in long bone diaphyses, in order to esti-mate the mechanical performance of the boneunder various types of loading. This provides amore detailed biomechanical analysis of thestrength of the skeleton than external osteo-metrics. Using a combination of these analyticalapproaches, it is possible to place the morphologyof the F-81 skeleton within the context of hunter-gatherer variability from other regions.
In the present paper, the term robusticity will beused to refer to indices of external diaphysis mor-phology, while the terms strength and diaphysealcircularity will be used to refer to various aspectsof long bone cross-sectional geometry. Descriptionsof variables used to quantify robusticity and cross-sectional geometry are provided in Table 1. Thetorsional strength of a diaphysis, expressed as J0.73
(Ruff, 2000b), is thought to be the most relevantindicator of the mechanical performance of a boneunder a variety of loading conditions. Bilateralasymmetry in strength characteristics of thehumerus appears to be directly related to activitypatterns (Ruff, 1992; Trinkaus et al., 1994;Haapasalo et al., 1996; Churchill and Formicola,1997; Schmitt et al., 2003). There is no reason tobelieve that the mechanical influences on asymme-try of the humerus would not extend to otherupper limb bones. Bilateral asymmetry in J0.73 ofthe humeri and ulnae are used to provide indica-tors of relative loading of one arm vs. the other inrelation to habitual activity.
Morphological comparisons are made betweenF-81 and pre-Natufian and Natufian skeletonswherever comparators are available. The analysisof skeletal biomechanics incorporates a broad com-parative approach to place the mechanical attri-butes of the F-81 skeleton within the context ofvariability in the Epipaleolithic of the Levant, andamong other well-documented groups of Holocenehunter-gatherers. Samples used in this comparisoninclude: Indigenous Andaman Islanders (AI), semi-sedentary, marine-adapted foragers who madeextensive use of watercraft and a variety of hunt-
ing technologies including bows and spears (Stockand Pfeiffer, 2001); and Later Stone Age SouthernAfricans (LSA), highly mobile terrestrially basedforagers who likely hunted using a combination ofspears and light bows, and foraged, often makinguse of weighted digging sticks (Stock and Pfeiffer,2004). A previous comparison of long bone cross-sectional geometry among these hunter-gatherersdemonstrated that their diaphyseal morphologycorresponds with patterns of habitual activity andterrestrial mobility (Stock and Pfeiffer, 2001). Com-parisons of the biomechanical properties of theF-81 skeleton to data from these groups allowed usto determine whether the long bone strengths ofF-81 demonstrate morphological characteristicsthat are analogous with specific patterns of habi-tual behavior among other well-documented groupsof hunter-gatherers.
RESULTS
Craniodental and mandibular morphology
Cranial and mandibular measurements of theF-81 skeleton are compared to other pre-Natufianremains, and mean values for a sample of Natufianskulls in Table 2. A comparison of the cranial mod-ule provides an approximation of differences incranial size among the pre-Natufian and Natufianperiods. The pre-Natufian remains have modulesthat are slightly lower than the Natufian mean,suggesting that there was a slight increase incranial size between these periods. The maximumcranial lengths and heights of F-81 and the otherpre-Natufian crania are consistently lower thanthe Natufian mean. With a basion-bregma heightof approximately 125 mm, F-81 and Ein Gev 1have very low cranial vaults, a feature that seemsto be characteristic of all pre-Natufian crania. Thistrend appears to be associated with slightly widercranial vaults, as reflected by the higher maximumcranial breadths and cranial indices of the pre-Nat-ufian remains. F-81 shares a Natufian pattern oflong parietal bones, as documented by the bregma-lambda chord and arc, yet the occipital chord fallswell below values for Ohalo II H2, Nahal Ein Gev1, and Natufian mean. The F-81 cranium alsofeatures a very low minimum frontal breadth
TABLE 1. Variables used to quantify postcranial robusticity
Symbol Definition Mechanical relevance
CA Cortical area Axial compressive and tensile strengthImax Maximum second moment of area Correlate of maximum bending strengthImin Minimum second moment of area Correlate of minimum bending strengthImax/Imin Bending strength circularity ratio Ratio of Imax to Imin
J Polar second moment of area Torsional strengthBA Bilateral asymmetry between left and
right values of a given variable(Greater side � lesser side/lesser)*100
Robusticity index (Maximum þ minimum diaphysealbreadth/bone length)*100
External quantification of long bonerobusticity
456 J.T. STOCK ET AL.

(Ft–Ft ¼ 94.5 mm), comparable with the NahalEin Gev 1 female.
Several nonmetric aspects of cranial morphologywere proposed to characterize the Natufian, includ-ing a high, rounded frontal squama, low, rectangu-lar orbits, and minimal expression of superciliaryarches. The Ohalo II skeleton features a high fron-tal bone, with well-developed superciliary arches.Most Natufian skulls share the characteristicallyraised, vertical forehead, but lack the prominentbrow ridges of Ohalo II. By comparison, the F-81skeleton lacks the higher, more rounded frontalsquama that is characteristic of most Natufianskulls, but it features a reduction in the promi-nence of the superciliary arches from the morphol-ogy of Ohalo II. The orbits of Ohalo II and Ein GevI are rectangular and narrow, but they are lowerthan the orbits of most Natufian skulls. The orbitsof F-81 appear to be low in height and rectangularin shape, but poor preservation of the inferior orbi-tal margin prohibits accurate measurement to sup-port direct comparisons.
The cranial vault of F-81 is thick (Fig. 3).Table 3 compares vault thicknesses of F-81 at theparietal eminence and asterion bilaterally, and atobelion, to the range among modern humans, andamong nine early modern humans from the sites ofSkhul and Qafzeh. Measurements of cranial thick-ness of 9 and 8.5 mm at the parietal eminences fallin the upper end of the range of modern human
variability, and lie above the means for the Skhul/Qafzeh crania. The thickness of the F-81 parietalbones at obelion is 7 and 8 mm for the right andleft sides, respectively. These values fall in theupper end of the modern human range. The vaultthickness of 4.5 mm at asterion provides furtherevidence that the F-81 cranium is robust in thecontext of modern human variation.
The mandible of F-81 has characteristics thatwould be unusual among Natufian populations. Com-pared to Ohalo II, there are similarities and differ-ences. The heights of the mandibular body at M2 forF-81 (32 mm) and Ohalo II (30 mm) are higher thanNahal Ein-Gev or the Natufian means. The F-81mandible is more gracile than that of Ohalo II, andcontrasts in terms of the consistency of the height ofthe mandibular corpus, and the gonial angle. Despitethese differences, both mandibles share a relativelybroad ascending ramus with a slight posterior con-cavity, and a vertically oriented and deeply concaveanterior border. It was suggested that there was areduction in the breadth and border concavity ofthe mandibular ramus in the Natufian period(Hershkovitz et al., 1995). The bigonial breadth ofF-81 is higher than the Natufian mean. This featureis shared with the Ohalo II and Qasr Kharaneh2 male skeletons, and appears to be characteristic ofpre-Natufian mandibular morphology.
The full dentition of F-81 shows excellent align-ment and occlusion. There is no evidence of caries
TABLE 2. F-81 cranial and mandibular metrics, compared to upper paleolithic and natufian remains
Measurement(Martin number)1
Wadi MatahaF-81
Ohalo IIH2 male2
QasrKharaneh2 male3
Ein Gev1 female4
Nahal Ein Gev1 female4
Natufianmean2
CraniumGl-Op (1) 182 182 180 186 177 191Eu-Eu (8) 143 144 150.5 146 138 139Ba-Br (17) 125 135 115? 125 130 144Cranial module 150 154 149 152 148 158Cranial index 78.6 79.7 83.6 78.0 76.8Cranial length-height index 76.9 82.8 69.6 75.3 82.5 87.3Na-Br chord (29) 119.5 110 109 125 115 114Na-Br arc (26) 136 133 131 140 135 133Br-La chord (30) 125.5 116 117 112 115 124Br-La arc (27) 145 127 136 125 128 142La-O chord (31) 86.9 99 946 98La-O arc (28) 113 123 1116 117La-In chord (31 (1)) 53.1 68 60La-In arc (28 (1)) 61 70 58In-O chord (31 (2)) 49.5 51 50In-O arc (28 (2)) 50 53 53Ft-Ft (9) 94.5 105 108 99 95 98Foramen mag br (16) 30.2 30 30
MandibleGo-Go 102 102 113.5 95 87 92Height at M2 32 30 (26)2 (29)5
Minimum ramus Br 37.5 40 35 33 36
1 Measurement definitions from Martin and Saller, (1957).2 Measurements from Hershkovitz et al. (1995).3 Measurements from Rolston (1992).4 Measurements from Arensburg (1977).5 Measurements from Belfer-Cohen et al. (1991).6 Estimated measurements from Arensburg (1977).
457WADI MATAHA F-81 SKELETAL MORPHOLOGY

or alveolar resorption. Tooth wear exposed thedentine on all anterior teeth, most molar cusps,and the higher of the premolar cusps, but there isno evidence of nonmasticatory habitual tooth use.Matrix adhering to the teeth confounds the obser-vation of calculus and hypoplasia, and precludessome anterior maxillary measurements. However,most teeth could be measured in both the mesio-distal and buccal-lingual dimensions (Table 4). Themesio-distal and bucco-lingual dimensions of theF-81 maxillary molars are relatively small com-pared to the Natufian mean reported by Smith(1970). The remaining teeth are generally similarin size to mean Natufian values, with a slighttrend toward greater size toward the anterior por-tion of the mandibular dentition.
Bone lengths, body size, and proportions
Postcranial osteometrics and indices represent-ing limb proportions are presented in Table 5. Allof the measured long bone lengths of F-81 suggestthat the stature of this individual was low com-pared to any Epipaleolithic or Natufian remainsfrom the Levant. All bone lengths fall below theNatufian means, and below any of the other pre-Natufian remains yet described. The estimated sta-ture of F-81, based on femur length, is 155 63.3~cm, using the formula of Trotter (1970) forwhite males. This is shorter than any of eight Nat-ufian male skeletons from Hayonim, Eynan, andNahal Oren, with a range of estimated staturefrom 158–175 cm (N ¼ 8; mean ¼ 166.2 cm; SD ¼5.4 cm), and shorter than any of the skeletonsdescribed by Belfer-Cohen et al. (1991). In con-
TABLE 4. F-81 dental matrics1
Tooth
Right maxillary Right mandibular
Mesio-distal
Bucco-lingual
Mesio-distal
Bucco-lingual
I1 8.6 6.8 5.1 6.9I2 7.2 8.8 5.8 6.8C 7.4 9.3 6.2 7.8PM3 6.8 9.3 6.5 8.7PM4 7.4 11.2 7.1 9.1M1 9.8 11.6 10.9 11M2 9.6 10.5 10.7 10.4M3 7.8 10.7 10.5
1 All measurements in millimeters.
TABLE 3. F-81 cranial vault thickness
Location F-81Modern mean
(range)1Skhul/Qafzehmean (range)2
Parietaleminence
9.0 R, 8.5 L 5.7 (3–9.5) 8.2, n ¼ 9(4.5 –11)
Obelion 7.0 R, 8.0 L 5.8 (3–8.7)Asterion 4.5 3.9 (1.5–7)
1 Comparative data from Olivier (1969).2 Data from Nawrocki (1992).
TABLE
5.F-81postcra
nialmetrics,comparedto
other
Epipaleolithic
remains
Wadi
MatahaF-81
Ohalo
IIH2male
1Qasr
Kharaneh
1male
2Qasr
Kharaneh
2male
2NahalEin
Gev
1female
3
Natufianmales4
Natufianall4
NMea
nSD
z-score5
NMea
nSD
z-score5
Clavicle
length
128.0
150.0
145
7143.9
4.1
�3.88
18
142.7
8.9
�1.65
Humeruslength
280.0
344.0
306.8
315
11311
.412.6
�2.49
24
307.5
20.2
�1.36
Ulnalength
233.0
277.0
9266.5
8.6
�3.90
24
258.7
18.1
�1.42
Radiuslength
214.5
217
9245.3
8.7
�3.54
20
242.1
16.1
�1.71
Fem
urlength
394.0
494.0
412
8440.2
22.6
�2.04
20
433.3
24.7
�1.59
Fem
urhea
ddiameter
43.0
49.0
43.5
47
745.8
2.7
�1.04
14
43.8
5.3
�0.15
FHD/FXL*100
10.9
9.9
610.3
0.5
1.20
12
10.4
0.5
1.00
Tibia
length
3306
387.0
372
8363.2
11.8
�2.81
19
355.2
17.6
�1.43
Brach
ialindex
76.6
73.0
878.3
1.8
�0.94
19
78.0
2.4
�0.58
Cru
ralindex
83.8
683.0
882.6
4.1
0.29
15
81.2
3.7
0.70
1Mea
suremen
tsfrom
Hersh
kov
itzet
al.(1995).
2Mea
suremen
tsfrom
Rolston
(1992).
3Mea
suremen
tsfrom
Arensb
urg
(1977).
4Natufianmea
suremen
tstaken
byJTS,includingremainsfrom
NahalOren,Hey
onim
,Eynan,andRakefet.
5z-scores
representrelation
ship
betwee
nWadiMatahaF-81anddistribution
ofsa
memea
suremen
tamon
gNatufiansa
mples.
6Estim
atedvalue.
458 J.T. STOCK ET AL.

trast, the stature estimate for Ohalo II is 173.5 cm,toward the upper end of the Natufian range.
While all long bone lengths show positive corre-lations with stature, the diameter of the femoralhead is more strongly correlated with body mass(Ruff et al., 1997). The F-81 femoral head diameterof 43 mm falls 2.8 mm below the mean for Natu-fian males, but only slightly below the overall Nat-ufian mean. Femoral head diameters are alsoavailable from Ohalo II H2 and the two male ske-letons from Qasr Kharaneh (QK). Of these, OhaloII and QK 2 fall above the Natufian means, whilethe diameter of the QK1 femoral head is a low43.5 mm, comparable to F-81. The ratio of femoralhead diameter (FHD) to femur length (FXL) canprovide a provisional estimate of the relationshipbetween body mass and stature. In this dimension,the F-81 skeleton is particularly high compared toOhalo II and the Natufian remains, suggestingthat although short in stature, the body mass tostature ratio of F-81 was high.
In terms of body proportions, the brachial indexof F-81 is 76.6. This is between the means forrecent North African populations (78.6) and Eur-opeans (75.1) reported by Holliday (1997), andbelow the mean for male Natufians from Hayonim(N ¼ 8; mean ¼ 78.1; SD ¼ 1.8). The brachialindex of the Ohalo II male is 73, which is wellbelow the mean of Natufians. The crural index of83.8 for the F-81 skeleton, based on estimated tibialength, is similar to the value of 83 calculated forOhalo II. Both are comparable to Natufian cruralindices (N ¼ 8; mean ¼ 82.6; SD ¼ 4.1).
Skeletal biomechanics and behavioralinterpretations
It has been asserted, on the basis of the rugosityof musculoskeletal stress markers, that there isa general gracility among Natufians (Peterson,
1997). An analysis of muscle insertion rugosity isnot entirely appropriate for the F-81 skeletonbecause of taphonomic damage to the epiphysealends of the long bones. However, the femora showminimal development of the linea aspera, and alow pilastric index comparable to Natufian femora.The shape of the diaphysis in the subtrochantericregion, as reflected by the meric index, featuressignificant antero-posterior flattening. This trait isshared with Ohalo II but differs from other Natu-fians. In the upper limb, the humeri show well-developed deltoid insertions. Although it remainsdifficult to adequately compare muscle insertionrugosity with other groups, below we will investi-gate the strength of the F-81 skeleton by compar-ing the cross-sectional geometry of long bonediaphyses to Holocene foragers. By comparing thepattern of postcranial strength of F-81 to well-documented hunter-gatherers from the AndamanIslands and Southern Africa, we can make preli-minary inferences about the relative skeletalstrength of the individual from Wadi Mataha.
Postcranial long bone strength
The pattern of cortical bone distribution and dia-physeal strength in the lower limb is correlatedwith the intensity of terrestrial mobility among theAndamanese and Later Stone Age Southern Afri-can hunter-gatherers (Stock and Pfeiffer, 2001;Stock, 2002b). When F-81 is compared to thesegroups (Table 6), the midshafts of the F-81 femurand tibia have high cortical areas and polar secondmoments of area (J0.73). The z-score for the com-parison of the F-81 femur J0.73 and the AndamanIslander mean is 2.83, while the comparison ofJ0.73 at the tibia midshaft yields a z-score of 3.95.This suggests that the F-81 femora and tibiae havesignificantly greater diaphyseal torsional strengthsthan the semisedentary Andaman Islanders. TheF-81 J0.73 values at these locations also exceed themeans of the terrestrially mobile LSA foragers, butto a somewhat lesser extent. The midshaft corticalareas of F-81 fall slightly above the LSA means,which are in turn greater than the AI, suggestingthat the compressive strengths of the F-81 femoraand tibiae are most similar to the highly mobileLater Stone Age Southern Africans.
The Imax/Imin and Ix/Iy circularity indices of theF-81 femur are relatively low (1.28 and 1.20,respectively), reflecting the minimal developmentof the linea aspera. It was argued that Ix/Iy (Ruffet al., 1984; Ruff, 1987) and Imax/Imin (Stock andPfeiffer, 2001) circularity indices correspond withterrestrial mobility. The diaphysis of the tibia atmidshaft has a high Imax/Imin ratio (3.0) when com-pared to the other groups, illustrating significantantero-posterior rigidity of the F-81 tibial diaphy-sis. Overall, the lower limb diaphyseal strengthsand tibial midshaft Imax/Imin of F-81 are higherthan the mean values for the LSA hunter-
Fig. 3. Lateral radiograph of F-81 cranial vault. P, posterior;A, anterior.
459WADI MATAHA F-81 SKELETAL MORPHOLOGY

gatherers, which seems to correspond with consid-erable terrestrial mobility. This interpretation issomewhat contradicted by the low diaphyseal cir-cularity values of the F-81 femoral midshaft. Toinvestigate this morphology further, canonical vari-ates analysis was used to compare the overall rela-tionship between femoral and tibial diaphysisstrength and shape characteristics of the F-81 ske-leton to the terrestrially mobile LSA and semise-dentary AI hunter-gatherers. Canonical and struc-ture coefficients for the first two axes producedby this analysis are presented in Table 6, whileFigure 4 presents a plot of individual scores for thefirst two axes of the analysis. The first axisaccounts for 83.6% of the variance in cross-sec-tional properties between groups. The variable thatshows the highest loading on this axis is femurmidshaft Imax/Imin, although the structure coeffi-cients for axis 1 demonstrate that this axis corre-lates strongly with femur and tibia midshaft J0.73.This axis clearly separates the LSA from the AImales, and identifies some differentiation of thefemales, with the LSA subsamples tending towardhigher scores in both cases. Based on previous ana-lyses of the AI and LSA samples (Stock and Pfeif-fer, 2001), this axis appears to reflect differences inrobusticity between groups that is related to ter-restrial mobility. Axis 2 only accounts for a further9.5% of variation in lower limb diaphyseal strengthand shape characteristics. It partially separatesthe LSA females from the other groups, primarilyon the basis of differences in J0.73 of the femoralmidshaft and tibial midshaft shape. Within thisanalysis, F-81 falls within one standard deviationof the Later Stone Age male centroid, at the lowerend of the range of LSA male scores for canonicalaxis 1. It is clearly outside the range of variationfound among the semisedentary Andaman Islan-ders. Thus, on the basis of femur and tibia mid-shaft strength and circularity, the F-81 individualis most similar to the highly mobile LSA male sub-sample.
The pattern of cortical bone distribution and dia-physis strength in the upper body can be corre-lated with the intensity of manual activity amongforagers (Ben-Itzhak et al., 1988; Churchill, 1994;Trinkaus and Churchill, 1999; Stock and Pfeiffer,2001; Stock, 2002c; Weiss, 2003). Comparing bodysize standardized cortical areas of the humeri andulnae of F-81 to other foragers (Table 7) illustratesthat these bones tend toward high compressivestrengths, particularly in the right arm. Thetorsional strength (J0.73) of the humeri and ulnaeof F-81, when standardized for the product of esti-mated body mass and bone length, falls over onestandard deviation above the Later Stone Agemeans for all locations, and above the canoe-depen-dent Andaman Islanders for the right side. The cir-cularity of strength properties (Imax/Imin) in theF-81 long bone diaphyses tends to be lower thanthe for other groups, suggesting more generalized
TABLE
6.F-81lower
limbdiaphysea
lstrength
andcircularity
comparedto
other
hunter-gatherers,
withcoefficien
tsfrom
canon
icalvariatesanalysis1
Property
Wadi
MatahaF-81
AndamanIslanders
LaterStoneAge,
Sou
thAfrica
Axis
1:83.6%
2Axis
2:95%
2
NMea
nSD
z-score
NMea
nSD
z-score
Canon
ical
coefficien
tStructure
coefficien
tCanon
ical
coefficien
tStructure
coefficien
t
Fem
ur50%
CA
702.4
31
592.1
70.8
1.56
49
669.3
65.4
0.51
�0.423
0.437
�0.107
�0.324
J0.73
1,615.6
31
1027.9
207.9
2.83
49
1,421.3
306.4
0.63
0.351
0.659
�1.223
�0.456
I x/Iy
1.20
31
1.25
0.14
�0.36
53
1.48
0.25
�1.12
�0.508
0.584
0.115
0.105
I max/Imin
1.28
31
1.30
0.12
�0.17
49
1.56
0.24
�1.17
1.109
0.684
�0.051
0.021
Tibia
50%
CA
657.3
29
496.0
70.7
2.28
39
581.9
84.1
0.90
0.331
0.613
0.339
0.075
J0.73
1,840.3
29
971.5
220.2
3.95
39
1,362.6
385.5
1.24
0.433
0.700
0.680
0.190
I max/Imin
3.00
29
2.48
0.41
1.27
39
2.48
0.50
1.04
�0.137
0.111
0.605
0.395
1CA,corticalarea(axialcompressivestrength).J0.73¼
polarsecondmom
entof
area(torsion
alstrength).I x/Iy,I m
ax/Imin
¼diaphysealben
dingstrength
circularity.z-scores
repre-
sentrelation
ship
betwee
nWadiMatahaF-81anddistribution
ofsa
memea
suremen
tamon
gcomparativehunter-gatherer
samples.
2Percentageof
variance
explained
bycanon
icalaxis,illustratedin
Figure
5.
460 J.T. STOCK ET AL.

loading patterns of the arms, rather than repeti-tive activity that emphasizes movement in a singleplane (Schmitt et al., 2003).
The level of bilateral asymmetry in upper limbmorphology (Fig. 5) shows F-81 to have consis-tently greater asymmetry compared to the AI andLSA hunter-gatherers. At both the mid-distal loca-tion of the humerus and the midshaft of the ulna,the bilateral asymmetry in torsional strength ofF-81 is over two standard deviations above the AImeans, and in the upper end of the distributionof the Southern African Later Stone Age, withz-scores above 1.
Robusticity compared to Natufian remains:upper limb
The previous analysis suggests that the F-81male skeleton from Wadi Mataha was a highlymobile forager with strong upper limbs and a
highly asymmetric loading pattern of the upperbody. Is this pattern of skeletal robusticity of F-81consistent with morphological variability through-out the Natufian? Cross-sectional geometric prop-erties of long bone diaphyses are not currentlyavailable for other Natufian and pre-Natufian sam-
TABLE 7. F-81 upper limb robusticity compared to other hunter-gatherers1
PropertyWadi
Mataha F-81
Andaman Islanders Later Stone Age, South Africa
N Mean SD z-score N Mean SD z-score
R. humerus 35% CA 371.1 25 293.5 66.9 1.16 27 288.1 35.0 2.37J0.73 547.2 25 393.2 147.5 1.04 27 352.6 111.7 1.74Imax/Imin 1.01 25 1.26 0.16 �1.56 27 1.27 0.16 �1.63
R. ulna 50% CA 183.6 10 160.1 23.1 1.02 32 154.5 21.3 1.37J0.73 184.3 10 114.3 34.6 2.02 32 112.0 33.7 2.15Imax/Imin 1.60 10 1.70 0.23 �0.43 32 1.44 0.33 0.48
L. humerus 35% CA 313.8 25 284.4 63.2 0.47 28 268.6 33.7 1.34J0.73 412.2 25 383.8 140.8 0.20 28 297.1 79.5 1.45Imax/Imin 1.14 25 1.31 0.18 �0.94 28 1.27 0.12 �1.08
L. ulna 50% CA 165.1 10 153.9 24.8 0.45 34 145.6 19.8 0.98J0.73 137.4 10 106.5 37.8 0.82 34 98.1 27.4 1.43Imax/Imin 1.28 10 1.87 0.22 �2.68 34 1.39 0.26 �0.42
Bilateral asymmetry Humerus J 32.7 23 12.7 9.0 2.22 30 17.1 15.6 1.00Ulna J 34.1 10 11.2 8.2 2.79 28 18.4 10.0 1.57
1 CA, cortical area (axial compressive strength). J0.73 ¼ polar second moment of area (torsional strength). Imax/Imin and Ix/Iy ¼ dia-physeal bending strength circularity.
Fig. 4. Plot of canonical variates scores for lower limb dia-physeal strength and circularity values. Ellipses represent onestandard deviation from subsample centroids.
Fig. 5. Antero-posterior radiographs of F-81 humeri. L, left;R, right.
461WADI MATAHA F-81 SKELETAL MORPHOLOGY

ples, but general levels of skeletal robusticity inthe eastern Mediterranean can be compared usingexternal osteometric data.
Smith et al. (1984) demonstrated that Natufianremains from the sites of Eynan and El Wad havegreater combined cortical thickness (CCT) in thehumeral diaphyses than later Bronze age, Byzan-tine, and Early Arab populations in the Near East.Table 8 provides a comparison between the com-bined cortical thickness of the F-81 humeri andthe data published by Smith et al. (1984). Themid-distal maximum shaft diameters for the rightand left humeri of F-81 fall well within the rangeof the Natufians, and between the male and femalemeans. The antero-posterior CCT of the F-81 lefthumerus falls between the male and female Natu-fian means, while the CCT of the right is approxi-mately equal to the male mean. When the CCT isscaled to maximum shaft diameters, the F-81 lefthumerus matches the average value for Natufianmales, while the z-score for the right humerus(2.79) suggests that the F-81 cortical thickness issignificantly higher than for Natufian males. Com-paring mid-distal robusticity in the same manner,the F-81 right humerus is more robust than thesample of Natufian males (z ¼ 1.53). In contrast,the robusticity of the left humerus falls slightlybelow the Natufian mean, within the range docu-mented for male Natufians. Bilateral asymmetryin mid-distal humerus robusticity (9.55%) is abovethe male Natufian mean (z ¼ 1.08), and is consid-erably higher than the two documented Natufianfemales. The mid-distal humerus asymmetry of theF-81 skeleton also falls above the range of the LSAand AI groups (Fig. 6). In general, the identifiedmale and female Natufian skeletons show levels ofasymmetry that are similar to the other foraginggroups, although several Natufians of indetermi-
nate sex tend toward greater asymmetry. This sug-gests a considerable variability in humerus dia-physeal strength among Natufians. One Natufianindividual, the fragmentary Homo 27 skeletonfrom Hayonim, appears to have over 14% asymme-try in mid-distal humerus robusticity. Assumingthat the paired Homo 27 humeri represent oneindividual, this would also be considered an outlierwhen compared to other Natufian males.
The above comparisons suggest that the bonestrengths of the right arm, and the associated highlevel of bilateral asymmetry of the F-81 skeleton,are atypical among Natufians and other recent for-agers. If this morphological condition were relatedto pathology, we would expect one side to show evi-dence of atrophy, with lower levels of bone strengthand robusticity than normal. This does not appearto be the case, as the strength and robusticity ofthe left arm appear to fall within the normal Natu-fian range of variability. Given the lack of evidencefor any pathological conditions associated with thismorphology, we conclude that the high level ofbilateral asymmetry is behavioral in origin. Thehigh level of bilateral asymmetry in torsionalstrength (J0.73) of the F-81 humeri and ulnae, com-bined with the relative circularity of the mid-distalhumeral diaphyses, likely reflect habitual activitythat was highly right arm-dominant and general-ized in loading trajectory.
Robusticity compared to Natufian remains:lower limb
The preceding analysis suggests that the lowerlimb strengths of F-81 are in the higher end of therange of modern foragers, comparable to the ter-restrially based and highly mobile Later Stone Agepeople of Southern Africa. Are robust lower limbbones a morphological characteristic that is consis-tent throughout the Epipaleolithic? A comparisonof the external dimensions of femur and tibiadiaphyses can provide preliminary information(Table 8). The maximum and minimum diametersof the F-81 femoral midshaft fall within the rangeof, although somewhat below, the mean for Natu-fian males. Comparable measurements for Ohalo IIare above the Natufian range. When these arescaled to femur length to calculate midshaft robus-ticity, the differences between F-81 and Ohalo IIdisappear, and both have robusticity indices of12.8. This demonstrates that despite great size dif-ferences between these two individuals, they sharea femoral robusticity that is somewhat higher thanthe Natufian means. The maximum and minimumdiameters of the F-81 tibia at the cnemic foramenfall within the range of male Natufians, but theindex of robusticity calculated when these valuesare scaled to bone length falls significantly abovethe Natufian male mean (z ¼ 3.0). These resultssuggest that the F-81 skeleton is more robust
Fig. 6. Bilateral asymmetry in mid-distal humerus robusti-city. þ, outliers (1.5–3x interquartile range); �, extreme cases(>3x interquartile range).
462 J.T. STOCK ET AL.

than would be expected for a Natufian of the samebody size.
DISCUSSION
Analysis of the F-81 male skeleton from WadiMataha highlights several aspects of its morphol-ogy that appear distinctive in the context of varia-bility in the Levant and among geographicallydispersed hunter-gatherers. The F-81 cranium andmandible have a combination of features that areshared with both pre-Natufian and Natufianskulls, including orbits that are low, square, andnarrow, rounded frontal squama, and reducedsuperciliary arches. However, F-81 differs in othermorphological characteristics shared with earlierremains, including aspects of cranial shape. Thepre-Natufian crania feature lower and wider cra-nial vaults when compared to the relatively highand long vaults of the Natufian crania. The cranialvault of F-81 is thick when compared to the rangefound within modern humans. It is unknownwhether other Epipaleolithic and Natufian remainsfrom the Levant share this characteristic.
The postcranial skeleton of F-81 is distinctive.The most apparent and pervasive of these uniquetraits are small body size combined with highlevels of skeletal strength/robusticity and bilateralasymmetry. The long bone lengths and resultingbody size estimates of the F-81 skeleton dramati-cally extend the known range of adult body size inthe Levantine Epipaleolithic. The F-81 postcranialskeleton shows some morphological similarities toOhalo II and Nahal Ein Gev 1, including signifi-cant skeletal robusticity and high levels of bilateralasymmetry.
The pattern of long bone diaphyseal strengthand circularity of F-81 is most similar to LaterStone Age Southern African males for the lowerlimb. As such, it appears to correspond with a highlevel of terrestrial mobility, reflecting rugged ter-rain, extensive geographic range, or both. The F-81humeral and ulnar diaphyseal strengths generallyexceed both the LSA and AI samples, suggestingmanually intense physical activity. However, themost pronounced morphological characteristic ofthe upper limbs is the high level of bilateral asym-metry. This suggests considerable lateralization ofhabitual loading of the arms. When the pattern ofpostcranial robusticity is considered with the evi-dence that the cranial vault of F-81 is also quitethick, it suggests a high level of systemic robusti-city, perhaps as a result of intense physical activ-ity. It appears that there may be a reduction in themanifestation of these characteristics from the pre-Natufian to Natufian periods.
The diaphyseal robusticity of F-81 appears to bea consequence of unusually high cortical thicknessin most regions of the skeleton, resulting from bothmedullary stenosis and periosteal apposition of cor-tical bone. Considerable humeral bilateral asym-metry appears to be characteristic of UpperPalaeolithic (UP) skeletons in general (Trinkauset al., 1994; Trinkaus and Churchill, 1999; Schmittet al., 2003), including individual remains from cir-cum-Mediterranean sites such as Barma Grande(Churchill and Formicola, 1997), Wadi Kubbaniya(Angel and Kelley, 1986) and Ohalo II (Herskovitzet al., 1993). While the significant bilateral asym-metry of Ohalo II and Barma Grande were inter-preted as correlates of pathological conditions(Herskovitz et al., 1993; Churchill and Formicola,1997), the general pattern of asymmetry among
TABLE 8. F-81 diaphyseal morphology compared to Ohalo II and Natufians
SideWadi
Mataha F-81 Ohalo II H23Natufian males Natufian females
N Mean SD z-score N Mean SD z-score
Humerus mid-distalMaximum shaft diameter1 R 18.1 10 18.5 1.2 �0.33 9 16.3 1.3 1.38
L 16.9 11 18.1 1.3 �0.92 12 16.3 1.0 0.60CCT1 R 12.3 10 12.1 1.2 0.17 9 10.0 1.7 1.35
L 10.5 11 11.3 1.9 �0.42 12 8.9 1.7 0.94CCT/diameter1 R 6.79 10 6.59 0.07 2.86 9 6.10 0.09 7.67
L 6.21 11 6.21 0.08 0.00 12 5.41 0.09 8.89Mid-distal robusticity2 R 12.73 10 11.89 0.55 1.53 4 11.63 0.90 1.22
L 11.63 7 11.91 0.64 �0.44 3 11.52 0.86 0.13BA 9.55 6 3.97 5.15 1.08 2 0.36 0.28 32.82
Femur midshaftMaximum diameter 26.8 37 9 29.8 1.5 �2.00 6 26.6 2.8 0.07Minimum diameter 23.5 26 9 24.3 0.7 �1.14 5 22.9 1.3 0.46Robusticity 12.8 12.8 8 12.4 0.8 0.50 4 12.3 1.4 0.36
TibiaCnemic maximum diameter 37.7 10 37.0 1.3 0.54 4 32.1 2.0 2.80Cnemic minimum diameter 22.7 10 22.9 2.3 �0.09 4 22.6 2.2 0.05Cnemic robusticity 18.3 10 16.5 0.6 3.00 4 16.5 0.5 3.60
1 Measurements from Smith et al. (1984).2 Natufian mid-distal robusticity measurements taken by I.T.S., including remains from Nahal Oren, Hayonim, Eynan, and Rakefet.3 A-P and M-L diaphyseal breadths from Hershkovitz et al. (1995).
463WADI MATAHA F-81 SKELETAL MORPHOLOGY

most UP skeletons appears to be a mechanical con-sequence of patterns of habitual hunting and gath-ering. The current study provides evidence thatthis pattern of high bilateral asymmetry mayextend into the Middle Epipaleolithic/GeometricKebaran period in the Levant, with a subsequentincrease in symmetry between humeral antimeresin the Natufian.
In the morphology of the postcrania and skull,the F-81 skeleton increases the range of variabilityknown from the periods that predate the Natufianin the Levant. The morphological similarities anddifferences among the remains from Ohalo II,Nahal Ein Gev, Ein Gev, Qasr Kharaneh, and WadiMataha suggest: 1) considerable morphologicalvariability prior to the Natufian; 2) continuitybetween the pre-Natufian and Natufian popula-tions; and 3) a general reduction in intensity ofhabitual behavior between these periods. Moredetailed analyses of UP and Epipaleolithic remainsfrom the Levant will shed light on the biologicaland behavioral relationships between these andsubsequent Natufian populations.
ACKNOWLEDGMENTS
The authors thank Baruch Arensburg for allow-ing J.T.S. to study the Natufian collections at theSackler School of Medicine in Tel Aviv, and forreading an earlier draft of this paper. We alsothank Cheryl Katzmarzyk for her work preparingthe skeleton, and two anonymous reviewers forcomments and suggestions that improved thispaper. This research was funded by a grant toM.C. from the Social Sciences and HumanitiesResearch Council of Canada.
LITERATURE CITED
Angel JL, JO Kelley. 1986. Description and comparison of the ske-leton. In: Wendorf F, Schild R, editors. The Wadi Kubbaniyaskeleton: a Late Paleolithic burial from southern Egypt.Volume 1. Dallas: Southern Methodist University Press. p 53–70.
Arensburg B. 1977. New Upper Paleolithic human remains fromIsrael. Eretz Israel 13:208–215.
Arensburg B. 1981. Recent evolution in Israel. Colloques inter-nationaux du C.N.R.S. N. 599—les processus d l’hominisation.16–20 Juin, 1980. Paris: Editions du CNRS. p 195–201.
Arensburg B, Bar-Yosef O. 1973. Human remains from EinGev I, Jordan Valley, Israel. Paleorient 1:201–206.
Arensburg B, Bar-Yosef O, Belfer-Cohen A, Rak Y. 1990. Mous-terian and Aurignacian human remains from Hayonim Cave,Israel. Paleorient 16:107–109.
Bar-Yosef O. 1998. The Natufian culture in the Levant: thresh-old to the origins of agriculture. Evol Anthropol 6:159–177.
Belfer-Cohen A, Schepartz LA, Arensburg B. 1991. New biologi-cal data for the Natufian populations in Israel. Ann Arbor:International Monographs in Prehistory.
Ben-Itzhak S, Smith P, Bloom RA. 1988. Radiographic study ofthe humerus in Neandertals and Homo sapiens sapiens. AmJ Phys Anthropol 77:231–242.
Bocquentin F. 2003. Burial practices, biological factors and cul-tural identities during the Natufian period: a bioarchaeologi-cal perspective. Ph.D. dissertation, Universite de Bordeaux.
Churchill SE. 1994. Human upper body evolution in the Eura-sian Later Pleistocene. Ph.D. dissertation, University of NewMexico.
Churchill SE, Formicola V. 1997. A case of marked bilateralasymmetry in the upper limbs of an Upper Palaeolithic malefrom Barma Grande (Liguria), Italy. Int J Osteoarch 7:18–38.
Ferembach D. 1977. Les Natoufiens de Palestine. In: Arensburg B,Bar-Yosef O, editors. Stekelis memorial volume. Jerusalem:Israel Exploration Society. p 241–252.
Garrod DAE. 1957. The Natufian culture: the life and economy of aMesolithic people in the Near East. Proc Br Acad 43:211–227.
Goring-Morris AN, Belfer-Cohen A. 1998. The articulation ofcultural processes and Late Quaternary environmentalchanges in Cisjordan. Paleorient 23:71–93.
Haapasalo H, Sievanen H, Kannus P, Heinonen A, Oja P, VuoriI. 1996. Dimensions and estimated mechanical characteristicsof the humerus after long-term tennis loading. J Bone MinerRes 11:864–872.
Hershkovitz I, Edelson G, Spiers M, Arensburg B, Nadel D, LeviB. 1993. Ohalo II man—unusual findings in the anterior ribcage and shoulder girdle of a 19,000-year-old specimen. Int JOsteoarch 3:177–188.
Hershkovitz I, Speirs MS, Frayer D, Nadel D, Wishbaratz S,Arensburg B. 1995. Ohalo-II H2—a 19,000-year-old skeletonfrom a water-logged site at the Sea of Galilee, Israel. AmJ Phys Anthropol 96:215–234.
Holliday TW. 1997. Body proportions in Late Pleistocene Europeand modern human origins. J Hum Evol 32:423–447.
Kaufman D, Ronen A. 1987. Le sepulture Kebarienne Geometri-que de Neve David, Haifa, Israel. Anthropologie 91:335–342.
Lahr MM, Arensburg B. 1995. Skeletal robusticity in the Epipa-leolithic of North Africa and the Levant. Paleorient 21:87–96.
Lieberman DE. 1996. How and why humans grow thin skulls:experimental evidence for systemic cortical robusticity. AmJ Phys Anthropol 101:217–236.
Martin R, Saller K. 1957. Lehrbuch der antropologie. Stuttgart:Gustav Fischer Verlag.
Nawrocki SP. 1992. A biomechanical model of cranial vaultthickness in archaic Homo. Ph.D. dissertation, State Univer-sity of New York at Binghampton.
Olivier G. 1969. Practical anthropology. Springfield, IL: CharlesC. Thomas.
Peterson J. 1997. Tracking activity patterns through skeletalremains: a case study from Jordan and Palestine. In: GebelHGK, Kafafi Z, Rollefson GO, editors. The prehistory of Jordan,II. Perspectives from 1997. Berlin: Ex Oriente. p 475–492.
Peterson J. 2002. Sexual revolutions: gender and labor at thedawn of agriculture. New York: Altamira Press.
Rolston SL. 1992. Two prehistoric burials from Qasr Kharaneh.Annual of the Department of Antiquities. Amman: HashemiteKingdom of Jordan. p 221–229.
Ruff CB. 1987. Sexual dimorphism in human lower limb bonestructure: relationship to subsistence strategy and sexualdivision of labour. J Hum Evol 16:391–416.
Ruff CB. 1992. Age changes in endosteal and periosteal sensitiv-ity to increased mechanical loading. Abstract. Transactions ofthe 38th Annual Meeting of the Orthopaedic Research Society,Washington, DC. 17:532.
Ruff CB. 2000a. Biomechanical analyses of archaeologicalhuman skeletons. In: Katzenberg MA, Saunders SR, editors.Biological anthropology of the human skeleton. Toronto:Wiley-Liss. p 71–102.
Ruff CB. 2000b. Body size, body shape, and long bone strengthin modern humans. J Hum Evol 38:269–290.
Ruff CB, Larsen CS, Hayes WC. 1984. Structural changes inthe femur with the transition to agriculture on the Georgiacoast. Am J Phys Anthropol 64:125–136.
Ruff CB, Trinkaus E, Holliday TW. 1997. Body mass and ence-phalization in Pleistocene Homo. Nature 387:173–176.
Schmitt D, Churchill SE, Hylander WL. 2003. Experimental evi-dence concerning spear use in Neandertals and early modernhumans. J Archaeol Sci 30:103–114.
Scott EC. 1979. Dental wear scoring technique. Am J PhysAnthropol 51:213–218.
464 J.T. STOCK ET AL.

Smith P. 1970. Dental morphology and pathology in the Natu-fians: the dental evidence for dietary specialization. Ph.D.dissertation, University of Chicago.
Smith P, Bloom RA, Berkowitz J. 1984. Diachronic trends inhumeral cortical thickness of Near Eastern populations.J Hum Evol 13:603–611.
Southon J. 2002. A first step to reconciling the GRIP andGISP2 ice-core chronologies, 0–14,500 yr BP. Quatern Res57:32–37.
Stock JT. 2002a. A test of two methods of radiographically derivinglong bone cross-sectional properties compared to direct section-ing of the diaphysis. Int J Osteoarch 12:335–342.
Stock JT. 2002b. Climate, terrestrial mobility, and the pattern-ing of lower limb robusticity among Holocene foragers. AmJ Phys Anthropol [Suppl]34:148–149.
Stock JT. 2002c.Climatic and behavioral influences on postcra-nial robusticity among Holocene foragers. Ph.D. dissertation,University of Toronto.
Stock J, Pfeiffer S. 2001. Linking structural variability in longbone diaphyses to habitual behaviors: foragers from theSouthern African Later Stone Age and the Andaman Islands.Am J Phys Anthropol 115:337–348.
Stock JT, Pfeiffer SK. 2004. Long bone robusticity and subsis-tence behaviour among Later Stone Age foragers of the forest
and fynbos biomes of South Africa. J Archaeol Sci 31:999–1013.
Stuiver M, Reimer PJ, Bard E, Beck JW, Burr GS, Hughen KA,Kromer B, McCormac G, Van der Plicht J, Spurk M. 1998.INTCAL98 radiocarbon age calibration, 24,000–0 cal BP. Radio-carbon 40:1041–1083.
Trinkaus E. 1997. Appendicular robusticity and the paleobiologyof modern human emergence. Proc Natl Acad Sci USA94:13367–13373.
Trinkaus E, Churchill SE. 1999. Diaphyseal cross-sectional geo-metry of Near Eastern Middle Paleolithic humans: thehumerus. J Archaeol Sci 26:173–184.
Trinkaus E, Ruff C. 1989. Diaphyseal cross-sectional morphol-ogy and biomechanics of the Fond-de-Foret 1 femur and theSpy 2 femur and tibia. Bull Soc R Belge Anthropol Prehist100:33–42.
Trinkaus E, Churchill SE, Ruff CB. 1994. Postcranial robusti-city in Homo. II: humeral bilateral asymmetry and bone plas-ticity. Am J Phys Anthropol 93:1–34.
Trotter M. 1970. Estimation of stature from intact long limbbones. In: Stewart TD, editor. Personal indentification in massdisasters. Washington, DC: Smithsonian Institute. p 71–83.
Weiss E. 2003. Effects of rowing on humeral strength. AmJ Phys Anthropol 121:293–302.
465WADI MATAHA F-81 SKELETAL MORPHOLOGY