ex vivo endothelin dependent contraction of the remodeled rat spiral artery
TRANSCRIPT

lable at ScienceDirect
Placenta xxx (2014) 1e7
Contents lists avai
Placenta
journal homepage: www.elsevier .com/locate/placenta
Ex vivo endothelin dependent contraction of the remodeled rat spiralartery
I. Ariel a, *, G. Skarzinski a, T. Kossovsky a, V. Belzer b, D. Knigin a, M. Khamaisi c, d,Z. Abassi d, 1, M. Bursztyn e, 1
a The Department of Pathology, Hadassah-Hebrew University Medical Center, Jerusalem, Israelb The Laboratory of Experimental Surgery, Hadassah-Hebrew University Medical Center, Jerusalem, Israelc Institute of Endocrinology, Diabetes & Metabolism & Internal Medicine C, Rambam Medical Center, Haifa, Israeld RB Rappaport Faculty of Medicine-Technion, Haifa, Israele The Department of Medicine, Hadassah-Hebrew University Medical Center, Jerusalem, Israel
a r t i c l e i n f o
Article history:Accepted 4 July 2014
Keywords:Rat placentaEndovascular trophoblastSmooth muscle differentiationEndothelin systemMyotrophoblastRemodeled spiral artery
* Corresponding author. Department of Pathology,Medical Center, Mount Scopus, PO Box 24035, Jerusal2 5844788; fax: þ972 2 5324168.
E-mail addresses: [email protected], ariel@ha1 Sharing equal contribution.
http://dx.doi.org/10.1016/j.placenta.2014.07.0050143-4004/© 2014 Elsevier Ltd. All rights reserved.
Please cite this article in press as: Ariel I, ethttp://dx.doi.org/10.1016/j.placenta.2014.07.
a b s t r a c t
Introduction: Similarities of the rat to the human placenta make rat pregnancy models relevant to thestudy of human gestational diseases. Understanding of species differences is necessary to extrapolatefrom animal models to humans. We observed alpha-smooth muscle actin (aSMA) expression in ratendovascular trophoblasts (EVasT) and investigated the spatial and temporal expression of smoothmuscle (SM) proteins and their potential function in remodeled spiral artery.Methods: Rat placentas were examined from gestational day 13 to term, and were immunostained forcytokeratin, aSMA, alpha heavy chain of SM myosin, non-muscle myosin, Rho proteins, regulators of SMgene expression, myocardin, an early marker of SM differentiation and endothelin receptors A and B(ETA, ETB). Transmission electron microscopy (TEM) was performed. Modified spiral artery rings werestudied ex vivo for endothelin-1- induced contraction.Results: EVasT expressed SM proteins co-localizing with cytokeratin confirming their trophoblasticorigin from gestational day 13 to term. Thin fibers, consistent with actin fibers, were observed by TEM, inthe cellular localization of aSMA in EVasT.Functional experiments revealed that addition of 10�7 M endothelin-1 ex vivo reduced vascular lumenarea by 11.1% ± 1.8% compared with control. This effect was reduced to only 1.0 ± 1.7% with ETAantagonist, and to 5.4 ± 1.7% contraction by ETB antagonist, p < 0.002, for all.Discussion: The expression of SM proteins in EVasT along with the contractibility of the rat remodeledspiral artery ex vivo, suggest that some vascular tone is potentially maintained by endothelin-1, and mayplay a role in situations of dysregulation of the vasoactive systems.
© 2014 Elsevier Ltd. All rights reserved.
1. Introduction
The conspicuous similarities of the rat to the human placenta, inparticular hemochorial placentation associated with deep tropho-blastic invasion [1,2] make rat models attractive to the study ofdiseases of pregnancy. The investigation of mechanisms controllinguteroplacental circulation in the rat may shed light on similarmechanisms in humans, or alternatively, may elucidate the
Hadassah-Hebrew Universityem il 91240, Israel. Tel.: þ972
dassah.org.il (I. Ariel).
al., Ex vivo endothelin depen005
differences and hence the limitations of this animal as a model forhuman gestational diseases.
Endovascular trophoblasts (EVasT) are a special subset of theinvasive trophoblast that have the ability to remodel the spiral ar-teries. In the human spiral artery, remodeling is essential forestablishing adequate uteroplacental blood supply, and its disrup-tion is associated with major gestational diseases, such as pre-eclampsia and intrauterine growth restriction (IUGR) [2e4].
In studying the vasoactive systems in the maternal hyper-insulinemia IUGR ratmodel, wewere intrigued by the expression ofendothelin converting enzyme-1 (ECE-1) and endothelin receptorA (ETA) in EVasT [5]. This was especially interesting in view of ourprevious findings of the expression of nitric oxide synthase (NOS)isoforms in these cells [6]. Studying the morphology of the
dent contraction of the remodeled rat spiral artery, Placenta (2014),
I. Ariel et al. / Placenta xxx (2014) 1e72
mesometrial triangle (MT) in our IUGR rat model [6] we observedexpression of alpha smooth muscle actin (aSMA) in EVasT thatreplace the vascular smooth muscle (SM) in the spiral artery. Thisled us to investigate the spatial and temporal expression of aSMAand other SM proteins and their potential biological role in EVasT.We hypothesized that in the rat, by acquiring SM protein expres-sion EVasT partially replace the function of SM cells in the wall ofthe spiral artery, thus contributing to dysregulation of the vasoac-tive systems in pathological conditions.
2. Materials and methods
2.1. Animal model
Female Wistar rats were handled and housed according to the guidelines andapproval of the Committee for the Care of Laboratory Animals of the Hebrew
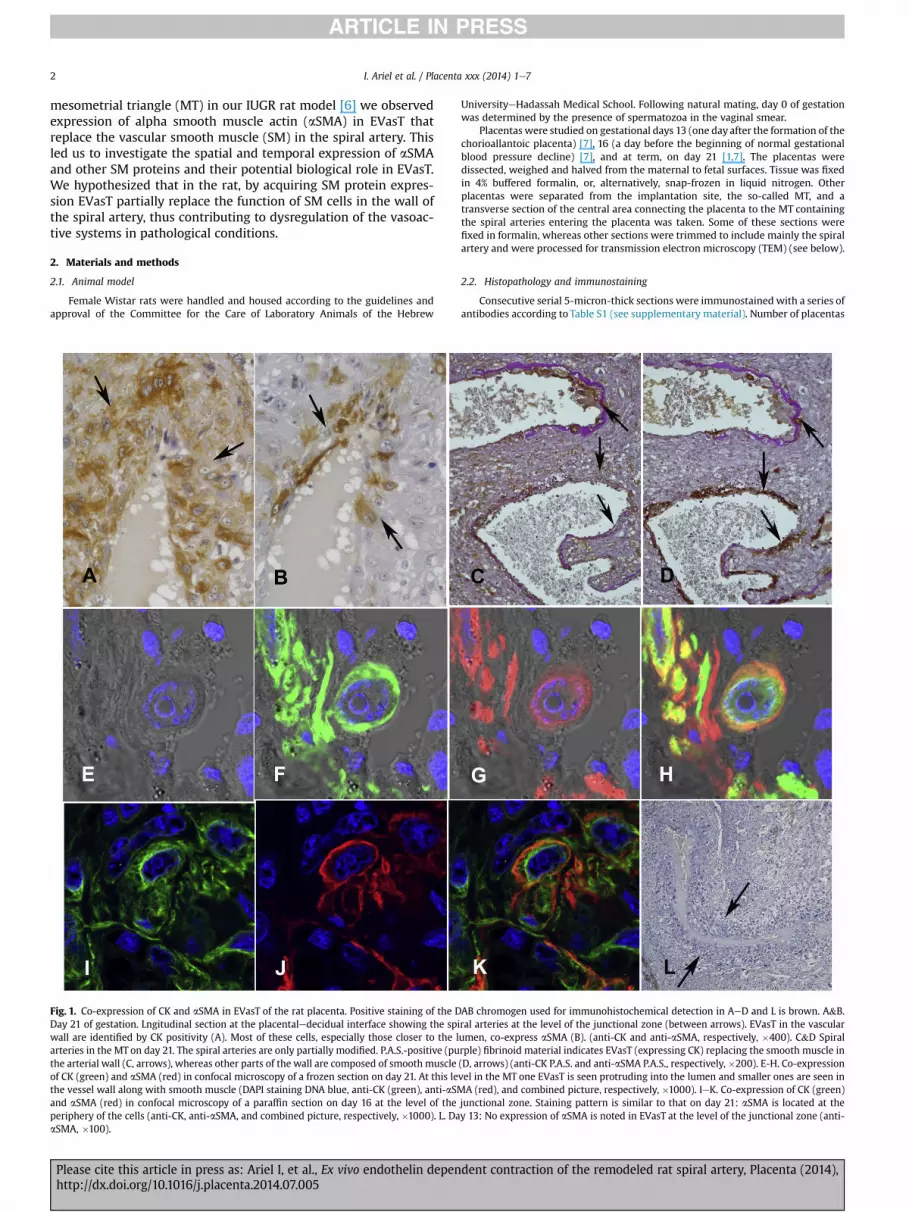
Fig. 1. Co-expression of CK and aSMA in EVasT of the rat placenta. Positive staining of the DDay 21 of gestation. Lngitudinal section at the placentaledecidual interface showing the spwall are identified by CK positivity (A). Most of these cells, especially those closer to the larteries in the MT on day 21. The spiral arteries are only partially modified. P.A.S.-positive (puthe arterial wall (C, arrows), whereas other parts of the wall are composed of smooth muscleof CK (green) and aSMA (red) in confocal microscopy of a frozen section on day 21. At this lethe vessel wall along with smooth muscle (DAPI staining DNA blue, anti-CK (green), anti-aSMand aSMA (red) in confocal microscopy of a paraffin section on day 16 at the level of theperiphery of the cells (anti-CK, anti-aSMA, and combined picture, respectively, �1000). L. DaaSMA, �100).
Please cite this article in press as: Ariel I, et al., Ex vivo endothelin depenhttp://dx.doi.org/10.1016/j.placenta.2014.07.005
UniversityeHadassah Medical School. Following natural mating, day 0 of gestationwas determined by the presence of spermatozoa in the vaginal smear.
Placentas were studied on gestational days 13 (one day after the formation of thechorioallantoic placenta) [7], 16 (a day before the beginning of normal gestationalblood pressure decline) [7], and at term, on day 21 [1,7]. The placentas weredissected, weighed and halved from the maternal to fetal surfaces. Tissue was fixedin 4% buffered formalin, or, alternatively, snap-frozen in liquid nitrogen. Otherplacentas were separated from the implantation site, the so-called MT, and atransverse section of the central area connecting the placenta to the MT containingthe spiral arteries entering the placenta was taken. Some of these sections werefixed in formalin, whereas other sections were trimmed to include mainly the spiralartery and were processed for transmission electron microscopy (TEM) (see below).
2.2. Histopathology and immunostaining
Consecutive serial 5-micron-thick sections were immunostained with a series ofantibodies according to Table S1 (see supplementary material). Number of placentas
AB chromogen used for immunohistochemical detection in AeD and L is brown. A&B.iral arteries at the level of the junctional zone (between arrows). EVasT in the vascularumen, co-express aSMA (B). (anti-CK and anti-aSMA, respectively, �400). C&D Spiralrple) fibrinoid material indicates EVasT (expressing CK) replacing the smooth muscle in(D, arrows) (anti-CK P.A.S. and anti-aSMA P.A.S., respectively, �200). E-H. Co-expressionvel in the MT one EVasT is seen protruding into the lumen and smaller ones are seen inA (red), and combined picture, respectively, �1000). IeK. Co-expression of CK (green)
junctional zone. Staining pattern is similar to that on day 21: aSMA is located at they 13: No expression of aSMA is noted in EVasT at the level of the junctional zone (anti-
dent contraction of the remodeled rat spiral artery, Placenta (2014),

I. Ariel et al. / Placenta xxx (2014) 1e7 3
immunostained with each antibody at different gestational ages is summarized inTable S1. Following heat induced epitope retrieval in citrate buffer pH 6.0, endoge-nous peroxidase and non-specific staining were blocked by 3% hydrogen peroxideand Background Buster (Innovex, USA), respectively.
Antibodies were detected with ZytoChem Plus HRP Polymer anti-Mouse andZytoChem Plus HRP Polymer anti-Rabbit (Zytomed, Germany), followed by dia-minobenzidine tetrahydrochloride (DAB) Chromogen System (SIGNET, Covance,England) or the Permanent AEC Kit (Zytomed, Germany), visualized as brownand red staining, respectively. Each staining procedure included a negativecontrol.
Double immunofluorescence staining was performed on frozen and paraffinsections using a mixture of anti-aSMA and anti-cytokeratin (CK) antibodies,mounted with mounting medium containing DAPI (SouthernBiotech, USA), andmicrographs were obtained with a confocal microscope Zeiss LSM 710 AxioObserver.Z1with a 63 � /1.4 Oil DIC M27 lens (Zeiss, Jena, Germany).
2.3. TEM
Tissues were fixed with 2.5% glutaraldehyde, embedded in Epon mix (SPI,USA), sectioned and mounted on copper grids. Grids were stained with uranylacetate and lead citrate and examined by TEM (JEM-100C) in parallel to SPI crossgrating grid (2160 lines/mm). Measurements were determined under differentmagnifications (�16,000, �33,000 and �50,000) by determining fiber diameter(mean of 10 separate fibers) and compared with the grating grid for accuratemagnification.
2.4. Ex vivo experiment
Placentas on day 21 were dissected, separated from the MT and a 1 mm thicktransverse section of the placenta at the maternal surface was dissected to include ashort segment of the spiral artery. The sections were fixed to a silicon-coated 8-wellchamber slide (Ibidi) in oxygenated Krebs solution. Under laser scanning confocalmicroscope (Olympus FluoView™ FV1000) ETA and ETB inhibitors (BQ-123 and BQ-788, respectively, AldricheSigma) at a concentration of 10�6 M were added to someof the wells, followed by the addition of endothelin-1 (ET-1)(AldricheSigma) at10�7 M to wells with and without inhibitors. Spiral artery and a segment of thefemoral artery with and without ET-1 were used as controls.
The concentration of ET-1 used in our study is within the range used in theliterature for similar ex vivo experiments with rat vascular rings [8]. Previous studieshave shown that comparable concentrations of the currently applied doses of ETAand ETB blockers were sufficient to block the biological actions of ET-1(10�11e10�7 M) on isolated perfused aortic arteries or pulmonary artery [9,10].Therefore, we utilized theses validated doses to ensure complete blockade of eachendothelin receptor subtype in a selective manner.
We measured the surface area of the lumen of 19 spiral arteries (from 8 rats)exposed to ET-1 only, 6 arteries (from 5 rats) exposed to ET-1 and ETA antagonist, 6arteries (from 3 rats) exposed to ET-1 and ETB antagonist, and 11 spiral arteries(from 8 animals) were used as controls.
Surface area of the lumen was measured using ImageJ software. Variance sta-tistical analysis was performed using 2-way ANOVA for repeated measures SSPSversion 20 software.
Following the ex vivo experiment the specimens were put in formalin, processedand embedded in paraffin, sectioned and subjected to double immunofluorescentstaining for CK and aSMA, as above, to verify the trophoblastic nature of the cells inthe vessel wall.
Fig. 2. Transmission electron microscopy of EVasT at the placental decidual interface. A. Tranvessel with fibrinoid material in the wall (�5100). B Bundles of thin fibers located at the perithe fibers (between arrows is compatible with actin (�125,000).
Please cite this article in press as: Ariel I, et al., Ex vivo endothelin depenhttp://dx.doi.org/10.1016/j.placenta.2014.07.005
3. Results
3.1. Histopathology, immunostaining and TEM
Transverse sections of the placenta at the placental mesometrialjunction on day 21 revealed 1-2 sets spiral arteries (also designatedcentral canals at this level) (Fig. 1AeB). The artery is easily recog-nized by thick wall at this level which is composed only of largecells identified as trophoblasts by positive immunostaining withanti-CK (Fig. 1A), enmeshed in fibrinoid material. In mid-verticalsections along the placentalemesometrial axis, EVasT in deeperbranches of the spiral artery within the MT usually replace onlypart of the vascular wall circumference, whereas the SM cells areretained in the rest of the vascular wall (Fig. 1CeD) [6,11].
Expression of aSMA co-localized with CK expression in EVasT(Fig. 1AeB). aSMAwas observed at the periphery of the cytoplasm,whereas CK was distributed more evenly in the cytoplasm andtherefore was more prominent at the center of the cell in thecombined immunofluorescence picture (Fig. 1EeH). Staining in-tensity for aSMA varied from cell to cell, and tended to be strongerin trophoblasts closer to the lumen (Fig. 1AeB). The expression ofaSMA in trophoblastic cells, which was performed on paraffinembedded and frozen tissue (Fig. 1EeH), was confirmed by im-munostaining with two different monoclonal antibodies. Furtherconfirmation was the electron microscopic detection of bundles ofintracellular thin fibers consistent with actin fibers at the cell pe-riphery in EVasT (Fig. 2) lining the vascular lumen, in parallel to thelocalization of aSMA as viewed by confocal microscopy (Fig. 1H).
Strong expression of aSMA was seen in the myometrium at themargins of the MT as well as in fetal vessels in the labyrinth andbranches of the spiral artery deep in the MT. Expression of aSMAwas not seen in any other type of trophoblast, notably in theinvading interstitial trophoblasts in the MT, except for a few scat-tered giant trophoblasts in the junctional zone (not shown).
On day 16, co-expression of CK and aSMA in EVasT in the wall ofthe central artery leading from theMT into the placentawas similarto that on day 21 (Fig. 1IeK). Expression of aSMA in EVasT demon-strated a gradient from the stronger immunostaining in and aroundthe central artery to weaker in EVasT deeper in the MT. On day 13weak to non-discernible expression of aSMAwas seen in the EVasTin the junctional zone (Fig. 1L). Deeper in the MT, weak expressionwas seen in theminority of EVasT relative to the strongexpression ofsmooth muscle fiber retained in the vascular wall (not shown).
Expression of other contractile proteins, i.e. smooth musclemyosin (SMM) and non-muscle myosin (NMM), was observed in
sverse section of the central artery showing large EVasT pave the luminal surface of thephery of the EVasT at the placental decidual interface (arrows) (�26,500). C. The size of
dent contraction of the remodeled rat spiral artery, Placenta (2014),
I. Ariel et al. / Placenta xxx (2014) 1e74
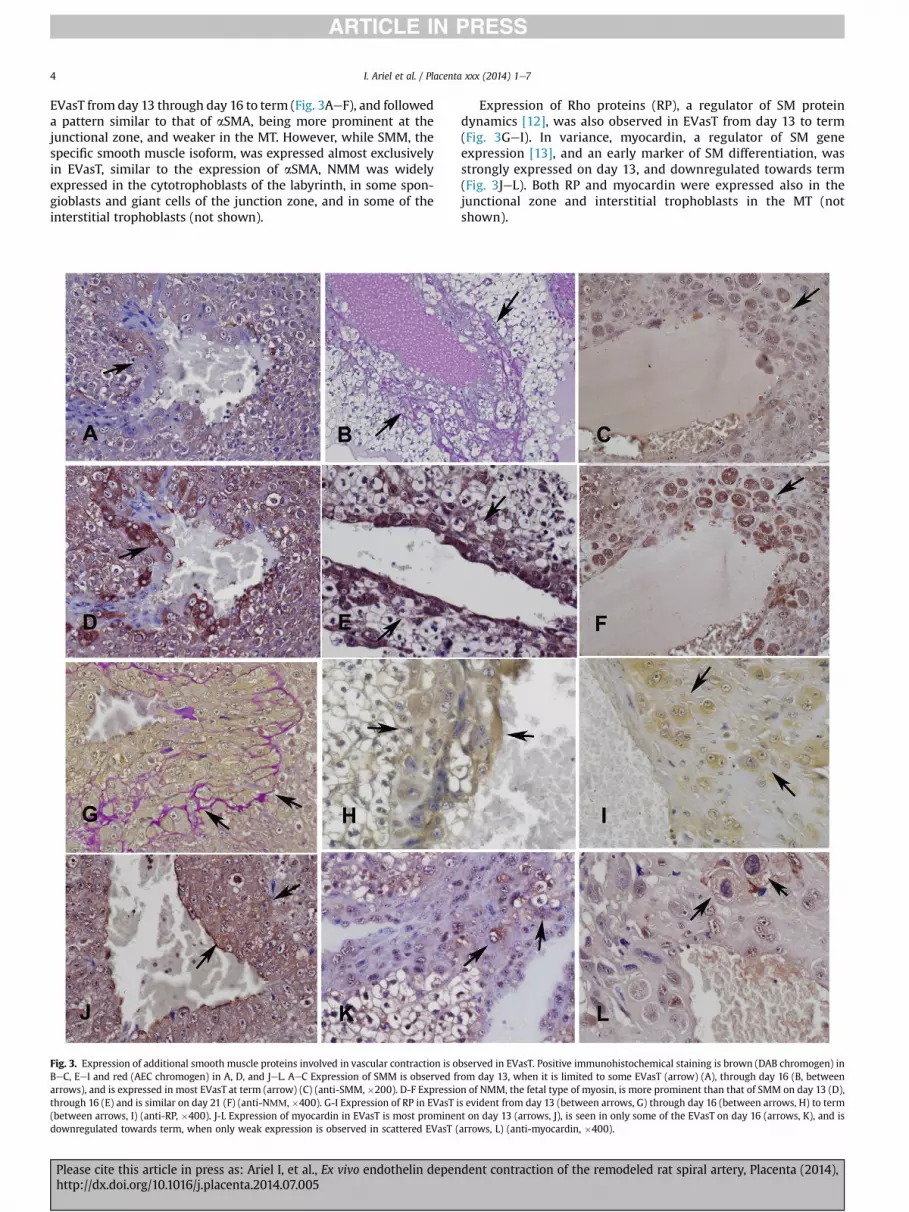
EVasT from day 13 through day 16 to term (Fig. 3AeF), and followeda pattern similar to that of aSMA, being more prominent at thejunctional zone, and weaker in the MT. However, while SMM, thespecific smooth muscle isoform, was expressed almost exclusivelyin EVasT, similar to the expression of aSMA, NMM was widelyexpressed in the cytotrophoblasts of the labyrinth, in some spon-gioblasts and giant cells of the junction zone, and in some of theinterstitial trophoblasts (not shown).
Fig. 3. Expression of additional smooth muscle proteins involved in vascular contraction is oBeC, EeI and red (AEC chromogen) in A, D, and JeL. AeC Expression of SMM is observed frarrows), and is expressed in most EVasT at term (arrow) (C) (anti-SMM, �200). D-F Expressiothrough 16 (E) and is similar on day 21 (F) (anti-NММ, �400). G-I Expression of RP in EVasT i(between arrows, I) (anti-RP, �400). J-L Expression of myocardin in EVasT is most prominendownregulated towards term, when only weak expression is observed in scattered EVasT (
Please cite this article in press as: Ariel I, et al., Ex vivo endothelin depenhttp://dx.doi.org/10.1016/j.placenta.2014.07.005
Expression of Rho proteins (RP), a regulator of SM proteindynamics [12], was also observed in EVasT from day 13 to term(Fig. 3GeI). In variance, myocardin, a regulator of SM geneexpression [13], and an early marker of SM differentiation, wasstrongly expressed on day 13, and downregulated towards term(Fig. 3JeL). Both RP and myocardin were expressed also in thejunctional zone and interstitial trophoblasts in the MT (notshown).
bserved in EVasT. Positive immunohistochemical staining is brown (DAB chromogen) inom day 13, when it is limited to some EVasT (arrow) (A), through day 16 (B, betweenn of NMM, the fetal type of myosin, is more prominent than that of SMM on day 13 (D),s evident from day 13 (between arrows, G) through day 16 (between arrows, H) to termt on day 13 (arrows, J), is seen in only some of the EVasT on day 16 (arrows, K), and isarrows, L) (anti-myocardin, �400).
dent contraction of the remodeled rat spiral artery, Placenta (2014),
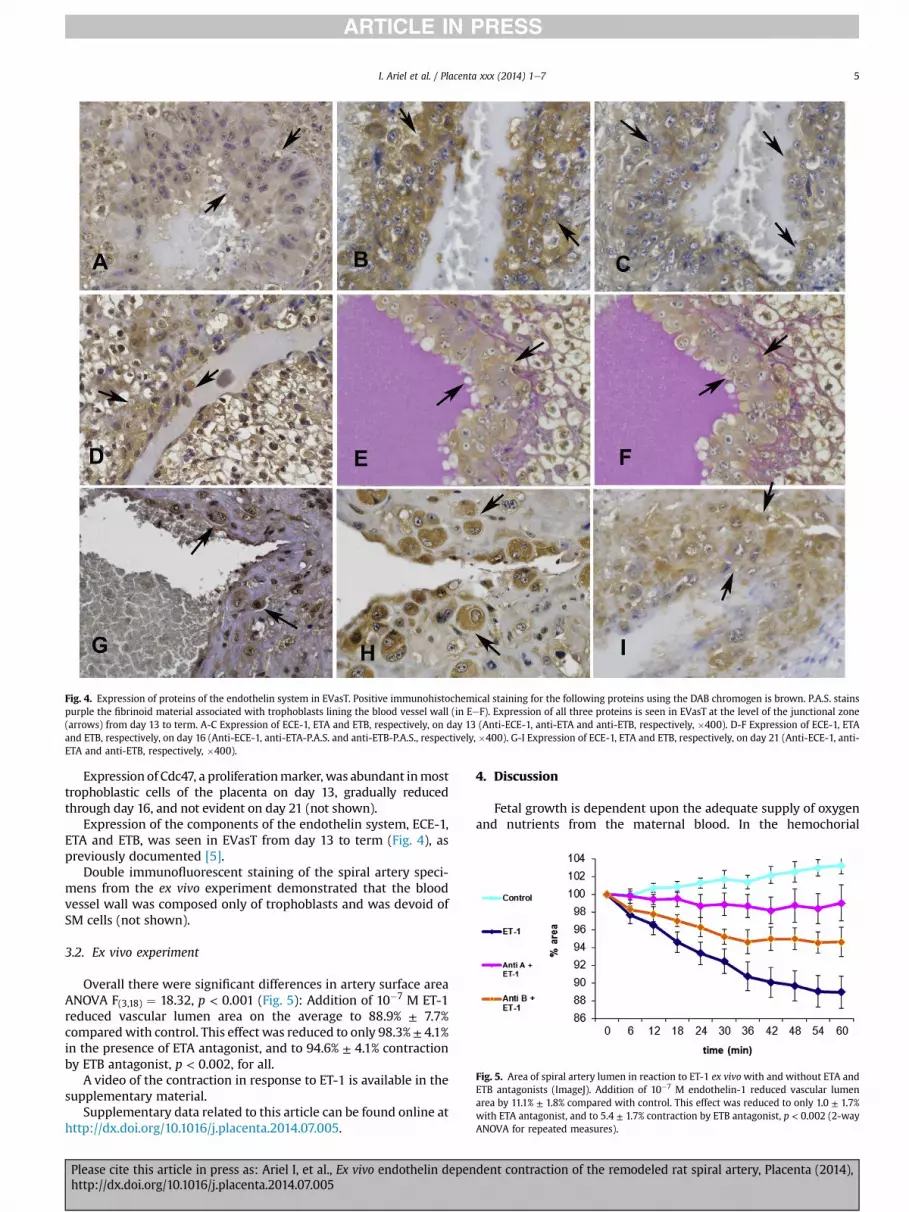
Fig. 4. Expression of proteins of the endothelin system in EVasT. Positive immunohistochemical staining for the following proteins using the DAB chromogen is brown. P.A.S. stainspurple the fibrinoid material associated with trophoblasts lining the blood vessel wall (in EeF). Expression of all three proteins is seen in EVasT at the level of the junctional zone(arrows) from day 13 to term. A-C Expression of ECE-1, ETA and ETB, respectively, on day 13 (Anti-ECE-1, anti-ETA and anti-ETB, respectively, �400). D-F Expression of ECE-1, ETAand ETB, respectively, on day 16 (Anti-ECE-1, anti-ETA-P.A.S. and anti-ETB-P.A.S., respectively, �400). G-I Expression of ECE-1, ETA and ETB, respectively, on day 21 (Anti-ECE-1, anti-ETA and anti-ETB, respectively, �400).
Fig. 5. Area of spiral artery lumen in reaction to ET-1 ex vivowith and without ETA andETB antagonists (ImageJ). Addition of 10�7 M endothelin-1 reduced vascular lumenarea by 11.1% ± 1.8% compared with control. This effect was reduced to only 1.0 ± 1.7%with ETA antagonist, and to 5.4 ± 1.7% contraction by ETB antagonist, p < 0.002 (2-wayANOVA for repeated measures).
I. Ariel et al. / Placenta xxx (2014) 1e7 5
Expression of Cdc47, a proliferationmarker, was abundant inmosttrophoblastic cells of the placenta on day 13, gradually reducedthrough day 16, and not evident on day 21 (not shown).
Expression of the components of the endothelin system, ECE-1,ETA and ETB, was seen in EVasT from day 13 to term (Fig. 4), aspreviously documented [5].
Double immunofluorescent staining of the spiral artery speci-mens from the ex vivo experiment demonstrated that the bloodvessel wall was composed only of trophoblasts and was devoid ofSM cells (not shown).
3.2. Ex vivo experiment
Overall there were significant differences in artery surface areaANOVA F(3,18) ¼ 18.32, p < 0.001 (Fig. 5): Addition of 10�7 M ET-1reduced vascular lumen area on the average to 88.9% ± 7.7%comparedwith control. This effect was reduced to only 98.3% ± 4.1%in the presence of ETA antagonist, and to 94.6% ± 4.1% contractionby ETB antagonist, p < 0.002, for all.
A video of the contraction in response to ET-1 is available in thesupplementary material.
Supplementary data related to this article can be found online athttp://dx.doi.org/10.1016/j.placenta.2014.07.005.
Please cite this article in press as: Ariel I, et al., Ex vivo endothelin depenhttp://dx.doi.org/10.1016/j.placenta.2014.07.005
4. Discussion
Fetal growth is dependent upon the adequate supply of oxygenand nutrients from the maternal blood. In the hemochorial
dent contraction of the remodeled rat spiral artery, Placenta (2014),

I. Ariel et al. / Placenta xxx (2014) 1e76
placenta, maternal blood from the uterine arteries perfuses a spacelined by trophoblastic cells. Adequate uteroplacental blood flowinto this space is achieved by structural modification of the spiralarteries. Following invasion of the arterial wall by trophoblasticcells the SM and endothelial cells degenerate and are replaced bytrophoblasts enmeshed in fibrinoid material.
Human and rat implantation sites notably share deep invasionof EVasT [1]. In the rat the spiral arteries entering from the MT intothe placental disc as central arteries, are populated by trophoblasticcells with rare to no SM cells, allowing for high capacity/lowpressure blood perfusion of the labyrinth. At a deeper level of theMT, branches of the spiral arteries are partially lined by tropho-blasts, whereas the rest of their circumference retains its normalmuscular wall [6,11].
Therefore, the amount and pressure of blood flowing throughthe central artery into the labyrinth could potentially be controlledby branches of the spiral artery in the MT. In cases of dysregulationof the vasoactive systems, such as decreased production of NO and/or increased production of endothelin, vessel contraction in thesesegments limits the amount and increases the pressure of bloodperfusing the placenta.
The expression of actin in extravillous trophoblasts was docu-mented in scattered in vitro studies both in humans and in themouse [14], but actin isoforms were not specified. Since actinexpression was observed in migrating cells, especially whenassociated with the lamellipodium [15], it is conceivable that thiswas b-actin, which is associated with cell migration in all celltypes. aSMA expression during placentation, which disappearedupon invasion of the spiral arteries was documented in theextravillous trophoblasts of a South American rodent, Octogondegus [16]. This is in variance to the expression of aSМА in the rat,which was seen only in EVasT. In the human placenta SMM andSMA isoforms are concentrated mainly in the blood vessel wall[17].
The finding of aSMA expression in EVasT of the rat placenta ledus to investigate the expression of other proteins involved in SMdifferentiation and contraction in these cells. These proteins havevarious cellular functions other than SM contraction, such as cellmigration, and therefore it is not surprising that expression ofsome, but not all, of these proteins was observed in interstitial andother types of trophoblast. It is noteworthy that SM specific iso-forms (SMM and aSMA) were almost exclusively expressed inEVasT at their end-point of migration in the vascular wall and werenot expressed by other migrating trophoblasts.
Myocardin is involved in SM differentiation in the cardio-vascular system and internal organs as well as in cell growth andmigration [18]. Its relative abundance in early placental devel-opment declining towards term may be associated both withinduction of SM proteins at an early placental developmentalstage as well as cessation of trophoblast migration towards term.
The RhoeRho kinase pathway, important in the regulation ofnon-muscle cell functionsmediated bymyosin II [19], plays a role incell motility including human trophoblast migration [20]. NMMand SMM comprise the developmentally regulated class II myosinsof SM [21]. In the rat, upon maturation, the NMM heavy chain isreplaced by SMM heavy chain [21]. NMM plays an important role invarious cellular functions including cell migration [22].
Therefore, the expression of ETA in EVasT at term (day 21), whenEVasT are neither migrating nor proliferating is intriguing. Themajor biological role of ETA is mediating the vasoconstrictive effectof endothelin-1 [23]. It plays a role in cell proliferation [23,24],which is not relevant at term.
We demonstrate here for the first time, EVasT expression ofmajor proteins required for SM differentiation and contractionalong with expression of major components of the vasoactive
Please cite this article in press as: Ariel I, et al., Ex vivo endothelin depenhttp://dx.doi.org/10.1016/j.placenta.2014.07.005
systems of endothelin and, previously documented, NOS. ET-1mediates the vasoconstrictive effect of endothelin via its re-ceptors ETA and ETB on SM cells and induces relaxation throughactivation of NOS via its ETB receptor on endothelial cells [25,26].By stripping the endothelial layer from the muscular layer onlycontraction is induced by ET-1 [27].
We have shown that by blocking ETA most of the vasocon-striction of the spiral artery ring populated only by EVasT wasabolished. Blocking of ETB only partially reduced the vasocon-striction in response to ET-1.
In contrast to SM of stripped arteries, EVasT express NOS [6]which could potentially produce NO in response to ET-1 medi-ated by ETB and cause muscle relaxation. It is therefore conceiv-able that by blocking ETB we abolish the net effect of ET-1 ofcontraction and relaxation mediated by ETB. The interplay be-tween these two vasoactive systems in EVasT requires furtherinvestigation.
We suggest the term “myotrophoblast” referring to EvasT in therat placenta, reflecting their dual differentiation and contractibilityupon exposure to Endothelin-1. We found that endothelin-inducedcontraction of the modified spiral artery ex vivo is mediated via itsreceptors, and suggest that this phenomenon may play a role insituations of dysregulation of the vasoactive systems.
Conflict of interest
None declared.
Acknowledgments
The authors are grateful to N. Melamed-Book, PhD, for herinvaluable assistance in the confocal microscopy of the ex vivoexperiment, and are indebted to M. Hanani, PhD, for his help. Theauthors also thank Wiessam Abu-Ahmad, MA, for performing thestatistical analysis, and to C. Kalcheim, PhD, D. Goldman-Wohl, PhDand G. Vainer, MD, PhD, for their comments.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.placenta.2014.07.005.
References
[1] Soares MJ, Chakraborty D, Karim Rumi MA, Konno T, Renaud SJ. Rat placen-tation: an experimental model for investigating the hemochorial maternal-fetal interface. Placenta 2012;33(4):233e43.
[2] Kaufmann P, Black S, Huppertz B. Endovascular trophoblast invasion: impli-cations for the pathogenesis of intrauterine growth retardation and pre-eclampsia. Biol Reprod 2003;69(1):1e7.
[3] Brosens I, Pijnenborg R, Vercruysse L, Romero R. The “Great Obstetrical Syn-dromes” are associated with disorders of deep placentation. Am J ObstetGynecol 2011;204(3):193e201.
[4] Lyall F, Robson SC, Bulmer JN. Spiral artery remodeling and trophoblast in-vasion in preeclampsia and fetal growth restriction: relationship to clinicaloutcome. Hypertension 2013;62(6):1046e54.
[5] Khamaisi M, Skarzinski G, Mekler J, Zreik F, Damouni R, Ariel I, et al. Hyper-insulinemia increases placenta endothelin-converting enzyme-1 expression introphoblasts. Am J Hypertens 2012;25(1):109e14.
[6] Skarzinski G, Khamaisi M, Bursztyn M, Mekler J, Lan D, Evdokimov P, et al.Intrauterine growth restriction and shallower implantation site in rats withmaternal hyperinsulinemia are associated with altered NOS expression.Placenta 2009;30(10):898e906.
[7] Beausejour A, Auger K, St-Louis J, Brochu M. High-sodium intake preventspregnancy-induced decrease of blood pressure in the rat. Am J Physiol HeartCirculator Physiol 2003;285(1):H375e83.
[8] Curzen NP, Mitchell JA, Jourdan KB, Griffiths MJ, Evans TW. Endothelin-1-induced contraction of pulmonary arteries from endotoxemic rats is attenu-ated by the endothelin-A receptor antagonist, BQ123. Crit Care Med1996;24(12):2007e13.
dent contraction of the remodeled rat spiral artery, Placenta (2014),

I. Ariel et al. / Placenta xxx (2014) 1e7 7
[9] Taddei S, Vanhoutte PM. Role of endothelium in endothelin-evoked contrac-tions in the rat aorta. Hypertension 1993;21(1):9e15.
[10] Warner TD, Allcock GH, Vane JR. Reversal of established responses toendothelin-1 in vivo and in vitro by the endothelin receptor antagonists, BQ-123 and PD 145065. Br J Pharmacol 1994;112(1):207e13.
[11] Geusens N, Verlohren S, Luyten C, Taube M, Hering L, Vercruysse L, et al.Endovascular trophoblast invasion, spiral artery remodelling and uteropla-cental haemodynamics in a transgenic rat model of pre-eclampsia. Placenta2008;29(7):614e23.
[12] Hilgers RH, Webb RC. Molecular aspects of arterial smooth muscle contrac-tion: focus on Rho. Exp Biol Med (Maywood) 2005;230(11):829e35.
[13] Parmacek MS. Myocardin-related transcription factors: critical coactivatorsregulating cardiovascular development and adaptation. Circ Res 2007;100(5):633e44.
[14] Sobel JS, Cooke R, Pedersen RA. Distribution of actin and myosin in mousetrophoblast: correlation with changes in invasiveness during developmentin vitro. Dev biol 1980;78(2):365e79.
[15] Harris LK, McCormick J, Cartwright JE, Whitley GS, Dash PR. S-nitrosylation ofproteins at the leading edge of migrating trophoblasts by inducible nitricoxide synthase promotes trophoblast invasion. Exp Cell Res 2008;314(8):1765e76.
[16] Bosco C, Buffet C. Immunohistochemical identification of the extravilloustrophoblast during the placentation of the degu (Octodon degus). J Exp ZoolPart B 2008;310B(6):534e9.
[17] Matsumura S, Sakurai K, Shinomiya T, Fujitani N, Key K, Ohashi M.Biochemical and immunohistochemical characterization of the isoformsof myosin and actin in human placenta. Placenta 2011;32(5):347e55.
Please cite this article in press as: Ariel I, et al., Ex vivo endothelin depenhttp://dx.doi.org/10.1016/j.placenta.2014.07.005
[18] Pipes GC, Creemers EE, Olson EN. The myocardin family of transcriptionalcoactivators: versatile regulators of cell growth, migration, and myogenesis.Genes Dev 2006;20(12):1545e56.
[19] Fukata Y, Amano M, Kaibuchi K. Rho-Rho-kinase pathway in smooth musclecontraction and cytoskeletal reorganization of non-muscle cells. TrendsPharmacol Sci 2001;22(1):32e9.
[20] Saso J, Shields SK, Zuo Y, Chakraborty C. Role of Rho GTPases in humantrophoblast migration induced by IGFBP1. Biology Reprod 2012;86(1):1e9.
[21] Eddinger TJ, Meer DP. Myosin II isoforms in smooth muscle: heterogeneityand function. Am J Physiol Cell Physiol 2007;293(2):C493e508.
[22] Wang A, Ma X, Conti MA, Liu C, Kawamoto S, Adelstein RS. Nonmuscle myosinII isoform and domain specificity during early mouse development. Proc NatlAcad Sci U S A 2010;107(33):14645e50.
[23] Luscher TF, Barton M. Endothelins and endothelin receptor antagonists:therapeutic considerations for a novel class of cardiovascular drugs. Circula-tion 2000;102(19):2434e40.
[24] Cervar-Zivkovic M, Dieber-Rotheneder M, Barth S, Hahn T, Kohnen G,Huppertz B, et al. Endothelin-1 stimulates proliferation of first-trimestertrophoblasts via the A- and B-type receptor and invasion via the B-type re-ceptor. J Clin Endocrinol Metabol 2011;96(11):3408e15.
[25] Kawanabe Y, Nauli SM. Endothelin. Cellular and molecular life sciences: CMLS2011;68(2):195e203.
[26] Remuzzi G, Perico N, Benigni A. New therapeutics that antagonize endothelin:promises and frustrations. Nat Rev Drug Discov 2002;1(12):986e1001.
[27] Ou M, Dang Y, Mazzuca MQ, Basile R, Khalil RA. Adaptive regulation ofendothelin receptor type A and type B in vascular smooth muscle cells duringpregnancy in rats. J Cell Physiol 2014;229(4):489e501.
dent contraction of the remodeled rat spiral artery, Placenta (2014),