expression of oxidative stress-related molecules in circulating leukocytes and urine in patients...
TRANSCRIPT

Clinical Studies
Expression of oxidative stress-relatedmolecules in circulating leukocytes andurine in patients with chronic viral hepatitis
Toshiya Saeki1, Miho Ichiba1,Naotada Tanabe1, Masaru Ueki2,Kinya Okamoto2, YoshikoMatsunaga2, Keiko Hosho2,Takamasa Kanbe1,3, HiroyukiTsuchiya1, Akihiro Kurimasa1,Sadako Yamada3, Yasuaki Hirooka4,Ichiro Hisatome5, YukihiroKishimoto6, Takeaki Suou3,Yoshikazu Murawaki2, HironakaKawasaki6, Junji Yodoi7 andGoshi Shiota1
1Division of Molecular and Genetic Medicine,
Department of Genetic Medicine and
Regenerative Therapeutics, Graduate School of
Medicine, 2Division of Medicine and Clinical
Science, Department of Multidisciplinary Internal
Medicine, Faculty of Medicine, 3Department of
Pathological Science and Technology, School of
Science, Faculty of Medicine, 4Division of
Surgery and Clinical Oncology, Department of
Surgery, 5Division of Regenerative Medicine,
Department of Genetic Medicine and
Regenerative Therapeutics, Graduate School of
Medicine, Tottori University, Yonago, Japan,6Department of Internal Medicine, San-in Rosai
Hospital, Yonago, Japan, 7Department of
Biological Responses, Laboratory of Infection
and Prevention Institute for Virus Research,
Kyoto University, Kyoto, Japan
Saeki T, Ichiba M, Tanabe N, Ueki M, Okamoto K, Matsunaga Y, Hosho K,Kanbe T, Tsuchiya H, Kurimasa A, Yamada S, Hirooka Y, Hisatome I,Kishimoto Y, Suou T, Murawaki Y, Kawasaki H, Yodoi J, Shiota G.Expression of oxidative stress-related molecules in circulating leukocytes andurine in patients with chronic viral hepatitis.Liver International 2006: 26: 157–165.r 2005 The Authors. Journal compilation r 2006 Blackwell Munksgaard
Abstract: Aims: Oxidative stress plays a role in pathogenesis of chronic viralhepatitis. Expression of oxidative stress-related molecules remains to beclarified. Methods: 4-hydroxy-2-nonenal (4-HNE), 4-hydroxy-2-hexenal (4-HHE), catalase, superoxide dismutase-1 (SOD-1), glutathione peroxidase-1,thioredoxin (TRX) in leukocytes and urinary 8-hydroxy-20-deoxyguanosine(8-OHdG) were examined in 164 persons, including 130 chronic viralhepatitis patients and 34 normal individuals, by Western blot analysis andenzyme-linked immunosorbent assay, respectively. Hepatic expression ofthese proteins was immunohistochemically examined in 12 patients withchronic viral hepatitis, compared with three persons without liver damage.Results: The 4-HNE/b-actin ratios in chronic viral hepatitis were significantlyhigher than those in normal individuals (Po0.01), and were significantlycorrelated with asparate aminotransferase (AST) and alanineaminotransferase (ALT) (Po0.01, each). The catalase/b-actin and SOD-1/b-actin ratios in chronic viral hepatitis were higher than those in normalindividuals, and were significantly correlated with 4-HNE/b-actin ratios(Po0.01, each). Hepatic expression of 4-HNE, 4-HHE, catalase, SOD-1 andTRX in chronic viral hepatitis was higher than that without liver damage.Urinary excretion of 8-OHdG was not changed in chronic viral hepatitis.Conclusions: The results of the present study suggest that expression ofoxidative stress-related molecules in leukocytes is upregulated in relationto serum aminotransferase levels.
Key words: antioxidant protein – chronic viral
hepatitis – leukocyte – lipid peroxidation
Goshi Shiota, MD, PhD, Division of Molecular
and Genetic Medicine, Department of Genetic
Medicine and Regenerative Therapeutics, Grad-
uate School of Medicine, Tottori University,
Yonago 683-8504, Japan.
Tel: 181 859 34 8464
Fax: 181 859 34 8139
e-mail: [email protected]
Received 8 July 2005,
accepted 3 October 2005
����������������������������������������������������������������������
����������������������������������������������������������������������
Oxidative stress is involved in a number ofpathological conditions such as inflammationand cancer (1). In the liver, oxidative stress has
Abbreviations: 4-HNE, 4-hydroxy-2-nonenal; 4-HHE, 4-hy-droxy-2-hexenal; SOD, superoxide dismutase; GPx, glutathioneperoxidase; TRX, thioredoxin; 8-OHdG, 8-hydroxy-20-deoxy-guanosine; ELISA, enzyme-linked immunosorbent assay; HCV,hepatitis C virus; HBV, hepatitis B virus; HCC, hepatocellularcarcinoma.
Liver International 2006: 26: 157–165 r 2005 The AuthorsJournal compilation r 2006 Blackwell Munksgaard
DOI: 10.1111/j.1478-3231.2005.01213.x
157

been reported to be involved in the progression ofchronic liver diseases and hepatocellular carci-noma (HCC) (2–5). 8-hydroxy-20-deoxyguano-sine (8-OHdG) formed by oxidative stress,mainly by hydroxyl radicals, causes specific typesof mutation, and it is associated with various liverdiseases (2, 5–7).Lipid peroxidation is an initial event that
results in liver damage (8). Lipid peroxidationstimulates procollagen a1(I) gene expression inhuman liver fat-storing cells (9). The breakdownof the hydroperoxides generates a variety ofaldehydes such as 4-hydroxy-2-nonenal (4-HNE) and 4-hydroxy-hexenal (4-HHE) from n-6 fatty acids and n-3 fatty acids, respectively (10).4-HNE and 4-HHE are considered as reliablemarkers of lipid peroxidation (11). In addition, 4-HNE protein was expressed in rat liver afterischemia–perfusion (12) and human liver tissueswith chronic hepatitis C (13), suggesting that lipidperoxidation plays an important role in liverdamage.Levels of antioxidant proteins such as super-
oxide dismutase (EC 1.15.1.1, SOD), catalase (EC1.11.1.6), glutathione peroxidase (EC 1.11.1.19,GPx) and thioredoxin (TRX) are closely linkedwith cellular responses to various oxidative stres-ses (14). SOD-1 (Cu–Zu SOD-1) is ubiquitouslyexpressed and is a cytosolic scavenger of oxygen-free radicals by facilitating the dismutation ofoxygen radicals to molecular oxygen and hydro-gen peroxide, which in turn is metabolized toharmless water and oxygen by catalase and GPx.Catalase is a heme-containing enzyme, whosesubcellular localization is in peroxisomes, wherecatalase catalyzes the dismutation of hydrogenperoxide to water and molecular oxygen andfunctions in lowering the risk of hydroxyl radicalformation from hydrogen peroxide via Fentonreaction (15). GPx, which are composed of fourdifferent isoforms, catalyzes the reduction ofhydrogen peroxide using glutathione as a sub-strate. They can also reduce lipid peroxides toalcohols. TRX is a small disulfide-reducing en-zyme that has a conserved consensus sequence –CXXC – at its active site. TRX is a potent proteindisulfide oxidoreductase, which is important inantioxidant defense (15). So far, there have been afew reports that describe the expression of anti-oxidant proteins in chronic liver diseases.Recent studies have reported that 8-OHdG
contents in circulating leukocytes of chronic liverdisease were increased in comparison with normalindividuals (16, 17). However, although urinary8-OHdG excretion was changed according to theinfluence of smoking, gender and body massindex in normal subjects (18), it remains to be
clarified whether urinary 8-OHdG excretion isaltered in chronic viral hepatitis. In addition,although 8-OHdG in leukocytes reflected liverdamage (16, 17), and serum lipid peroxidationand antioxidants were increased in human non-alcoholic fatty liver diseases (19, 20), it remains tobe clarified whether expression of oxidativestress-related molecules in leukocytes is asso-ciated with liver damage of chronic viral hepati-tis. In order to clarify the significance of lipidperoxidation in leukocytes and urinary DNAoxidative damage in human liver diseases, weexamined expression of 4-HNE, 4-HHE, catalase,SOD-1, GPx-1 and TRX in leukocytes of chronicviral hepatitis, and measured urinary 8-OHdGcontents by an enzyme-linked immunosorbentassay (ELISA) assay.
Materials and methods
Patients
One hundred and thirty patients with chronicviral hepatitis, who were seen in Tottori Univer-sity hospital between April 2003 and July 2005and whose informed consent was obtained, wereprospectively enrolled in the study. The inclusioncriteria for the study were positivity of hepatitisvirus infection, presence of chronic liver diseaseand absence of active substance abuse or othersystemic illness. They included 27 with chronichepatitis (CH), 14 with liver cirrhosis (LC) and 89with HCC. All the CH patients were diagnosedby liver biopsy under laparoscopy or ultrasoundusing a 17-gauge needle. Seven LC patients werealso diagnosed by liver biopsy under laparoscopy,and the other seven LC patients were diagnosedby clinical symptoms, biochemical examinations,ultrasonography and computed tomography. TenHCC patients were diagnosed by biopsy underultrasonography, and the other HCC patientswere diagnosed by ultrasonography, computedtomography and tumor markers such as a-feto-protein (AFP) and PIVKAII. All HCC patientshad LC as non-cancerous counterparts. Tissuesamples of HCC for immunohistochemical studywere taken under surgical operation. Thirty-fourhealthy individuals served as normal controls.The inclusion criteria for normal controls werethat their serum levels of AST and ALT werewithin the normal range and they did not havediabetes mellitus, cancer, active substance abuseor other systemic illness. Informed consent wasobtained from each patient according to theethical guidelines approved by the Tottori Uni-versity research committee. The clinical para-meters include age, sex, diagnosis, etiology,
158
Saeki et al.

height, body weight, body mass index (BMI),smoking, alcohol drinking, number and size ofHCC, platelet, BUN, creatinine, AST, ALT,alkaline phosphatase (ALP), g-GTP, ChE, indo-cyanine green (ICG) retention test, AFP, PIV-KAII, CA19-9, type IV collagen and hyaluronate.The etiology was classified by HBs antigen(LPIA-200, Diatron Laboratories Inc., Tokyo,Japan) and hepatitis C virus (HCV) antibody(Immunocheck-HCV Ab, International ReagentCorporation, Kobe, Japan). Serum ALT, AST,bilirubin, albumin, LDH, g-GTP, ICG and AFPwere measured as described previously (5). Thetype IV collagen in the serum was measured byPanasay IV C latex (Daiichi Fine Chemical Co.Ltd., Toyama, Japan). Serum hyaluronate wasmeasured by an HA standard set (MitsubishiKagaku Latron Inc., Tokyo, Japan).
Separation of leukocytes and Western blot analysis
After obtaining informed consent, a 4ml of bloodsample was taken from each of the patients andnormal subjects. Collection of blood samples andpreparation of leukocyte protein were performedas described previously (20, 21). Fractionatedleukocytes were lysed in a lysis buffer consistingof PBS (pH 7.2) with 1% NP-40, 10 nmol/lEDTA, 0.2% sodium dodecyl sulfate (SDS) andthe following inhibitors: 10mmol/l benzamidine,1mmol/l phenylmethylsulfonyl fluoride (PMSF),100 mmol/l Na-p-tosylL-lysine chloromethyl ke-tone, 100mmol/l leupeptin and 1 mmol/l pepstatin.After separation with a microfuge, the proteinhomogenates (20 mg) were separated by SDS/polyacrylamide gel electrophoresis (SDS-PAGE), and proteins were transferred on to apolyvinylidene difluoride membrane followed byimmunoblot analysis with a monoclonal anti-4-HNE (NOF Corp. Tokyo, Japan), a monoclonalanti-4-HHE (NOF Corp.), a rabbit polyclonalanti-SOD1(Santa Cruz Biotechnologies, SantaCruz, CA), a goat polyclonal anti-GPx-1 (SantaCruz Biotechnologies), a rabbit anti-catalase(Lab Frontier, Seoul, Korea), a monoclonalanti-TRX and a monoclonal anti-b-actin (SantaCruz Biotechnologies) antibody, respectively.Membranes were incubated with horseradishperoxidase-conjugated secondary antibodies fol-lowed by detection with enhanced chemilumines-cence (ECL) (Amersham Bioscience Corp.,Piscataway, NJ). The intensity of the bands wascalculated by Image J1.30v (National Institutes ofHealth), and was expressed as the ratio to b-actin.For 4-HNE expression, the 42 kD band wasselected for measurement of intensity since the42 kDa band is prominent using the anti-4-HNE
antibody (12). For 4-HHE expression, the 53 kDaband was selected for measurement of intensity asthe 53 kDa band was prominent in Western blotusing anti-4HHE antibody.
8-OHdG determination
Urine was obtained after at least 4 h urine sam-ples from all the participants. Urinary 8-OHdGwas measured using the 8-OHdG ELISA kit(NOF Science Division, Tokyo, Japan), as theUVC-induced product was detected by a mono-clonal antibody recognizing 8-OHdG, with astrong correlation between the measurement ofthe UVC produced by high-performance liquidchromatography with electrochemical detection(HPLC-EC) and ELISA (22). The measurementof 8-OHdG determination by ELISA was per-formed according to the manufacturer’s instruc-tion. Urinary 8-OHdG was expressed as the ratioto creatinine. Urinary creatinine was measured byan autoanalyzer after treatment with DETERMI-NARLE CRE (Kyowa Medix Co. Ltd., Tokyo,Japan).
Immunohistochemistry
The liver sections from 12 patients including fourwith CH, four with LC and 4 with HCC wereimmunohistochemically examined. For immuno-histochemical study, the tissue specimens of HCCwere taken under surgical operation, and those ofCH and LC were taken under laparoscopy- orultrasonography-guided liver biopsy. For controlliver tissues, three normal liver tissues of thepatients who underwent partial hepatectomy be-cause of colon cancer metastasis were examined.De-waxed paraffin sections were immunostainedusing the avidin–biotin–peroxidase complex. Thesections were incubated with 0.3% H2O2 inmethanol for 30min. They were heated in10mmol/l sodium citrate buffer (pH 6.0) at600W for 15min. They were incubated withnormal horse serum. After washing, they wereincubated with the following primary antibodies:a monoclonal anti-4-HNE (NOF Corp.), a mono-clonal anti-4-HHE (NOF Corp.), a rabbit poly-clonal anti-SOD1 (Santa Cruz Biotechnologies),a goat polyclonal anti-GPx-1 (Santa Cruz Bio-technologies), a rabbit anti-catalase antibody(Lab Frontier, Seoul, Korea) and a monoclonalanti-TRX antibody at 4 1C overnight. Then, theywere incubated with biotin-conjugated anti-mouse immunoglobulin G (Vector Laboratories,Burlingame, CA) for 60min. Immunoreactivecells were visualized using a Vectastain ABC-POKit (Vector Laboratories).
159
Oxidative stress-related molecules in chronic viral hepatitis

Statistics
Values were expressed as mean� SD. Meanswere compared using Wilcoxon’s rank test. Therelationships between two data were examined byusing Pearson’s correlation coefficient. Probabil-ity levels o0.05 were considered statistically sig-nificant.
Results
Clinical characteristics of the patients withchronic viral hepatitis
The clinical characteristics of the patients areshown in Table 1. The HCC patients were olderthan normal individuals, CH and LC patients(Po0.01, each). There were no statistical differ-ences in sex, etiology or AST among them. Thevalue of serum ALT in CH was higher than thatin HCC (Po0.01). The platelet numbers inchronic viral hepatitis were smaller than thosein normal individuals (Po0.01, each). ICG reten-tion rates at 15min in LC and HCC were greaterthan those in CH (Po0.01, each). The type IVcollagen levels in LC and HCC were higher than
those in CH (Po0.05 and Po0.01, respectively).The serum hyaluronate level in LC was higherthan that in CH (Po0.05).
Oxidative stress-related molecules in patients withchronic viral hepatitis
There were no differences in urinary 8-OHdGexcretion among liver diseases (Table 2). However,there was a relationship between urinary 8-OHdGexcretion and age (r5 0.258, Po0.05). Interest-ingly, the HCV-related patients had higher urinary8-OHdG excretion than the hepatitis B virus(HBV)-related patients (14.4 ng/mg in HCV vs.10.8 ng/mg in HBV, Po0.01). However, as theHCV-related patients were older than the HBV-related patients (Po0.01), the difference in 8-OHdG excretion may not be because of etiology,but age. No relationship was observed amongurinary 8-OHdG, liver function tests, and otheroxidative stress-related molecules. These data in-dicate that urinary 8-OHdG does not reflecthepatic oxidative stress and liver damage.
Expression of reactive oxygen species-relatedmolecules such as 4-HNE, 4-HHE, catalase,
Table 1. Characteristics of the patients
Normal CH LC HCC
Patient no. 34 27 14 89Age 59 � 181 52 � 151 60 � 121 67 � 9Sex (M/F) 13/21 10/17 7/7 55/34Etiology (B/C/B1C/others) ND 8/18/1/0 5/7/1/1 22/65/2/0AST (IU/l) 26 � 9 85 � 782 74 � 562 77 � 682
ALT (IU/l) 29 � 11 111 � 1111,2 74 � 111 61 � 392
ALP (IU/l) 204 � 37 295 � 902,3 319 � 742 383 � 2182
g-GTP (IU/l) 33 � 9 73 � 69 72 � 67 93 � 105Plt (109/ml) 221 � 52 126 � 622 135 � 602 110 � 512
ICG (%) ND 15 � 7 27 � 124 28 � 154
AFP (ng/ml) ND 15 � 20 9 � 7 7651 � 26 415Type IV collagen ND 5.0 � 1.6 7.5 � 3.05 7.8 � 2.74
Hyaluronate ND 120 � 89 760 � 11235 486 � 1241
Data are expressed as mean � SD. 1Po0.01, compared with HCC. 2Po0.01, compared with normal subjects. 3Po0.05, compared with HCC.4Po0.01, compared with CH. 5Po0.05, compared with CH. AST, asparate aminotransferase; ALT, alanine aminotransferase; ALP, alkalinephosphatase; ICG, indocyanine green; AFP, a-fetoprotein; ND; not determined; HCC, hepatocellular carcinoma; CH, chronic hepatitis.
Table 2. Summary of oxidative stress-related molecules in patients with chronic viral hepatitis and normal individuals
N CH LC HCC
8-OHdG(ng/mg)1 11.4 � 5.3 13.7 � 5.3 11.5 � 4.8 13.3 � 5.64-HNE 0.9 � 0.4 3.4 � 3.32 1.9 � 1.92 3.1 � 3.62
4-HHE 0.8 � 0.6 5.8 � 11.43 ND 2.0 � 4.4Catalase 1.6 � 1.1 5.8 � 9.83 8.9 � 9.92 8.1 � 11.32
SOD-1 3.5 � 2.0 12.3 � 13.72 13.0 � 13.72 12.4 � 18.43
GPx-1 0.01 � 0.06 0.18 � 0.423,4 0.08 � 0.204 0TRX 1.1 � 1.4 1.9 � 2.1 1.0 0.7 � 1.1
Data are expressed as mean � SD. All data except 8-OHdG were expressed as arbitrary units. 1Urinary 8-OHdG expressed as 8-OHdG (ng/ml) dividedby urinary creatinine (mg/ml). 2Po0.01, compared with normal individuals. 3Po0.05, compared with normal individuals. 4Po0.01,compared withHCC. ND, not determined; 8-OHdG, 8-hydroxy-2 0-deoxyguanosine; 4-HNE, 4-hydroxy-2-nonenal; 4-HHE, 4-hydroxy-2-hexenal; SOD-1, superoxidedismutase-1;GPx, glutathione peroxidase-1; TRX, thioredoxin; HCC; hepatocellular carcinoma; LC, liver cirrhosis; CH, chronic hepatitis.
160
Saeki et al.

SOD-1, GPx-1, TRX and b-actin in leukocyteswas examined by Western blot analysis (Fig. 1).The samples, of which b-actin expression wasvery low, were excluded from further analysis
since the quality of the sample proteins does notseem to be suitable for further analysis. Not allWestern blot analyses using seven antibodieswere performed in some patients when the vo-lume of protein samples was not sufficient for allWestern blot analyses. The 4-HNE/b-actin ratiosin CH, LC and HCC were higher than those innormal individuals (Po0.01, each, Table 2). The4-HNE/b-actin ratio was correlated with ASTand ALT (Po0.01, each, Fig. 2A, B). The 4-HHE/b-actin was higher in CH than in normalindividuals (Po0.05). The catalase/b-actin ratiosin CH, LC and HCC were higher than those innormal individuals (Po0.05, Po0.01 andPo0.01, respectively). The catalase/b-actin ratiowas correlated with ALP (Po0.01, Fig. 2C). TheSOD-1/b-actin ratios in CH, LC and HCC weresignificantly increased in comparison with normalindividuals (Po0.01, Po0.01 and Po0.05, re-spectively). The SOD-1/b-actin ratio was corre-lated with ALP (Po0.01, Fig. 2D). The GPx-1/b-actin ratios in CH and LC were higher than thosein HCC (Po0.01, each, Table 2). The GPx-1/b-actin ratio was not related to any liver functiontests or other protein expression. For the TRX/b-actin ratio, no differences were observed amongliver diseases and normal individuals (Table 2).The 4-HNE/b-actin ratio was significantly cor-
related with catalase/b-actin and SOD-1/b-actin
4-HNE
4-HHE
Catalase
SOD
GPx
N
TRX
CH LC HCC
ββ-actin
Fig. 1. Western blot analysis of leukocyte protein by anti-4-HNE, anti-4-HHE, anti-catalase, anti-SOD, anti-GPx, anti-TRX and b-actin antibodies. N, normal subjects; CH, chronichepatitis; LC, liver cirrhosis; HCC, hepatocellular carcinoma; 4-HNE, 4-hydroxy-2-nonenal; 4-HHE, 4-hydroxy-2-hexenal;SOD, superoxide dismutase; Gpx, glutathione peroxidase-1;TRX, thioredoxin.
AS
T(I
U/l)
ALT
(IU
/l)
Catalase/β-actin
ALP
(IU
/l)
r=0.351P<0.01 r=0.333
P<0.01
P<0.01P<0.01r=0.356 r=0.369
400
600550500450400350300250200150100
ALP
(IU
/l)
600550500450400350300250200150100
350300250200150100500
–50
500450400350300250200150100500
–50
–5 0 5 10 15 20 25 30 35 40 45
4-HNE/ β-actin–2 0 2 4 6 8 10 12 14 16 18
4-HNE/ β-actin–2 0 2 4 6 8 10 12 14 16 18
SOD-1/β -actin
–10 0 10 20 30 40 50 60 70 80 90 100
(A) (B)
(C) (D)
Fig. 2. Correlation of oxidative stress-related molecules in leukocytes with liver function tests. (A) 4-HNE/b-actin vs. AST, (B) 4-HNE/b-actin vs. ALT, (C) catalase/b-actin vs. ALP and (D) SOD-1/b-actin vs. ALP. The intensity of the bands was calculated byImageJ1.30v (National Institutes of Health), and was expressed as the ratio to b-actin. 4-HNE, 4-hydroxy-2-nonenal; SOD-1,superoxide dismutase-1; AST, asparate aminotransferase; ALT, alanine aminotransferase; ALP, alkaline phosphatase.
161
Oxidative stress-related molecules in chronic viral hepatitis
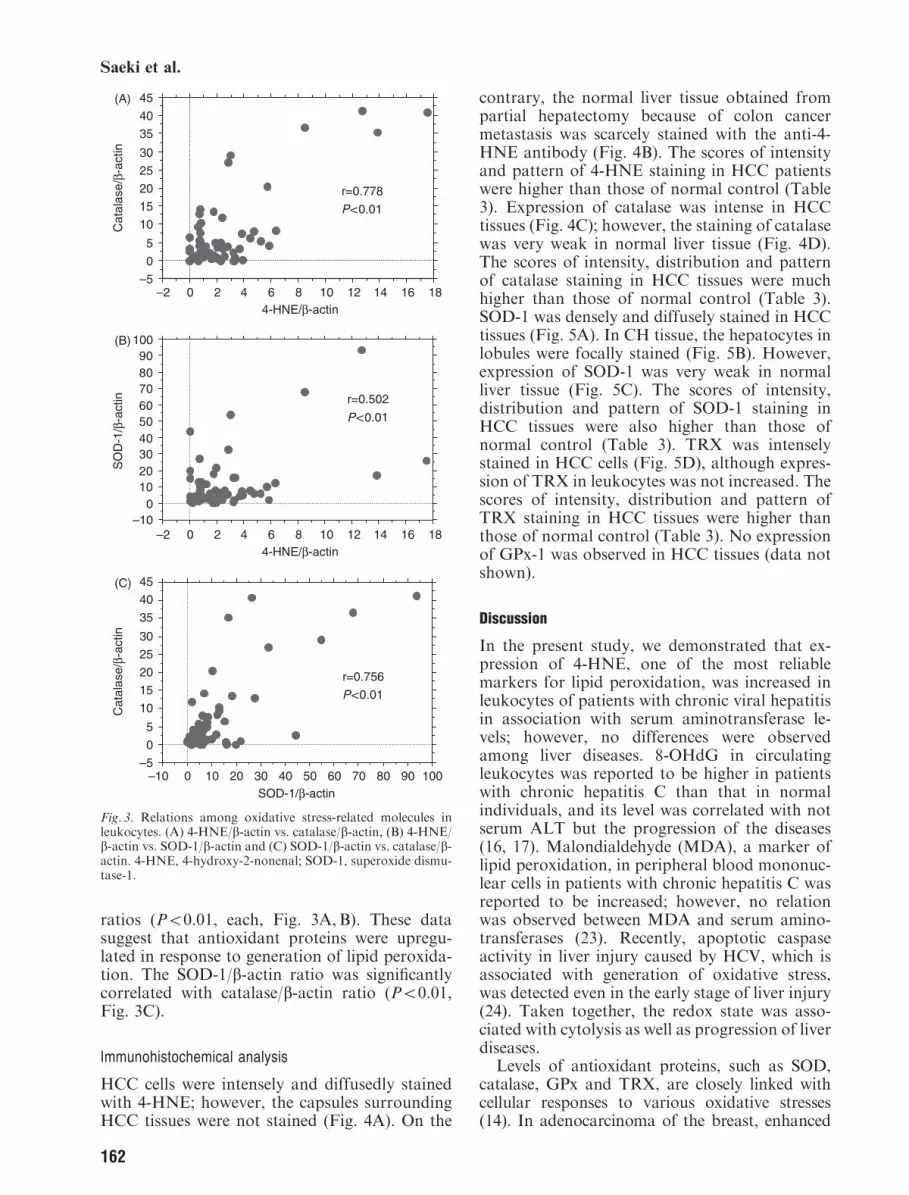
ratios (Po0.01, each, Fig. 3A, B). These datasuggest that antioxidant proteins were upregu-lated in response to generation of lipid peroxida-tion. The SOD-1/b-actin ratio was significantlycorrelated with catalase/b-actin ratio (Po0.01,Fig. 3C).
Immunohistochemical analysis
HCC cells were intensely and diffusedly stainedwith 4-HNE; however, the capsules surroundingHCC tissues were not stained (Fig. 4A). On the
contrary, the normal liver tissue obtained frompartial hepatectomy because of colon cancermetastasis was scarcely stained with the anti-4-HNE antibody (Fig. 4B). The scores of intensityand pattern of 4-HNE staining in HCC patientswere higher than those of normal control (Table3). Expression of catalase was intense in HCCtissues (Fig. 4C); however, the staining of catalasewas very weak in normal liver tissue (Fig. 4D).The scores of intensity, distribution and patternof catalase staining in HCC tissues were muchhigher than those of normal control (Table 3).SOD-1 was densely and diffusely stained in HCCtissues (Fig. 5A). In CH tissue, the hepatocytes inlobules were focally stained (Fig. 5B). However,expression of SOD-1 was very weak in normalliver tissue (Fig. 5C). The scores of intensity,distribution and pattern of SOD-1 staining inHCC tissues were also higher than those ofnormal control (Table 3). TRX was intenselystained in HCC cells (Fig. 5D), although expres-sion of TRX in leukocytes was not increased. Thescores of intensity, distribution and pattern ofTRX staining in HCC tissues were higher thanthose of normal control (Table 3). No expressionof GPx-1 was observed in HCC tissues (data notshown).
Discussion
In the present study, we demonstrated that ex-pression of 4-HNE, one of the most reliablemarkers for lipid peroxidation, was increased inleukocytes of patients with chronic viral hepatitisin association with serum aminotransferase le-vels; however, no differences were observedamong liver diseases. 8-OHdG in circulatingleukocytes was reported to be higher in patientswith chronic hepatitis C than that in normalindividuals, and its level was correlated with notserum ALT but the progression of the diseases(16, 17). Malondialdehyde (MDA), a marker oflipid peroxidation, in peripheral blood mononuc-lear cells in patients with chronic hepatitis C wasreported to be increased; however, no relationwas observed between MDA and serum amino-transferases (23). Recently, apoptotic caspaseactivity in liver injury caused by HCV, which isassociated with generation of oxidative stress,was detected even in the early stage of liver injury(24). Taken together, the redox state was asso-ciated with cytolysis as well as progression of liverdiseases.
Levels of antioxidant proteins, such as SOD,catalase, GPx and TRX, are closely linked withcellular responses to various oxidative stresses(14). In adenocarcinoma of the breast, enhanced
Cat
alas
e/ β-
actin
SO
D-1
/β -a
ctin
SOD-1/β -actin
4-HNE/ β-actin
–10 0 10 20 30 40 50 60 70 80 90 100
–5
0
5
10
15
20
25
30
35
40
45
Cat
alas
e/ β-
actin
–5
0
5
10
15
20
25
30
35
40
45
–2 0 2 4 6 8 10 12 14 16 18
4-HNE/ β-actin–2 0 2 4 6 8 10 12 14 16 18
–100
102030405060708090
100
r=0.778
P<0.01
r=0.502
P<0.01
r=0.756
P<0.01
(A)
(B)
(C)
Fig. 3. Relations among oxidative stress-related molecules inleukocytes. (A) 4-HNE/b-actin vs. catalase/b-actin, (B) 4-HNE/b-actin vs. SOD-1/b-actin and (C) SOD-1/b-actin vs. catalase/b-actin. 4-HNE, 4-hydroxy-2-nonenal; SOD-1, superoxide dismu-tase-1.
162
Saeki et al.

Fig. 4. Expression of 4-HNE and catalase in liver tissues. (A) 4-HNE expression in HCC (patient no. 336, original magnification� 100). HCC is a well-differentiated HCC. 4-HNE was diffusely stained in the cytoplasm of HCC cells. The arrow indicates the HCCtissue. (B) Weak expression of 4-HNE in normal control, whose liver tissues were obtained at partial hepatectomy because of metastaticliver cancer of the colon (patient no. 1416, original magnification � 40). (C) Catalase expression in HCC (patient no. 403, originalmagnification � 100). HCC is a well-differentiated HCC. Expression of catalase was diffuse in the cytoplasm (indicated by the arrow).(D) Weak expression of catalase in normal control, whose liver tissues were obtained at partial hepatectomy because of metastatic livercancer of the colon (patient no. 1435, original magnification � 40). 4-HNE, 4-hydroxy-2-nonenal; HCC, hepatocellular carcinoma.
Fig. 5. Expression of antioxidant proteins in liver tissues. (A) SOD-1 expression in HCC tissue (patient no. 3351, originalmagnification � 100). HCC is a well-differentiated HCC. SOD-1 was intensely expressed in HCC tissues (indicated by the arrows).(B) SOD-1 expression in CH tissue (patient no. 3351, original magnification � 40). The spotty expression of SOD-1 was observed inthe lobule (indicated by the arrow). (C) Weak expression of SOD-1 in normal control, whose liver tissues were obtained at partialhepatectomy because of metastatic liver cancer of the colon (patient no. 1435, original magnification � 40). (D) Thioredoxinexpression in HCC tissue (patient no. 3351, original magnification � 100). Thioredoxin was highly expressed in HCC cells. SOD-1,superoxide dismutase-1; HCC, hepatocellular carcinoma; CH, chronic hepatitis.
163
Oxidative stress-related molecules in chronic viral hepatitis

lipid peroxidation was accompanied by significantelevation in enzymatic and nonenzymatic antiox-idants, such as SOD, catalase and GPx, wereobserved in tumor tissues (25). In HepG2 cancercells, the activities of SOD, catalase and GPx-1were significantly lower than those of Chang non-cancerous cells (14). However, expression of GPx-1 was reduced in cancer cells while that of TRXreductase was not (26). These data may supportthe following hypotheses. First, excess generationof oxygen-free radicals can cause oxidative da-mage to biomolecules, resulting in lipid peroxida-tion, mutagenesis and carcinogenesis. Second, theantioxidant proteins may have their specific reg-ulatory machinery that scavenges lipid peroxida-tion in various pathological conditions.TRX is a protein with oxidoreductase activity
and is ubiquitous in both mammalian and prokar-yotic cells. A number of clinical conditions involve
TRX (27). Increased plasma levels of TRX havebeen found in HCC and non-alcoholic steatohe-patitis (28, 29). Indeed, overexpression of TRXprevents liver injury caused by thioacetamide orlipopolysaccaride in mice, suggesting that TRXacts as an antiapoptotic factor (30). In the presentstudy, higher levels of TRX expression in HCCtissues was observed; however, expression of TRXin leukocytes was not altered. These findingssuggest that the expression of TRX in liver tissuesis not identical to that in circulating leukocytes.
In the present study, GPx-1 was scarcely ex-pressed in liver tissues including HCC. It isknown that selenium protects against certain hu-man cancers. GPx-1 is the most extensively stu-died mammalian selenoprotein. Expression ofselenoprotein is regulated by selenium itself, andnutritional selenium deficit leads to reduced ex-pression of levels (31). GPx-1 has a low affinityfor selenium incorporation when selenium islimiting for selenoprotein synthesis, and expres-sion of this protein is significantly decreased atmoderate selenium levels while expression ofother selenoproteins under these conditions isunaffected (32). On the other hand, humanGPx-1 is transcriptionally regulated by p53 (33).Taken together, little expression of GPx-1 in livertissues may be due to the nutritional deficit ofselenium or p53 inactivation.
How were the markers of oxidative stress incirculating leukocytes of chronic viral hepatitisincreased? There may be two reasons responsiblefor this phenomenon. The first reason is thatcirculating leukocytes are exposed to free radicalswhen flowing through the liver as suggested pre-viously (16). In our previous study, hepatic 8-OHdG was significantly correlated with serumaminotransferases (5). Hepatic generation of oxida-tive stress may affect the redox state of circulatingleukocytes. The second reason is a direct effect ofhepatitis virus on lymphocytes as suggested recently(23). The nonstructural protein 3 of HCV has beenreported to trigger an oxidative burst in humanmonocytes (34). This damage may, in turn, besomehow involved with chronic infection of HCVand development of chronic lymphoproliferativedisorders associated with HCV infection.
In conclusion, a higher expression of lipidperoxidation-related molecules and antioxidantproteins was observed in circulating leukocytesof chronic viral hepatitis.
References
1. Kasai H. Analysis of a form of oxidative DNA damage, 8-hydroxy-20-deoxyguanosine, as a marker of cellular oxidativestress during carcinogenesis. Mutation Res 1997; 387: 147–63.
Table 3. Expression of oxidative stress-related proteins in liver tissues
Intensityn Distributionw Patternz
4-HNEControl 1.0 � 0.0 1.0 � 0.0 1.0 � 0.0CH 2.5 � 0.62 2.0 � 0.0 1.5 � 0.62
LC 2.0 � 0.0 2.0 � 0.0 1.5 � 0.63
HCC 2.5 � 0.62 2.0 � 0.0 2.5 � 0.61,5
CatalaseControl 1.0 � 0.0 1.0 � 0.0 1.0 � 0.0CH 2.5 � 0.62 2.0 � 0.02,6 2.0 � 0.0LC 2.0 � 0.02 1.0 � 0.0 1.0 � 0.0HCC 3.0 � 0.02 2.0 � 0.02,6 2.5 � 0.62,6
GPx-1Control 1.0 � 0.0 1.0 � 0.0 1.0 � 0.0CH 1.0 � 0.0 1.0 � 0.0 1.0 � 0.0LC 1.0 � 0.0 1.0 � 0.0 1.0 � 0.0HCC 1.0 � 0.0 1.0 � 0.0 1.0 � 0.04-HHEControl 1.0 � 0.0 1.0 � 0.0 1.0 � 0.0CH 2.0 � 0.0 1.3 � 0.5 1.3 � 0.55
LC 1.5 � 0.6 2.0 � 0.03 1.0 � 0.0HCC 2.0 � 0.0 2.0 � 0.03 1.8 � 0.5SOD-1Control 1.3 � 0.6 1.0 � 0.0 1.0 � 0.0CH 2.5 � 0.61 2.0 � 0.02 2.0 � 0.02
LC 2.0 � 0.0 1.0 � 0.02 2.0 � 0.02
HCC 2.5 � 0.01 2.0 � 0.02,3,5 2.5 � 0.62,6
TRXControl 1.3 � 0.6 1.0 � 0.0 1.0 � 0.0CH 2.5 � 0.62 2.0 � 0.02,4 2.0 � 0.02,4
LC 2.0 � 0.02,6 2.0 � 0.02 1.5 � 0.6HCC 3.0 � 0.02 3.0 � 0.02,3,5 3.0 � 0.02,6
Data are expressed as mean � SD. nThe intensity of staining wasevaluated in the following way: 1, negative; 2, positive; 3, stronglypositive. wThe distribution of staining was evaluated as follows: 1,staining o10% of cells; 2, staining in 10–70% of cells; 3, staining in470% of cells. zThe pattern of staining was evaluated as follows: 1,sporadic; 2, focal or clustered staining; 3, diffuse or widespread staining.1Po0.05, compared with control. 2Po0.01, compared with control.3Po0.05, compared with CH. 4Po0.01, compared with CH. 5Po0.05,compared with LC. 6Po0.01, compared with LC. 4-HNE, 4-hydroxy-2-nonenal; 4-HHE, 4-hydroxy-2-hexenal; SOD-1, superoxide dismutase-1;GPx-1, glutathione peroxidase-1; TRX, thioredoxin; HCC; hepatocellu-lar carcinoma; LC, liver cirrhosis; CH, chronic hepatitis.
164
Saeki et al.

2. Shimoda R, Nagashima M, Sakamoto M, YamaguchiN, Hirohashi S, Yokota J, et al. Increased formation ofoxidative DNA damage, 8-hydroxydeoxyguanosine, in hu-man livers with chronic hepatitis. Cancer Res 1994; 54:3171–2.
3. Farinati F, Cardin R, De Maria N, Della Libera G,Marafin C, Lecis E, et al. Iron storage, lipid peroxidationand glutathione turnover in chronic anti-HCV positivehepatitis. J Hepatol 1995; 22: 449–56.
4. De Maria N, Colantoni A, Fagiuoli S, Liu G J, Ro-gers B K, Farinati F, et al. Association between reactiveoxygen species and disease activity in chronic hepatitis C.Free Radic Biol Med 1996; 21: 291–5.
5. Ichiba M, Maeta Y, Mukoyama T, Saeki T, Yasui S,KanbeT, et al. Expression of 8-hydroxy-20-deoxyguanosinein chronic liver disease and hepatocellular carcinoma. LiverInt 2003; 23: 338–45.
6. Kasai H, Nishimura S. Hydroxylation of deoxyguanosineat the C-8 position by ascorbic acid and other reducingagents. Nucleic Acids Res 1984; 12: 2137–45.
7. Floyd RA,West M S, Eneff KL, Hogsett W E, TingeyD T. Hydroxyl free radical mediated formation of 8-hydro-xyguanosine in isolated DNA. Arch Biochem Biophys 1988;262: 266–72.
8. Kamimura S, Gaal K, Britton R S, Bacon B R, Tria-daifilopoulos G, Tsukamoto H. Increased 4-hydroxyno-nenal levels in experimental alcoholic liver disease:association of lipid peroxidation with liver fibrogenesis.Hepatology 1992; 16: 448–53.
9. Parola M, Pinzani M, Casini A, Albano E, Poli G,Gentilini A, et al. Stimulation of lipid peroxidation or 4-hydroxynonenal treatment increases procollagen a1(I) geneexpression in human liver fat-storing cells. Biochem BiophysRes Commun 1993; 194: 1044–50.
10. Bacot S, Bernoud-Hubac N, Baddas N, Chantegrel B,Deshayes C, Doutheau A, et al. Covalent binding tohydroxyl-alkenals 4-HDDE, 4-HHE, and 4-HNE to ethano-lamine phospholipids subclasses. J Lipid Res 2003; 44: 917–26.
11. Guichardant M, Chantegrel B, Deshayes C,Doutheau A, Moliere P, Lagarde M. Specific markersof lipid peroxidation issued from n-3 and n-6 fatty acids.Biochem Soc Trans 2004; 32: 139–40.
12. Yamagami K, Yamamoto Y, Kume M, Ishikawa Y, Ya-maoka Y, Hiai H, et al. Formation of 8-hydroxy-20-deoxyguanosine and 4-hydroxy-2-nonenal-modified pro-teins in rat liver after ischemia-reperfusion: distinct localiza-tion of the two oxidatively modified products. AntioxidantsRedox Signal 2000; 2: 127–36.
13. Mahmood S, Kawanaka M, Kamei A, Izumi A, Na-
kata K, Niyama G, et al. Immunohistochemical evaluationof oxidative stress markers in chronic hepatitis C. Antiox-idant Redox Signal 2004; 6: 19–24.
14. Lee Y-Y, Kim H-G, Jung H-I, Shin Y-H, Hong S-M,park E-H, et al. Activities of antioxidant and redoxenzymes in human normal hepatic and hepatoma cell lines.Mol Cells 2002; 14: 305–11.
15. Nordberg J, Arner S J. Reactive oxygen species, antiox-idants, and the mannmalian thioredoxin system. Free Ra-dical Biol Med 2001; 31: 1287–312.
16. Farinati F, Cardin R, Dgan P, De Maria N, Floyd R
A, Van Thiel D H, et al. Oxidative DNA damage incirculating leukocytes occurs as an early event in chronicHCV infection. Free Radic Biol Med 1999; 27: 1284–91.
17. Cardin R, Saccoccio G, Masutti F, Bellentani S,Farinati F, Tiribelli C. DNA oxidative damage in leu-kocytes correlates with the severity of HCV-related liverdisease: validation in an open population study. J Hepatol2001; 34: 587–92.
18. Loft S, Vistisen K, Ewertz M, Tjonneland A, Overad
K, Poulsen H E. Oxidative DNA damage estimated by 8-hydroxyguansoine excretion in humans: influence of smoking,gender and body mass index. Carcinogenesis 1992; 13: 2241–7.
19. Sumida Y, Nakashima T, Yoh T, Furutani M, Hiro-
hama A, Kakisaka Y, et al. Serum thioredoxin levels asa predictor of steatohepatitis in patients with nonalcoholicfatty liver diseases. J Hepatol 2003; 38: 32–8.
20. Yesilova Z, Yaman H, Oktenli C, Ozcan A,Uygun A,Cakir E, et al. Systemic markers of lipid peroxidation andantioxidants in patients with nonalcoholic fatty liver dis-ease. Am J Gastroenterol 2005; 100: 850–5.
21. Kumar A, etzler E, Berger M. Isolation and character-ization of complement receptor type 1 (CR1) storagevesicles from human neutrophils using antibodies to thecytoplasmic tail of CR1. Blood 1997; 89: 4555–65.
22. Evans M K, Cooke M S, Podmore I D, Zheng Q, Her-
bert K E, Lunec J. Discrepancies in the measurement ofUV-induced 8-oxo-20-deoxyguanosine: implications for theanalysis of oxidative DNA damage. Biochem Biophys ResCommun 1999; 259: 374–8.
23. Boya P, de la Pena A, Beloqui O, Conchillo M, Castel-ruiz Y, Civerira M-P, et al. Antioxidant status and gluta-thione metabolism in peripheral blood mononuclear cells frompatients with chronic hepatitis C. J Hepatol 1999; 31: 808–14.
24. Bantel H, Lugering A, Heidemann J, Volkmann X,Poremba C, Strassburg C P, et al. Detection of apoptoticcaspase activity in sera from patients with chronic HCVinfection is associated with fibrotic liver injury. Hepatology2004; 40: 1078–87.
25. Kumaraguruparan R, Kabalimoorthy J, Nagini S.
Correlation of tissue lipid peroxidation and antioxidantswith clinical stage and menopausal status in patients withadenocarcinoma of the breast. Clin Biochem 2005; 38: 154–8.
26. Gladyshev V N, FactorV M, Housseau F, Hatfield D
L. Contrasting patterns of regulation of the antioxidantselenoproteins, thioredoxin reductase, and glutathione per-oxidase, in cancer cells. Biochem Biophys Res Commun1998; 251: 488–93.
27. Nordberg J, Arner E S J. Reactive oxygen species,antioxidants, and the mammalian thioredoxin system.Free Radical Biol Med 2001; 31: 1287–312.
28. Rubartelli A, Bonifaci N, Sitia R. High rates ofthioredoxin secretion correlate with growth arrest in hepa-toma cells. Cancer Res 1995; 55: 675–80.
29. Sumida Y, Nakashima T, Yoh T, Furutani M, Hiro-
hama A, Kakisaka Y, et al. Serum thioredoxin levels asa predictor of steatohepatitis in patients with nonalcoholicfatty liver disease. J Hepatol 2003; 38: 32–8.
30. Okuyama H, Nakamura H, Shimahara Y, Araya S,Kawada N, Yamaoka Y, et al. Overexpression of thior-edoxin prevents acute hepatitis caused by thioacetamide orlipopolysaccharide in mice. Hepatology 2003; 37: 1015–25.
31. Allan C B, Lacourciere G M, Stadtman T C. Respon-siveness of selenoproteins to dietary selenium. Ann RevNutr 1999; 19: 1–16.
32. Al-Taie OH, Uceyler N, EuSner U, Jakob F, Mork H,Scheurlen M, et al. Expression profiling and geneticalterations of the selenoproteins GI-gpx and sepp in color-ectal carcinogenesis. Nutr Cancer 2004; 48: 6–14.
33. Tan M, Li S, Swaroop M, Guan K, Oberleys LW, SunY. Trancriptional activation of the human glutathione prox-idase promoter by p53. J Biol Chem 1999; 274: 12061–6.
34. Bureau C, Bernard J, Chaouche N, Orfila C, BeraudM, Gonindard C, et al. Nonstructural 3 protein ofhepatitis C virus triggers an oxidative burst in humanmonocytes via activation of NADPH oxidase. J BiolChem 2001; 76: 23077–83.
165
Oxidative stress-related molecules in chronic viral hepatitis