evaluation of physiological traits for improving drought tolerance in faba bean (vicia faba l.)
TRANSCRIPT

REGULAR ARTICLE
Evaluation of physiological traits for improving droughttolerance in faba bean (Vicia faba L.)
Habib ur Rahman Khan Æ W. Link ÆT. J. Hocking Æ F. L. Stoddard
Received: 25 September 2006 / Accepted: 6 February 2007 / Published online: 3 March 2007� Springer Science+Business Media B.V. 2007
Abstract Among grain legumes, faba bean is
becoming increasingly popular in European agri-
culture due to recent economic and environmen-
tal interests. Faba bean can be a highly productive
crop, but it is sensitive to drought stress and yields
can vary considerably from season to season.
Understanding the physiological basis of drought
tolerance would indicate traits that can be used as
indirect selection criteria for the development of
cultivars adapted to drought conditions. To assess
genotypic variation in physiological traits associ-
ated with drought tolerance in faba bean and to
determine relationships among these attributes,
two pot experiments were established in a growth
chamber using genetic materials that had previ-
ously been screened for drought response in the
field. Nine inbred lines of diverse genetic back-
grounds were tested under adequate water supply
and limited water conditions. The genotypes
showed substantial variation in shoot dry matter,
water use, stomatal conductance, leaf tempera-
ture, transpiration efficiency, carbon isotope dis-
crimination (D13C), relative water content (RWC)
and osmotic potential, determined at pre-flower-
ing vegetative stage. Moisture deficits decreased
water usage and consequently shoot dry matter
production. RWC, osmotic potential, stomatal
conductance and D13C were lower, whereas leaf
temperature and transpiration efficiency were
higher in stressed plants, probably due to re-
stricted transpirational cooling induced by stoma-
tal closure. Furthermore, differences in stomatal
conductance, leaf temperature, D13C and transpi-
ration efficiency characterized genotypes that
were physiologically more adapted to water
deficit conditions. Correlation analysis also
showed relatively strong relationships among
these variables under well watered conditions.
The drought tolerant genotypes, ILB-938/2 and
Melodie showed lower stomatal conductance
associated with warmer leaves, whereas higher
stomatal conductance and cooler leaves were
observed in sensitive lines (332/2/91/015/1 and
Aurora/1). The lower value of D13C coupled with
higher transpiration efficiency in ILB-938/2, rel-
ative to sensitive lines (Aurora/1 and Condor/3),
is indeed a desirable characteristic for water-
limited environments. Finally, the results showed
that stomatal conductance, leaf temperature and
H. R. Khan (&) � T. J. HockingSchool of Applied Sciences, University ofWolverhampton, Wulfruna Street, WolverhamptonWV1 1SB, UKe-mail: [email protected]
W. LinkInstitute of Agronomy and Plant Breeding, vonSiebold Straße 8, 37075 Gottingen, Germany
F. L. StoddardDepartment of Applied Biology, University ofHelsinki, P.O. Box 27, 00014 Helsinki, Finlande-mail: [email protected]
123
Plant Soil (2007) 292:205–217
DOI 10.1007/s11104-007-9217-5

D13C are promising physiological indicators for
drought tolerance in faba bean. These variables
could be measured in pot-grown plants at ade-
quate water supply and may serve as indirect
selection criteria to pre-screen genotypes.
Keywords Carbon isotope discrimination �Leaf temperature � Stomatal conductance �Transpiration efficiency
Abbreviations
D13C Carbon isotope discrimination
RWC Relative water content
TE Transpiration efficiency
Introduction
Production systems in European agriculture have
been predominantly cereals-based but in the
recent past, economic and environmental devel-
opments have revived interest in grain legumes as
break crops that break cereal disease cycles, fix
nitrogen and yield pulses for feed and food uses
(Plies-Balzer et al. 1995; Robson et al. 2002).
Among the grain legumes, faba bean is adapted to
a broad range of environmental conditions and is
the subject of considerable attention in Europe.
The cultivated area of faba bean in the UK, for
example, has increased by 87% during the last
10 years (DEFRA Statistics 2005). Nevertheless,
yields have remained lower and more variable
than those of cereals. Drought is a major abiotic
constraint responsible for heavy production losses
(Link et al. 1999; Amede et al. 1999; Ricciardi
et al. 2001). Furthermore, the climatic-change
models predict that in many regions of Europe,
drought losses and yield variability of field crops
will increase (Marsh 1996).
Grain legumes respond to drought differently
and express various drought tolerance strategies
(Subbarao et al. 1995). Faba bean is more sensi-
tive to drought than some other grain legumes
including common bean, pea and chickpea
(McDonald and Paulsen 1997; Amede and Schu-
bert 2003). Although genotypic differences in the
response of faba bean to drought have been
documented (Heringa et. al. 1984; Grzesiak et al.
1997; Abdelmula et al. 1999; Amede et al. 1999;
Link et al. 1999), the physiological processes
associated with drought tolerance are less under-
stood than for other crop species.
The development of drought-tolerant cultivars
is essential to improve the yield stability of faba
bean. Plant breeders have considerable capacity
to evaluate breeding material for drought toler-
ance under field conditions based on grain yield at
drought-prone sites (Grzesiak et al. 1997; Link
et al. 1999), but the routine evaluation of drought
tolerance in the field is expensive, prolonged and
often produces variable results due to seasonal
variation. Blum (1984) suggested that genotypes
that show better performance under hostile envi-
ronments generally possess some unidentified
physiological attributes of tolerance to environ-
mental stresses in good conditions. It is impera-
tive for breeding drought resistant cultivars to
identify the specific physiological traits that
improve adaptation to water-limited environ-
ments (Subbarao et al. 1995). So far, progress
using this approach has been slow mainly due to
lack of an efficient screening technique (Wery
et al. 1994; Stoddard et al. 2006). An ideal
screening technique should be non-destructive,
rapid, accurate and able to handle many samples
(Reynolds et al. 1994). Therefore, understanding
the physiological basis of drought tolerance in
faba bean is important to identify traits, which can
be used as indirect selection criteria. The objec-
tives of the present investigation were first, to
determine the magnitude of genetic diversity in
physiological traits related to drought tolerance in
faba bean inbred lines and second, to explore
relationships among potentially useful traits to be
used in breeding programs for drought tolerance.
Materials and methods
The experiments were designed as completely
randomized factorial designs with four replica-
tions. Plants were grown in 4 L plastic pots filled
with 3.0 kg potting mix containing all essential
nutrients. The soil moisture was kept at field
capacity (20% w/w) in all pots by applying water
on alternate days until the induction of moisture
stress treatment. Water use increased with plant
206 Plant Soil (2007) 292:205–217
123

growth. Four inbred lines were used in experiment
1 and six inbred lines in experiment 2 (Table 1).
Lines of diverse genetic backgrounds were chosen
from those that Abdelmula et al. (1999) and Link
et al. (1999) characterized for drought response in
field conditions. Seeds were germinated on moist
Whatman filter paper at 20�C for 72 h in an
incubator. Two radicle-emerged seeds were sown
at 3-cm depth in each pot and were later thinned to
one plant per pot. Plants were not inoculated with
Rhizobium in these experiments. A layer of 3-mm
white polypropylene beads ~1 cm deep was placed
over the soil surface as mulch and reduced evapo-
ration to 5–7 mL day–1 in unplanted pots. Plants
were grown in a growth chamber (1400 Plant Growth
Room, Fisons Environmental Equipment, Lough-
borough, UK). Pots were 15 cm apart and photope-
riod was adjusted to 14 h light and 10 h dark, and the
temperature was 20�C day/10�C night ± 2�C. Pho-
tosynthetic photon flux density (PPFD) was about
300 lmol m–2 s–1 at the canopy level.
Moisture stress was induced in half of the pots
43 days after sowing (experiment 1) or 41 days
after sowing (experiment 2) by decreasing water
application by 2% (w/w) per day to bring the
moisture level down from field capacity (20% w/
w) to moisture stress (2–4% w/w). Pots were
weighed daily and where water use exceeded 2%,
water was applied. The aim was to control the
development of moisture stress to a slow rate that
was kept to a consistent level across accessions.
Water use was estimated from the loss in weight
by weighing the pots at watering. Relative water
content (RWC%) was determined on five leaf discs
of 6 mm diameter each, punched from the upper-
most fully expanded leaf of the main stem, 7–
8 days after the induction of water stress. After the
fresh weight was determined, then the discs were
floated for 20 h at room temperature on deionized
water in a covered Petri dish at light near the
compensation point (10–12 lE m–2 s–1). After
rehydration, the discs were surface-dried using tissue
paper, transferred into glass vial and turgid weight
was obtained. The final dry weight was determined
after drying in a pre-heated oven at 80�C for 24 h.
RWC was calculated according to Turner (1981).
Water relation measurements were taken
between 1100 and 1300 hours. Stomatal conduc-
tance was determined 6 days after induction of
water stress using a porometer (Mk 3, Delta- T
Devices, Cambridge, UK) on the abaxial surface
of the leaflets of the uppermost fully expanded
leaf. Leaf temperature readings of the three
leaflets of a fully illuminated youngest expanded
leaf were taken with a non-contact infrared
thermometer (Raynger ST60 ProPlus, Raytek,
Santa Cruz, CA, USA), held at an appropriate
angle about 10 cm above the leaf, focussing the
laser point in the centre of the leaflet. To measure
osmotic potential (wp), a leaflet from the same
leaf used for RWC was excised directly into a
plastic microcentrifuge tube and immediately
frozen in liquid nitrogen. The tubes were later
transferred to –30�C and kept frozen until mea-
surements. The frozen leaf material was allowed
to thaw at room temperature for 30 min and sap
was extracted using a 2.5-mL syringe. A filter
paper disc was placed in the sampling chamber of
a vapour pressure osmometer (model 5100C;
Wescor, Logan, UT, USA) and 8 lL sap was
applied to saturate the disc to measure osmotic
potential. At the end of 10-day drying cycle, the
youngest expanded leaf (usually the one following
the leaf chosen for osmotic potential analysis) was
collected from water stressed and well watered
plants and dried in an oven at 80�C for 24 h for
carbon isotope determination. The stable carbon
isotope composition in the leaf tissue was deter-
mined by elemental analyser isotope ratio mass
spectrometry (Iso-analytical Ltd., Sandbach,
Cheshire, UK) using standard procedures and
techniques. Carbon isotope discrimination (D13C)
was calculated according to Farquhar and Rich-
ards (1984), assuming an isotope composition for
Table 1 Names and origins of accessions used to evaluatedrought response in faba bean
Accession Origin Experiment
332/2/91/015/1 Interpoola 2Aurora/1 Sweden 1BB686WN/1 Germany 2Condor/3 Germany 1ILB 2282/2 Ecuador 2ILB 938/2 ICARDA 1, 2Melodie France 1, 2Victor/2 Netherlands 2
a Inbred line derived from Mediterranean · CentralEuropean cross
Plant Soil (2007) 292:205–217 207
123
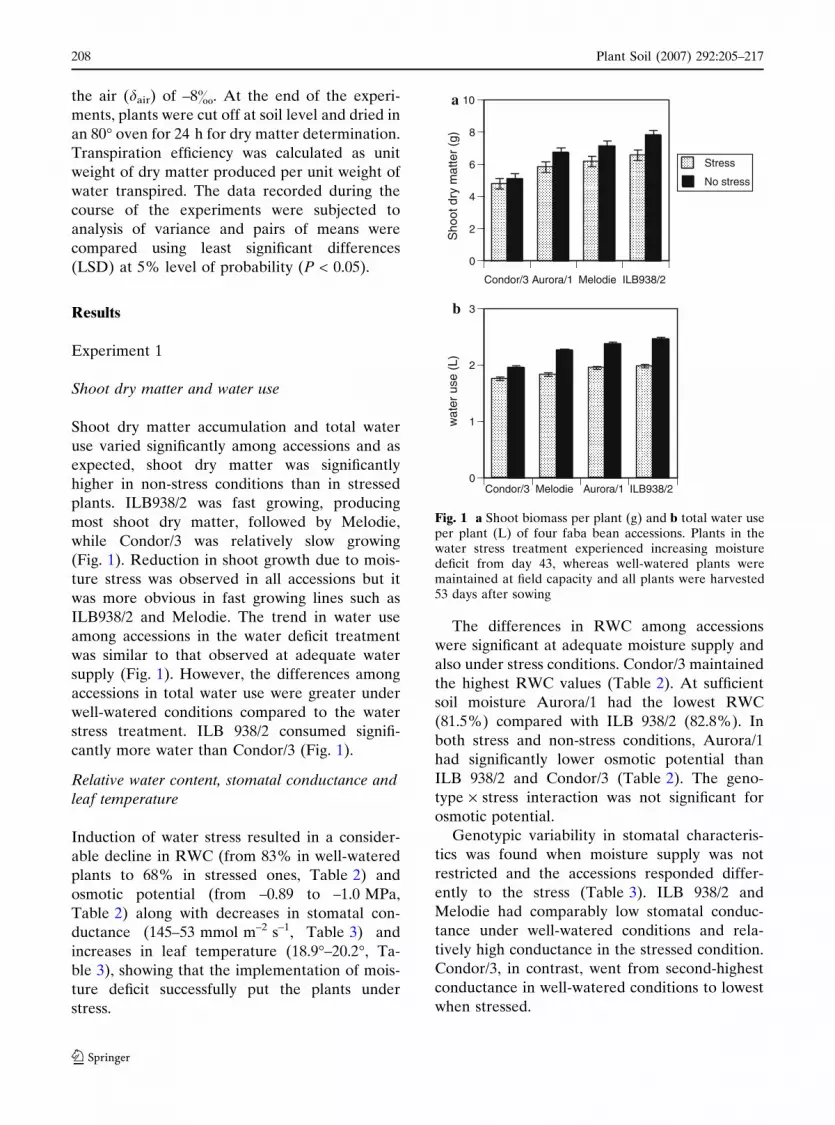
the air (dair) of –8&. At the end of the experi-
ments, plants were cut off at soil level and dried in
an 80� oven for 24 h for dry matter determination.
Transpiration efficiency was calculated as unit
weight of dry matter produced per unit weight of
water transpired. The data recorded during the
course of the experiments were subjected to
analysis of variance and pairs of means were
compared using least significant differences
(LSD) at 5% level of probability (P < 0.05).
Results
Experiment 1
Shoot dry matter and water use
Shoot dry matter accumulation and total water
use varied significantly among accessions and as
expected, shoot dry matter was significantly
higher in non-stress conditions than in stressed
plants. ILB938/2 was fast growing, producing
most shoot dry matter, followed by Melodie,
while Condor/3 was relatively slow growing
(Fig. 1). Reduction in shoot growth due to mois-
ture stress was observed in all accessions but it
was more obvious in fast growing lines such as
ILB938/2 and Melodie. The trend in water use
among accessions in the water deficit treatment
was similar to that observed at adequate water
supply (Fig. 1). However, the differences among
accessions in total water use were greater under
well-watered conditions compared to the water
stress treatment. ILB 938/2 consumed signifi-
cantly more water than Condor/3 (Fig. 1).
Relative water content, stomatal conductance and
leaf temperature
Induction of water stress resulted in a consider-
able decline in RWC (from 83% in well-watered
plants to 68% in stressed ones, Table 2) and
osmotic potential (from –0.89 to –1.0 MPa,
Table 2) along with decreases in stomatal con-
ductance (145–53 mmol m–2 s–1, Table 3) and
increases in leaf temperature (18.9�–20.2�, Ta-
ble 3), showing that the implementation of mois-
ture deficit successfully put the plants under
stress.
The differences in RWC among accessions
were significant at adequate moisture supply and
also under stress conditions. Condor/3 maintained
the highest RWC values (Table 2). At sufficient
soil moisture Aurora/1 had the lowest RWC
(81.5%) compared with ILB 938/2 (82.8%). In
both stress and non-stress conditions, Aurora/1
had significantly lower osmotic potential than
ILB 938/2 and Condor/3 (Table 2). The geno-
type · stress interaction was not significant for
osmotic potential.
Genotypic variability in stomatal characteris-
tics was found when moisture supply was not
restricted and the accessions responded differ-
ently to the stress (Table 3). ILB 938/2 and
Melodie had comparably low stomatal conduc-
tance under well-watered conditions and rela-
tively high conductance in the stressed condition.
Condor/3, in contrast, went from second-highest
conductance in well-watered conditions to lowest
when stressed.
Condor/3 Aurora/1 Melodie ILB938/2
0
2
4
6
8
10
Stress
No stress
Sho
ot d
ry m
atte
r (g
)w
ater
use
(L)
Condor/3 Melodie Aurora/1 ILB938/20
1
2
3
a
b
Fig. 1 a Shoot biomass per plant (g) and b total water useper plant (L) of four faba bean accessions. Plants in thewater stress treatment experienced increasing moisturedeficit from day 43, whereas well-watered plants weremaintained at field capacity and all plants were harvested53 days after sowing
208 Plant Soil (2007) 292:205–217
123

Leaf temperature by contrast, showed no
significant difference among the lines in the stress
conditions, but significant difference under the
non-stress conditions, with Aurora/1 and Condor/
3 having significantly lower leaf temperatures
than ILB 938/2 and Melodie (Table 3).
Carbon isotope discrimination and transpiration
efficiency
Carbon isotope discrimination varied significantly
among accessions and was also affected by stress
levels, while the accession · stress interaction was
not significant (Table 4). Aurora/1 and Condor/3
had equally high values of D13C (23.3 · 10–3),
while ILB 938/2 had the lowest (21.6 · 10–3). The
values of D13C were significantly lower in water
stressed plants but the ranking of accessions was
not altered by water regimes.
Transpiration efficiency increased when plants
suffered from moisture stress but the acces-
sion · stress interaction was not significant. Melo-
die and ILB-938/2 were more efficient in water
use and maintained significantly higher transpira-
tion efficiency then Aurora/1 and Condor/3,
regardless of water stress levels (Table 4).
Relationships among physiological traits
Correlation analysis showed some strong rela-
tionships among physiological traits. As expected,
shoot dry matter and transpiration efficiency were
Table 2 Effect of water deficit on relative water content (%) and osmotic potential (-MPa) of four faba bean accessions
Genotypes Relative water content (%) Osmotic potential (-MPa)
Stress No stress Stress No stress
Aurora/1 68.1 81.5 1.02 0.83Condor/3 70.2 83.4 0.98 0.76ILB 938/2 66.4 82.8 0.91 0.78Melodie 68.3 82.7 1.09 0.81Mean 68.2 82.6 1.00 0.79LSD (P £ 0.05)Genotype 1.0** 0.04**Stress 0.7** 0.03**Genotype · stress 1.4** NS
Relative water content and osmotic potential was recorded 8 and 9 days after the induction of water stress, respectively
NS non significant
** P < 0.01
Table 3 Stomatal conductance (mmol m–2 s–1) and Leaf temperature (�C) as influenced by water deficit in four faba beanaccessions
Genotypes Stomatal conductance (mmol m–2 s–1) Leaf temperature (�C)
Stress No stress Stress No stress
Aurora/1 58 176 20.3 18.3Condor/3 30 152 20.2 18.6ILB 938/2 56 125 20.1 19.3Melodie 68 127 20.2 19.4Mean 53 145 20.2 18.9LSD (P £ 0.05)Genotype 13** NSStress 10** 0.3**Genotype · stress 19** 0.6*
Measurements were taken 6 days after the induction of water stress
NS non significant
*, ** P < 0.05, 0.01, respectively
Plant Soil (2007) 292:205–217 209
123

negatively correlated with D13C, showing parallel
lines for the two water treatments (Fig. 2). In the
well-watered plants, leaf temperature and specific
leaf weight exhibited strong negative correlation
with D13C (Fig. 3), while significant positive rela-
tionships were observed between stomatal con-
ductance and D13C. In stress conditions these
relationships were generally non-significant.
Experiment 2
Relative water content and osmotic potential
Induction of water stress was again successful at
reducing RWC, osmotic potential (Table 5). ILB
938/2 and Melodie had significantly higher RWC%
than Victor/2 and line 332/2/91/015/1, which con-
sistently was the lowest (Table 5). Differences in
osmotic potential were also found at adequate
moisture supply, and Melodie and 332/2/91/015/1
had lower osmotic potential than Victor/2 and BB
686WN/1 (Table 5). Moisture stress resulted in a
significant decline in osmotic potential in all entries
except ILB938/2; the highest decrease (0.17 MPa)
was recorded in Victor/2.
Transpiration efficiency, stomatal conductance
and leaf temperature
Transpiration efficiency based on shoot dry mat-
ter accumulation varied significantly among
accessions (Fig. 4). ILB 938/2 was once again
most efficient in water use (4.03 g L–1), while the
lowest value of transpiration efficiency was
Table 4 Effect of water stress on carbon isotope discrimination (D · 10–3) and transpiration efficiency (g L–1) in four fababean genotypes
Genotypes Carbon isotope discrimination (D · 10–3) Transpiration efficiency (g L–1)
Stress No stress Stress No stress
Aurora/1 21.4 23.3 2.99 2.82Condor/3 21.7 23.3 2.73 2.61ILB 938/2 20.1 21.6 3.30 3.16Melodie 20.5 22.4 3.37 3.15Mean 20.9 22.7 3.10 2.94LSD (P £ 0.05)Genotype 0.4** 0.13**Stress 0.3** 0.09**Genotype · stress NS NS
Plants were harvested 53 days after sowing and subjected to water deficit for the last 10 days
NS non significant
** P < 0.01
15
17
19
21
23
25
2 3 4 5 6 7 8 9
Car
bon
isot
ope
disc
rimin
atio
n C
arbo
n is
otop
e di
scrim
inat
ion
Stress
Well-watered
y = 24.3 - 0.57 x
r = - 0.68**
y = 25.9 - 0.48 x
r = - 0.64**
15
17
19
21
23
25
2.0 2.5 3.0 3.5 4.0
Transpiration efficiency
Shoot dry matter
y = 27.6 - 2.15 x
r = - 0.81**
y = 28.9 - 2.08 x
r = - 0.84**
a
b
Fig. 2 Relationship of carbon isotope discrimination(D · 10–3) with a shoot dry matter production (g) and btranspiration efficiency (g L–1). Plants in the stresstreatment received restricted supply of water for the last10 days whereas the other plants were well watered
210 Plant Soil (2007) 292:205–217
123

recorded in line 332/2/91/015/1. The difference in
transpiration efficiency between water treatments
was not significant.
Induction of water stress reduced stomatal
conductance (Table 6) along with increasing leaf
temperature. At adequate moisture supply,
ILB938/2 and Melodie had significantly lower
stomatal conductance than lines 332/2/91/015/1,
ILB2282/2 and Victor/2 (Table 6). The decline of
stomatal conductance in ILB 938/2 in response to
water stress was much more rapid than in 332/2/
91/015/1 (Fig. 5). The decrease in stomatal con-
ductance was significantly higher in BB686WN/1,
ILB 938/2 and Melodie, compared to 332/2/91/
015/1 and ILB2282/2.
The entries had significant variation in leaf
temperature even when moisture supply was
adequate (Table 6). ILB938/2 had warmer leaves
than 332/2/91/015/1 and ILB2282/2. Although
moisture stress raised the leaf temperature in all
genotypes, lines 332/2/91/015/1, ILB2282/2 and
BB686WN/1 showed less change than ILB938/2.
Relationships among physiological traits
In this experiment, stomatal resistance under
well-watered conditions was again positively cor-
related with transpiration efficiency and leaf
temperature, and unlike experiment 1 with
RWC (data not shown). At high water supply,
leaf temperature had a significant and positive
correlation with leaf RWC (not significant in
experiment 1) and, as before, transpiration effi-
ciency (Fig. 6). Transpiration efficiency was pos-
itively correlated with RWC but the slope was not
significantly different between water treatments
(data not shown).
Discussion
The present study showed wide variation in the
transpiration characteristics of faba bean lines.
These traits were correlated with each other, to a
greater or lesser degree, and the variation in some
traits in well-watered conditions was indicative of
the variation in drought response in the field.
The chosen faba bean lines varied considerably
in shoot dry matter production and water use
pattern. The imposed reduction in water supply
was sufficient to induce moisture stress, which
consequently reduced plant growth. After 10 days
of restricted water supply, leaves had wilted,
particularly in Aurora/1 and line 332/2/91/015/1.
Bond et al. (1994) reported that the faba bean
18
20
22
24
26
50 100 150 200 250
Stomatal conductance
Car
bon
isot
ope
disc
rimin
atio
nC
arbo
n is
otop
e di
scrim
inat
ion
Car
bon
isot
ope
disc
rimin
atio
n
y = 20.2 + 0.017 *X
r = 0.62*
16
18
20
22
24
26
18 19 20 21 22
Leaf temperature
y = 38.5 - 0.79 x
r = - 0.74**
18
20
22
24
2.0 2.5 3.0 3.5 4.0
Specific leaf weight
y = 8.36 - 0.23 x
r = - 0.78**
a
b
c
Fig. 3 Relationship of carbon isotope discrimination(D · 10–3) with a stomatal conductance (mmol m–2 s–1), bleaf temperature (�C), and c specific leaf weight (mg cm–2)in well watered plants
Plant Soil (2007) 292:205–217 211
123

Table 5 Effect of water deficit on relative water content (%) 7 days after induction of water stress and osmotic potential(-MPa) 8 days after induction in six faba bean genotypes
Genotypes Relative water content (%) Osmotic potential (-MPa)
Stress No stress Stress No stress
332/2/91/015/1 61.1 68.2 0.90 0.81BB 686 WN/1 73.1 77.6 0.86 0.75ILB 2282/2 68.3 77.3 0.86 0.77ILB 938/2 74.2 77.7 0.79 0.78Victor/2 70.3 73.9 0.91 0.74Melodie 71.0 78.6 0.91 0.82Mean 69.6 75.6 0.87 0.78LSD (P £ 0.05)Genotype 2.3** 0.04**Stress 1.3** 0.02**Genotype · stress NS 0.05**
NS non significant
** P < 0.01
332/2/91/015/1 ILB2282/2 Victor/2 Melodie
Tra
nspi
ratio
n ef
ficie
ncy
BB686 WN ILB938/2
0
1
2
3
4
Fig. 4 Variation intranspiration efficiency(g L–1) among six fababean inbred lines. Plantswere harvested at pre-flowering stage, 50 daysafter sowing
Table 6 Stomatal conductance (mmol m–2 s–1) 5 days after induction of water stress and leaf temperature (�C) 6 days afterinduction in six faba bean genotypes
Genotypes Stomatal conductance (mmol m–2 s–1) Leaf temperature (�C)
Stress No stress Stress No stress
332/2/91/015/1 79 175 18.7 16.2BB 686 WN/1 35 119 18.7 17.0ILB 2282/2 67 141 18.2 16.9ILB 938/2 39 99 19.2 17.6Victor/2 45 137 18.9 17.4Melodie 41 106 18.9 17.1Mean 519 129 18.8 17.0LSD (P £ 0.05)Genotype 19** 0.3**Stress 11** 0.2**Genotype · stress NS 0.4**
NS non significant
** P < 0.01
212 Plant Soil (2007) 292:205–217
123

plant requires a large amount of water to retain
turgor in its fleshy leaves, making it relatively
sensitive.
In our experiments, plants were at a consistent
vegetative stage (pre-flowering) when water stress
was induced. We found inherent differences in
growth rate among the lines, so care was taken to
induce an even moisture stress in the pots by
monitoring water use daily and applying water if
the water use was > 2% w/w. Pot experiments
have been successfully used previously with
legumes to facilitate identification of drought
tolerant breeding materials (Amede et al. 1999).
Balko et al. (1995) reported that the rank order of
faba bean lines subjected to moisture stress in
pots was reproducible and strongly correlated
with their relative yield in field trials. Ricciardi
et al. (2001) used controlled environments to
effectively induce water stress in faba bean and
it was also possible to simulate stress of different
intensities for evaluation of breeding material for
moisture deficit tolerance.
In the present study, numerous physiological
traits that can contribute towards yield under
drought conditions were assessed in entries of
diverse genetic backgrounds, representing a wide
range of origins. The accessions differed in their
water usage and the trend in water use in the
moisture deficit treatment was very similar to that
observed at adequate water supply. Furthermore,
the differences among genotypes were more obvi-
ous and consistent in the well-watered conditions
than in the water stress treatment. ILB 938/2 and
Melodie exhibited high transpiration efficiency in
both experiments indicating that these are more
efficient in water use. These results are in agree-
ment with the field observations, which character-
ized ILB 938/2 and Melodie as drought-tolerant,
with high relative yield under simulated drought
conditions (Abdelmula et al. 1999; Tolera and
Link 2002). In contrast, Aurora was sensitive to
0Day 1 Day 3 Day 5
50
100
150
200S
tom
atal
con
duct
ance
332/2/91/015/1 Stressed
332/2/91/015/1 Well watered
ILB 938/2 Stressed
ILB 938/2 Well watered
Fig. 5 Stomatal response of ILB 938/2 and 332/2/91/015/1to the development of moisture stress or well watered inpots under controlled condition (Experiment 2). Stomatalconductance measurements are in mmol m–2 s–1
123.0 3.5 4.0 4.5
14
16
18
Transpiration efficiency
y = 11.3 + 1.3 x
r = 0.66**
12
14
16
18
60 70 80 90Relative water content
Leaf
tem
pera
ture
Le
af te
mpe
ratu
re
y = 10.9 + 0.068 x
r = 0.59**
a
b
Fig. 6 Relationship of leaf temperature (�C) with atranspiration efficiency (g L–1), and b relative watercontent (%) in well watered plants. Transpiration effi-ciency is based on shoot biomass of plants, harvested atpre-flowering stage
Plant Soil (2007) 292:205–217 213
123

drought in field conditions (Amede et al. 1999) as
well as in our experiments.
Carbon isotope composition has been used to
determine the genotypic and environmental
responses of water use efficiency in several
legume species, including lentil (Matus et al.
1996), alfalfa (Johnson and Rumbaugh 1995),
common bean (Zacharisen et al. 1999) and chick-
pea (Khan et al. 2004) and now faba bean.
Our experiments showed considerable genetic
variation for D13C in the leaf material among faba
bean genotypes. The drought tolerant accessions,
ILB 938/2 and Melodie, had significantly lower
D13C than sensitive varieties such as Aurora/1 and
332/2/91/015/1 at adequate moisture supply.
Selection for low D13C has been proposed as a
method to select for improved water use effi-
ciency in breeding programs for C3 crop species
(Johnson and Rumbaugh 1995) and the present
data support this. Correlation analysis conducted
using genotypic means demonstrated that D13C
was negatively associated with transpiration effi-
ciency, stomatal resistance and leaf temperature.
Similarly, lentil showed genetic variation for D13C
(Matus et al. 1996), which was strongly and
negatively correlated with water use efficiency
(Johnson et al. 1995). Water stress also led to
reduced D13C and increased transpiration effi-
ciency in faba bean, indicating that drought
altered the balance between the stomatal con-
ductance and internal leaf capacity for photosyn-
thesis, which determine the internal leaf CO2
concentration and D13C, as observed in alfalfa
(Johnson and Tieszen 1994). The utility of D13C
for selection in a plant breeding program is
strongly enhanced by the consistency of genotypic
ranking for different water regimes (Hall et al.
1992). Our results suggest that D13C may be a
useful parameter for selecting faba bean geno-
types that use water efficiently under water-
limited conditions. Additional data concerning
the physiological basis of D13C response at
restricted moisture supply and association with
grain yield in faba bean, are required to confirm
that selection for low D13C would lead to signif-
icant gains in water use efficiency.
Variation in drought response between faba
bean lines was mainly associated with dehydra-
tion avoidance through stomatal control (Amede
et al. 1999). Nerkar et al. (1981) found consider-
able differences in water loss and transpiration in
faba bean genotypes having contrasting stomatal
features, and suggested that lower transpiration
rate would contribute to drought tolerance. Fur-
thermore, excessive water use in the drought-
sensitive faba bean cultivar Adriewaalse from
Netherlands was associated with its higher tran-
spiration, while L-7, a breeding line of Mediter-
ranean background was modest in water use and
hence drought tolerant (Amede et al. 1999).
The extent of physiological variability in the
stomatal characteristics among genotypes is of
primary importance, as it constitutes an efficient
control mechanism of water loss that can be used in
breeding programmes (Tanzarella et al. 1984).
Our results revealed marked variation in stomatal
conductance among inbred lines, with lower sto-
matal conductance and higher transpiration effi-
ciency in ILB938/2 and Melodie at adequate water
supply compared with lines 332/2/91/015/1 and
Victor/2. This confirms the physiological signifi-
cance of lower stomatal conductance in regulating
the water loss from leaves. Stomatal characteristics
vary widely in faba bean (Ricciardi 1989). Fre-
quency and size of stomata also differed between
drought-resistant faba bean cultivar Gobo and
sensitive cultivar Victor (Grzesiak et al. 1997).
Therefore, lower stomatal conductance seems a
useful physiological attribute for evaluation of
breeding material for superior water use efficiency.
We found that leaf temperature at adequate
water availability differentiated genotypes for
drought response. In the presence of drought
stress, when the stomatal conductance data indi-
cated that the stomata were largely closed, there
was little variation in leaf temperature. In well-
watered conditions, however, warmer leaves were
associated with low stomatal conductance and
high-transpiration efficiency, which are favourable
attributes for drought adaptation. Idso et al. (1981)
used the increase in average canopy temperature
following drought stress as a key component of a
crop water stress index and O’Neill et al. (2006)
considered leaf temperature as a potential indica-
tor of plant water stress, since increasing plant
water deficit leads to stomatal closure, decreases
transpirational cooling and consequently increases
leaf temperature.
214 Plant Soil (2007) 292:205–217
123

The application of infrared thermometry to
irrigated wheat demonstrated that canopy tem-
perature measurably decreased as stomatal con-
ductance increased (Fischer et al. 1998). Wheat
cultivars with the warmest canopy temperatures
under well watered conditions not only had the
lowest leaf conductance and the lowest seasonal
water use under normal irrigation practices but
they also had the most favourable yield response
when subjected to water deficit conditions (Pinter
et al. 1990). The difference between leaf temper-
ature and air temperature was linearly related to
stomatal conductance in spring wheat and there-
fore, it might serve as surrogate for stomatal
conductance (Amani et al. 1996). Pea cultivars
showing higher transpiration rate had higher
stomatal density that effectively reduced canopy
temperature due to transpirational cooling (San-
chez et al. 2001). Hence, non-invasive approaches
using infrared thermometry to classify genotypes
according to canopy temperature appear feasible
for the efficient screening for water use charac-
teristics and tolerance to drought (Pinter et al.
1990).
This study demonstrates the potential useful-
ness of leaf temperature data for identifying the
water use characteristics of faba bean genotypes.
A simple hand-held infrared thermometer can be
used for measuring leaf temperature relatively
easily and precisely in faba bean, owing to its
broad leaves. Furthermore, genotypic differences
can be detected in controlled environments,
reducing error due to subtle weather conditions.
Although it is possible to evaluate leaf tempera-
ture differences during the vegetative stage at
different water regimes, our results show that
measurements should be taken at ample soil
water to safeguard against differential rates of
water use confounding the results. Similarly,
Singh and Kanemasu (1983) observed leaf tem-
perature differences of 5.0�C in pearl millet
genotypes under well-watered conditions,
whereas differences in leaf temperature were less
in water stressed plants, and they suggested that
canopy temperature in a non-stressed environ-
ment would be a valid criterion for screening
genotypes for their yield stability.
Cost is important when considering a useful
indirect selection criterion for drought tolerance.
Determination of D13C is expensive and the non-
contact infrared sensing system offers exciting
possibilities for cost reduction (Fischer et al.
1998). Canopy temperature measurement using
the infrared thermometer has been recommended
as the physiological trait of choice for evaluating
cereal drought response, for the obvious reasons
that it is very rapid, giving instant measurements,
and can be measured on single leaves inexpen-
sively and quickly, unlike many other physiolog-
ical parameters (Reynolds et al. 1994). The
infrared thermometry may have potential utility
in water-limited environments because relatively
small differences in plant transpiration are gen-
erally translated into canopy temperature differ-
ences of several degrees (Pinter et al. 1990). Our
results confirm that leaf temperature measure-
ments show promise for identifying desirable
traits among genotypes in breeding nurseries,
preferably at adequate water supply, and infrared
thermometers may be used to pre-screen for
physiological responses related with drought tol-
erance under controlled conditions prior to the
execution of extensive yield trials.
Conclusion
This study has shown genotypic diversity for
drought-associated physiological traits in a di-
verse set of European and Mediterranean faba
bean inbred lines. Furthermore, the variation in
stomatal conductance, leaf temperature, transpi-
ration efficiency and D13C characterized geno-
types that are physiologically more adapted to
water deficits. A correlation analysis showed
strong relationships among these variables. Inter-
estingly, drought tolerant genotypes, ILB-938/2
and Melodie showed lower stomatal conductance
than drought-sensitive lines (332/2/91/015/1 and
Aurora/1), probably a drought avoidance mech-
anism for minimizing water loss through re-
stricted transpiration. The cooler canopy of
sensitive lines relative to ILB-938/2 and Melodie
further explained genotypic response to stomatal
characteristics, as wide and open stomata may
enhance transpirational cooling. The lower value
of D13C associated with higher transpiration
efficiency in ILB-938/2 would be regarded as a
Plant Soil (2007) 292:205–217 215
123

desirable characteristic in breeding programs for
water-limited environments.
Finally, it appears that stomatal conductance;
leaf temperature and D13C are potential indica-
tors for drought tolerance. Although these char-
acteristics were measured in pot-grown plants at a
pre-flowering vegetative stage, there is a possibil-
ity of exploiting these differences in a breeding
program to develop lines less susceptible to
drought stress. The study was conducted within
the framework of a faba bean breeding for
sustainable agriculture program geared mainly
towards identifying genotypes with physiological
attributes of superior drought adaptation. These
results obtained in pot experiments supported by
pre-existing field data will help assess the genetic
determination of differences in drought tolerance.
Acknowledgements This work was carried out with thefinancial support of EU that was provided under projectQLK5-CT-2002-02307 ‘‘Faba bean Breeding forSustainable Agriculture’’, acronym EUFABA. Technicalassistance of Robert Hooton and Fiona Bowers for thiswork is gratefully acknowledged.
References
Abdelmula AA, Link W, Kittlitz EV, Stelling D (1999)Heterosis and inheritance of drought tolerance in fababean, Vicia faba L. Plant Breed 118:485–490
Amani I, Fischer RA, Reynolds MP (1996) Canopy tem-perature depression association with yield of irrigatedspring wheat cultivars in a hot climate. J Agron CropSci 176:119–129
Amede T, Kittlitz EV, Schubert S (1999) Differentialdrought responses of faba bean (Vicia faba L.) inbredlines. J Agron Crop Sci 183:35–45
Amede T, Schubert S (2003) Mechanisms of droughtresistance in grain legumes I. Osmotic adjustment.Ethiop J Sci 26:37–46
Balko C, Stelling D, Laher F, Seddig S, Kittlitz EV, Jur-gens HU (1995) Investigations into selection fordrought tolerance in Vicia faba L. In Chapter 2.International Congress on Integrated Studies onDrought Tolerance of Higher Plants, August 31–September 2, Montpellier, France
Bond DA, Jellis GJ, Rowland GG, Guen J Le, RobertsonLD, Khalil SA, Li-Juan L (1994) Present status andfuture strategy in breeding faba beans (Vicia faba L.)for resistance to biotic and abiotic stresses. Euphytica73:151–166
Blum A (1984) Methods of selection for plant tolerance toenvironmental stresses. In: Selection in mutationbreeding. Proceedings of the Consultants’ Meeting,
Vienna, 21–25 June 1982. pp. 85–96. InternationalAtomic Energy Agency, Vienna, Austria
DEFRA (2005) Agriculture in the United Kingdom. GreatBritain Department for Environment, Food andRural Affairs, UK, pp. 143
Farquhar GD, Richards RA (1984) Isotope composition ofplant carbon correlates with water use efficiency ofwheat genotypes. Aust J Plant Physiol 11:539–552
Fischer RA, Rees D, Sayre KD, Lu ZM, Condon AG,Saavedra AL (1998) Wheat yield progress associatedwith higher stomatal conductance and photosyntheticrate, and cooler canopies. Crop Sci 38:1467–1475
Grzesiak S, Iijima M, Kono Y, Yamauchi A (1997) Dif-ferences in drought tolerance between cultivars offield bean and field pea. A comparison of drought-resistant and drought-sensitive cultivars. Acta PhysiolPlant 19:349–357
Hall AE, Mutters RG, Farquhar GD (1992) Genotypicand drought induced differences in carbon isotopediscrimination and gas exchange in cowpea. Crop Sci32:1–6
Heringa RJ, Norel A, Post J (1984) Searching for toleranceto drought in Vicia faba. FABIS Newslett 8:13–14
Idso SB, Jackson RD, Pinter PJ, Reginato RJ, Hatfield JL(1981) Normalizing the stress degree day parameterfor environmental variability. Agric Meteorol 24:45–55
Johnson RC, Muehlbauer FJ, Simon CJ (1995) Geneticvariation in water use efficiency and its relation tophotosynthesis and productivity in lentil germplasm.Crop Sci 35:457–463
Johnson DA, Rumbaugh MD (1995) Genetic variationand inheritance characteristics for carbon isotopediscrimination in alfalfa. J Range Manag 48:126–131
Johnson RC, Tieszen LL (1994) Variation for water useefficiency in alfalfa germplasm. Crop Sci 34:452–458
Khan HR, McDonald GK, Rengel Z (2004) Zinc fertil-ization and water stress affects plant water relations,stomatal conductance and osmotic adjustment inchickpea (Cicer arietinum L.). Plant Soil 267:271–284
Link W, Abdelmula AA, Kittlitz EV, Bruns S, Riemer H,Stelling D (1999) Genotypic variation for droughttolerance in Vicia faba. Plant Breed 118:477–483
Marsh TJ (1996) The 1995 UK drought—a signal of cli-matic instability? Proc Inst Civ Eng Water MaritEnergy 118:189–195
Matus A, Slinkard AE, Kessel CV (1996) Carbon isotopediscrimination and indirect selection for transpirationefficiency at flowering in lentil (Lens culinaris Medi-kus), spring wheat (Triticum aestivum L.), durumwheat (Triticum turgidum L.) and canola (Brassicanapus L.). Euphytica 87:141–145
McDonald GK, Paulsen GM (1997) High temperatureeffects on photosynthesis and water relations of grainlegumes. Plant Soil 196:47–58
Nerkar YS, Wilson D, Lawes DA (1981) Genetic variationin stomatal characteristics and behaviour, water useand growth of five Vicia faba L. genotypes undercontrasting soil moisture regimes. Euphytica 30:335–345
216 Plant Soil (2007) 292:205–217
123

O’Neill PM, Shanahan JF, Schepers JS (2006) Use ofchlorophyll fluorescence assessments to differentiatecorn hybrid response to variable water conditions.Crop Sci 46:681–687
Pinter PJ, Zipoli G, Reginato RJ, Jackson RD, Idso SB,Hohman JP (1990) Canopy temperature as an indi-cator of differential water use and yield performanceamong wheat cultivars. AgricWater Manag 18:35–48
Plies-Balzer E, Kong T, Schubert S, Mengel K (1995) Ef-fect of water stress on plant growth, nitrogenaseactivity and nitrogen economy of four different culti-vars of Vicia faba L. Eur J Agron 4:167–173
Reynolds MP, Balota M, Delgado MIB, Amani I, FischerRA (1994) Physiological and morphological traitsassociated with spring wheat yield under hot, irrigatedconditions. Aust J Plant Physiol 21:717–730
Robson MC, Fowler SM, Lampkin NH, Leifert C, LeitchM, Robinson D, Watson CA, Litterick AM (2002)The agronomic and economic potential of break cropsfor ley/arable rotations in temperate organic agricul-ture. Adv Agron 77:369–427
Ricciardi A (1989) Plant breeding for resistance to droughtII. Relationships between stomatal and agronomictraits in Vicia faba L. genotypes. Agric Mediterr119:424–434
Ricciardi L, Polignano GB, Giovanni CD (2001) Geno-typic response of faba bean to water stress. Euphytica118:39–46
Sanchez FJ, Manzanares M, de Andres EF, Tenorio JL,Ayerbe L (2001) Residual transpiration rate, epicu-ticular wax load and leaf colour of pea plants in
drought conditions. Influence on harvest index andcanopy temperature. Eur J Agron 15:57–70
Singh P, Kanemasu ET (1983) Leaf and canopy tempera-ture of pearl millet genotypes under irrigated andnon-irrigated conditions. Agron J 75:497–501
Stoddard FL, Balko C, Erskine W, Khan HR, Link W,Sarker A (2006) Screening techniques and sources ofresistance to abiotic stresses in cool-season food le-gumes. Euphytica 147:167–186
Subbarao GV, Johansen C, Slinkard AE, Nageswara RaoRC, Saxena NP, Chauhan YS (1995) Strategies forimproving drought resistance in grain legumes. CritRev Plant Sci 14:469–523
Tanzarella OA, De Pace C, Filippetti A (1984) Stomatalfrequency and size in Vicia faba L. Crop Sci 24:1070–1076
Tolera T, Link W (2002) Genotypische Unterschiedezwischen Europaischen Elitesorten von Vicia faba L.im Merkmal Trockenheitstoleranz. Vortrage furPflanzenzuchtung 54:465–468 (In German)
Turner NC (1981) Techniques and experimental ap-proaches for the measurement of plant water status.Plant Soil 58:339–366
Wery J, Silim SN, Knights EJ, Malhotra RS, Cousin S(1994) Screening techniques and sources and toler-ance to extremes of moisture and air temperature incool season food legumes. Euphytica 73:73–83
Zacharisen MH, Brick MA, Fisher AG, Ogg JB, Ehlerin-ger JR (1999) Relationships between productivity andcarbon isotope discrimination among dry bean linesand F2 progeny. Euphytica 105:239–250
Plant Soil (2007) 292:205–217 217
123